Recombinant Adeno-Associated Viral Vectors in the Nervous System
11
HUMAN GENE THERAPY 16:781–791 (July 2005) © Mary Ann Liebert, Inc. Review Recombinant Adeno-Associated Viral Vectors in the Nervous System CORINNA BURGER, 1–3 KEVIN NASH, 1,2 and RONALD J. MANDEL 2–4 ABSTRACT Recombinant adeno-associated virus 2 (rAAV2) has been extensively used as a gene delivery vector for the nervous system. It targets primarily neurons in the nervous system and results in sustained long-term ex- pression of transgenes. New rAAV serotypes have been characterized and demonstrated to have improved transduction efficiencies in various regions of the brain and spinal cord. This review discusses some proper- ties of rAAV that have been studied in the nervous system such as cell tropism, duration of transgene ex- pression, and distribution of viral transduction, as well as immunity and regulation of transgene expression issues, all of which are important for optimization of the use of rAAV in the nervous system. 781 INTRODUCTION R ECOMBINANT ADENO-ASSOCIATED VIRUS (rAAV) has been an attractive gene therapy vehicle since the wild-type virus was genetically engineered in the early 1980s (Carter, 2004). The features of this human parvovirus that make it an attrac- tive tool for gene therapy include the fact that the wild-type virus is nonpathogenic, that all viral genes can be removed from the recombinant virus, and that replication is dependent on coin- fection with a lytic helper virus (Muzyczka and Berns, 2001) and therefore reversion to replication-competent virus is an un- likely event in the context of gene therapy. Furthermore, rela- tively new purification methods have resulted in almost com- pletely pure rAAV vector stocks (Kaludov et al., 2002; Zolotukhin et al., 2002; Smith et al., 2003; Blouin et al., 2004; Davidoff et al., 2004). All these features, together with the fact that rAAV can target neurons in the nervous system, have made this vector a valuable system for the delivery of genes to the brain for the treatment of neurological disorders and for the study of nervous system function. VIRAL CELL TROPISM IN THE NERVOUS SYSTEM: EFFICIENCY OF EXPRESSION IN DIVERSE NEUROANATOMICAL REGIONS There are approximately 35 known serotypes of AAV. To date, at least 10 have been engineered into recombinant viral vectors (Gao et al., 2003; Grimm and Kay, 2003). rAAV2 has been the most widely used AAV serotype to deliver genes to the CNS (Kaplitt et al., 1994; Ali et al., 1996; McCown et al., 1996; Flannery et al., 1997; Peel et al., 1997; Klein et al., 1998; Glatzel et al., 2000). rAAV2 transduces mostly neurons (Mc- Cown et al., 1996; Peel et al., 1997; Bartlett et al., 1998; Klein et al., 1998; Mandel et al., 1998), although it can infect some nonneuronal cell types at a significantly lower efficiency (Kaplitt et al., 1994; McCown et al., 1996; Peel et al., 1997; Klein et al., 1998; Lo et al., 1999; Davidson et al., 2000; Cuc- chiarini et al., 2003). AAV2 has been shown to efficiently trans- duce almost exclusively neurons in a number of brain structures including the substantia nigra (Kaplitt et al., 1994; Mandel et al., 1997; Klein et al., 1998; Bjorklund et al., 2000; Furler et 1 Department of Molecular Genetics and Microbiology, College of Medicine, University of Florida, Gainesville, FL 32610. 2 Powell Gene Therapy Center, College of Medicine, University of Florida, Gainesville, FL 32610. 3 McKnight Brain Institute, College of Medicine, University of Florida, Gainesville, FL 32610. 4 Department of Neuroscience, College of Medicine, University of Florida, Gainesville, FL 32610.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Recombinant Adeno-Associated Viral Vectors in the Nervous System

HUMAN GENE THERAPY 16:781–791 (July 2005)© Mary Ann Liebert, Inc.
Review
Recombinant Adeno-Associated Viral Vectors in the Nervous System
CORINNA BURGER,1–3 KEVIN NASH,1,2 and RONALD J. MANDEL2–4
ABSTRACT
Recombinant adeno-associated virus 2 (rAAV2) has been extensively used as a gene delivery vector for thenervous system. It targets primarily neurons in the nervous system and results in sustained long-term ex-pression of transgenes. New rAAV serotypes have been characterized and demonstrated to have improvedtransduction efficiencies in various regions of the brain and spinal cord. This review discusses some proper-ties of rAAV that have been studied in the nervous system such as cell tropism, duration of transgene ex-pression, and distribution of viral transduction, as well as immunity and regulation of transgene expressionissues, all of which are important for optimization of the use of rAAV in the nervous system.
781
INTRODUCTION
RECOMBINANT ADENO-ASSOCIATED VIRUS (rAAV) has been anattractive gene therapy vehicle since the wild-type virus
was genetically engineered in the early 1980s (Carter, 2004).The features of this human parvovirus that make it an attrac-tive tool for gene therapy include the fact that the wild-typevirus is nonpathogenic, that all viral genes can be removed fromthe recombinant virus, and that replication is dependent on coin-fection with a lytic helper virus (Muzyczka and Berns, 2001)and therefore reversion to replication-competent virus is an un-likely event in the context of gene therapy. Furthermore, rela-tively new purification methods have resulted in almost com-pletely pure rAAV vector stocks (Kaludov et al., 2002;Zolotukhin et al., 2002; Smith et al., 2003; Blouin et al., 2004;Davidoff et al., 2004). All these features, together with the factthat rAAV can target neurons in the nervous system, have madethis vector a valuable system for the delivery of genes to thebrain for the treatment of neurological disorders and for thestudy of nervous system function.
VIRAL CELL TROPISM IN THE NERVOUSSYSTEM: EFFICIENCY OF EXPRESSION INDIVERSE NEUROANATOMICAL REGIONS
There are approximately 35 known serotypes of AAV. Todate, at least 10 have been engineered into recombinant viralvectors (Gao et al., 2003; Grimm and Kay, 2003). rAAV2 hasbeen the most widely used AAV serotype to deliver genes tothe CNS (Kaplitt et al., 1994; Ali et al., 1996; McCown et al.,1996; Flannery et al., 1997; Peel et al., 1997; Klein et al., 1998;Glatzel et al., 2000). rAAV2 transduces mostly neurons (Mc-Cown et al., 1996; Peel et al., 1997; Bartlett et al., 1998; Kleinet al., 1998; Mandel et al., 1998), although it can infect somenonneuronal cell types at a significantly lower efficiency(Kaplitt et al., 1994; McCown et al., 1996; Peel et al., 1997;Klein et al., 1998; Lo et al., 1999; Davidson et al., 2000; Cuc-chiarini et al., 2003). AAV2 has been shown to efficiently trans-duce almost exclusively neurons in a number of brain structuresincluding the substantia nigra (Kaplitt et al., 1994; Mandel etal., 1997; Klein et al., 1998; Bjorklund et al., 2000; Furler et
1Department of Molecular Genetics and Microbiology, College of Medicine, University of Florida, Gainesville, FL 32610.2Powell Gene Therapy Center, College of Medicine, University of Florida, Gainesville, FL 32610.3McKnight Brain Institute, College of Medicine, University of Florida, Gainesville, FL 32610.4Department of Neuroscience, College of Medicine, University of Florida, Gainesville, FL 32610.

al., 2001; Wang et al., 2002; Burger et al., 2004; Paterna et al.,2004), septal area (Mandel et al., 1999a; Klein et al., 2000;Kaspar et al., 2002b; Landgraf et al., 2003), and globus pal-lidus (Tenenbaum et al., 2000; Burger et al., 2004), and slightlyless efficiently the neostriatum (Fan et al., 1998; Burger et al.,2004; Hadaczek et al., 2004; Dass et al., 2004).
rAAV2 has also been shown to strongly transduce neuronsin the dorsal root ganglia (DRG) and peripheral axons (Flem-ing et al., 2001; Xu et al., 2003). rAAV2 transduction in thehippocampus has been preferentially observed in the dentategyrus, with only weak transduction in the CA1 and CA3 fields(McCown et al., 1996; Freese et al., 1997; Bartlett et al., 1998;Klein et al., 1998; Shimazaki et al., 2000; Ehrengruber et al.,2001; Kaspar et al., 2002a; Klein et al., 2002; Passini et al.,2002, 2003; Haberman et al., 2003; Lin et al., 2003; Scammellet al., 2003; Ahmed et al., 2004; Burger et al., 2004).
