
The Bosporus Strait: Exchange Fluxes, Currents and Sea-Level Changes

REVIEW
From Membrane Pores to Aquaporins: 50 YearsMeasuring Water Fluxes
Mario Parisi & Ricardo A. Dorr & Marcelo Ozu &
Roxana Toriano
Received: 30 October 2007 /Accepted: 13 March 2008 /Published online: 9 May 2008# Springer Science + Business Media B.V. 2008
Abstract This review focuses on studies of water movement across biological membranesperformed over the last 50 years. Different scientific approaches had tried to elucidate suchintriguing mechanism, from hypotheses emphasizing the role of the lipid bilayer to thecloning of aquaporins, the ubiquitous proteins described as specific water channels.Pioneering and clarifying biophysical work are reviewed beside results obtained with thehelp of recent sophisticated techniques, to conclude that great advances in the subject livetogether with old questions without definitive answers.
Keywords Water channels . Aquaporins . Single file . Water diffusion .
Osmotic permeability . Water pathway
1 Introduction
The mechanism by which water passes through biological membranes has been a subject ofintense research over the last 50 years. Different hypotheses were proposed (sometimescomplementary, sometimes contradictory) to explain such crucial machinery inherent tolife. The discovery of the proteins known as aquaporins (AQPs) had ended to the debate onthe controversial existence of specific water channels. Still, at the present time, there are nodefinitive conclusions to explain important topics as, for example, the microscopicdevelopment of forces that govern the osmosis mechanism inside the channel.
This review will outline the pioneering ideas and discoveries about the mechanisms ofwater movements across cellular membranes.
It must be here stressed that the net water movements across biological membranes canbe observed under two fundamentally different experimental conditions: (1) associated with
J Biol Phys (2007) 33:331–343DOI 10.1007/s10867-008-9064-5
M. Parisi :M. OzuUnidad de Biomembranas, Universidad Favaloro, Buenos Aires, Argentina
R. A. Dorr : R. Toriano (*)Laboratorio de Biomembranas, Departamento de Fisiología, Facultad de Medicina,Universidad de Buenos Aires, Buenos Aires, Argentinae-mail: [email protected]

a net solute transfer, in the absence of at least a macroscopic osmotic gradient and (2) in thepresence of a trans-membrane osmotic gradient. The representative examples of these twosituations are the so-called isosmotic reabsorption in the proximal kidney tubule and thefacultative reabsorption in the kidney collecting duct. In the next paragraphs, we will beconsidering case 2, except where specified.
2 The “Water Pore” Hypothesis
2.1 Theoretical Bases
Biophysical studies in the 1950s and 1960s led to the hypothesis that the waterpermeability of certain biological membranes had to be due to the presence of waterpores in the cells [1].
In general, the following parameters were taken into account to explore that hypothesis:
1. The diffusional water permeability coefficient (Pdw) and the osmotic waterpermeability coefficient (Posm). Pdw considers the rate of water transfer across amembrane in the absence of an osmotic gradient, while Posm reflects the watertransport in the presence of such gradient. The ratio of osmotic-to-diffusional waterpermeability (ΔPosm/ΔPdw) is considered a parameter that gives information aboutthe existence of water pores: In pure lipid membranes, the ΔPosm/ΔPdw ratio is equalto unity. On the other hand, when water channels are present, ΔPosm/ΔPdw will begreater than one.
2. The activation energy (Ea) for both parameters. Ea provides a measure of the energybarrier to water movement across a membrane. It is usually assumed that Ea for watermovement across a lipid membrane must be higher than Ea for water movementthrough water channels.
3. The Staverman’s reflection coefficient of the membrane (σ) for a series of non-electrolytic nonlipid-soluble molecules [2, 3], which takes into account the propertiesof a membrane to be more or less permeable to a solute, with the limits being zerowhen a solute has no difficulties to go through the membrane and unity when themembrane is impermeable to it.
2.2 Experimental Support for the Pore Hypothesis
Pioneering work on water permeability across the cell membrane was performed, at the endof the 1950s, by Solomon and co-workers [4–6]. The experimental methodology usedhuman red cells with measurements of tritiated water fluxes to estimate Pdw and light-scattering analyses to evaluate cell volume changes to calculate Posm. Experimentally, light-scattering implies a rapid mix of a cell suspension with an anisosmotic solution to create atransmembrane osmotic gradient. Osmotic water flux produces a change in cell volume andscattered light intensity, and then Posm can be calculated from the temporal changes.
According to Ussing, a difference between the permeability constants for diffusion(Pdw) and “filtration” (Posm) of water is indicative of the presence of a water-filled channelin the membrane. According to this model, water was considered as moving acrosscylindrical and rectilinear pores. The picture of a membrane perforated by cylindrical anduniform pores is an idealized geometrical approximation but was considered as aconvenient model, rather than a structural reality [4].
332 M. Parisi et al.

Equations based on Fick’s law were commonly used to model diffusion processes;meanwhile, Poiseuille’s law was applied to study nondiffusional fluid movements. Inaddition, studies of the entrance rate of a number of nonelectrolytic lipid-insolublemolecules of graded size into the human red cell and the generation of theoretical curves,with the pore radius as a parameter, correlated Staverman’s reflection coefficient σ to anequivalent pore radius of 4.2 Å [6].
A similar formalism was applied in those early years to measure water permeabilityproperties in different cells, for example in the isolated single fibers of frog muscle [7] andin the giant nerve fiber of the squid [8], complementing the observations with optical orelectron microscope microphotography, respectively.
3 Unstirred Layers and Artificial Lipid Bilayers
In the mid-1960s, determinations of Pdw made in frog skin demonstrated that the values ofthis coefficient, measured using tritiated water, changed with the stirring rate of the mediumbathing the epithelium. These experiments demonstrated that Pdw values were under-estimated because of the artifactual influence of unstirred layers [9, 10]. Inside a thin regionnext to the membrane, the tagged species moving through the membrane had aconcentration that was not equal to that in the bulk solution. Therefore, corrections forunstirred layers had to be made in determinations of Pdw.
