
In-vitromodellering van de linkerhelft van het hart In-Vitro ...
Upload
independentCategory
view
2download
0
Vleermuizenonderzoek in de
omgeving van het kasteeldomein
Klaverblad en de vallei van de
Kleine Struisbeek te Wilrijk
Johan De Ridder
Daniel Sanders
Luc De Bruyn
2013

Auteurs: Johan De Ridder, Daniel Sanders en Luc De Bruyn
Contact: Natuurpunt Zuidrand Antwerpen Frans De Cortlaan 42 2610 Wilrijk Afgiftekantoor Wilrijk 1 www.zuidrand.be
e-mail: [email protected]
Wijze van citeren:
De Ridder, J., Sanders, D. & De Bruyn, L. (2013). Vleermuizenonderzoek in de omgeving van het kasteeldomein Klaverblad en de vallei van de Kleine Struisbeek te Wilrijk. Rapport Natuurpunt Zuidrand Antwerpen werkgroep Studie 2013/1.
Verantwoordelijke uitgever: Guido Callaerts
Foto cover: Johan De Ridder
Deze studie kwam tot stand met financiële ondersteuning van de Provincie Antwerpen en ANKONA. INBO en Universiteit Antwerpen gaven ondersteuning bij de statistische verwerking.
Provincie Antwerpen Dienst Duurzaam Milieu- en Natuurbeleid
Team Natuur & Landschap, Cel Beleid K. Elisabethlei 22,2018 Antwerpen
T 03 240 66 87

Vleermuizenonderzoek in de omgeving
van het kasteeldomein Klaverblad en de
vallei van de Kleine Struisbeek te Wilrijk
Coördinatie: Johan De Ridder
Veldwerk en geluidanalyses: Johan De Ridder en Daniel Sanders
Statistische analyses: Luc De Bruyn (INBO; Universiteit Antwerpen)
Tekst: Johan De Ridder, Daniel Sanders en Luc De Bruyn
Eindredactie en vormgeving: Johan De Ridder
Foto’s: Johan De Ridder
Met dank aan Wout Willems voor het ter beschikking stellen van rapporten en wintertelgegevens, Michel Barataud voor controle van geluidsanalyses.

Pagina | 1
Inhoudsopgave
Inhoudsopgave ....................................................................................................... 1
1. Inleiding .......................................................................................................... 2
2. Situering ......................................................................................................... 3
3. Onderzoeksvragen en te verwachten resultaten ....................................................... 5
4. Materiaal en methoden ....................................................................................... 5
4.1. Telmethode in lente en zomer ........................................................................ 5
4.2. Wintertelling ............................................................................................... 7
4.3. Soortdeterminatie aan de hand van ultrasone geluiden ........................................ 7
4.4. Verwerking en statistische analyse .................................................................. 8
5. Resultaten ....................................................................................................... 9
5.1. Algemeen .................................................................................................. 9
5.2. Vlieg- en jachtactiviteit gewone dwergvleermuis .............................................. 11
5.3. Vlieg- en jachtactiviteit van vleermuizen in de omgeving van het domein Klaverblad 16
5.4. Vlieg- en jachtactiviteit van vleermuizen langs de Kleine Struisbeek .................... 17
5.5. Wintertelling Klaverblad .............................................................................. 18
6. Bespreking..................................................................................................... 19
6.1. Vliegactiviteit en jachtactiviteit ..................................................................... 19
6.2. Landschapsgebruik Klaverblad...................................................................... 25
6.3. Landschapsgebruik Kleine Struisbeek............................................................. 27
7. Aanbevelingen voor verder onderzoek ................................................................. 29
8. Aanbevelingen voor beheer ............................................................................... 30
8.1. Voorzorgsmaatregelen ................................................................................ 30
8.2. Beheermaatregelen voor de omgeving van het kasteeldomein Klaverblad .............. 30
8.3. Beheermaatregelen voor de omgeving van de Kleine Struisbeek .......................... 33
9. Literatuurlijst.................................................................................................. 35
10. Bijlagen......................................................................................................... 40
10.1. Overzicht van tel-data, tijdstippen en weersomstandigheden ........................... 40
10.2. Analyse geluidsopnamen .......................................................................... 44
10.3. Kaarten ................................................................................................ 49
10.4. Beeldmateriaal ....................................................................................... 88

Pagina | 2
1. Inleiding
De Europese vleermuizen zijn insectenetende zoogdieren met een complexe levenswijze. Ze stellen hoge eisen aan overwinteringsplaatsen, zomerverblijfplaatsen en jachtgebieden. Hierdoor zijn ze bedreigd in een steeds grootschaliger en verstedelijkt cultuurlandschap (Wickramasinghe et al., 2003; Dietz et al., 2010). De belangrijkste bedreigingen in Vlaanderen zijn gebruik van bestrijdingsmiddelen, vernietiging van leefomgeving en verstoring tijdens belangrijke fasen in de levenscyclus (Verkem, 1998; Paelinckx et al., 2009). Vleermuispopulaties zijn vaak gelimiteerd door de aanwezigheid van jachtmogelijkheden. De jachtactiviteit van vleermuizen is sterk afhankelijk van de vliegactiviteit van insecten (Rydell et al., 1996; Kusch et al., 2004; Cel’uch & Kaňuch, 2004; Lee, Y. & McCracken, 2004). Grote wijzigingen of verstoringen in jachtgebieden die een veranderde insectendensiteit met zich meebrengen kunnen populatieaantallen rechtstreeks beïnvloeden (Vaughan et al., 1997). Kennis en bescherming van jachthabitat is daarom zeer belangrijk voor de bescherming van vleermuizen. In Vlaanderen leven momenteel 17 soorten vleermuizen van de 23 die voorkomen in België (Verkem, 1988; Paelinckx et al., 2009). Het merendeel van deze soorten is opgenomen in de rode lijst van zoogdieren voor Vlaanderen (Criel et al., 1994). Voor Brandts/baardvleermuis, watervleermuis, laatvlieger, ingekorven vleermuis en franjestaart wordt een vooruitgang in Vlaanderen aangegeven. Van de andere soorten is de trend onbekend, of blijft de situatie stabiel. Dit wil nog niet zeggen dat deze soorten effectief veiliggesteld zijn (NARA 2005, Demolder et al., 2012). De bescherming van vleermuizen krijgt daarom zowel op internationaal als nationaal niveau meer en meer aandacht. Alle in België voorkomende soorten vleermuizen zijn beschermd door de conventie van Bern en Bonn. Vleermuizen zijn opgenomen in de bijlage II of IV van de habitatrichtlijn. Vleermuizen zijn ook prioritaire soorten voor de provincie Antwerpen (Dienst Duurzaam Milieu- en Natuurbeleid, 2010).
Het is bekend dat de Antwerpse forten voor de Vlaamse vleermuispopulaties tijdens de winter een belangrijke rol spelen. Er overwinteren jaarlijks 12 soorten vleermuizen in de forten. Voor zeven soorten zijn de forten ronduit cruciaal voor hun voortbestaan in Vlaanderen (Verkem, 2008; Paelinckx et al., 2009; Verkem, 2009; Boers et al., 2010; Boers et al., 2013). Een aantal forten werd daarom opgenomen binnen habitatrichtlijngebied. Vlaanderen maakte gewestelijke instandhoudingsdoelstellingen op voor de vleermuissoorten die voorkomen in Vlaanderen (Adriaens et al., 2008; Paelinckx et al., 2009). Voor het gebied “BE2100045 Historische Fortengordels van Antwerpen als vleermuizenhabitat” werden specifieke instandhouding-doelstellingen opgesteld. De voorgestelde maatregelen dienen gerealiseerd te worden binnen de Speciale Beschermingszone (SBZ) zelf, terwijl de vleermuizen eveneens behoefte hebben aan een (na)zomerhabitat, verbindingen tussen zomer- en winterhabitat, en verbindingen tussen de verschillende forten onderling. Door de nadruk te leggen op winterhabitat bestaat er het risico dat op termijn de forten ontwikkelen tot kwalitatief hoogstaand winterhabitat, waar de aantallen vleermuizen in dalende lijn gaan omdat ze als geïsoleerde eilanden in een vleermuisonvriendelijk omgeving liggen. Er vindt immers jaarlijks migratie plaats van zomerverblijfplaats naar winterverblijfplaats. De afstanden die hierbij worden afgelegd bedragen bij de meeste soorten vaak niet meer dan enkele tientallen kilometers (Lange et al., 1994; Limpens et al., 1997; Twisk et al., 2010; Dietz et al., 2011). Andere soorten zoals ruige dwergvleermuis en rosse vleermuis vertonen trekgedrag en seizoensmigratie. Vleermuizen leggen dagelijks ook een afstand af tussen hun dagverblijfplaats en jachtgebieden. De afstand is afhankelijk van de soort en van de kwaliteit van het jachthabitat. Vleermuizen jagen tot op een afstand van 500 meter tot 5 kilometer van de

