
A consistent pattern of RIN1 rearrangements in oral squamous cell carcinoma cell lines supports a...
11
A Consistent Pattern of RIN1 Rearrangements in Oral Squamous Cell Carcinoma Cell Lines Supports a Breakage-Fusion-Bridge Cycle Model for 11q13 Amplification Michele I. Shuster, 1 Limin Han, 2 Michelle M. Le Beau, 3 Elizabeth Davis, 3 Mark Sawicki, 4 Christa M. Lese, 1 No-Hee Park, 5 John Colicelli, 2 and Susanne M. Gollin 1 * 1 Department of Human Genetics, University of Pittsburgh Graduate School of Public Health and the University of Pittsburgh Cancer Institute, Pittsburgh, Pennsylvania 2 Department of Biological Chemistry and Molecular Biology Institute, UCLA School of Medicine, Los Angeles, California 3 Section of Hematology/Oncology, University of Chicago, Chicago, Illinois 4 Department of Surgery, UCLA School of Medicine, Los Angeles, California 5 Department of Dentistry, UCLA School of Medicine, Los Angeles, California Gene amplification is a common feature of tumors. Overexpression of some amplified genes plays a role in tumor progression. Gene amplification can occur either extrachromosomally as double-minute chromosomes (dmin) or intrachromosomally in the form of homogeneously staining regions (hsrs). Approximately one-half of our oral squamous cell carcinomas (OSCCs) are characterized by amplification of band 11q13, usually as an hsr located entopically (occurring or situated at the normal chromosomal site, as opposed to ectopically). Using chromosomal fluorescence in situ hybridization (FISH), we confirmed the amplification of the cyclin D1 (CCND1/PRAD1) and fibroblast growth factor types 3 and 4 (FGF3/INT2 and FGF4/HSTF1) genes within the 11q13 amplicon in our series of primary OSCCs and derived cell lines. The human RIN1 gene was isolated as an RAS interaction/interference protein in a genetic selection in yeast and has been described as a putative effector of both the RAS and ABL oncogenes. We mapped RIN1 to 11q13.2. FISH analysis of 10 11q13-amplified OSCC cell lines revealed high-level RIN1 amplification in two cell lines. Three additional cell lines have what appear to be duplications and/or low-level amplification of RIN1, visible in both interphase and metaphase cells. The hybridization pattern of RIN1 on the metaphase chromosomes is particularly revealing; RIN1 signals flank the 11q13 hsr, possibly as a result of an inverted duplication. The gene amplification model of Coquelle et al. (1997) predicted that gene amplification occurs by breakage-fusion-bridge (BFB) cycles involving fragile sites. Our data suggest that the pattern of gene amplification at 11q13 in OSCC cell lines is consistent with a BFB model. RIN1 appears to be a valuable probe for investigating the process of gene amplification in general and, specifically, 11q13 amplification in oral cancer. Genes Chromosomes Cancer 28:153–163, 2000. © 2000 Wiley-Liss, Inc. INTRODUCTION Gene amplification is a common finding in many human cancers. The amplification and subsequent overexpression of critical genes may play an impor- tant role in the loss of cell cycle control in cancer. Amplification of chromosomal band 11q13 is fre- quently observed in head and neck squamous cell carcinoma (HNSCC), its subset, oral squamous cell carcinoma (OSCC), other aerodigestive tract can- cers, and breast carcinomas (Berenson et al., 1989; Roelofs et al., 1993; Williams et al., 1993; Jin et al., 1995, 1998; Lese et al., 1995; Rubin et al., 1995; Speicher et al., 1995; Åkervall et al., 1997; Muller et al., 1997; Bekri et al., 1998). A number of genes have been shown to be amplified in this region, including CCND1 (encoding cyclin D1), EMS1 (en- coding human cortactin, an actin-binding protein possibly involved in the organization of the cy- toskeleton and cell adhesion structures), and FGF3 and FGF4 (fibroblast growth factors 3 and 4, also called INT2 and HSTF1; Fig. 1) (Schuuring et al., 1998). A proximal gene, VEGFB, was not found to be amplified in mammary carcinoma cell lines (Paavonen et al., 1996). However, more distal This study was carried out in part in the University of Pittsburgh Cancer Institute Cytogenetics Facility. Christa M. Lese’s current address is Department of Human Ge- netics, University of Chicago, Chicago, Illinois. Michele I. Shuster’s current address is Department of Biological Sciences, University of Pittsburgh, Pittsburgh, Pennsylvania. Supported by: NIH; Grant numbers: F32DE05707 (to MS), RO1DE10513 and RO1DE12008 (to SMG), RO1CA56301 (to JC), DE10598 (to N-HP), and P30CA47049019 and S10RR11879; Smokeless Tobacco Research Council; Grant numbers: 0466, 0660 (to SMG); PHS; Grant number: CA40046 (to MML). *Correspondence to: Dr. Susanne M. Gollin, Department of Hu- man Genetics, University of Pittsburgh Graduate School of Public Health, 130 De Soto Street, Pittsburgh, PA 15261. E-mail: [email protected] Received 25 August 1999; Accepted 4 November 1999 GENES, CHROMOSOMES & CANCER 28:153–163 (2000) © 2000 Wiley-Liss, Inc.
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of A consistent pattern of RIN1 rearrangements in oral squamous cell carcinoma cell lines supports a...

A Consistent Pattern of RIN1 Rearrangements inOral Squamous Cell Carcinoma Cell Lines Supportsa Breakage-Fusion-Bridge Cycle Model for 11q13Amplification
Michele I. Shuster,1 Limin Han,2 Michelle M. Le Beau,3 Elizabeth Davis,3 Mark Sawicki,4 Christa M. Lese,1
No-Hee Park,5 John Colicelli,2 and Susanne M. Gollin1*
1Department of Human Genetics, University of Pittsburgh Graduate School of Public Health and the University of Pittsburgh CancerInstitute, Pittsburgh, Pennsylvania2Department of Biological Chemistry and Molecular Biology Institute, UCLA School of Medicine, Los Angeles, California3Section of Hematology/Oncology, University of Chicago, Chicago, Illinois4Department of Surgery, UCLA School of Medicine, Los Angeles, California5Department of Dentistry, UCLA School of Medicine, Los Angeles, California
Gene amplification is a common feature of tumors. Overexpression of some amplified genes plays a role in tumor progression.Gene amplification can occur either extrachromosomally as double-minute chromosomes (dmin) or intrachromosomally inthe form of homogeneously staining regions (hsrs). Approximately one-half of our oral squamous cell carcinomas (OSCCs) arecharacterized by amplification of band 11q13, usually as an hsr located entopically (occurring or situated at the normalchromosomal site, as opposed to ectopically). Using chromosomal fluorescence in situ hybridization (FISH), we confirmed theamplification of the cyclin D1 (CCND1/PRAD1) and fibroblast growth factor types 3 and 4 (FGF3/INT2 and FGF4/HSTF1) geneswithin the 11q13 amplicon in our series of primary OSCCs and derived cell lines. The human RIN1 gene was isolated as anRAS interaction/interference protein in a genetic selection in yeast and has been described as a putative effector of both theRAS and ABL oncogenes. We mapped RIN1 to 11q13.2. FISH analysis of 10 11q13-amplified OSCC cell lines revealed high-levelRIN1 amplification in two cell lines. Three additional cell lines have what appear to be duplications and/or low-levelamplification of RIN1, visible in both interphase and metaphase cells. The hybridization pattern of RIN1 on the metaphasechromosomes is particularly revealing; RIN1 signals flank the 11q13 hsr, possibly as a result of an inverted duplication. The geneamplification model of Coquelle et al. (1997) predicted that gene amplification occurs by breakage-fusion-bridge (BFB) cyclesinvolving fragile sites. Our data suggest that the pattern of gene amplification at 11q13 in OSCC cell lines is consistent witha BFB model. RIN1 appears to be a valuable probe for investigating the process of gene amplification in general and, specifically,11q13 amplification in oral cancer. Genes Chromosomes Cancer 28:153–163, 2000. © 2000 Wiley-Liss, Inc.
