
Position Controller Design for Quad-rotor under Perturbed Condition
Upload
independentCategory
view
0download
0
Tc
GPRa
b
c
a
ARRAA
KEUEA
1
lpVcaFat
hw1h
-B
0d
Tissue and Cell 44 (2012) 47– 53
Contents lists available at SciVerse ScienceDirect
Tissue and Cell
jou rn al h om epage: www.elsev ier .com/ locate / t i ce
he expression of aquaporins 1 and 9 in adult rat epididymis is perturbed byhronic exposure to ethanol
iovana R. Teixeiraa,∗, Luiz Gustavo A. Chuffaa, Otávio A. Martinsa, Rafael Kremerb,atrícia Fernanda F. Pinheiroa, Wilson de Mello Júniora, Marcelo Martinezc, Francisco E. Martineza,aquel F. Domeniconia
Department of Anatomy, Institute of Biosciences, UNESP - Univ Estadual Paulista, Botucatu 18618-970, SP, BrazilDepartment of Natural Sciences, FURB - Universidade Regional de Blumenau, Blumenau 89012-900, SC, BrazilDepartment of Morphology and Pathology, UFSCar - Univ Federal de São Carlos, São Carlos 13565-905, SP, Brazil
r t i c l e i n f o
rticle history:eceived 31 August 2011eceived in revised form 31 October 2011ccepted 3 November 2011vailable online 30 November 2011
eywords:thanol
a b s t r a c t
Aquaporins (AQPs), notably AQP-1 and AQP-9, may contribute to reabsorption of fluid and solute acrossthe epididymis. Ethanol is related to be a toxicant affecting directly or indirectly the epididymis and thesperm motility. This study examined the expression of AQP-1 and AQP-9 in adult epididymis of the UChAand UChB 10% (v/v) ethanol-preferring rats, focusing the ethanol-induced hormonal disturbances uponthe regulation of these AQPs. Chronic ethanol intake significantly decreased body weight, while UChAand UChB rats displayed a marked loss of epididymal weights. Both ethanol-consuming animals had asevere reduction of testosterone levels, whereas LH and 17�-estradiol were unchanged. Throughout the
Ch ratspididymisQP-1 and AQP-9
epididymis, a strong reaction to AQP-1 was observed in myoid and endothelial cells of the UChB ethanol-preferring rats, differently from a moderate intensity in the initial segment of the UChA rats. In addition,AQP-9 showed a strong immunoreaction in the apical membrane of principal cells at initial segment. Incauda epididymis, the level of AQP-9 was reduced along the microvillus projections in both UChA andUChB rats compared to controls. We conclude that chronic ethanol consumption modulates the androgenlevels, thereby modifying the expression pattern of AQP-1 and 9 in the epididymis.
. Introduction
The absorptive and secretory capacity of the epithelial cellsining the lumen of the male excurrent duct creates the appro-riate environment for spermatozoa (Turner, 1984; Robaire andiger, 1995). The epididymis plays an essential role in sperm con-entration, maturation, transport, protection and storage (Robairend Hermo, 1988; Hermo and Robaire, 2002; Robaire et al., 2006).luid movement through epididymal epithelium is necessary tochieve significant sperm concentration, which is required for fer-ility (Robaire and Viger, 1995; Da Silva et al., 2006).
Aquaporins (AQPs) are responsible for the regulation of wateromeostasis by providing a sustained and rapid permeability of
ater with low activation energy across the epithelium (Agre et al.,993; Brown et al., 1993; King and Agre, 1996; Wintour, 1997). Itas been previously assumed that the transport of water is due to
∗ Corresponding author at: Department of Anatomy, Biosciences Institute, UNESP Univ Estadual Paulista, P.O. Box 510, 18618-000 Rubião Júnior, s/n, Botucatu, SP,razil. Tel.: +55 14 3811 6040; fax: +55 14 3811 6361.
E-mail address: [email protected] (G.R. Teixeira).
040-8166/$ – see front matter © 2011 Elsevier Ltd. All rights reserved.oi:10.1016/j.tice.2011.11.001
© 2011 Elsevier Ltd. All rights reserved.
simple diffusion through the lipid bilayer membrane surroundingcells (Huang et al., 2006). AQPs are proteins corresponding toaround 30 kDa (its non-glycosylated form) which present bothcytoplasmic N-terminal and C-terminal domains. They act asfunctional tetramers in the plasma membrane, specifically on theapical or basolateral counterpart (Agre et al., 1995).
AQPs are a family of small, hydrophobic and integral membranechannel proteins that facilitate rapid passive movement of water(Huang et al., 2006). AQPs are located in a wide variety of cells,such as kidney, lung, pancreas, brain, gastrointestinal tract, eye,ear, immune system, skin, adipocytes, muscles, uterus and testis(Li et al., 1994; Ishibashi et al., 1997; Frigeri et al., 1998; Page et al.,1998; Beitz and Schultz, 1999; Shanahan et al., 1999).
It has been clearly demonstrated that multiple AQPs are active inthe testis (AQP-0, AQP-1, AQP-7, AQP-8 and AQP-9), efferent ducts(AQP-1, AQP-9 and AQP-10), epididymis (AQP-1, AQP-3, AQP-9 andAQP-10), vas deferens (AQP-1, AQP-2 and AQP-9) and accessoryglands (AQP-1 and AQP-9) of adult mammals (Huang et al., 2006).