Pagina | 3
zomerverblijfplaats (Lange et al., 1994; Limpens et al., 1997; Twisk et al., 2010; Dietz et al., 2011). Momenteel is er echter weinig geweten over de forten als zomerhabitat voor vleermuizen, en nog minder over de tussenliggende jachtgebieden en zomerverblijfplaatsen. Er zijn enkele onderzoeken uitgevoerd naar vleermuizenactiviteit in de omgeving van forten van de buitengordel (Lefevre en Boeckx in Nieuwborgh, 2001; Thomaes & Hofman, 2009; Boers et al., 2012). Verbeylen et al. (2010) onderzochten foerageergebieden, vliegroutes, kraamkolonies en zomerverblijfplaatsen van vleermuizen in Fort 8 en omgeving. Willems et al. (2011) onderzochten zwermgedrag in Fort 8. Via een gerichte studie van potentieel zomerhabitat gelegen tussen enkele forten van de Brialmontfortengordel, willen we een leemte in kennis gedeeltelijk opvangen.
2. Situering
Deze studie onderzoekt de samenstelling van de vleermuizenpopulatie in twee open ruimte gebieden tussen de forten 6, 7 en 8: het kasteeldomein Klaverblad en de vallei van de Kleine Struisbeek. Het studiegebied situeert zich hoofdzakelijk op het grondgebied van de stad Antwerpen in het district Wilrijk op de grens met de gemeenten Aartselaar, Edegem en Hemiksem. Fort 6, 7 en 8 maken deel uit van de Brialmontfortengordel rond Antwerpen. In de 3 forten werden de afgelopen 10 jaar in totaal 10 soorten vleermuizen waargenomen tijdens wintertellingen. Watervleermuis (Myotis daubentonii), baard- en Brandtvleermuis (Myotis
mystacinus; Myotis brandtii) en gewone dwergvleermuis (Pipistrellus pipistrellus) overwinteren in de grootste aantallen. Laatvlieger (Eptesicus serotinus), rosse vleermuis (Nyctalus noctula), ruige dwergvleermuis (Pipistrellus nathusii), gewone grootoorvleermuis (Plecotus auritus), franjestaart (Myotis nattereri), ingekorven vleermuis (Myotis emarginatus) en meervleermuis (Myotis dasycneme) worden sporadisch tot zelden waargenomen (Baetens et al., 2003; Verkem, 2008; Beers et al., 2009; Verkem, 2009; Boers et al., 2010; Verbeylen G. & Lefevre
A., 2010.Willems W., Verbeylen G. & Lefevre A., 2011, Waeterschoot, H., mondelinge mededeling, 2013). De wintertellingen gebeuren door de Vleermuizenwerkgroep van Natuurpunt. Zwermgedrag van o.a. laatvlieger en gewone grootoorvleermuis werd waargenomen in Fort 6 (Ben Vanderwijden, mondelinge mededeling). In Fort 7 werd foerageergedrag waargenomen van watervleermuis, baardvleermuis, gewone dwergvleermuis, laatvlieger, rosse vleermuis en zwermgedrag van grootoorvleermuis, laatvlieger, gewone dwergvleermuis en baardvleermuis (Baetens et al., 2003; Beers et al., 2009; De Ridder, J. & Sanders, D., 2014, in voorbereiding). Fort 7 is een erkend natuurreservaat en opgenomen in het habitatrichtlijngebied. Fort 7 moet volgens de instandhoudingsdoelstellingen ontwikkelen naar een kwaliteitsvolle overwinterings- en zwermplaats voor watervleermuis, franjestaart, baard– en Brandts vleermuis. In Fort 8 wordt jachtgedrag waargenomen van gewone dwergvleermuis, laatvlieger, watervleermuis, rosse vleermuis en zwermgedrag van gewone grootoorvleermuis, watervleermuis, baardvleermuis en gewone dwergvleermuis (Johan De Ridder, persoonlijke observaties; Dekeukeleire D., Janssen R., Boers K. & W. Willems, 2011; Willems W., Verbeylen G. & Lefevre A., 2011).
Het Kasteel Klaverblad is een van de laatste hoven van plaisantie in Wilrijk en is nog bewoond door de adellijke familie Moretus. De eerste vermelding dateert uit 1577 (Prims, 1952). Het kasteeldomein is gelegen in een sterk landelijk gebied met afwisseling van weilanden, akkers en bossen. Er zijn heel wat dreven waarvan de Klaverbladdreef ten noordoosten van het kasteel de meest bekende is. Het kasteeldomein is niet vrij toegankelijk. Daarom werd voornamelijk de omgeving onderzocht. In de omgeving van het kasteel bevindt zich het oorsprongsgebied van de Hollebeek. De Hollebeek vormt hier de grens tussen het district

Pagina | 4
Wilrijk en Hoboken. De weilanden ten noorden van de Moerelei worden gekenmerkt door een toenemend hobbygebruik door paardenliefhebbers. De overige percelen zijn in (al dan niet extensief) landbouwgebruik. Het gebied wordt gekenmerkt door de aanwezigheid van kleine landschapselementen zoals knotbomen, populierenrijen, opgaande wilgen en houtkanten. Door het afwisselend landgebruik (bos, weiland, akkerland, dreven en waterpartijen) wordt het gebied ingeschat als potentieel waardevol zomer- en paarverblijfplaats voor vleermuizen. Er zijn nauwelijks vleermuizengegevens van het gebied bekend.
In de omgeving van het domein Klaverblad liggen een aantal waardevolle cultuurhistorische relicten. Nabij het kasteel Klaverblad, aan de Reigershoek, ligt de commandobunker van het voormalige schijnvliegveld in Wilrijk. Het is de enige constructie van het schijnvliegveld die bewaard is gebleven. De bunker is, samen met een bijna identieke bunker in Deurne, uniek in Europa. Het vliegveld van Wilrijk werd rond 1939 aangelegd in het kader van de Algemene Mobilisatie als één van de vele uitwijkvliegvelden. Het Duitse leger richtte dit vliegveld in als een schijnvliegveld, om verkenningsvliegtuigen te misleiden. Het werd "Flugplatz Hemiksem" genoemd (Philippart, 2011). De schansen 16 en 17 werden aangelegd rond 1907 en maakten onderdeel uit van de veiligheidsomwalling rond Antwerpen. De schansen bestaan uit een betonnen traditore batterij overdekt met een aarden wal en een gevechtskoepel. De schansen zijn omringd met een gracht (Gils, 1998). Schans 16 ligt op openbaar domein. Schans 16 was tijdens het onderzoek vrij toegankelijk. De gracht staat in de zomer droog tot halfdroog, maar de contouren zijn dan herkenbaar in het landschap. Het glacis is vergraven en wordt gebruikt als (motor)crossterrein. Het glacis is gedeeltelijk begroeid met opgaand hout en wilgenstruwelen. De traditore batterij is relatief droog en in vrij goede staat. Sommige lokalen vertonen sporen van vochtinsijpeling. De schietgaten zijn nog deels open, net zoals de toegangen. De betonnen onderbouw van de koepel is aanwezig, maar werd zwaar beschadigd bij de verwijdering van de koepel door Duitse militairen. Ten zuiden van de schans ligt het ‘Mastenbos’ het enige naaldbos in Wilrijk. Schans 17 is ontoegankelijk en particuliere eigendom. Het wordt gebruikt als kleinveestal.
Het stroomgebied van de Grote Struisbeek en de Benedenvliet is onderwerp van een speerpuntgebied in het kader van het stroomgebiedbeheerplan voor de Schelde. Langsheen de Kleine Struisbeek werd in kader hiervan een signaalgebied voor potentiele waterberging aangeduid. De Kleine Struisbeek is een waterloop van 2e categorie. De beek vormt de grens tussen het district Wilrijk en de gemeente Edegem. De beekvallei werd door de provincie Antwerpen aangeduid als potentiële provinciale natuurverbinding. Langs het gebied loopt een recreatief fietspad, het Geitenpad. Tijdens de studie werd het segment tussen de Kleine Doornstraat en de Edegemsesteenweg – Wilrijkstraat onderzocht. Het omvat een poelenrijk parkachtig gebied waar kamsalamander wordt teruggevonden (Baetens et al., 2007), en delen van de campus Drie Eiken van de Universiteit Antwerpen. Het segment tussen de Kleine Doornstraat en Doornstraat wordt gekenmerkt door een oud loofbosrelict, poelen, houtkanten en extensieve graslanden en akkers. Het bosgebied Mariënborgh op het grondgebied Edegem maakt onderdeel uit van een voormalig hof van plaisantie. Het is een oud eiken- en beukenbos met voorjaarsflora-relicten. Er lopen enkele monumentale oude beukendreven doorheen het gebied. De universiteitscampus is relatief open met een afwisseling van gebouwen, parkings, wandelwegen, weiden, kleine bossen en vijvers. De vijvers zijn via een grachtenstelsel met elkaar verbonden. De Kleine Struisbeek loopt doorheen de campus. In de onmiddellijke omgeving van de Vallei van de Kleine Struisbeek werden in het gebied “Groen Neerland” tijdens een inventarisatieronde in 2007 gewone dwergvleermuis, ruige dwergvleermuis en laatvlieger waargenomen (Baetens et al., 2007).

Pagina | 5
3. Onderzoeksvragen en te verwachten resultaten
Welke soorten vleermuizen komen voor in de omgeving van het kasteeldomein Klaverblad en langs de loop van de Kleine Struisbeek? Zijn er verschillen tussen de gebieden in het voorkomen van soorten? Welke functies voor vleermuizen kunnen toegekend worden aan de gebieden? Hoe verloopt de activiteit van vleermuizen over het seizoen?
Het antwoord op deze vragen wordt onderzocht aan de hand van het meten van vlieg- en jachtactiviteit van vleermuizen op vaste meetpunten in 2011 en 2012. De tellingen en soortdeterminatie gebeurd met behulp van batdetectoren. Het belang van Schans 16 als winterverblijfplaats wordt ingeschat door de uitvoering van wintertellingen
De inventarisatie wil een verkennend beeld geven van de verspreiding en soortenrijkdom van vleermuizen in het onderzoeksgebied. Uit de vaststellingen op terrein en de verzamelde gegevens worden conclusies getrokken over de verschillende vleermuizenfuncties in het gebied. Er wordt getracht om o.a. vliegroutes en jachtgebieden van vleermuizen in kaart te brengen. De rapportering wordt afgesloten met het formuleren van bijkomende of nieuwe onderzoeksvragen, en aanbevelingen voor het beleid en het beheer van de onderzoeksgebieden in functie van behoud en versterken van de vleermuizenpopulaties.
4. Materiaal en methoden
4.1. Telmethode in lente en zomer
De inventarisatiemethode werd uitgewerkt volgens de basisprincipes van het vleermuisprotocol van de Gegevensautoriteit Natuur (2009; 2012). Het vleermuisprotocol bestaat uit een tabel met veldcondities en werkwijzen per vleermuissoort en per gebiedsfunctie, op grond waarvan de aanwezigheid van soorten en gebiedsfuncties met voldoende zekerheid kunnen worden vastgesteld. De goede praktijken van de Bat Conservation Trust (UK, 2007) en de werkwijze van Barataud (Barataud, 2012) werden in de mate van het mogelijke gevolgd.
Langs het kasteeldomein Klaverblad werd een transect met een lengte van 4,5 km uitgestippeld. Langs de vallei van de Kleine Struisbeek werd een transect met een lengte van 2,1 km uitgestippeld. Er werden respectievelijk 22 en 15 meetpunten langs de transecten aangeduid (kaart 1). De meetpunten lagen langs toegankelijke wegen. De meetpunten lagen minstens 50 meter uit elkaar. Elk transect kon op 1 avond worden geïnventariseerd en werd telkens in dezelfde richting gelopen.
In 2011 werd een verkennende inventarisatieronde uitgevoerd. Hoofddoel was een eerste inzicht krijgen in de soortensamenstelling van de gebieden, en de inventarisatiemethodiek in te oefenen. Het feitelijke veldwerk werd uitgevoerd tussen mei en oktober 2012, zodat de verzamelde gegevens betrekking hebben op een zo breed mogelijk aspect van de seizoenactiviteit van de vleermuizen (met uitzondering van de winterslaap). De vlieg- en jachtactiviteit van vleermuizen is immers onderhevig aan seizoensgebonden variatie (Encarnação et al., 2006; Ciechanowski et al., 2010). Tijdens de studie werd vlieg- en jachtactiviteit bepaald in trek-, balts- en paartijd en kraamperiode (figuur 1).

