
3D nuclear architecture reveals coupled cell cycle dynamics of chromatin and nuclear pores in the...
11
3D nuclear architecture reveals coupled cell cycle dynamics of chromatin and nuclear pores in the malaria parasite Plasmodium falciparum Allon Weiner, 2† Noa Dahan-Pasternak, 1† Eyal Shimoni, 3 Vera Shinder, 3 Palle von Huth, 3 Michael Elbaum 2 and Ron Dzikowski 1 * 1 Department of Microbiology and Molecular Genetics, The Kuvin Center for the Study of Infectious and Tropical Diseases, IMRIC, The Hebrew University-Hadassah Medical School, Jerusalem 91120, Israel. 2 Department of Materials and Interfaces and 3 Electron Microscopy Unit, Weizmann Institute of Science, Rehovot 76100, Israel. Summary The deadliest form of human malaria is caused by the protozoan parasite Plasmodium falciparum. The complex life cycle of this parasite is associ- ated with tight transcriptional regulation of gene expression. Nuclear positioning and chromatin dynamics may play an important role in regulating P. falciparum virulence genes. We have applied an emerging technique of electron microscopy to construct a 3D model of the parasite nucleus at distinct stages of development within the infected red blood cell. We have followed the distribution of nuclear pores and chromatin throughout the intra- erythrocytic cycle, and have found a striking cou- pling between the distributions of nuclear pores and chromatin organization. Pore dynamics involve clustering, biogenesis, and division among daughter cells, while chromatin undergoes stage- dependent changes in packaging. Dramatic changes in heterochromatin distribution coincide with a previously identified transition in gene expression and nucleosome positioning during the mid-to-late schizont phase. We also found a corre- lation between euchromatin positioning at the nuclear envelope and the local distribution of nuclear pores, as well as a dynamic nuclear polarity during schizogony. These results suggest that cyclic patterns in gene expression during parasite development correlate with gross changes in cellular and nuclear architecture. Introduction The spatial organization of DNA into nuclear sub- compartments is a dynamic process that involves changes in chromatin organization during the cell cycle (Chakalova et al., 2005; Hakim et al., 2010; Babu et al., 2008). Many studies have demonstrated a role of nuclear organization in gene expression (Sexton et al., 2007). Gene positioning in particular has been implicated in tran- scriptional regulation. At one level this involves the con- version or movement of silenced genes from regions of condensed heterochromatin to less compact euchromatin that permits access to the transcription machinery. At a second level, it may involve reorganization of the euchromatin/heterochromatin mixture spatially within the nucleus. Furthermore, it was proposed that the nucleus possesses multiple foci where the transcriptional machin- ery resides, and that active loci may move into these ‘transcriptional factories’. Special attention was given to the active foci that reside at the nuclear periphery (Brown and Silver, 2007; Akhtar and Gasser, 2007). The nuclear envelope (NE) of eukaryotic organisms is composed of two juxtaposed bilayer lipid membranes that separate the nucleus from the cytoplasm. Nuclear pore complexes (NPCs) are embedded in the NE and allow exchange of macromolecules between these compart- ments. It has become clear that components of the nuclear periphery serve as more than a passive boundary and are also involved in chromatin organization and gene expression (Goldman et al., 2002; Gruenbaum et al., 2003). In yeast and animal cells the inner face of the NE is typically occupied by heterochromatic genetic matter that is inactive for transcription. Within the perinuclear heterochromatin there are also islands of loose euchro- matin. Transcriptionally active foci are found adjacent to NPCs, and indeed active genes were shown to interact with components of the NPC (Akhtar and Gasser, 2007). The typical occupancy of the nuclear periphery with Received 20 October, 2010; revised 2 February, 2011; 9 March, 2011; accepted 9 March, 2011. *For correspondence. E-mail rond@ ekmd.huji.ac.il; Tel. (+972) 2 675 8095; Fax (+972) 2 675 7425. † These authors have equal contribution to this manuscript. Cellular Microbiology (2011) 13(7), 967–977 doi:10.1111/j.1462-5822.2011.01592.x First published online 28 Apr 2011 © 2011 Blackwell Publishing Ltd cellular microbiology
Transcript of 3D nuclear architecture reveals coupled cell cycle dynamics of chromatin and nuclear pores in the...

3D nuclear architecture reveals coupled cell cycledynamics of chromatin and nuclear pores in themalaria parasite Plasmodium falciparum
Allon Weiner,2† Noa Dahan-Pasternak,1†
Eyal Shimoni,3 Vera Shinder,3 Palle von Huth,3
Michael Elbaum2 and Ron Dzikowski1*1Department of Microbiology and Molecular Genetics,The Kuvin Center for the Study of Infectious andTropical Diseases, IMRIC, The HebrewUniversity-Hadassah Medical School, Jerusalem 91120,Israel.2Department of Materials and Interfaces and 3ElectronMicroscopy Unit, Weizmann Institute of Science,Rehovot 76100, Israel.
Summary
The deadliest form of human malaria is caused bythe protozoan parasite Plasmodium falciparum.The complex life cycle of this parasite is associ-ated with tight transcriptional regulation of geneexpression. Nuclear positioning and chromatindynamics may play an important role in regulatingP. falciparum virulence genes. We have applied anemerging technique of electron microscopy toconstruct a 3D model of the parasite nucleus atdistinct stages of development within the infectedred blood cell. We have followed the distribution ofnuclear pores and chromatin throughout the intra-erythrocytic cycle, and have found a striking cou-pling between the distributions of nuclear poresand chromatin organization. Pore dynamicsinvolve clustering, biogenesis, and division amongdaughter cells, while chromatin undergoes stage-dependent changes in packaging. Dramaticchanges in heterochromatin distribution coincidewith a previously identified transition in geneexpression and nucleosome positioning during themid-to-late schizont phase. We also found a corre-lation between euchromatin positioning at thenuclear envelope and the local distribution ofnuclear pores, as well as a dynamic nuclear
polarity during schizogony. These results suggestthat cyclic patterns in gene expression duringparasite development correlate with grosschanges in cellular and nuclear architecture.
Introduction
The spatial organization of DNA into nuclear sub-compartments is a dynamic process that involveschanges in chromatin organization during the cell cycle(Chakalova et al., 2005; Hakim et al., 2010; Babu et al.,2008). Many studies have demonstrated a role of nuclearorganization in gene expression (Sexton et al., 2007).Gene positioning in particular has been implicated in tran-scriptional regulation. At one level this involves the con-version or movement of silenced genes from regions ofcondensed heterochromatin to less compact euchromatinthat permits access to the transcription machinery. At asecond level, it may involve reorganization of theeuchromatin/heterochromatin mixture spatially within thenucleus. Furthermore, it was proposed that the nucleuspossesses multiple foci where the transcriptional machin-ery resides, and that active loci may move into these‘transcriptional factories’. Special attention was given tothe active foci that reside at the nuclear periphery (Brownand Silver, 2007; Akhtar and Gasser, 2007).