As a next step on this direction, careful measurements of tritiated unidirectional fluxesand net water movements were performed in artificial pure lipid membranes. The results ofthose experiments demonstrated that, after correction of unstirred layers effects, Pdw wasequal to Posm in these systems [11, 12]. Consequently, the difference between Posm andPdw was explained in terms of unstirred layers, and hence the “pore model” was stronglyquestioned. The alternative hypothesis (the “partition–diffusion model”) established that thewater molecules partitioned between the water phase and the lipid phase and then diffusedthrough the membrane, crossing it until the next interphase where they left the lipid andreturned to an aqueous phase.
4 Artificial Water Channels
The addition of artificial channels to lipid bilayers allowed the second birth of the “specificwater channels hypothesis”. One of these channels was gramicidin A, a polypeptidicantibiotic that forms pores when it is incorporated into the membrane. Experiments byFinkelstein’s group confirmed that, in pure lipid membranes, Posm was equal to Pdw,supporting in this case the “partition–diffusion model”. Nevertheless, if a “channel-forming” molecule was added, it was possible to demonstrate that ΔPosm/ΔPdw=n, wheren was a number characterizing the inserted channel, indicating that the increase in the watermovement occurred through the pores [13, 14]. These results revitalized the pore theory andgave rise to the “single-file hypothesis” [15]. Because the pore radius of the gramicidinchannel is about 2 Å, then the water molecules can cross it one after the other withoutpassing each other inside the channel. The difference between Posm and Pdw was explainedby the different pathways the water takes to cross a membrane with pores, and theΔPosm/ΔPdwrate was equal to a constant that is characteristic of the particular channel involved. This constantswitched from 1 in pure lipid membranes to 5 when the polypeptide gramicidin Awas insertedand indicated the number of water molecules moving in single file inside the channel [13, 14].
From membrane pores to aquaporins 333

During the 1990s, computational simulations were done to study the behavior of watermoving across a bilayer membrane containing pores [16, 17]. Considering gramicidin A asthe channel incorporated into the membrane, the water permeability was calculated usingthe formal correspondence between diffusion and random walk: The results agreed closelywith those obtained experimentally by Rosenberg and Finkelstein in 1978 [13].
Assuming water movement through rectilinear pores, the discussion at the end of the1970s was centered on elucidating the mechanism that propels the water molecules throughthe channel.
5 The Antidiuretic Hormone-controlled Water Channel
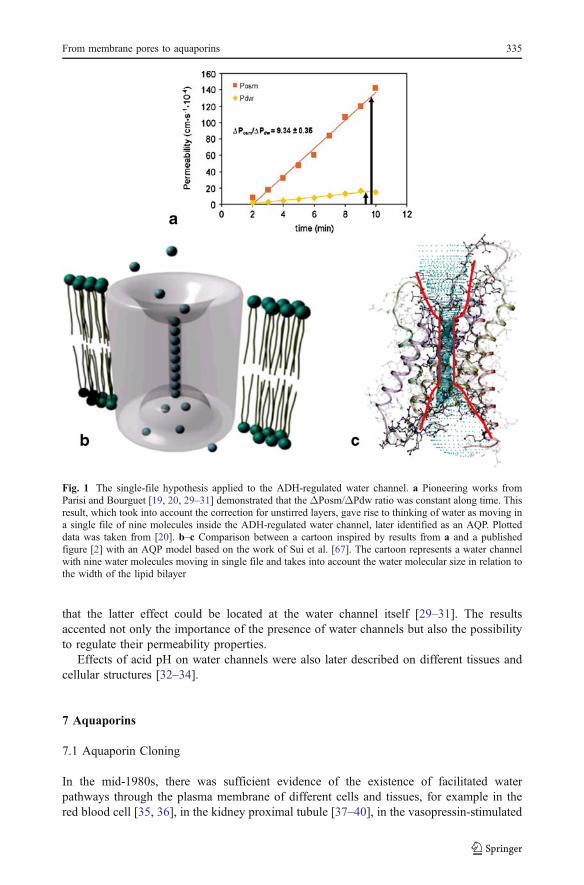
The ideas described above (i.e., the impact of the existence of an unstirred layer, theconsequences of the value of the ΔPosm/ΔPdw ratio, and the idea of water moleculesmoving in a single file) were applied to study the water channel controlled by theantidiuretic hormone (ADH). ADH induces a large increase in the water permeability of theluminal membrane of the toad urinary bladder [18]. Using an experimental setup allowingthe simultaneous minute-by-minute measurement of both the unidirectional transepithelialflux of tritiated water and the net volume of fluid traversing urinary bladder tissue, it waspostulated in a first step that the increase in water permeability induced by the hormoneaction was a “quantum” phenomenon (i.e., the addition of “permeability units” that increasein number during the development of the hormonal action) [19]. In a subsequent study, the“single-file model” was proposed to explain the water molecules’ transfer across the ADH-controlled channel [20]. The changes in both Pdw and Posm parameters were followedduring the action of the ADH at different temperatures and stirring conditions. Aftercorrection for external unstirred layer effects, the ΔPosm/ΔPdw ratio remained remarkablyconstant (n~9) under ADH action. On the other hand, in nonstimulated preparations, thisratio was not different from 1. These results, together with previous observations,demonstrating that other small molecules (like urea) are excluded from the ADH-inducedchannel [21], indicated that probably nine water molecules move in a single file throughthis structure [20]. The observed results are summarized in Fig. 1 (inspired by results in[20]). Levine et al. proposed a similar model from independent results [22].
Nowadays, it is known that ADH affects water transfer across epithelial barriers via twodifferent mechanisms: (1) directly, regulating the number of AQP2 water channels insertedin the membrane [23–25], and (2) indirectly, inducing water transfer associated with anincrease in Na+ active transport [26].
6 Water Permeability and pH of the Media
Inhibitory effects of low pH on the ADH-induced response were reported in earlier studies[27, 28].
While ADH regulates the number of permeability units (channels), an on–off mechanismwas proposed to explain this acidic pH effect on toad urinary bladder cells. The inhibitionwas only observed in the presence of a permeant buffer that acidified the intracellularmedium, even when the mucosal pH was kept constant. These results could be explained bypostulating intracellular loci sensitive to the medium acidification. The fact that the increasein the number of intramembranous particle aggregates (associated with the hormone-induced water permeability) did not disappear after flow inhibition by acid pH, suggested
334 M. Parisi et al.
that the latter effect could be located at the water channel itself [29–31]. The resultsaccented not only the importance of the presence of water channels but also the possibilityto regulate their permeability properties.
