
Low-frequency stimulation evokes serotonin release in the nucleus accumbens and induces long-term...
10
Low-frequency stimulation evokes serotonin release in the nucleus accumbens and induces long-term depression via production of endocannabinoid Costanza Burattini, 1 Giulia Battistini, 1 Francesco Tamagnini, 1 and Giorgio Aicardi 1,2 1 Department for Life Quality Studies, University of Bologna, Bologna, Italy; and 2 Interdepartmental Center “Luigi Galvani” for the Study of Biophysics, Bioinformatics and Biocomplexity, University of Bologna, Bologna, Italy Submitted 10 July 2013; accepted in final form 11 December 2013 Burattini C, Battistini G, Tamagnini F, Aicardi G. Low-fre- quency stimulation evokes serotonin release in the nucleus accumbens and induces long-term depression via production of endocannabinoid. J Neurophysiol 111: 1046 –1055, 2014. First published December 11, 2013; doi:10.1152/jn.00498.2013.—The nucleus accumbens (NAc), a major component of the mesolimbic system, is involved in the mediation of reinforcing and addictive properties of many depen- dence-producing drugs. Glutamatergic synapses within the NAc can express plasticity, including a form of endocannabinoid (eCB)-long- term depression (LTD). Recent evidences demonstrate cross talk between eCB signaling pathways and those of other receptor systems, including serotonin (5-HT); the extensive colocalization of CB1 and 5-HT receptors within the NAc suggests the potential for interplay between them. In the present study, we found that 20-min low- frequency (4 Hz) stimulation (LFS-4Hz) of glutamatergic afferences in rat brain slices induces a novel form of eCB-LTD in the NAc core, which requires 5-HT 2 and CB1 receptor activation and L-type volt- age-gated Ca 2 channel opening. Moreover, we found that exogenous 5-HT application (5 M, 20 min) induces an analogous LTD (5-HT- LTD) at the same synapses, requiring the activation of the same receptors and the opening of the same Ca 2 channels; LFS-4Hz-LTD and 5-HT-LTD were mutually occlusive. Present results suggest that LFS-4Hz induces the release of 5-HT, which acts at 5-HT 2 postsyn- aptic receptors, increasing Ca 2 influx through L-type voltage-gated channels and 2-arachidonoylglycerol production and release; the eCB travels retrogradely and binds to presynaptic CB1 receptors, causing a long-lasting decrease of glutamate release, resulting in LTD. These observations might be helpful to understand the neurophysiological mechanisms underlying drug addiction, major depression, and other psychiatric disorders characterized by dysfunction of 5-HT neu- rotransmission in the NAc. long-term depression; serotonin; endocannabinoids; nucleus accum- bens THE NUCLEUS ACCUMBENS (NAc), the ventral extension of the striatal complex, is a chief component of the mesolimbic dopamine (DA) system, receiving DAergic inputs from the ventral tegmental area (Le Moal and Simon 1991) and gluta- matergic afferents from cortical and sub cortical limbic struc- tures, such as the prefrontal cortex (PFC) (Christie et al. 1985), the amygdala, and the hippocampus (Brog et al. 1993; Pennartz et al. 1994), catecholaminergic afferents from the locus coer- uleus, and a dense serotonergic innervation from dorsal raphe nucleus neurons (Van Bockstaele and Pickel 1993). Behav- ioral, anatomic, and physiological studies have shown that the NAc is critically involved in reward and in the translation of motivation to action (Koob and Bloom 1988; Marty and Spigelman 2012; Mogenson et al. 1980), as well as in other functions, including feeding and pain, and in pathologies such as depression, schizophrenia, and attention-deficit and motor disorders (Altier and Stewart 1999; Belujon et al. 2012; Ohno et al. 2012; Salamone et al. 2005; Shirayama and Chaki 2006; Wickelgren 1997). As observed in other brain regions, glutamatergic synapses within the NAc can express plasticity: both long-term poten- tiation (LTP) and long-term depression (LTD) can be reliably elicited at these synapses (Ji and Martin 2012; Kombian and Malenka 1994; Pennartz et al. 1993). Recently, more attention has been focused on LTD in the NAc, due to experimental evidence of its possible role as a cellular correlate of complex behaviors, such as sensitization and addiction (see Marty and Spigelman 2012). Distinct forms of LTD at glutamatergic synapses have been observed, including endocannabinoid (eCB) receptor 1 (CB1)-mediated LTD induced by different stimulation protocols: 13 Hz/10 min (Robbe et al. 2002b), 5 Hz/10 min (Hoffman et al. 2003), or 10 Hz/5 min (Grueter et al. 2010). Anatomic and electrophysiological studies have shown that CB1 receptors are located presynaptically on glu- tamatergic neurons arising in the PFC and projecting to the NAc, and that they can participate in the regulation of trans- mitter release (Robbe et al. 2001). Induction of eCB-LTD by 13-Hz/10-min or 10-Hz/5-min stimulation requires the activa- tion of postsynaptic metabotropic glutamate receptor (mGluR) 5, which triggers the release of Ca 2 from the intracellular stores, finally resulting in the release of eCBs; eCBs are known to mainly act at presynaptic CB1 receptors, inhibiting the release of glutamate (Grueter et al. 2010; Robbe et al. 2002b). Data obtained with the 10-Hz/5-min protocol suggest that the eCB mainly involved in this process is anandamide (AEA), which also triggers the endocytosis of postsynaptic AMPA receptors via the activation of transient receptor potential vanilloid 1 (TRPV1) channels (Grueter et al. 2010). In recent years, compelling evidence has demonstrated cross talk between CB signaling pathways and those of other recep- tor systems, such as the DA D2 (Blume et al. 2013), -opioid and serotonin (5-HT) 2 receptors (Best and Regehr 2008). The extensive colocalization of CB1 and 5-HT receptors within the NAc suggests the potential for interplay between the signaling cascades activated by these two receptors, which belong to the 7-transmembrane G protein-coupled receptors superfamily (Barnes and Sharp 1999; Pertwee 1997). Interestingly, 5-HT 2 receptor activation strongly suppresses excitatory postsynaptic currents in the inferior olive by releasing eCBs that suppress glutamate release through presynaptic CB1 receptor activation (Best and Regehr 2008). Address for reprint requests and other correspondence: C. Burattini, Dept. for Life Quality Studies, Univ. of Bologna, Via San Donato 19/2, 40127 Bologna, Italy (e-mail: [email protected]). J Neurophysiol 111: 1046 –1055, 2014. First published December 11, 2013; doi:10.1152/jn.00498.2013. 1046 0022-3077/14 Copyright © 2014 the American Physiological Society www.jn.org
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Low-frequency stimulation evokes serotonin release in the nucleus accumbens and induces long-term...

Low-frequency stimulation evokes serotonin release in the nucleus accumbensand induces long-term depression via production of endocannabinoid
Costanza Burattini,1 Giulia Battistini,1 Francesco Tamagnini,1 and Giorgio Aicardi1,2
1Department for Life Quality Studies, University of Bologna, Bologna, Italy; and 2Interdepartmental Center “Luigi Galvani”for the Study of Biophysics, Bioinformatics and Biocomplexity, University of Bologna, Bologna, Italy
Submitted 10 July 2013; accepted in final form 11 December 2013
Burattini C, Battistini G, Tamagnini F, Aicardi G. Low-fre-quency stimulation evokes serotonin release in the nucleus accumbensand induces long-term depression via production of endocannabinoid.J Neurophysiol 111: 1046–1055, 2014. First published December 11,2013; doi:10.1152/jn.00498.2013.—The nucleus accumbens (NAc), amajor component of the mesolimbic system, is involved in themediation of reinforcing and addictive properties of many depen-dence-producing drugs. Glutamatergic synapses within the NAc canexpress plasticity, including a form of endocannabinoid (eCB)-long-term depression (LTD). Recent evidences demonstrate cross talkbetween eCB signaling pathways and those of other receptor systems,including serotonin (5-HT); the extensive colocalization of CB1 and5-HT receptors within the NAc suggests the potential for interplaybetween them. In the present study, we found that 20-min low-frequency (4 Hz) stimulation (LFS-4Hz) of glutamatergic afferencesin rat brain slices induces a novel form of eCB-LTD in the NAc core,which requires 5-HT2 and CB1 receptor activation and L-type volt-age-gated Ca2� channel opening. Moreover, we found that exogenous5-HT application (5 �M, 20 min) induces an analogous LTD (5-HT-LTD) at the same synapses, requiring the activation of the samereceptors and the opening of the same Ca2� channels; LFS-4Hz-LTDand 5-HT-LTD were mutually occlusive. Present results suggest thatLFS-4Hz induces the release of 5-HT, which acts at 5-HT2 postsyn-aptic receptors, increasing Ca2� influx through L-type voltage-gatedchannels and 2-arachidonoylglycerol production and release; the eCBtravels retrogradely and binds to presynaptic CB1 receptors, causinga long-lasting decrease of glutamate release, resulting in LTD. Theseobservations might be helpful to understand the neurophysiologicalmechanisms underlying drug addiction, major depression, and otherpsychiatric disorders characterized by dysfunction of 5-HT neu-rotransmission in the NAc.
long-term depression; serotonin; endocannabinoids; nucleus accum-bens
THE NUCLEUS ACCUMBENS (NAc), the ventral extension of thestriatal complex, is a chief component of the mesolimbicdopamine (DA) system, receiving DAergic inputs from theventral tegmental area (Le Moal and Simon 1991) and gluta-matergic afferents from cortical and sub cortical limbic struc-tures, such as the prefrontal cortex (PFC) (Christie et al. 1985),the amygdala, and the hippocampus (Brog et al. 1993; Pennartzet al. 1994), catecholaminergic afferents from the locus coer-uleus, and a dense serotonergic innervation from dorsal raphenucleus neurons (Van Bockstaele and Pickel 1993). Behav-ioral, anatomic, and physiological studies have shown that theNAc is critically involved in reward and in the translation ofmotivation to action (Koob and Bloom 1988; Marty and
Spigelman 2012; Mogenson et al. 1980), as well as in otherfunctions, including feeding and pain, and in pathologies suchas depression, schizophrenia, and attention-deficit and motordisorders (Altier and Stewart 1999; Belujon et al. 2012; Ohnoet al. 2012; Salamone et al. 2005; Shirayama and Chaki 2006;Wickelgren 1997).