INTRODUCTION
Gene amplification is a common finding in manyhuman cancers. The amplification and subsequentoverexpression of critical genes may play an impor-tant role in the loss of cell cycle control in cancer.Amplification of chromosomal band 11q13 is fre-quently observed in head and neck squamous cellcarcinoma (HNSCC), its subset, oral squamous cellcarcinoma (OSCC), other aerodigestive tract can-cers, and breast carcinomas (Berenson et al., 1989;Roelofs et al., 1993; Williams et al., 1993; Jin et al.,1995, 1998; Lese et al., 1995; Rubin et al., 1995;Speicher et al., 1995; Åkervall et al., 1997; Muller etal., 1997; Bekri et al., 1998). A number of geneshave been shown to be amplified in this region,including CCND1 (encoding cyclin D1), EMS1 (en-coding human cortactin, an actin-binding proteinpossibly involved in the organization of the cy-toskeleton and cell adhesion structures), and FGF3
and FGF4 (fibroblast growth factors 3 and 4, alsocalled INT2 and HSTF1; Fig. 1) (Schuuring et al.,1998). A proximal gene, VEGFB, was not found tobe amplified in mammary carcinoma cell lines(Paavonen et al., 1996). However, more distal
This study was carried out in part in the University of PittsburghCancer Institute Cytogenetics Facility.
Christa M. Lese’s current address is Department of Human Ge-netics, University of Chicago, Chicago, Illinois.
Michele I. Shuster’s current address is Department of BiologicalSciences, University of Pittsburgh, Pittsburgh, Pennsylvania.
Supported by: NIH; Grant numbers: F32DE05707 (to MS),RO1DE10513 and RO1DE12008 (to SMG), RO1CA56301 (to JC),DE10598 (to N-HP), and P30CA47049019 and S10RR11879;Smokeless Tobacco Research Council; Grant numbers: 0466, 0660(to SMG); PHS; Grant number: CA40046 (to MML).
*Correspondence to: Dr. Susanne M. Gollin, Department of Hu-man Genetics, University of Pittsburgh Graduate School of PublicHealth, 130 De Soto Street, Pittsburgh, PA 15261.E-mail: [email protected]
Received 25 August 1999; Accepted 4 November 1999
GENES, CHROMOSOMES & CANCER 28:153–163 (2000)
© 2000 Wiley-Liss, Inc.

markers and genes, such as GARP at 11q13.5–14.1,have been shown to be amplified in breast carcino-mas (Bekri et al., 1998). Of the amplified genes,CCND1 is thought to play a direct role in oralcancer. This conclusion is based on the observationthat CCND1 is overexpressed at both the transcriptand protein levels (Bartkova et al., 1995), and am-plification and/or overexpression are associatedwith a more rapid and frequent recurrence of dis-ease (Michalides et al., 1995) and poorer survival(Åkervall et al., 1997). CCND1 is also overexpressedin some parathyroid adenomas as the result of achromosomal inversion (Motokura et al., 1991) andin some B-cell lymphomas as the result of a trans-location (Withers et al., 1991). Overexpression ofCCND1 may lead to premature cycling through theG1/S transition, resulting in propagation of unre-paired DNA damage, accumulation of genetic er-rors, and a selective growth advantage for the al-tered cells. Cyclin D1 overexpression also appearsto alter the sensitivity of tumor cells to ionizingradiation (Coco Martin et al., 1999). It is still un-known whether other, as yet uncharacterized genesin the 11q13 region may be overexpressed and con-tribute to oral tumor development and/or progression.
Human RIN1 was identified in a yeast-basedselection for suppressors of activated RAS (Coli-celli et al., 1991; Han and Colicelli, 1995). Subse-quent analysis demonstrated that RIN1 directlyand specifically binds activated RAS in vitro and invivo (Han et al., 1997). Through a separate domain,RIN1 has also been shown to interact with andserve as a substrate for the tyrosine kinase ABL. Inaddition, overexpression of RIN1 can enhance thetransforming and leukemogenic properties of theCML- and ALL-associated BCR/ABL fusion pro-teins (Afar et al., 1997). The intimate relationshipof RIN1 with both the RAS and ABL oncoproteinssuggested the possibility of a direct or indirect rolefor RIN1 in naturally occurring tumors and
prompted studies into possible chromosomal alter-ations affecting the RIN1 gene. Here we report themapping of RIN1 to 11q13.2 and provide additionallocalization information relative to other genes inthis region. Examination of the RIN1 locus andRIN1 protein expression levels in OSCC cell linesis also presented.
Although gene amplification is observed fre-quently in cancer, the mechanisms underlying theprocess are still not well understood (Schwab,1998). The outcome of gene amplification can beobserved as either extrachromosomal amplification,in the form of double-minute chromosomes(dmin), or intrachromosomal amplification, in theform of homogeneously staining regions (hsrs). Inthe case of 11q13 amplification in HNSCC, intra-chromosomal amplification at the entopic 11q13site seems to be the preferred route, as evidencedby the presence of hsrs containing 11q13 se-quences localized to chromosome 11 (Roelofs et al.,1993; Lese et al., 1995; Jin et al., 1998) or frequentduplication of 11q13 (Jin et al., 1993; Van Dyke etal., 1994). Double-minute chromosomes are occa-sionally observed in HNSCC/OSCC (Jin et al.,1995). Although many models of gene amplifica-tion involve chromosome breakage as an initiatingevent (Windle et al., 1991; Stark, 1993; Coquelle etal., 1997; Pipiras et al., 1998), models involvingsubsequent reintegration of extrachromosomal ele-ments (Windle et al., 1991) are less likely to ac-count for the observation of entopic 11q13 ampli-fication in HNSCC (Roelofs et al., 1993).Nevertheless, such models suggesting homologousreintegration of ectopically amplified sequenceshave been suggested (Windle et al., 1991). Studiesof gene amplification following selection by lowlevels of clastogenic drugs have suggested a modelfor intrachromosomal gene amplification involvingan initial distal breakage event, perhaps at a fragilesite, followed by sister-chromatid fusion at thebreak and proximal breakage of the dicentricbridge at mitosis (Coquelle et al., 1997). Cycles ofbreakage-fusion-bridge (BFB) events would resultin intrachromosomal gene amplification, in theform of inverted duplications of the amplicon, withthe size of the amplicon determined by the sites ofbreakage. Markers near the site of breakage of thebridge may be amplified to a lesser extent than theselected gene. Cycles of BFB with random bridgebreakage can result in low-level amplification ofone marker flanking high-level amplification of anadjacent marker (Toledo et al., 1992). According toCoquelle et al. (1997), three fragile sites have beenlocalized to the region of 11q13 amplification:
Figure 1. The linear order of selected genes and markers from the11q13–q14 region and their corresponding chromosomal band loca-tions from the recent chromosome 11 integrated map (Gerhard andDebelenko, 1999). Asterisk: RIN1 is not on the integrated map, but iscontained on a cosmid that overlaps with HNP36, which is on the map;double asterisks: not on the integrated map.