The nuclear envelope (NE) of eukaryotic organisms iscomposed of two juxtaposed bilayer lipid membranes thatseparate the nucleus from the cytoplasm. Nuclear porecomplexes (NPCs) are embedded in the NE and allowexchange of macromolecules between these compart-ments. It has become clear that components of thenuclear periphery serve as more than a passive boundaryand are also involved in chromatin organization and geneexpression (Goldman et al., 2002; Gruenbaum et al.,2003). In yeast and animal cells the inner face of the NEis typically occupied by heterochromatic genetic matterthat is inactive for transcription. Within the perinuclearheterochromatin there are also islands of loose euchro-matin. Transcriptionally active foci are found adjacent toNPCs, and indeed active genes were shown to interactwith components of the NPC (Akhtar and Gasser, 2007).The typical occupancy of the nuclear periphery with
Received 20 October, 2010; revised 2 February, 2011; 9 March, 2011;accepted 9 March, 2011. *For correspondence. E-mail [email protected]; Tel. (+972) 2 675 8095; Fax (+972) 2 675 7425.†These authors have equal contribution to this manuscript.
Cellular Microbiology (2011) 13(7), 967–977 doi:10.1111/j.1462-5822.2011.01592.xFirst published online 28 Apr 2011
© 2011 Blackwell Publishing Ltd
cellular microbiology

heterochromatin may differ in specific cell types, forexample in the retinal cells of nocturnal mammals in whichheterochromatin is concentrated at the centre of thenucleus (Solovei et al., 2009), yet euchromatin remainsassociated with the NPCs.
Nuclear dynamics may play a key role in the pathoge-nicity of P. falciparum, the causative agent of the deadliestform of human malaria. Parasite virulence, as well as itsability to maintain long-term chronic infections, is attrib-uted to tight regulation of several clonally variant genefamilies. There is mounting evidence that dynamics innuclear architecture and positioning at the nuclear periph-ery are pivotal in transcriptional regulation of these genefamilies (Duraisingh et al., 2005; Ralph et al., 2005; Vosset al., 2006; Dzikowski et al., 2007; Howitt et al., 2009;Lopez-Rubio et al., 2009). It was suggested that a peri-nuclear expression site exists and that virulence genesmove into this focus upon activation (Ralph et al., 2005;Dzikowski et al., 2007, Dzikowski and Deitsch, 2009).However, very little is known about the structure andcomponents of the nuclear periphery of this organism,and moreover, the dynamics of nuclear architecture havenever been extensively studied in any eukaryotic organ-ism that undergoes schizogony. Given the extensivemodifications required to properly regulate and segregateup to ~ 32 copies of the genome in closed mitosis, nucleardynamics of this organism might prove to be particularlyinteresting and informative. Here we have applied severalultrastructural techniques to investigate dynamics innuclear architecture of P. falciparum throughout the intra-erythrocytic cycle. We show that dramatic changes inchromatin organization during parasite development areclosely coupled with changes in NPC number and positionat the NE. These observations are highlighted by anemerging EM technology that allowed us to create com-plete 3D models of nuclei in various stages. The dynamicsin nuclear architecture correlate with previously identifiedcyclic patterns of gene expression and genome organiza-tion in P. falciparum (Bozdech et al., 2003; Ponts et al.,2010).
Results
A first look at the organization of NPC in P. falciparum
As a first step towards understanding nuclear structureand dynamics in malaria-causing parasites, we were inter-ested to identify and characterize the organization of P.falciparum nuclear pores during the intra-erythrocyticdevelopment cycle (IDC). To our knowledge, structuralanalysis of the NPCs of blood stage parasites has notbeen extensively studied or described previously. Trans-mission electron microscopy (TEM) of early schizonts pre-pared by conventional chemical fixation revealed the
classic NPC structure as an electron-dense disk perforat-ing the double membrane of the NE (Fig. 1A and B), yetwe were interested to have a broader view of their orga-nization at the NE than could be afforded by isolated thinsections. Infected red blood cell (iRBC) samples werevitrified by high pressure freezing and subjected to freezefracture, then imaged by cryogenic scanning electronmicroscopy (cryoSEM). This method provides a surfaceview of the fracture plane, which in numerous instancescoincided with the parasite NE. In some fractures we wereable to view the cytoplasmic face of the nuclear pores(Fig. 1C and D), while in others the chromatin wasremoved, revealing the nucleoplasmic face and thedouble membrane in section (Fig. 1E and F). The size ofthe pores is ~ 80 nm, which is slightly smaller than whathas been described in yeast (~ 96 nm) (Yang et al., 1998).Interestingly, the pores seemed not to be evenly distrib-uted around the NE.
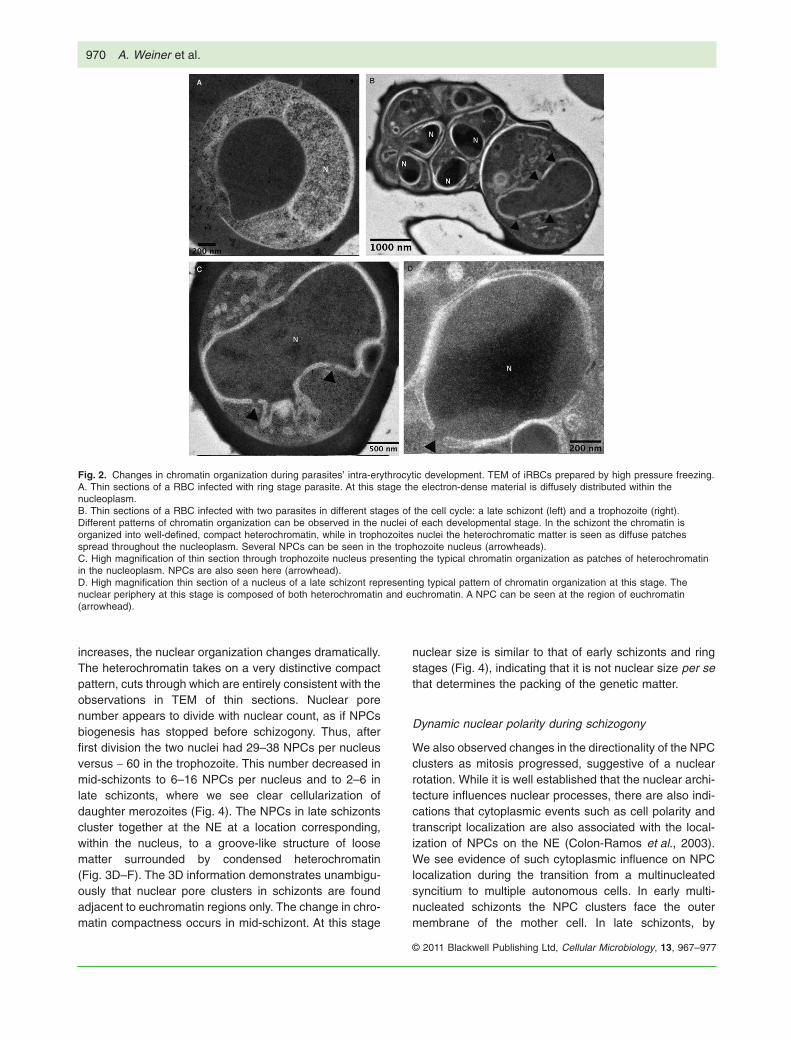
Dramatic changes in chromatin organization duringintra-erythrocytic development
In budding yeast the nuclear pores are distributed aroundthe NE and create islands of loose euchromatin that mayallow transcription within the condensed heterochromaticregion of the nuclear periphery (Capelson and Hetzer,2009). Therefore, we were interested to determine howthe uneven distribution of nuclear pores observed byfreeze fracture may be correlated to the chromatin orga-nization within the nucleus, specifically the distribution ofheterochromatin and euchromatin. Heterochromatin isrecognized in TEM by electron density of the heavy metalstains that associate to it preferentially. We first preparedthin sections of parasites by high pressure freezing andfreeze substitution. This method provides superior ultra-structural preservation of chromatin as compared withconventional chemical fixation, due to cryogenic immobi-lization and gentle dehydration of the sample. As seen inFig. 2, there are distinct differences in chromatin organi-zation between parasites at different stages of the IDC. Inrings (Fig. 2A) electron-dense (dark) genetic matter wasscattered throughout the nucleoplasm and there was noclear distinction between regions of heterochromatin andeuchromatin. In the trophozoite stage (Fig. 2B and C), theorganization of chromatin shows patches of dark hetero-chromatin scattered throughout the nucleoplasm. In sharpcontrast, in late schizonts one can clearly distinguishbetween the densely stained heterochromatic regions andthe lighter euchromatic regions in the nuclei. In one tellingexample, a single RBC was infected by two parasites atdifferent stages, a trophozoite and a late schizont side byside (Fig. 2B). We noted that while numerous nuclearpores could be seen in trophozoites, very few could bedetected in thin sections through schizonts and none were