Nevertheless, the exact mechanism(s) controlling their differentialexpressions remains to be investigated.AQP-1 (also known as CHIP28) can modulate the bulk flowacross erythrocytes and plasma membrane (Nielsen et al., 1993;

4 ue and
Mlada(
ieiectet2g(r(9Aicsse
cicdstAmro
io1ft(
A(h
2
2
t–ta2Uwetl
8 G.R. Teixeira et al. / Tiss
a et al., 1998). Moreover, while AQP-1 is absent from the epithe-ium of the epididymis, it is present around the smooth musclend endothelial cells of vascular channels throughout the epi-idymis (Badran and Hermo, 2002; Domeniconi et al., 2008) as wells in other epithelial segments displaying reabsorptive functionsBrown et al., 1993).
AQP-9 expression has been reported in several tissues includ-ng the male reproductive tract, where it localizes in efferent ductspithelium, epididymis and vas deferens and may be involvedn sperm maturation, concentration and storage (Pastor-Solert al., 2001). These findings indicate that AQP-9 is a major api-al aquaporin in these epithelia, providing a potential route whichransepithelial fluid and solute transport can occur (Pastor-Solert al., 2002). In addition, AQP-9 is expressed in principal cells ofhe epididymis in a region-specific manner (Badran and Hermo,002) allowing the influx of a wide range of molecules, such aslycerol, urea, mannitol and sorbitol gradients beyond the waterTsukaguchi et al., 1998). Reabsorption of luminal fluid in maleeproductive tract is regulated by both androgens and estrogensHess et al., 2000). Furthermore, it has been emphasized that AQP-
is under the control of androgens (Pastor-Soler et al., 2002) whileQP-1 expression in peritubular cells of efferent duct appears to be
nhibited by androgen replacement (Oliveira et al., 2005). Despiteonsiderable effort for understanding gene expression pattern andpecific activity related to AQPs, the effects of certain compoundsuch as drug or toxicant, e.g.: alcohol, upon the selective aquaporinsxpression are not discussed.
It is currently known that ethanol consumption leads to harmfulonsequences to reproductive function. Ethanol affects directly orndirectly the male reproduction and its deleterious effects mayause testicular and epididymal atrophy, stromal and germ cellamages, reduction of prostate and seminal vesicle weights and lowperm motility (Anderson et al., 1985). Recently, it was observedhat the effects of acetaldehyde in the liver, while mediated byQP9, are probably influenced by binding to cell membrane pro-oting changes in their permeability (Potter et al., 2011). The
elationship between AQP-1 and AQP-9 and chronic ethanol intaken the epididymis has never been described.
Ethanol drinker strains (UChA and UChB) are derived from orig-nal Wistar rats and have been selectively bred at the Universityf Chile for almost sixty years (Mardones and Segovia-Riquelme,983). These ethanol-preferring rats are considered a special modelor the understanding of the basis of alcoholism-linked charac-eristics such as those found in alcohol-related human diseasesMardones and Segovia-Riquelme, 1983; Chuffa et al., 2009, 2011).
Therefore, this study aimed to verify the possible alterations ofQP-1 and AQP-9 in adult epididymis of the UChA and UChB rats
10% (v/v) ethanol-preferring rats), focusing the ethanol-inducedormonal disturbances on the regulation of these aquaporins.
. Materials and methods
.1. Animals
Thirty adult male rats, 60 days old (±250 g) were obtained fromhe Department of Anatomy, Bioscience Institute/Botucatu, UNESP
Univ Estadual Paulista. The animals were randomly divided intohree groups (n = 10/group). UChA group: rats fed 10% (v/v) ethanold libitum (free choice for water or ethanol) drinking less than.0 mL/100 g body weight (BW)/day (low ethanol consumption);ChB group: rats fed 10% (v/v) ethanol ad libitum (free choice for
ater or ethanol) drinking from 2.0 to 6.0 mL/100 g BW/day (highthanol consumption) and Control group: naïve rats without accesso ethanol. All animals were individually housed in polypropy-ene cages with laboratory-grade pine shavings as bedding and
Cell 44 (2012) 47– 53
also maintained under controlled room temperature (23 ± 1 ◦C) andlighting conditions (12L, 12D photoperiod, lights switched on at6 a.m.). The rats received a standard rodent chow (3074 SIF, PurinaLtd., Campinas, SP, Brazil) and filtered tap water ad libitum. Experi-mental protocols followed the Ethical Principles in Animal Researchand were approved by Ethics Committee on Animal Experimenta-tion.
2.2. Procedures for selection of ethanol-preferring rats
When the UChA and UChB rats reached 65 days of age, theywere given during 15 consecutive days, a choice between twobottles containing either water ad libitum or 10% (v/v) ethanolsolution. All animals were monitored and 10 animals per groupdisplaying ethanol consumption less than 2.0 mL (UChA) or higherthan 2.0 mL (UChB) of ethanol/100 g BW/day (ranging from 2to 5 g ethanol/kg/day) were finally selected in accordance toMardones and Segovia-Riquelme (1983). For this study, the prefer-ence ratio associated to ethanol-seeking behavior was about 60%.Besides that, to ensure more efficiency and maintenance of con-stant consumption throughout the experiment, the animals werekept under observation for 10 days when they started with ethanolconsumption. After 60 days of chronic ethanol intake, totaling 120days old, all animals were weighed and euthanized by decapitationat 09:00 a.m.
2.3. Morphometrical analysis
After decapitation, the left epididymides were collected, sepa-rately weighed (caput + corpus and cauda) and then fixed in Bouin’ssolution for 24 h. Afterwards, the samples were maintained in a70% alcohol solution and dehydrated in graded alcohol series fol-lowed by embedding in paraplast (Paraplast1 plus Oxford, St. Louis,USA). Tissue sections were obtained after each 4 �m thicknessand slides were prepared for morphometrical and immunohisto-chemical analysis. For morphometric data, 16 repetitions/animalwere evaluated based on cross-sectioned layers stained withhematoxylin-eosin (HE). The height of the epithelium and internaldiameters of the tubules (initial segment, caput, corpus and caudaepididymis) were measured at 100× magnification using an ImageAxion Vision 4.1 software. All tissue sections were analyzed andphotographed under a Zeiss Axiophot 2 microscope.