Pagina | 6
Figuur 1. Seizoencyclus vleermuizen
Er werd van meetpunt naar meetpunt gewandeld. Per meetpunt werden gedurende 3 minuten in tijdsintervallen van 5 seconden genoteerd of er vliegactiviteit was en van welke soort dit was. Elk ononderbroken signaal werd geteld als 1 passage. Indien een signaal langer dan 5 seconden aanhield, werd per bijkomende tijdsperiode van 5 seconden 1 extra passage geteld (Barataud, 2012). Met batdetectors kunnen passages geteld worden, maar geen aantallen of individuen (Flaquer et al, 2007). Het aantal passages is indicatief voor zowel vleermuisdensiteit als individuele foerageeractiviteit (Verboom en Huitema, 1997; Wickramasinghe et al., 2003). Aantallen werden enkel geteld bij visuele waarneming van de vleermuizen. Bijkomend werden, indien mogelijk, gegevens over gedrag verzameld (foerageren, zwermen, baltsactiviteiten aan de hand van sociaal geluid). Per meetpunt werd het aantal vangstmomenten geteld op dezelfde manier als vliegactiviteit. De vangstmomenten werden uitsluitend vastgesteld door een hoorbare vangstbuzz op de heterodyne batdetector. De aan- of afwezigheid van sociale geluiden op een meetpunt werd genoteerd, dus tijdens de 3 minuten waren er sociale geluiden of niet. Bij de verplaatsing tussen meetpunten werden van de dan waargenomen vleermuizen de soort en eventuele gedragsaanduidingen genoteerd op kaart. De veldnotities werden genoteerd op een inventarisatiefiche. De tellingen werden uitgevoerd door minstens twee personen, omwille van veiligheidsredenen en voor controle van de determinaties. Er werd steeds getracht om minimaal 20 dagen tussen elk opeenvolgend bezoek aan een gebied te laten. De tellingen werden uitgevoerd in goede weeromstandigheden. De starttemperatuur was voor alle tellingen, met uitzondering van de telling van 26 oktober, hoger dan 10°C (Bat Conservation Trust, 2007; Gegevensautoriteit Natuur, 2012). Alle tellingen, met uitzondering van de telling op 25 augustus en 26 september, werden gestart binnen het uur na de officiële zonsondergang om een zo representatief mogelijk beeld van de activiteit te krijgen. De vliegactiviteit van vleermuizen is gerelateerd tot de vliegactiviteit van insecten (Cel’uch & Kaňuch, 2004). Vleermuizenactiviteit en de beschikbaarheid van vliegende insecten die dienen als potentiele prooi volgen immers een bimodale curve met maxima tijdens de avond- en ochtendschemering (Kusch, 2005). Vleermuizen zoals gewone dwergvleermuis, die jachtvlucht (aerial hawking) als jachtstrategie gebruiken zijn bv. afhankelijk van de avondpiek van beschikbare insecten. De jachtactiviteit van deze soorten neemt af later op de avond (Kapfer & Aron, 2007). Kapfer et al. (2005) toonden aan dat de vliegactiviteit van gewone dwergvleermuis in Brussel sterk afnam afhankelijk van het tijdstip na zonsondergang. De vliegactiviteit van watervleermuis en ruige dwergvleermuis bleef echter gelijk. Kusch en Idelberger (2005) kwamen tot vergelijkbare conclusies bij een onderzoek in Duitsland.
J F M A M J J A S O N D
activiteit migratie
functie
tussen-
verblijf
kraamtijd
kraamverblijfplaats/
zomerverblijfplaats
winterslaap
winterverblijf
plaats
winterslaap
winterverblijf
plaats
paartijd/migratie
paarverblijfplaats

Pagina | 7
4.2. Wintertelling
Het opvolgen van soorten en aantallen vleermuizen in Schans 16 gebeurde door visuele inspectie. Er werd gebruik gemaakt van een goede zaklamp en een spiegeltje om in moeilijke holten en spleten te zoeken (Dijkstra en Korsten, 2005). De te onderzoeken locatie werd 1 maal per winter onderzocht. De beste periode voor het tellen van vleermuizen is van 15/12 tot 01/03. Waarnemingen buiten deze periode blijken in de praktijk niet goed vergelijkbaar (Willems et al, 2009).
4.3. Soortdeterminatie aan de hand van ultrasone geluiden
De activiteitstellingen werden uitgevoerd met twee heterodyne batdetectoren Pettersson D100 (Pettersson Elektronics AB, Uppsala, Zweden). De Pettersson D100 werd afgesteld op 20 kHz en 45 kHZ. Vleermuizen werden zoveel als mogelijk op het terrein gedetermineerd met één of twee time-expansie batdetector Pettersson D240x (Pettersson Elektronics AB, Uppsala, Zweden). De Pettersson D240X werd ingesteld op het heterodyne kanaal op 35 kHz. Gedurende de telling en bij waarneming van vleermuizenactiviteit, werd met de D240x over een brede range van frequenties gescand om de nulfrequentie te achterhalen. Werken met de Pettersson D240x maakt het mogelijk geluiden op te nemen en vertraagd terug af te spelen zodat determinaties op het terrein correcter kunnen gebeuren (o.a. door te luisteren naar piekfrequentie en bandbreedte). De opnames werden gemaakt in .wav-formaat met een Roland R05 Edirol. De geluidsopnames werden gedigitaliseerd en geanalyseerd met Batsound 4 (Pettersson Elektronik AB, Zweden). Bij twijfelgevallen werd enkel gedetermineerd tot op genus-niveau. Van de signalen werd piekfrequentie (FME), initiële frequentie (FI), eindfrequentie (FT) en pulslengte (ms) bepaald van een tiental representatieve signalen. De soorten werden gedetermineerd op basis van referentiewerken en -geluiden (Barataud, 2012; Russ, 2012; Van De Sijpe, 1999) en determinatiegrafieken (Barataud, 2012). Het is tot op zekere hoogte mogelijk om vleermuizen te determineren aan de hand van ultrasone geluiden, zeker indien er een combinatie van heterodyne en time-expansie technieken wordt gebruikt. Visuele waarnemingen (grootte, kleur, gedrag) kunnen extra informatie bieden om tot determinatie te komen (Ahlén en Baagøe, 1999). Voor de analyse van de opnames en de soortdeterminatie werd teruggevallen op de methode beschreven door Barataud (2012). De vleermuizen van het genus Myotis lijken sterk op elkaar, zowel uiterlijk als wat ultrasone geluiden betreft. De Myotis–geluiden werden in deze studie meestal slechts (gedetermineerd en) verwerkt op genus-niveau. Op basis van de geluidsopnames werd toch getracht te determineren op soortniveau.
Uit onderzoek in Spanje en de Verenigde Staten bleek dat de resultaten van inventarisaties aan de hand van ultrasone geluiden een goede correlatie opleveren met de gekende soorten in een gebied. Er werden over het algemeen meer soorten gevonden dan bij het gebruik van vangsttechnieken met mistnetten (O’Farrel & Gannon, 1999; Flaquer et al., 2007). Het aantal contacten per meetpunt was eveneens hoger bij inventarisaties met batdetectors dan met vangststechnieken (Flaquer et al., 2007). Er zijn echter ook nadelen verbonden aan soortdeterminatie met ultrasone methoden. Afhankelijk van de gebruikte methode werden soorten over- of ondergeïnventariseerd. Dit heeft onder andere te maken met het landschapsgebruik, vlieghoogte en roepsterkte van vleermuizen en de kenmerken en positionering van de batdetector (O’Farrel & Gannon, 1999; Brigham et al., 2004; Flaquer et al., 2007).