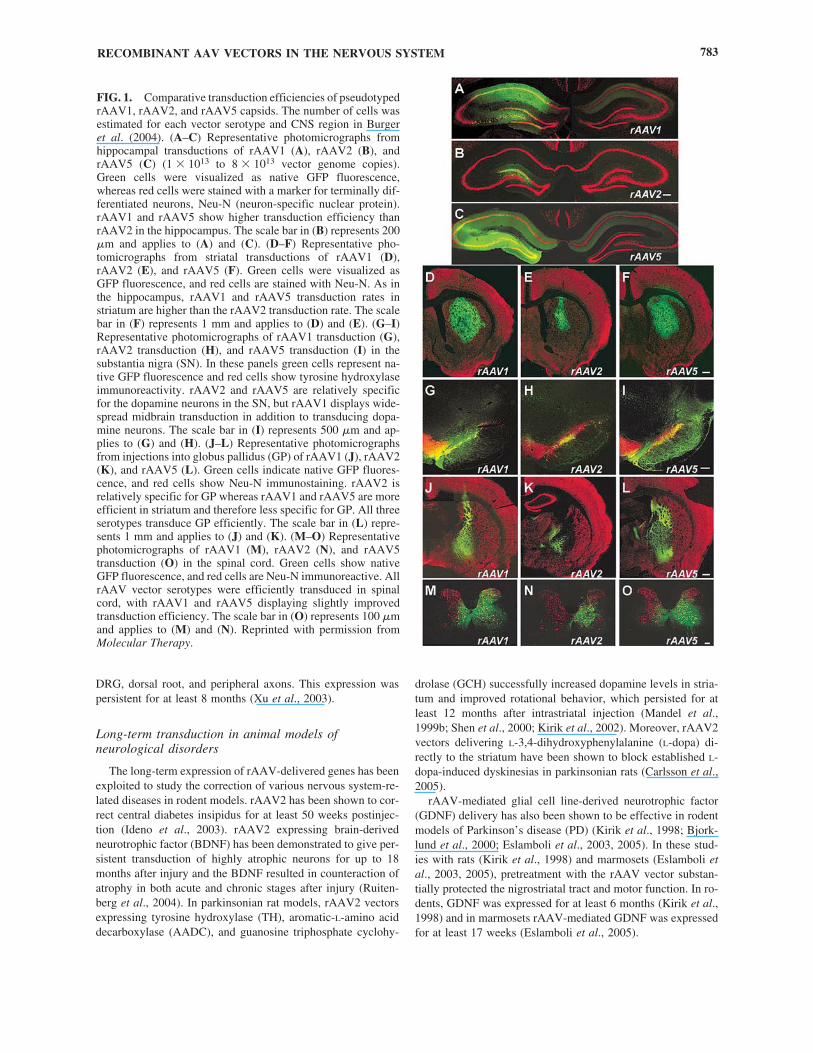
Because different AAV serotypes enter cells via distinct cellsurface receptors (Summerford and Samulski, 1998; Qing et al.,1999; Summerford et al., 1999; Walters et al., 2001; DiPasquale et al., 2003), it was hypothesized that differentserotypes would target diverse cell types in the nervous system.Therefore, the identification of other serotypes and their de-velopment into recombinant AAV vectors sparked interest intesting these serotypes in the CNS. To date, rAAV1, rAAV2,rAAV4, rAAV5, and rAAV6 have been tested in the nervoussystem (Alisky et al., 2000; Davidson et al., 2000; Yang et al.,2002; Passini et al., 2003; Vite et al., 2003; Burger et al., 2004)(Fig. 1).
rAAV1 and rAAV5 have demonstrated a higher distributionand number of neurons transduced than rAAV2 in the CNS(Alisky et al., 2000; Davidson et al., 2000; Auricchio et al.,2001; Yang et al., 2002; Passini et al., 2003; Wang et al., 2003;Weber et al., 2003; Burger et al., 2004) (see Fig. 1). rAAV4has been shown to target ependymal cells exclusively; rAAV1rAAV2, and rAAV5 transduce both neurons and ependymalcells (Davidson et al., 2000; Wang et al., 2003). In the retina,rAAV2 and rAAV5 transduce retinal pigment epithelium (RPE)and photoreceptor cells, with higher transduction in the pho-toreceptor cells, whereas rAAV4 transduces RPE cells only(Weber et al., 2003).
rAAV1 and rAAV5 have been shown to have higher trans-duction efficiency and higher levels of transgene expressionthan rAAV2 in striatum, hippocampus, globus pallidus, sub-stantia nigra, spinal cord, and cerebellum (Alisky et al., 2000;Davidson et al., 2000; Wang et al., 2003; Burger et al., 2004;Paterna et al., 2004) (see Fig. 1). rAAV2 transduction of thehippocampus is limited to the hilar region; however, rAAV1and rAAV5 transduce the entire hippocampus including thepyramidal neurons in the CA1–CA3 region (Di Pasquale et al.,2003; Burger et al., 2004; Richichi et al., 2004; Klugmann etal., 2005) (see Fig. 1). rAAV1 has also been shown to have bet-ter transduction than rAAV2 in the neonatal mouse brain(Passini et al., 2003), and in the cat brain (where rAAV5 didnot transduce neurons to any significant level) (Passini et al.,2003).
rAAV serotypes 6–8 have shown greater transduction thanrAAV2 in other tissue organs such as lung, liver, pancreas, andmuscle (Halbert et al., 2001; Gao et al., 2002; Louboutin et al.,2004; Wang et al., 2004, 2005) and transduction in the liver
has been shown to be stable for more than 2 years with rAAV8(Wang et al., 2005). Indeed, rAAV8 appears to transduce stria-tum at a higher efficiency than rAAV1, rAAV2, and rAAV5(R.J. Mandel, C. Burger, N. Muzyczka, W.W. Hauswirth, F.Manfredsson, and S. Reimsnider, unpublished observations).
LONG-TERM PERSISTENCE OF TRANSGENEEXPRESSION IN VIVO
Early studies of long-term rAAV transduction in theCNS
rAAV2, the first and most commonly used serotype until afew years ago, was found to support long-term transgene ex-pression in the CNS (Kaplitt et al., 1994; McCown et al., 1996;Mandel et al., 1997; Peel et al., 1997; Bartlett et al., 1998; Kleinet al., 1998), although initial experiments with rAAV2 ex-pressing �-galactosidase (�-Gal) in the rat brain yielded vari-able results. Stable expression was observed in some regionsof the brain, such as the inferior colliculus; however, after only3 months, significant reduction of transgene expression levelswas observed in other regions such as the hippocampus (Mc-Cown et al., 1996). Similarly, Lo et al. (1999) injected rAAV2with �-Gal into BALB/c mouse caudate and observed expres-sion for as long as 1 year, although the amount of positive cellsdecreased significantly compared with levels seen 2 monthspostinjection. The low levels of expression in all these studieswere believed to be due to silencing of the cytomegalovirus(CMV) promoter, most likely due to hypermethylation (Proschet al., 1996; Lo et al., 1999).
This led to the investigation of other promoters, includingthe neuron-specific enolase (NSE) promoter, the platelet-de-rived growth factor � (PDGF-�) promoter, the myelin basicprotein (MBP) promoter, and two synthetic promoters (theCMV plus human growth hormone first intron enhancer, andthe hybrid CMV-chicken �-actin [CBA] promoter) (Niwa et al.,1991; Peel et al., 1997; Klein et al., 1998; Chen et al., 1999;Paterna et al., 2000; Fu et al., 2002; Owen et al., 2002; Idenoet al., 2003). In several studies, the MD promoter (CMV witha human �-globin intron) was also shown to support expres-sion for at least 1 year (Mandel et al., 1998; Leff et al., 1999;Szczypka et al., 1999). These promoters were found to be sig-nificantly better at consistent long-term expression (at least6–12 months) in the brain and therefore better candidates thanthe CMV promoter. Paterna et al. (2000) also tested the wood-chuck hepatitis virus posttranscriptional regulatory element(WPRE) (Donello et al., 1998; Loeb et al., 1999) in combina-tion with the PDGF-� promoter. The authors showed that greenfluorescent protein (GFP) expression was more robust with theWPRE and was stable for as long as 41 weeks in dopaminer-gic neurons and their projections in the striatum of rats.
In addition to the brain, rAAV has been found to transduceneurons in the spinal cord (Peel et al., 1997; Burger et al., 2004)(see Fig. 1) and cells in the peripheral nervous system (PNS)(Martinov et al., 2002; Xu et al., 2003). Long-term expressionin dorsal root ganglia (DRG) has been observed after injectionof rAAV2 into the sciatic nerve or DRG, which resulted instrong GFP expression of a large number of neurons of the
BURGER ET AL.782
DRG, dorsal root, and peripheral axons. This expression waspersistent for at least 8 months (Xu et al., 2003).
Long-term transduction in animal models ofneurological disorders
The long-term expression of rAAV-delivered genes has beenexploited to study the correction of various nervous system-re-lated diseases in rodent models. rAAV2 has been shown to cor-rect central diabetes insipidus for at least 50 weeks postinjec-tion (Ideno et al., 2003). rAAV2 expressing brain-derivedneurotrophic factor (BDNF) has been demonstrated to give per-sistent transduction of highly atrophic neurons for up to 18months after injury and the BDNF resulted in counteraction ofatrophy in both acute and chronic stages after injury (Ruiten-berg et al., 2004). In parkinsonian rat models, rAAV2 vectorsexpressing tyrosine hydroxylase (TH), aromatic-L-amino aciddecarboxylase (AADC), and guanosine triphosphate cyclohy-
drolase (GCH) successfully increased dopamine levels in stria-tum and improved rotational behavior, which persisted for atleast 12 months after intrastriatal injection (Mandel et al.,1999b; Shen et al., 2000; Kirik et al., 2002). Moreover, rAAV2vectors delivering L-3,4-dihydroxyphenylalanine (L-dopa) di-rectly to the striatum have been shown to block established L-dopa-induced dyskinesias in parkinsonian rats (Carlsson et al.,2005).
rAAV-mediated glial cell line-derived neurotrophic factor(GDNF) delivery has also been shown to be effective in rodentmodels of Parkinson’s disease (PD) (Kirik et al., 1998; Bjork-lund et al., 2000; Eslamboli et al., 2003, 2005). In these stud-ies with rats (Kirik et al., 1998) and marmosets (Eslamboli etal., 2003, 2005), pretreatment with the rAAV vector substan-tially protected the nigrostriatal tract and motor function. In ro-dents, GDNF was expressed for at least 6 months (Kirik et al.,1998) and in marmosets rAAV-mediated GDNF was expressedfor at least 17 weeks (Eslamboli et al., 2005).