As observed in other brain regions, glutamatergic synapseswithin the NAc can express plasticity: both long-term poten-tiation (LTP) and long-term depression (LTD) can be reliablyelicited at these synapses (Ji and Martin 2012; Kombian andMalenka 1994; Pennartz et al. 1993). Recently, more attentionhas been focused on LTD in the NAc, due to experimentalevidence of its possible role as a cellular correlate of complexbehaviors, such as sensitization and addiction (see Marty andSpigelman 2012). Distinct forms of LTD at glutamatergicsynapses have been observed, including endocannabinoid(eCB) receptor 1 (CB1)-mediated LTD induced by differentstimulation protocols: 13 Hz/10 min (Robbe et al. 2002b), 5Hz/10 min (Hoffman et al. 2003), or 10 Hz/5 min (Grueter etal. 2010). Anatomic and electrophysiological studies haveshown that CB1 receptors are located presynaptically on glu-tamatergic neurons arising in the PFC and projecting to theNAc, and that they can participate in the regulation of trans-mitter release (Robbe et al. 2001). Induction of eCB-LTD by13-Hz/10-min or 10-Hz/5-min stimulation requires the activa-tion of postsynaptic metabotropic glutamate receptor (mGluR)5, which triggers the release of Ca2� from the intracellularstores, finally resulting in the release of eCBs; eCBs are knownto mainly act at presynaptic CB1 receptors, inhibiting therelease of glutamate (Grueter et al. 2010; Robbe et al. 2002b).Data obtained with the 10-Hz/5-min protocol suggest that theeCB mainly involved in this process is anandamide (AEA),which also triggers the endocytosis of postsynaptic AMPAreceptors via the activation of transient receptor potentialvanilloid 1 (TRPV1) channels (Grueter et al. 2010).
In recent years, compelling evidence has demonstrated crosstalk between CB signaling pathways and those of other recep-tor systems, such as the DA D2 (Blume et al. 2013), �-opioidand serotonin (5-HT) 2 receptors (Best and Regehr 2008). Theextensive colocalization of CB1 and 5-HT receptors within theNAc suggests the potential for interplay between the signalingcascades activated by these two receptors, which belong to the7-transmembrane G protein-coupled receptors superfamily(Barnes and Sharp 1999; Pertwee 1997). Interestingly, 5-HT2receptor activation strongly suppresses excitatory postsynapticcurrents in the inferior olive by releasing eCBs that suppressglutamate release through presynaptic CB1 receptor activation(Best and Regehr 2008).
Address for reprint requests and other correspondence: C. Burattini, Dept.for Life Quality Studies, Univ. of Bologna, Via San Donato 19/2, 40127Bologna, Italy (e-mail: [email protected]).
J Neurophysiol 111: 1046–1055, 2014.First published December 11, 2013; doi:10.1152/jn.00498.2013.
1046 0022-3077/14 Copyright © 2014 the American Physiological Society www.jn.org

In the present study, performed in a rat slice preparationcontaining the NAc, we found that prolonged (20 min) low-frequency (4 Hz) stimulation (LFS-4Hz) of glutamatergic af-ferences induces a novel form of eCB-LTD in the NAc core,which requires 5-HT2 and CB1 receptor activation and L-typevoltage-gated Ca2� channel (VGCC) opening.
MATERIALS AND METHODS
Animals. Young (21- to 38-day-old) male Sprague-Dawley CD IGSrats (Charles River, Calco, Italy) were used in this study. Animalswere treated in accordance with the European Community guidelineson animal care, and the experimental protocols were approved by theEthical Committee of the University of Bologna. Rats were individ-ually housed under controlled conditions (temperature: 24 � 1°C;humidity: 50 � 5%), maintained on a 12:12-h light-dark cycle, andfed ad libitum.
Slice preparation. The experiments were carried out in parasagittalbrain slices including the NAc. The slices were prepared as previouslydescribed by Li and Kauer (2004). Briefly, rats were deeply anaes-thetized using halothane (Sigma-Aldrich, Milan, Italy) and quicklydecapitated. Their brains were rapidly removed and immerged inice-cold artificial cerebrospinal fluid (aCSF) containing (in mM) 119NaCl, 2.5 KCl, 1 NaH2PO4, 26 NaHCO3, 1.3 MgSO4, 2.5 CaCl2, 10glucose, and 1 kynurenic acid, saturated with 95% O2-5% CO2 (295mOsm/l, pH 7.4). Parasagittal midbrain slices (400 �m thick) wereobtained from the blocked brain with the use of a vibrating microtomeand then transferred to a tissue bath containing aCSF withoutkynurenic acid at room temperature until use.
Electrophysiological recordings. After recovering for at least 60min, a single slice was transferred to a submersion recording chamberand perfused at the rate of 2 ml/min with aCSF without kynurenic acidmaintained at room temperature, saturated with 95% O2-5% CO2, andadded with 50 �M picrotoxin (Sigma-Aldrich) to block GABAA
receptors. Recordings started 30 min after the slice was placed into thechamber. Field excitatory postsynaptic potentials (fEPSPs) evoked byglutamatergic afferent pathway stimulation were recorded in the NAccore with an extracellular microelectrode (glass micropipette filledwith aCSF, 2–8 M�) connected to a DC amplifier by an Ag-AgClelectrode. Constant-current square pulses (0.2 ms, 40–120 �A, 0.033Hz) were applied using the stimulus generator Master 8 (AMPI,Jerusalem, Israel), connected through a stimulus isolation unit to aconcentric bipolar electrode (70–80 k�) positioned at the rostral edgeof the NAc, bordering the cortex, just below the anterior commissureat �400–600 �m from the recording electrode in the rostral direction.Stimulus intensity was adjusted to induce �50% of the maximalsynaptic response; after this adjustment, the fEPSP average size was0.6 � 0.06 mV. After stable baseline recording, LTD was induced byLFS-4Hz, consisting of a single train of 4,800 pulses delivered at 4 Hzin 20 min at test stimulus intensity. A weaker stimulation protocol wasalso used, consisting of 2,400 pulses at 4 Hz (LFS-weak) delivered in10 min at test stimulus intensity.
Drug application. Drugs were applied by switching from the controlsolution to one that differed only for the presence of the added drug. Thefollowing pharmacological compounds were applied in this study: theGABAA receptors blocker picrotoxin (50 �M), the L-type VGCCblocker nifedipine hydrochloride (10 �M), and the ionotropic glutamatereceptor antagonist kynurenic acid (1 mM) were purchased from Sigma-Aldrich; the N-methyl-D-aspartate receptor (NMDAR) antagonist D-amino-5-phosphonovaleric acid (D-AP5), the CB1 receptor antagonists N-(pip-eridin-1-yl)-5-(4-iodophenyl)-1-(2,4-dichlorophenyl)-4-methyl-1H-pyrazole-3-carboxamide (AM251; 2 �M) and 4-[[6-methoxy-2-(4-methoxyphenyl)-3-benzofuranyl]carbonyl]benzonitrile (LY-320135; 10�M), the mGluR1 antagonist (S)-(�)-�-amino-4-carboxy-2-methyl-benzeneacetic acid (LY-367385), the mGluR5 antagonist 2-methyl-6-(phenylethynyl)-pyridine (MPEP), the mGluR2/3 antagonist 2-amino-2-(2-carboxycyclopropan-1-yl)-3-(dibenzopyran-4-yl) propanoic acid (LY-
341495), 5-hydroxytryptamine hydrobromide (5-HT; 5 �M), the 5-HT2
receptor antagonist ketanserin tartrate (10 �M), the 5-HT1 receptor an-tagonist pindolol (10 �M), the TRPV1 antagonist capsazepine (10 �M),the D1 receptor antagonist SCH-23390 (25 �M), the D2 receptor antag-onist sulpiride (25 �M), the diacylglycerol lipase � (DGL) inhibitortetrahydrolipstatin (THL; 10 �M), the fatty acid amide hydrolase(FAAH) inhibitor JNJ-1661010 (1 �M), the selective 5-HT reuptakeinhibitor (SSRI) citalopram hydrobromide (4 �M), the 5-HT2 selectiveagonist �-methyl-5-hydroxytryptamine maleate (�-methyl 5-HT; 20�M), and the selective monoacylglycerol lipase (MAGL) inhibitor4-[bis(1,3-benzodioxol-5-yl)hydroxyl-methyl]-1-piperidinecarboxylicacid 4-nitrophenyl ester (JZL-184; 50 �M) were purchased from TocrisCookson (Bristol, UK). Drugs were made up in dimethyl sulfoxide(DMSO) or bidistilled water stock solutions and diluted 1,000 times intothe superfusing aCSF, yielding their final concentrations. In a subset ofcontrol experiments (4 slices), 0.1% DMSO was also added to theslice-perfusing aCSF; LFS-4Hz elicited normal LTD in each of theseslices, and therefore data obtained were pooled with those obtained in thecontrol slices in all following experiments and defined as “control.” Withthe exception of kynureate, 5-HT, and �-methyl 5-HT, none of the drugsused affected basal synaptic transmission.
Data analysis. Sweeps of 1,000 ms comprising the artifact, thenonsynaptic components, and the fEPSP were digitally acquired usingthe software Axoscope 9.0 (Axon Instruments, Downington, PA) andanalyzed off-line using the software Clampfit (Axon Instruments); thepeak amplitude values (mV) were exported in an Excel file. Eachamplitude value was normalized to the baseline (calculated as themean of the fEPSP amplitudes recorded in the last 10 min beforeapplication of LFS-4Hz). All data are plotted as means � SE. In eachexperimental condition, statistical comparison was made betweennormalized (%) amplitudes (means � SE) of fEPSPs recorded duringthe last 5 min of baseline and in the time points of interest (e.g., mean55–60 min after the end of LFS-4Hz); paired Student’s t-test was usedfor binary comparison (e.g., baseline vs. follow-up), whereas one-wayrepeated-measures analysis of variance (ANOVA) with Bonferronipost hoc analysis was used for comparing multiple time points of theexperiments compared with baseline (e.g., follow-up after 2nd stim-ulation vs. follow-up after 1st stimulation vs. baseline). One-wayANOVA with Bonferroni post hoc analysis was used for statisticalcomparison between normalized (%) amplitude (means � SE) offEPSP recorded 55–60 min after the end of LFS-4Hz in controlconditions and in the presence of different drug treatments. Differ-ences were considered significant for P � 0.05.