2.4. Immunohistochemistry analysis
For immunohistochemistry, paraplast-embedded epididymistissues were cut into 4 �m-thick and mounted on silanized slides(DAKO, Carpinteria, California). After tissue dehydration, they wereplaced in sodium citrate buffer (10 mM sodium citrate, 0.05%tween 20, pH 6.0) for antigen retrieval using a microwave heatingmethod. Briefly, the endogenous peroxidase activity was blocked byimmersing the slides in a mixed solution of 0.3% H2O2 and methanoland then, they were incubated with primary polyclonal antibod-ies anti-AQP1 and AQP9 (Chemicon Temecula, CA, USA) diluted1:100 in 1% BSA overnight at 4 ◦C. After washing three times in PBS(5 min), the tissues were incubated with secondary biotinylatedantibody goat anti-rabbit IgG (Santa Cruz Biotechnology, CA, USA)diluted 1:70 in 1% BSA at room temperature for 1 h. After sequentialwashes, the reaction was finally detected with diaminobenzidinetetrachloride-DAB (Sigma Chemical Company, St. Louis, MO, USA)
as a chromogen and sections were counter-stained with Harris’hematoxylin for 1 min 15 s, and dehydrated in 70%, 90% and 100%ethanol. Coverslips were mounted on the slides with Permount.Negative controls were performed without primary antibody. All
ue and Cell 44 (2012) 47– 53 49
at
2
rcumRtMeiaaIv4ipR
2
efwwv
3
gmoavirlsd
Table 1Diameter (�m) of the ductus epididymis (initial segment, caput, corpus and cauda)in the UChA, UChB and control rats.
Epididymis Groups
UChA UChB Control
Initial segment 243.56 ± 3.0 253.37 ± 19.1 222.63 ± 6.1Caput 360.36 ± 6.8 323.96 ± 6.6a 348.76 ± 8.4Corpus 357.61 ± 4.3 323.65 ± 3.6a 380.55 ± 4.1a,b
Cauda 427.17 ± 11.1 242.33 ± 5.1a 388.81 ± 8.4b
Results are expressed as mean ± SEM. One-way ANOVA with post hoc Tukey’s test.a p < 0.05 vs. UChA group.b p < 0.05 vs. UChB group.
Table 2Immunostaining scores of intensity for AQP-1 in all epididymis regions (initial seg-ment, caput and cauda).
Epididymis Groups
UChA UChB Control
Initial segment ++/++ ++/++ ++/+Caput +/+++ +++/+++ +/+Cauda +/++ ++/+++ +/+
The relative intensities of the immunoreaction for epididymis were graded as fol-lows: negative (−) absence of reaction; low grade (+; positive >10 and <25%);moderate (++; positive for >25 and <50%) and high (+++, positive for >50%). The first
F(
G.R. Teixeira et al. / Tiss
nalyses were carried out in an Olympus BX 41 microscope coupledo Olympus DP12 camera for images digitalization.
.5. Hormone assay
Blood samples were collected from the trunk of decapitatedats into heparinized tubes. Afterwards, plasma was obtained byentrifugation at 1200 × g for 15 min at 4 ◦C and stored at −20 ◦Cntil assayed by radioimmunoassay (RIA). Plasma luteinizing hor-one (LH) concentrations were determined by double-antibody
IA with specific kits provided by the “National Institute of Arthri-is, Diabetes, Digestive and Kidney Diseases” (NIADDK, Baltimore,
D, USA). The antiserum for LH was LH-S10 using RP3 as refer-nce. The lower limit of detection for LH was 0.2 ng/mL and thentra-assay coefficient of variation was 4%. Plasma 17-� estradiolnd testosterone concentrations were determined using Estradiolnd Testosterone Maia kits (Biochem Immunosystems, Serotec,taly). The lower detection limit and the intra-assay coefficient ofariation were respectively 7.5 pg/mL and 2.5% for estradiol and.1 ng/mL and 3.7% for testosterone. All samples were measured
n duplicate and at different dilutions, when necessary. In order torevent interassay variation, all samples were assayed in the sameIA.
.6. Statistical analysis
Values are presented as mean ± SEM. Data of plasma LH, 17-�stradiol and testosterone, and morphometric data were per-ormed by One-way ANOVA analysis of variance. Significant resultsere subjected to post hoc Tukey’s test and statistical significanceas set at p < 0.05. The statistical software was GraphPad Instat
ersion 4 and Sigma Plot version 11.0 for graphic design.
. Results
Chronic and high ethanol consumption showed a reduced BWain (UChB 315.7 ± 10.2 g vs. Control 385.0 ± 9.9 g) and also aarked loss of relative epididymal weight (caput + corpus) was
bserved in both UChA and UChB animals (UChA 0.28 ± 0.01nd UChB 0.26 ± 0.02 vs. Control 0.34 ± 0.01 g/100 g BW). Con-ersely, the cauda epididymis weight was not affected by ethanolntake after the experiment. In addition, morphometric analysis
evealed no significant results in the height of the epithelial cellayer throughout the epididymis (data not shown). There wereignificant reductions (p < 0.05) in the diameter of UChB ductus epi-idymis (caput, corpus and cauda) when compared to UChA ratsig. 1. Plasma hormone profiles of the rats in the different experimental groups. Determng/mL) levels (C). One-way ANOVA with post hoc Tukey’s test. *p < 0.05 vs. UChA and UCh
and second signal indicates AQP-1 reaction for myoid cells and endothelial vascularchannels, respectively.
and only the diameters of ductus epididymis (corpus and cauda)were found to be lower in the UChB rats than controls (Table 1).