Effects of acid pH on water channels were also later described on different tissues andcellular structures [32–34].
7 Aquaporins
7.1 Aquaporin Cloning
In the mid-1980s, there was sufficient evidence of the existence of facilitated waterpathways through the plasma membrane of different cells and tissues, for example in thered blood cell [35, 36], in the kidney proximal tubule [37–40], in the vasopressin-stimulated
Fig. 1 The single-file hypothesis applied to the ADH-regulated water channel. a Pioneering works fromParisi and Bourguet [19, 20, 29–31] demonstrated that the ΔPosm/ΔPdw ratio was constant along time. Thisresult, which took into account the correction for unstirred layers, gave rise to thinking of water as moving ina single file of nine molecules inside the ADH-regulated water channel, later identified as an AQP. Plotteddata was taken from [20]. b–c Comparison between a cartoon inspired by results from a and a publishedfigure [2] with an AQP model based on the work of Sui et al. [67]. The cartoon represents a water channelwith nine water molecules moving in single file and takes into account the water molecular size in relation tothe width of the lipid bilayer
From membrane pores to aquaporins 335

kidney collecting duct [41, 42], and its functional analogous epithelium, the amphibianurinary bladder [19, 20].
The idea that water permeability in tissues was strongly associated with a proteinstructure was rapidly gaining strength [43]. In 1990, Verkman’s experiments injecting totalmessenger ribonucleic acid (RNA) of different tissues into Xenopus laevis oocytesdemonstrated functional expression of water channels, indicating that water channels areeffectively proteins [44].
In 1992, Agre and co-workers reported the appearance of water channels in X. laevisoocytes, expressing a red cell membrane protein called CHIP28 [45], in a success thatrepresents the first cloning of water channels, later called AQPs. It is interesting enough tonote that the measured parameters used to establish the expression of water channels afterinjection of genetic material were again those used during the previous 40 years: the Pdw,the Posm, the ΔPosm/ΔPdw value, and the Ea for both parameters.
In 1985, Parisi and Bourguet proposed that water channels are widely present in cellmembranes of animal cells [46]. It then was a conservative idea, and now an impressivenumber of AQPs have been cloned, going from bacteria to humans, through all kingdomsof life [47]. In plants, for instance, only in Arabidopsis thaliana, 35 AQP genes have beendetected. In mammals, the AQP family has grown to 13 members (AQP0 to AQP12).Based on their permeability characteristics, this AQP family has traditionally been dividedinto two classes: the AQPs that only conduct water (AQP0, AQP1, AQP2, AQP4, AQP5,AQP6, and AQP8) and the aquaglyceroporins that also transport small solutes, in particular,glycerol (AQP3, AQP7, AQP9, and AQP10, reviewed in [48, 49]). The measuredparameter to differentiate aquaglyceroporins from AQPs, related to their permeability tosmall solutes, is the reflection coefficient (σ) for a series of nonelectrolytic nonlipid-solublemolecules. AQP11 and AQP12 appear to be more distantly related to the other mammalianAQPs and aquaglyceroporins in their transport function. Although AQP0 is included in thefirst group, the water permeability of this protein, originally named major intrinsicpolypeptide (MIP), is approximately 100-fold lower than AQP1 [50], conferring only four- tofivefold increase in water permeability over the intrinsic permeability of lipid bilayers. Sincethese earlier physiological studies, AQP0 was thought to have another physiological function[51]. Based on the structural determination and molecular dynamic simulation studies, it wasrecently proposed that AQP0 could be an “adhennel”, a term that describes a membraneprotein that function both as a cell adhesion molecule and a membrane channel [52, 53].
7.2 Aquaporin Structure
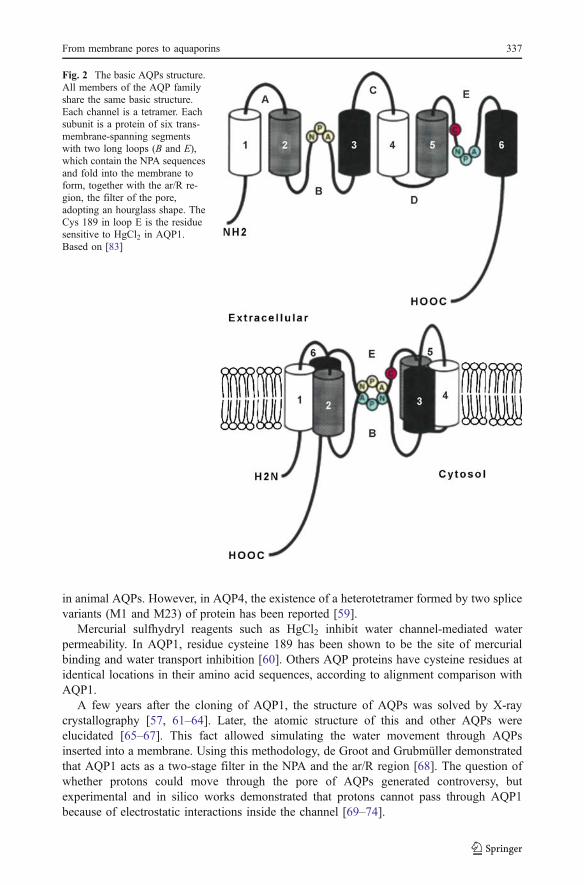
The AQPs are small, very hydrophobic, intrinsic membrane proteins of about 270 aminoacids, having six membrane-spanning α-helices and two long loops, one cytosolic (loop B)and the other extracytosolic (loop E). The N and C termini of the proteins are intracellular.Predicted sizes for the monomer of the mammalian AQPs range from 27 to 31 kDa [54].The sequence alignment of the AQPs shows several highly conserved motifs, including tworepeating Asn-Pro-Ala (NPA) [55], the signature sequence motif of these proteins, each onelocalized in a pointed loop. These NPA motifs are juxtaposed to one another and, togetherwith a highly conserved aromatic/arginine (ar/R) region located above the NPAs, form thenarrowest constriction sites (Fig. 2).
