
Intrathymic adeno-associated virus gene transfer rapidly ...
Upload
independentCategory
view
0download
0
Gene Therapy (2002) 9, 1576–1588 2002 Nature Publishing Group All rights reserved 0969-7128/02 $25.00
www.nature.com/gt
RESEARCH ARTICLE
Adeno-associated virus vector-mediated gene transferinto dystrophin-deficient skeletal muscles evokesenhanced immune response against the transgeneproduct
K Yuasa1, M Sakamoto1, Y Miyagoe-Suzuki1, A Tanouchi1, H Yamamoto2, J Li3, JS Chamberlain4,X Xiao3 and S Takeda1
1Department of Molecular Therapy, National Institute of Neuroscience, National Center of Neurology and Psychiatry, Tokyo, Japan;2Department of Immunology, Graduate School of Pharmaceutical Sciences, Osaka University, Osaka, Japan; 3Department ofMolecular Genetics and Biochemistry, Gene Therapy Center, and Duchenne Muscular Dystrophy Research Center, University ofPittsburgh, Pittsburgh, PA, USA; and 4Department of Neurology, University of Washington School of Medicine, Seattle, WA, USA
Duchenne muscular dystrophy (DMD) is an X-linked, lethalmuscular disorder caused by a defect in the DMD gene. AAVvector-mediated micro-dystrophin cDNA transfer is anattractive approach to treatment of DMD. To establish effec-tive gene transfer into skeletal muscle, we examined thetransduction efficiency of an AAV vector in skeletal musclesof dystrophin-deficient mdx mice. When an AAV vectorencoding the LacZ gene driven by a CMV promoter (AAV-CMVLacZ) was introduced, �-galactosidase expressionmarkedly decreased in mdx muscle 4 weeks after injectiondue to immune responses against the transgene product.We also injected AAV-CMVLacZ into skeletal muscles ofmini-dystrophin-transgenic mdx mice (CVBA3’), which showameliorated phenotypes without overt signs of muscle
Keywords: AAV vector; gene transfer; skeletal muscle; mdx mouse; immune response; micro-dystrophin
IntroductionDuchenne muscular dystrophy (DMD) is an X-linked,lethal disorder of skeletal muscle caused by a defect inthe dystrophin gene, which encodes a large sub-sarco-lemmal cytoskeletal protein. The absence of dystrophinaccompanied the loss of dystrophin–glycoprotein com-plex at the sarcolemma and results in progressive muscleweakness, cardiomyopathy and early death. DMD ischaracterized by a high incidence (one among 3500 boys)and a high frequency of de novo mutation1 and currentlyhas no effective treatment.
Several treatment modalities aimed at correcting thedystrophic phenotype have been researched using thedystrophin-deficient mdx mouse and the canine X-linkedmuscular dystrophy (CXMD), and tried on human DMDpatients. These include the adenoviral vector-mediatedintroduction of functional dystrophin,2–6 naked DNA,7
Correspondence: S Takeda, Department of Molecular Therapy, NationalInstitute of Neuroscience, National Center of Neurology and Psychiatry,4-1-1 Ogawa-higashi, Kodaira, Tokyo 187-8502, JapanReceived 22 January 2002; accepted 16 June 2002
degeneration. AAV vector administration, however, evokedsubstantial immune responses in CVBA3’ muscle.Importantly, AAV vector using muscle-specific MCK pro-moter also elicited responses in mdx muscle, but at a con-siderably later period. These results suggested that neo-anti-gens introduced by AAV vectors could evoke immunereactions in mdx muscle, since increased permeabilityallowed a leakage of neo-antigens from the dystrophin-deficient sarcolemma of muscle fibers. However, residentantigen-presenting cells, such as myoblasts, myotubes andregenerating immature myofibers, might also play a role inthe immune response.Gene Therapy (2002) 9, 1576–1588. doi:10.1038/sj.gt.3301829
and primary myoblast8 or stem cell9 transplantation.Alternatively, correction of the endogenous dystrophingene was attempted using chimeric RNA/DNA oligonu-cleotides10,11 or antisense 2’-O-methyl oligoribonucleo-tides.12 The antibiotic gentamicin has been reported toincrease read-through dystrophin protein over a prema-ture stop codon,13 and clinical trials have been conductedrecently.14 On the other hand, compensation for dystro-phin lack by up-regulation of endogenous utrophin15 isanother possible strategy. These innovative strategieshave not been successful because of several difficulties,including host immune response and low efficiency.
Adeno-associated virus (AAV) vector is a potential toolof gene therapy for treatment of genetic neuromusculardisorders. It is a non-pathogenic and replication-defectiveviral vector without any viral open reading frame that isable to infect effectively non-dividing cells, such as thoseof skeletal muscle16–18 and the central nervous system.19
It is widely believed that AAV vectors offer long-termexpression of the transferred gene without immuneresponse against the transferred gene product, and it hasbeen demonstrated that intramuscular injection of AAVvectors expressing immunogenic proteins does not

AAV vector-mediated gene transfer into mdx muscleK Yuasa et al
1577stimulate humoral and cellular immune response totransgene products in immunocompetent mice.16–18,20
Thus, AAV vectors may be applicable to treatment ofinherited neuromuscular disorders such as DMD. In fact,the AAV vector has been successfully introduced into �-sarcoglycan-deficient hamsters (Bio 14.6)21,22 and �-sarco-glycan-deficient mice,23 animal models of limb girdlemuscular dystrophies (LGMD). A recent report, however,showed lower levels of transgene expression togetherwith substantial immune response to the therapeutictransgene product in �-sarcoglycan-null mice. Restrictionof transgene expression to the muscle is important toavoid transgene expression in antigen-presenting cells(APCs).24
Although AAV vector-mediated gene transfer is anattractive option for treatment of DMD, the size of theforeign gene is limited to 4.7–4.9 kb of exogenous DNA.Therefore, full-length dystrophin (14 kb) and mini-dys-trophin (6.4 kb) cDNAs are too large to be incorporatedinto AAV vector as the therapeutic gene.25,26 We pre-viously reported that rod-truncated micro-dystrophin,which is encoded by 3.7 kb cDNA, could effectivelyaccumulate at the sarcolemma and recover dystrophin-associated proteins after adenovirus-mediated genetransfer into mdx skeletal muscles.27 Later, Wang et al28
reported the effective transfer of two kinds of micro-dys-trophin cDNAs (�4.2 kb) into mdx skeletal muscle usingAAV vectors. The AAV vector treatment ameliorateddystrophic pathology in mdx muscle, demonstratingmicro-dystrophin cDNA to be a good candidate forinsertion into the AAV vector.
To establish effective gene transfer into dystrophicskeletal muscle, we examined the transduction efficiencyof the AAV vector in the skeletal muscles of mdx mice,using the �-galactosidase (�-gal) gene as a reporter. AAVvector-mediated gene transfer elicited strong immuneresponses and resulted in poor and short-term expressionof the transferred gene in mdx muscles. Furthermore, anAAV vector elicited immune responses in transgenic mdxmuscle expressing mini-dystrophin, despite ameliorationof dystrophic phenotypes. We also demonstrate thatenhancement of immune responses was associatedclosely with membrane permeability. In addition, use ofmuscle-specific promoter could not prevent the activationof the immune system, although the activation was sig-nificantly delayed. Our results suggest the importance oftransient immunosuppression before the transgeneproduct accumulates in the affected muscle.