Plasma LH and 17-� estradiol levels did not shown significantdifferences among the groups (Fig. 1A and B), whereas testosteronelevels were markedly decreased (p < 0.05) after ethanol consump-tion in the UChA and UChB rats compared to controls (Fig. 1C). Thisis consistent with our previous results (Tables 2 and 3).
The immunoreactivity for AQP-1 and AQP-9 in the epididymiswas detected in many regions. Regarding to initial segment, allanimals presented a strong labeling for AQP-1 in myoid cells(Fig. 2A–C), while both UChA and UChB rats evidenced a moder-ate AQP-1 reaction in endothelial cells compared to controls (weakdetection). Unlike the intense immunostaining noted in the caputepididymis of UChB rats, extensively in myoid cells (Fig. 2E), ahigh positive reaction was also seen in the endothelial cell mem-
brane of the UChA and UChB rats, differently from control animals(Fig. 2D–F). In cauda epididymis, a moderate and strong positivereaction at the myoid cell and apical surface of endothelial cellwas observed in UChB ethanol-preferring rats, while displaying aination of LH (ng/mL) levels (A); 17-� estradiol (pg/mL) levels (B) and testosteroneB groups, respectively.
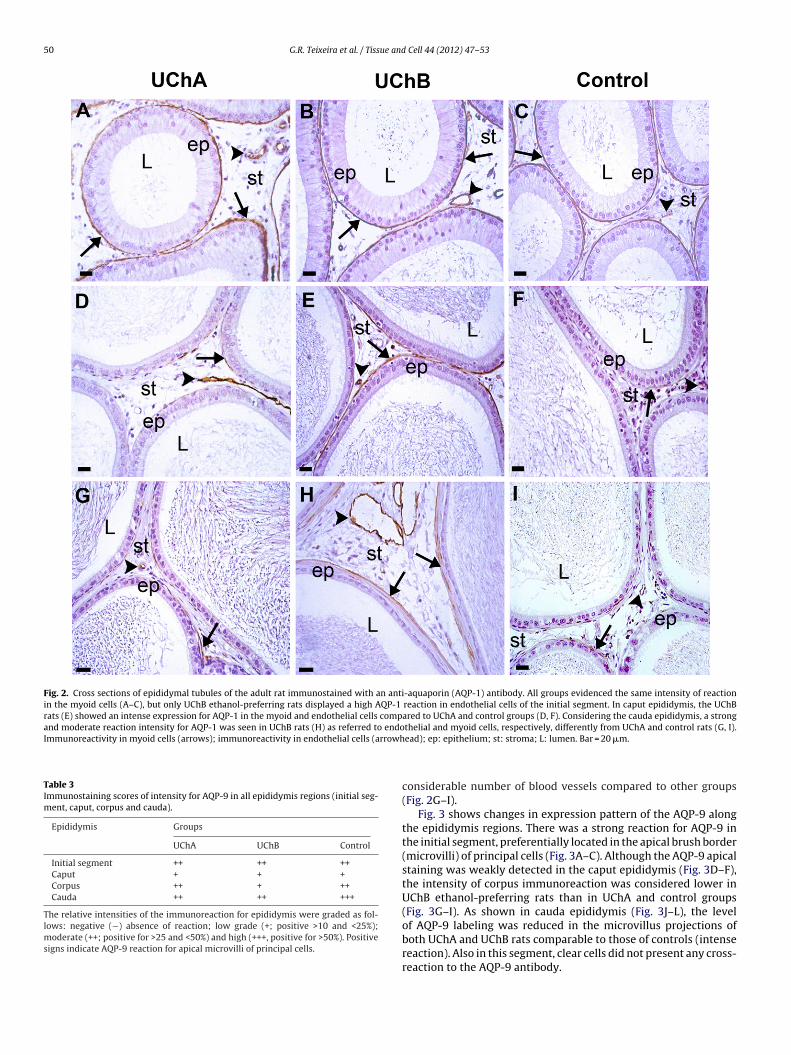
50 G.R. Teixeira et al. / Tissue and Cell 44 (2012) 47– 53
Fig. 2. Cross sections of epididymal tubules of the adult rat immunostained with an anti-aquaporin (AQP-1) antibody. All groups evidenced the same intensity of reactionin the myoid cells (A–C), but only UChB ethanol-preferring rats displayed a high AQP-1 reaction in endothelial cells of the initial segment. In caput epididymis, the UChBrats (E) showed an intense expression for AQP-1 in the myoid and endothelial cells compared to UChA and control groups (D, F). Considering the cauda epididymis, a strongand moderate reaction intensity for AQP-1 was seen in UChB rats (H) as referred to endothelial and myoid cells, respectively, differently from UChA and control rats (G, I).Immunoreactivity in myoid cells (arrows); immunoreactivity in endothelial cells (arrowh
Table 3Immunostaining scores of intensity for AQP-9 in all epididymis regions (initial seg-ment, caput, corpus and cauda).
Epididymis Groups
UChA UChB Control
Initial segment ++ ++ ++Caput + + +Corpus ++ + ++Cauda ++ ++ +++
The relative intensities of the immunoreaction for epididymis were graded as fol-lows: negative (−) absence of reaction; low grade (+; positive >10 and <25%);ms
oderate (++; positive for >25 and <50%) and high (+++, positive for >50%). Positiveigns indicate AQP-9 reaction for apical microvilli of principal cells.
ead); ep: epithelium; st: stroma; L: lumen. Bar = 20 �m.
considerable number of blood vessels compared to other groups(Fig. 2G–I).