968 A. Weiner et al.
© 2011 Blackwell Publishing Ltd, Cellular Microbiology, 13, 967–977

seen in ring stage parasites. In both trophozoites andschizonts we noted that the nuclear pores in these thinsections are always located nearby regions of looseeuchromatin and not adjacent to the electron-dense het-erochromatin.
3D analysis of P. falciparum nucleus reveals coupleddynamics of chromatin and NPC organization
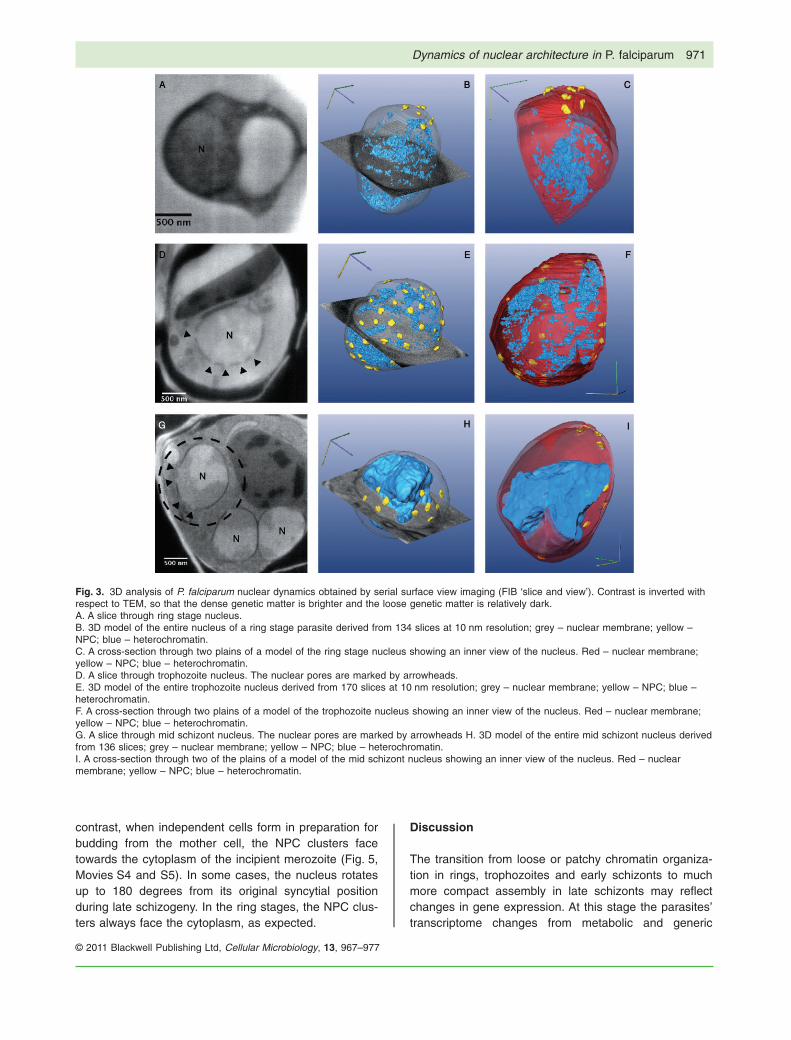
A full verification of this suggestive association is notpossible using isolated thin sections, but requires a full 3Dview of multiple nuclei at each developmental stage. Tothat end, we applied the emerging technique of serialsurface view imaging by focused ion beam (FIB) millingwithin a scanning electron microscope (SEM) (Bennettet al., 2009). Known as ‘slice and view’, this method pro-vides automated acquisition of serial surface images withspacing as thin as 10 nm. Thus a single nucleus isspanned typically by ~ 150 sections. A 3D model is gen-erated directly, without the need for tomographic recon-struction and without the limitations imposed by sample tiltgeometry. We applied this method to investigate nucleardynamics in rings, trophozoites and through schizogonyfrom early to late schizont stage parasites (Fig. 3;Movies S1–5; Data sets S1–3). We were able to create
and analyse 3D models of 4 nuclei from rings, 4 fromtrophozoites and 24 of nuclei at different stages ofschizogony (Fig. 4; Table S1). Analysis of these 3Dmodels of whole nuclei (in fact, whole parasites) showsdistinctly different patterns of sub-nuclear organizationamong the different stages. Nuclei of ring stage parasiteshave very few nuclear pores (3–7 pores per nucleus)clustered closely together. This can explain why nuclearpores were not detected in isolated thin sections; arandom cut is not likely to cross the cluster. Similar to whatwas observed by thin sections, the 3D models of ringsindicate that chromatin organization seems diffuse withinthe nucleoplasm with no significant differentiationbetween heterochromatic and euchromatic sub-nuclearcompartments. In trophozoites the number of pores pernucleus increases (12–58 pores per nucleus), corre-sponding with biogenesis of new pores as the parasitesdevelops within the RBC. At this stage the pores seemuniformly distributed around the NE, similar to what isobserved in other eukaryotic organisms. In addition, theheterochromatic regions are distributed as small patchesthroughout the nucleoplasm. At early schizogony, whenthe parasite has only two nuclei, the nuclear architectureis similar to that of trophozoites (Fig. S1). However, asschizogony progresses and the number of nuclei
Fig. 1. The nuclear pores of P. falciparum.N (nucleus); RBC (red blood cell); FV (foodvacuole).A and B. Cross-section through a red bloodcell infected with an early schizont obtainedby TEM of chemically fixed iRBC. The framedregion in (A) is presented at highermagnification in (B) and shows a nuclear porewith its inner ring seen as electron-densematter between the double membranes of thenuclear envelope.C and D. Freeze fracture cryoSEM exposingthe outer surface of an early schizont nucleuswith a cluster of NPCs (arrowhead).E and F. Freeze fracture cryoSEM through anearly schizont exposing the inner part of thenuclear membrane. The framed region in (E)is presented at higher magnification in F andshows two NPCs (arrowed). Two more NPCswere exposed in a longitude break where thesurrounding nuclear envelope can beobserved (arrowhead).