Results
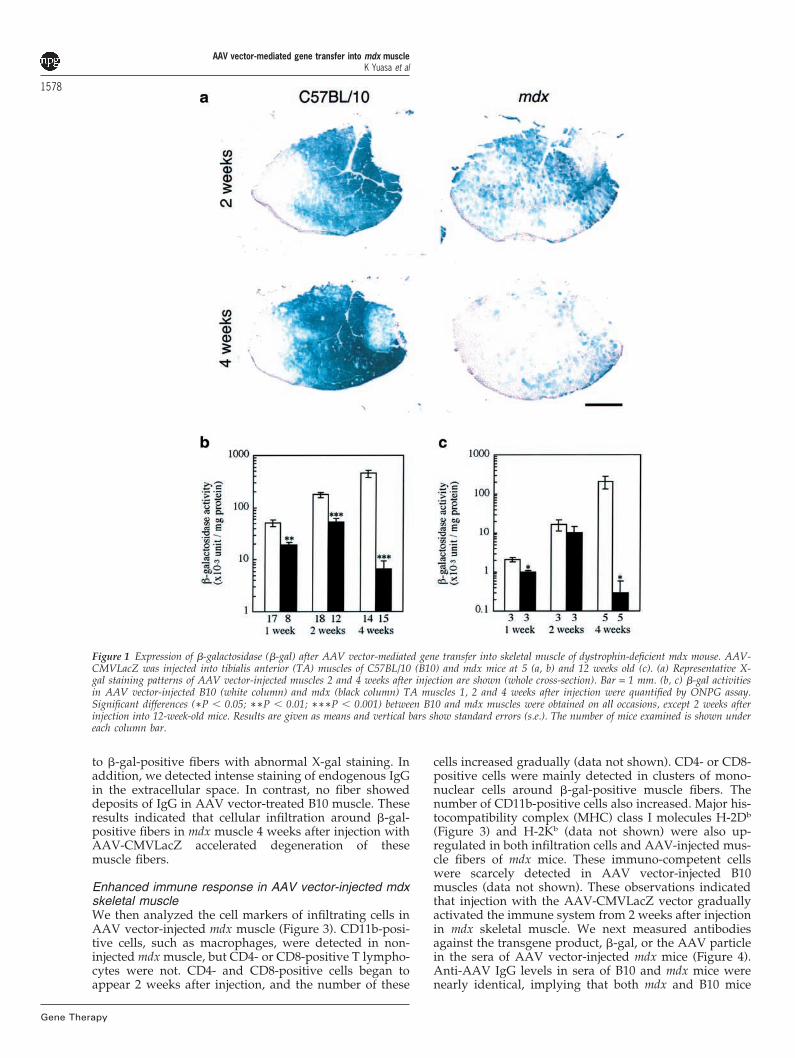
�-Gal expression in mdx skeletal muscle afterintramuscular injection of an AAV vectorTo investigate the transduction efficiency of AAV vector-mediated gene transfer in skeletal muscles of dystrophin-deficient mdx mice, we injected the AAV vector encoding�-galactosidase driven by a ubiquitous CMV promoter(AAV-CMVLacZ) into the tibialis anterior (TA) musclesof 5-week-old C57BL/10 (B10) and mdx mice. One, 2 and4 weeks after injection, the expression of �-galactosidase(�-gal) was evaluated by X-gal staining and ONPG (o-nitrophenyl �-D-galactopyranoside) assay. X-gal stainingshowed that the number of �-gal-positive fibers in AAVvector-injected muscles of control B10 mice increased
Gene Therapy
with time, and high levels of �-gal were expressed 4weeks after injection (Figure 1a). In contrast, �-galactivity and the number of �-gal-positive fibers in AAVvector-injected mdx muscle was much lower than that inB10 mice, and had decreased significantly 4 weeks afterinjection (Figure 1a and b).
Poor and reduced �-gal expression in the late stage inmdx muscle after AAV vector-mediated gene transfermay be due to the elimination of large numbers of �-gal-positive fibers by active muscle degeneration–regener-ation cycles. Muscle fibers of 5-week-old mdx mice gener-ally degenerate at a high rate. To test this hypothesis, weinjected AAV-CMVLacZ into the skeletal muscles of 12-week-old mdx mice, when muscle degeneration hasalmost ceased. Importantly, �-gal expression in musclesinjected at 12 weeks was much lower than that at 5 weeksin both mdx and B10 mice. The expression of transferredgene products showed a similar pattern: it increased for2 weeks after injection and at 2 weeks there was littledifference in �-gal activities between mdx and B10 skel-etal muscles, but it reduced quickly in mdx mice at 4weeks after injection, as seen in 5-week-old mdx mice(Figure 1c). This result supports our hypothesis thatactive degeneration of mdx muscle is largely responsiblefor poor expression of the transgene in the early stage ofthe AAV vector administration. Decreased expression ofthe LacZ gene products was also observed at 4 weeksafter AAV vector administration when introduced into12-week-old mdx muscles. This observation suggests thatthe remarkable reduction of �-gal expression at the laterstage in AAV vector-injected mdx muscle was caused bymechanisms other than the dystrophic process.
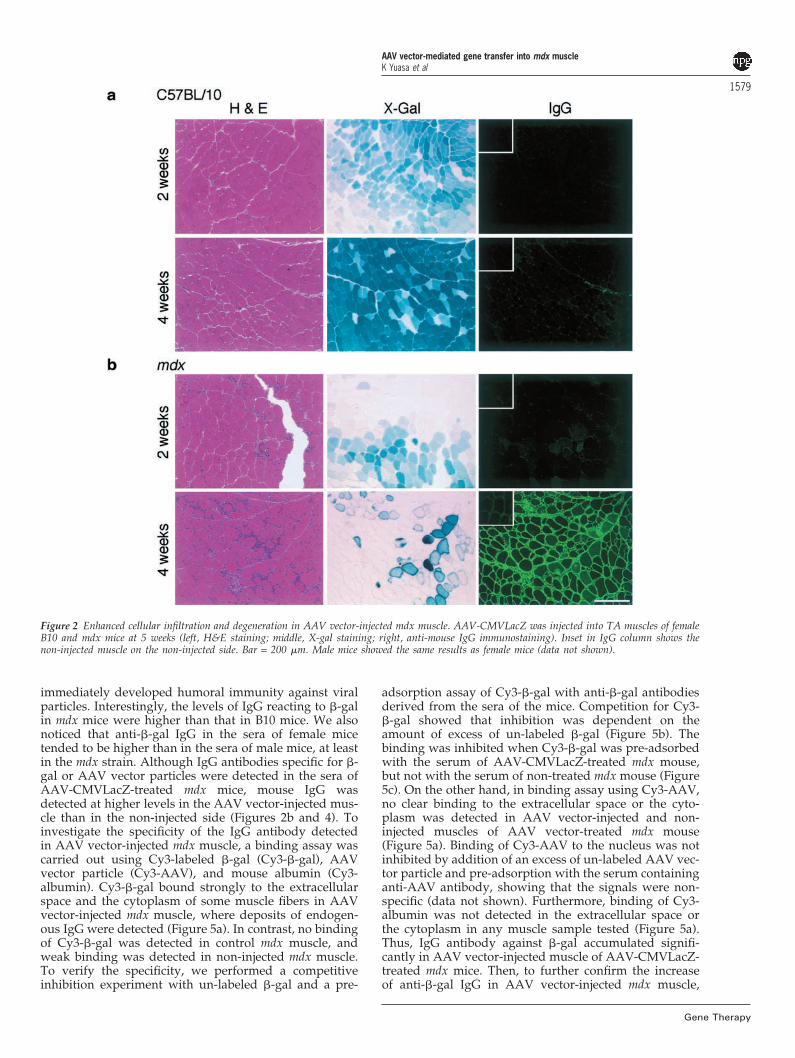
Histology of mdx skeletal muscle after AAV vector-mediated gene transferTo investigate why �-gal expression decreased quickly inmdx muscle 4 weeks after AAV-CMVLacZ injection, wescrutinized the patterns of X-gal staining and histologyof AAV vector-injected skeletal muscles of mdx and B10mice. X-gal staining in AAV vector-injected mdx musclesshowed a mosaic pattern at 4 weeks after injection(Figure 1a): a fiber strongly positive for X-gal stainingoften neighbors totally negative muscle fibers. Next, weanalyzed the histology of the AAV vector-injected mdxmuscle (Figure 2, column H&E and X-Gal). The histologyof skeletal muscles of B10 mice seemed to be normal even4 weeks after injection, as previously reported.16–18,20 InAAV vector-injected mdx muscle, clusters of infiltratingcells were often observed 1 and 2 weeks after injection,but such infiltrates are observed even in the non-injectedmuscle. Four weeks after injection, however, a large num-ber of mononuclear cells appeared around the �-gal-posi-tive fibers, and the cellular infiltration was more apparentin AAV vector-injected mdx muscle than in untreated mdxmuscle. Further, X-gal staining in muscle fibers sur-rounded by mononuclear cells in mdx muscle tended tobe intense both in the cytoplasm and at the sarcolemma(Figure 2b, 4 weeks), suggesting that these fibers were inthe process of degeneration.
We next immunostained AAV vector-injected mdxmuscle using an antibody against mouse IgG (Figure 2,column IgG) because IgG uptake by degenerated fibershas been reported in dystrophic muscles.29 In mdx muscle4 weeks after AAV vector injection, mouse IgG wasdetected weakly within some fibers, which corresponded
AAV vector-mediated gene transfer into mdx muscleK Yuasa et al
1578
Gene Therapy
Figure 1 Expression of �-galactosidase (�-gal) after AAV vector-mediated gene transfer into skeletal muscle of dystrophin-deficient mdx mouse. AAV-CMVLacZ was injected into tibialis anterior (TA) muscles of C57BL/10 (B10) and mdx mice at 5 (a, b) and 12 weeks old (c). (a) Representative X-gal staining patterns of AAV vector-injected muscles 2 and 4 weeks after injection are shown (whole cross-section). Bar = 1 mm. (b, c) �-gal activitiesin AAV vector-injected B10 (white column) and mdx (black column) TA muscles 1, 2 and 4 weeks after injection were quantified by ONPG assay.Significant differences (∗P � 0.05; ∗∗P � 0.01; ∗∗∗P � 0.001) between B10 and mdx muscles were obtained on all occasions, except 2 weeks afterinjection into 12-week-old mice. Results are given as means and vertical bars show standard errors (s.e.). The number of mice examined is shown undereach column bar.
to �-gal-positive fibers with abnormal X-gal staining. Inaddition, we detected intense staining of endogenous IgGin the extracellular space. In contrast, no fiber showeddeposits of IgG in AAV vector-treated B10 muscle. Theseresults indicated that cellular infiltration around �-gal-positive fibers in mdx muscle 4 weeks after injection withAAV-CMVLacZ accelerated degeneration of thesemuscle fibers.