RECOMBINANT AAV VECTORS IN THE NERVOUS SYSTEM 783
FIG. 1. Comparative transduction efficiencies of pseudotypedrAAV1, rAAV2, and rAAV5 capsids. The number of cells wasestimated for each vector serotype and CNS region in Burgeret al. (2004). (A–C) Representative photomicrographs fromhippocampal transductions of rAAV1 (A), rAAV2 (B), andrAAV5 (C) (1 � 1013 to 8 � 1013 vector genome copies).Green cells were visualized as native GFP fluorescence,whereas red cells were stained with a marker for terminally dif-ferentiated neurons, Neu-N (neuron-specific nuclear protein).rAAV1 and rAAV5 show higher transduction efficiency thanrAAV2 in the hippocampus. The scale bar in (B) represents 200�m and applies to (A) and (C). (D–F) Representative pho-tomicrographs from striatal transductions of rAAV1 (D),rAAV2 (E), and rAAV5 (F). Green cells were visualized asGFP fluorescence, and red cells are stained with Neu-N. As inthe hippocampus, rAAV1 and rAAV5 transduction rates instriatum are higher than the rAAV2 transduction rate. The scalebar in (F) represents 1 mm and applies to (D) and (E). (G–I)Representative photomicrographs of rAAV1 transduction (G),rAAV2 transduction (H), and rAAV5 transduction (I) in thesubstantia nigra (SN). In these panels green cells represent na-tive GFP fluorescence and red cells show tyrosine hydroxylaseimmunoreactivity. rAAV2 and rAAV5 are relatively specificfor the dopamine neurons in the SN, but rAAV1 displays wide-spread midbrain transduction in addition to transducing dopa-mine neurons. The scale bar in (I) represents 500 �m and ap-plies to (G) and (H). (J–L) Representative photomicrographsfrom injections into globus pallidus (GP) of rAAV1 (J), rAAV2(K), and rAAV5 (L). Green cells indicate native GFP fluores-cence, and red cells show Neu-N immunostaining. rAAV2 isrelatively specific for GP whereas rAAV1 and rAAV5 are moreefficient in striatum and therefore less specific for GP. All threeserotypes transduce GP efficiently. The scale bar in (L) repre-sents 1 mm and applies to (J) and (K). (M–O) Representativephotomicrographs of rAAV1 (M), rAAV2 (N), and rAAV5transduction (O) in the spinal cord. Green cells show nativeGFP fluorescence, and red cells are Neu-N immunoreactive. AllrAAV vector serotypes were efficiently transduced in spinalcord, with rAAV1 and rAAV5 displaying slightly improvedtransduction efficiency. The scale bar in (O) represents 100 �mand applies to (M) and (N). Reprinted with permission fromMolecular Therapy.

Various rAAV serotypes (rAAV1, rAAV2, and rAAV5)have also been effective in long-term (4–12 months) correctionof mucopolysaccharidosis (MPS) type I, II, and VII lysosomalstorage disease (Bosch et al., 2000; Fu et al., 2002; Passini etal., 2003; Desmaris et al., 2004). In transgenic knockout micefor Canavan’s disease, rAAV2 carrying the gene for asparto-acylase remarkably reduced the spongiform degeneration ob-served in the thalamus up to 5 months postinjection (Matalonet al., 2003).
Large animals have also been examined as a better model ofhuman viral transduction. Stable GFP expression has been ob-served in both RPE and photoreceptor cells in the eyes of bothdogs and cynomolgus macaques for at least 6 months postinjec-tion of rAAV2, rAAV4, and rAAV5 (Weber et al., 2003). Sub-retinal injections of rAAV2 carrying Rpe65 into Rpe65–/– dogsresulted in correction of eyesight for at least 9 months postinjec-tion (Narfstrom et al., 2003). In the treatment of Parkinson’s dis-ease, persistent behavioral recovery for up to 10 months postin-jection of rAAV2 vectors (encoding TH, AADC, and GCH) hasbeen observed in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-treated monkeys (Muramatsu et al., 2002). In conclusion, the var-ious rAAVs appear to sustain transgene expression for greatlengths of time, sometimes the lifetime of the animal, with no ob-vious side effects.
METHODS TO ENHANCE DISTRIBUTION OF TRANSDUCTION
Many neurological disorders caused by single-gene muta-tions such as leukodystrophies or lysosomal storage disorderswill require either global brain transduction or at least wide-spread transduction for successful treatment. One of the currentchallenges of virus-mediated gene therapy is the ability to trans-duce sufficient cells in the CNS to successfully treat disordersrequiring widespread transduction and, for the most part, rAAVcannot meet this challenge. As mentioned above, capsidserotypes such as rAAV1 and rAAV5 display increased trans-duction efficiency in many brain regions (Davidson et al., 2000;Burger et al., 2004) (see Fig. 1). In addition, some ancillarymethods to further increase distribution of rAAV transductionin the CNS have been studied. Infusion of rAAV2 with hep-arin, presumably to block one of the coreceptors for AAV bind-ing, has been reported to increase the distribution of rAAVtransduction (Nguyen et al., 2001; Mastakov et al., 2002). Asimilar approach has been used with basic fibroblast growthfactor, the other AAV coreceptor (Hadaczek et al., 2004). Amethod of slow, long-term infusion of rAAV, termed convec-tion-enhanced delivery, which essentially induces a controllededema in the brain, has also been demonstrated to greatly en-hance the distribution of rAAV-mediated transduction in thebrain (Nguyen et al., 2001). Finally, mannitol, a complex sugarthat induces hyperosmolality in the brain, thereby shrinking theinterstitial space and reversing the flow of cerebrospinal fluid(CSF), has been reported to increase both the efficiency and thedistribution of transduction when mixed with the vector stock(Mastakov et al., 2001) or after systemic injection (Fu et al.,2003; Burger et al., 2005) (Fig. 2).
BRAIN IMMUNE RESPONSE TO AAV
Long-term transgene expression is apparently not accompa-nied by any significant toxicity in naive animals with low trans-duction rates and relatively weak promoters (Mandel et al.,1997, 1998). However, there is evidence of immune responseswhen the animals have been preexposed to rAAV2 (Peden etal., 2004; Sanftner et al., 2004). Thus, peripheral preimmu-nization with either rAAV2 (Sanftner et al., 2004) or wild-typeAAV2 (Peden et al., 2004) can completely block brain trans-duction. However, preimmunization with rAAV2 does notblock subsequent striatal transduction with rAAV5-pseudo-typed vectors (Peden et al., 2004).
Furthermore, intrastriatal rAAV2 injections are sufficient toprime an immune response because a second injection ofrAAV2 in the opposite striatum either 2 weeks (C.S. Peden andR.J. Mandel, unpublished observations) or 4 weeks after thefirst injection led to highly significant inflammatory responseand infiltration of CD8a� cells (Peden et al., 2004). If the trans-gene is not immunogenic, transgene expression appears to berelatively unaffected, even at the second injection site (Pedenet al., 2004). However, if the expressed transgene is immuno-genic, such as humanized jellyfish GFP, there is significantclearance of transduced cells in both injection sites (C.S. Pe-den and R.J. Mandel, unpublished observations).
The immune effects of readministration in the brain are un-expected because no viral proteins are expressed in rAAV2-transduced cells. Without viral gene expression and the ex-pression of a nonimmunogenic protein, no presentation ofimmunogenic peptides on MHC class I should occur. More-over, readministration studies with high-capacity adenoviralvectors does not result in clearance of transduced cells after aninitial reduction of transgene expression associated with theearly inflammatory events peculiar to adenoviral infection(Lowenstein and Castro, 2003). It has been shown that rAAV2uncoats slowly, leading to MHC presentation of capsid peptidesover a prolonged period (Hauck et al., 2004; Thomas et al.,2004). It has been hypothesized that it is this slow uncoatingthat accounts for the unexpected immune reaction to readmin-istration of rAAV2 in the brain (Lowenstein, 2004).
In addition, these results regarding the brain immune re-sponse point out that the brain is not as immune privileged aspreviously believed and requires more detailed study as hasbeen performed for adenoviral vectors (Lowenstein and Castro,2003). Therefore, clinical protocols aimed at neurological dis-orders should probably include determination of peripheral cir-culating anti-AAV antibodies before intracerebral injection ofrAAV.
REGULATION OF TRANSGENE EXPRESSIONIN VIVO
The subject of regulation systems and rAAV in the CNS hasbeen discussed in detail (Haberman and McCown, 2002), andtherefore we briefly discuss only a few issues. As described inearlier sections, AAV can produce therapeutic proteins at rela-tively high levels for extended periods of time in the brain. Theconstitutive high-level expression of therapeutic proteins can
BURGER ET AL.784

cause undesirable side effects, such as in the case of growthfactors (Lang et al., 2004), which need to be expressed at lowlevels to produce physiological effects. Drug-regulated gene ex-pression systems provide the ability to control the levels of theprotein product quantitatively and temporally. Artificially reg-ulated gene expression systems could potentially avoid side ef-fects due to constitutive expression of the transgene and allowpharmacological control of expression. A number of artificialregulation systems have been reported for the control of trans-
gene expression in vivo, including tetracycline and its deriva-tive doxycycline (Gossen and Bujard, 1992; Gossen et al.,1995), ecdysone (No et al., 1996), the progesterone antagonistmifepristone (Wang et al., 1994), and rapamycin (Rivera et al.,1996). These systems rely on chimeric transcription factors thatbind promoters directing the expression of the transgene of in-terest. The transcription factor in turn responds to a drug thatcan be administered orally and the transgene expression can bedosed in this manner. Regulation of rAAV expression in the
RECOMBINANT AAV VECTORS IN THE NERVOUS SYSTEM 785
FIG. 2. Systemic administration of mannitol significantly enhances the transduction rate of rAAV2 in striatum. (a) Stereolog-ical estimation of the number of GFP-positive cells transduced with either systemic mannitol or saline control. (b) Volume ofGFP expression estimated by stereological method. (c) Low-power micrographs of representative serial coronal sections show-ing the anterior-to-posterior extent of transduction in the striatum. Error bars represent �1 SEM. Sections show GFP im-munoreactivity and illustrate the enhanced transgene distribution in the mannitol-infused brain (left), versus the saline controlbrain (right). The anterior–posterior (AP) measurements that correspond most closely to coronal sections in the Paxinos and Wat-son atlas (Paxinos and Watson, 1997) are indicated next to each section. Scale bar, 1 mm. For systemic injection of mannitol, 3ml of sterile 25% mannitol in 0.9% saline per 100 g body weight was injected intraperitoneally 15 min before intracerebral vec-tor injections (n � 5), as described above. Control animals received identical preinjections of 3 ml of 0.9% saline per 100 g bodyweight instead of mannitol (n � 5). Unbiased stereological estimation of the total number of cells in striatum was obtained bythe optical fractionator method, as previously described in detail (Burger et al., 2004; Peden et al., 2004). Actual cell countingwas performed with a �100 oil objective. The total number of neurons was estimated according to the optical fractionator for-mula (West et al., 1991) A coefficient of error (CE) was calculated according to Gundersen and Jensen (1987) for each animaland the estimation for each animal was accepted only when the CE was �0.1. Parametric analysis of variance (ANOVA) wasused to assess significant differences between treatments. Significance was accepted at the 95% probability level. Reprinted withpermission from Molecular Therapy.