RESULTS
We recorded fEPSPs evoked in the NAc core by the stim-ulation of glutamatergic afferences. We found that LFS-4Hz(20-min stimulation at 4 Hz) induces a robust LTD (n � 8; Fig. 1, Aand D; Table 1): when the recording was protracted for 180min after the end of LFS-4Hz stimulation, the mean of fEPSPamplitudes recorded between 175 and 180 min was 74.87%, avalue still significantly (P � 0.05) different from baseline (n �3; data not shown).
We first tested whether this novel form of LTD requiredactivation of the glutamate NMDAR, as previously demon-strated in other forms of LTD in the NAc (Grueter et al. 2010;Thomas et al. 2000). Thus the competitive NMDAR antagonistD-AP5 (50 �M) was added to the slice-perfusing medium 20min before and during LFS-4Hz stimulation: this treatment didnot affect LTD (n � 6; Fig. 1D; Table 1). To evaluate theinvolvement of postsynaptic ionotropic glutamate receptors,the slices were perfused with the non-subtype-selective iono-tropic glutamate receptor antagonist kynureate (1 mM) 20 minbefore and during LFS-4Hz stimulation. As expected, kynure-
1047SEROTONIN MEDIATES ECB-LTD IN THE NAC
J Neurophysiol • doi:10.1152/jn.00498.2013 • www.jn.org
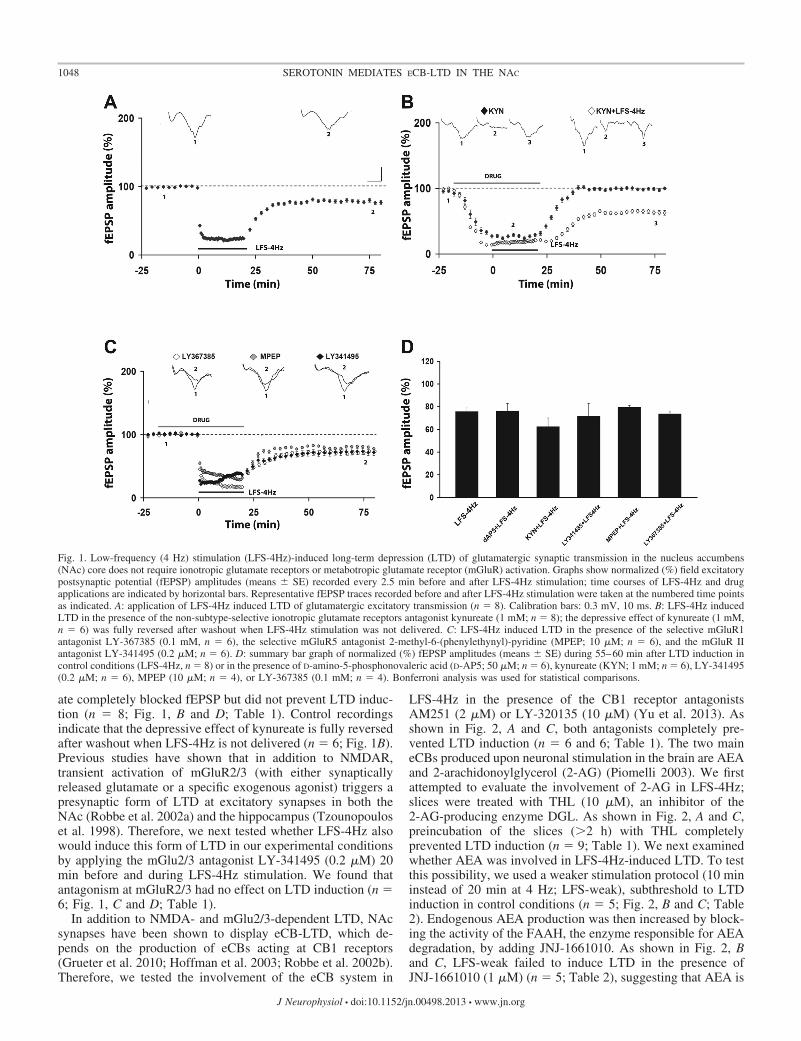
ate completely blocked fEPSP but did not prevent LTD induc-tion (n � 8; Fig. 1, B and D; Table 1). Control recordingsindicate that the depressive effect of kynureate is fully reversedafter washout when LFS-4Hz is not delivered (n � 6; Fig. 1B).Previous studies have shown that in addition to NMDAR,transient activation of mGluR2/3 (with either synapticallyreleased glutamate or a specific exogenous agonist) triggers apresynaptic form of LTD at excitatory synapses in both theNAc (Robbe et al. 2002a) and the hippocampus (Tzounopouloset al. 1998). Therefore, we next tested whether LFS-4Hz alsowould induce this form of LTD in our experimental conditionsby applying the mGlu2/3 antagonist LY-341495 (0.2 �M) 20min before and during LFS-4Hz stimulation. We found thatantagonism at mGluR2/3 had no effect on LTD induction (n �6; Fig. 1, C and D; Table 1).
In addition to NMDA- and mGlu2/3-dependent LTD, NAcsynapses have been shown to display eCB-LTD, which de-pends on the production of eCBs acting at CB1 receptors(Grueter et al. 2010; Hoffman et al. 2003; Robbe et al. 2002b).Therefore, we tested the involvement of the eCB system in
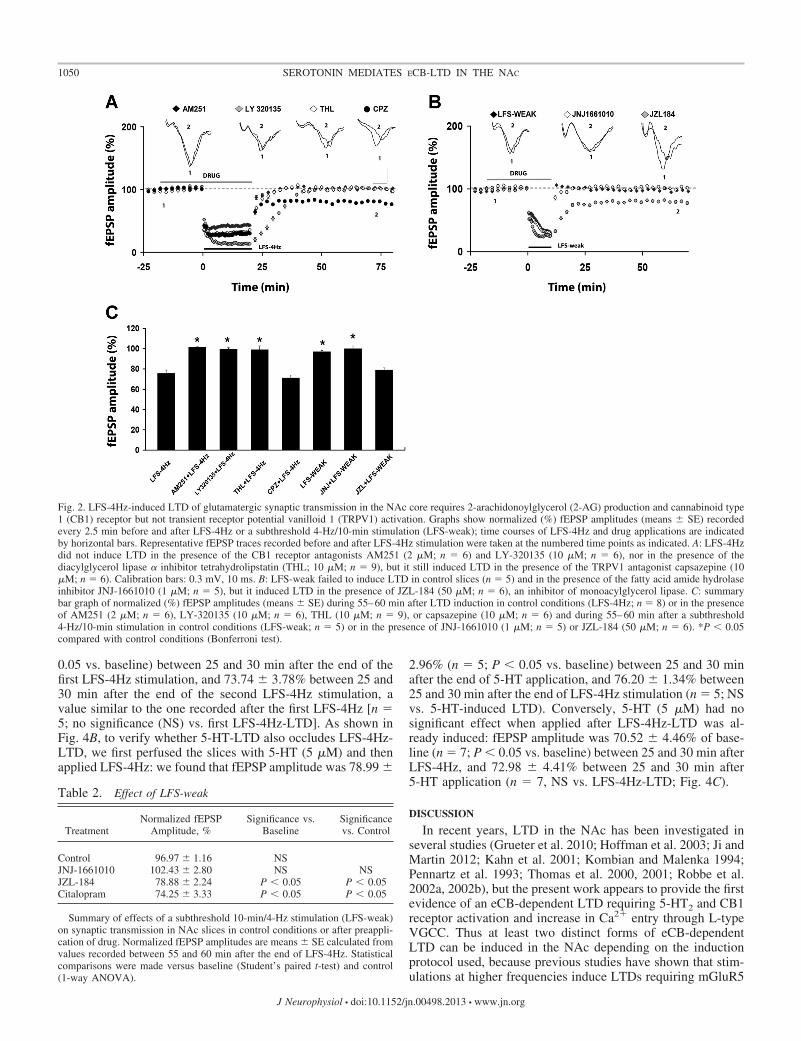
LFS-4Hz in the presence of the CB1 receptor antagonistsAM251 (2 �M) or LY-320135 (10 �M) (Yu et al. 2013). Asshown in Fig. 2, A and C, both antagonists completely pre-vented LTD induction (n � 6 and 6; Table 1). The two maineCBs produced upon neuronal stimulation in the brain are AEAand 2-arachidonoylglycerol (2-AG) (Piomelli 2003). We firstattempted to evaluate the involvement of 2-AG in LFS-4Hz;slices were treated with THL (10 �M), an inhibitor of the2-AG-producing enzyme DGL. As shown in Fig. 2, A and C,preincubation of the slices (�2 h) with THL completelyprevented LTD induction (n � 9; Table 1). We next examinedwhether AEA was involved in LFS-4Hz-induced LTD. To testthis possibility, we used a weaker stimulation protocol (10 mininstead of 20 min at 4 Hz; LFS-weak), subthreshold to LTDinduction in control conditions (n � 5; Fig. 2, B and C; Table2). Endogenous AEA production was then increased by block-ing the activity of the FAAH, the enzyme responsible for AEAdegradation, by adding JNJ-1661010. As shown in Fig. 2, Band C, LFS-weak failed to induce LTD in the presence ofJNJ-1661010 (1 �M) (n � 5; Table 2), suggesting that AEA is
Fig. 1. Low-frequency (4 Hz) stimulation (LFS-4Hz)-induced long-term depression (LTD) of glutamatergic synaptic transmission in the nucleus accumbens(NAc) core does not require ionotropic glutamate receptors or metabotropic glutamate receptor (mGluR) activation. Graphs show normalized (%) field excitatorypostsynaptic potential (fEPSP) amplitudes (means � SE) recorded every 2.5 min before and after LFS-4Hz stimulation; time courses of LFS-4Hz and drugapplications are indicated by horizontal bars. Representative fEPSP traces recorded before and after LFS-4Hz stimulation were taken at the numbered time pointsas indicated. A: application of LFS-4Hz induced LTD of glutamatergic excitatory transmission (n � 8). Calibration bars: 0.3 mV, 10 ms. B: LFS-4Hz inducedLTD in the presence of the non-subtype-selective ionotropic glutamate receptors antagonist kynureate (1 mM; n � 8); the depressive effect of kynureate (1 mM,n � 6) was fully reversed after washout when LFS-4Hz stimulation was not delivered. C: LFS-4Hz induced LTD in the presence of the selective mGluR1antagonist LY-367385 (0.1 mM, n � 6), the selective mGluR5 antagonist 2-methyl-6-(phenylethynyl)-pyridine (MPEP; 10 �M; n � 6), and the mGluR IIantagonist LY-341495 (0.2 �M; n � 6). D: summary bar graph of normalized (%) fEPSP amplitudes (means � SE) during 55–60 min after LTD induction incontrol conditions (LFS-4Hz, n � 8) or in the presence of D-amino-5-phosphonovaleric acid (D-AP5; 50 �M; n � 6), kynureate (KYN; 1 mM; n � 6), LY-341495(0.2 �M; n � 6), MPEP (10 �M; n � 4), or LY-367385 (0.1 mM; n � 4). Bonferroni analysis was used for statistical comparisons.