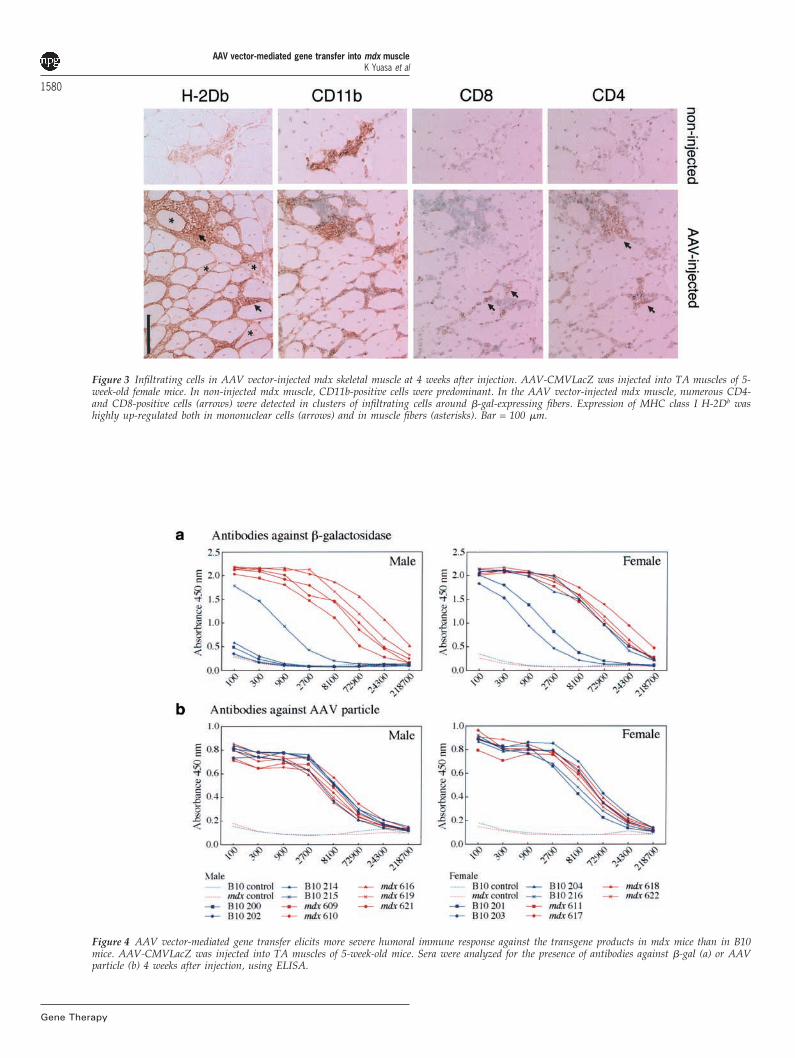
Enhanced immune response in AAV vector-injected mdxskeletal muscleWe then analyzed the cell markers of infiltrating cells inAAV vector-injected mdx muscle (Figure 3). CD11b-posi-tive cells, such as macrophages, were detected in non-injected mdx muscle, but CD4- or CD8-positive T lympho-cytes were not. CD4- and CD8-positive cells began toappear 2 weeks after injection, and the number of these
cells increased gradually (data not shown). CD4- or CD8-positive cells were mainly detected in clusters of mono-nuclear cells around �-gal-positive muscle fibers. Thenumber of CD11b-positive cells also increased. Major his-tocompatibility complex (MHC) class I molecules H-2Db
(Figure 3) and H-2Kb (data not shown) were also up-regulated in both infiltration cells and AAV-injected mus-cle fibers of mdx mice. These immuno-competent cellswere scarcely detected in AAV vector-injected B10muscles (data not shown). These observations indicatedthat injection with the AAV-CMVLacZ vector graduallyactivated the immune system from 2 weeks after injectionin mdx skeletal muscle. We next measured antibodiesagainst the transgene product, �-gal, or the AAV particlein the sera of AAV vector-injected mdx mice (Figure 4).Anti-AAV IgG levels in sera of B10 and mdx mice werenearly identical, implying that both mdx and B10 mice
AAV vector-mediated gene transfer into mdx muscleK Yuasa et al
1579
Figure 2 Enhanced cellular infiltration and degeneration in AAV vector-injected mdx muscle. AAV-CMVLacZ was injected into TA muscles of femaleB10 and mdx mice at 5 weeks (left, H&E staining; middle, X-gal staining; right, anti-mouse IgG immunostaining). Inset in IgG column shows thenon-injected muscle on the non-injected side. Bar = 200 �m. Male mice showed the same results as female mice (data not shown).
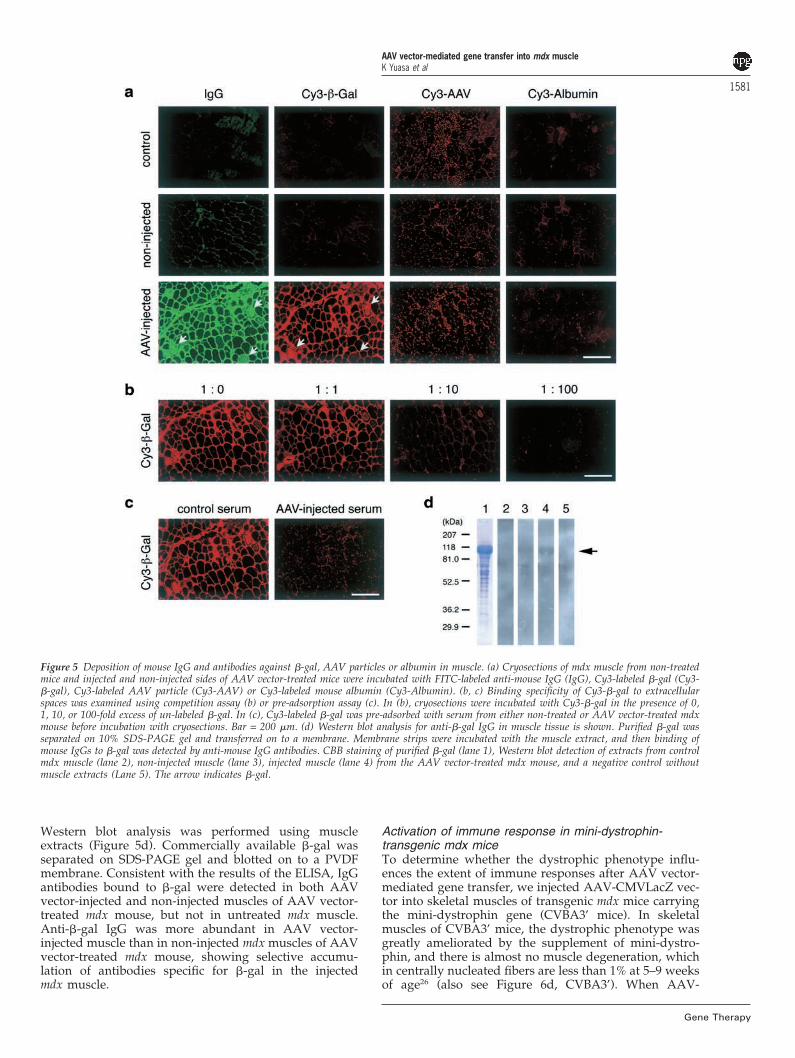
immediately developed humoral immunity against viralparticles. Interestingly, the levels of IgG reacting to �-galin mdx mice were higher than that in B10 mice. We alsonoticed that anti-�-gal IgG in the sera of female micetended to be higher than in the sera of male mice, at leastin the mdx strain. Although IgG antibodies specific for �-gal or AAV vector particles were detected in the sera ofAAV-CMVLacZ-treated mdx mice, mouse IgG wasdetected at higher levels in the AAV vector-injected mus-cle than in the non-injected side (Figures 2b and 4). Toinvestigate the specificity of the IgG antibody detectedin AAV vector-injected mdx muscle, a binding assay wascarried out using Cy3-labeled �-gal (Cy3-�-gal), AAVvector particle (Cy3-AAV), and mouse albumin (Cy3-albumin). Cy3-�-gal bound strongly to the extracellularspace and the cytoplasm of some muscle fibers in AAVvector-injected mdx muscle, where deposits of endogen-ous IgG were detected (Figure 5a). In contrast, no bindingof Cy3-�-gal was detected in control mdx muscle, andweak binding was detected in non-injected mdx muscle.To verify the specificity, we performed a competitiveinhibition experiment with un-labeled �-gal and a pre-
Gene Therapy
adsorption assay of Cy3-�-gal with anti-�-gal antibodiesderived from the sera of the mice. Competition for Cy3-�-gal showed that inhibition was dependent on theamount of excess of un-labeled �-gal (Figure 5b). Thebinding was inhibited when Cy3-�-gal was pre-adsorbedwith the serum of AAV-CMVLacZ-treated mdx mouse,but not with the serum of non-treated mdx mouse (Figure5c). On the other hand, in binding assay using Cy3-AAV,no clear binding to the extracellular space or the cyto-plasm was detected in AAV vector-injected and non-injected muscles of AAV vector-treated mdx mouse(Figure 5a). Binding of Cy3-AAV to the nucleus was notinhibited by addition of an excess of un-labeled AAV vec-tor particle and pre-adsorption with the serum containinganti-AAV antibody, showing that the signals were non-specific (data not shown). Furthermore, binding of Cy3-albumin was not detected in the extracellular space orthe cytoplasm in any muscle sample tested (Figure 5a).Thus, IgG antibody against �-gal accumulated signifi-cantly in AAV vector-injected muscle of AAV-CMVLacZ-treated mdx mice. Then, to further confirm the increaseof anti-�-gal IgG in AAV vector-injected mdx muscle,
AAV vector-mediated gene transfer into mdx muscleK Yuasa et al
1580
Gene Therapy
Figure 3 Infiltrating cells in AAV vector-injected mdx skeletal muscle at 4 weeks after injection. AAV-CMVLacZ was injected into TA muscles of 5-week-old female mice. In non-injected mdx muscle, CD11b-positive cells were predominant. In the AAV vector-injected mdx muscle, numerous CD4-and CD8-positive cells (arrows) were detected in clusters of infiltrating cells around �-gal-expressing fibers. Expression of MHC class I H-2Db washighly up-regulated both in mononuclear cells (arrows) and in muscle fibers (asterisks). Bar = 100 �m.