Fig. 3 shows changes in expression pattern of the AQP-9 alongthe epididymis regions. There was a strong reaction for AQP-9 inthe initial segment, preferentially located in the apical brush border(microvilli) of principal cells (Fig. 3A–C). Although the AQP-9 apicalstaining was weakly detected in the caput epididymis (Fig. 3D–F),the intensity of corpus immunoreaction was considered lower inUChB ethanol-preferring rats than in UChA and control groups(Fig. 3G–I). As shown in cauda epididymis (Fig. 3J–L), the levelof AQP-9 labeling was reduced in the microvillus projections of
both UChA and UChB rats comparable to those of controls (intensereaction). Also in this segment, clear cells did not present any cross-reaction to the AQP-9 antibody.
G.R. Teixeira et al. / Tissue and Cell 44 (2012) 47– 53 51
Fig. 3. Cross sections of epididymal tubules of the adult rat immunostained with an anti-aquaporin (AQP-9) antibody. In initial segment, the reaction in the long apicalmicrovilli was similar at different groups (A–C). In caput epididymis, AQP-9 showed an extensive diffused reaction associated to microvilli (D–F). Immunoreactivity of corpuse trast tc ymis)(
4
t(aI
pididymis in the UChB ethanol-preferring rats (H) presented low intensity in cononsuming animals (J, K) showed a decrease in apical AQP-9 staining (cauda epididarrows); clear cell (arrowhead); ep: epithelium; st: stroma; L: lumen. Bar = 20 �m.
. Discussion
Ethanol is thought to be a toxicant affecting the body composi-
ion and nutritional status. In addition, the energy value of ethanolempty calories) did not contribute to body weight gain as wells the lipid synthesis is generally blocked (Ayene et al., 2002).n this study, ethanol-preferring rats showed reduced BW ando moderate intensity (G, I) over the entire epithelium toward the lumen. Ethanol- when compared to controls (L). Immunoreactivity in apical epithelium microvilli
epididymal weights compared to controls. There seem to be nodoubt that UCh male rats display a low BW gain (Cândido et al.,2007) and epididymal weight (Martinez et al., 2000). It was also
previously described that testosterone levels tends to drop with thechronic ethanol consumption (Montico et al., 2010; Cândido et al.,2007), and furthermore, every androgen-dependent reproductivetissue may have its morphophysiological function compromised.
5 ue and
ItaorTwUdtedifire1
sm2sttdam
wSatmiebtOtssrreewliptfldpAsawfci
pPpd2
2 G.R. Teixeira et al. / Tiss
t is widely accepted that a severe reduction of serum testos-erone levels in animals submitted to various alcohol doses isttributed to an indirect action on the LH surge through imbalancef the hypothalamus–pituitary–gonadal axis or directly by down-egulating LH receptors in leydig cells (Van Thiel and Lester, 1979;adic et al., 2000). The representative reduction in the epididymaleight (mainly in the caput segment) and epididymis tubules ofChB rats could be associated, at least in part, with testosteroneisturbances and other related factors. O’Hara et al. (2011) statedhat mice failing to express androgen receptor did not developpididymal initial segment and otherwise, the caput epididymisisplays a decrease in lumen diameter, thus proving that this organ
s androgen-responsive. These harmful consequences were con-rmed in our study. For this purpose, UChA and UChB rat lines bestepresent a model for studying the ethanol effects as those found inthanol-related human diseases (Mardones and Segovia-Riquelme,983).
Fluid and electrolyte transport processes in epithelial cellsignificantly modify the luminal microenvironment in which sper-atozoa acquire maturation and are retained (Pastor-Soler et al.,
002). Specific AQPs are expressed in the postnatal epididymal cellso that they can exert a transepithelial water movement acrosshe organ (Huang et al., 2006). Curiously, this study examined forhe first time the immunolocalization of AQP-1 and AQP-9 in epi-idymis of UChA and UChB ethanol-preferring rats, namely, lownd high ethanol consumption, thus correlating to the steroid hor-one levels.It has been reported that AQP-1 is abundantly expressed in
hole excurrent duct of several species (Fisher et al., 1998; Pastor-oler et al., 2001; Domeniconi et al., 2008). In accordance to Badrannd Hermo (2002) and Oliveira et al. (2005), AQP-1 was not found inhe epididymal epithelium, but instead, it was expressed in smooth
uscle as well as endothelial cells lining vascular channels situatedn the intertubular space of the epididymis. Interestingly, AQP-1xpression in myoid cells restricted to initial segment is inducedy orchidectomy and after anti-estrogen therapies or exposureso ethinyl oestradiol (Fisher et al., 1999; McKinnell et al., 2001;liveira et al., 2005). Animals consuming ethanol and presenting
he lowest circulating testosterone levels, preferentially UChB rats,howed more intensity for AQP-1 reaction in the peritubular tis-ue of the epididymis, similarly to those described in castratedats (Oliveira et al., 2005). Taking into account that androgens caneverse the effects of AQP-1 overexpression in myoid cells (Oliveirat al., 2005), our UCh rats could have a stimulatory effect on itsxpression due to the lack of testosterone. More recently, AQP-1as found to be ubiquitously expressed in tumor vascular endothe-
ium or tumor angiogenesis and it appears that their increasedmmunostaining is linked to the plasma membrane osmotic waterermeability (Verkman et al., 2008). These data argue for an impor-ant involvement of epididymal AQP-1 in regulating transcellularuid flow and tissue hydration considering the UChB ethanol-rinking rats. Additionally, our results pointed AQP-1 as a keylayer in the fluid reabsorption of UChB ethanol-preferring rats.
large amount of AQP-1 in the epididymal vascular network andtroma suggest that this water channel is an important crossing for
rapid fluid absorption, mainly during prolonged ethanol intake,hich in turn may accumulates more fluid, thus increasing the need
or absorption through epididymis. Moreover, there remain manyoncerns about the role of aquaporins into the endothelium-stromanterface.