1048 SEROTONIN MEDIATES ECB-LTD IN THE NAC
J Neurophysiol • doi:10.1152/jn.00498.2013 • www.jn.org

not involved in this form of LTD. To further test the involve-ment of 2-AG in LFS-4Hz-induced LTD, slices were incubated(�2 h) with JZL-184 (50 �M) (Puente et al. 2011), an inhibitorof MAGL, the enzyme responsible for 2-AG degradation, andLFS-weak was applied. As shown in Fig. 2, B and C, elevationof 2-AG levels rescued LTD, confirming the involvement of2-AG in LFS-4Hz-induced LTD (n � 6; Table 2).
A recent study suggested that LTD induced in the NAc coreby 10-Hz/5-min stimulation involves TRPV1 activation (Gru-eter et al. 2010); therefore, we performed a series of experi-ments in the presence of the TRPV1 antagonist capsazepine(10 �M) and found that capsazepine did not affect LTDinduced by LFS-4Hz (n � 6; Fig. 2, A and C; Table 1).Moreover, we performed a series of experiments using the10-Hz/5-min stimulation protocol and found that, in our ex-perimental conditions, this protocol does not induce LTD(mean fEPSP was 106.52 � 4.67% at 55–60 min after 10-Hz/5-min LFS; n � 6; data not shown).
Because it has been shown that eCB-LTD in the NAcrequires activation of mGluR5 (Robbe et al. 2002b), we alsotested its involvement in the LTD obtained in our experimentalconditions: application of the mGluR5 antagonist MPEP (10�M) during LFS-4Hz did not affect LTD induction (n � 4;Fig. 1, C and D; Table 1). We further investigated the involve-ment of group I mGluRs (which comprise both mGluR1 andmGluR5) by applying the mGluR1 antagonist LY-367385 (0.1mM). Figure 1, C and D, shows that LFS-4Hz still elicitedLTD (n � 4; Table 1).
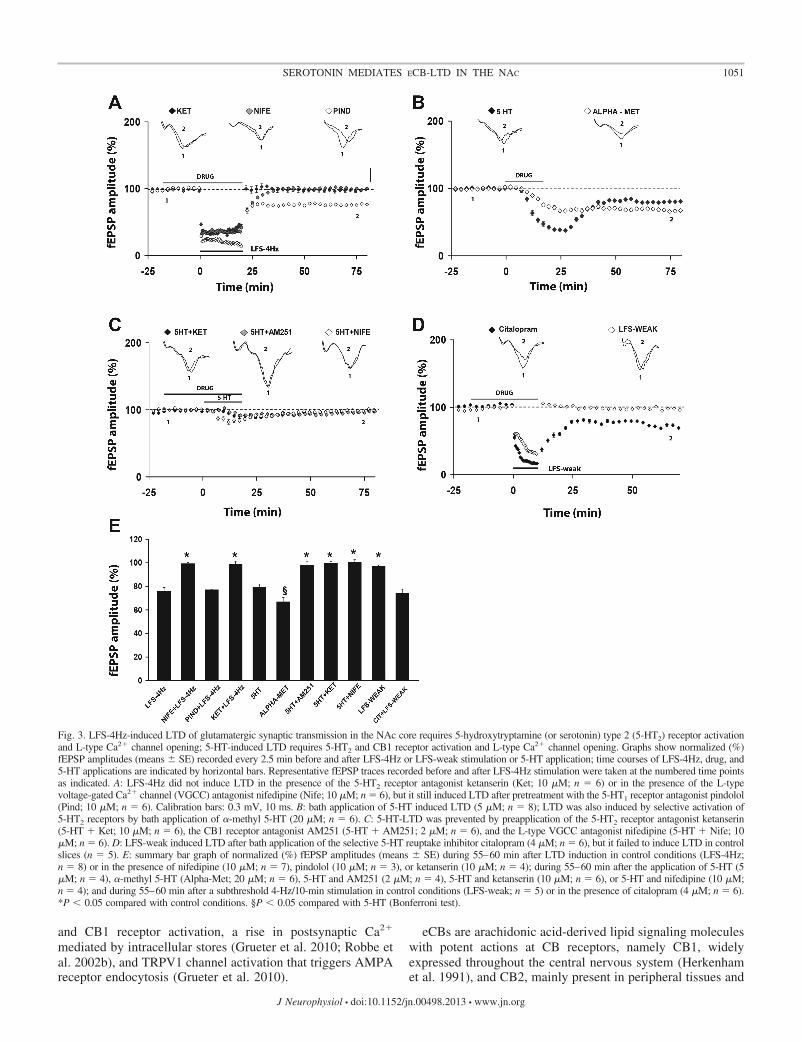
To further clarify the mechanisms underlying LFS-4Hz-induced LTD, we measured the effect of the L-type VGCCblocker nifedipine (10 �M) on LFS-4Hz-LTD. As depicted inFig. 3, A and E, nifedipine completely prevented LTD induc-tion. According to the observation that the dense afferent DAinnervation in the NAc regulates the activity of the mediumspiny neurons (MSN), which comprise the great majority ofNAc neurons, as well as their ability to pursue plastic changes(Kreitzer and Malenka 2005; Schotanus and Chergui 2008;
Thomas et al. 2001), we next examined whether DA is alsoinvolved in LFS-4Hz-induced LTD. To this aim, the DA D1and D2 receptor antagonists SCH-23390 (25 �M) and sulpiride(25 �M) were added to the slice-perfusing medium 20 minbefore and during LFS-4Hz stimulation. We found that block-ade of both D1 and D2 DA receptors did not prevent LTDinduction (n � 6; data not shown).
Because CB1 and 5-HT2 receptors are both expressed in theNAc (Barnes and Sharp 1999; Pertwee 1997), and activation of5-HT2 receptors has been shown to cause production andrelease of eCBs leading to LTD in the inferior olive (Best andRegehr 2008), we investigated the involvement of 5-HT2receptors in LFS-4Hz-induced LTD. As shown in Fig. 3, A andE, the 5-HT2 receptor blocker ketanserin (10 �M) completelyblocked LTD. Data obtained in the presence of the 5-HT1receptor antagonist pindolol (10 �M) suggest that this receptorsubtype does not contribute to LFS-4Hz-induced LTD (n � 4;Fig. 1, A and E; Table 1).
To confirm the involvement of 5-HT in this form of eCB-LTD, in the next set of experiments LFS-4Hz was replaced bybath application of 5-HT (5 �M) for 20 min. Consistent withprevious observations at corticostriatal synapses (Mathur et al.2011), 5-HT added to the slice-perfusing medium (5 �M, 20min) produced an initial, large depression and a significant(P � 0.05) and stable LTD following washout (n � 4; Fig. 3,B and E; Table 3). To test if the effect of 5-HT-LTD wasmediated by 5-HT2 receptors, slices were perfused with thespecific 5-HT2 receptor agonist �-methyl 5-HT (20 �M). Asshown in Fig. 3, B and E, the agonist mimicked the effect of5-HT (n � 6) to induce LTD. A comparison of the amplitudeof the LTD obtained in the presence of 5-HT vs. �-methyl5-HT revealed that the magnitude of LTD was significantlyincreased by the latter treatment. To further test the involve-ment of 5-HT2 receptors in 5-HT-LTD, slices were perfusedwith ketanserin (10 �M) for at least 20 min before and duringapplication of 5-HT (5 �M). As depicted in Fig. 3, C and E,ketanserin completely prevented the effect of 5-HT on fEPSPamplitude (n � 6; Table 3). We then tested whether the effectsof 5-HT on synaptic transmission was mediated by the CB1receptors: coapplication of 5-HT and the CB1 receptors antag-onist AM251 (2 �M) completely prevented 5-HT-LTD (n � 4;Fig. 3, C and E; Table 3). We next examined the involvementof the L-type VGCC in 5-HT-LTD by applying 5-HT (5 �M)along with the specific antagonist nifedipine (10 �M). Asdepicted in Fig. 3, C and E, nifedipine completely prevented5-HT-LTD (n � 4; Table 3). To test whether endogenous 5-HTin the acute slice preparation would facilitate the induction ofLTD, the subthreshold stimulation protocol LFS-weak wasapplied in the presence of the SSRI citalopram (4 �M). Figure3, D and E, shows that this treatment enabled LFS-weak toinduce LTD (n � 6; Table 2), indicating the involvement ofendogenous 5-HT in this process.