Figure 4 AAV vector-mediated gene transfer elicits more severe humoral immune response against the transgene products in mdx mice than in B10mice. AAV-CMVLacZ was injected into TA muscles of 5-week-old mice. Sera were analyzed for the presence of antibodies against �-gal (a) or AAVparticle (b) 4 weeks after injection, using ELISA.
AAV vector-mediated gene transfer into mdx muscleK Yuasa et al
1581
Figure 5 Deposition of mouse IgG and antibodies against �-gal, AAV particles or albumin in muscle. (a) Cryosections of mdx muscle from non-treatedmice and injected and non-injected sides of AAV vector-treated mice were incubated with FITC-labeled anti-mouse IgG (IgG), Cy3-labeled �-gal (Cy3-�-gal), Cy3-labeled AAV particle (Cy3-AAV) or Cy3-labeled mouse albumin (Cy3-Albumin). (b, c) Binding specificity of Cy3-�-gal to extracellularspaces was examined using competition assay (b) or pre-adsorption assay (c). In (b), cryosections were incubated with Cy3-�-gal in the presence of 0,1, 10, or 100-fold excess of un-labeled �-gal. In (c), Cy3-labeled �-gal was pre-adsorbed with serum from either non-treated or AAV vector-treated mdxmouse before incubation with cryosections. Bar = 200 �m. (d) Western blot analysis for anti-�-gal IgG in muscle tissue is shown. Purified �-gal wasseparated on 10% SDS-PAGE gel and transferred on to a membrane. Membrane strips were incubated with the muscle extract, and then binding ofmouse IgGs to �-gal was detected by anti-mouse IgG antibodies. CBB staining of purified �-gal (lane 1), Western blot detection of extracts from controlmdx muscle (lane 2), non-injected muscle (lane 3), injected muscle (lane 4) from the AAV vector-treated mdx mouse, and a negative control withoutmuscle extracts (Lane 5). The arrow indicates �-gal.
Western blot analysis was performed using muscleextracts (Figure 5d). Commercially available �-gal wasseparated on SDS-PAGE gel and blotted on to a PVDFmembrane. Consistent with the results of the ELISA, IgGantibodies bound to �-gal were detected in both AAVvector-injected and non-injected muscles of AAV vector-treated mdx mouse, but not in untreated mdx muscle.Anti-�-gal IgG was more abundant in AAV vector-injected muscle than in non-injected mdx muscles of AAVvector-treated mdx mouse, showing selective accumu-lation of antibodies specific for �-gal in the injectedmdx muscle.
Gene Therapy
Activation of immune response in mini-dystrophin-transgenic mdx miceTo determine whether the dystrophic phenotype influ-ences the extent of immune responses after AAV vector-mediated gene transfer, we injected AAV-CMVLacZ vec-tor into skeletal muscles of transgenic mdx mice carryingthe mini-dystrophin gene (CVBA3’ mice). In skeletalmuscles of CVBA3’ mice, the dystrophic phenotype wasgreatly ameliorated by the supplement of mini-dystro-phin, and there is almost no muscle degeneration, whichin centrally nucleated fibers are less than 1% at 5–9 weeksof age26 (also see Figure 6d, CVBA3’). When AAV-
AAV vector-mediated gene transfer into mdx muscleK Yuasa et al
1582
Gene Therapy
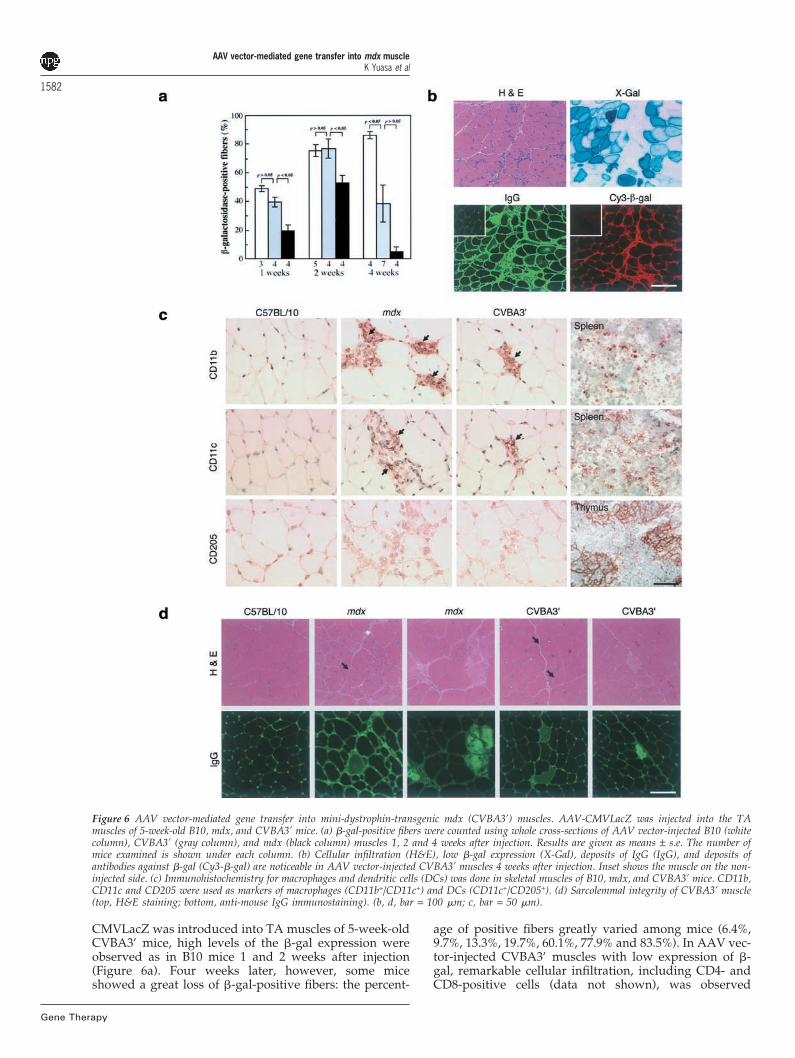
Figure 6 AAV vector-mediated gene transfer into mini-dystrophin-transgenic mdx (CVBA3’) muscles. AAV-CMVLacZ was injected into the TAmuscles of 5-week-old B10, mdx, and CVBA3’ mice. (a) �-gal-positive fibers were counted using whole cross-sections of AAV vector-injected B10 (whitecolumn), CVBA3’ (gray column), and mdx (black column) muscles 1, 2 and 4 weeks after injection. Results are given as means ± s.e. The number ofmice examined is shown under each column. (b) Cellular infiltration (H&E), low �-gal expression (X-Gal), deposits of IgG (IgG), and deposits ofantibodies against �-gal (Cy3-�-gal) are noticeable in AAV vector-injected CVBA3’ muscles 4 weeks after injection. Inset shows the muscle on the non-injected side. (c) Immunohistochemistry for macrophages and dendritic cells (DCs) was done in skeletal muscles of B10, mdx, and CVBA3’ mice. CD11b,CD11c and CD205 were used as markers of macrophages (CD11b+/CD11c+) and DCs (CD11c+/CD205+). (d) Sarcolemmal integrity of CVBA3’ muscle(top, H&E staining; bottom, anti-mouse IgG immunostaining). (b, d, bar = 100 �m; c, bar = 50 �m).