AQPs are present in the membrane as tetramers, but the channel for water permeabilitydoes not reside in the center of the tetramer (as it does for ion channels). Instead, eachmonomer contains a channel [55]. Structural studies provided insights into the requirementfor homotetramer formation for the expression of water transport activity [56–58], at least
336 M. Parisi et al.
in animal AQPs. However, in AQP4, the existence of a heterotetramer formed by two splicevariants (M1 and M23) of protein has been reported [59].
Mercurial sulfhydryl reagents such as HgCl2 inhibit water channel-mediated waterpermeability. In AQP1, residue cysteine 189 has been shown to be the site of mercurialbinding and water transport inhibition [60]. Others AQP proteins have cysteine residues atidentical locations in their amino acid sequences, according to alignment comparison withAQP1.
A few years after the cloning of AQP1, the structure of AQPs was solved by X-raycrystallography [57, 61–64]. Later, the atomic structure of this and other AQPs wereelucidated [65–67]. This fact allowed simulating the water movement through AQPsinserted into a membrane. Using this methodology, de Groot and Grubmüller demonstratedthat AQP1 acts as a two-stage filter in the NPA and the ar/R region [68]. The question ofwhether protons could move through the pore of AQPs generated controversy, butexperimental and in silico works demonstrated that protons cannot pass through AQP1because of electrostatic interactions inside the channel [69–74].
Fig. 2 The basic AQPs structure.All members of the AQP familyshare the same basic structure.Each channel is a tetramer. Eachsubunit is a protein of six trans-membrane-spanning segmentswith two long loops (B and E),which contain the NPA sequencesand fold into the membrane toform, together with the ar/R re-gion, the filter of the pore,adopting an hourglass shape. TheCys 189 in loop E is the residuesensitive to HgCl2 in AQP1.Based on [83]
From membrane pores to aquaporins 337

The crystallography of the AQPs and the determination of their structures at a 2.2-Åresolution [67] confirmed the model of the channel proposed prior to the AQP cloning(Fig. 2), based on previous biophysical measurements [2]. Furthermore, moleculardynamics simulations also confirmed previous hypotheses like the single-file movement.
8 Physiological Significance of Aquaporin Expression in Animal Cells
The role of AQP2 in the renal collecting duct is a clearer situation with regard to AQP’sphysiological function. The absence of AQP2 expression in the human kidney induces thedevelopment of diabetes insipidus. Nevertheless, the role of AQPs in the isotonic watertransfer, associated with solute transport, is not so clearly established. For instance, activefluid transport is AQP-independent in many tissues, as it occurs during intestinal fluidsecretion stimulated with different agonists (for example, cholera toxin) [75, 76]. In ahuman intestinal cell line (T84), Escherichia coli STa toxin induced a water secretory flux,despite the lack of AQPs in these cells [77]. In the same direction, the loss of the ADH-induced hydrosmotic response was observed in a rat cortical collecting renal cell line(RCCD1) not expressing AQPs, while the water net transfer associated to sodium transportwas preserved [78]. Nonetheless, after RCCD1 transfection with AQP2, the hydrosmoticresponse was restored [79].
In the same way, Hill et al. [80] and Fischbarg et al. [81] cited specific cases where thegenetic elimination or the reduction in AQP expression does not cause the pathologicaleffects attributed to water transport reduction. Moreover, Hill et al. also proposed ahypothesis considering that the main function of AQPs is to perform as osmotic and turgorpressure sensors.
Other proteins besides AQPs appear to be able to transport water [82–84], but themechanism is still controversial [85–87]. For instance, a water transport pathway has beendescribed to be associated with the glucose transporter [88–90] and different Na–solutecotransporters [91, 92].
9 Measurement of Water Permeability in Cells Expressing Aquaporins
Different biophysical approaches were used to study transport activity and its regulation inAQP-transfected cells. Those studies were performed in isolated cells or in monolayer-forming culture cells. The techniques included stopped-flow light scattering [93], totalinternal reflection fluorescence microscopy [94], laser interferometry [95], Fourier opticsdark-field/phase-contrast microscopy [96], and “optical water flux bidirectional registra-tion”, a technique that allows the measurement of real-time bidirectional transepithelialwater flux [97]. Nevertheless, the classic technique to evaluate AQP activity is the Xenopusoocytes osmotic swelling assay. In this, X. laevis oocytes expressing an AQP complementaryRNA are subjected to a sudden decrease in the extracellular solution osmolarity. Quantitativeimage analysis is then used to measure the time course of oocyte volume changes and toestimate Posm [44, 98].
However, the previously mentioned approach using Xenopus oocytes is limited becauseof the uncertainty of the oocyte’s internal composition, since cytoplasmic content is farfrom being an ideal water reservoir. The internal side of the oocyte is a heterogeneous andviscous medium with the presence of diffusional barriers and areas that may be osmoticallyinsensitive. A further step is the use of the emptied-out Xenopus oocyte technique, which
338 M. Parisi et al.
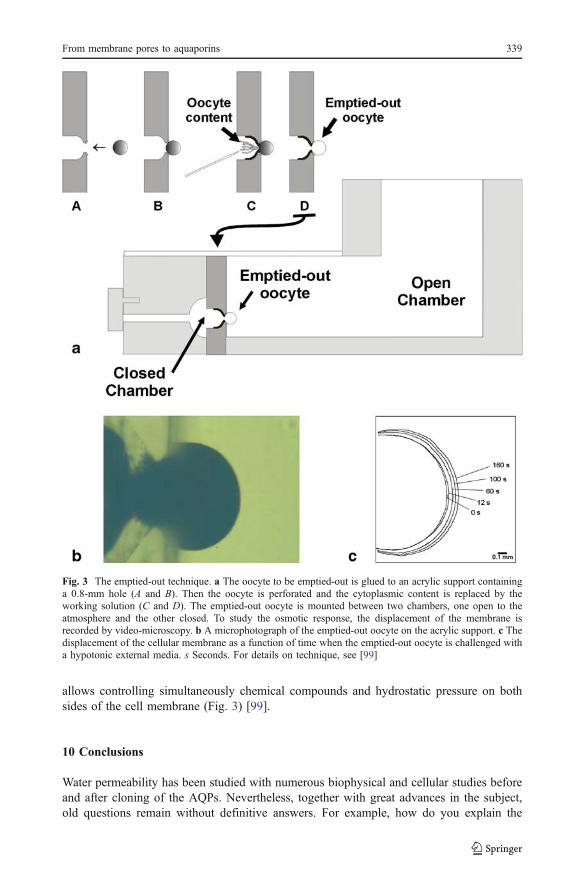
allows controlling simultaneously chemical compounds and hydrostatic pressure on bothsides of the cell membrane (Fig. 3) [99].