154 SHUSTER ET AL.

FRA11A at 11q13.3 (Perucca-Lostanlen et al.,1997), FRA11F at 11q14.2, and FRA11H at 11q13.FRA11A maps between RIN1 and CCND1 (Fig. 1).The study of gene amplification in cancer thus maynot only identify potentially important genes, butmay also contribute to an understanding of theunderlying in vivo mechanisms. Analysis of thefrequency and pattern of RIN1 amplification inOSCC cell lines indicates that RIN1 may be avaluable marker for investigating the mechanism ofgene amplification.
MATERIALS AND METHODS
Cell Culture and Cytogenetics
OSCC cell lines were established from tumorbiopsies obtained from patients undergoing surgi-cal resection of their tumors after informed consentunder a protocol approved by the University ofPittsburgh Biomedical Institutional Review Board(data not shown). The clinical and personal char-acteristics of the patients from whom the tumorswere excised and the histopathologic features ofthe tumors from which the cell lines were derivedare listed in Table 1. Tumors were washed inHanks’ Balanced Salt Solution (Irvine Scientific,Santa Ana, CA) containing antibiotics, then mincedinto 1-mm3 pieces that were cultured at 37°C in ahumidified 5% CO2 incubator in MEM medium(Gibco-BRL, Gaithersburg, MD) with 10% fetalbovine serum (Irvine Scientific, Santa Ana, CA)and antibiotics. Normal human oral keratinocytes(NHOKs) were obtained from healthy volunteersand cultured as previously described (Park et al.,1991).
The OSCC cell lines used in this study havebeen assessed previously for 11q13 amplificationby FISH with an INT2, HSTF1, or CCND1 probe
(Oncor, Gaithersburg, MD; D.J. Demetrick, Uni-versity of Calgary, Canada) (Lese et al., 1995).OSCC cell lines were harvested for cytogeneticstudies following exposure to 0.1 or 0.02 mg/mlColcemid (Irvine Scientific, Santa Ana, CA) for 5 or16 hr, respectively. After removal from flasks, cellswere treated with a hypotonic 0.075-M KCl solu-tion for 18 min at 37°C, then fixed with a 3:1methanol:acetic acid fixative for 20 min at roomtemperature. Cells were washed two more times inthis fixative, and fixed cell pellets were stored at220°C prior to use.
Initial Mapping of RIN1
Fluorescence in situ chromosomal hybridization(FISH) was performed on human metaphase cellsprepared from phytohemagglutinin (PHA)-stimu-lated peripheral blood lymphocytes. The RIN1probe was a 5-kb cDNA probe, isolated from ahuman glioblastoma cDNA library (Colicelli et al.,1991). FISH was performed as described previ-ously (Rowley et al., 1990). The RIN1 probe wasbiotinylated by nick-translation using bio-11-dUTP (Enzo Diagnostics, Farmingdale, NY). Hy-bridization was detected with fluorescein-conju-gated avidin (Vector Laboratories, Burlingame,CA), and chromosomes were identified after stain-ing with 4,6-diamidino-2-phenylindole-dihydro-chloride (DAPI).
Isolation of RIN1 Cosmids
Primers were designed to amplify a 150-bp se-quence within the 39 UTR of human RIN1(RIN.1U-CTGAGCCTGCTGAGAAATCC andRIN.1D-CGAAGGCCCCTCATCTTTAT). PCRamplification of RIN1 from a chromosome 11 so-matic cell hybrid mapping panel DNA using pre-
TABLE 1. Clinical and Personal Characteristics of the Patients and Pathologic Features of Their Tumors From Which the CellLines Were Derived
Patient(UPCI:) Age (yrs) Gender Smoker Site
Pathologicalstage Differentiation
SCC125 77 Female Yes Alveolar ridge 4 moderateSCC040 50 Male No Tongue 4 moderateSCC056 76 Male Yes Tongue 4 moderateSCC070 34 Female Yes Retromolar trigone 4 moderateSCC074 51 Female No Mandible 4 poorSCC078 60 Male No Floor of mouth 2 moderateSCC081 87 Female Yes Alveolar ridge 4 wellSCC084 52 Male Yes Retromolar trigone 4 moderateSCC089 58 Male Yes Tongue 4 moderateSCC131 73 Male Yes Floor of mouth 4 wellSCC136 64 Female Yes Retromolar trigone 4 moderate
155GENE AMPLIFICATION IN ORAL CANCER

viously described protocols (Sawicki et al., 1997)confirmed localization to 11q13. This analysis alsodemonstrated that the primers were human-spe-cific and did not span a large intron. These primerswere used to screen an arrayed chromosome 11cosmid library (Wood et al., 1996). PCR amplifica-tion of DNA pools identified the 96-well platescontaining the RIN1 cosmids. Filters stamped fromthe plates were screened by hybridization with anRIN1 cDNA fragment to identify the address of thespecific RIN1 cosmids (LA11NC01-9C6 andLA11NC01-41C9). Cosmid DNA was isolated byanion exchange chromatography (Qiagen, Chats-worth, CA). Restriction fragment analysis and PCRamplification revealed two overlapping cosmidsthat contained the RIN1 sequence.
RIN1 Copy Number Assessment and RelativeMapping by FISH
Within 48 hr of a scheduled FISH hybridization,slides were prepared from the fixed cell pellets. Acontrol slide prepared from normal PHA-stimu-lated lymphocytes was hybridized in parallel withevery set of experimental FISH slides. Cytogeneticpreparations of normal lymphocytes were used forthe relative mapping of RIN1, CCND1, and INT2.
RIN1 and CCND1 probes were prepared fromcosmids (RIN1-LA11NC01-41C9; the CCND1 cos-mid was generously provided by D.J. Demetrick,University of Calgary, Canada) by nick translationin the presence of biotin-16-dUTP or digoxigenin-11-dUTP (Boehringer-Mannheim, Indianapolis,IN) using standard protocols (Lichter and Ried,1994), and then precipitated in the presence ofCot-1 DNA (Gibco-BRL, Gaithersburg, MD). Bi-otin- or digoxigenin-labeled INT2 and chromosome11 centromere (D11Z1) probes were obtained fromOncor (Gaithersburg, MD). Dual- or three-colorFISH was carried out as described by Lichter andRied (1994). Briefly, slides were aged in 2 3 SCCat 37°C, pretreated with RNase for 30 min at 37°C,then with pepsin for 5 min at 37°C, followed bypostfixation in 1% formaldehyde. Slides were de-natured for 2 min at 70°C in 70% formamide/2 3SSC, then dehydrated and air-dried prior to hybrid-ization; 100 ng of each probe was used in a 10-mlhybridization mixture of 50% formamide/2 3 SSC/10% dextran sulfate, and denatured and prean-nealed prior to overnight hybridization at 37°C.Slides were washed in 50% formamide/2 3 SSC at45°C, followed by 2 3 SSC at 37°C, prior to de-tection with 5 mg/ml each FITC-conjugated avidin(Vector Laboratories) and rhodamine-conjugatedantidigoxigenin Fab fragments (Boehringer-Mann-
heim). Slides were counterstained with DAPI inantifade solution (Oncor) and stored at 220°C priorto viewing.