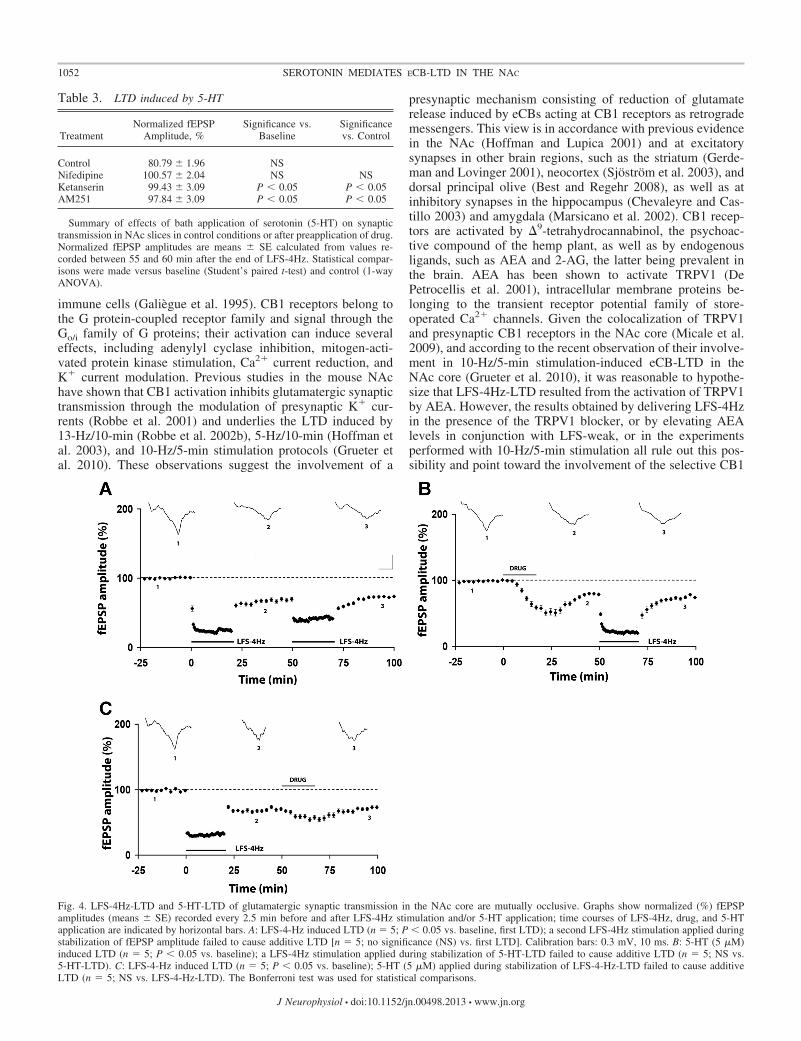
Presynaptic CB1 receptors have been shown to inhibit glu-tamate release at the prelimbic cortex-NAc synapses (Robbe etal. 2001); therefore, if 5-HT receptor activation and LFS-4Hzshare common mechanisms to inhibit excitatory synaptic trans-mission, LFS-4Hz-LTD and 5-HT-LTD may be mutually oc-clusive. We first verified whether repetition of LFS-4Hz causedsaturation of LTD. As shown in Fig. 4A, a second LFS-4Hzapplied during LTD consolidation failed to induce a furtherdepression: fEPSP amplitude was 74.02 � 5.44% (n � 5; P �
Table 1. LTD induced by LFS-4Hz
TreatmentNormalized fEPSP
amplitude, %Significance vs.
BaselineSignificance vs.
Control
Control 75.64 � 3.26 P � 0.05 —D-AP5 76.10 � 6.79 P � 0.05 NSKynureate 62.41 � 7.68 P � 0.05 NSLY-341495 71.55 � 11.41 P � 0.05 NSMPEP 79.48 � 1.70 P � 0.05 NSLY-367385 73.63 � 1.03 P � 0.05 NSAM251 101.36 � 0.51 NS P � 0.05LY-320135 99.39 � 1.53 NS P � 0.05Capsazepine 80.87 � 2.90 P � 0.05 NSTHL 98.91 � 3.89 NS P � 0.05Ketanserin 98.88 � 2.51 NS P � 0.05Pindolol 77.09 � 0.53 P � 0.05 NSNifedipine 99.28 � 0.99 NS P � 0.05SCH-2339 �
sulpiride 67.48 � 9.38 P � 0.05 NS
Summary of effects of 20-min low-frequency (4 Hz) stimulation (LFS-4Hz)on synaptic transmission in nucleus accumbens (NAc) slices in control con-ditions or in the presence of drug. Normalized field excitatory postsynapticpotential (fEPSP) amplitudes are means � SE calculated from values recordedbetween 55 and 60 min after the end of LFS-4Hz. Statistical comparisons weremade versus baseline (Student’s paired t-test) and control (1-way ANOVA).NS, not significant.
1049SEROTONIN MEDIATES ECB-LTD IN THE NAC
J Neurophysiol • doi:10.1152/jn.00498.2013 • www.jn.org
0.05 vs. baseline) between 25 and 30 min after the end of thefirst LFS-4Hz stimulation, and 73.74 � 3.78% between 25 and30 min after the end of the second LFS-4Hz stimulation, avalue similar to the one recorded after the first LFS-4Hz [n �5; no significance (NS) vs. first LFS-4Hz-LTD]. As shown inFig. 4B, to verify whether 5-HT-LTD also occludes LFS-4Hz-LTD, we first perfused the slices with 5-HT (5 �M) and thenapplied LFS-4Hz: we found that fEPSP amplitude was 78.99 �
2.96% (n � 5; P � 0.05 vs. baseline) between 25 and 30 minafter the end of 5-HT application, and 76.20 � 1.34% between25 and 30 min after the end of LFS-4Hz stimulation (n � 5; NSvs. 5-HT-induced LTD). Conversely, 5-HT (5 �M) had nosignificant effect when applied after LFS-4Hz-LTD was al-ready induced: fEPSP amplitude was 70.52 � 4.46% of base-line (n � 7; P � 0.05 vs. baseline) between 25 and 30 min afterLFS-4Hz, and 72.98 � 4.41% between 25 and 30 min after5-HT application (n � 7, NS vs. LFS-4Hz-LTD; Fig. 4C).
DISCUSSION
In recent years, LTD in the NAc has been investigated inseveral studies (Grueter et al. 2010; Hoffman et al. 2003; Ji andMartin 2012; Kahn et al. 2001; Kombian and Malenka 1994;Pennartz et al. 1993; Thomas et al. 2000, 2001; Robbe et al.2002a, 2002b), but the present work appears to provide the firstevidence of an eCB-dependent LTD requiring 5-HT2 and CB1receptor activation and increase in Ca2� entry through L-typeVGCC. Thus at least two distinct forms of eCB-dependentLTD can be induced in the NAc depending on the inductionprotocol used, because previous studies have shown that stim-ulations at higher frequencies induce LTDs requiring mGluR5
Fig. 2. LFS-4Hz-induced LTD of glutamatergic synaptic transmission in the NAc core requires 2-arachidonoylglycerol (2-AG) production and cannabinoid type1 (CB1) receptor but not transient receptor potential vanilloid 1 (TRPV1) activation. Graphs show normalized (%) fEPSP amplitudes (means � SE) recordedevery 2.5 min before and after LFS-4Hz or a subthreshold 4-Hz/10-min stimulation (LFS-weak); time courses of LFS-4Hz and drug applications are indicatedby horizontal bars. Representative fEPSP traces recorded before and after LFS-4Hz stimulation were taken at the numbered time points as indicated. A: LFS-4Hzdid not induce LTD in the presence of the CB1 receptor antagonists AM251 (2 �M; n � 6) and LY-320135 (10 �M; n � 6), nor in the presence of thediacylglycerol lipase � inhibitor tetrahydrolipstatin (THL; 10 �M; n � 9), but it still induced LTD in the presence of the TRPV1 antagonist capsazepine (10�M; n � 6). Calibration bars: 0.3 mV, 10 ms. B: LFS-weak failed to induce LTD in control slices (n � 5) and in the presence of the fatty acid amide hydrolaseinhibitor JNJ-1661010 (1 �M; n � 5), but it induced LTD in the presence of JZL-184 (50 �M; n � 6), an inhibitor of monoacylglycerol lipase. C: summarybar graph of normalized (%) fEPSP amplitudes (means � SE) during 55–60 min after LTD induction in control conditions (LFS-4Hz; n � 8) or in the presenceof AM251 (2 �M; n � 6), LY-320135 (10 �M; n � 6), THL (10 �M; n � 9), or capsazepine (10 �M; n � 6) and during 55–60 min after a subthreshold4-Hz/10-min stimulation in control conditions (LFS-weak; n � 5) or in the presence of JNJ-1661010 (1 �M; n � 5) or JZL-184 (50 �M; n � 6). *P � 0.05compared with control conditions (Bonferroni test).
Table 2. Effect of LFS-weak
TreatmentNormalized fEPSP
Amplitude, %Significance vs.
BaselineSignificancevs. Control
Control 96.97 � 1.16 NSJNJ-1661010 102.43 � 2.80 NS NSJZL-184 78.88 � 2.24 P � 0.05 P � 0.05Citalopram 74.25 � 3.33 P � 0.05 P � 0.05
Summary of effects of a subthreshold 10-min/4-Hz stimulation (LFS-weak)on synaptic transmission in NAc slices in control conditions or after preappli-cation of drug. Normalized fEPSP amplitudes are means � SE calculated fromvalues recorded between 55 and 60 min after the end of LFS-4Hz. Statisticalcomparisons were made versus baseline (Student’s paired t-test) and control(1-way ANOVA).
1050 SEROTONIN MEDIATES ECB-LTD IN THE NAC
J Neurophysiol • doi:10.1152/jn.00498.2013 • www.jn.org
and CB1 receptor activation, a rise in postsynaptic Ca2�
mediated by intracellular stores (Grueter et al. 2010; Robbe etal. 2002b), and TRPV1 channel activation that triggers AMPAreceptor endocytosis (Grueter et al. 2010).
eCBs are arachidonic acid-derived lipid signaling moleculeswith potent actions at CB receptors, namely CB1, widelyexpressed throughout the central nervous system (Herkenhamet al. 1991), and CB2, mainly present in peripheral tissues and
Fig. 3. LFS-4Hz-induced LTD of glutamatergic synaptic transmission in the NAc core requires 5-hydroxytryptamine (or serotonin) type 2 (5-HT2) receptor activationand L-type Ca2� channel opening; 5-HT-induced LTD requires 5-HT2 and CB1 receptor activation and L-type Ca2� channel opening. Graphs show normalized (%)fEPSP amplitudes (means � SE) recorded every 2.5 min before and after LFS-4Hz or LFS-weak stimulation or 5-HT application; time courses of LFS-4Hz, drug, and5-HT applications are indicated by horizontal bars. Representative fEPSP traces recorded before and after LFS-4Hz stimulation were taken at the numbered time pointsas indicated. A: LFS-4Hz did not induce LTD in the presence of the 5-HT2 receptor antagonist ketanserin (Ket; 10 �M; n � 6) or in the presence of the L-typevoltage-gated Ca2� channel (VGCC) antagonist nifedipine (Nife; 10 �M; n � 6), but it still induced LTD after pretreatment with the 5-HT1 receptor antagonist pindolol(Pind; 10 �M; n � 6). Calibration bars: 0.3 mV, 10 ms. B: bath application of 5-HT induced LTD (5 �M; n � 8); LTD was also induced by selective activation of5-HT2 receptors by bath application of �-methyl 5-HT (20 �M; n � 6). C: 5-HT-LTD was prevented by preapplication of the 5-HT2 receptor antagonist ketanserin(5-HT � Ket; 10 �M; n � 6), the CB1 receptor antagonist AM251 (5-HT � AM251; 2 �M; n � 6), and the L-type VGCC antagonist nifedipine (5-HT � Nife; 10�M; n � 6). D: LFS-weak induced LTD after bath application of the selective 5-HT reuptake inhibitor citalopram (4 �M; n � 6), but it failed to induce LTD in controlslices (n � 5). E: summary bar graph of normalized (%) fEPSP amplitudes (means � SE) during 55–60 min after LTD induction in control conditions (LFS-4Hz;n � 8) or in the presence of nifedipine (10 �M; n � 7), pindolol (10 �M; n � 3), or ketanserin (10 �M; n � 4); during 55–60 min after the application of 5-HT (5�M; n � 4), �-methyl 5-HT (Alpha-Met; 20 �M; n � 6), 5-HT and AM251 (2 �M; n � 4), 5-HT and ketanserin (10 �M; n � 6), or 5-HT and nifedipine (10 �M;n � 4); and during 55–60 min after a subthreshold 4-Hz/10-min stimulation in control conditions (LFS-weak; n � 5) or in the presence of citalopram (4 �M; n � 6).*P � 0.05 compared with control conditions. §P � 0.05 compared with 5-HT (Bonferroni test).