10 Conclusions
Water permeability has been studied with numerous biophysical and cellular studies beforeand after cloning of the AQPs. Nevertheless, together with great advances in the subject,old questions remain without definitive answers. For example, how do you explain the
Fig. 3 The emptied-out technique. a The oocyte to be emptied-out is glued to an acrylic support containinga 0.8-mm hole (A and B). Then the oocyte is perforated and the cytoplasmic content is replaced by theworking solution (C and D). The emptied-out oocyte is mounted between two chambers, one open to theatmosphere and the other closed. To study the osmotic response, the displacement of the membrane isrecorded by video-microscopy. b A microphotograph of the emptied-out oocyte on the acrylic support. c Thedisplacement of the cellular membrane as a function of time when the emptied-out oocyte is challenged witha hypotonic external media. s Seconds. For details on technique, see [99]
From membrane pores to aquaporins 339

development of forces that govern the mechanism of osmosis inside the channel at themolecular level? Which is the molecular mechanism that differentiates diffusion fromosmosis in a “single-file” channel? At present, some approaches that cannot be usedbecause of technical impossibilities could be probed thanks to molecular dynamicssimulations, but new experimental developments are needed to support significant “insilico” results.
Acknowledgments The authors want to thank to Dr Adrian Roitberg (University of Florida) for reading andcorrecting the original manuscript of this paper. This work was supported by grants from UniversidadNacional de Buenos Aires, Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) andFondo Nacional para la Ciencia y la Tecnología (FONCYT), República Argentina.
References
1. Hohmann, S., Nielsen, S.: Preface. In: Hohmann, S., Nielsen, S. (eds.) Molecular Biology andPhysiology of Water and Solute Transport, pp. v–vi. Plenum, New York (2000)
2. Parisi, M., Amodeo, G., Capurro, C., Dorr, R., Ford, P., Toriano, R.: Biophysical properties of epithelialwater channels. Biophys. Chem. 68(1–3), 255–263 (1997)
3. Verkman, A.S.: Water permeability measurement in living cells and complex tissues. J. Membr. Biol. 173(2), 73–87 (2000)
4. Sidel, V.W., Solomon, A.K.: Entrance of water into human red cells under an osmotic pressure gradient.J. Gen. Physiol. 41(2), 243–257 (1957)
5. Paganelli, C.V., Solomon, A.K.: The rate of exchange of tritiated water across the human red cellmembrane. J. Gen. Physiol. 41(2), 259–277 (1957)
6. Goldstein, D.A., Solomon, A.K.: Determination of equivalent pore radius for human red cells by osmoticpressure measurement. J. Gen. Physiol. 44, 1–17 (1960)
7. Zadunaisky, J.A., Parisi, M.N., Montoreano, R.: Effect of antidiuretic hormone on permeability of singlemuscle fibres. Nature 200, 365–366 (1963)
8. Villegas, R., Villegas, G.M.: Characterization of the membranes in the giant nerve fiber of the squid. J.Gen. Physiol. 43, 73–103 (1960)
9. Dainty, J., House, C.R.: Unstirred layers in frog skin. J. Physiol. 182(1), 66–78 (1966)10. Dainty, J., House, C.R.: An examination of the evidence for membrane pores in frog skin. J. Physiol. 185
(1), 172–184 (1966)11. Hanai, T., Haydon, D.A., Redwood, W.R.: The water permeability of artificial bimolecular leaflets: a
comparison of radio-tracer and osmotic methods. Ann. NY Acad. Sci. 137(2), 731–739 (1966)12. Hanai, T., Haydon, D.A.: The permeability to water of bimolecular lipid membranes. J. Theor. Biol. 11
(3), 370–382 (1966)13. Rosenberg, P.A., Finkelstein, A.: Water permeability of gramicidin a-treated lipid bilayer membranes. J.
Gen. Physiol. 72(3), 341–350 (1978)14. Finkelstein, A., Andersen, O.S.: The gramicidin A channel: a review of its permeability characteristics
with special reference to the single-file aspect of transport. J. Membr. Biol. 59(3), 155–171 (1981)15. Urban, B.W., Hladky, S.B., Haydon, D.A.: Ion movements in gramicidin pores. An example of single-
file transport. Biochim. Biophys. Acta 602(2), 331–354 (1980)16. Chui, S.W., Jakobsson, E., Subramahiam, S., McCammon, J.A.: Time-correlation analysis of simulated
water motion in flexible and rigid gramicidin channels. Biophys. J. 60, 273–285 (1991)17. Chui, S.W., Subramahiam, S., Jakobsson, E.: Simulation study of a gramicidin/lipid bilayer system in
excess water and lipid. I. Rates and mechanisms of water transport. Biophys. J. 76, 1929–1938 (1999)18. Muller, J., Kachadorian, W.A., Discala, V.A.: Evidence that ADH-stimulated intramembrane particle
aggregates are transferred from cytoplasmic to luminal membranes in toad bladder epithelial cells. J. CellBiol. 85, 83–95 (1980)
19. Parisi, M., Bourguet, J., Ripoche, P., Chevalier, J.: Simultaneous minute by minute determination ofunidirectional and net water fluxes in frog urinary bladder. A reexamination of the two barriers in serieshypothesis. Biochim. Biophys. Acta 556(3), 509–523 (1979)