Slides were analyzed with an Olympus BHS flu-orescence microscope and images were capturedusing a CytoVision Ultra (Applied Imaging, SantaClara, CA). For the relative mapping studies, hy-bridization was carried out using dual-color hybrid-izations to metaphase chromosomes and three-color hybridizations to interphase nuclei. In thethree-color experiments, the RIN1 cosmid was la-beled with digoxigenin and detected with rhoda-mine, a red fluorochrome; the CCND1 cosmid waslabeled with biotin and detected with FITC, agreen fluorochrome; and the INT2 probe consistedof a 1:1 mixture of digoxigenin- and biotin-labeledprobes, resulting in a yellow-orange color followingdetection. The relative interphase distance be-tween the centers of these hybridization signalswas measured using the CytoVision Ultra measure-ment software (Applied Imaging).
For the RIN1 amplification studies, the copynumber of chromosome 11 was determined by thenumber of signals from the centromeric probe, andthe relative copy number of RIN1 was then as-sessed. If the number of RIN1 signals (either sin-glets or closely apposed G2 doublets) was the sameas the number of D11Z1 signals, then no amplifi-cation was scored. For copy number gain of RIN1,two patterns were observed. Low-level amplifica-tion/duplication was scored when there were moreRIN1 signals than D11Z1 signals. To assess thelevel of amplification, RIN1 and D11Z1 signalswere counted in 100 cells from each of the threetetrasomic cell lines. These three cell lines werechosen for quantitation because they appeared toexhibit low-level amplification of RIN1. The aver-age number of RIN1 signals plus and minus twostandard deviations was found to range from 3.2 to11.6, with RIN1/D11Z1 ratios ranging from a low of1.29 to a high of 1.845. Cell lines were then scoredby inspection, as this pattern of low-level amplifi-cation was easily distinguishable from high-levelamplification. High-level amplification was quanti-tated by counting the number of INT2 spots in 100cells from one tetrasomic, INT2-amplified cell line.The average number of INT2 signals plus and mi-nus two standard deviations was found to rangefrom 14.36 to 33.04, with an INT2/D11Z1 ratio of5.9. The pattern of hybridization on visual inspec-tion was also distinct, consisting of a large array ofspots in interphase nuclei, and a large, tight clusteron metaphase chromosomes. Thus, any similar pat-tern of hybridization observed for RIN1 was scored
156 SHUSTER ET AL.

as high-level amplification. RIN1 duplication wasassessed only on metaphase chromosomes.
Western Blot Analysis
Confluent 100-mm plates of cells were rinsedwith cold PBS and lysed with 1-ml lysis buffer asdescribed previously (Han et al., 1997). The totalprotein concentration of each extract was measuredby Bradford assay and all were then adjusted to 1mg per ml with lysis buffer. One-mg total proteinfrom each extract was immunoprecipitated with acombination of 2.5 mg of polyclonal anti-RIN1(Transduction Laboratories, Lexington, KY) and15 mg of a separate polyclonal antibody directed tothe amino terminus of RIN1 (Han et al., 1997).After immunoprecipitation with protein A beads(Pierce, Rockford, IL), each entire sample wasloaded on an 8% polyacrylamide gel and, followingtransfer to nitrocellulose, probed with monoclonalanti-RIN1 (Transduction Laboratories).
RESULTS
RIN1 Maps to 11q13.2
To localize the RIN1 gene, we performed fluo-rescence in situ hybridization of a biotin-labeledcDNA probe to normal human metaphase chromo-somes. Hybridization of this probe resulted in spe-cific labeling only of chromosome 11 (Fig. 2). Spe-cific labeling of 11q13 was observed on four (4cells), three (11 cells), two (12 cells), one (2 cells),or no (1 cell) chromatid(s) of the chromosome 11homologues in 30 cells examined. Of 82 signalsobserved, 75 (91%) were located at 11q13 (75 of120 11q chromatids from 30 metaphase cells were
labeled). Of these, 15 (20%) signals were located at11q13.1, 59 (79%) signals were located at 11q13.2,and 1 (1%) signal was located at 11q13.3. Sevensinglet signals were observed at other chromosomalsites (1q42, 7q11, 7q22, 10p12, 10p14, 14p13, and15p13); no doublet signals were observed at thesebackground sites. We observed specific signal at11q13.2 in a second hybridization experiment us-ing this cDNA probe. These results suggest thatthe RIN1 gene is localized to chromosome 11, bandq13.2.
In an effort to map the position of RIN1 relativeto known amplified genes in 11q13, normal meta-phase chromosomes and interphase nuclei werecohybridized with different combinations of aRIN1 cosmid probe with INT2 and CCND1 probes.Dual-color hybridization of RIN1 and INT2 tometaphase chromosomes shows RIN1 to be proxi-mal to INT2 on 11 of 19 hybridized (58%) chroma-tids from five cells. Seven (37%) chromatidsshowed RIN1 and INT2 to be either superimposedor at the same position on the chromatid, and one(5%) chromatid showed INT2 proximal to RIN1.These results are consistent with RIN1 being prox-imal and closely apposed to INT2 (Fig. 3A). Simi-larly, RIN1 is adjacent and proximal to CCND1, asshown in dual-color hybridizations with RIN1,CCND1, and an 11p telomere probe (Oncor), toconfirm the orientation of chromosome 11 (Fig.3B). Fifty-two of 78 (67%) chromatids from 25 cellsshowed RIN1 proximal to CCND1, 19 (24%)showed the two probes to be superimposed orhybridized to the same location on the chromatid,and 7 (9%) showed CCND1 to be proximal to RIN1.
Figure 2. In situ hybridization of a biotin-labeled RIN1 probe to humanmetaphase cells from phytohemagglutinin-stimulated peripheral blood lym-phocytes. The chromosome 11 homologues are identified with arrows;specific labeling was observed at 11q13.2. The inset shows partial karyo-types of three chromosome 11 homologues illustrating specific labeling at11q13.2. Images were obtained using a Zeiss Axiophot microscope attachedto a charge-coupled device (CCD) camera. Separate images of DAPI-stainedchromosomes and the hybridization signal were merged using image analysissoftware (NU200 and Image 1.57).