AQP-9 has been earlier recognized in the microvilli of princi-al cells along the four epididymal regions (Elkjaer et al., 2000;
astor-Soler et al., 2001). A growing body of evidences strongly sup-orts AQP-9 as the first major aquaporin detected in epididymisisplaying apical and basolateral immunoreaction (Elkjaer et al.,000; Pastor-Soler et al., 2001, 2002; Badran and Hermo, 2002;Cell 44 (2012) 47– 53
Oliveira et al., 2005; Da Silva et al., 2006). AQP-9 is a transmem-brane protein allowing both selective neutral solute and water flow(Ishibashi et al., 1998; Tsukaguchi et al., 1999). In initial segment,a moderate intensity reaction was obtained for all groups, provingthat ethanol consumption has no damaging effect on this segment,despite the reduced testosterone levels. It has been proposed thatDHT but not testosterone and estrogens can lead to a decrease inAQP-9 expression at this segment in a dependent way (Oliveiraet al., 2005). In fact, there is a crucial step that allows sperm con-centration in the epididymal initial segment enabling the secretorycompounds of the principal cells to interact more efficiently withsperm surface (Robaire and Hermo, 1988; Cornwall et al., 2002;Hess et al., 2000). Following to androgen ablation, it was statedthat AQP-9 is absent from microvilli of principal cells in the initialsegment (Badran and Hermo, 2002), however, expression of AQP-9 in principal cells of the corpus and cauda epididymis was notaffected. Our findings related to AQP-9 immunostaining in caputregion were similar after chronic ethanol consumption, while itwas decreased over the corpus of UChB ethanol-preferring rats.More directly, ethanol per se has no significant effect on glyceroluptake through AQP-9 channels, whereas acetaldehyde seems tomodify AQP-9 by disrupting membrane permeability to enhanceboth glycerol influx and water transport (Potter et al., 2011). How-ever, the exact related mechanism(s) are still unknown. Regardingthe corpus, it was evidenced a distinct regulatory role not presum-ably linked to a fall in testosterone levels, since AQP-9 expression incorpus epididymis seems to be unresponsive to androgens (Hermoand Smith, in press). In cauda epididymis, both UChA and UChBrats exhibited lower expression of AQP-9 compared to controls.Thus, it would lead us to believe that the microvilli of principal cellsin this region are quite sensitive to testosterone and/or lumicrinefactor(s) as previously mentioned by Badran and Hermo (2002).Other study has also indicated that treatment with anti-androgenflutamide causes significant reductions in levels of AQP-9 in theadult epididymis, which can be restored in the cauda epididymisafter testosterone administration (Pastor-Soler et al., 2002). On theother hand, the 17-� estradiol levels were unchanged with ethanolconsumption, preventing any estradiol-induced alterations AQP-related. In our study, there was no immunoreaction regarding toclear cells in the cauda epididymis. In this region, immunolabel-ing of clear cells have been more inconsistent with some authorswho found intense AQP-9 reactions over the cytoplasm of such cells(Badran and Hermo, 2002; Hermo et al., 2008) while others havestated no reactivity to clear cells (Pastor-Soler et al., 2001). Thesereports are controversial and some points need to be clarified.
In summary, chronic ethanol consumption is able to causechanges in the pattern of AQPs expression throughout the epi-didymis. In the peritubular space, AQP-1 expression is enhancedwhen ethanol intake rises, acting as channel removing fluid andthereby keeping constant the water balance. Although AQP-1 hasbeen positively regulated by high ethanol consumption, AQP-9has differential regulation in corpus and cauda epididymis, beingseverely influenced by testosterone. Further studies with othersteroid hormones underlying the mechanisms related to ethanolconsumption via its specific receptors are greatly encouraged.
Financial support
FAPESP – São Paulo Research Foundation, Grant number:2005/04712-1.
Conflict of interest
All authors declare that there is no conflict of interest

ue and
A
bp0a
R
A
A
A
A
B
B
B
C
C
C
C
D
D
E
F
F
F
H
H
H
H
tion. Clin. Endocrinol. Metab. 8, 499–510.
G.R. Teixeira et al. / Tiss
cknowledgements
All authors express their grateful to Ms. Gelson Rodriguezy the excellent technical assistance. This work was also sup-orted by FAPESP – São Paulo Research Foundation, Process n.4/10661-5/05/04194-8 and by the National Council for Scientificnd Technological Development, Process n. 470297/2004-2.
eferences
gre, P., Brown, D., Nielsen, S., 1995. Aquaporin water channels: unanswered ques-tions and unresolved controversies. Curr. Opin. Cell Biol. 7, 472–483.
gre, P., Preston, G.M., Smith, B.L., Jung, J.S., Raina, S., Moon, C., Guggino, W.B.,Nielsen, S., 1993. Aquaporin CHIP: the archetypal molecular water channel. Am.J. Physiol. 34, 463–476.
nderson Jr., R.A., Willis, B.R., Oswald, C., Zaneveld, L.J., 1985. Partial reversal ofethanol-induced male reproductive pathology following abstinence. AlcoholAlcohol. 20, 273–286.
yene, I.S., Stamato, T.D., Mauldin, S.K., Biaglow, J.E., Tuttle, S.W., Jenkins, S.F., Koch,C.J., 2002. Mutation in the glucose-6-phosphate dehydrogenase gen leads toinactivation of ku DNA end binding during oxidative stress. J. Bio. Chem. 277,9929–9935.
adran, H.H., Hermo, L., 2002. Expression and regulation of aquaporins 1, 8, and 9in the testis, efferent ducts, and epididymis of adult rats and during postnataldevelopment. J. Androl. 23, 358–373.
eitz, E., Schultz, J.E., 1999. The mammalian aquaporin water channel family: apromising new drug target. Curr. Med. Chem. 6, 457–467.
rown, D., Verbavatz, J.M., Valenti, G., Lui, B., Sabolic, I., 1993. Localization of theCHIP28 water channel in reabsorptive segments of the rat male re-productivetract. Eur. J. Cell Biol. 61, 264–273.