Dynamics of nuclear architecture in P. falciparum 969
© 2011 Blackwell Publishing Ltd, Cellular Microbiology, 13, 967–977
increases, the nuclear organization changes dramatically.The heterochromatin takes on a very distinctive compactpattern, cuts through which are entirely consistent with theobservations in TEM of thin sections. Nuclear porenumber appears to divide with nuclear count, as if NPCsbiogenesis has stopped before schizogony. Thus, afterfirst division the two nuclei had 29–38 NPCs per nucleusversus ~ 60 in the trophozoite. This number decreased inmid-schizonts to 6–16 NPCs per nucleus and to 2–6 inlate schizonts, where we see clear cellularization ofdaughter merozoites (Fig. 4). The NPCs in late schizontscluster together at the NE at a location corresponding,within the nucleus, to a groove-like structure of loosematter surrounded by condensed heterochromatin(Fig. 3D–F). The 3D information demonstrates unambigu-ously that nuclear pore clusters in schizonts are foundadjacent to euchromatin regions only. The change in chro-matin compactness occurs in mid-schizont. At this stage
nuclear size is similar to that of early schizonts and ringstages (Fig. 4), indicating that it is not nuclear size per sethat determines the packing of the genetic matter.
Dynamic nuclear polarity during schizogony
We also observed changes in the directionality of the NPCclusters as mitosis progressed, suggestive of a nuclearrotation. While it is well established that the nuclear archi-tecture influences nuclear processes, there are also indi-cations that cytoplasmic events such as cell polarity andtranscript localization are also associated with the local-ization of NPCs on the NE (Colon-Ramos et al., 2003).We see evidence of such cytoplasmic influence on NPClocalization during the transition from a multinucleatedsyncitium to multiple autonomous cells. In early multi-nucleated schizonts the NPC clusters face the outermembrane of the mother cell. In late schizonts, by
Fig. 2. Changes in chromatin organization during parasites’ intra-erythrocytic development. TEM of iRBCs prepared by high pressure freezing.A. Thin sections of a RBC infected with ring stage parasite. At this stage the electron-dense material is diffusely distributed within thenucleoplasm.B. Thin sections of a RBC infected with two parasites in different stages of the cell cycle: a late schizont (left) and a trophozoite (right).Different patterns of chromatin organization can be observed in the nuclei of each developmental stage. In the schizont the chromatin isorganized into well-defined, compact heterochromatin, while in trophozoites nuclei the heterochromatic matter is seen as diffuse patchesspread throughout the nucleoplasm. Several NPCs can be seen in the trophozoite nucleus (arrowheads).C. High magnification of thin section through trophozoite nucleus presenting the typical chromatin organization as patches of heterochromatinin the nucleoplasm. NPCs are also seen here (arrowhead).D. High magnification thin section of a nucleus of a late schizont representing typical pattern of chromatin organization at this stage. Thenuclear periphery at this stage is composed of both heterochromatin and euchromatin. A NPC can be seen at the region of euchromatin(arrowhead).
970 A. Weiner et al.
© 2011 Blackwell Publishing Ltd, Cellular Microbiology, 13, 967–977
contrast, when independent cells form in preparation forbudding from the mother cell, the NPC clusters facetowards the cytoplasm of the incipient merozoite (Fig. 5,Movies S4 and S5). In some cases, the nucleus rotatesup to 180 degrees from its original syncytial positionduring late schizogeny. In the ring stages, the NPC clus-ters always face the cytoplasm, as expected.
Discussion
The transition from loose or patchy chromatin organiza-tion in rings, trophozoites and early schizonts to muchmore compact assembly in late schizonts may reflectchanges in gene expression. At this stage the parasites’transcriptome changes from metabolic and generic
Fig. 3. 3D analysis of P. falciparum nuclear dynamics obtained by serial surface view imaging (FIB ‘slice and view’). Contrast is inverted withrespect to TEM, so that the dense genetic matter is brighter and the loose genetic matter is relatively dark.A. A slice through ring stage nucleus.B. 3D model of the entire nucleus of a ring stage parasite derived from 134 slices at 10 nm resolution; grey – nuclear membrane; yellow –NPC; blue – heterochromatin.C. A cross-section through two plains of a model of the ring stage nucleus showing an inner view of the nucleus. Red – nuclear membrane;yellow – NPC; blue – heterochromatin.D. A slice through trophozoite nucleus. The nuclear pores are marked by arrowheads.E. 3D model of the entire trophozoite nucleus derived from 170 slices at 10 nm resolution; grey – nuclear membrane; yellow – NPC; blue –heterochromatin.F. A cross-section through two plains of a model of the trophozoite nucleus showing an inner view of the nucleus. Red – nuclear membrane;yellow – NPC; blue – heterochromatin.G. A slice through mid schizont nucleus. The nuclear pores are marked by arrowheads H. 3D model of the entire mid schizont nucleus derivedfrom 136 slices; grey – nuclear membrane; yellow – NPC; blue – heterochromatin.I. A cross-section through two of the plains of a model of the mid schizont nucleus showing an inner view of the nucleus. Red – nuclearmembrane; yellow – NPC; blue – heterochromatin.
Dynamics of nuclear architecture in P. falciparum 971
© 2011 Blackwell Publishing Ltd, Cellular Microbiology, 13, 967–977

cellular function into specialized functions needed foregress and re-invasion (Bozdech et al., 2003). This hasbeen correlated recently with an overall decrease in thetotal transcriptional activity per nucleus (Sims et al.,2009), as well as changes in the nucleosome landscapeat that stage (Ponts et al., 2010). Epigenome analysis ofP. falciparum revealed unique plasticity in the use ofhistone modifications during the IDC. The vast majority ofthe genome is enriched in H3K4me3 and H3K9ac duringearly stages, while in schizonts these euchromatinmarkers are enriched at the 5′ of active genes (Salcedo-Amaya et al., 2009). These data coincide precisely withthe ultrastructural chromatin dynamics described here,where until late schizont stages the nucleoplasm containsmainly loose genetic matter with scattered patches ofcondensed heterochromatin.