157GENE AMPLIFICATION IN ORAL CANCER
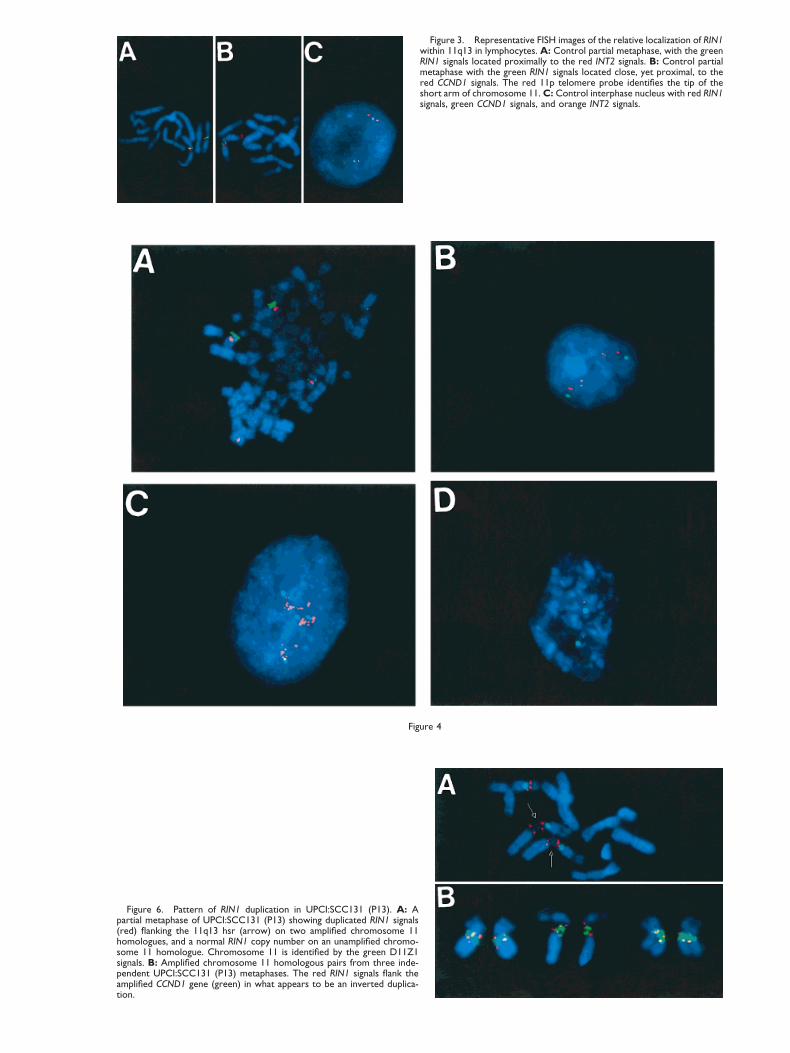
Figure 3. Representative FISH images of the relative localization of RIN1within 11q13 in lymphocytes. A: Control partial metaphase, with the greenRIN1 signals located proximally to the red INT2 signals. B: Control partialmetaphase with the green RIN1 signals located close, yet proximal, to thered CCND1 signals. The red 11p telomere probe identifies the tip of theshort arm of chromosome 11. C: Control interphase nucleus with red RIN1signals, green CCND1 signals, and orange INT2 signals.
Figure 4
Figure 6. Pattern of RIN1 duplication in UPCI:SCC131 (P13). A: Apartial metaphase of UPCI:SCC131 (P13) showing duplicated RIN1 signals(red) flanking the 11q13 hsr (arrow) on two amplified chromosome 11homologues, and a normal RIN1 copy number on an unamplified chromo-some 11 homologue. Chromosome 11 is identified by the green D11Z1signals. B: Amplified chromosome 11 homologous pairs from three inde-pendent UPCI:SCC131 (P13) metaphases. The red RIN1 signals flank theamplified CCND1 gene (green) in what appears to be an inverted duplica-tion.

The close proximity of these three probes pre-cludes determination of relative gene order inthree-probe experiments to metaphase chromo-somes. However, three-color FISH on interphasenuclei permits a relative ordering of RIN1, INT2,and CCND1 (Fig. 3C). Images of interphase nucleihybridized with RIN1, INT2, and CCND1 werecaptured on the CytoVision Ultra (Applied Imag-ing), and the distance between each pair of probeswas measured. The pair-wise distance between sig-nals from 25 hybridized homologues was measuredin 22 nuclei. In 72% of the measurements, therelative gene order was found to be RIN1-CCND1-INT2 and the distance between RIN1 and CCND1was greater than the distance between CCND1 andINT2, and the distance between RIN1 and INT2was approximately the sum of the distances be-tween RIN1 and CCND1 and CCND1 and INT2.Twenty-four percent of the homologues showed arelative gene order of RIN1-INT2-CCND1, and 4%showed superimposition of the INT2 and CCND1signals, such that no order could be determined.These results are consistent with a gene order ofcen-RIN1-CCND1-INT2-tel, with RIN1 being themost centromeric locus, based on the hybridizationpattern on metaphase chromosomes. The place-ment of CCND1 centromeric to INT2 in band11q13.3 is in agreement with the physical mapping
by Brookes et al. (1993) and with the integratedmap illustrated in Figure 1.
RIN1 Copy Number in OSCC
The localization of RIN1 to 11q13.2, its closeproximity to CCND1 and INT2, genes known to beamplified at high frequency in OSCC, as well as itsknown interaction with RAS and ABL suggestedthat RIN1 may also be a target for gene amplifica-tion and overexpression. Our collection of OSCCcell lines has been tested for 11q13 amplificationby FISH using INT2, HSTF1, and/or CCND1probes, along with a chromosome 11 centromereprobe as a control for chromosome copy number.Approximately 50% of our cell lines were found tohave 11q13 amplification (Fig. 4A). Amplificationwas scored as a tight cluster of INT2 and/or CCND1signals on metaphase chromosomes or a more dis-persed array of signals in interphase nuclei. TheOSCC cell lines are frequently polyploid, and inthese cell lines not every homologue of chromo-some 11 exhibits 11q13 amplification. 11q13 am-plification was observed predominantly on chromo-some 11, consistent with the observation of hsrs at11q13 in the karyotypes of these cell lines (data notshown) and reports of intrachromosomal 11q13 am-plification in HNSCC and HNSCC cell lines (Ro-elofs et al., 1993; Lese et al., 1995; Jin et al., 1998).
Ten 11q13-amplified cell lines and one cell linewith three copies of chromosome 11, but no 11q13amplification, were tested for RIN1 amplificationby FISH analysis. Normal lymphocytes were usedas a diploid control. The results are presented inTable 2. A RIN1 cosmid was labeled with digoxi-genin and cohybridized to cytogenetic preparationswith a biotin-labeled D11Z1 centromeric probe.The RIN1 signals were examined for amplificationin approximately 100 cells for each cell line, pre-dominantly in interphase. In order to be considered
Figure 5. Overexpression of RIN1 protein in an OSCC cell line. The leftand right panels represent separate experiments, each including a NHOKcontrol lane. Molecular weight markers are indicated in kDa at left, and theposition of RIN1 protein is indicated at right. The large band of about 50kDa is antibody protein from the immunoprecipitation. RIN1 overexpres-sion in UPCI:SCC089 cells compared to NHOKs was 2.1- and 2.4-fold inthe two experiments performed, as determined by the use of a Phospho-imager. The accuracy of the immunoprecipitation, Western blot procedure,and quantitation were verified using multiple loadings (with increasingamounts) of a positive control sample.