CMVLacZ was introduced into TA muscles of 5-week-oldCVBA3’ mice, high levels of the �-gal expression wereobserved as in B10 mice 1 and 2 weeks after injection(Figure 6a). Four weeks later, however, some miceshowed a great loss of �-gal-positive fibers: the percent-
age of positive fibers greatly varied among mice (6.4%,9.7%, 13.3%, 19.7%, 60.1%, 77.9% and 83.5%). In AAV vec-tor-injected CVBA3’ muscles with low expression of �-gal, remarkable cellular infiltration, including CD4- andCD8-positive cells (data not shown), was observed

AAV vector-mediated gene transfer into mdx muscleK Yuasa et al
1583around the �-gal-positive fibers, which were stainedabnormally with X-gal. Abundant IgG antibodies against�-gal were also detected in the extracellular space andwithin some �-gal-positive fibers (Figure 6b). In contrast,no or little cellular infiltration and negligible anti-�-galantibodies were detected in AAV vector-injected CVBA3’muscles with high �-gal expression (data not shown).Thus, AAV vector elicited immune responses against thetransgene product, even in the nearly normal muscles ofCVBA3’ mice.
It has been reported that immature dendritic cells(DCs) infected with AAV vector play an important rolein activation of cytotoxic T lymphocytes (CTLs), leadingto elimination of transduced cells.30 To assess the possi-bility that antigen-presenting cells (APCs) are involvedin activation of immune responses against the transgeneproduct, we detected macrophages (CD11b+/CD11c+) orDCs (CD11c+/CD205+) by immunohistochemistry tocharacterize the infiltrates in native skeletal muscles ofmdx and CVBA3’ mice (Figure 6c). A large number ofCD11b- and CD11c-positive cells were detected withinand around degenerating fibers in mdx muscle, butCD11b- and CD11c-positive cells were much fewer inCVBA3’ muscle. In addition, strong staining for CD205was detected in the T cell areas of the spleen and corticalepithelium of the thymus,31 but not in skeletal muscles ofmdx and CVBA3’ mice. These observations indicate thatCVBA3’ muscle is devoid of DCs at the time of the AAVvector injection.
Dystrophin forms a complex with dystrophin-associa-ted proteins (DAPs) linking the cytoskeleton to the extra-cellular matrix at the inner surface of the sarcolemma toprotect the membrane from stress. The absence of dystro-phin increases membrane permeability.26 One activator ofthe immune response in mdx and CVBA3’ mice, but notin wild-type B10 mice, might be the release of cellularproteins into the extracellular space, so we investigatedthe sarcolemmal integrity of mdx and CVBA3’ musclesusing anti-mouse IgG staining. Depositions of IgG withinfibers were observed in apparently necrotic fibers withphagocytosis in mdx and CVBA3’ muscles (Figure 6d).Furthermore, slight IgG uptake into some fibers wasdetected in both mdx and CVBA3’ muscles although thesemuscle fibers often lack signs of degeneration on H&E-stained sections (Figure 6d). These observations indicatethat membrane integrity of CVBA3’ muscle is not as com-plete as that of normal B10 muscle, supporting ourhypothesis that release of cellular proteins might stimu-late the humoral and cellular immune responses againstthe transgene products.
Effects of immunosuppression of AAV vector-injectedmdx mice using anti-CD4 antibodyTo test whether immunosuppression prolongs theexpression of the transgene in AAV vector-mediatedgene transfer into mdx skeletal muscle, we administeredantibodies against CD4 (GK1.5) intraperitoneally to 5-week-old mdx mice on day �2, �1, 0 and +1 of AAV-CMVLacZ vector injection and then weekly until theywere killed. �-Gal activities in anti-CD4 antibody-treatedmdx muscles remained high 4 weeks after injection(Figure 7a). Furthermore, anti-CD4 treatment inhibitedboth deposition of IgG in the extracellular space anduptake into muscle fibers in AAV vector-injected mdxmuscle (Figure 7b). These results indicate that immune
Gene Therapy
response markedly reduced transgene expression in AAVvector-mediated gene transfer into mdx skeletal muscle.
Skeletal muscle in 5-week-old mdx mouse shows vigor-ous degeneration and regeneration. This might lead topoor expression of the transgene in AAV-injected mdxmuscle because AAV-transduced cells can be replaced byregenerating fibers at a high rate. We also noticed thatanti-CD4-treated mdx muscles showed milder dystrophicchanges than non-treated age-matched mdx mice (datanot shown).
Gene transfer with muscle-specific MCK promoter intomdx skeletal muscleTo clarify the mechanism of humoral and cellularimmune responses in mdx mice, we next investigatedwhether �-gal expression driven by a muscle-specificMCK promoter could suppress the immune response ornot. The depressed expression of highly immunogenic �-gal in potent antigen-presenting cells, such as dendriticcells or macrophages, might greatly attenuate theimmune response. Expression of �-gal continued toincrease at least for 4 weeks in AAV-MCKLacZ-injectedmdx muscles (Figure 8a), but it significantly decreased 8weeks after injection. Four weeks after injection, cellularinfiltration and deposition of IgG were not detected inAAV-MCKLacZ-injected mdx muscle (Figure 8b),whereas it was obviously detected in AAV-CMVLacZ-injected mdx muscles.
DiscussionThe AAV vector is a powerful tool to introduce thera-peutic genes into non-dividing cells, such as skeletalmuscle fibers. It evokes little immune reaction andenables long-term expression of the transferred genes,although full-length dystrophin cDNA is too large to beaccommodated in an AAV vector. Several micro-dystro-phins, in which the rod domain is largely deleted, intro-duced into mdx mice,27 were successfully expressed at thesarcolemma together with the recovery of the dystro-phin–glycoprotein complex that protects muscle fibersfrom degeneration.28 In order to establish an AAV vector-mediated gene therapy for DMD, we investigated theeffect of AAV vector-mediated gene transfer into dys-trophic skeletal muscles from mdx mice.
When an AAV vector (AAV-CMVLacZ) encoding theLacZ gene driven by CMV promoter was introduced intoskeletal muscle, earlier transduction efficiency was sig-nificantly lower in mdx muscle than in control C57BL/10muscle. Poor transduction efficiency in mdx muscle in theearly stage after AAV vector injection could be due toelimination of AAV vector-transduced cells by the spon-taneous degeneration–regeneration cycles that arecharacteristic of young mdx mice. Interestingly, continu-ous immunosuppression by anti-CD4 antibody in mdxmice resulted in efficient transduction of the transferredgene 2 weeks, as well as 4 weeks after injection of theAAV vector (Figure 7). This result leads us to the hypoth-esis that immunosuppression could limit the degener-ation by inhibiting immunocompetent cells that attackdystrophic muscle fibers. In fact, prednisone therapy hasbeen confirmed to be of value in enhancing musclestrength and function in DMD.32 The anti-inflammatoryand immunosuppressive effect of the steroid suppressesthe progression of muscular dystrophy. In this respect,

AAV vector-mediated gene transfer into mdx muscleK Yuasa et al
1584
Gene Therapy
Figure 7 Effect of immunosuppression with anti-CD4 antibody on �-gal expression in AAV vector-injected mdx muscle. AAV-CMVLacZ was injectedinto TA muscles of 5-week-old mdx mice and a monoclonal anti-CD4 antibody (GK1.5) was administered as described in Materials and methods. (a)�-gal activity in AAV-CMVLacZ-injected muscles of B10 mice (white column), anti-CD4-treated mdx mice (gray column) and non-treated mdx mice(black column) at 2 and 4 weeks after injection. Results are given as means ± s.e. The number of mice examined is shown under each column. (b) �-gal expression and humoral immune response in anti-CD4-treated and non-treated mdx muscles 4 weeks after injection. (top, X-gal staining, bar = 1mm; bottom, anti-mouse IgG immunostaining, bar = 200 �m). Inset shows the muscle on the non-injected side.
transient immunosuppression could be useful not onlyfor efficient transduction of the transgene in AAV vector-mediated gene transfer into dystrophic skeletal muscle,but also for prevention of progressive muscular degener-ation itself.