1051SEROTONIN MEDIATES ECB-LTD IN THE NAC
J Neurophysiol • doi:10.1152/jn.00498.2013 • www.jn.org
immune cells (Galiègue et al. 1995). CB1 receptors belong tothe G protein-coupled receptor family and signal through theGo/i family of G proteins; their activation can induce severaleffects, including adenylyl cyclase inhibition, mitogen-acti-vated protein kinase stimulation, Ca2� current reduction, andK� current modulation. Previous studies in the mouse NAchave shown that CB1 activation inhibits glutamatergic synaptictransmission through the modulation of presynaptic K� cur-rents (Robbe et al. 2001) and underlies the LTD induced by13-Hz/10-min (Robbe et al. 2002b), 5-Hz/10-min (Hoffman etal. 2003), and 10-Hz/5-min stimulation protocols (Grueter etal. 2010). These observations suggest the involvement of a
presynaptic mechanism consisting of reduction of glutamaterelease induced by eCBs acting at CB1 receptors as retrogrademessengers. This view is in accordance with previous evidencein the NAc (Hoffman and Lupica 2001) and at excitatorysynapses in other brain regions, such as the striatum (Gerde-man and Lovinger 2001), neocortex (Sjöström et al. 2003), anddorsal principal olive (Best and Regehr 2008), as well as atinhibitory synapses in the hippocampus (Chevaleyre and Cas-tillo 2003) and amygdala (Marsicano et al. 2002). CB1 recep-tors are activated by 9-tetrahydrocannabinol, the psychoac-tive compound of the hemp plant, as well as by endogenousligands, such as AEA and 2-AG, the latter being prevalent inthe brain. AEA has been shown to activate TRPV1 (DePetrocellis et al. 2001), intracellular membrane proteins be-longing to the transient receptor potential family of store-operated Ca2� channels. Given the colocalization of TRPV1and presynaptic CB1 receptors in the NAc core (Micale et al.2009), and according to the recent observation of their involve-ment in 10-Hz/5-min stimulation-induced eCB-LTD in theNAc core (Grueter et al. 2010), it was reasonable to hypothe-size that LFS-4Hz-LTD resulted from the activation of TRPV1by AEA. However, the results obtained by delivering LFS-4Hzin the presence of the TRPV1 blocker, or by elevating AEAlevels in conjunction with LFS-weak, or in the experimentsperformed with 10-Hz/5-min stimulation all rule out this pos-sibility and point toward the involvement of the selective CB1
Table 3. LTD induced by 5-HT
TreatmentNormalized fEPSP
Amplitude, %Significance vs.
BaselineSignificancevs. Control
Control 80.79 � 1.96 NSNifedipine 100.57 � 2.04 NS NSKetanserin 99.43 � 3.09 P � 0.05 P � 0.05AM251 97.84 � 3.09 P � 0.05 P � 0.05
Summary of effects of bath application of serotonin (5-HT) on synaptictransmission in NAc slices in control conditions or after preapplication of drug.Normalized fEPSP amplitudes are means � SE calculated from values re-corded between 55 and 60 min after the end of LFS-4Hz. Statistical compar-isons were made versus baseline (Student’s paired t-test) and control (1-wayANOVA).
Fig. 4. LFS-4Hz-LTD and 5-HT-LTD of glutamatergic synaptic transmission in the NAc core are mutually occlusive. Graphs show normalized (%) fEPSPamplitudes (means � SE) recorded every 2.5 min before and after LFS-4Hz stimulation and/or 5-HT application; time courses of LFS-4Hz, drug, and 5-HTapplication are indicated by horizontal bars. A: LFS-4-Hz induced LTD (n � 5; P � 0.05 vs. baseline, first LTD); a second LFS-4Hz stimulation applied duringstabilization of fEPSP amplitude failed to cause additive LTD [n � 5; no significance (NS) vs. first LTD]. Calibration bars: 0.3 mV, 10 ms. B: 5-HT (5 �M)induced LTD (n � 5; P � 0.05 vs. baseline); a LFS-4Hz stimulation applied during stabilization of 5-HT-LTD failed to cause additive LTD (n � 5; NS vs.5-HT-LTD). C: LFS-4-Hz induced LTD (n � 5; P � 0.05 vs. baseline); 5-HT (5 �M) applied during stabilization of LFS-4-Hz-LTD failed to cause additiveLTD (n � 5; NS vs. LFS-4-Hz-LTD). The Bonferroni test was used for statistical comparisons.
1052 SEROTONIN MEDIATES ECB-LTD IN THE NAC
J Neurophysiol • doi:10.1152/jn.00498.2013 • www.jn.org

receptor ligand 2-AG, which has no activity at TRPV1 (DePetrocellis et al. 2000). 2-AG is the most abundant eCB in thebrain (Mechoulam et al. 1995; Sugiura et al. 1995); in partic-ular, its concentration in the mesencephalon is 100 timeshigher than that of AEA (Bisogno et al. 1999). In addition,2-AG acts as a potent and full agonist at CB1 receptors,whereas AEA is only a partial agonist (Savinainen et al. 2001;Stella et al. 1997; Sugiura et al. 1995, 1999). Biochemicalstudies indicate that the major enzymes involved in 2-AGbiosynthesis are phospholipase C (PLC) and DGL (Piomelli2003), although other pathways have also been suggested(Sugiura et al. 2006). The involvement of 2-AG in LFS-4Hz-LTD was confirmed by the experiments performed in thepresence of the DGL inhibitor THL, which was proven to haveno direct effect on CB1 receptors (Szabo et al. 2006; Uchi-gashima et al. 2007); moreover, it is further supported by theevidence that LTD can be induced by a subthreshold stimula-tion protocol when endogenous 2-AG levels are increased byan inhibitor of MAGL, the 2-AG-hydrolyzing enzyme. Thisfinding is consistent with previous observations in the hip-pocampus (Ameri and Simmet 2000; Edwards et al. 2006;Hashimotodani et al. 2007; Stella et al. 1997), cerebellum(Maejima et al. 2005; Safo and Regehr 2005), and prefrontalcortex (Melis et al. 2004), supporting the involvement of2-AG.
In addition to eCB, present results point toward the involve-ment of 5-HT, acting at 5-HT2 receptors, in LFS-4Hz-LTD, asdemonstrated by LFS-4Hz experiments performed in the pres-ence of the 5-HT2 receptor blocker and by the evidence thatLTD can be induced by the subthreshold stimulation protocolwhen the endogenous 5-HT levels are elevated by the SSRIcitalopram. This evidence is further supported by the observa-tion that LTD can also be elicited by bath application of 5-HTor a specific 5-HT2 receptor agonist.
It was recently shown that 5-HT increases the activity ofGABAergic interneurons in the NAc through 5-HT2C recep-tors, resulting in inhibitory action on MSN (Blomeley andBracci 2009). One might argue that the 5-HT2-dependent LTDobserved in the current study could derive from increasedGABAergic signaling; however, this possibility seems un-likely, because the experiments were carried out in the constantpresence of the GABAA receptor antagonist picrotoxin. Rather,present data support the view that 5-HT2 receptor activationmight trigger eCB production and release, in accordance witha previous observation in the dorsal principal olive (Best andRegehr 2008).
We also investigated the possible involvement of L-typeVGCC in both LFS-4Hz- and 5-HT-induced LTD, becausethey are particularly important for down- to up-state transitionof MSN (Carter and Sabatini 2004). Moreover, in vivo studieshave demonstrated that these channels play a critical role inpsychostimulant-induced behavioral and neuronal plasticity invarious regions of the mesolimbic DA system, including theNAc (Gnegy 2000; Licata and Pierce 2003). We found thatnifedipine prevented both LFS-4Hz- and 5-HT-induced LTD,indicating that these channels play a critical role in this pro-cess, as previously observed in eCB-induced LTD in dorsalstriatum MSN (Kreitzer and Malenka 2005), as well as in otherforms of synaptic plasticity in various brain areas, includingthe hippocampus (Bolshakov and Siegelbaum 1994; Groverand Teyler 1992; Morgan and Teyler 1999), lateral amygdala
(Weisskopf et al. 1999), and perirhinal cortex (Seoane et al.2009). It has been shown that large increases in intracellularCa2� (10–50 �M) are needed to activate phospholipase D andAEA biosynthesis, whereas lower intracellular Ca2� elevationscan activate PLC and DAG, yielding 2-AG production (Bre-nowitz and Regehr 2003; Hashimotodani et al. 2005). It seemsreasonable to suppose that 5-HT2 activation by 5 HT (eitherendogenously released in response to LFS-4Hz stimulation orexogenously applied) might stimulate 2-AG production byincreasing both DGL activity and L-type VGCC permeability,resulting in intracellular Ca2� level elevation compatible with2-AG production and release.
Altogether, present results suggest that LFS-4Hz stimulationof glutamatergic afferences induces a novel form of eCB-LTDin the NAc core involving the release of 5-HT, which acts at5-HT2 postsynaptic receptors, increasing Ca2� influx throughL-type channels and 2-AG production and release; the eCBwould travel retrogradely and bind to presynaptic CB1 recep-tors, causing a long-lasting decrease of glutamate release,resulting in LTD (Fig. 5).