Figure 4. Patterns of 11q13 duplication and amplification in theOSCC cell lines. A: UPCI:SCC040 (P9; passage 9) metaphase with fourhomologues of chromosome 11 (red D11Z1 signals). Two homologueshave entopic CCND1 amplification (large green signals), while twohomologues have a normal CCND1 copy number (two small greensignals). B: Duplication/low-level amplification of RIN1 (red) relative tochromosome 11 (green D11Z1 signals) in a UPCI:SCC089 (P32) di-somic interphase nucleus. C: High-level amplification of RIN1 (red)relative to the copy number of chromosome 11 (green D11Z1 signals)in a UPCI:SCC089 (P32) tetrasomic interphase nucleus. D: Normaldisomic RIN1 copy number (red) relative to chromosome 11 (greenD11Z1 signals) in a UPCI:SCC081 (P2) interphase nucleus.
159GENE AMPLIFICATION IN ORAL CANCER

significant in the analysis, at least 10% of the cellsexamined had to show an abnormal RIN1 copynumber. Two categories of abnormal RIN1 copynumber were identified: low-level amplification/duplication and high-level amplification. Unequiv-ocal duplication of RIN1 was only possible to as-certain on metaphase chromosomes, and thepattern of duplication will be described in moredetail below. The criteria for scoring low- and high-level amplification are described above. Of the 10cell lines with INT2 amplification, five (50%)showed duplication or low-level amplificationof RIN1 (UPCI:SCC040, UPCI:SCC074, UPCI:SCC084, UPCI:SCC131, and UPCI:SCC136; Fig.4B). The duplication of RIN1 in UPCI:SCC084appears to be the result of an extra copy of theRIN1 locus on a chromosome other than chromo-some 11, based on the hybridization pattern ob-served on metaphase spreads. This most likelyrepresents a mechanism of RIN1 gene copy num-ber increase distinct from intrachromosomal 11q13amplification or duplication. Two (20%) OSCC celllines (UPCI:SCC070 and UPCI:SCC089) showedheterogeneity for RIN1 copy number, with sub-populations of cells in each cell line exhibiting low-and high-level (Fig. 4C) RIN1 amplification. Theremaining three (30%) cell lines showed the ex-pected copy number of RIN1 (1:1) relative to thecentromere (UPCI:SCC056, UPCI:SCC078, andUPCI:SCC081; Fig. 4D).
RIN1 Expression in OSCC Cell Lines
To determine whether RIN1 protein is overex-pressed as a result of the observed chromosomal
alterations, we carried out Western blot analysis. Asshown in Figure 5, the cell line UPCI:SCC089showed a moderate but significant increase inRIN1 expression (2.1-fold and 2.4-fold in two sep-arate experiments) compared to NHOKs. This lineshowed RIN1 alterations (both low- and high-levelamplification) in more than 80% of the cells ana-lyzed by FISH. Other OSCC cell lines examined(UPCI:SCC074 and UPCI:SCC136), all with dupli-cation/low-level amplification, did not show anyincrease in RIN1 levels (Fig. 5). Another cell line,UPCI:SCC131, also had no RIN1 overexpression(data not shown). Although the OSCC cell linesdisplayed some aneuploidy, the elevated RIN1 ex-pression was not the result of extra copies of chro-mosome 11, as UPCI:SCC089 is primarily disomicfor this chromosome (Table 2), while one of thenegative lines (UPCI:SCC136) was tetrasomic (Ta-ble 2).
Pattern of RIN1 Duplication: Mechanismof Amplification
During our assessment of RIN1 copy number byFISH in the OSCC cell lines, we observed thatcertain cell lines with what appeared to be dupli-cation or low-level amplification showed a distinctpattern of RIN1 duplication on metaphase chromo-somes. The RIN1 signals appeared to flank the hsrat 11q13 (Fig. 6A), which is consistent with aninverted RIN1 duplication. This pattern was seenon metaphase chromosomes from four cell lines(UPCI:SCC040, UPCI:SCC070, UPCI:SCC074,and UPCI:SCC131). Cohybridization of CCND1with RIN1 confirmed that the RIN1 signals lie
TABLE 2. RIN1 Amplification and Expression in OSCC Cell Lines
Cell Line (UPCI:)(P; passage number)
Chromosome 11 copynumber (%)a
11q13amplificationb
Number of cellsscored RIN1
% RIN1 Duplication/low-level
amplification
% RIN1high-level
amplificationRIN1 proteinexpression
Lymphocytes 2 (99%) 2 212 0 0SCC125 P8 3 (92%) 2 123 0 0 NDd
SCC040 P9 4 (85%) 1 99 90 0 NDSCC056 P5 2 (99%) 1 120 0 0 NDSCC070 P20 4 (86%) 1 96 70 19 NDSCC074 P5 2 (92%) 1 109 30 0 1Xe
SCC078 P40 3 (96%) 1 103 0 0 NDSCC081 P2 2 (99%) 1 102 0 0 NDSCC084 P3 2 (70%) 1 119 27c 0 NDSCC089 P32 2 (72%) 1 97 40 43 ; 2XSCC131 P13 4 (92%) 1 103 10 0 1XSCC136 P13 4 (99%) 1 96 26 0 1X
aChromosome 11 copy number determined by alpha-satellite enumeration in RIN1/D11Z1 hybridizations.b11q13 amplification previously determined by INT2, HSTF1, and/or CCND1 FISH.cDuplication of RIN1 due to an extra copy on a chromosome other than 11.dND, not done.eLevel of RIN1 protein relative to RIN1 levels in normal human oral keratinocytes (1X 5 same level as oral keratinocytes).
160 SHUSTER ET AL.

immediately adjacent to either side of the 11q13hsr in UPCI:SCC131, which includes the CCND1gene (Fig. 6B). In UPCI:SCC070, there were sub-populations of cells with both duplication/low-levelamplification and high-level RIN1 amplification.The high-level amplification of RIN1 in this cellline appeared to be localized to the 11q13 hsr,based on the hybridization pattern to metaphasechromosomes. Thus, RIN1 is included in the am-plification process at 11q13 in some instances,while in other cases, RIN1 appears not to be coam-plified with INT2/CCND1.
DISCUSSION
FISH analysis of RIN1 copy number in 11q13-amplified OSCC cell lines showed moderately highlevels of amplification in 20%, and low levels ofamplification (duplication) in 40%. Western blot-ting showed only an approximately 2–3-fold eleva-tion in RIN1 levels in one of the cell lines exam-ined (UPCI:SCC089). This is consistent with atwofold increase in the RIN1 gene (duplication)found in approximately 40% of the cell population(Table 2), but not commensurate with the RIN1amplification that was detected in 43% of the samepopulation. This apparent discrepancy could resultfrom a disruption of the amplified RIN1 gene thatprevents its normal expression. Feedback mecha-nisms that keep RIN1 expression below a thresh-old level may also be involved. Further analysis ofthe RIN1 promoter may shed light on this obser-vation. However, analysis of the RIN1 hybridiza-tion pattern on metaphase chromosomes preparedfrom OSCC cell lines revealed a striking pattern.The RIN1 signals appear to flank the 11q13 hsr, inan inverted duplication, very similar to the patternobserved with the P3C4 probe flanking the se-lected ampd2 amplification in GMA32 cells (Tole-do et al., 1992). Thus, although not overexpressedto high levels in OSCC, RIN1 may serve as amarker for endogenous sites of chromosomalbreakage resulting in gene amplification in cancer.