The minimal immune response that the AAV vectorelicits in the host enables long-term expression of thetransferred gene. We observed stable long-termexpression of the LacZ gene products in normal B10 mus-cle. These muscles showed almost no signs of immunereactions against �-gal. However, enhanced immuneresponses were observed in mdx skeletal muscles afterAAV-CMVLacZ-mediated gene transfer. We detected alarge number of CD4- or CD8-positive T lymphocytesand up-regulation of MHC class I in mononuclear cellssurrounding �-gal-expressing muscle fibers 4 weeks afterinjection. At this stage, �-gal expression remarkablydecreased, and the remaining �-gal-positive muscle fibersshowed signs of ongoing degeneration, implying thatantigen-specific MHC class I-restricted CTLs play a majorrole in elimination of �-gal-expressing muscle fibers. Inaddition, we detected high levels of antibodies specificfor �-gal in the sera of AAV vector-treated mdx mice anddense deposits of IgG including antibodies specific to �-gal in muscle tissues. The results of immunosupressionin later stages by anti-CD4 antibody treatment also sup-port the hypothesis that immune response has been con-cerned in the AAV-vector mediated gene transfer intomdx skeletal muscle. Several groups have previouslydemonstrated effective AAV vector-mediated gene trans-fer into skeletal muscles of mdx mice,28 �-sarcoglycan-deficient hamsters (Bio 14.6)21,22 and �-sarcoglycan-deficient mice.23 On the other hand, Cordier et al24 haveshown that an AAV vector driven by CMV promoter elic-its strong cellular and humoral immune responses to thetransgene product after intramuscular injection into �-sarcoglycan-null mice.
Infection of potent antigen-presenting cells (APCs),such as immature dendritic cells, by AAV vector with aubiquitous promoter results in production of neo-anti-gens in APCs. Infected APCs would in turn activate CTLsto a great extent and eliminate the transduced muscle
cells.24,30 Indeed, the number of DCs and macrophagesdramatically increased in degenerating and regeneratingskeletal muscle after artificial crush injury.33 It has beenalso reported that mdx muscles contained 20 times moremacrophage and seven times more DCs than normalmuscles.34 We could not detect CD205-positive dendriticcells in mdx muscle, but the decrease in �-gal expressionwas considerably delayed, when AAV-MCKLacZ wasintroduced into mdx muscle (Figure 8). This result indi-cates that APCs play a critical role in developing immun-ity against the AAV vector infection. Importantly,enhanced immune responses were observed in laterstages of AAV-MCKLacZ-injected mdx muscles, as wellas AAV-CMVLacZ-injected muscles. This finding couldbe explained by neo-antigens that were continuouslyreleased from dystrophin-deficient myofibers due tospontaneous degeneration. However, there is anotherpossibility that myotubes and immature myofiberspresented �-gal as a neo-antigen to immuno-competentcells. Indeed, myogenic cell, such as myoblast, myotubesand immature myofibers have been reported to functionas local APCs.35 Therefore, myoblasts and regeneratingmyofibers in dystrophic skeletal muscle might also con-siderably enhance the immune responses in mdx muscle.
CVBA3’ is a transgenic mdx mouse line carrying themini-dystrophin gene isolated from a mild Becker MDpatient that presents very mild symptoms of musculardystrophy.25,26 In fact, CVBA3’ skeletal muscle showedonly a few degenerating fibers and infiltrating cells.When the AAV-CMVLacZ vector was introduced intoCVBA3’ mice at the age of 5 weeks, the expression levelof the transferred gene products was as high as in normalmice until 2 weeks after injection, without overt sign ofimmune responses. We noticed, however, a reduction of�-gal expression 4 weeks after AAV-CMVLacZ injection,together with the enhanced cellular infiltration and exten-sive deposits of antibodies specific for the transgene pro-duct. Why were the immune responses activated inCVBA3’ muscle in spite of their ameliorated phenotype?Cordier et al24 suggested that a minimum number of DCswould be required to capture the AAV vector and/or itstransduced gene products in order to elicit T cell immun-

AAV vector-mediated gene transfer into mdx muscleK Yuasa et al
1585
Figure 8 �-gal expression in mdx skeletal muscle after injection with an AAV vector driven by muscle-specific MCK promoter. (a) �-gal activity inTA muscles of mdx mice at 1, 2, 4 and 8 weeks after injection with AAV-CMVLacZ (white column) and AAV-MCKLacZ (black column). The micewere injected with AAV vectors at 5 weeks of age. Results are given as means ± s.e. The number of mice examined is shown under each column. (b)Cellular infiltration (H&E), �-gal expression (X-Gal) and deposits of IgG (IgG) in AAV-MCKLacZ-injected mdx muscles and AAV-CMVLacZ-injectedmdx muscles 4 weeks or 8 weeks after injection. Inset shows the muscle on the non-injected side. Bar = 100 �m.
ity. Therefore, the number of inflammatory cells presentat the time of the AAV injection would be critical in mob-ilizing sufficient APCs.24,30 We, however, observed verylittle infiltration of macrophages in CVBA3’ muscle, andimmunostaining with CD205 detected no potent DCs. Inaddition, proliferating myoblasts and regeneratingmyofibers were rarely observed in CVBA3’ muscles.These observations suggested that immune responses inCVBA3’ muscle would not be directly related to thetransgene expression in resident APCs including myog-enic cells. A remaining possibility is that impaired mem-brane integrity of CVBA3’ led to immune reaction againstAAV vector-infected muscle fibers. We saw some fiberswith high membrane permeability in CVBA3’ muscle,although membrane integrity of the muscle was greatlyimproved over the original mdx muscle. Intracellular pro-teins could be leaking from dystrophic fibers into theextracellular space, as indicated by the increased serum
Gene Therapy
creatine kinase level in mdx and CVBA3’ mice.26 Thetransgene product released from AAV vector-transducedmuscle cells into circulation might be trapped by APCsand stimulate both cellular and humoral immune sys-tems. In mdx muscles, neo-antigens on APCs, in parti-cular immature myofibers themselves, might stimulateCD4+ helper or CD8+ cytotoxic T cells. We need to furtherclarify the molecular mechanisms which underlie theunexpected immune response observed in CVBA3’muscle.
We clearly demonstrated that immune suppressionimproved the efficiency of the gene expression after AAVvector-mediated gene transfer, and our results suggestthe importance of transient immunosuppression beforethe transgene product accumulates in the affected mus-cle. Nevertheless, we should independently consider howto minimize immune response for future approaches toDMD treatment. First of all, a muscle-specific promoter

AAV vector-mediated gene transfer into mdx muscleK Yuasa et al
1586
Gene Therapy
should be used to suppress the transgene expression inAAV vector-transduced APCs, as suggested by this paperand others.24,34 Further, a fully functional micro-dystro-phin would effectively protect muscle fibers fromdegeneration and reduce the release of neo-antigens orself-antigens into the extracellular space. A truncatedmicro-utrophin may be an alternative molecule for genetherapy of DMD. Utrophin has been shown to amelioratedystrophic phenotypes when expressed exogenously15 orendogenously36 in dystrophin-deficient muscle. We mayavoid undesirable activation of immune response by useof micro-utrophin instead of micro-dystrophin becausethe utrophin gene is intact in DMD patients.
Materials and methods
Production of AAV vectorsWe used the AAV type2 vector named AAV-CMVLacZ37
and AAV-MCKLacZ, which contain the LacZ gene underthe control of the cytomegalovirus (CMV) promoter andthe truncated muscle creatine kinase (MCK) promoter,respectively. The truncated MCK promoter was shownto drive muscle-specific expression of the reporter gene38
and constructed by PCR from the mouse genomic DNA.Briefly, 206 bp of the enhancer region (from �1256 to�1050 relative to the transcriptional start site, AccessionNo. M21390) and 358 bp of the proximal regulatoryregion (from �358 to +7) were amplified separately byPCR reactions, subcloned into pCR2.1 (Invitrogen, Gron-ingen, The Netherlands), sequenced and connecteddirectly, based on the report described.38 The recombi-nant viral vector stocks were produced by cotransfectionmethods as already described.39 The AAV virions werepurified twice by CsCl density gradient centrifugation,dialyzed against sterile phosphate-buffered saline (PBS)and titrated by a quantitative DNA dot blot assay,according to the previously published protocol.16,39 Titerswere routinely in the range of 1 × 1013 to 2 × 1013 viralgenomes per ml.