340 M. Parisi et al.

20. Parisi, M., Bourguet, J.: The single file hypothesis and the water channels induced by antidiuretichormone. J. Membr. Biol. 71(3), 189–193 (1983)
21. Hays, R.M., Carvounis, C.P., Franki, N., Levine, S.D.: Water permeation in epithelial tissues: currentconcepts. In: Bourguet, J., Chevalier, J., Parisi, M., Ripoche, P. (eds.) Hormonal Control of EpithelialTransport, vol 85, pp. 281–288. INSERM, Paris (1979)
22. Levine, S.D., Jacoby, M., Finkelstein, A.: The water permeability of toad urinary bladder. II. The valueof Pf/Pd(w) for the antidiuretic hormone-induced water permeation pathway. J. Gen. Physiol. 83(4), 543–61 (1984)
23. Fushimi, K., Uchida, S., Hara, Y., Hirata, Y., Marumo, F., Sasaki, S.: Cloning and expression of apicalmembrane water channel of rat kidney collecting tubule. Nature 361, 549–552 (1993)
24. Nielsen, S., Chou, C.L., Marples, D., Christensen, E.I., Kishore, B.K., Knepper, M.A.: Vasopressinincreases water permeability of kidney collecting duct by inducing translocation of aquaporin-CD waterchannels to plasma membrane. Proc. Natl. Acad. Sci. USA 92, 1013–1017 (1995)
25. Toriano, R., Ford, P., Rivarola, V., Tamarappoo, B.K., Verkman, A.S., Parisi, M.: Reconstitution of aregulated transepithelial water pathway in cells transfected with AQP2 and an AQP1/2 hybrid containingthe AQP2-C-terminus. J. Membr. Biol. 161, 141–149 (1998)
26. Johnsen, A.H., Nielsen, R.: Enhanced sensitivity to stimulation of sodium transport and cyclic AMP byantidiuretic hormone after Ca2+ depletion of isolated frog skin epithelium. J. Membr. Biol. 69, 137–143(1982)
27. Gulyassy, P.F., Edelman, I.S.: Hydrogen ion dependence of the antidiuretic action of vasopressin,oxytocin, and deaminooxytocin. Biochim. Biophys. Acta 102, 185–197 (1965)
28. Carvounis, C.P., Levine, S.D., Hays, R.M.: pH dependence of water and solute transport in toad urinarybladder. Kidney Int. 15, 513–519 (1979)
29. Parisi, M., Chevalier, J., Bourguet, J.: Influence of mucosal and serosal pH on antidiuretic action in frogurinary bladder. Am. J. Physiol. 237(6), F483–F489 (1979)
30. Parisi, M., Montoreano, R., Chevalier, J., Bourguet, J.: Cellular pH and water permeability control infrog urinary bladder. A possible action on the water pathway. Biochim. Biophys. Acta 648(2), 267–274(1981)
31. Parisi, M., Bourguet, J.: Effects of cellular acidification on ADH-induced intramembrane particleaggregates. Am. J. Physiol. 246(1 Pt 1), C157–C159 (1984)
32. Amodeo, G., Sutka, M., Dorr, R., Parisi, M.: Protoplasmic pH modifies water and solute transfer in Betavulgaris root vacuoles. J. Membr. Biol. 187(3), 175–184 (2002)
33. Hedfalk, K., Törnroth-Horsefield, S., Nyblom, M., Johanson, U., Kjellbom, P., Neutze, R.: Aquaporingating. Curr. Opin. Struct. Biol. 16(4), 447–456 (2006)
34. Chaumont, F., Moshelion, M., Daniels, M.J.: Regulation of plant aquaporin activity. Biol. Cell 97(10),749–764 (2005)
35. Macey, R.I.: Transport of water and urea in red blood cells. Am. J. Physiol. 246, C195–C203 (1984)36. Solomon, A.K., Chasan, B., Dix, J.A., Lukacovic, M.F., Toon, M.R., Verkman, A.S.: The aqueous pore
in the red cell membrane: band 3 as a channel for anions, cations, nonelectrolytes, and water. Ann. NYAcad. Sci. 414, 79–124 (1984)
37. Verkman, A.S., Weyer, P., Brown, D., Ausiello, D.A.: Functional water channels are present in clathrin-coated vesicles from bovine kidney but not from brain. J. Biol. Chem. 264, 20608–20613 (1989)
38. Meyer, M.M., Verkman, A.S.: Evidence for water channels in renal proximal tubule cell membranes. J.Membr. Biol. 96, 107–119 (1987)
39. Whittembury, G., Carpi-Medina, P., Gonzalez, E., Linares, H.: Effect of para-chloromercuribenzene-sulfonic acid and temperature on cell water osmotic permeability of proximal straight tubules. Biochim.Biophys. Acta 775(3), 365–373 (1984)
40. Ye, R., Shi, L.B., Lencer, W., Verkman, A.S.: Functional colocalization of water channels and protonpumps in endosomes from kidney proximal tubule. J. Gen. Physiol. 93, 885–902 (1989)
41. Al-Zahid, G., Schafer, J.A., Troutman, S.L., Andreoli, T.E.: Effect of antidiuretic hormone on water andsolute permeation, and the activation energies for these processes, in mammalian cortical collectingtubules: evidence for parallel ADH-sensitive pathways for water and solute diffusion in luminal plasmamembranes. J. Membr. Biol. 31, 103–129 (1977)
42. Verkman, A.S., Lencer, W.I., Brown, D., Ausiello, D.A.: Endosomes from kidney collecting tubule cellscontain the vasopressin-sensitive water channel. Nature 333, 268–269 (1988)
43. Brown, D.: Membrane recycling and epithelial cell function. Am. J. Physiol. 256, F1–F12 (1989)44. Zhang, R., Logee, K.A., Verkman, A.S.: Expression of mRNA coding for kidney and red cell water
channels in Xenopus oocytes. J. Biol. Chem. 265(26), 15375–15378 (1990)45. Preston, G.M., Carroll, T.P., Guggino, W.B., Agre, P.: Appearance of water channels in Xenopus oocytes
expressing red cell CHIP28 protein. Science 256(5055), 385–387 (1992)
From membrane pores to aquaporins 341

46. Parisi, M., Bourguet, J.: Water channels in animal cells: a widespread structure? Biol. Cell. 55(3), 155–157 (1985)
47. Agre, P.: Nobel Lecture. Aquaporin water channels. Biosci. Rep. 24(3), 127–163 (2004)48. King, L.S., Kozono, D., Agre, P.: From structure to disease: the evolving tale of aquaporin biology. Nat.