Intrachromosomal amplification has been specu-lated to depend on breakage of chromosomes at aminimum of two sites (Coquelle et al., 1997; Pipi-ras et al., 1998), with the distance between thesesites determining the size and extent of the ampli-fied region. Preamplification breakage has beenaccomplished in experimental models with clasto-genic drugs and introduction of sites for rare-cut-ting restriction enzymes, but models of in vivogene amplification have suggested that breakagemay be the result of fragile site expression. In vitroanalysis of gene amplification induced by clasto-
genic drugs in GMA32 cells has mapped sites ofdrug-induced chromosomal breakage adjacent toamplified mdr1 and ampd2 genes and identified amarker (P3C4) near these sites (Toledo et al., 1992;Coquelle et al., 1997). The hybridization patternobserved for the P3C4 marker at early stages ofampd2 amplification in GMA32 cells is strikinglysimilar to that seen with RIN1 and 11q13 amplifi-cation in our OSCC cell lines, with P3C4 flankingthe region of amplification (Toledo et al., 1992). Amodel for breakage-fusion-bridge cycles that couldlead to the chromosomes 11 shown in Figure 6Aand B is shown in Figure 7. The model wouldpredict that an initial break distal to CCND1, pos-
Figure 7. A model for 11q13 amplification in OSCC cell lines. A:Chromosome 11 is represented by a line, with RIN1 (filled rectangle)and CCND1 (open rectangle) shown on the long arm (filled circlerepresents the centromere). An initial break (arrow) distal to CCND1could lead to chromosome fusion after replication, resulting in a dicen-tric chromosome with two copies each of RIN1 and CCND1. B: Thisdicentric chromosome could then undergo one cycle of BFB, with thebreak (arrow) occurring between RIN1 and CCND1. Replication andfusion would create a dicentric chromosome with four copies ofCCND1 and two copies of RIN1. C: Continued cycles of BFB would leadto amplification of CCND1, flanked single copies of RIN1. As the chro-mosome 11 homologues do not appear to be dicentric by DAPI stainingand have a large amount of chromatin distal to the amplicon, a trans-location may have occurred to introduce chromatin distal to the am-plicon. Although chromosome breaks have been illustrated as occurringduring the G1 phase of the cell cycle, chromosome breaks occurringduring the G2 phase, followed by chromatid fusion, could also give riseto breakage-fusion-bridge cycles.
161GENE AMPLIFICATION IN ORAL CANCER

sibly at a fragile site, could initiate the process, andthen breakage at a fragile site between RIN1 andCCND1 would promote the BFB cycles. Based onthe most recent chromosome 11 map, FRA11A islocated less than 100 to 200 kb distal to RIN1,between RIN1 and CCND1, which are approxi-mately 4 to 5 Mb apart (Gerhard and Debelenko,1999). Thus, FRA11A may be involved in geneamplification in oral cancer. Further evidence tosupport the occurrence of a second break distal toCCND1 is the observation that 11q13 amplificationis usually accompanied by distal deletions (Jin etal., 1998). Studies designed to map the 11q13 am-plicon further and examine the mechanism of geneamplification in OSCC cells are warranted.
ACKNOWLEDGMENTS
The authors are grateful to Ms. Seia Comsa, Ms.Jaya Reddy, and Mr. Apoor Patel for excellenttechnical assistance with the cell culture and cellharvesting; Dr. Daniela S. Gerhard for clarificationof the chromosome 11 gene map; and Drs. Jonas T.Johnson and Eugene N. Myers for their enthusias-tic support of the project and oral cancer genetics.
REFERENCES
Afar DE, Han L, McLaughlin J, Wong S, Dhaka A, Parmar K,Rosenberg N, Witte ON, Colicelli J. 1997. Regulation of theoncogenic activity of BCR-ABL by a tightly bound substrateprotein RIN1. Immunity 6:773–782.
Åkervall J, Michalides RJAM, Mineta H, Balm A, Borg Å, DictorMR, Jin Y, Loftus B, Mertens F, Wennerberg JP. 1997. Amplifi-cation of cyclin D1 in squamous cell carcinoma of the head andneck and the prognostic value of chromosomal abnormalities andcyclin D1 overexpression. Cancer 79:380–389.
Bartkova J, Lukas J, Muller H, Strauss M, Gusterson B, Bartek J.1995. Abnormal patterns of D-type cyclin expression and G1regulation in human head and neck cancer. Cancer Res 55:949–956.
Bekri S, Adelaıde J, Merscher S, Grosgeorge J, Caroli-Bosc J, Pe-rucca-Lostanlen D, Kelley PM, Pebusque MJ, Theillet C, Birn-baum D, Gaudray P. 1998. Detailed map of a region commonlyamplified at 11q13–q14 in human breast carcinoma. CytogenetCell Genet 79:125–131.
Berenson JR, Yang J, Mickel RA. 1989. Frequent amplification ofthe bcl-1 locus in head and neck squamous cell carcinoma. Onco-gene 4:1111–1116.
Brookes S, Lammie A, Schuuring E, de Boer C, Michalides R,Dickson C, Peters G. 1993. Amplified region of chromosome band11q13 in breast and squamous cell carcinomas encompasses threeCpG islands telomeric of FGF3, including the expressed geneEMS1. Genes Chromosomes Cancer 6:222–231.
Coco Martin JM, Balkenende A, Verschoor T, Lallemand F, Micha-lides R. 1999. Cyclin D1 overexpression enhances radiation-in-duced apoptosis and radiosensitivity in a breast tumor cell line.Cancer Res 59:1134–1140.
Colicelli J, Nicolette C, Birchmeier C, Rodgers L, Riggs M, WiglerM. 1991. Expression of three mammalian cDNAs that interferewith RAS function in Saccharomyces cerevisiae. Proc Natl AcadSci USA 88:2913–2917.
Coquelle A, Pipiras E, Toledo F, Buttin G, Debatisse M. 1997.Expression of fragile sites triggers intrachromosomal mammaliangene amplification and sets boundaries to early amplicons. Cell89:215–225.
Gaudray P, Carle GF, Gerhard DS, et al. 1999. Report of the SixthInternational Workshop on human chromosome 11 mapping 1998.
Nice, France, May 2–5, 1998. Cytogenet Cell Genet 86(3–4):167–186.
Han L, Colicelli J. 1995. A human protein selected for interferencewith Ras function interacts directly with Ras and competes withRaf1. Mol Cell Biol 15:1318–1323.
Han L, Wong D, Dhaka A, Afar D, White M, Xie W, Herschman H,Witte O, Colicelli J. 1997. Protein binding and signaling proper-ties of RIN1 suggest a unique effector function. Proc Natl AcadSci USA 94:4954–4959.