Early ultrastructural studies of Plasmodium nuclei usingconventional EM thin sections pointed to differences inthe electron density of stained regions within the nucleus(Aikawa, 1966; Aikawa et al., 1966a,b; 1967; Ladda et al.,1969, Aikawa and Ward, 1974, Bannister and Sinden,1982). Electron-dense (dark) regions were associatedwith heterochromatin while electron sparse (light) regionswere associated loose genetic matter. This sub-nuclearcompartmentalization was recently supported with thefinding that chromatin modifying enzymes such as PfSIR2and PfHP1 are located in these heterochromatic regions(Duraisingh et al., 2005; Freitas-Junior et al., 2005;Flueck et al., 2009). The rich prior literature of EM studiesshows great diversity in the patterns by which theseregions are organized within the nucleoplasm, as could beobserved in thin sections. This variety could representdifferences between Plasmodium species, the stagesduring development, or technical differences as a conse-quence of chemical fixation and staining. In classicalTEM, heterochromatin is recognized by strong absorptionof the illumination by electron-dense, heavy metal stains,
creating dark patches. These usually corresponds to chro-matin density detected optically by fluorescence staining(Schermelleh et al., 2008); however, less dense hetero-chromain might not be seen by this method but only withspecific molecular markers for histone modifications.Some of the previous EM images (Freitas-Junior et al.,2005; Ralph et al., 2005) showed that in nuclei of lateschizonts there are large regions of electron-dense mate-rial at the inner perimeter of the NE. In some of the nucleithe heterochromatin is distributed all around the periph-ery, while other nuclei in the same schizont have differentpatterns (Freitas-Junior et al., 2005). It is possible that thedifferences between these patterns and those describedhere are simply the result of different fixation protocols.However, in light of our 3D analysis, one may also expectto observe such different patterns in nuclei of the sameschizont due to the various positions of nuclei being ran-domly sampled and displayed in a single isolated thinsection.
In addition to these chromatin dynamics we observedchanges in the number and the distribution of NPCs asparasite development progresses. The parasite beginsthe IDC with a few clustered NPCs at the ring stage.Biogenesis of NPCs occurs as it develops into trophozoiteand the NPCs are spread all around the NE, reaching acount of ~ 60. The number of NPCs per nucleusdecreases gradually during schizogony as the number ofnuclei increases, until in nuclei of late schizonts there areonly few NPCs on each nucleus. The few NPCs remainingin late schizonts cluster again at distinct foci on the NE.The number of NPCs per NE surface can vary amongspecies, and with changes in nuclear size and activity(Lim et al., 2008). In certain plants and eukaryotesdynamics in NPC organization was associated with devel-opmental changes of the cell (Maeshima et al., 2006;Fiserova et al., 2009). For example, in mammalianspermatids, which undergo significant morphological
Fig. 4. Graphical summary of the number ofNPCs per nucleus and nuclear volume ofparasites at different stages during IDC. Eachcolumn represents the number of NPCscounted in each of the 3D models that werecreated for a single nucleus (left axis). Thevolume of each nucleus is marked with blackdots (right axis). The numbers given for NPCsat schizont stage represent the nuclei inwhich we were able to create and analyse 3Dmodels. Altogether we analysed 2/2 nuclei ofone early schizont; 4/6, 7/8, 4/9 nuclei of 3mid schizont, and 8/14 of one late schizont.
972 A. Weiner et al.
© 2011 Blackwell Publishing Ltd, Cellular Microbiology, 13, 967–977

modifications during spermatogenesis, the NPC organiza-tion changes from even distribution on the NE at earlyphase into clustering in high densities at the posterior sideof the nucleus in mature spermatids (Ho, 2010). Similarly,dynamics in the number and distribution of NPCs duringthe cell cycle were reported also for the budding yeastSaccharomyces cervisiae (Winey et al., 1997, Bucci andWente, 1997) and the unicellular alga Chlamydomonasreinhardtii (Colon-Ramos et al., 2003). Very little wasknown about dynamics of NPCs organization in Plasmo-dium spp., although early freeze fracture of trophozoitesof the avian malaria parasite P. gallinaceum suggestedlong ago that NPCs are not equally spread on the surfaceof the NE (Seed et al., 1971). However, the clusteringobserved in P. falciparum is associated with a significant
decrease in the number of NPCs. This may indicate thatbiogenesis of nuclear pores is halted at the trophozoitestage, so that a fixed number of pores are then distributedamong the daughter nuclei. NPCs also disappear frommost of the NE, explaining the difficulty to find them byTEM in thin sections of late or early stage parasites. Theclustered distribution of NPCs seems to maintain specificcellular directionality. In early schizogony they faceoutward, whereas upon completion of cytokinesis theyface the cytoplasm of the daughter cells. The observednuclear rotation is consistent with efficient transport ofmRNA transcripts into polysome-rich cytoplasmic regions.Apicomplexan parasites maintain their NE during mitosisand use an intranuclear spindle to interact with the cyto-plasmic centrosome through an opening in the NE
Fig. 5. 3D analysis of P. falciaprum nuclei in mid and late schizonts by serial surface view imaging shows reorientation of nuclear pores (FIB‘slice and view’). Contrast is inverted with respect to TEM, so that the dense genetic matter is brighter and the loose genetic matter isrelatively dark.A. A slice through a parasite in mid schizont stage. NPCs are also seen here (arrowheads).B. 3D model of the entire parasite in mid schizont stage. Red – nuclear membrane; yellow – NPC; blue – heterochromatin; purple – foodvacuole; orange – haemozoin; green – RBC membrane. NPCs are oriented outward, facing the parasite membrane.C. A slice through a parasite in late schizont stage. NPC is also seen here (arrowhead).D. 3D model of the entire parasite in late schizont stage; red – nuclear membrane; yellow – NPC; blue – heterochromatin; green – merozoitemembrane. NPCs are oriented to face the cytoplasm of each incipient merozoite.
Dynamics of nuclear architecture in P. falciparum 973
© 2011 Blackwell Publishing Ltd, Cellular Microbiology, 13, 967–977

(Schrevel et al., 1977; Striepen et al., 2007). Dynamics inthe polarity of the centrosome during mitosis is verysimilar to what we observed here in the clusters of NPCs.Although mitotic spindles could not been observed by FIBone can speculate that these changes are directed bynuclear microtubule centres that traverse the NE throughthese clusters.
Considering the dramatic changes in chromatin organi-zation during parasite development, it is interesting thatthe NPCs are found in regions of loose euchromatin.Recent advances in our understanding of how the nucleararchitecture is related to the control of gene transcriptionat the nuclear periphery suggested that, in addition to theirrole in nuclear transport, the NPC can serve as chromatininsulators, which create the platforms that enable activetranscription (Akhtar and Gasser, 2007, Kohler and Hurt,2010). Nuclear dynamics and compartmentalization playkey roles in virulence gene expression in P. falciparum,possibly by movement of active genes into a sub-nuclearexpression site at the nuclear periphery where the tran-scriptional machinery that is required for their activation islocated. Many years ago the ‘gene gating’ hypothesisenvisioned that a non-random distribution of NPCs at thenuclear periphery may suggest that they can serve as agating organelle capable of interacting specifically withtranscribable portions in the genome (Blobel, 1985). Inview of our current 3D data one can hypothesize that thepostulated expression site at the nuclear periphery of P.falciparum may be related to the positions of NPCs orNPC clusters.