Rev. Mol. Cell Biol. 5, 687–698 (2004)49. Takata, K., Matsuzaki, T., Tajika, Y.: Aquaporins: water channel proteins of the cell membrane. Prog.
Histochem. Cytochem. 39, 1–83 (2004)50. Zampighi, G.A., Kreman, M., Boorer, K.J., Loo, D.D., Bezanilla, F., Chandy, G., Hall, J.E., Wright,
E.M.: A method for determining the unitary functional capacity of cloned channels and transportersexpressed in Xenopus laevis oocytes. J. Membr. Biol. 148(1), 65–78 (1995)
51. Mulders, S.M., Preston, G., Deen, P., Guggino, W., Van Os, C., Agre, P.: Water channel properties ofmajor intrinsic protein of lens. J. Biol. Chem. 270, 9010–9016 (1995)
52. Engel, A., Fujiyoshi, Y., Gonen, T., Walz, T.: Junction-forming aquaporins. Curr. Opin. Struct. Biol.2008 Jan 12 [Epub ahead of print] In press
53. Han, B.G., Guliaev, A.B., Walian, P.J., Jap, B.K.: Water transport in AQP0 aquaporin: moleculardynamics studies. J. Mol. Biol. 360(2), 285–96 (2006)
54. Verkman, A.S., Mitra, A.K.: Structure and function of aquaporin water channels. Am. J. Physiol. 278,F13–F28 (2000)
55. Jung, J.S., Preston, G.M., Smith, B.L., Guggino, W.B., Agre, P.: Molecular structure of the water channelthrough aquaporin CHIP. The hourglass model. J. Biol. Chem. 269, 14648–14654 (1994)
56. Smith, B.L., Agre, P.: Erythrocyte Mr 28,000 transmembrane protein exists as a multisubunit oligomersimilar to channel proteins. J. Biol. Chem. 266, 6407–6415 (1991)
57. Walz, T., Smith, B.L., Zeidel, M.L., Engel, A., Agre, P.: Biologically active two-dimensional crystals ofaquaporin CHIP. J. Biol. Chem. 269, 1583–1586 (1994)
58. Mathai, J.C., Agre, P.: Hourglass pore-forming domains restrict aquaporin-1 tetramer assembly.Biochemistry 38, 923–928 (1999)
59. Neely, J.D., Christensen, B.M., Nielsen, S., Agre, P.: Heterotetrameric composition of aquaporin-4 waterchannels. Biochemistry 38, 11156–11163 (1999)
60. Preston, G.M., Jung, J.S., Guggino, W.B., Agre, P.: The mercury-sensitive residue at cysteine 189 in theCHIP28 water channel. J. Biol. Chem. 268, 17–20 (1993)
61. Walz, T., Hirai, T., Murata, K., Heymann, J.B., Mitsuoka, K., Fujiyoshi, Y., Smith, B.L., Agre, P., Engel,A.: The three-dimensional structure of aquaporin-1. Nature 387, 624–627 (1997)
62. Cheng, A., van Hoek, A.N., Yeager, M., Verkman, A.S., Mitra, A.K.: Three-dimensional organization ofa human water channel. Nature 387, 627–630 (1997)
63. Mitsuoka, K., Murata, K., Walz, T., Hirai, T., Agre, P., Heymann, J.B., Engel, A., Fujiyoshi, Y.: Thestructure of aquaporin-1 at 4.5-Å resolution reveals short alpha-helices in the center of the monomer. J.Struct. Biol. 128, 34–43 (1999)
64. Ren, G., Cheng, A., Reddy, V., Melnyk, P., Mitra, A.K.: Three-dimensional fold of the human AQP1water channel determined at 4 Å resolution by electron crystallography of two-dimensional crystalsembedded in ice. J. Mol. Biol. 301, 369–387 (2000)
65. Murata, K., Mitsuoka, K., Hirai, T., Walz, T., Agre, P., Heymann, J.B., Engel, A., Fujiyoshi, Y.:Structural determinants of water permeation through aquaporin-1. Nature 407, 599–605 (2000)
66. Fu, D., Libson, A., Miercke, L.J.W., Weitzman, C., Nollert, P., Krucinski, J., Stroud, R.M.: Structure of aglycerol-conducting channel and the basis for its selectivity. Science 290, 481–486 (2000)
67. Sui, H., Han, B.G., Lee, J.K., Walian, P., Jap, B.K.: Structural basis of water-specific transport throughthe AQP1 water channel. Nature 414, 872–878 (2001)
68. de Groot, B.L., Grubmüller, H.: Water permeation across biological membranes: mechanism anddynamics of aquaporin-1 and GlpF. Science 294, 2353–2357 (2001)
69. Agre, P., Lee, M.D., Devidas, S., Guggino, W.B.: Aquaporins and ion conductance. Science 275, 1490–1492 (1997)
70. Tsunoda, S.P., Wiesner, B., Lorenz, D., Rosenthal, W., Pohl, P.: Aquaporin-1, nothing but a waterchannel. J. Biol. Chem. 279, 11364–11367 (2004)
71. Jensen, M.Ø., Tajkhorshid, E., Schulten, K.: Electrostatic tuning of permeation and selectivity inaquaporin water channels. Biophys. J. 85, 2884–2899 (2003)
72. Burykin, A., Warshel, A.: What really prevents proton transport through aquaporin? Charge self-energyversus proton wire proposals. Biophys. J. 85, 3696–3706 (2003)
73. de Groot, B.L., Engel, A., Grubmüller, H.: The structure of the aquaporin-1 water channel: a comparisonbetween cryo-electron microscopy and X-ray crystallography. J. Mol. Biol. 325, 485–493 (2003)
74. Chakrabarti, N., Roux, B., Pomès, R.: Structural determinants of proton blockage in aquaporins. J. Mol.Biol. 343, 493–510 (2004)