Jin Y, Mertens F, Mandahl N, Heim S, Olegård C, Wennerberg J,Biorklund A, Mitelman F. 1993. Chromosome abnormalities ineighty-three head and neck squamous cell carcinomas: influenceof culture conditions on karyotypic pattern. Cancer Res 53:2140–2146.
Jin Y, Mertens F, Jin C, Åkervall J, Wennerberg J, Gorunova L,Mandahl N, Heim S, Mitelman F. 1995. Nonrandom chromosomeabnormalities in short-term cultured primary squamous cell car-cinomas of the head and neck. Cancer Res 55:3204–3210.
Jin Y, Hoglund M, Jin C, Martins C, Wennerberg J, Åkervall J,Mandahl N, Mitelman F, Mertens F. 1998. FISH characterizationof head and neck carcinomas reveals that amplification of band11q13 is associated with deletion of distal 11q. Genes Chromo-somes Cancer 22:312–320.
Lese CM, Rossie KM, Appel BN, Reddy JK, Johnson JT, MyersEN, Gollin SM. 1995. Visualization of INT2 and HST1 amplifica-tion in oral squamous cell carcinoma. Genes Chromosomes Can-cer 12:288–295.
Lichter P, Ried T. 1994. Molecular analysis of chromosome aberra-tions-in situ hybridization. Methods Mol Biol 29:449–478.
Michalides R, van Veelen N, Hart A, Loftus B, Wientjens E, BalmA. 1995. Overexpression of cyclin D1 correlates with recurrence ina group of forty-seven operable squamous cell carcinomas of thehead and neck. Cancer Res 55:975–978.
Motokura T, Bloom T, Goo Kim H, Juppner H, Ruderman J,Kronenberg HM, Arnold A. 1991. A novel cyclin encoded by abcl1-linked candidate oncogene. Nature 350:512–515.
Muller D, Millon R, Velten M, Bronner G, Jung G, Engelman A,Flesch H, Eber M, Methlin G, Abecassis J. 1997. Amplification of11q13 DNA markers in head and neck squamous cell carcinomas:correlation with clinical outcome. Eur J Cancer 33:2203–2210.
Paavonen K, Horelli-Kuitunen N, Chilov D, Kukk E, Pennanen S,Kallioniemi O-P, Pajusola K, Olofsson B, Eriksson U, Joukov V,Palotie A, Alitalo K. 1996. Novel human vascular endothelialgrowth factor genes VEGF-B and VEGF-C localize to chromo-somes 11q13 and 4q34, respectively. Circulation 93:1079–1082.
Park N-H, Min B-M, Li S-L, Huang MZ, Cherick HM, Doniger J.1991. Immortalization of normal human oral keratinocytes withtype 16 human papillomavirus. Carcinogenesis 12:1627–1631.
Perucca-Lostanlen D, Hecht BK, Courseaux A, Grosgeorge J, HechtF, Gaudray P. 1997. Mapping FRA11A, a folate-sensitive fragilesite in human chromosome band 11q13.3. Cytogenet Cell Genet79:88–91.
Pipiras E, Coquelle A, Bieth A, Debatisse M. 1998. Interstitialdeletions and intrachromosomal amplification initiated from adouble-strand break targeted to a mammalian chromosome.EMBO J 17:325–333.
Roelofs H, Schuuring E, Wiegant J, Michalides R, Giphart-GasslerM. 1993. Amplification of the 11q13 region in human carcinomacell lines: a mechanistic view. Genes Chromosomes Cancer 7:74–84.
Rowley JD, Diaz MO, Espinosa R, Patel YD, van Melle E, ZieminS, Taillon-Miller P, Lichter P, Evans GA, Kersey JD, Ward DC,Domer PH, Le Beau MM. 1990. Mapping chromosome band11q23 in human acute leukemia with biotinylated probes: Iden-tification of 11q23 translocation breakpoints with a yeast artificialchromosome. Proc Natl Acad Sci USA 87:9358–9362.
Rubin JS, Qiu L, Etkind P. 1995. Amplification of the Int-2 gene inhead and neck squamous cell carcinoma. J Laryngol Otol 109:72–76.
Sawicki M, Arnold E, Ebrahimi S, Duell T, Jin S, Wood T, Chakra-bati R, Peters J, Wan Y, Samara G, Weier HU, Udar N, Passaro EJr, Srivatsan ES. 1997. A transcript map encompassing the multi-ple endocrine neoplasia type-1 (MEN1) locus on chromosome11q13. Genomics 42:405–412.
Schuuring E, van Damme H, Schurring-Scholtes E, Verhoeven E,Michalides R, Geelen E, de Boer C, brok H, van Buuren C, KluinP. 1998. Characterization of the EMS1 gene and its product,human Cortactin. Cell Adhesion Commu 6:185–209.
Schwab M. 1998. Amplification of oncogenes in human cancer cells.Bioessays 20:473–479.
162 SHUSTER ET AL.

Serrano M, Hannon GJ, Beach D. 1993. A new regulatory motif incell-cycle control causing specific inhibition of cyclin D/CDK4.Nature 366:704–707.
Speicher MR, Howe C, Crotty P, du Manoir S, Costa J, Ward DC.1995. Comparative genomic hybridization detects novel deletionsand amplifications in head and neck squamous cell carcinoma.Cancer Res 55:1010–1013.
Stark G. 1993. Regulation and mechanisms of mammalian geneamplification. Adv in Cancer Res 61:87–113.
Toledo F, Le Roscouet D, Buttin G, Debatisse M. 1992. Co-amplifiedmarkers alternate in megabase long chromosomal inverted repeatsand cluster independently in interphase nuclei and early steps ofmammalian gene amplification. EMBO J 11:2665–2673.
Van Dyke DL, Worsham MJ, Benninger MS, Krause CJ, Baker SR,Wolf GT, Drumheller T, Tilley BC, Carey TE. 1994. Recurrentcytogenetic abnormalities in squamous cell carcinomas of thehead and neck region. Genes Chromosomes Cancer 9:192–206.
Williams ME, Gaffey MJ, Weiss LM, Wilczynski SP, Schuuring E,Levine PA. 1993. Chromosome 11q13 amplification in head andneck squamous cell carcinoma. Arch Otolaryngol Head Neck Surg119:1238–1243.
Windle B, Draper BW, Yin Y, O’Gorman S, Wahl GM. 1991. Acentral role for chromosome breakage in gene amplification, de-letion formation, and amplicon integration. Genes Dev 5:160–174.
Withers DA, Harvey RC, Faust JB, Melnyk O, Carey K, MeekerTC. 1991. Characterization of a candidate bcl-1 gene. Mol CellBiol 11:4846–4853.
Wood TF, Srivatsan ES, Chakrabarti R, Ma GC, Kuan N, SamaraGJ, Higgins MJ, Shows TB, Johnson CL, Wan YJ, Passaro EP Jr,Sawicki MP. 1996. A 1.5-megabase physical map encompassingthe multiple endocrine neoplasia type-1 (MEN1) locus on chro-mosome 11q13. Genomics 38:166–173.
163GENE AMPLIFICATION IN ORAL CANCER




