Experimental procedure
Parasite culture
All parasites used were derivatives of the NF54 parasite line,cultivated at 5% haematocrit in RPMI 1640 medium, 0.5%Albumax II (Invitrogen), 0.25% sodium bicarbonate and0.1 mg ml-1 gentamicin. Parasites were incubated at 37°C in anatmosphere of 5% oxygen, 5% carbon dioxide and 90% nitrogen.Parasite cultures were synchronized using percoll/sorbitol gradi-ent centrifugation as previously described (Aley et al., 1984;Calderwood et al., 2003). Briefly, infected RBCs were layered ona step gradient of 40%/70% percoll containing 6% sorbitol. Thegradients were then centrifuged at 12 000 g for 20 min at roomtemperature. Highly synchronized, late stage parasites wererecovered from the 40%/70% interphase, washed twice withcomplete culture media and collected for the different fixation forEM. After synchronization RBCs infected with ring stage parasitewere isolated by lysing the un-infected RBCs with streptolysin O(Sigma No. S5265) at a concentration of 5 u ml-1 of packed RBCsas described (Jackson et al., 2007).
Chemical fixation and embedding
Parasites were pelleted and fixed with 2% paraformaldehyde and2.5% glutaraldehyde in 0.1 M Cacodylate buffer (pH 7.4) at room
temperature for 2–3 h, washed and post-fixed with 1% osmiumtetroxide in the same buffer for 1 h at room temperature. After enbloc staining with 2% uranyl acetate in water for 2 h at roomtemperature, the samples were dehydrated in graded ethanolseries and embedded in Epon 812.
Cryo-fixation and embedding
Parasites were pelleted and cryo-fixed by high pressure freezing(HPF) using a HPM 010 high-pressure freezer (Bal-Tec).Samples were freeze-substituted (Leica EM AFS) in dry acetonecontaining 2% glutaraldehyde and 0.1% tannic with a gradualtemperature increase of -90°C to -30°C over 90 h. Sampleswere rinsed in acetone and incubated with 0.1% uranyl acetate(UA) and 1% OsO4 for 1 h on ice, then rinsed again, infiltratedgradually with increasing concentrations of Resin (Epon by AgarScientific) over 6 days, and polymerized at 60°C.
TEM thin sections
Thin sections (60–90 nm) were prepared using an Ultracut UCTmicrotome (Leica). Following post staining with 1%–2% uranylacetate and Reynold’s lead citrate, sections were viewed in aTecnai Spirit BioTWIN (FEI) operating at 120 kV. Imagesrecorded with an Eagle 2K*2K CCD camera (FEI).
Freeze fracture cryoSEM
Freeze fracture cryoSEM was performed according to a previ-ously described protocol (Wild et al., 2009) with some modifica-tions. Briefly, parasites were centrifuged and a drop of pellet wasplaced on a 200 mesh EM grid, which was then sandwichedbetween the flat surfaces of two type B aluminum platelets forhigh pressure freezing (Wohlwend Engineering Office). Thesample was then high-pressure frozen in a HPM 010 high-pressure freezer (Bal-Tec). High pressure freezing leads to vitri-fication of the aqueous medium, and so to physical fixation of theconstituent structures. The frozen samples were mounted on aholder and transferred to a BAF 60 freeze fracture device (Bal-Tec) using a VCT 100 Vacuum Cryo Transfer device (Bal-Tec).After fracturing at a temperature of -120°C, samples were imme-diately coated with 4 nm Pt/C by double axis rotary shadowing.Samples were then transferred to an Ultra 55 SEM (Zeiss) usingthe VCT 100 transfer device and observed using the secondaryelectrons in-lens detector at 2.5 kV at a temperature of -120°C.
Serial surface view imaging by dual head focused ionbeam/scanning electron microscopy (‘slice and view’)
Resin blocks were handled according to a previously describedprotocol (Bennett et al., 2009) with some modifications. Samplesembedded in resin were first exposed at one edge of a pyramidalshape using a razor blade. The top surface was smoothed usinga 45° diamond knife (Diatome). The pyramid was sawed off andmounted on a SEM stub using UV glue with the top surfacepointing upwards. The stub was then sputtered with gold-palladium (Edwards) and placed in a Helios Nanolab Dual beam(FEI). The stage was tilted to 52° such that the top surface isorthogonal to the ion beam. An aggressive cleaning cross-section
974 A. Weiner et al.
© 2011 Blackwell Publishing Ltd, Cellular Microbiology, 13, 967–977

(21 nA at 30 kV) was used to expose the tissue at the edge of thepyramid. Once a suitable cell was exposed, a protective layer ofplatinum (1 mm thickness) was deposited above it on the topsurface. Two trenches were milled on the sides of the sample anda gentle ion beam current (0.92 nA) was used to polish thesurface. The Auto Slice and View G2 (FEI) program was used toproduce serial views of freshly exposed surfaces. Combined SEand BSE SEM images were recorded with 1.4 nA at 2 kV in theimmersion lens mode, with pixel sizes ranging between 1.9 nm to5 nm depending on the sample. For milling slices an ion beamcurrent of 0.92 nA at 30 kV was used with a step size of 10 nm.Images were aligned in ImageJ (Rasband, W.S., ImageJ, USNational Institutes of Health, Bethesda, Maryland, USA, http://rsb.info.nih.gov/ij/, 1997–2009) using StackReg plugin (PhilippeThevenaz, Biomedical Imaging Group, Swiss Federal institute oftechnology, Lausanne, Switzerland; http://bigwww.epfl.ch/thevenaz/stackreg/). Amira (Visage Imaging) was used for 3Dvisualization, rendering, and manual segmentation using themagic wand tool. Note the inversion of contrast with respect toTEM. Whereas heavy metal stained regions appear dark in TEMdue to absorption of the incident electrons, in SEM the stainedregions emit secondary electrons and so appear bright.
Acknowledgements
This work was supported by a grant from the United States-IsraelBinational Science Foundation [2007350]. R.D. is a Gaia LuoniScholar and supported by the Marie Curie International Reinte-gration Grant (IRG) [203675], the German Israel Foundation[997/2008] and the Israel Science Foundation [660/09]. R.D. isalso supported by the Jacob and Lena Joels Memorial Founda-tion Senior Lectureship for Excellence in the Life and MedicalSciences. Contributions of A.W. and M.E. were supported in partby the Curwen-Lowy Fellowship of the Weizmann Institute ofScience, by the Gerhardt M.J. Schmidt Minerva Center forSupramolecular Architecture, and by the historical generosity ofthe Harold Perlman family. Electron microscopy was performed inthe Irving and Cherna Moscowitz Center for Nano and Bio-nanoImaging of the Weizmann Institute.
References
Aikawa, M. (1966) The fine structure of the erythrocyticstages of three avian malarial parasites, Plasmodiumfallax, P. lophurae, and P. cathemerium. Am J Trop MedHyg 15: 449–471.
Aikawa, M., and Ward, R.A. (1974) Intraspecific variation inPlasmodium falciparum. Am J Trop Med Hyg 23: 570–573.