Pagina | 8
4.4. Verwerking en statistische analyse
In deze studie werd vliegactiviteit gedefinieerd als het aantal tijdsintervallen met passages per tijdseenheid (i.e. gedurende de 3 minuten waarnemingstijd). Bij de verwerking werd geen onderscheid gemaakt tussen aantallen passages afkomstig van 1 vleermuis of van meerdere vleermuizen. Vleermuizen zijn zeer opportunistisch in hun jachtgedrag. De foerageeractiviteit van vleermuizen hangt af van de verschillende jachtstrategieën van vleermuizensoorten en van de beschikbaarheid van insecten. Als de activiteit van insecten toeneemt, neemt de vliegactiviteit van vleermuizen toe (Rydell et al., 1996; Kusch et al., 2004; Cel’uch & Kaňuch, 2004; Lee, Y. & McCracken, 2004). Jachtactiviteit werd eveneens gedefinieerd als het aantal tijdsintervallen met vangstmomenten per tijdseenheid (i.e. gedurende de 3 minuten waarnemingstijd). Vleermuisactiviteit wordt beïnvloed door de reproductieve status van de vleermuizen. De verzamelde gegevens werden daarom voor de bespreking onderverdeeld in 3 perioden: april – tot midden juni, midden juni tot einde juli en augustus tot oktober. Deze perioden komen overeen met de onderscheiden fasen in de voorplantingscyclus van de meeste algemene vleermuissoorten in Europa: drachtperiode of kraamtijd, lactatiefase en post-lactatiefase. In deze studie werden de begrippen vliegroute en jachtgebied gehanteerd. Een vliegroute is een vaste route van een vleermuis of een groep van vleermuizen vanaf een verblijfplaats naar een foerageergebied of tussen verblijfplaatsen vice versa. Het jachtgebied is een gebied waar een vleermuis of een groep van vleermuizen foerageert (Gegevensautoriteit Natuur, 2012). Foerageergebied werd gebruikt als synoniem voor jachtgebied.
De telgegevens en puntwaarnemingen werden verwerkt in een GIS-omgeving (ESRI ArcGIS 10) voor ruimtelijke interpretatie.
Variatie in vliegactiviteit en jachtactiviteit werden geanalyseerd met “generalised linear mixed models” (GLMM) (Zuur et al., 2009). Beide activiteiten werden gekwantificeerd als het aantal tijdsintervallen van 5 seconden over een periode van 3 minuten dat er activiteit werd waargenomen. Activiteit is dus de proportie van tijdsintervallen waarin activiteit plaatsvindt en varieert tussen 0 (geen activiteit) en 36 (activiteit tijdens alle intervallen). De afhankelijke variabele in de analyse is een proportie en dus niet normaal maar binomiaal verdeeld (= generalised model). Om activiteitsvariatie in de tijd te bepalen werden de opnamedatums als verklarende factor in het model gestoken. Vermits opnames gedaan op een bepaald meetpunt niet onafhankelijk zijn (een bepaald punt kan immers meer activiteit hebben omdat het een beter foerageerplaats is) moet daarvoor gecorrigeerd worden. Daarom wordt meetpunt als een random effect in het statistisch model gestoken (= mixed model). Wanneer verschillen in activiteit tussen verschillende meetpunten wordt geanalyseerd zijn opnames gemaakt op eenzelfde dag niet onafhankelijk. Bij deze analyse wordt dan ook opnamedag als random effect in het statistisch model gestoken. Wanneer variatie in functie van de tijd geanalyseerd wordt gaan we er van uit dat dit geen lineair verband is. We verwachten bijvoorbeeld minder activiteit net na of net voor de winter en meer activiteit in de lente en/of zomer. Daarom wordt hier gebruik gemaakt van “generalised additive mixed models” (GAMM) (Zuur et al., 2009). Additive models gebruiken “splines”, een aaneenschakeling van stukjes polynomen, om de regressielijn te bepalen waardoor die een vloeiende curve kan beschrijven. Bij het vergelijken van meetpunten is dit niet nodig vermits meetpunt een categorische variabele is.
De hoger beschreven theorie gaat ook op voor de analyse van de sociale geluiden. Vermits sociale geluiden echter gekwantificeerd werd als aanwezig (1) of afwezig (0) maken we hier gebruik van “mixed logistic models”, een specifieke vorm van GLMM. Alle analyses werden uitgevoerd met het statistisch programma R (versie 3.0.2, R Development Core Team 2013). Voor GLMM’s werd gebruik gemaakt van de glmer procedure van het lme4 pakket (Bates et al 2013), voor de GAMM van de gamm procedure uit het mgcv pakket (Wood, 2014).

Pagina | 9
5. Resultaten
5.1. Algemeen
In 2011 werden 13 tellingen uitgevoerd tussen 18 mei en 13 oktober (zie kaart 2 t.e.m. 7 en 16 t.e.m. 21). Er waren in totaal 25,4 actieve waarnemingsuren in het studiegebied in 2011. Op de meetpunten werd enkel vliegactiviteit geteld. Bij de telling in mei en juni werd slechts gedurende 2 minuten op de meetpunten geteld. Bij de latere tellingen werd een bijgestuurde telmethode gebruikt, en werd er geteld gedurende een periode van 3 minuten. De activiteitstellingen van 2011 werden daarom niet systematisch en kwantitatief besproken in dit rapport, behalve indien grote verschillen met de gegevens van 2012 werd waargenomen. In 2012 werden 15 tellingen uitgevoerd, waarvan 1 voortijdig afgebroken wegens te slechte weersomstandigheden (zie kaart 8 t.e.m. 15 en 22 t.e.m. 35). Er waren in totaal 28,4 actieve waarnemingsuren, waarvan 13,7 uur activiteitstelling, in het studiegebied in 2012. De waarnemingsdata, locatie, start- en eind-uur, start-temperatuur en weersomstandigheden worden in tabellen 3 en 4 in de bijlage weergegeven.
Tabel 1. geeft de tijdens de inventarisatieperiode waargenomen soorten, het aantal passages per soort op de meetpunten en de passages tussen de meetpunten weer. Er werden in 2012 op de meetpunten 1832 passages van vleermuizen geteld, verdeeld over minstens 7 soorten vleermuizen: gewone dwergvleermuis (Pipistrellus pipistrellus), ruige dwergvleermuis (Pipistrellus nathusii), laatvlieger (Eptesicus serotinus), rosse vleermuis (Nyctalus noctula), baardvleermuis (Myotis mystacinus), watervleermuis (Myotis daubentonii) en gewone grootoorvleermuis (Plecotus auritus). Er werden 269 passages tussen de meetpunten geregistreerd (zie kaart 8 t.e.m. 15 en 22 t.e.m. 28).
Tabel 1. Overzicht van de waargenomen soorten en aantal passages tijdens de totale inventarisatieperiode (3 mei tot 26 oktober 2012)
Soort # passages op meetpunten
% passages op meetpunten
# passages
tussen meetpunten
% passages
tussen meetpunten
Pipistrellus pipistrellus 1599 87,28 210 78,07
Pipistrellus nathusii 128 6,99 7 2,6
Eptesicus serotinus 62 3,38 23 8,55
Myotis daubentonii 11 0,60 7 2,6
Nyctalus noctula 4 0,22 4 1,49
Myotis mystacinus 2 0,11 5 1,86
Plecotus auritus 2 0,11 0 0
Myotis spp. 13 0,71 8 2,97
P. auritus/P. austriacus 7 0,38 2 0,74
Eptesicus/Nyctalus 3 0,16 3 1,12
Ongedetermineerd 1 0,05 0 0
Totaal 1832 100 269 100

Pagina | 10
Er werden in beide gebieden 1 tot 3 soorten per meetpunt waargenomen. Figuur 2. en 3. geven de relatieve verdeling van de vliegactiviteit en de waargenomen soorten per meetpunt weer. Gewone dwergvleermuis werd op alle meetpunten waargenomen. De waarnemingen van de andere soorten of soortengroepen was beperkt tot specifieke meetpunten. Gewone dwergvleermuis vertoonde voldoende vlieg- en jachtactiviteit om kwantitatieve analyses mogelijk te maken. De andere soorten worden uitsluitend kwalitatief besproken.
Figuur 2. Relatieve verdeling van de vliegactiviteit en de waargenomen soorten per meetpunt in het gebied Klaverblad (Es: laatvlieger, Msp: Myotis-soorten, Nn: rosse vleermuis, Pn: ruige dwergvleermuis; Pp: gewone dwergvleermuis).
Figuur 3. Relatieve verdeling van de activiteit en de waargenomen soorten per meetpunt langs de Kleine Struisbeek (Es: laatvlieger, Msp: Myotis-soorten, Plsp: grootoorvleermuis, Pn: ruige dwergvleermuis; Pp: gewone dwergvleermuis).
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
KL0
1
KL0
2
KL0
3
KL0
4
KL0
5
KL0
6
KL0
7
KL0
8
KL0
9
KL1
0
KL1
1
KL1
2
KL1
3
KL1
4
KL1
5
KL1
6
KL1
7
KL1
8
KL1
9
KL2
0
KL2
1
KL2
2
Es
Msp
Nn
Pn
Pp
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Es
Msp
Plsp
Pn
Pp

Pagina | 11
Niet alle vleermuizen werden op soort- of op genus-niveau gedetermineerd. De grootoor-vleermuizen werden via geluidsanalyse meestal niet tot op soortniveau gedetermineerd. Het betreft gewone grootoorvleermuis of grijze grootoorvleermuis. Beide soorten komen in Vlaanderen voor, maar gewone grootoorvleermuis is het meest algemeen van de twee soorten (Verkem et al., 2003, Paelinckx et al., 2009). In de studie worden deze waarnemingen verder als grootoorvleermuis benoemd. Sommige vleermuizen op route konden op basis van geluidskenmerken niet tot op genus-niveau bepaald worden. Het ging hoofdzakelijk over laatvlieger en rosse vleermuis. In de studie worden deze verder weergegeven als laatvlieger/rosse vleermuis. In kader van deze studie werden 25 opnames gemaakt van Myotis-soorten. Een aantal van deze opnames konden tot op soort gedetermineerd worden. De determinatie gebeurde op basis van auditieve kenmerken of op basis van een combinatie van auditieve kenmerken en visuele waarnemingen van kenmerken en gedrag. Bijna alle opnames van vleermuizen langs de Kleine Struisbeek behoorden tot het akoestisch type abs moy (afwezigheid van energiepiek met gemiddelde eindfrequentie). Ze konden worden gedetermineerd als watervleermuis (identificatieniveau: zeker of waarschijnlijk) door aanvullende visuele waarnemingen en door de aanwezigheid van overgangen in akoestisch type (abs moy naar abs ht) en de aanwezigheid van een sinusoïdaal verloop in de fundamentele puls. In één opname werd uitsluitend het akoestisch type am moy (explosieve start met gemiddelde eindfrequentie) waargenomen. Volgens Barataud (2012) wordt dit akoestisch type gebruikt door baardvleermuis in een vrij open milieu met een traag ritme en door watervleermuis in een korte fase bij benadering van het bladerdak, of tijdens jacht in een dichte begroeiing. De vleermuis vloog in een open milieu. De pulsduur was meer dan 5 ms. en het pulsinterval (meer dan 90 ms.) was relatief lang. De vleermuis werd geïdentificeerd als baardvleermuis (identificatieniveau: zeker). De opnames van Myotis-soorten in de omgeving van het Klaverblad behoorden tot een aantal types. In een aantal opnames werd uitsluitend het akoestisch type am moy waargenomen. De vleermuizen vlogen in een relatief open milieu. De pulsduur was meer dan 5 ms. en het pulsinterval (meer dan 90 ms.) was relatief lang. De vleermuizen werden geïdentificeerd als baardvleermuis (identificatieniveau: zeker). In een aantal opnames werd een overgang van het akoestisch type am moy naar am ht (explosieve start met hoge eindfrequentie) waargenomen. De pulsduur was bovendien meer dan 5 ms. De vleermuizen werden geïdentificeerd als baardvleermuis (identificatieniveau: zeker). De andere waarnemingen van Myotis-soorten konden niet tot op soort gedetermineerd worden. Er werd uitsluitend het akoestisch type abs moy waargenomen zonder overgangen. Ze werden in de bespreking verder als Myotis-soorten opgenomen. Eén van deze opnames werd ter controle aan Michel Barataud overgemaakt. Hij identificeerde de vleermuis als M.
bechsteinii/dasycneme/mystacinus.
5.2. Vlieg- en jachtactiviteit gewone dwergvleermuis
Vliegactiviteit van gewone dwergvleermuis werd waargenomen van begin mei tot einde oktober (zie kaart 8 t.e.m. 15 en kaart 22 t.e.m. 28). De vliegactiviteit was niet gelijkmatig verdeeld over de inventarisatieperiode. Om dit te testen werd gewerkt via een “generalised additive mixed model” (GAMM) met binomiale error structuur. De afhankelijke variabele is activiteit en de verklarende variabele is datum. Meetpunt werd als random effect in het model ingebracht om te corrigeren van de afhankelijkheid van de data binnen een meetpunt.