Administration of AAV vectors to murine skeletalmusclesNormal C57BL/10 (B10), C57BL/10 mdx and mini-dys-trophin-transgenic mdx (CVBA3’)26 mice were injectedwith AAV-CMVLacZ or AAV-MCKLacZ vectors (5–8 ×1011 viral genomes) in 50 �l of PBS into the right tibialisanterior (TA) muscles at 5 or 12 weeks of age. To sup-press the immune response, we treated other AAV-CMVLacZ-injected mdx mice with rat monoclonal anti-body against mouse CD4 (GK1.5). The anti-CD4 antibodywas administered intraperitoneally at a dose of 100 �gper animal on days �2, �1, 0 and +1 at AAV-CMVLacZinjection, and then weekly until they were killed.
Histological analysisThe AAV vector-injected mdx and normal B10 mice werekilled by cervical dislocation 1, 2, 4 and 8 weeks afterthe AAV vector injection. The AAV vector-injected andcontralateral non-injected TA muscles were isolated andfrozen in liquid nitrogen-cooled isopentane. Cryosections(10 �m) from a third of the muscles were stained withhematoxylin/eosin (H&E). They were also stained with5-bromo-4-chloro-3-indolyl-�-D-galactopyranoside (X-gal; Wako Pure Chemical Industries, Osaka, Japan), as
described elsewhere.40 Photographs were taken with aDAS Mikroskop LEITZ DMRB (Leica, Wetzlar,Germany), using a digital still camera system HC-2500(Fujifilm, Tokyo, Japan). We counted the number of �-galactosidase (�-gal)-positive fibers in whole cross-sections of the AAV vector-injected TA muscles of atleast three mice at each time point.
Measurement of �-galactosidase activityA cell extract was prepared from frozen muscle and usedto measure �-gal activity by ONPG assay, as describedelsewhere,40 with some modifications. Approximately 40cryosections (each 10 �m thick) were suspended in 100�l of sample buffer (250 mM Tris-HCl (pH 7.4), 2 mMMgCl2, 0.1 % Triton X-100) and then subjected to threecycles of freezing and thawing. After centrifugation at18 000 g for 10 min, the supernatant was collected as thecell extract. For normalization of activity, we estimatedthe protein concentrations using the Bradford CoomassieBrilliant Blue binding assay (BioRad, Hercules, CA,USA). The cell extract was assayed in 1.0 ml of reactionbuffer (60 mM Na2HPO4, 40 mM NaH2PO4, 10 mM KCl,1 mM MgCl2) containing 0.8 mg/ml ONPG (Sigma, StLouis, MO, USA) and 4 �l/ml �-mercaptoethanol. Thesolutions were then incubated at 37°C until they turnedyellow, the reactions were halted by adding 0.5 ml of 1M Na2CO3, and their optical densities were measured at420 nm. The results were expressed as �-gal units, incomparison with a standard enzymatic plot obtainedfrom purified E. coli �-galactosidase (Roche Diagnostic,Mannheim, Germany).
ImmunohistochemistryFor immunohistochemical study, we prepared transversecryosections (6 �m) from AAV vector-injected and non-injected TA muscles. Sections from each muscle wereplaced on the same slide, then dried and fixed in acetonefor 10 minutes at �20°C. We used the following anti-bodies: biotin-conjugated rat monoclonal antibodiesagainst mouse CD4 (L3T4; clone H129.19, PharMingen,San Diego, CA, USA) or against mouse CD8a (Ly-2; clone53-6.7, PharMingen); rat monoclonal antibodies againstmouse CD11b (Mac-1, Cedarlane, Ontario, Canada) andagainst mouse CD205 (DEC-205; clone NLDC-145, Sero-tec, Raleigh, NC, USA); hamster monoclonal antibodyagainst mouse CD11c (clone N418), biotin-conjugatedmouse monoclonal antibodies against mouse major histo-compatibility complex (MHC) class I alloantigens H-2Db
(clone 28-14-8, PharMingen) or H-2Kb (clone AF6-88.5,PharMingen); a fluorescein-conjugated F(ab’)2 fragmentof goat antibody against mouse IgG (H&L) (Leinco Tech-nologies, St Louis, MO, USA). The primary antibodiesagainst CD11b, CD11c and CD205 were detected usingbiotinylated rabbit anti-rat immunoglobulins (Dako,Glostrup, Denmark) or biotinylated goat anti-hamsterIgG (Cedarlane). The biotin molecules were detectedusing the avidin-biotinylated peroxidase supplied in theVECTASTAIN ABC kit (Vector Laboratories, Burlingame,CA, USA). The immune complex labeled with horserad-ish peroxidase was visualized with diaminobenzidine(DAB) (Vector Laboratories), then counterstained withmethyl green, and photographed with a DAS MikroskopLeitz DMRB using a digital still camera system HC-2500.The signals from fluorescein-conjugated antibody againstmouse IgG were recorded photographically under the

AAV vector-mediated gene transfer into mdx muscleK Yuasa et al
1587same conditions using the confocal laser scanning micro-scope system FLUOVIEW (Olympus Optical, Tokyo,Japan).
Measurement of humoral immune responseThe titer of the antibody to the transgene product (�-gal)or to the AAV vector particle was measured by ELISA.A SUMILON ELISA plate H (Sumitomo Bakelite, Tokyo,Japan) was coated overnight at 4°C with �-gal (0.2�g/well; Roche Diagnostic) or the AAV vector particle (2× 109 genomes/well) in 100 �l of PBS. The microtiterplate was washed and blocked overnight at 4°C withblocking buffer (Block Ace; Dainippon Pharmaceutical,Osaka, Japan). After washing, serial dilutions of mouseserum were applied to the wells and incubated 2 h atroom temperature. Reactivity to �-gal and the AAV vec-tor particle was determined by incubation with a 1/5000dilution of horseradish peroxidase-conjugated goat anti-mouse IgG (H+L) (BioRad, Hercules, CA, USA) for 1 hat room temperature, followed by an extensive wash withPBS-T (0.05% Tween 20 in PBS). The Peroxidase SubstrateSystem T (Sumitomo Bakelite) consisting of 3, 3’, 5, 5’-tetramethylbenzidine and H2O2 was used for the colorreaction and absorbance was read at 450 nm.
Binding assay using the fluorescence-conjugated proteinOne milligram of �-gal (Roche), 0.11 mg of recombinantAAV-CMVLacZ viron (5.2 × 1012 genomes), and 1 mg ofmouse albumin (Sigma) were labeled with Cy3 fluor-escence dye, using a FluoroLink-Ab Cy3 labeling kit(Amersham Pharmacia Biotech, Buckinghamshire, UK)according to the supplemental protocol. After the labe-ling reaction, approximately 1 ml of the labeled �-gal andalbumin was separated from non-conjugated dye by thesupplied column, and stored in PBS containing 0.1%sodium azide at �80°C. One milliliter of the labeled AAVvector viron was separated from non-conjugated dye byextensive dialysis against PBS, and stored at �80°C. Todetect the antibody against each antigen, the muscle sec-tion was incubated with Cy3-labeled proteins. Briefly,transverse cryosections (6 �m) from AAV vector-injectedand non-injected TA muscles were fixed for 2 h at 4°C in4% paraformaldehyde. The sections were blocked with5% goat serum in PBS, and then incubated with Cy3-lab-eled protein (1/200 dilution for �-gal and AAV vector,1/80 for albumin) for 1 h at room temperature. For pre-adsorption of Cy3-labeled protein, a dilution was incu-bated for 1 h with an equal volume of serum from eithera non-injected or an AAV vector-injected mdx mouse (4weeks after injection), before reaction with the cryosec-tions. After extensive washing, binding of Cy3-labeledprotein on muscle sections was detected by a confocallaser scanning microscope system FLUOVIEW(Olympus).
Western blot analysis for detection of antibodies inmuscle tissueFor detection of anti-�-gal IgG antibody in the musclesof non-injected and AAV vector-injected mice, an alterna-tive Western blot analysis was carried out, as describedelsewhere18 with some modifications. To prepare muscleextracts, 100 cryosections (each 10 �m thick) of non-injected or AAV vector-injected muscle were suspendedin 200 �l of PBS with 0.1% Triton X-100 and subjected tothree cycles of freezing and thawing. After centrifugation,
Gene Therapy
the supernatant was removed as the muscle extract andthe protein concentrations estimated. Aliquots (5�g/lane) of purified �-gal protein (Roche) were separ-ated on 10% SDS-PAGE and electrotransferred on to aPVDF membrane (Immobilon; Millipore, New Bedford,MA, USA). Individual lanes were cut and blocked for 2h at room temperature with Block Ace (DainipponPharmaceutical). The blots were then incubated over-night at 4°C in 30 �g/ml of the muscle extracts. IgG anti-bodies bound to �-gal were detected by horseradish per-oxidase-conjugated goat anti-mouse IgG (1/2000, Bio-Rad) and ECL Western blotting detection reagents(Amersham Pharmacia Biotech, Uppsala, Sweden). Theblots were also stained with Coomassie Brilliant BlueR-250.