Aikawa, M., Hepler, P.K., Huff, C.G., and Sprinz, H. (1966a)The feeding mechanism of avian malarial parasites. J CellBiol 28: 355–373.
Aikawa, M., Huff, C.G., and Spinz, H. (1966b) Comparativefeeding mechanisms of avian and primate malarial para-sites. Mil Med 131 (Suppl.): 969–983.
Aikawa, M., Huff, C.G., and Sprinz, H. (1967) Fine structureof the asexual stages of Plasmodium elongatum. J Cell Biol34: 229–249.
Akhtar, A., and Gasser, S.M. (2007) The nuclear envelopeand transcriptional control. Nat Rev Genet 8: 507–517.
Aley, S.B., Sherwood, J.A., and Howard, R.J. (1984) Knob-positive and knob-negative Plasmodium falciparum differ in
expression of a strain-specific malarial antigen on thesurface of infected erythrocytes. J Exp Med 160: 1585–1590.
Babu, M.M., Janga, S.C., de Santiago, I., and Pombo, A.(2008) Eukaryotic gene regulation in three dimensions andits impact on genome evolution. Curr Opin Genet Dev 18:571–582.
Bannister, L.H., and Sinden, R.E. (1982) New knowledge ofparasite morphology. Br Med Bull 38: 141–145.
Bennett, A.E., Narayan, K., Shi, D., Hartnell, L.M., Gousset,K., He, H., et al. (2009) Ion-abrasion scanning electronmicroscopy reveals surface-connected tubular conduits inHIV-infected macrophages. PLoS Pathog 5: e1000591.
Blobel, G. (1985) Gene gating: a hypothesis. Proc Natl AcadSci USA 82: 8527–8529.
Bozdech, Z., Llinas, M., Pulliam, B.L., Wong, E.D., Zhu, J.C.,and Derisi, J.L. (2003) The transcriptome of the intraeryth-rocytic developmental cycle of Plasmodium falciparum.PLoS Biology 1: 85–100.
Brown, C.R., and Silver, P.A. (2007) Transcriptional regula-tion at the nuclear pore complex. Curr Opin Genet Dev 17:100–106.
Bucci, M., and Wente, S.R. (1997) In vivo dynamics of nuclearpore complexes in yeast. J Cell Biol 136: 1185–1199.
Calderwood, M.S., Gannoun-Zaki, L., Wellems, T.E., andDeitsch, K.W. (2003) Plasmodium falciparum var genesare regulated by two regions with separate promoters, oneupstream of the coding region and a second within theintron. J Biol Chem 278: 34125–34132.
Capelson, M., and Hetzer, M.W. (2009) The role of nuclearpores in gene regulation, development and disease. EMBORep 10: 697–705.
Chakalova, L., Debrand, E., Mitchell, J.A., Osborne, C.S.,and Fraser, P. (2005) Replication and transcription:shaping the landscape of the genome. Nat Rev Genet 6:669–677.
Colon-Ramos, D.A., Salisbury, J.L., Sanders, M.A., Shenoy,S.M., Singer, R.H., and Garcia-Blanco, M.A. (2003)Asymmetric distribution of nuclear pore complexes and thecytoplasmic localization of beta2-tubulin mRNA in Chlamy-domonas reinhardtii. Dev Cell 4: 941–952.
Duraisingh, M.T., Voss, T.S., Marty, A.J., Duffy, M.F., Good,R.T., Thompson, J.K., et al. (2005) Heterochromatin silenc-ing and locus repositioning linked to regulation of virulencegenes in Plasmodium faiciparum. Cell 121: 13–24.
Dzikowski, R., and Deitsch, K.W. (2009) Genetics of anti-genic variation in Plasmodium falciparum. Curr Genet 55:103–110.
Dzikowski, R., Li, F., Amulic, B., Eisberg, A., Frank, M., Patel,S., et al. (2007) Mechanisms underlying mutually exclusiveexpression of virulence genes by malaria parasites. EMBORep 8: 959–965.
Fiserova, J., Kiseleva, E., and Goldberg, M.W. (2009)Nuclear envelope and nuclear pore complex structure andorganization in tobacco BY-2 cells. Plant J 59: 243–255.
Flueck, C., Bartfai, R., Volz, J., Niederwieser, I., Salcedo-Amaya, A.M., Alako, B.T., et al. (2009) Plasmodium falci-parum heterochromatin protein 1 marks genomic locilinked to phenotypic variation of exported virulence factors.PLoS Pathog 5: e1000569.
Freitas-Junior, L.H., Hernandez-Rivas, R., Ralph, S.A.,
Dynamics of nuclear architecture in P. falciparum 975
© 2011 Blackwell Publishing Ltd, Cellular Microbiology, 13, 967–977

Montiel-Condado, D., Ruvalcaba-Salazar, O.K., Rojas-Meza, A.P., et al. (2005) Telomeric heterochromatin propa-gation and histone acetylation control mutually exclusiveexpression of antigenic variation genes in malaria para-sites. Cell 121: 25–36.
Goldman, R.D., Gruenbaum, Y., Moir, R.D., Shumaker, D.K.,and Spann, T.P. (2002) Nuclear lamins: building blocks ofnuclear architecture. Genes Dev 16: 533–547.
Gruenbaum, Y., Goldman, R.D., Meyuhas, R., Mills, E., Mar-galit, A., Fridkin, A., et al. (2003) The nuclear lamina and itsfunctions in the nucleus. Int Rev Cytol 226: 1–62.
Hakim, O., Sung, M.H., and Hager, G.L. (2010) 3D shortcutsto gene regulation. Curr Opin Cell Biol 22: 305–313.
Ho, H.C. (2010) Redistribution of nuclear pores during forma-tion of the redundant nuclear envelope in mouse sperma-tids. J Anat 216: 525–532.
Howitt, C.A., Wilinski, D., Llinas, M., Templeton, T.J.,Dzikowski, R., and Deitsch, K.W. (2009) Clonally variantgene families in Plasmodium falciparum share a commonactivation factor. Mol Microbiol 73: 1171–1185.
Jackson, K.E., Spielmann, T., Hanssen, E., Adisa, A.,Separovic, F., Dixon, M.W., et al. (2007) Selective per-meabilization of the host cell membrane of Plasmodiumfalciparum-infected red blood cells with streptolysin O andequinatoxin II. Biochem J 403: 167–175.
Kohler, A., and Hurt, E. (2010) Gene regulation by nucleopor-ins and links to cancer. Mol Cell 38: 6–15.
Ladda, R., Aikawa, M., and Sprinz, H. (1969) Penetration oferythrocytes by merozoites of mammalian and avianmalarial parasites. J Parasitol 55: 633–644.
Lim, R.Y., Ullman, K.S., and Fahrenkrog, B. (2008) Biologyand biophysics of the nuclear pore complex and its com-ponents. Int Rev Cell Mol Biol 267: 299–342.
Lopez-Rubio, J.-J., Mancio-Silva, L., and Scherf, A. (2009)Genome-wide analysis of heterochromatin associatesclonally variant gene regulation with perinuclear repressivecenters in malaria parasites. Cell Host Microbe 5: 179–190.