Pagina | 12
Figuur 4. Gemiddelde vliegactiviteit van gewone dwergvleermuis over het vliegseizoen. De zwarte lijn is de trend (± SE) voor alle meetpunten gecombineerd. Elke rode lijn is een meetpunt.
Figuur 5. Gemiddelde vliegactiviteit van gewone dwergvleermuis op meetpunten in de omgeving van het Klaverblad (gemiddelde ± SE)
Figuur 6. Gemiddelde vliegactiviteit van gewone dwergvleermuis op meetpunten langs de Kleine Struisbeek (gemiddelde ± SE)
0
10
20
30
0
10
20
30
Kla
verb
lad
Stru
isbee
k
mei jun jul aug sep okt nov
# pa
ssag
es / 3
min
ute
n
0
5
10
KL01
KL02
KL03
KL04
KL05
KL06
KL07
KL08
KL09
KL10
KL11
KL12
KL13
KL14
KL15
KL16
KL17
KL18
KL19
KL20
KL21
KL22
Meetpunt
# pa
ssag
es /
3 m
inut
en
0
5
10
15
GS
01
GS
02
GS
03
GS
04
GS
05
GS
06
GS
07
GS
08
GS
09
GS
10
GS
11
GS
12
GS
13
GS
14
GS
15
Meetpunt
# p
assa
ges
/ 3
min
uten

Pagina | 13
In een eerste stap werd nagegaan of het verloop van vliegactiviteit in beide gebieden verschillende was. Het model met één activiteitscurve (AIC = 1104.52) voor beide gebieden gaf een significant beter resultaat dan deze met een curve voor ieder gebied apart (AIC = 1121.20). Het activiteitspatroon over het seizoen verliep voor beide gebieden dus gelijkmatig. Het model fit significant (F4,179 = 4.40, p = 0.004). Uit figuur 4. kan afgeleid worden dat er een geleidelijke toename van de vliegactiviteit is vanaf ongeveer eind mei tot begin oktober waarna de activiteit terug daalt. De grootste vliegactiviteit werd dus waargenomen in de post-lactatie fase. Globaal genomen was er geen verschil in vliegactiviteit tussen Klaverblad en Kleine Struisbeek (χ�
� = 0.004, p = 0.951).
Binnen een gebied verschilde de vliegactiviteit wel tussen de meetpunten. Figuur 5. geeft de gemiddelde vliegactiviteit op elk meetpunt in de omgeving van het kasteeldomein Klaverblad weer. Er is een significant verschil in gemiddelde vliegactiviteit tussen de meetpunten van het Klaverblad (χ��
� = 285.36, p < 0.001). De meetpunten KL01 en KL11 hebben de hoogste gemiddelde vliegactiviteit. Op deze punten werd gedurende het hele seizoen hoge vliegactiviteit waargenomen. Einde juni werden op KL01 o.a. minstens 4 dwergvleermuizen waargenomen. Op andere meetpunten werd soms ook hoge vliegactiviteit geteld, maar deze vertoonden meer variatie in de loop van het jaar. Dit werd niet statistisch getest in deze studie, maar de verschillen in punten worden indicatief aangegeven. In de kraamperiode hadden o.a. meetpunten KL05, KL07, KL13 en KL21 een hoge vliegactiviteit. In de lactatiefase waren dit de meetpunten KL02, KL03, KL15, KL18, KL19 en KL21. In de post-lactatiefase werden enkel op de meetpunten KL03, KL09, KL12, KL20 en KL22 minder dan 10 passages per 3 minuten geteld. Op 25 oktober werd op de punten KL12, KL16 en KL20 nog hoge vliegactiviteit waargenomen.
Figuur 6. geeft de gemiddelde vliegactiviteit op elk meetpunt langs de Kleine Struisbeek. Er is een significant verschil in gemiddelde activiteit tussen de meetpunten van de Kleine Struisbeek (χ��� = 466.41, p < 0.001). Er is een hogere gemiddelde vliegactiviteit rond de meetpunten
GS05-GS06 en op meetpunt GS14. Op meetpunt GS09 is de minste gemiddelde vliegactiviteit. Op andere meetpunten werd soms ook hoge vliegactiviteit geteld, maar deze vertoonden meer variatie in de loop van het jaar. Dit werd niet statistisch getest in deze studie, maar de verschillen in punten worden indicatief aangegeven. In de kraamperiode waren dit o.a. de meetpunten GS03, GS07 en GS15. Einde mei werd op meetpunt GS03 minstens 3 foeragerende dwergvleermuizen simultaan waargenomen. In de lactatiefase waren dit o.a. de punten GS01, GS06 en GS11. In de post-lactatiefase was er hoge vliegactiviteit op de punten GS08 en GS15 en zeer hoge vliegactiviteit op de punten GS04, GS05, GS06, GS07, GS13 en GS14. Einde augustus waren er meerdere punten waarop minimaal twee exemplaren werden waargenomen.
Figuur 7. geeft de gemiddelde jachtactiviteit van de beide onderzoeksgebieden weer. De gemiddelde jachtactiviteit vertoonde een gelijkaardig patroon als de vliegactiviteit. Ook hier is de AIC van het model met één gezamenlijke jachtactiviteitscurve (1298.53) lager dan die met een verschillende curve per plaats (1336.11). De gemiddelde jachtactiviteit neemt geleidelijk toe vanaf eind mei tot september, waarna de jachtactiviteit terug daalt. De grootste jachtactiviteit werd dus waargenomen in de post-lactatie fase. Het finale model fit significant (F3,179 = 6.50, p < 0.001). Er werd geen globaal verschil in jachtactiviteit tussen beide gebieden vastgesteld (χ��
� = 1.04, p = 0.308). In figuur 8 en 9 wordt de gemiddelde

Pagina | 14
Figuur 7. Gemiddelde jachtactiviteit van gewone dwergvleermuis over het vliegseizoen. De zwarte lijn is de trend (± SE) voor alle meetpunten gecombineerd. Elke rode lijn is een meetpunt.
Figuur 8. Jachtactiviteit van gewone dwergvleermuis op meetpunten in de omgeving van het Klaverblad (gemiddelde ± SE).
Figuur 9. Gemiddelde Jachtactiviteit van gewone dwergvleermuis op meetpunten langs de Kleine Struisbeek (gemiddelde ± SE).
0
10
20
30
0
10
20
30
Klaverblad
Struisbeek
mei jun jul aug sep okt nov
# va
ngst
mom
ente
n /
3 m
inut
en
0.0
2.5
5.0
7.5
10.0
KL01
KL02
KL03
KL04
KL05
KL06
KL07
KL08
KL09
KL10
KL11
KL12
KL13
KL14
KL15
KL16
KL17
KL18
KL19
KL20
KL21
KL22
Meetpunt
# va
ngst
mom
ent
en/3
min
0
5
10
15
GS
01
GS
02
GS
03
GS
04
GS
05
GS
06
GS
07
GS
08
GS
09
GS
10
GS
11
GS
12
GS
13
GS
14
GS
15
Meetpunt
# va
ngst
mom
ente
n/3
min

Pagina | 15
jachtactiviteit per meetpunt van beide onderzoeksgebieden weergegeven. In beide gebieden (Klaverblad: χ��
� = 242.70, p < 0.001; Kleine Struisbeek: χ��� = 404.89, p < 0.001) was er een
significant verschil in activiteit tussen de meetpunten. In het gebied Klaverblad werd op de meetpunten KL01 en KL11 gedurende het hele seizoen hogere jachtactiviteit waargenomen. De hoge jachtactiviteit was soms het gevolg van meerdere jagende vleermuizen (bv. 4 jagende dwergvleermuizen op KL01 in juni). Op 25 oktober was de gemiddelde jachtactiviteit beperkt, maar er werd nog hoge jachtactiviteit waargenomen rond de meetpunten KL11 en KL20. De jachtactiviteit aan de Kleine Struisbeek was gemiddeld het hoogst op meetpunt GS05, GS06 en GS14. Het patroon is duidelijker dan bij vliegactiviteit. Op sommige tijdstippen werd op meetpunten de maximale jachtactiviteit geteld (36 vangstmomenten op 3 minuten). Vermoedelijk is dit een overschatting van de jachtactiviteit als gevolg van de telmethode. De overgangen van heterodyne signalen van meerdere te gelijk jagende vleermuizen klinken vaak zoals vangstmomenten (mondelinge mededeling Ralf Gyselings).
Figuur 10 geeft het verband tussen vliegactiviteit en jachtactiviteit voor beide onderzoeksgebieden weer. Het verband is gelijk voor beide onderzoekgebieden (χ�
� = 0.417, p = 0.518). Bij een vliegactiviteit van 10 of minder passages gedurende 3 minuten bleef de jachtactiviteit beperkt (0 – 2 vangstmomenten per 3 minuten). Bij hogere vliegactiviteiten nam de gemiddelde jachtactiviteit zeer sterk toe als de totale activiteit ook steeg (χ�
� = 67.08, p < 0.001).
Figuur 10. Verband tussen activiteit en voedingsactiviteit voor beide onderzoeksgebieden
De aan- of afwezigheid van sociale geluiden werd geteld op de meetpunten. Voor de omgeving van het kasteeldomein Klaverblad is er een duidelijk significant verschil tussen de tijdstippen (χ�� = 49.89, p < 0.001). Er is een kleine piek in mei en een grotere piek in september –
november (vooral oktober). Begin mei zijn er twee punten waar sociale geluiden werden gedetecteerd: KL10 en KL15. Van einde mei tot einde juli werd op de telpunten geen sociale geluiden waargenomen. Tijdens de telling van 26 augustus werd op 6 telpunten sociale geluiden waargenomen. Vanaf augustus neemt de sociale activiteit toe. Er wordt op 12 telpunten sociale geluiden waargenomen. Op 5 telpunten (KL01, KL10, KL17, KL21 en KL22)
0
10
20
30
0 10 20 30# passages / 3 minuten
# va
ngst
mom
ente
n /
3 m
inut
en
PlaatsKlaverblad
Struisbeek