A
C
B

brain, for gene therapy has focused mainly on the tetracycline-regulated system.
The tetracycline-regulated transcription system as developedby Gossen and Bujard (1992) has been the most studied as aneffective system for tight regulation of transgene expression forgene therapy applications (Urlinger et al., 2000). Other systemsare becoming more popular for gene therapy applications in thenervous system. The rapamycin system (Rivera et al., 1996)has been engineered into AAV and successfully used in mus-cle (Rivera et al., 1999; Ye et al., 1999; Johnston, 2003) andliver (Auricchio et al., 2002a). In the eye, intravitreous or sub-retinal injections of erythropoietin (Epo) (Auricchio et al.,2002b), allowed the control of expression of the transgene in areversible manner, although basal levels of Epo took severalweeks to decline after withdrawal of the drug. Tetracycline (Tet)regulation is the only other artificial regulation system that hasbeen engineered into rAAV to date.
One of the biggest issues in the regulation of gene expres-sion, using these artificial systems, concerns “leakiness” (or in-complete shutdown of transgene expression in the off-state).This is important when basal levels of expression of the trans-gene are unacceptable because expression of the gene can betoxic to the system. In the context of rAAV it has been arguedthat expression of the transgene in the absence of a transcrip-tional activator is probably due to the fact that AAV2 terminalrepeats (TRs) have endogenous enhancer/promoter functionthat is directing expression in the absence of a Tet activator.This has been demonstrated (Flotte et al., 1993; Haberman etal., 2000) and also discussed in Haberman and McCown (2002).Various expression cassettes have been tested to resolve thisbasal expression issue (Bohl et al., 1998; Fitzsimons et al.,2001; Haberman et al., 2002; Samakoglu et al., 2002; Chtartoet al., 2003; Folliot et al., 2003; Chenuaud et al., 2004; Gafniet al., 2004; Jiang et al., 2004), but no leak-proof constructshave been convincingly demonstrated in vivo. Until an optimalcassette that results in good induction and negligible basal ex-pression is achieved, it will not be safe to deliver genes to thenervous system for the cure of human neurological disorders.Therefore, in order for a gene therapy approach to reach theclinic, tight regulation of transgene expression needs to beachieved.
CONCLUDING REMARKS
Given that the safety and efficacy profile of rAAV2 vectorshas been encouraging, especially in the CNS, it is not surpris-ing that this vector has entered human clinical trials. There arecurrently five rAAV2-based clinical trials either underway orlate in the approval process (Mandel and Burger, 2004). Todate, none of these trials incorporate a regulated rAAV vector,nor have any of the trials explicitly dealt with immune re-sponses. Further development of the rAAV system for enhancedtransduction efficiency and targeting of various cellular phe-notypes is still highly warranted to improve the clinical utilityof this vector system. The capsid structure of AAV2 has beenresolved by electron cryomicroscopy (Kronenberg et al., 2001)and by X-ray crystallography (Xie et al., 2002); in addition,mutagenesis studies of the capsid protein have revealed the pu-tative cell receptor-binding sites (Wu et al., 2000; Opie et al.,
2003). These studies have been key to potential retargeting ofthe recombinant viral vector (Warrington et al., 2004). Contin-ued advancements in this field of study may allow targeting ofrAAV to specific neuronal phenotypes, or to glial cells. Also,improved delivery techniques will allow increasing retrogradetransport for better treatment of motoneuron disorders, or mas-sively augmenting transduction efficiency in the nervous sys-tem. Therefore, future advances in the areas discussed in thisreview may hold exciting developments in the field of genetherapy in the CNS.
REFERENCES
AHMED, B.Y., CHAKRAVARTHY, S., EGGERS, R., HERMENS,W.T., ZHANG, J.Y., NICLOU, S.P., LEVELT, C., SABLITZKY,F., ANDERSON, P.N., LIEBERMAN, A.R., and VERHAAGEN, J.(2004). Efficient delivery of Cre-recombinase to neurons in vivo andstable transduction of neurons using adeno-associated and lentiviralvectors. BMC Neurosci. 5, 4.
ALI, R.R., REICHEL, M.B., THRASHER, A.J., LEVINSKY, R.J.,KINNON, C., KANUGA, N., HUNT, D.M., and BHAT-TACHARYA, S.S. (1996). Gene transfer into the mouse retina me-diated by an adeno-associated viral vector. Hum. Mol. Genet. 5,591–594.
ALISKY, J.M., HUGHES, S.M., SAUTER, S.L., JOLLY, D., DUBEN-SKY, T.W., Jr., STABER, P.D., CHIORINI, J.A., and DAVIDSON,B.L. (2000). Transduction of murine cerebellar neurons with re-combinant FIV and AAV5 vectors. Neuroreport 11, 2669–2673.
AURICCHIO, A., KOBINGER, G., ANAND, V., HILDINGER, M.,O’CONNOR, E., MAGUIRE, A.M., WILSON, J.M., and BEN-NETT, J. (2001). Exchange of surface proteins impacts on viral vec-tor cellular specificity and transduction characteristics: The retina asa model. Hum. Mol. Genet. 10, 3075–3081.
AURICCHIO, A., GAO, G.P., YU, Q.C., RAPER, S., RIVERA, V.M.,CLACKSON, T., and WILSON, J.M. (2002a). Constitutive and reg-ulated expression of processed insulin following in vivo hepatic genetransfer. Gene Ther. 9, 963–971.
AURICCHIO, A., RIVERA, V.M., CLACKSON, T., O’CONNOR,E.E., MAGUIRE, A.M., TOLENTINO, M.J., BENNETT, J., andWILSON, J.M. (2002b). Pharmacological regulation of protein ex-pression from adeno-associated viral vectors in the eye. Mol. Ther.6, 238–242.
BARTLETT, J.S., SAMULSKI, R.J., and MCCOWN, T.J. (1998). Se-lective and rapid uptake of adeno-associated virus type 2 in brain.Hum. Gene Ther. 9, 1181–1186.
BJORKLUND, A., KIRIK, D., ROSENBLAD, C., GEORGIEVSKA,B., LUNDBERG, C., and MANDEL, R.J. (2000). Towards a neu-roprotective gene therapy for Parkinson’s disease: Use of adenovi-rus, AAV and lentivirus vectors for gene transfer of GDNF to thenigrostriatal system in the rat Parkinson model. Brain Res. 886,82–98.
BLOUIN, V., BRUMENT, N., TOUBLANC, E., RAIMBAUD, I.,MOULLIER, P., and SALVETTI, A. (2004). Improving rAAV pro-duction and purification: Towards the definition of a scaleable pro-cess. J. Gene Med. 6(Suppl. 1), S223–S228.
BOHL, D., SALVETTI, A., MOULLIER, P., and HEARD, J.M. (1998).Control of erythropoietin delivery by doxycycline in mice after in-tramuscular injection of adeno-associated vector. Blood 92,1512–1517.
BOSCH, A., PERRET, E., DESMARIS, N., and HEARD, J.M. (2000).Long-term and significant correction of brain lesions in adult mu-copolysaccharidosis type VII mice using recombinant AAV vectors.Mol. Ther. 1, 63–70.
BURGER ET AL.786

BURGER, C., GORBATYUK, O.S., VELARDO, M.J., PEDEN, C.S.,WILLIAMS, P., ZOLOTUKHIN, S., REIER, P.J., MANDEL, R.J.,and MUZYCZKA, N. (2004). Recombinant AAV viral vectorspseudotyped with viral capsids from serotypes 1, 2, and 5 displaydifferential efficiency and cell tropism after delivery to different re-gions of the central nervous system. Mol. Ther. 10, 302–317.
BURGER, C., NGUYEN, F.N., DENG, J., and MANDEL, R.J. (2005).Systemic mannitol-induced hyperosmolality amplifies rAAV2-me-diated striatal transduction to a greater extent than local co-infusion.Mol. Ther. 11, 327–331.
CARLSSON, T., WINKLER, C., BURGER, C., MUZYCZKA, N.,MANDEL, R.J., CENCI, A., BJORKLUND, A., and KIRIK, D.(2005). Reversal of dyskinesias in an animal model of Parkinson’sdisease by continuous L-DOPA delivery using rAAV vectors. Brain128, 559–569.
CARTER, B.J. (2004). Adeno-associated virus and the development ofadeno-associated virus vectors: A historical perspective. Mol. Ther.10, 981–989.
CHEN, H., MCCARTY, D.M., BRUCE, A.T., and SUZUKI, K. (1999).Oligodendrocyte-specific gene expression in mouse brain: Use of amyelin-forming cell type-specific promoter in an adeno-associatedvirus. J. Neurosci. Res. 55, 504–513.
CHENUAUD, P., LARCHER, T., RABINOWITZ, J.E., PROVOST,N., JOUSSEMET, B., BUJARD, H., SAMULSKI, R.J., FAVRE, D.,and MOULLIER, P. (2004). Optimal design of a single recombinantadeno-associated virus derived from serotypes 1 and 2 to achievemore tightly regulated transgene expression from nonhuman primatemuscle. Mol. Ther. 9, 410–418.