Maeshima, K., Yahata, K., Sasaki, Y., Nakatomi, R.,Tachibana, T., Hashikawa, T., et al. (2006) Cell-cycle-dependent dynamics of nuclear pores: pore-free islandsand lamins. J Cell Sci 119: 4442–4451.
Ponts, N., Harris, E.Y., Prudhomme, J., Wick, I., Eckhardt-Ludka, C., Hicks, G.R., et al. (2010) Nucleosome land-scape and control of transcription in the human malariaparasite. Genome Res 20: 228–238.
Ralph, S.A., Scheidig-Benatar, C., and Scherf, A. (2005)Antigenic variation in Plasmodium falciparum is associatedwith movement of var loci between subnuclear locations.Proc Natl Acad Sci USA 102: 5414–5419.
Salcedo-Amaya, A.M., van Driel, M.A., Alako, B.T., Trelle,M.B., van den Elzen, A.M., Cohen, A.M., et al. (2009)Dynamic histone H3 epigenome marking during theintraerythrocytic cycle of Plasmodium falciparum. Proc NatlAcad Sci USA 106: 9655–9660.
Schermelleh, L., Carlton, P.M., Haase, S., Shao, L., Winoto,L., Kner, P., et al. (2008) Subdiffraction multicolor imagingof the nuclear periphery with 3D structured illuminationmicroscopy. Science 320: 1332–1336.
Schrevel, J., Asfaux-Foucher, G., and Bafort, J.M. (1977)[Ultrastructural study of multiple mitoses during sporogony
of Plasmodium b. berghei]. J Ultrastruct Res 59: 332–350.
Seed, T., Pfister, R., Kreier, J., and Johnson, A. (1971) Plas-modium gallinaceum: fine structure by freeze-etch tech-nique. Exp Parasitol 30: 73–81.
Sexton, T., Schober, H., Fraser, P., and Gasser, S.M. (2007)Gene regulation through nuclear organization. Nat StructMol Biol 14: 1049–1055.
Sims, J.S., Militello, K.T., Sims, P.A., Patel, V.P., Kasper, J.M.,and Wirth, D.F. (2009) Patterns of gene-specific and totaltranscriptional activity during the Plasmodium falciparumintraerythrocytic developmental cycle. Eukaryot Cell 8:327–338.
Solovei, I., Kreysing, M., Lanctot, C., Kosem, S., Peichl, L.,Cremer, T., et al. (2009) Nuclear architecture of rod photo-receptor cells adapts to vision in mammalian evolution. Cell137: 356–368.
Striepen, B., Jordan, C.N., Reiff, S., and van Dooren, G.G.(2007) Building the perfect parasite: cell division in apicom-plexa. PLoS Pathog 3: e78.
Voss, T.S., Healer, J., Marty, A.J., Duffy, M.F., Thompson,J.K., Beeson, J.G., et al. (2006) A var gene promoter con-trols allelic exclusion of virulence genes in Plasmodiumfalciparum malaria. Nature 439: 1004–1008.
Wild, P., Senn, C., Manera, C.L., Sutter, E., Schraner, E.M.,Tobler, K., et al. (2009) Exploring the nuclear envelope ofherpes simplex virus 1-infected cells by high-resolutionmicroscopy. J Virol 83: 408–419.
Winey, M., Yarar, D., Giddings, T.H., Jr, and Mastronarde,D.N. (1997) Nuclear pore complex number and distributionthroughout the Saccharomyces cerevisiae cell cycle bythree-dimensional reconstruction from electron micro-graphs of nuclear envelopes. Mol Biol Cell 8: 2119–2132.
Yang, Q., Rout, M.P., and Akey, C.W. (1998) Three-dimensional architecture of the isolated yeast nuclear porecomplex: functional and evolutionary implications. Mol Cell1: 223–234.
Supporting information
Additional Supporting Information may be found in the onlineversion of this article:
Fig. S1. A slice through an early schizont that was obtained byserial surface view imaging (‘slice & view’). At this stage theparasite has only two nuclei. The dense genetic matter is morereflective and therefore looks brighter in this method than loosegenetic matter which is darker. The pattern of chromatin organi-zation at this stage is similar to that of the trophozoite.Table S1. Details of nuclear data obtained from the 3D modelsthat were generated for each nucleus.Movie S1. This movie demonstrates the generation of a 3Dmodel of a nucleus of a ring stage parasite that is presented inFig. 3. Data were collected from 10 nm FIB section through thenucleus, aligned and visualized with Amira software. Yellow –NPC; blue – heterochromatin (QuickTime).Movie S2. This movie demonstrates the generation of a 3Dmodel of a nucleus of a trophozoite that is presented in Fig. 3.The model was generated similar to Movie S1 (QuickTime).
976 A. Weiner et al.
© 2011 Blackwell Publishing Ltd, Cellular Microbiology, 13, 967–977

Movie S3. This movie demonstrates the generation of a 3Dmodel of a nucleus of a schizont that is presented in Fig. 3. Themodel was generated similar to Movie S1 (QuickTime).Movie S4. This movie demonstrates the generation of a 3Dmodel of an entire parasite at mid schizont stage that ispresented in Fig. 4. Data were collected from 10 nm FIB sectionthrough the infected red blood cell, aligned and visualized withAmira software. Red – nuclear membrane; yellow – NPC; blue –heterochromatin; purple – food vacuole; orange – haemozoin;green – RBC membrane (QuickTime).Movie 5. This movie demonstrates the generation of a 3D modelof an entire parasite at late schizont stage that is presented inFig. 4. The model was generated similar to Movie S3. Red –nuclear membrane; yellow – NPC; blue – heterochromatin;orange – haemozoin; green – merozoite membrane (QuickTime).Data set S1. This movie shows the data set of images collectedby serial surface view imaging (‘slice & view’) from RBC infectedwith ring stage parasite. These data were used to create the 3D
model of the ring nucleus presented in Fig. 3B and C and theMovie S1. Voxel size: x,y – 10 nm, z – 20 nm.Data set S2. This movie shows the data set of images collectedby serial surface view imaging (‘slice & view’) from RBC infectedwith trophozoite. These data were used to create the 3D model ofthe trophozoite nucleus presented in Fig. 3E and F and theMovie S2. Voxel size: x,y – 7.14 nm, z – 20 nm.Data set S3. This movie shows the data set of nucleus imagescollected by serial surface view imaging (‘slice & view’) from RBCinfected with schizont. These data were used to create the 3Dmodel of the schizont nucleus presented in Fig. 3H and I and theMovie S3. Voxel size: x,y – 3.84 nm, z – 16 nm.
Please note: Wiley-Blackwell are not responsible for the contentor functionality of any supporting materials supplied by theauthors. Any queries (other than missing material) should bedirected to the corresponding author for the article.
Dynamics of nuclear architecture in P. falciparum 977
© 2011 Blackwell Publishing Ltd, Cellular Microbiology, 13, 967–977




