Pagina | 16
werd zowel in augustus als oktober sociale geluiden waargenomen. Op het telpunt KL17 werden duidelijke baltsvluchten waargenomen.
Het verschil tussen de opnametijdstippen langs de Kleine Struisbeek is niet zo sterk en eigenlijk maar een tendens, bijna significant (χ�
� = 11.46, p = 0.074). In de kraamperiode werden sociale geluiden waargenomen op de telpunten GS03, GS07 en GS14. In de lactatieperiode werden sociale geluiden waargenomen op de telpunten GS05 en GS11. Tijdens de telling van 25 augustus werd op 7 telpunten (GS05 – GS08; GS12 – GS14) sociale geluiden waargenomen. Einde september was dit nog op 3 telpunten (GS06, GS14, GS15). Op het telpunt GS05 en GS14 werden duidelijke baltsvluchten waargenomen.
5.3. Vlieg- en jachtactiviteit van vleermuizen in de omgeving van het domein
Klaverblad
De vlieg- en jachtactiviteit van andere soorten was vaak beperkt tot enkele punten (zie figuur 2). Er waren onvoldoende telgegevens om een statistische analyse te kunnen uitvoeren. De bespreking is beperkt tot kwalitatieve bespreking.
Ruige dwergvleermuis werd op de meetpunten enkel waargenomen in oktober. Op 25 oktober was er beperkte activiteit op 2 meetpunten (KL15 en KL20). Het waren telkens dieren op route. Als de passages tussen de meetpunten mee in rekening worden gebracht, werd ruige dwergvleermuis vanaf einde augustus in het gebied waargenomen. De dieren werden op route in de straat Groenenhoek tussen punten KL15 en KL16, Asterlaan en Leemweg waargenomen. Er werd geen jachtgedrag waargenomen, op een enkel vangstmoment na. Er werden geen sociale geluiden van ruige dwergvleermuis waargenomen. Het aantal waarnemingen was beperkter ten opzicht van de tellingen in 2011. In 2011 werd ruige dwergvleermuis op 7 meetpunten (KL02, KL08-KL11, KL15, KL21), verspreid over 3 tijdstippen (24/08, 14/09 en 10/10) waargenomen. De activiteit concentreerde zich eveneens in de straat Groenenhoek en Leemweg. Op 10 oktober 2011 werd eveneens op de meetpunten KL09 – KL11 in de Klaverblad-dreef hoge vliegactiviteit en jachtactiviteit van ruige dwergvleermuis waargenomen (Johan De Ridder & Daniel Sanders, ongepubliceerd).
Laatvlieger werd gedurende de hele inventarisatieperiode op 12 meetpunten verspreid over het gebied waargenomen. Er werden op de meetpunten 61 passages van laatvlieger geteld. Er waren 14 passages van laatvlieger tussen de meetpunten. Door de aard van de waarneming was het niet altijd duidelijk te bepalen of dit hetzelfde of een ander individu is. De vliegactiviteit vertoonde een maximum in augustus. De vliegactiviteit was verdeeld over het hele gebied, met uitzondering van de Groenenhoek. Op 26 augustus werd een foeragerende laatvlieger visueel waargenomen op de weilanden ten noorden van de Moerelei (ter hoogte van KL01). De laatvlieger joeg 30-50 cm boven de grond en tussen de populieren. De hoge vliegactiviteit op het punt wordt verklaard door continue aanwezig van de vleermuis, waardoor, conform de methodiek, 36 passages werden geteld. Er werd één vangstmoment per 5 seconden geteld, wat wijst op een zeer hoge jachtactiviteit. Op 23 mei werd op de Leemweg (ter hoogte van KL20) een foeragerende laatvlieger visueel waargenomen. De laatvlieger joeg nabij een verlichtingspaal op meikevers. Het aantal vangstmomenten was beperkt tot 1 per minuut. De andere waarnemingen betreft voornamelijk dieren op route (op basis van type signaal). Er werd meer activiteit van laatvlieger waargenomen in 2012 dan in 2011. In 2011 werd er dan wel activiteit ter hoogte van de Groenenhoek waargenomen (Johan De Ridder & Daniel Sanders, ongepubliceerd).
De vliegactiviteit van rosse vleermuis was beperkt tot juni en augustus. Er werd slechts op 2 meetpunten (KL07 en KL10) vliegactiviteit waargenomen. Op het meetpunt KL10 werden op

Pagina | 17
27 juni 3 passages geteld. Op 23 mei werden enkele passages tussen de meetpunten geregistreerd. De vliegactiviteit situeerde zich boven de weilanden ten noorden van de Moerelei en in de Klaverbladdreef. Er werd voor de start van de telperiode, gedurende minimaal 20 minuten vliegactiviteit waargenomen. Dit wijst vermoedelijk op jachtgedrag. In 2011 werd er vanaf juli vliegactiviteit waargenomen van rosse vleermuis. De vliegactiviteit situeerde zich op de Klaverbladdreef tussen meetpunten KL09 en KL11. Op 10 oktober 2011 was er een waarneming ter hoogte van meetpunt KL18. Op 13 juni 2011 werd vliegactiviteit en jachtactiviteit van minimaal twee exemplaren waargenomen hoog boven de weilanden rond de Moerelei (Johan De Ridder & Daniel Sanders, ongepubliceerd).
Myotis-soorten werden van begin mei tot einde augustus bijna uitsluitend waargenomen tussen en op de meetpunten KL05 en KL06 ter hoogte van de Klaverbladreef. Het waren exemplaren op route en foeragerend, en er werden vaak meerdere passages gedurende de 3 minuten periode geteld. De meeste waarnemingen konden niet op soort geïdentificeerd worden. Op 25 juli 2012 werd op de Moerelei tussen meetpunt KL04 en KL05 een opname van een baardvleermuis (identificatieniveau: zeker) gemaakt. De opname van 26 augustus 2012 op meetpunt KL06 in de Klaverbladdreef kon eveneens gedetermineerd worden als baardvleermuis (identificatieniveau: zeker). Op 23 mei 2012 werd een Myotis op route waargenomen ter hoogte van KL13. De opname was niet van voldoende kwaliteit om verder te kunnen determineren. Op 1 oktober werd tussen punt KL01 en KL02 en tussen punt KL02 en KL03 een baardvleermuis (determinatieniveau: zeker) op route waargenomen. Op 25 oktober werd in de Klaverbladdreef ter hoogte van meetpunt KL09 een baardvleermuis (determinatieniveau: zeker) op route waargenomen. In 2011 werden bij de telling Myotis-soorten waargenomen ter hoogte van KL06 op 18 mei, 14 september en 10 oktober en ter hoogte van KL08 op 10 oktober 2011. Passages tussen de meetpunten van Myotis-soorten werden waargenomen op 18 mei tussen de meetpunten KL05 en KL07 en op 14 september op meetpunt KL02 en tussen meetpunten KL05 en KL06. (Johan De Ridder & Daniel Sanders, ongepubliceerd). De vleermuizen werden slechts op genus-niveau gedetermineerd.
Grootoorvleermuis werd op 25 juli tussen meetpunten KL11 en KL12 waargenomen. De vleermuis kon niet tot op soortniveau gedetermineerd worden. In 2011 werden er geen grootoorvleermuizen waargenomen (Johan De Ridder & Daniel Sanders, ongepubliceerd).
5.4. Vlieg- en jachtactiviteit van vleermuizen langs de Kleine Struisbeek
De vlieg- en jachtactiviteit van andere soorten was vaak beperkt tot enkele punten (zie figuur 3). Er waren onvoldoende telgegevens om een statistische analyse te kunnen uitvoeren. De bespreking is beperkt tot kwalitatieve bespreking.
Ruige dwergvleermuis werd tijdens de inventarisatieperiode geteld op 4 punten langs de route. De activiteit was bijna volledige geconcentreerd op meetpunt GS05, waar in mei en augustus tot september hoge vliegactiviteit en hoge tot zeer hoge jachtactiviteit werd waargenomen. Het waren foeragerende dieren boven de vijver aan de cafetaria. 4 mei was er een waarneming op meetpunt GS13 van een individu op route. 26 september waren er waarnemingen op de meetpunten GS02 en GS09 van een individu op route. De waarnemingen werden aangevuld met enkele passages tussen de meetpunten op de campus Drie Eiken. In juni en juli waren er geen waarnemingen van ruige dwergvleermuis. De waarnemingen zijn in overeenstemming met de resultaten van 2011. Ruige dwergvleermuis werd toen op 5 meetpunten waargenomen. Vanaf midden augustus werd ruige dwergvleermuis foeragerend boven de vijver van het cafetaria (GS05) en ter hoogte van de vijver aan meetpunt GS07 waargenomen. De passages tussen de meetpunten situeerden zich op de campus Drie Eiken