342 M. Parisi et al.

75. Yang, B., Song, Y., Zhao, D., Verkman, A.S.: Phenotype analysis of aquaporin-8 null mice. Am. J.Physiol. Cell. Physiol. 288, C1161–C1170 (2005)
76. Wang, K.S., Ma, T., Filiz, F., Verkman, A.S., Bastidas, J.A.: Colon water transport in transgenic micelacking aquaporin-4 water channels. Am. J. Physiol. Gastrointest. Liver Physiol. 279, G463–G670(2000)
77. Toriano, R., Kierbel, A., Ramirez, M.A., Malnic, G., Parisi, M.: Spontaneous water secretion in T84cells: effects of STa enterotoxin, bumetanide, VIP, forskolin, and A-23187. Am. J. Physiol. Gastrointest.Liver Physiol. 281, 816–822 (2001)
78. Capurro, C., Rivarola, V., Kierbel, A., Escoubet, B., Farman, N., Blot-Chabaud, M., Parisi, M.:Vasopressin regulates water flow in a rat cortical collecting duct cell line not containing knownaquaporins. J. Membr. Biol. 179, 63–70 (2001)
79. Chara, O., Ford, P., Rivarola, V., Parisi, M., Capurro, C.: Asymmetry in the osmotic response of a ratcortical collecting duct cell line: role of aquaporin-2. J. Membr. Biol. 207, 143–150 (2005)
80. Hill, A.E., Shachar-Hill, B., Shachar-Hill, Y.: What are aquaporins for? J. Membr. Biol. 197, 1–32 (2004)81. Fischbarg, J., Diecke, F.P., Iserovich, P., Rubashkin, A.: The role of the tight junction in paracellular fluid
transport across corneal endothelium. Electro-osmosis as a driving force. J. Membr. Biol. 210(2), 117–130 (2006)
82. Agre, P., Nielsen, S., Ottersen, O.P.: Towards a molecular understanding of water homeostasis in thebrain. Neuroscience 12, 849–850 (2004)
83. Agre, P., King, L.S., Yasui, M., Guggino, W.B., Ottersen, O.P., Fujiyoshi, Y., Engel, A., Nielsen, S.:Aquaporin water channels: from atomic structure to clinical medicine. J. Physiol. 542, 3–16 (2002)
84. MacAulay, N., Hamann, S., Zeuthen, T.: Water transport in the brain: role of cotransporters.Neuroscience 129, 1031–1044 (2004)
85. Gagnon, M.P., Bissonnette, P., Deslandes, L.M., Wallendorff, B., Lapointe, J.Y.: Glucose accumulationcan account for the initial water flux triggered by Na+/glucose cotransport. Biophys. J. 86, 125–133(2004)
86. Duquette, P.P., Bisonnette, P., Lapointe, J.Y.: Local osmotic gradients drive the water flux associatedwith Na+/glucose cotransport. Proc. Natl. Acad. Sci. USA 98, 3796–3801 (2001)
87. Charron, F.M., Blanchard, M.G., Lapointe, J.Y.: Intracellular hypertonicity is responsible for water fluxassociated with Na+/glucose cotransport. Biophys. J. 90, 3546–3554 (2006)
88. Fischbarg, J., Kunyan, K., Vera, J.C., Arant, S., Silverstein, S., Loike, J., Rosen, O.M.: Glucosetransporters serve as water channels. Proc. Natl. Acad. Sci. USA 87, 3244–3247 (1990)
89. Iserovich, P., Wang, D., Ma, L., Zuniga, F.A., Pascual, J.M., Kuang, K., De Vivo, D.C., Fischbarg, J.:Changes in glucose transport and water permeability resulting from the T310I pathogenic mutation inGlut1 are consistent with two transport channels per monomer. J. Biol. Chem. 277, 30991–30997 (2002)
90. Zeuthen, T., Zeuthen, E., MacAulay, N.: Water transport by GLUT2 expressed in Xenopus laevisoocytes. J. Physiol. 579, 345–361 (2007)
91. Loo, D.D., Hirayama, B.A., Meinild, A.K., Chandy, G., Zeuthen, T., Wright, E.M.: Passive water and iontransport by cotransporter. J. Physiol. 518, 195–202 (1999)
92. Zeuthen, T., Belhage, B., Zeuthen, E.: Water transport by Na+-coupled cotransporters of glucose(SGLT1) and of iodide (NIS). The dependence of substrate size studied at high resolution. J. Physiol.570, 485–499 (2006)
93. Ma, T., Frigeri, A., Tsai, S.T., Verbavatz, J.M., Verkman, A.S.: Localization and functional analysis ofCHIP28k water channels in stably transfected CHO cells. J. Biol. Chem. 268, 22756–22764 (1993)
94. Farinas, J., Simenak, V., Verkman, A.S.: Cell volume measured in adherent cells by total internalreflection microfluorimetry: application to permeability in cells transfected with water channelhomologs. Biophys. J. 68, 1613–1620 (1995)
95. Farinas, J., Verkman, A.S.: Measurement of cell volume and water permeability in epithelial cell layersby interferometry. Biophys. J. 71, 3511–3522 (1996)
96. Farinas, J., Kneen, M., Moore, M., Verkman, A.S.: Plasma membrane water permeability of culturedcells and epithelia measured by light microscopy with spatial filtering. J. Gen. Physiol. 110, 283–296(1997)
97. Dorr, R.A., Kierbel, A., Vera, J., Parisi, M.: A new data-acquisition system for the measurement of thenet water flux across epithelia. Comput. Methods Programs Biomed. 53(1), 9–14 (1997)
98. Dorr, R., Ozu, M., Parisi, M.: Simple and inexpensive hardware and software method to measure volumechanges in Xenopus oocytes expressing aquaporins. J. Neurosci. Methods 161(2), 301–5 (2007)
99. Ozu, M., Dorr, R., Parisi, M.: New method to measure water permeability in emptied-out Xenopusoocytes controlling conditions on both sides of the membrane. J. Biochem. Biophys. Methods 63(3),187–200 (2005)
From membrane pores to aquaporins 343
Copyright © 2022 FDOKUMEN




