Dysfunction in 5-HT neurotransmission plays a critical rolein a variety of psychiatric pathologies, including drug addic-tion, major depression, food intake disorders, emesis, chronicpain, sleep disorders, autism, and schizophrenia (Malison et al.1998; Mann et al. 1995; Pacher et al. 2006; Shahsvand-Ananloo et al. 2013). Present observations might be helpful tounderstand the mechanisms underlying these disorders, hope-fully leading to the development of new pharmacological tools.Moreover, the present findings add a bit of knowledge to thegrowing body of evidence pointing toward a major role of theeCB system in short- and long-term synaptic plasticity in brainareas relevant to the mediation of reward.
ACKNOWLEDGMENTS
Present address of F. Tamagnini: School of Physiology and Pharmacology,Medical Resource Council Centre of Synaptic Plasticity, Bristol University,Bristol, UK.
Fig. 5. Diagram of the proposed signaling cascade underlying LFS-4Hz-LTD ofglutamatergic synaptic transmission in the NAc core. LFS-4Hz stimulation ofglutamatergic fibers in the NAc core increases the release of 5-HT acting at 5-HT2
receptors, causing an increase in Ca2� influx through L-type VGCC and leadingto 2-AG production and release; the endocannabinoid travels retrogradely andbinds to presynaptic CB1 receptors, causing a long-lasting decrease of glutamaterelease, resulting in LTD. Glu, glutamate; AMPA, �-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid glutamate receptor; NMDA, N-methyl-D-aspartate re-ceptor; G, G protein; PIP2, phosphatidylinositol 4,5-bisphosphate; DAG, diacyl-glycerol; PLC, phospholipase C.
1053SEROTONIN MEDIATES ECB-LTD IN THE NAC
J Neurophysiol • doi:10.1152/jn.00498.2013 • www.jn.org

GRANTS
This work was supported by Ricerca Fondamentale Orientata (RFO) grantsfrom the University of Bologna and by a fundamental contribution from the“Fondazione del Monte di Bologna e Ravenna” (to G. Aicardi).
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
AUTHOR CONTRIBUTIONS
C.B. and G.A. conception and design of research; C.B. and G.B. performedexperiments; C.B., G.B., and F.T. analyzed data; C.B. and G.A. interpretedresults of experiments; C.B. and G.B. prepared figures; C.B. drafted manu-script; C.B., G.B., F.T., and G.A. edited and revised manuscript; C.B., G.B.,F.T., and G.A. approved final version of manuscript.
REFERENCES
Altier N, Stewart J. The role of dopamine in the nucleus accumbens inanalgesia. Life Sci 65: 2269–2287, 1999.
Ameri A, Simmet T. Effects of 2-arachidonylglycerol, an endogenous can-nabinoid, on neuronal activity in rat hippocampal slices. Naunyn Schmie-debergs Arch Pharmacol 361: 265–272, 2000.
Barnes NM, Sharp T. A review of central 5-HT receptors and their function.Neuropharmacology 38: 1083–1152, 1999.
Belujon P, Patton MH, Grace AA. Role of the prefrontal cortex in alteredhippocampal-accumbens synaptic plasticity in a developmental animalmodel of schizophrenia. Cereb Cortex. First published December 12, 2012;doi:10.1093/cercor/bhs380.
Best AR, Regehr WG. Serotonin evokes endocannabinoid release and retro-gradely suppresses excitatory synapses. J Neurosci 28: 6508–6515, 2008.
Bisogno T, Berrendero F, Ambrosino G, Cebeira M, Ramos JA, Fernan-dez-Ruiz JJ, Di Marzo V. Brain regional distribution of endocannabinoids:implications for their biosynthesis and biological function. Biochem BiophysRes Commun 256: 377–380, 1999.
Blomeley CP, Bracci E. Serotonin excites fast-spiking interneurons in thestriatum. Eur J Neurosci 29: 1604–1614, 2009.
Blume LC, Bass CE, Childers SR, Dalton GD, Roberts DC, RichardsonJM, Xiao R, Selley DE, Howlett AC. Striatal CB1 and D2 receptorsregulate expression of each other, CRIP1A and delta opioid systems. JNeurochem 124: 808–820, 2013.
Bolshakov VY, Siegelbaum SA. Postsynaptic induction and presynapticexpression of hippocampal long-term depression. Science 264: 1148–1152,1994.
Brenowitz SD, Regehr WG. Calcium dependence of retrograde inhibition byendocannabinoids at synapses onto Purkinje cells. J Neurosci 23: 6373–6384, 2003.
Brog JS, Salyapongse A, Deutch AY, Zahm DS. The patterns of afferentinnervation of the core and shell in the “accumbens” part of the rat ventralstriatum: immunohistochemical detection of retrogradely transported fluoro-gold. J Comp Neurol 338: 255–278, 1993.
Carter AG, Sabatini BL. State-dependent calcium signaling in dendriticspines of striatal medium spiny neurons. Neuron 44: 483–493, 2004.
Chevaleyre V, Castillo PE. Heterosynaptic LTD of hippocampal GABAergicsynapses: a novel role of endocannabinoids in regulating excitability. Neu-ron 38: 461–472, 2003.
Christie MJ, James LB, Beart PM. An excitant amino acid projection fromthe medial prefrontal cortex to the anterior part of nucleus accumbens in therat. J Neurochem 45: 477–482, 1985.
De Petrocellis L, Bisogno T, Maccarrone M, Davis JB, Finazzi-Agro A, DiMarzo V. The activity of anandamide at vanilloid VR1 receptors requiresfacilitated transport across the cell membrane and is limited by intracellularmetabolism. J Biol Chem 276: 12856–12863, 2001.
De Petrocellis L, Bisogno T, Davis JB, Pertwee RG, Di Marzo V. Overlapbetween the ligand recognition properties of the anandamide transporter andthe VR1 vanilloid receptor: inhibitors of anandamide uptake with negligiblecapsaicin-like activity. FEBS Lett 483: 52–56, 2000.
Edwards DA, Kim J, Alger BE. Multiple mechanisms of endocannabinoidresponse initiation in hippocampus. J Neurophysiol 95: 67–75, 2006.
Galiègue S, Mary S, Marchand J, Dussossy D, Carrière D, Carayon P,Bouaboula M, Shire D, Le Fur G, Casellas P. Expression of central and
peripheral cannabinoid receptors in human immune tissues and leukocytessubpopulation. Eur J Biochem 232: 54–61, 1995.
Gerdeman G, Lovinger DM. CB1 cannabinoid receptor inhibits synapticrelease of glutamate in rat dorsolateral striatum. J Neurophysiol 85: 468–467, 2001.
Gnegy ME. Ca2�/calmodulin signaling in NMDA-induced synaptic plasticity.Crit Rev Neurobiol 14: 91–129, 2000.
Grover LM, Teyler TJ. N-methyl-D-aspartate receptor-independent long-term potentiation in area CA1 of rat hippocampus: input-specific inductionand preclusion in a non-tetanized pathway. Neuroscience 49: 7–11, 1992.
Grueter BA, Brasnjo G, Malenka RC. Postsynaptic TRPV1 triggers celltype-specific long-term depression in the nucleus accumbens. Nat Neurosci13: 1519–1525, 2010.
Hashimotodani Y, Ohno-Shosaku T, Tsubokawa H, Ogata H, Emoto K,Maejima T, Araishi K, Shin HS, Kano M. Phospholipase Cbeta serves asa coincidence detector through its Ca2� dependency for triggering retro-grade endocannabinoid signal. Neuron 45: 257–268, 2005.
Hashimotodani Y, Ohno-Shosaku T, Kano M. Presynaptic monoacylglyc-erol lipase activity determines basal endocannabinoid tone and terminatesretrograde endocannabinoid signaling in the hippocampus. J Neurosci 27:1211–1219, 2007.
Herkenham M, Lynn AB, Johnson MR, Melvin LS, de Costa BR, RiceKC. Characterization and localization of cannabinoid receptors in rat brain:a quantitative in vitro autoradiographic study. J Neurosci 11: 563–583,1991.
Hoffman AF, Lupica CR. Direct actions of cannabinoids on synaptic trans-mission in the nucleus accumbens: a comparison with opioids. J Neuro-physiol 85: 72–83, 2001.
Hoffman AF, Oz M, Caulder T, Lupica CR. Functional telerance andblockade of long-term depression at synapses in the nucleus accumbens afterchronic cannabinoid exposure. J Neurosci 23: 4815–4820, 2003.
Ji X, Martin GE. New rules governing synaptic plasticity in core nucleusaccumbens in medium spiny neurons. Eur J Neurosci 36: 3615–3627, 2012.
Kahn L, Alonso G, Robbe D, Bockaert J, Manzoni OJ. Group 2 metabo-tropic glutamate receptors induced long term depression in mouse striatalslices. Neurosci Lett 316: 178–182, 2001.
Koob GF, Bloom FE. Cellular and molecular mechanisms of drug depen-dence. Science 242: 715–723, 1988.
Kombian SB, Malenka RC. Simultaneous LTP of non-NMDA and LTD ofNMDA-receptor-mediated responses in the nucleus accumbens. Nature 368:242–246, 1994.
Kreitzer AC, Malenka RC. Dopamine modulation of state-dependent endo-cannabinoid release and long-term depression in the striatum. J Neurosci 25:10537–10545, 2005.
Le Moal M, Simon H. Mesocorticolimbic dopaminergic network: functionaland regulatory roles. Physiol Rev 71: 155–234, 1991.
Li Y, Kauer JA. Repeated exposure to amphetamine disrupts dopaminergicmodulation of excitatory synaptic plasticity and neurotransmission in nu-cleus accumbens. Synapse 51: 1–10, 2004.
Licata SC, Pierce RC. The roles of calcium/calmodulin-dependent andRas/mitogen-activated protein kinases in the development of psychostimu-lant-induced behavioral sensitization. J Neurochem 85: 14–22, 2003.
Maejima T, Oka S, Hashimotodani Y, Ohno-Shosaku T, Aiba A, Wu D,Waku K, Sugiura T, Kano M. Synaptically driven endocannabinoidrelease requires Ca2�-assisted metabotropic glutamate receptor subtype 1 tophospholipase C�4 signaling cascade in the cerebellum. J Neurosci 25:6826–6835, 2005.