Pagina | 18
(meetpunten GS03, GS04 en GS06), en enkele waarnemingen langs het Geitenpad ter hoogte van de punten GS15 en GS13 (Johan De Ridder & Daniel Sanders, ongepubliceerd).
Laatvlieger werd gedurende de hele inventarisatieperiode op slechts 2 meetpunten (GS08 en GS14) waargenomen. Het waren voornamelijk dieren op route (op basis van de activiteitsgraad en signaaltype). Er werden 7 waarnemingen gedaan van laatvlieger tussen de meetpunten, en één waarneming werd benoemd als Eptesicus/Nyctalus. De waarnemingsperiode liep van einde mei tot einde september. De waarnemingen zijn in overeenstemming met de resultaten van 2011. Er werd toen op 5 meetpunten laatvlieger op route waargenomen. Er werden 5 passages tussen de meetpunten waargenomen (Johan De Ridder & Daniel Sanders, ongepubliceerd).
Rosse vleermuis werd niet waargenomen tijdens de inventarisatieperiode in 2012. In 2011 werd rosse vleermuis ter hoogte van meetpunten GS06 en GS07 waargenomen (Johan De Ridder & Daniel Sanders, ongepubliceerd).
Myotis-soorten werden waargenomen op de meetpunten GS03, GS05, GS06 en GS15. Het waren zowel foeragerende vleermuizen als individuen op route. Het betreft watervleermuis, m.u.v. de opname van 26 oktober op GS15. Deze werd gedetermineerd als baardvleermuis (identificatieniveau: zeker). Er werden 5 passages tussen de meetpunten van Myotis-soorten waargenomen. Ze werden geïdentificeerd als watervleermuis (identificatieniveau: waarschijnlijk en zeker). De waarnemingen van watervleermuis op de campus Drie Eiken zijn vergelijkbaar met de waarnemingen in 2011. In 2011 werden geen baardvleermuizen waargenomen (Johan De Ridder & Daniel Sanders, ongepubliceerd).
Grootoorvleermuizen werden van mei tot augustus waargenomen op 7 meetpunten. Het hoofdaandeel van de vliegactiviteit werd voornamelijk waargenomen op de campus Drie Eiken en ter hoogte van het bosgebied Mariënborgh. De opname van 25 augustus kon gedetermineerd worden als gewone grootoorvleermuis. De overige opname waren gewone grootoorvleermuis/grijze grootoorvleermuis. Grootoorvleermuis werd in 2011 enkel ter hoogte van GS05 waargenomen. Het was een onzekere waarneming (Johan De Ridder & Daniel Sanders, ongepubliceerd).
5.5. Wintertelling Klaverblad
In kader van deze studie werden twee wintertellingen uitgevoerd in Schans 16 ter hoogte van de Moerelei. Onafhankelijk van deze studie werd de schans ook bezocht door Wout Willems in de periode 2011 – 2013. Figuur 11 geeft de lokalen van Schans 16 aan die geteld werden. De betonnen fundering van de gevechtskoepel was in 2013 dichtgestort met steenpuin. Een telling kon niet meer uitgevoerd worden.
De eerste wintertelling had plaats op 10 november 2011. Dit was buiten de aan te bevelen telperiode. Er werden twee exemplaren watervleermuis (Myotis daubentonii) gevonden vrij hangend aan het gewelf van de vierde kamer van de traditore batterij. Het gewelf vertoonde hier barsten en vochtinsijpeling zorgde voor natte wanden.
De tweede wintertelling van Schans 16 gebeurde op 12 januari 2013. Er werd 1 exemplaar watervleermuis (Myotis daubentonii) gevonden vrij hangend aan een zijmuur van de vierde kamer van de traditore batterij. Het gewelf was vochtig en water droop van de muren. De vleermuis hing op een stuk van de muur waarvan de beton gedeeltelijk was weg geërodeerd, en de onderliggende bakstenen voor een ruw oppervlak zorgden. Het overzicht van de gekende wintertelresultaten worden weergegeven in tabel 2.

Pagina | 19
Tabel 2. Overzicht van de wintertelgegevens van Schans 16
Datum Myotis daubentonii
Myotis mystacinus
Myotis Mystacinus /brandtii
Teller
10/11/2011 2 0 0 Johan De Ridder
08/02/2011 0 1 1 Wout Willems
20/02/2012 1 1 0 Wout Willems
12/01/2013 1 0 0 Johan De Ridder
27/01/2013 1 0 0 Wout Willems, Kamila Wawrocka
Figuur 11. Grondplan van Schans 16 met aanduiding van lokalen die werden geteld (geel: geteld in 2011 en 2013, groen: geteld in 2011, rood: niet geteld).
6. Bespreking
6.1. Vliegactiviteit en jachtactiviteit
De samenstelling van de vleermuizenpopulatie werd ingeschat op basis van de batdetector-waarnemingen. Er werden tijdens de inventarisatie 7 soorten vleermuizen waargenomen: gewone dwergvleermuis (Pipistrellus pipistrellus), ruige dwergvleermuis (Pipistrellus nathusii), laatvlieger (Eptesicus serotinus), rosse vleermuis (Nyctalus noctula), baardvleermuis (Myotis

Pagina | 20
mystacinus), watervleermuis (Myotis daubentonii) en gewone grootoorvleermuis (Plecotus
auritus). Dit zijn soorten die met zekerheid in de forten rond Antwerpen overwinteren. De vliegactiviteit van gewone dwergvleermuis was het hoogst en was verantwoordelijk voor meer dan 87% van de totale activiteit in het studiegebied. Ruige dwergvleermuis en laatvlieger vertoonden resp. 6,99% en 3,38% van de vliegactiviteit. Barataud (2012) geeft aan dat in Europa 50 – 80% van de waarnemingen met batdetectors uit dwergvleermuizen bestaan. In andere studies van gebieden in Vlaanderen werden gelijkaardige aantallen soorten waargenomen en was het aandeel gewone dwergvleermuis vergelijkbaar (Boeckx & Lefevre, 2002; Van der Wijden, 2003, Dekeukeleire en Nicaise, 2006; Dekeukeleire en Galens, 2011). Onderzoek van Natura 2000 gebieden in Wallonië in de periode 2007 – 2012 gaf bij een vergelijkbare akoestische methode eveneens meer dan 80% contacten met gewone dwergvleermuis (Nyssen, 2012). Een onderzoek van landbouwgebied in Groot – Brittannië met een akoestische methode gaf meer dan 70% activiteit van dwergvleermuizen aan (Wickramasinghe et al., 2003). Het vleermuizenonderzoek in Groen Neerland in Wilrijk gaf 85% activiteit van gewone dwergvleermuis aan (Baetens et al., 2007).
Tabel 3. geeft een overzicht van de waargenomen soorten en vliegactiviteit in de beide deelgebieden. In beide deelgebieden was het grootste aandeel van de vliegactiviteit toe te schrijven aan gewone dwergvleermuis. Er zijn verschillen waar te nemen in de vliegactiviteit van soorten tussen de deelgebieden. In Klaverblad waren de meest waargenomen soorten gewone dwergvleermuis (92,4%), laatvlieger (5,7%) en Myotis-soorten (1,3%). In de omgeving van de Kleine Struisbeek waren gewone dwergvleermuis (80,3%), ruige dwergvleermuis (16,3%) en Myotis-soorten (1,6%) de meest waargenomen soorten. Laatvlieger en Myotis-soorten werden in verhouding ook relatief veel tussen de meetpunten waargenomen (respectievelijk 8,6% en 7,4% van de waarnemingen tussen meetpunten). De vliegactiviteit in 2011 in het Klaverblad werd voor 89,7% toegeschreven aan gewone dwergvleermuis, 4,9% aan laatvlieger en 3,6% aan ruige dwergvleermuis (Johan De Ridder & Daniel Sanders, ongepubliceerd).
Aangezien enkel een ultrasone methode werd gebruikt voor de inventarisatie zal er een zekere overschatting en onderschatting van de aanwezigheid van bepaalde soorten zijn. Soorten die QCF-signalen gebruiken zijn luider dan soorten die exclusief FM-signalen gebruiken. Soorten die tussen vegetatie jagen gebruiken steilere signalen, die over kortere afstanden worden waargenomen (Brigham et al., 2004). Flaquer et al. (2007) stelden bij een vergelijking van methoden vast dat bij inventarisatie met batdetectors holte-bewonende vleermuizen werden onderschat ten opzichte van bijvoorbeeld soorten die in menselijke structuren leven. Een groot deel van de meetpunten is bovendien gesitueerd langs de rand van het gebied, langs overgangen van habitats en langs potentieel foerageergebied van vleermuizen. Dit zou ook een bias van de gegevens kunnen geven. Lesiński et al. (2011) onderzochten de hypothese dat de vliegactiviteit van vleermuizen in bossen hoger is in delen dichter bij de randen o.a. door de aanwezigheid van kraamkolonies in bomen en door foerageergedrag langs de randen van vleermuizen van binnen en buiten het bos. Bij laatvlieger werd een tendens waargenomen, maar enkel bij gewone grootoorvleermuis kon door hen een significant verschil aangetoond worden. Onderzoek in Nederland toonde een verband aan tussen activiteit van gewone dwergvleermuis en bomenrijen (Verboom & Huitema, 1997 & 1999). In het onderzoek werd een verminderende activiteit waargenomen naarmate de afstand tot bomenrijen toenam.

Pagina | 21
Tabel 3. Overzicht van de waargenomen soorten en aantal passages tijdens de totale inventarisatieperiode (3 mei tot 26 oktober 2012) in het gebied Klaverblad en langsheen de Kleine Struisbeek.
Klaverblad Kleine
Struisbeek
Soort # passages op
meetpunten
% passages op meetpunten
# passages
op
meetpunten
% passages
op
meetpunten
Pipistrellus pipistrellus 979 92,36 620 80,31
Pipistrellus nathusii 2 0,19 126 16,32
Eptesicus serotinus 60 5,66 2 0,26
Nyctalus noctula 4 0,38 0 0
Plecotus auritus 0 0 2 0,26
Myotis mystacinus 1 0,09 1 0,13
Myotis daubentonii 0 0 11 1,42
Myotis spp. 13 1,23 0 0
P. auritus/P. austriacus 0 0 7 0,91
Eptesicus/Nyctalus 1 0,09 2 0,26
Ongedetermineerd 0 0 1 0,13
Totaal 1060 100 772 100
Gewone dwergvleermuis is een soort van uiteenlopende biotopen, zoals agrarische gebieden, bossen en stedelijke gebieden. Gewone dwergvleermuizen zijn plaatsgetrouw, maar gebruiken meerdere verblijfplaatsen en wisselen relatief vaak tussen de verblijfplaatsen. Zomer- en winterverblijfplaatsen zijn voornamelijk gebouwen. Kraamkolonies worden hoofdzakelijk in gebouwen gevormd, meestal in een spouwmuur of onder houten dakbedekking (achter betimmering en daklijsten) en dakpannen (Lange et al., 1994; Limpens et al., 1997; Schober, 2001; Zukal & Ŕehák, 2006; Adriaens et al., 2008; Twisk et al., 2010; Dietz et al., 2011). Het jachtgebied is zeer divers en ligt vaak in de beschutting van opgaande elementen en veelal in een groene bebouwde omgeving, o.m. rond gebouwen en straatlantaarns, langs uiteenlopende lijnstructuren zoals kanalen, bomenrijen, dreven, houtwallen en holle wegen en tussen de boomkruinen en boven open plekken van bossen (vooral oude voedselrijke loofbossen), tuinen en parken (Lange et al., 1994; Limpens et al., 1997; Schober, 2001; Zukal & Ŕehák, 2006; Adriaens et al., 2008; Twisk et al., 2010; Dietz et al., 2011). Bij voorkeur jagen ze op een hoogte tussen 2,5 en 4,5 m boven de grond, en op enige afstand (1 tot 8 m) van de vegetatie. Gewone dwergvleermuis jaagt hoofdzakelijk binnen een straal van 2 tot 5 km van de verblijfplaats (Limpens et al., 1997, Verboom & Huitema, 2010). Vliegroutes volgen zoveel mogelijk lijnvormige structuren. Gewone dwergvleermuis vertoonde vlieg en jachtactiviteit gedurende de ganse inventarisatieperiode. Er werd een geleidelijke toename van de vlieg- en jachtactiviteit waargenomen vanaf ongeveer eind mei tot begin oktober waarna de