CHTARTO, A., BENDER, H.U., HANEMANN, C.O., KEMP, T.,LEHTONEN, E., LEVIVIER, M., BROTCHI, J., VELU, T., andTENENBAUM, L. (2003). Tetracycline-inducible transgene expres-sion mediated by a single AAV vector. Gene Ther. 10, 84–94.
CUCCHIARINI, M., REN, X.L., PERIDES, G., and TERWILLIGER,E.F. (2003). Selective gene expression in brain microglia mediatedvia adeno-associated virus type 2 and type 5 vectors. Gene Ther. 10,657–667.
DASS, B., IRAVANI, M.M., HUANG, C., BARSOUM, J., ENGBER,T.M., GALDES, A., and JENNER, P. (2004). Sonic hedgehog de-livered by an adeno-associated virus protects dopaminergic neuronesagainst 6-OHDA toxicity in the rat. J. Neural Transm. (Dec. 10; epubahead of print).
DAVIDOFF, A.M., NG, C.Y., SLEEP, S., GRAY, J., AZAM, S.,ZHAO, Y., MCINTOSH, J.H., KARIMIPOOR, M., and NATH-WANI, A.C. (2004). Purification of recombinant adeno-associatedvirus type 8 vectors by ion exchange chromatography generates clin-ical grade vector stock. J. Virol. Methods 121, 209–215.
DAVIDSON, B.L., STEIN, C.S., HETH, J.A., MARTINS, I., KOTIN,R.M., DERKSEN, T.A., ZABNER, J., GHODSI, A., and CHIORINI,J.A. (2000). Recombinant adeno-associated virus type 2, 4, and 5 vec-tors: Transduction of variant cell types and regions in the mammaliancentral nervous system. Proc. Natl. Acad. Sci. U.S.A. 97, 3428–3432.
DESMARIS, N., VEROT, L., PUECH, J.P., CAILLAUD, C., VANIER,M.T., and HEARD, J.M. (2004). Prevention of neuropathology inthe mouse model of Hurler syndrome. Ann. Neurol. 56, 68–76.
DI PASQUALE, G., DAVIDSON, B.L., STEIN, C.S., MARTINS, I.,SCUDIERO, D., MONKS, A., and CHIORINI, J.A. (2003). Identi-fication of PDGFR as a receptor for AAV-5 transduction. Nat. Med.9, 1306–1312.
DONELLO, J.E., LOEB, J.E., and HOPE, T.J. (1998). Woodchuck hep-atitis virus contains a tripartite posttranscriptional regulatory element.J. Virol. 72, 5085–5092.
EHRENGRUBER, M.U., HENNOU, S., BUELER, H., NAIM, H.Y.,DEGLON, N., and LUNDSTROM, K. (2001). Gene transfer intoneurons from hippocampal slices: Comparison of recombinant Sem-liki Forest virus, adenovirus, adeno-associated virus, lentivirus, andmeasles virus. Mol. Cell. Neurosci. 17, 855–871.
ESLAMBOLI, A., CUMMINGS, R.M., RIDLEY, R.M., BAKER, H.F.,MUZYCZKA, N., BURGER, C., MANDEL, R.J., KIRIK, D., andANNETT, L.E. (2003). Recombinant adeno-associated viral vector(rAAV) delivery of GDNF provides protection against 6-OHDA le-sion in the common marmoset monkey (Callithrix jacchus). Exp.Neurol. 184, 536–548.
ESLAMBOLI, A., GEORGIEVSKA, B., RIDLEY, R.M., BAKER,H.F., MUZYCZKA, N., BURGER, C., MANDEL, R.J., ANNETT,L., and KIRIK, D. (2005). Continuous low-level glial cell line-de-rived neurotrophic factor delivery using recombinant adeno-associ-ated viral vectors provides neuroprotection and induces behavioralrecovery in a primate model of Parkinson’s disease. J. Neurosci. 25,769–777.
FAN, D.S., OGAWA, M., FUJIMOTO, K.I., IKEGUCHI, K., OGA-SAWARA, Y., URABE, M., NISHIZAWA, M., NAKANO, I.,YOSHIDA, M., NAGATSU, I., ICHINOSE, H., NAGATSU, T.,KURTZMAN, G.J., and OZAWA, K. (1998). Behavioral recoveryin 6-hydroxydopamine-lesioned rats by cotransduction of striatumwith tyrosine hydroxylase and aromatic L-amino acid decarboxylasegenes using two separate adeno-associated virus vectors. Hum. GeneTher. 9, 2527–2535.
FITZSIMONS, H.L., MCKENZIE, J.M., and DURING, M.J. (2001).Insulators coupled to a minimal bidirectional tet cassette for tightregulation of rAAV-mediated gene transfer in the mammalian brain.Gene Ther. 8, 1675–1681.
FLANNERY, J.G., ZOLOTUKHIN, S., VAQUERO, M.I., LAVAIL,M.M., MUZYCZKA, N., and HAUSWIRTH, W.W. (1997). Effi-cient photoreceptor-targeted gene expression in vivo by recombinantadeno-associated virus. Proc. Natl. Acad. Sci. U.S.A. 94, 6916–6921.
FLEMING, J., GINN, S.L., WEINBERGER, R.P., TRAHAIR, T.N.,SMYTHE, J.A., and ALEXANDER, I.E. (2001). Adeno-associatedvirus and lentivirus vectors mediate efficient and sustained trans-duction of cultured mouse and human dorsal root ganglia sensoryneurons. Hum. Gene Ther. 12, 77–86.
FLOTTE, T.R., AFIONE, S.A., SOLOW, R., DRUMM, M.L.,MARKAKIS, D., GUGGINO, W.B., ZEITLIN, P.L., and CARTER,B.J. (1993). Expression of the cystic fibrosis transmembrane con-ductance regulator from a novel adeno-associated virus promoter. J.Biol. Chem. 268, 3781–3790.
FOLLIOT, S., BRIOT, D., CONRATH, H., PROVOST, N., CHEREL,Y., MOULLIER, P., and ROLLING, F. (2003). Sustained tetracy-cline-regulated transgene expression in vivo in rat retinal ganglioncells using a single type 2 adeno-associated viral vector. J. Gene Med.5, 493–501.
FREESE, A., KAPLITT, M.G., O’CONNOR, W.M., ABBEY, M.,LANGER, D., LEONE, P., O’CONNOR, M.J., and DURING, M.J.(1997). Direct gene transfer into human epileptogenic hippocampaltissue with an adeno-associated virus vector: Implications for a genetherapy approach to epilepsy. Epilepsia 38, 759–766.
FU, H., SAMULSKI, R.J., MCCOWN, T.J., PICORNELL, Y.J.,FLETCHER, D., and MUENZER, J. (2002). Neurological correctionof lysosomal storage in a mucopolysaccharidosis IIIB mouse modelby adeno-associated virus-mediated gene delivery. Mol. Ther. 5,42–49.
FU, H., MUENZER, J., SAMULSKI, R.J., BREESE, G., SIFFORD,J., ZENG, X., and MCCARTY, D.M. (2003). Self-complementaryadeno-associated virus serotype 2 vector: Global distribution andbroad dispersion of AAV-mediated transgene expression in mousebrain. Mol. Ther. 8, 911–917.
FURLER, S., PATERNA, J.C., WEIBEL, M., and BUELER, H. (2001).Recombinant AAV vectors containing the foot and mouth diseasevirus 2A sequence confer efficient bicistronic gene expression in cul-tured cells and rat substantia nigra neurons. Gene Ther. 8, 864–873.
GAFNI, Y., PELLED, G., ZILBERMAN, Y., TURGEMAN, G., AP-PARAILLY, F., YOTVAT, H., GALUN, E., GAZIT, Z., JOR-GENSEN, C., and GAZIT, D. (2004). Gene therapy platform for
RECOMBINANT AAV VECTORS IN THE NERVOUS SYSTEM 787

bone regeneration using an exogenously regulated, AAV-2-basedgene expression system. Mol. Ther. 9, 587–595.
GAO, G., ALVIRA, M.R., SOMANATHAN, S., LU, Y., VANDEN-BERGHE, L.H., RUX, J.J., CALCEDO, R., SANMIGUEL, J., AB-BAS, Z., and WILSON, J.M. (2003). Adeno-associated viruses un-dergo substantial evolution in primates during natural infections.Proc. Natl. Acad. Sci. U.S.A. 100, 6081–6086.
GAO, G.P., ALVIRA, M.R., WANG, L., CALCEDO, R., JOHNSTON,J., and WILSON, J.M. (2002). Novel adeno-associated viruses fromrhesus monkeys as vectors for human gene therapy. Proc. Natl. Acad.Sci. U.S.A. 99, 11854–11859.
GLATZEL, M., FLECHSIG, E., NAVARRO, B., KLEIN, M.A., PA-TERNA, J.C., BUELER, H., and AGUZZI, A. (2000). Adenoviraland adeno-associated viral transfer of genes to the peripheral ner-vous system. Proc. Natl. Acad. Sci. U.S.A. 97, 442–447.
GOSSEN, M., and BUJARD, H. (1992). Tight control of gene expres-sion in mammalian cells by tetracycline-responsive promoters. Proc.Natl. Acad. Sci. U.S.A. 89, 5547–5551.
GOSSEN, M., FREUNDLIEB, S., BENDER, G., MULLER, G.,HILLEN, W., and BUJARD, H. (1995). Transcriptional activationby tetracyclines in mammalian cells. Science 268, 1766–1769.
GRIMM, D., and KAY, M.A. (2003). From virus evolution to vectorrevolution: Use of naturally occurring serotypes of adeno-associatedvirus (AAV) as novel vectors for human gene therapy. Curr. GeneTher. 3, 281–304.