ândido, E.M., Carvalho, C.A., Martinez, F.E., Cagnon, V.H.A., 2007. Experimental alco-holism and pathogenesis of prostatic diseases in UChB rats. Cell Biol. Int. 31,459–472.
huffa, L.G., Amorim, J.P., Teixeira, G.R., Mendes, L.O., Fioruci, B.A., Pinheiro, P.F.,Seiva, F.R., Novelli, E.L., de Mello Júnior, W., Martinez, M., Almeida-Francia,C.C., Martinez, F.E., 2011. Long-term exogenous melatonin treatment modu-lates overall feed efficiency and protects ovarian tissue against injuries causedby ethanol-induced oxidative stress in adult UChB rats. Alcohol. Clin. Exp. Res.35, 1498–1508.
huffa, L.G., Padovani, C.R., Martinez, F.E., 2009. Ovarian structure and hormonalstatus of the UChA and UChB adult rats in response to ethanol. Maturitas 62,21–29.
ornwall, G.A., Lareyre, J.J., Matusik, R.J., Hinton, B.T., Orgebin-Crist, M.C., 2002.Gene expression and epididymal function. In: Robaire, B., Hinton, B.T. (Eds.),The Epididymis: From Molecules to Clinical Practice. Kluwer Academic/PlenumPublishers, New York, NY, pp. 169–199.
a Silva, N., Silberstein, C., Beaulieu, V., Piétrement, C., Van Hoek, A.N., Brown, D.,Breton, S., 2006. Postnatal expression of aquaporins in epithelial cells of the ratepididymis. Biol. Reprod. 74, 427–438.
omeniconi, R.F., Orsi, A.M., Justulin, L.A., Leme Beu, C.C., Felisbino, S.L., 2008.Immunolocalization of aquaporins 1, 2 and 7 in rete testis, efferent ducts, epi-didymis and vas deferens of adult dog. Cell Tissue Res. 332, 329–335.
lkjaer, M., Vajda, Z., Nejsum, L.N., Kwon, T., Jensen, U.B., Amiry-Moghaddam, M.,Frokiaer, J., Nielsen, S., 2000. Immunolocalization of AQP9 in liver, epididymis,testis, spleen, and brain. Biochem. Biophys. Res. Commun. 276, 1118–1128.
isher, J.S., Turner, K.J., Brown, D., Sharpe, R.M., 1999. Effect of neonatal exposure toestrogenic compounds on development of the excurrent ducts of the rat testisthrough puberty to adulthood. Environ. Health Perspect. 107, 397–405.
isher, J.S., Turner, K.J., Fraser, H.M., Saunders, P.T., Brown, D., Sharpe, R.M., 1998.Immunoexpression of aquaporin-1 in the efferent ducts of the rat and marmosetmonkey during development, its modulation by estrogens, and its possible rolein fluid resorption. Endocrinology 139, 3935–3945.
rigeri, A., Nicchia, G.P., Verbavatz, J.M., Valenti, G., Svelto, M., 1998. Expression ofaquaporin-4 in fast-twitch fibers of mammalian skeletal muscle. J. Clin. Invest.102, 695–703.
ermo, L., Robaire, B., 2002. Epididymal cell types and their functions. In: Robaire, B.,Hinton, B.T. (Eds.), The Epididymis: From Molecules to Clinical Practice. KluwerAcademic/Plenum Publishers, New York, pp. 81–98.
ermo, L., Schellenberg, M., Liu, L.Y., Dayanandan, B., Zhang, T., Mandato, C.A., Smith,C.E., 2008. Membrane domain specificity in the spatial distribution of aquaporins5, 7, 9, and 11 in efferent ducts and epididymis of rats. J. Histochem. Cytochem.56, 1121–1135.
ess, R.A., Bunick, D., Lubahn, D.B., Zhou, Q., Bouma, J., 2000. Morphologic changes
in efferent ductules and epididymis in estrogen receptor-alpha knockout mice.J. Androl. 21, 107–121.uang, H.F., He, R.H., Sun, C.C., Zhang, Y., Meng, Q.X., Ma, Y.Y., 2006. Function ofaquaporins in female and male reproductive systems. Hum. Reprod. Update 12,785–795.
Cell 44 (2012) 47– 53 53
Ishibashi, K., Kuwahara, M., Gu, Y., Kageyama, Y., Tohsaka, A., Suzuki, F., Marumo,F., Sasaki, S., 1997. Cloning and functional expression of a new water channelabundantly expressed in the testis permeable to water, glycerol, and urea. J. Biol.Chem. 272, 20782–20786.
Ishibashi, K., Kuwahara, M., Gu, Y., Tanaka, Y., Marumo, F., Sasaki, S., 1998. Cloningand functional expression of a new aquaporin (AQP9) abundantly expressedin the peripheral leukocytes permeable to water and urea, but not to glycerol.Biochem. Biophys. Res. Commun. 244, 268–274.
King, L.S., Agre, P., 1996. Pathophysiology of the aquaporin water channels. Annu.Rev. Physiol. 58, 619–648.
Li, J., Nielsen, S., Dai, Y., Lazowski, K.W., Christensen, E.I., Tabak, L.A., Baum, B.J.,1994. Examination of rat salivary glands for the presence of the aquaporin CHIP.Pflugers Arch. 428, 455–460.
Hermo, L., Smith, C.E. Thirsty Business: Cell, Region and Membrane Specificity ofAquaporins (AQPs) in the Testis, Efferent Ducts and Epididymis and FactorsRegulating Their Expression. J. Androl. March 25, in press.