Statistical analysisIn this report, data are expressed as means ± s.e.m. Stud-ent’s t tests were used to evaluate the difference betweentwo groups.
AcknowledgementsWe are grateful to Dr M Yoshida for providing mdx mice.We also thank colleagues in our laboratory for useful dis-cussion and suggestions on this work. This work is sup-ported by Grants-in-Aid for Scientific Research for Centerof Excellence, Research on Nervous and Mental Dis-orders (10B-1, 13B-1) Health Sciences Research Grants forResearch on the Human Genome and Gene Therapy(H10-genome-015, H13-genome-001), for Research onBrain Science (H12-Brain-028) from the Ministry ofHeath, Labor and Welfare, Grant-in-Aids for ScientificResearch (10557065, 11470153, 11170264 and 14657158)from the Ministry of Education, Culture, Sports, Scienceand Technology, and a Research Grant from HumanFrontier Science Project.
References1 Emery AEH. Duchenne Muscular Dystrophy 2nd edn. Oxford
University Press: Oxford, 1993.2 Rogot T et al. Efficient adenovirus-mediated transfer of a human
minidystrophin gene to skeletal muscle of mdx mice. Nature1993; 361: 647–650.
3 Acsadi G et al. Dystrophin expression in muscles of mdx miceafter adenovirus-mediated in vivo gene transfer. Hum Gene Ther1996; 7: 129–140.
4 Kochanek S et al. A new adenoviral vector: replacement of allviral coding sequences with 28 kb of DNA independentlyexpressing both full-length dystrophin and �-galactosidase. ProcNatl Acad Sci USA 1996; 93: 5731–5736.
5 Tuner G, Duncley MG, Dickson G. Gene therapy of Duchennemuscular dystrophy. In: Brown SC, Lucy JA (eds). Dystrophin:Gene, Protein and Cell Biology. Cambridge University Press: Cam-bridge, 1997, pp 274–309.
6 Chen H-H et al. DNA from both high-capacity and first gener-ation adenoviral vectors remains intact in skeletal muscle. HumGene Ther 1999; 10: 365–373.
7 Acsadi G et al. Human dystrophin expression in mdx mice afterintramuscular injection of DNA constructs. Nature 1991; 352:815–818.
8 Gussoni E, Blau HM, Kunkel LM. The fate of individual myo-blasts after transplantation into muscles of DMD patients. Nat-ure Med 1997; 3: 970–977.
9 Gussoni E et al. Dystrophin expression in mdx mouse restoredby stem cell transplantation. Nature 1999; 40: 390–394.

AAV vector-mediated gene transfer into mdx muscleK Yuasa et al
1588
Gene Therapy
10 Rando TA, Disatnik MH, Zhou LZ. Rescue of dystrophinexpression in mdx mouse muscle by RNA/DNA oligonucleot-ides. Proc Natl Acad Sci USA 2000; 97: 5363–5368.
11 Bartlett RJ et al. In vivo targeted repair of a point mutation inthe canine dystrophin gene by a chimeric RNA/DNA oligonu-cleotide. Nat Biotechnol 2000; 18: 615–622.
12 Dunckley MG et al. Modification of splicing in dystrophin genein cultured Mdx muscle cells by antisense oligonucleotides. HumMol Genet 1998; 7: 1083–1090.
13 Barton-Davis ER et al. Aminoglycoside antibiotics restore dys-trophin function to skeletal muscles of mdx mice. J Clin Invest1999; 104: 375–381.
14 Wagner KR et al. Gentamicin treatment of Duchenne and Beckermuscular dystrophy due to nonsense mutations. Ann Neurol2001; 49: 706–711.
15 Yamamoto K et al. Immune response to adenovirus-deliveredantigens upregulates utrophin and results in mitigation of mus-cle pathology in mdx mice. Hum Gene Ther 2000; 11: 669–680.
16 Xiao X, Li J, Samulski RJ. Efficient long-term gene transfer intomuscle tissue of immunocompetent mice by adeno-associatedvirus vector. J Virol 1996; 70: 8098–8108.
17 Kessler PD et al. Gene delivery to skeletal muscle results in sus-tained expression and systemic delivery of a therapeutic protein.Proc Natl Acad Sci USA 1996; 93: 14082–14087.
18 Fisher KJ et al. Recombinant adeno-associated virus for muscledirected gene therapy. Nat Med 1997; 3: 306–312.
19 Kaplitt MG et al. Long-term gene expression and phenotypiccorrection using adeno-associated virus vectors in mammalianbrain. Nat Genet 1994; 8: 148–153.
20 Jooss K, Yang Y, Fisher KJ, Wilson JM. Transduction of dendriticcells by DNA viral vectors directs the immune response to trans-gene products in muscle cells. J Virol 1998; 72: 4212–4223.
21 Greelish JP et al. Stable restoration of the sarcoglycan complex indystrophic muscle perfused with histamine and a recombinantadeno-associated viral vector. Nat Med 1999; 5: 439–443.
22 Li J et al. rAAV vector-mediated sarcoglycan gene transfer in ahamster model for limb girdle muscular dystrophy. Gene Ther-apy 1999; 6: 74–82.
23 Cordier L et al. Rescue of skeletal muscles of �-sarcoglycan-deficient mice with adeno-associated virus-mediated gene trans-fer. Mol Ther 2000; 1: 119–129.
24 Cordier L et al. Muscle-specific promoters may be necessary foradeno-associated virus-mediated gene transfer in the treatmentof muscular dystrophies. Hum Gene Ther 2001; 12: 205–215.
25 England SB et al. Very mild muscular dystrophy associated withthe deletion of 46% of dystrophin. Nature 1990; 343: 180–182.
26 Phelps SF et al. Expression of full-length and truncated dystro-phin mini-gene in transgenic mdx mice. Hum Gene Ther 1995; 4:1251–1258.
27 Yuasa K et al. Effective restoration of dystrophin-associated pro-teins in vivo by adenovirus-mediated transfer of truncated dys-trophin cDNAs. FEBS Lett 1998; 425: 329–336.
28 Wang B, Li J, Xiao X. Adeno-associated virus vector carryinghuman minidystrophin genes effectively ameliorates musculardystrophy in mdx mouse model. Proc Natl Acad Sci USA 2000;97: 13714–13719.
29 Straub V, Rafael JA, Chamberlain JS, Campbell KP. Animalmodels for muscular dystrophy show different patterns of sar-colemmal disruption. J Cell Biol 1997; 139: 375–385.
30 Zhang Y, Chirmule N, Gao G-P, Wilson J. CD40 ligand-depen-dent activation of cytotoxic T lymphocytes by adeno-associatedvirus vectors in vivo: role of immature dendritic cells. J Virol2000; 74: 8003–8010.
31 Witmer-Pack MD et al. Tissue distribution of the DEC-205 pro-tein that is detected by the monoclonal antibody NLDC-145. CellImmunol 1995; 163: 157–162.
32 Khan MA. Corticosteroid therapy in Duchenne muscular dys-trophy. J Neurol Sci 1993; 120: 8–14.
33 Pimorady-Esfahani A, Grounds MD, McMenamin PG. Macro-phages and dendritic cells in normal and regenerating murineskeletal muscle. Muscle Nerve 1997; 20: 158–166.
34 Hartigan-O’Connor D et al. Immune evasion by muscle-specificgene expression in dystrophic muscle. Mol Ther 2001; 4: 525–533.
35 Hohlfeld R, Engel AG. The immunobiology of muscle. ImmunolToday 1994; 15: 269–274.
36 Tinsley JM et al. Amelioration of the dystrophic phenotype ofmdx mice using a truncated utrophin transgene. Nature 1996; 28:349–353.
37 Xiao X, Li J, Samulski RJ. Production of high-titer recombinantadeno-associated virus vectors in the absence of helper aden-ovirus. J Virol 1998; 72: 2224–2232.
38 Shield MA et al. E-box sites and a proximal regulatory regionof the muscle creatine kinase gene differentially regulateexpression in diverse skeletal muscles and cardiac muscle oftransgenic mice. Mol Cell Biol 1996; 16: 5058–5068.
39 Snyder R, Xiao X, Samulski RJ. Production of recombinantadeno-associated viral vectors. In: Dracopoli N et al (ed.). Cur-rent Protocols in Human Genetics. John Wiley: New York, 1996,pp 1–2.
40 Ishii A et al. Effective adenovirus-mediated gene expression inadult murine skeletal muscle. Muscle Nerve 1999; 22: 592–599.
Copyright © 2022 FDOKUMEN




