GUNDERSEN, H.J., and JENSEN, E.B. (1987). The efficiency of sys-tematic sampling in stereology and its prediction. J. Microsc. 147,229–263.
HABERMAN, R.P., and MCCOWN, T.J. (2002). Regulation of geneexpression in adeno-associated virus vectors in the brain. Methods28, 219–226.
HABERMAN, R.P., MCCOWN, T.J., and SAMULSKI, R.J. (2000).Novel transcriptional regulatory signals in the adeno-associated virusterminal repeat A/D junction element. J. Virol. 74, 8732–8739.
HABERMAN, R., CRISWELL, H., SNOWDY, S., MING, Z., BREESE,G., SAMULSKI, R., and MCCOWN, T. (2002). Therapeutic liabilitiesof in vivo viral vector tropism: Adeno-associated virus vectors, NM-DAR1 antisense, and focal seizure sensitivity. Mol. Ther. 6, 495–500.
HABERMAN, R.P., SAMULSKI, R.J., and MCCOWN, T.J. (2003).Attenuation of seizures and neuronal death by adeno-associated virusvector galanin expression and secretion. Nat. Med. 9, 1076–1080.
HADACZEK, P., MIREK, H., BRINGAS, J., CUNNINGHAM, J., andBANKIEWICZ, K. (2004). Basic fibroblast growth factor enhancestransduction, distribution, and axonal transport of adeno-associatedvirus type 2 vector in rat brain. Hum. Gene Ther. 15, 469–479.
HALBERT, C.L., ALLEN, J.M., and MILLER, A.D. (2001). Adeno-associated virus type 6 (AAV6) vectors mediate efficient transduc-tion of airway epithelial cells in mouse lungs compared with that ofAAV2 vectors. J. Virol. 75, 6615–6624.
HAUCK, B., ZHAO, W., HIGH, K., and XIAO, W. (2004). Intracel-lular viral processing, not single-stranded DNA accumulation, is cru-cial for recombinant adeno-associated virus transduction. J. Virol.78, 13678–13686.
IDENO, J., MIZUKAMI, H., HONDA, K., OKADA, T., HANAZONO,Y., KUME, A., SAITO, T., ISHIBASHI, S., and OZAWA, K. (2003).Persistent phenotypic correction of central diabetes insipidus usingadeno-associated virus vector expressing arginine-vasopressin inBrattleboro rats. Mol. Ther. 8, 895–902.
JIANG, L., RAMPALLI, S., GEORGE, D., PRESS, C., BREMER,E.G., O’GORMAN, M.R., and BOHN, M.C. (2004). Tight regula-tion from a single tet-off rAAV vector as demonstrated by flow cy-tometry and quantitative, real-time PCR. Gene Ther. 11, 1057–1067.
JOHNSTON, J., TAZELAAR, J., RIVERA, V.M., CLACKSON, T.,GAO, G.P., and WILSON, J.M. (2003). Regulated expression of ery-thropoietin from an AAV vector safely improves the anemia of �-thalassemia in a mouse model. Mol. Ther. 7, 493–497.
KALUDOV, N., HANDELMAN, B., and CHIORINI, J.A. (2002).Scalable purification of adeno-associated virus type 2, 4, or 5 usingion-exchange chromatography. Hum. Gene Ther. 13, 1235–1243.
KAPLITT, M.G., LEONE, P., SAMULSKI, R.J., XIAO, X., PFAFF,D.W., O’MALLEY, K.L., and DURING, M.J. (1994). Long-termgene expression and phenotypic correction using adeno-associatedvirus vectors in the mammalian brain. Nat. Genet. 8, 148–154.
KASPAR, B.K., ERICKSON, D., SCHAFFER, D., HINH, L., GAGE,F.H., and PETERSON, D.A. (2002a). Targeted retrograde gene de-livery for neuronal protection. Mol. Ther. 5, 50–56.
KASPAR, B.K., VISSEL, B., BENGOECHEA, T., CRONE, S., RAN-DOLPH-MOORE, L., MULLER, R., BRANDON, E.P., SCHAF-FER, D., VERMA, I.M., LEE, K.F., HEINEMANN, S.F., andGAGE, F.H. (2002b). Adeno-associated virus effectively mediatesconditional gene modification in the brain. Proc. Natl. Acad. Sci.U.S.A. 99, 2320–2325.
KIRIK, D., ROSENBLAD, C., and BJORKLUND, A. (1998). Char-acterization of behavioral and neurodegenerative changes followingpartial lesions of the nigrostriatal dopamine system induced by in-trastriatal 6-hydroxydopamine in the rat. Exp. Neurol. 152, 259–277.
KIRIK, D., GEORGIEVSKA, B., BURGER, C., WINKLER, C.,MUZYCZKA, N., MANDEL, R.J., and BJORKLUND, A. (2002).Reversal of motor impairments in parkinsonian rats by continuousintrastriatal delivery of L-dopa using rAAV-mediated gene transfer.Proc. Natl. Acad. Sci. U.S.A. 99, 4708–4713.
KLEIN, R.L., MEYER, E.M., PEEL, A.L., ZOLOTUKHIN, S., MEY-ERS, C., MUZYCZKA, N., and KING, M.A. (1998). Neuron-spe-cific transduction in the rat septohippocampal or nigrostriatal path-way by recombinant adeno-associated virus vectors. Exp. Neurol.150, 183–194.
KLEIN, R.L., HIRKO, A.C., MEYERS, C.A., GRIMES, J.R., MUZY-CZKA, N., and MEYER, E.M. (2000). NGF gene transfer to intrin-sic basal forebrain neurons increases cholinergic cell size and pro-tects from age-related, spatial memory deficits in middle-aged rats.Brain Res. 875, 144–151.
KLEIN, R.L., HAMBY, M.E., SONNTAG, C.F., MILLARD, W.J.,KING, M.A., and MEYER, E.M. (2002). Measurements of vector-derived neurotrophic factor and green fluorescent protein levels inthe brain. Methods 28, 286–292.
KLUGMANN, M., WYMOND SYMES, C., LEICHTLEIN, C.B.,KLAUSSNER, B.K., DUNNING, J., FONG, D., YOUNG, D., andDURING, M.J. (2005). AAV-mediated hippocampal expression ofshort and long Homer 1 proteins differentially affect cognition andseizure activity in adult rats. Mol. Cell. Neurosci. 28, 347–360.
KRONENBERG, S., KLEINSCHMIDT, J.A., and BOTTCHER, B.(2001). Electron cryo-microscopy and image reconstruction of adeno-associated virus type 2 empty capsids. EMBO Rep. 2, 997–1002.
LANDGRAF, R., FRANK, E., ALDAG, J.M., NEUMANN, I.D.,SHARER, C.A., REN, X., TERWILLIGER, E.F., NIWA, M., WIG-GER, A., and YOUNG, L.J. (2003). Viral vector-mediated genetransfer of the vole V1a vasopressin receptor in the rat septum: Im-proved social discrimination and active social behaviour. Eur. J. Neu-rosci. 18, 403–411.
LANG, A.E., GILL, S., BROOKS, D., BRODSKY, M.A., BURCHIEL,K., COFFEY, R.J., DALVI, A., DHAWAN, V., ELIAS, W.J., HEY-WOOD, P., HOTTON, G., HUTCHINSON, M., KELLY, P., LAWS,E.R., LOZANO, A., MATCHAM, J., MORO, E., NUTT, J., PATEL,N.K., PENN, R., SCOTT, B., STACY, M., STOESSL, A.J., TRAUB,M., TURNER, D., and WOOTEN, G.F. (2004). Multicenter, double-blind, randomized, placebo-controlled, parallel group trial of liater-min (rmetHuGDNF) administered by bilateral intraputaminal (IPu)infusion to subjects with idiopathic Parkinson’s disease. In 129th An-nual Meeting of the American Neurological Association, Toronto,Canada (American Neurological Association, Minneapolis, MN).
LEFF, S.E., SPRATT, S.K., SNYDER, R.O., and MANDEL, R.J.(1999). Long-term restoration of striatal L-aromatic amino acid de-
BURGER ET AL.788

carboxylase activity using recombinant adeno-associated viral vec-tor gene transfer in a rodent model of Parkinson’s disease. Neuro-science 92, 185–196.
LIN, E.J., RICHICHI, C., YOUNG, D., BAER, K., VEZZANI, A., andDURING, M.J. (2003). Recombinant AAV-mediated expression ofgalanin in rat hippocampus suppresses seizure development. Eur. J.Neurosci. 18, 2087–2092.
LO, W.D., QU, G., SFERRA, T.J., CLARK, R., CHEN, R., and JOHN-SON, P.R. (1999). Adeno-associated virus-mediated gene transfer tothe brain: Duration and modulation of expression. Hum. Gene Ther.10, 201–213.
LOEB, J.E., CORDIER, W.S., HARRIS, M.E., WEITZMAN, M.D.,and HOPE, T.J. (1999). Enhanced expression of transgenes fromadeno-associated virus vectors with the woodchuck hepatitis virusposttranscriptional regulatory element: Implications for gene therapy.Hum. Gene Ther. 10, 2295–2305.
LOUBOUTIN, J.P., WANG, L., and WILSON, J.M. (2004). Genetransfer into skeletal muscle using novel AAV serotypes. J. GeneMed. 7, 442–451.
LOWENSTEIN, P.R. (2004). Input virion proteins: Cryptic targets ofantivector immune responses in preimmunized subjects. Mol. Ther.9, 771–774.
LOWENSTEIN, P.R., and CASTRO, M.G. (2003). Inflammation andadaptive immune responses to adenoviral vectors injected into thebrain: Peculiarities, mechanisms, and consequences. Gene Ther. 10,946–954.