Ma, T., Yang, B., Gillespie, A., Carlson, E.J., Epstein, C.J., Verkman, A.S., 1998. Severelyimpaired urinary concentrating ability in transgenic mice lacking aquaporin-1water channels. J. Biol. Chem. 273, 4296–4299.
Mardones, J., Segovia-Riquelme, N., 1983. Thirty-two years of selection of rats byethanol preference: UChA and UChB strains. Neurobehav. Toxicol. Teratol. 5,171–178.
Martinez, F.E., Martinez, M., Padovani, C.R., 2000. Morphology of testis and epi-didymis in an ethanol-drinking rat strain (UChA and UChB). J. Submicrosc. Cytol.Pathol. 32, 175–184.
McKinnell, C., Atanassova, N., Williams, K., Fisher, J.S., Walker, M., Turner, K.J., Saun-ders, T.K., Sharpe, R.M., 2001. Suppression of androgen action and the inductionof gross abnormalities of the reproductive tract in male rats treated neonatallywith diethylstilbestrol. J. Androl. 22, 323–338.
Montico, F., Fávaro, W.J., Cândido, E.M., Martinez, M., Pinheiro, P.F., Martinez, F.E.,Cagnon, V.H., 2010. Alcoholism and coagulating gland: androgen and insulin likegrowth factor-1 receptor features. Tissue Cell 42, 203–210.
Nielsen, S., Smith, B.L., Christensen, E.I., Knepper, M.A., Agre, P., 1993. CHIP28water channels are localized in constitutively water-permeable segments of thenephron. J. Cell Biol. 120, 371–383.
O’Hara, L., Welsh, M., Saunders, P.T., Smith, L.B., 2011. Androgen receptor expressionin the caput epididymal epithelium is essential for development of the initialsegment and epididymal spermatozoa transit. Endocrinology 152, 718–729.
Oliveira, C.A., Carnes, K., Franca, L.R., Hermo, L., Hess, R.A., 2005. Aquaporin-1 and -9are differentially regulated by oestrogen in the efferent ductile epithelium andinitial segment of the epididymis. Biol. Cell. 97, 385–395.
Page, E., Winterfield, J., Goings, G., Bastawrous, A., Upshaw-Earley, J., 1998.Water channel proteins in rat cardiac myocyte caveolae: osmolarity-dependentreversible internalization. Am. J. Physiol. Heart Circ. Physiol. 274, 1988–2000.
Pastor-Soler, N., Bagnis, C., Sabolic, I., Tyszkowski, R., McKee, M., Van Hoek, A., Breton,S., Brown, D., 2001. Aquaporin 9 expression along the male reproductive tract.Biol. Reprod. 65, 384–393.
Pastor-Soler, N., Isnard-Bagnis, C., Herak-Kramberger, C., Sabolic, I., Van Hoek, A.,Brown, D., Breton, S., 2002. Expression of aquaporin 9 in the adult rat epididymalepithelium is modulated by androgens. Biol. Reprod. 66, 1716–1722.
Potter, J.J., Koteish, A., Hamilton, J., Liu, X., Liu, K., Agre, P., Mezey, E., 2011. Effects ofacetaldehyde on hepatocyte glycerol uptake and cell size: implication of aqua-porin 9. Alcohol. Clin. Exp. Res. 35, 939–945.
Robaire, B., Hinton, B.T., Orgebin-Crist, M.C., 2006. The epididymis. In: Knobil, E.,Neill, J. (Eds.), Physiology of Reproduction. Elsevier, New York, pp. 1071–1148.
Robaire, B., Hermo, L., 1988. Efferent ducts, epididymis, and vas deferens: structure,functions and their regulation. In: Knobil, E., Neill, J.D. (Eds.), The physiology ofreproduction. Raven Press, New York, pp. 999–1080.
Robaire, B., Viger, R.S., 1995. Regulation of epididymal epithelial cell functions. Biol.Reprod. 52, 226–236.
Shanahan, C.M., Connolly, D.L., Tyson, K.L., Cary, N.R., Osbourn, J.K., Agre, P., Weiss-berg, P.L., 1999. Aquaporin-1 is expressed by vascular smooth muscle cells andmediates rapid water transport across vascular cell membranes. J. Vasc. Res. 36,353–362.
Tadic, S.D., Elm, M.S., Subbotin, V.M., Eagon, P.K., 2000. Hypogonadism precedes liverfeminization in chronic alcohol-fed male rats. Hepatology 31, 1135–1140.
Tsukaguchi, H., Shayakul, C., Berger, U.V., Mackenzie, B., Devidas, S., Guggino, W.B.,Van Hoek, A.N., Hediger, M.A., 1998. Molecular characterization of a broad selec-tivity neutral solute channel. J. Biol. Chem. 273, 24737–24743.
Tsukaguchi, H., Weremowicz, S., Morton, C.C., Hediger, M.A., 1999. Functional andmolecular characterization of the human neutral solute channel aquaporin-9.Am. J. Physiol. 277, 685–696.
Turner, T.T., 1984. Resorption versus secretion in the rat epididymis. J. Reprod. Fertil.72, 509–514.
Van Thiel, D.H., Lester, R., 1979. The effect of chronic alcohol abuse on sexual func-
Verkman, A.S., Hara-Chikuma, M., Marios, C., 2008. Papadopoulos Aquaporins—newplayers in cancer biology. J. Mol. Med. 86, 523–529.
Wintour, E.M., 1997. Water channels and urea transporters. Clin. Exp. Pharmacol.Physiol. 24, 1–9.
Copyright © 2022 FDOKUMEN




