Malison RT, Price LH, Berman R, van Dyck CH, Pelton GH, CarpenterL, Sanacora G, Owens MJ, Nemeroff CB, Rajeevan N, Baldwin RM,Seibyl JP, Innis RB, Charney DS. Reduced brain serotonin transporteravailability in major depression as measured by [123I]-2 beta-carbome-thoxy-3 beta-(4-iodophenyl)tropane and single photon emission computedtomography. Biol Psychiatry 44: 1090–1098, 1998.
Mann JJ, McBride PA, Malone KM, DeMeo M, Keilp J. Blunted seroto-nergic responsivity in depressed inpatients. Neuropsychopharmacology 13:53–64, 1995.
Marsicano G, Moosmann B, Hermann H, Lutz B, Behl C. Neuroprotectiveproperties of cannabinoids against oxidative stress: role of the cannabinoidreceptor CB1. J Neurochem 80: 448–456, 2002.
Marty VN, Spigelman I. Effects of alcohol on the membrane excitability andsynaptic transmission of medium spiny neurons in the nucleus accumbens.Alcohol 46: 317–327, 2012.
1054 SEROTONIN MEDIATES ECB-LTD IN THE NAC
J Neurophysiol • doi:10.1152/jn.00498.2013 • www.jn.org

Mathur BN, Capik NA, Alvarez VA, Lovinger DM. Serotonin induceslong-term depression at corticostriatal synapses. J Neurosci 31: 7402–7411,2011.
Mechoulam R, Ben-Shabat S, Hanus L, Ligumsky M, Kaminski NE,Schatz AR, Gopher A, Almog S, Martin BR, Compton DR. Identificationof an endogenous 2-monoglyceride, present in canine gut, that binds tocannabinoid receptors. Biochem Pharmacol 50: 83–90, 1995.
Melis M, Perra S, Muntoni AL, Pillolla G, Lutz B, Marsicano G, Di MarzoV, Gessa GL, Pistis M. Prefrontal cortex stimulation induces 2-arachi-donoyl-glycerol-mediated suppression of excitation in dopamine neurons. JNeurosci 24: 10707–10715, 2004.
Micale V, Cristino L, Tamburella A, Petrosino S, Leggio GM, Drago F, DiMarzo V. Altered responses of dopamine D3 receptor null mice to excito-toxic or anxiogenic stimuli: Possible involvement of the endocannabinoidand endovanilloid systems. Neurobiol Dis 36: 70–80, 2009.
Mogenson GJ, Jones DL, Yim CY. From motivation to action: functionalinterface between the limbic system and the motor system. Prog Neurobiol14: 69–97, 1980.
Morgan SL, Teyler TJ. VDCCs and NMDARs underlie two forms of LTP inCA1 hippocampus in vivo. J Neurophysiol 82: 736–740, 1999.
Ohno Y, Okano M, Masui A, Imaki J, Egawa M, Yoshihara C, Tatara A,Mizuguchi Y, Sasa M, Shimizu S. Region-specific elevation of D1 recep-tor-mediated neurotransmission in the nucleus accumbens of SHR, a ratmodel of attention deficit/hyperactivity disorder. Neuropharmacology 63:547–554, 2012.
Pacher P, Bátkai S, Kunos G. The endocannabinoid system as an emergingtarget of pharmacotherapy. Pharmacol Rev 58: 389–462, 2006.
Pennartz CM, Ameerun RF, Groenewegen HJ, Lopes da Silva FH. Syn-aptic plasticity in an in vitro slice preparation of the rat nucleus accumbens.Eur J Neurosci 5: 107–117, 1993.
Pennartz CM, Groenewegen HJ, Lopes da Silva FH. The nucleus accum-bens as a complex of functionally distinct neuronal ensembles: an integra-tion of behavioural, electrophysiological and anatomical data. Prog Neuro-biol 42: 719–761, 1994.
Pertwee RG. Pharmacology of cannabinoid CB1 and CB2 receptors. Phar-macol Ther 74: 129–180, 1997.
Piomelli D. The molecular logic of endocannabinoid signalling. Nat RevNeurosci 4: 873–884, 2003.
Puente N, Cui Y, Lassalle O, Lafourcade M, Georges F, Venance L,Grandes P, Manzoni OJ. Polymodal activation of the endocannabinoidsystem in the extended amygdala. Nat Neurosci 6: 1542–1547, 2011.
Robbe D, Alonso G, Chaumont S, Bockaert J, Manzoni OJ. Role ofp/q-Ca2� channels in metabotropic glutamate receptor 2/3-dependent pre-synaptic long-term depression at nucleus accumbens synapses. J Neurosci22: 4346–4356, 2002a.
Robbe D, Alonso G, Duchamp F, Bockaert J, Manzoni OJ. Localizationand mechanisms of action of endocannabinoid receptors at the glutamatergicsynapses of the mouse nucleus accumbens. J Neurosci 21: 109–116, 2001.
Robbe D, Kopf M, Remaury A, Bockaert J, Manzoni OJ. Endogenouscannabinoids mediate long-term synaptic depression in the nucleus accum-bens. Proc Natl Acad Sci USA 99: 8384–8388, 2002b.
Safo PK, Regehr WG. Endocannabinoids control the induction of cerebellarLTD. Neuron 48: 647–659, 2005.
Salamone JD, Correa M, Mingote SM, Weber SM. Beyond the rewardinghypothesis: alternative functions of nucleus accumbens dopamine. CurrOpin Pharmacol 5: 34–41, 2005.
Savinainen JR, Jarvinen T, Laine K, Laitinen JT. Despite substantialdegradation, 2-arachidonoylglycerol is a potent full efficacy agonist medi-
ating CB(1) receptor-dependent G-protein activation in rat cerebellar mem-branes. Br J Pharmacol 134: 664–672, 2001.
Schotanus SM, Chergui K. Dopamine D1 receptors and group I metabotropicglutamate receptors contribute to the induction of long-term potentiation inthe nucleus accumbens. Neuropharmacology 54: 837–844, 2008.
Seoane A, Massey PV, Keen H, Bashir ZI, Brown MW. L-type voltage-dependent calcium channel antagonists impair perirhinal long-term recog-nition memory and plasticity processes. J Neurosci 29: 9534–9544, 2009.
Shahsavand-Ananloo E, Berenji F, Sadeghniiat K, Alimadadi A, Zahirod-din AR, Tabatabaee M, Abbasi-Asl M, Ghaeli P. Comparing effects ofcitalopram with fluoxetine on sleep quality in patients with major depressivedisorders. Eur Rev Med Pharmacol Sci 17: 1155–1161, 2013.
Shirayama Y, Chaki S. Neurochemistry of the nucleus accumbens and itsrelevance to depression and antidepressant action in rodents. Curr Neuro-pharmacol 4: 277–291, 2006.
Sjöström PJ, Turrigiano GG, Nelson SB. Neocortical LTD via coincidentactivation of presynaptic NMDA and cannabinoid receptors. Neuron 39:641–654, 2003.
Stella N, Schweitzer P, Piomelli D. A second endogenous cannabinoid thatmodulates long-term potentiation. Nature 388: 773–778, 1997.
Sugiura T, Kishimoto S, Oka S, Gokoh M. Biochemistry, pharmacology andphysiology of 2-arachidonoylglycerol, an endogenous cannabinoid receptorligand. Prog Lipid Res 45: 405–446, 2006.
Sugiura T, Kondo S, Sukagawa A, Nakane S, Shinoda A, Itoh K, Ya-mashita A, Waku K. 2-Arachidonoylglycerol: a possible endogenouscannabinoid receptor ligand in brain. Biochem Biophys Res Commun 215:89–97, 1995.
Sugiura T, Kodaka T, Nakane S, Miyashita T, Kondo S, Suhara Y,Takayama H, Waku K, Seki C, Baba N, Ishima Y. Evidence that thecannabinoid CB1 receptor is a 2-arachidonoylglycerol receptor. Structure-activity relationship of 2-arachidonoylglycerol, ether-linked analogues, andrelated compounds. J Biol Chem 274: 2794–2801, 1999.
Szabo B, Urbanski MJ, Bisogno T, Di Marzo V, Mendiguren A, Baer WU,Freiman I. Depolarization-induced retrograde synaptic inhibition in themouse cerebellar cortex is mediated by 2-arachidonoylglycerol. J Physiol577: 263–280, 2006.
Thomas MJ, Beurrier C, Bonci A, Malenka RC. Long-term depression inthe nucleus accumbens: a neural correlate of behavioral sensitization tococaine. Nat Neurosci 4: 1217–1223, 2001.
Thomas MJ, Malenka RC, Bonci A. Modulation of long-term depression bydopamine in the mesolimbic system. J Neurosci 20: 5581–5586, 2000.
Tzounopoulos T, Janz R, Südhof TC, Micoll RA, Malenka RC. A role forcAMP in long-term depression at hippocampal mossy fiber synapses. Neu-ron 21: 837–845, 1998.
Uchigashima M, Narushima M, Fukaya M, Katona I, Kano M, WatanabeM. Subcellular arrangement of molecules for 2-arachidonoyl-glycerol-me-diated retrograde signaling and its physiological contribution to synapticmodulation in the striatum. J Neurosci 27: 3663–3676, 2007.
Van Bockstaele EJ, Pickel VM. Ultrastructure of serotonin-immunoreactiveterminals in the core and shell of the rat nucleus accumbens: cellularsubstrates for interactions with catecholamine afferents. J Comp Neurol 34:603–617, 1993.
Weisskopf MG, Bauer EP, LeDoux JE. L-type voltage-gated calcium chan-nels mediate NMDA-independent associative long-term potentiation at tha-lamic input synapses to the amygdala. J Neurosci 19: 10512–10519, 1999.
Wickelgren I. Getting the brain’s attention. Science 278: 35–37, 1997.Yu W, Calos M, Pilitsis J, Shin DS. Deconstructing the neural and ionic
involvement of seizure-like events in the striatal network. Neurobiol Dis 52:128–136, 2013.
1055SEROTONIN MEDIATES ECB-LTD IN THE NAC
J Neurophysiol • doi:10.1152/jn.00498.2013 • www.jn.org







![Dopamine release in nucleus accumbens during rewarded task switching measured by [¹¹C]raclopride](https://static.fdokumen.com/doc/165x107/633ae2b3ea31206fd207b694/dopamine-release-in-nucleus-accumbens-during-rewarded-task-switching-measured-by.jpg)