MANDEL, R.J., and BURGER, C. (2004). Clinical trials in neurolog-ical disorders using AAV vectors: Promises and challenges. Curr.Opin. Mol. Ther. 6, 482–490.
MANDEL, R.J., SPRATT, S.K., SNYDER, R.O., and LEFF, S.E.(1997). Midbrain injection of recombinant adeno-associated virus en-coding rat glial cell line-derived neurotrophic factor protects nigralneurons in a progressive 6-hydroxydopamine-induced degenerationmodel of Parkinson’s disease in rats. Proc. Natl. Acad. Sci. U.S.A.94, 14083–14088.
MANDEL, R.J., RENDAHL, K.G., SPRATT, S.K., SNYDER, R.O.,COHEN, L.K., and LEFF, S.E. (1998). Characterization of intrastri-atal recombinant adeno-associated virus-mediated gene transfer ofhuman tyrosine hydroxylase and human GTP-cyclohydrolase I in arat model of Parkinson’s disease. J. Neurosci. 18, 4271–4284.
MANDEL, R.J., GAGE, F.H., CLEVENGER, D.G., SPRATT, S.K.,SNYDER, R.O., and LEFF, S.E. (1999a). Nerve growth factor ex-pressed in the medial septum following in vivo gene delivery usinga recombinant adeno-associated viral vector protects cholinergic neu-rons from fimbria–fornix lesion-induced degeneration. Exp. Neurol.155, 59–64.
MANDEL, R.J., RENDAHL, K.G., SNYDER, R.O., and LEFF, S.E.(1999b). Progress in direct striatal delivery of L-dopa via gene ther-apy for treatment of Parkinson’s disease using recombinant adeno-associated viral vectors. Exp. Neurol. 159, 47–64.
MARTINOV, V.N., SEFLAND, I., WALAAS, S.I., LOMO, T., NJA,A., and HOOVER, F. (2002). Targeting functional subtypes of spinalmotoneurons and skeletal muscle fibers in vivo by intramuscular in-jection of adenoviral and adeno-associated viral vectors. Anat. Em-bryol. (Berl.) 205, 215–221.
MASTAKOV, M.Y., BAER, K., XU, R., FITZSIMONS, H., and DUR-ING, M.J. (2001). Combined injection of rAAV with mannitol en-hances gene expression in the rat brain. Mol. Ther. 3, 225–232.
MASTAKOV, M.Y., BAER, K., KOTIN, R.M., and DURING, M.J.(2002). Recombinant adeno-associated virus serotypes 2- and 5-me-diated gene transfer in the mammalian brain: Quantitative analysisof heparin co-infusion. Mol. Ther. 5, 371–380.
MATALON, R., SURENDRAN, S., RADY, P.L., QUAST, M.J.,CAMPBELL, G.A., MATALON, K.M., TYRING, S.K., WEI, J., PE-DEN, C.S., EZELL, E.L., MUZYCZKA, N., and MANDEL, R.J.(2003). Adeno-associated virus-mediated aspartoacylase gene trans-
fer to the brain of knockout mouse for Canavan disease. Mol. Ther.7, 580–587.
MCCOWN, T.J., XIAO, X., LI, J., BREESE, G.R., and SAMULSKI,R.J. (1996). Differential and persistent expression patterns of CNSgene transfer by an adeno-associated virus (AAV) vector. Brain Res.713, 99–107.
MURAMATSU, S., FUJIMOTO, K., IKEGUCHI, K., SHIZUMA, N.,KAWASAKI, K., ONO, F., SHEN, Y., WANG, L., MIZUKAMI,H., KUME, A., MATSUMURA, M., NAGATSU, I., URANO, F.,ICHINOSE, H., NAGATSU, T., TERAO, K., NAKANO, I., andOZAWA, K. (2002). Behavioral recovery in a primate model ofParkinson’s disease by triple transduction of striatal cells with adeno-associated viral vectors expressing dopamine-synthesizing enzymes.Hum. Gene Ther. 13, 345–354.
MUZYCZKA, N., and BERNS, K.I. (2001). Parvoviridae: The virusesand their replication. In Fields Virology. D.M. Knipe and Howley, P.M.,eds. (Lippincott, Williams & Wilkins, New York) pp. 2327–2360.
NARFSTROM, K., KATZ, M.L., BRAGADOTTIR, R., SEELIGER,M., BOULANGER, A., REDMOND, T.M., CARO, L., LAI, C.M.,and RAKOCZY, P.E. (2003). Functional and structural recovery ofthe retina after gene therapy in the RPE65 null mutation dog. Invest.Ophthalmol. Vis. Sci. 44, 1663–1672.
NGUYEN, J.B., SANCHEZ-PERNAUTE, R., CUNNINGHAM, J.,and BANKIEWICZ, K.S. (2001). Convection-enhanced delivery ofAAV-2 combined with heparin increases TK gene transfer in the ratbrain. Neuroreport 12, 1961–1964.
NIWA, H., YAMAMURA, K., and MIYAZAKI, J. (1991). Efficientselection for high-expression transfectants with a novel eukaryoticvector. Gene 108, 193–199.
NO, D., YAO, T.P., and EVANS, R.M. (1996). Ecdysone-induciblegene expression in mammalian cells and transgenic mice. Proc. Natl.Acad. Sci. U.S.A. 93, 3346–3351.
OPIE, S.R., WARRINGTON JR, K.H., JR., AGBANDJE-MCKENNA,M., ZOLOTUKHIN, S., and MUZYCZKA, N. (2003). Identificationof amino acid residues in the capsid proteins of adeno-associatedvirus type 2 that contribute to heparan sulfate proteoglycan binding.J. Virol. 77, 6995–7006.
OWEN, R.T., MANDEL, R.J., AMMINI, C.V., CONLON, T.J., KERR,D.S., STACPOOLE, P.W., and FLOTTE, T.R. (2002). Gene therapyfor pyruvate dehydrogenase E1� deficiency using recombinantadeno-associated virus 2 (rAAV2) vectors. Mol. Ther. 6, 394–399.
PASSINI, M.A., LEE, E.B., HEUER, G.G., and WOLFE, J.H. (2002).Distribution of a lysosomal enzyme in the adult brain by axonal trans-port and by cells of the rostral migratory stream. J. Neurosci. 22,6437–6446.
PASSINI, M.A., WATSON, D.J., VITE, C.H., LANDSBURG, D.J.,FEIGENBAUM, A.L., and WOLFE, J.H. (2003). Intraventricularbrain injection of adeno-associated virus type 1 (AAV1) in neonatalmice results in complementary patterns of neuronal transduction toAAV2 and total long-term correction of storage lesions in the Brainsof �-glucuronidase-deficient mice. J. Virol. 77, 7034–7040.
PATERNA, J.C., MOCCETTI, T., MURA, A., FELDON, J., andBUELER, H. (2000). Influence of promoter and WHV post-tran-scriptional regulatory element on AAV-mediated transgene expres-sion in the rat brain. Gene Ther. 7, 1304–1311.
PATERNA, J.C., FELDON, J., and BUELER, H. (2004). Transductionprofiles of recombinant adeno-associated virus vectors derived fromserotypes 2 and 5 in the nigrostriatal system of rats. J. Virol. 78,6808–6817.
PAXINOS, G., and WATSON, C. (1997). The Rat Brain in StereotaxicCoordinates. (Academic Press, Orlando, FL).
PEDEN, C.S., BURGER, C., MUZYCZKA, N., and MANDEL, R.J.(2004). Circulating anti-wild-type adeno-associated virus type 2(AAV2) antibodies inhibit recombinant AAV2 (rAAV2)-mediated,but not rAAV5-mediated, gene transfer in the brain. J. Virol. 78,6344–6359.
RECOMBINANT AAV VECTORS IN THE NERVOUS SYSTEM 789

PEEL, A.L., ZOLOTUKHIN, S., SCHRIMSHER, G.W., MUZY-CZKA, N., and REIER, P.J. (1997). Efficient transduction of greenfluorescent protein in spinal cord neurons using adeno-associatedvirus vectors containing cell type-specific promoters. Gene Ther. 4,16–24.
PROSCH, S., STEIN, J., STAAK, K., LIEBENTHAL, C., VOLK,H.D., and KRUGER, D.H. (1996). Inactivation of the very strongHCMV immediate early promoter by DNA CpG methylation in vitro.Biol. Chem. Hoppe Seyler 377, 195–201.
QING, K., MAH, C., HANSEN, J., ZHOU, S., DWARKI, V., and SRI-VASTAVA, A. (1999). Human fibroblast growth factor receptor 1is a co-receptor for infection by adeno-associated virus 2. Nat. Med.5, 71–77.
RICHICHI, C., LIN, E.J., STEFANIN, D., COLELLA, D., RAVIZZA,T., GRIGNASCHI, G., VEGLIANESE, P., SPERK, G., DURING,M.J., and VEZZANI, A. (2004). Anticonvulsant and antiepilepto-genic effects mediated by adeno-associated virus vector neuropep-tide Y expression in the rat hippocampus. J. Neurosci. 24,3051–3059.
RIVERA, V.M., CLACKSON, T., NATESAN, S., POLLOCK, R.,AMARA, J.F., KEENAN, T., MAGARI, S.R., PHILLIPS, T.,COURAGE, N.L., CERASOLI, F., Jr., HOLT, D.A., and GILMAN,M. (1996). A humanized system for pharmacologic control of geneexpression. Nat. Med. 2, 1028–1032.
RIVERA, V.M., YE, X., COURAGE, N.L., SACHAR, J., CERASOLI,F., Jr., WILSON, J.M., and GILMAN, M. (1999). Long-term regu-lated expression of growth hormone in mice after intramuscular genetransfer. Proc. Natl. Acad. Sci. U.S.A. 96, 8657–8662.
RUITENBERG, M.J., BLITS, B., DIJKHUIZEN, P.A., TE BEEK,E.T., BAKKER, A., VAN HEERIKHUIZE, J.J., POOL, C.W., HER-MENS, W.T., BOER, G.J., and VERHAAGEN, J. (2004). Adeno-associated viral vector-mediated gene transfer of brain-derived neu-rotrophic factor reverses atrophy of rubrospinal neurons followingboth acute and chronic spinal cord injury. Neurobiol. Dis. 15,394–406.
SAMAKOGLU, S., BOHL, D., and HEARD, J.M. (2002). Mechanismsleading to sustained reversion of �-thalassemia in mice by doxycy-cline-controlled Epo delivery from muscles. Mol. Ther. 6, 793–803.
SANFTNER, L.M., SUZUKI, B.M., DOROUDCHI, M.M., FENG, L.,MCCLELLAND, A., FORSAYETH, J.R., and CUNNINGHAM, J.(2004). Striatal delivery of rAAV-hAADC to rats with preexistingimmunity to AAV. Mol. Ther. 9, 403–409.
SCAMMELL, T.E., ARRIGONI, E., THOMPSON, M.A., RONAN,P.J., SAPER, C.B., and GREENE, R.W. (2003). Focal deletion ofthe adenosine A1 receptor in adult mice using an adeno-associatedviral vector. J. Neurosci. 23, 5762–5770.
SHEN, Y., MURAMATSU, S.I., IKEGUCHI, K., FUJIMOTO, K.I.,FAN, D.S., OGAWA, M., MIZUKAMI, H., URABE, M., KUME,A., NAGATSU, I., URANO, F., SUZUKI, T., ICHINOSE, H., NA-GATSU, T., MONAHAN, J., NAKANO, I., and OZAWA, K. (2000).Triple transduction with adeno-associated virus vectors expressingtyrosine hydroxylase, aromatic-L-amino-acid decarboxylase, andGTP cyclohydrolase I for gene therapy of Parkinson’s disease. Hum.Gene Ther. 11, 1509–1519.
SHIMAZAKI, K., URABE, M., MONAHAN, J., OZAWA, K., andKAWAI, N. (2000). Adeno-associated virus vector-mediated bcl-2gene transfer into post-ischemic gerbil brain in vivo: Prospects forgene therapy of ischemia-induced neuronal death. Gene Ther. 7,1244–1249.
SMITH, R.H., DING, C., and KOTIN, R.M. (2003). Serum-free pro-duction and column purification of adeno-associated virus type 5. J.Virol. Methods 114, 115–124.
SUMMERFORD, C., and SAMULSKI, R.J. (1998). Membrane-asso-ciated heparan sulfate proteoglycan is a receptor for adeno-associ-ated virus type 2 virions. J. Virol. 72, 1438–1445.
SUMMERFORD, C., BARTLETT, J.S., and SAMULSKI, R.J. (1999).
�V�5 integrin: A co-receptor for adeno-associated virus type 2 in-fection. Nat. Med. 5, 78–82.
SZCZYPKA, M.S., MANDEL, R.J., DONAHUE, B.A., SNYDER,R.O., LEFF, S.E., and PALMITER, R.D. (1999). Viral gene deliv-ery selectively restores feeding and prevents lethality of dopamine-deficient mice. Neuron 22, 167–178.
TENENBAUM, L., JURYSTA, F., STATHOPOULOS, A., PUSCHBAN,Z., MELAS, C., HERMENS, W.T., VERHAAGEN, J., PICHON, B.,VELU, T., and LEVIVIER, M. (2000). Tropism of AAV-2 vectorsfor neurons of the globus pallidus. Neuroreport 11, 2277–2283.
THOMAS, C.E., STORM, T.A., HUANG, Z., and KAY, M.A. (2004).Rapid uncoating of vector genomes is the key to efficient liver trans-duction with pseudotyped adeno-associated virus vectors. J. Virol.78, 3110–3122.
URLINGER, S., BARON, U., THELLMANN, M., HASAN, M.T., BU-JARD, H., and HILLEN, W. (2000). Exploring the sequence spacefor tetracycline-dependent transcriptional activators: Novel muta-tions yield expanded range and sensitivity. Proc. Natl. Acad. Sci.U.S.A. 97, 7963–7968.
VITE, C.H., PASSINI, M.A., HASKINS, M.E., and WOLFE, J.H.(2003). Adeno-associated virus vector-mediated transduction in thecat brain. Gene Ther. 10, 1874–1881.
WALTERS, R.W., YI, S.M., KESHAVJEE, S., BROWN, K.E.,WELSH, M.J., CHIORINI, J.A., and ZABNER, J. (2001). Bindingof adeno-associated virus type 5 to 2,3-linked sialic acid is requiredfor gene transfer. J. Biol. Chem. 276, 20610–20616.
WANG, A.Y., PENG, P.D., EHRHARDT, A., STORM, T.A., andKAY, M.A. (2004). Comparison of adenoviral and adeno-associatedviral vectors for pancreatic gene delivery in vivo. Hum. Gene Ther.15, 405–413.
WANG, C., WANG, C.M., CLARK, K.R., and SFERRA, T.J. (2003).Recombinant AAV serotype 1 transduction efficiency and tropismin the murine brain. Gene Ther. 10, 1528–1534.
WANG, L., MURAMATSU, S., LU, Y., IKEGUCHI, K., FUJIMOTO,K., OKADA, T., MIZUKAMI, H., HANAZONO, Y., KUME, A.,URANO, F., ICHINOSE, H., NAGATSU, T., NAKANO, I., andOZAWA, K. (2002). Delayed delivery of AAV-GDNF prevents ni-gral neurodegeneration and promotes functional recovery in a ratmodel of Parkinson’s disease. Gene Ther. 9, 381–389.
WANG, L., CALCEDO, R., NICHOLS, T.C., BELLINGER, D.A.,DILLOW, A., VERMA, I.M., and WILSON, J.M. (2005). Sustainedcorrection of disease in naive and AAV2-pretreated hemophilia Bdogs: AAV2/8 mediated, liver-directed gene therapy. Blood 105,3079–3086.
WANG, Y., O’MALLEY, B.W., JR., TSAI, S.Y., and O’MALLEY,B.W. (1994). A regulatory system for use in gene transfer. Proc. Natl.Acad. Sci. U.S.A. 91, 8180–8184.
WARRINGTON, K.H., Jr., GORBATYUK, O.S., HARRISON, J.K.,OPIE, S.R., ZOLOTUKHIN, S., and MUZYCZKA, N. (2004).Adeno-associated virus type 2 VP2 capsid protein is nonessential andcan tolerate large peptide insertions at its N terminus. J. Virol. 78,6595–6609.
WEBER, M., RABINOWITZ, J., PROVOST, N., CONRATH, H.,FOLLIOT, S., BRIOT, D., CHEREL, Y., CHENUAUD, P.,SAMULSKI, J., MOULLIER, P., and ROLLING, F. (2003). Re-combinant adeno-associated virus serotype 4 mediates unique andexclusive long-term transduction of retinal pigmented epithelium inrat, dog, and nonhuman primate after subretinal delivery. Mol. Ther.7, 774–781.
WEST, M.J., SLOMIANKA, L., and GUNDERSEN, H.J. (1991). Un-biased stereological estimation of the total number of neurons in thesubdivisions of the rat hippocampus using the optical fractionator.Anat. Rec. 231, 482–497.
WU, P., XIAO, W., CONLON, T., HUGHES, J., AGBANDJE-MCKENNA, M., FERKOL, T., FLOTTE, T., and MUZYCZKA, N.(2000). Mutational analysis of the adeno-associated virus type 2
BURGER ET AL.790

(AAV2) capsid gene and construction of AAV2 vectors with alteredtropism. J. Virol. 74, 8635–8647.
XIE, Q., BU, W., BHATIA, S., HARE, J., SOMASUNDARAM, T.,AZZI, A., and CHAPMAN, M.S. (2002). The atomic structure ofadeno-associated virus (AAV-2), a vector for human gene therapy.Proc. Natl. Acad. Sci. U.S.A. 99, 10405–10410.
XU, Y., GU, Y., WU, P., LI, G.W., and HUANG, L.Y. (2003). Effi-ciencies of transgene expression in nociceptive neurons through dif-ferent routes of delivery of adeno-associated viral vectors. Hum.Gene Ther. 14, 897–906.
YANG, G.S., SCHMIDT, M., YAN, Z., LINDBLOOM, J.D., HARD-ING, T.C., DONAHUE, B.A., ENGELHARDT, J.F., KOTIN, R.,and DAVIDSON, B.L. (2002). Virus-mediated transduction ofmurine retina with adeno-associated virus: Effects of viral capsid andgenome size. J. Virol. 76, 7651–7660.
YE, X., RIVERA, V.M., ZOLTICK, P., CERASOLI, F., Jr.,SCHNELL, M.A., GAO, G., HUGHES, J.V., GILMAN, M., andWILSON, J.M. (1999). Regulated delivery of therapeutic proteinsafter in vivo somatic cell gene transfer. Science 283, 88–91.
ZOLOTUKHIN, S., POTTER, M., ZOLOTUKHIN, I., SAKAI, Y.,LOILER, S., FRAITES, T.J., Jr., CHIODO, V.A., PHILLIPSBERG,
T., MUZYCZKA, N., HAUSWIRTH, W.W., FLOTTE, T.R.,BYRNE, B.J., and SNYDER, R.O. (2002). Production and purifica-tion of serotype 1, 2, and 5 recombinant adeno-associated viral vec-tors. Methods 28, 158–167.
Address reprint requests to:Dr. Corinna Burger
Department of Molecular Genetics and Microbiologyand Powell Gene Therapy Center
College of MedicineP.O. Box 100266
University of FloridaGainesville, FL 32610
E-mail: [email protected]
Received for publication March 10, 2005; accepted March 10,2005.
Published online: May 11, 2005.
RECOMBINANT AAV VECTORS IN THE NERVOUS SYSTEM 791