Pagina | 22
activiteit terug daalt (figuur 12 en 13). Op de meetpunten KL01, KL11, GS05, GS06 en GS14 werd de hoogste gemiddelde vliegactiviteit waargenomen. De meetpunten met hoge vlieg- en jachtactiviteit liggen vermoedelijk in of langs jachtbiotoop en vliegroutes. De zeer beperkte vliegactiviteit van gewone dwergvleermuis op 26 oktober is waarschijnlijk te verklaren door de lage begintemperatuur (<10° C). Ter hoogte van de meetpunten GS05, GS06, GS07, GS14 en KL17 werden zangvluchten van gewone dwergvleermuis waargenomen. De piek van sociale geluiden werd waargenomen einde augustus. Wijzigingen in de seizoenactiviteit van gewone dwergvleermuis werd door verschillende onderzoekers vastgesteld. Er worden verschillende verklaringen gegeven aan de variatie in de vliegactiviteit over het seizoen, o.a. omwille van wijzigingen in de reproductieve cyclus, verschuivingen van jachthabitat en nazomerzwermactiviteit. In het onderzoek van Kusch en Idelberger (2005) waren er indicaties dat dwergvleermuizen van jachthabitat wisselen in de loop van het seizoen. Kapfer et al. (2007) stelden een significant lagere vliegactiviteit vast van gewone dwergvleermuis tijdens de kraamtijd dan in de lactatie en post-lactatie fase. Jonker et al. (2010) namen een duidelijk patroon waar in de seizoenactiviteit van gewone dwergvleermuis. De vliegactiviteit nam toe gedurende de zomer, en was het laagst in de nazomer. De toename wordt verklaard door de hogere energiebehoeften van lacterende vrouwtjes, en doordat juveniele vleermuizen individueel beginnen te jagen. De positieve correlatie tussen vliegactiviteit en jachtactiviteit die we vaststelden in deze studie, kan deze hypothese ondersteunen. De toename kan ook verklaard worden door de aanwezigheid van zomerzwermgedrag in de omgeving van een winterverblijfplaats. Dit werd o.a. vastgesteld door Sendor et al. (2002).
Ruige dwergvleermuis is een soort van half open, bosrijke landschappen. (Lange et al., 1994; Limpens et al., 1997; Schober, 2001; Zukal & Ŕehák, 2006; Twisk et al., 2010, Dietz et al., 2011). De kraamkolonies zijn hoofdzakelijk gevestigd in Noord-, Midden- en Oost-Europa. Van ruige dwergvleermuis is seizoensmigratie gekend. In het najaar trekken de vleermuizen naar westelijk Europa (Limpens et al., 1997; Russ et al., 1998; Ciechanowski en Jarzembowski, 2009), waar ze paren en overwinteren. De winterverblijfplaatsen zijn bomen, soms gebouwen. De vliegroutes liggen langs lijnvormige landschapselementen. De jachtgebieden liggen tot 5 – 6 kilometer van de verblijfplaatsen. De jachtactiviteit van ruige dwergvleermuis is het hoogst boven waterpartijen en vochtige bossen (Gelhaus & Zahn, 2010). De mannetjes bezetten in de herfst een paarverblijf dicht bij de kraamkamers, langs de migratieroutes of winterverblijven. Met baltsroepen lokken zij langskomende dieren (Dietz et al., 2011). Er werd in het studiegebied activiteit van ruige dwergvleermuis waargenomen op een beperkt aantal punten. De vliegactiviteit was onvoldoende groot om kwantitatieve analyses op uit te voeren. In het Klaverblad bleef de activiteit beperkt tot enkele passages van individuen op route. Op de campus Drie Eiken werd foerageergedrag waargenomen ter hoogte van de vijver aan de cafetaria (meetpunt GS05). Er was een continue vliegactiviteit van vermoedelijk meerdere exemplaren op dit punt waardoor de totale vliegactiviteit van ruige dwergvleermuis in verhouding hoog is. De vliegactiviteit was beperkt tot mei en van augustus tot oktober. Dit komt overeen met de volgens de literatuur te verwachten periode van aanwezigheid van de soort. Er werden beperkt sociale geluiden van ruige dwergvleermuis waargenomen. Het betreft het typische B-segment uit de roep (Russ et al., 1998; Middleton, 2006; Jahelková, 2010). De sociale geluiden werden waargenomen bij exemplaren op route en bij foeragerende exemplaren. Er werden geen zangvluchten waargenomen. In 2011 werd eveneens vliegactiviteit van ruige dwergvleermuis waargenomen. De activiteit was dan meer verspreid over verschillende locaties op de campus Drie Eiken (meetpunt GS02, GS03, GS04, GS06 en GS07). In 2011 werden zangvluchten waargenomen ter hoogte van meetpunt GS05 (De Ridder & Sanders, ongepubliceerd).

Pagina | 23
Laatvlieger is een soort van parken, bosranden, open gebied zoals vochtige graslanden en akkers en randen van stedelijk gebied (Lange et al., 1994; Limpens et al., 1997; Schober, 2001; Zukal & Ŕehák, 2006; Adriaens et al., 2008; Twisk et al., 2010; Dietz et al., 2011). De zomer- en kraamverblijven zijn gesitueerd in gebouwen. Over de winterverblijfplaatsen is weinig gekend, laatvlieger overwintert o.a. in beperkte mate in de forten (Baetens et al., 2003; Verkem, 2008; Beers et al., 2009; Verkem, 2009; Boers et al., 2010; De Ridder, ongepubliceerd). De vliegroutes liggen langs lijnvormige landschapselementen. Laatvliegers foerageren in open en halfopen gebieden, boven weilanden en op enkele meters van opgaande begroeiing. Laatvliegers jagen op een hoogte van 3 – 4 m, soms lager tot 30 cm boven het oppervlak (Simon et al., 2004). Het jachtgebied ligt in de regel binnen 1 (maximaal 3 tot 4) kilometer van de verblijfplaatsen (Limpens et al., 1997). Laatvlieger is een standsoort. De vliegactiviteit was onvoldoende groot om kwantitatieve analyses op uit te voeren. De activiteit was in het gebied Klaverblad gelijkmatig verdeeld over het gebied, maar langs de Kleine Struisbeek beperkt tot een aantal locaties. Laatvlieger vertoonde de grootste vliegactiviteit van begin mei tot einde augustus. Het ging meestal over dieren op route waarvan een éénmalige passage werd geregistreerd. In juli en augustus werd jachtactiviteit waargenomen van laatvlieger boven de weilanden van het Klaverblad. De dieren vertoonden duidelijk langzame korteafstand jachtvlucht (slow hawking). Dit is een typische jachtstrategie voor laatvlieger (Limpens et al., 1997; Cel’uch et al., 2004). De seizoenactiviteit van laatvlieger is geconcentreerd in de late lente en de zomer (Ciechanowksi et al., 2010). Bartonička (2003) registreerde in de lactatieperiode (tweede helft juni en begin juli) een verhoging in activiteit bij laatvlieger. Seizoenale variatie in de jachtactiviteit van laatvlieger kan verklaard worden door de fenologie van de prooien. Laatvlieger profiteert van insectenpopulaties die uitkomen in een korte periode van de nacht of het seizoen. Het betreft vaak grote prooien die een energetisch voordeel opleveren (Kervyn, 2007). Op 23 mei werd o.a. op de Leemweg (ter hoogte van KL20) een foeragerende laatvlieger visueel waargenomen. De laatvlieger joeg nabij een verlichtingspaal op meikevers. Jachtactiviteit van laatvlieger aan verlichtingspalen wordt vaker waargenomen (Bartonička, 2002; Bartonička & Zukal, 2003; Zukal & Ŕehák, 2006). Het afnemen of wegvallen van de activiteit van laatvlieger wordt verklaard door het uiteenvallen van kraamkolonies (Catto et al., 1995) en door verplaatsing naar andere jachtgebieden, o.a. van bosranden naar meer urbane habitats (De Jong en Ahlén, 1991). Dit kan een verklaring zijn voor de beperkte activiteit van laatvlieger in het gebied vanaf september. Laatvlieger wordt in een aantal Antwerpse stedelijke parken nog steeds waargenomen in de periode september – oktober (Johan De Ridder, persoonlijk observatie).
Rosse vleermuis is een soort van oeverbossen, of water- en bosrijke gebieden (Lange et al., 1994; Limpens et al., 1997; Twisk et al., 2010). De zomer- en winterverblijven en paarplaatsen zijn voornamelijk bomen (met name eiken en beuken, maar ook in grote populieren) op oude landgoederen, in parken, buitenplaatsen en bomenrijen langs wegen. Er wordt een netwerk van verblijfplaatsen bewoond. Rosse vleermuis maakt geen gebruik van vaste vliegroutes. Het jachtgebied zijn open wateren, moeras en weiland. Jachtgebied is gelegen binnen een straal van 3-6 km, een enkele keer tot tientallen km rondom de kolonieplaats. In de zomer leven vrouwtjes in kraamkolonies (20-70 dieren, soms enkele honderden). Mannetjes leven zomers apart van de vrouwtjes. Vanaf begin augustus bezetten mannetjes paarplaatsen in boomholten, en lokken van hieruit vrouwtjes. Rosse vleermuis is een migrerende soort die van begin september tot in de late herfst voornamelijk naar het zuidwesten trekt. Rosse vleermuis werd in 2012 enkel met zekerheid waargenomen in de omgeving van het domein Klaverblad.

Pagina | 24
Watervleermuis is een soort van half open tot gesloten, water- en bosrijk landschap. De zomerverblijven en kraamverblijven zijn voornamelijk gevestigd in (loof)bomen. Er wordt een netwerk van verblijfplaatsen bewoond, die om de 2 à 5 dagen worden gewisseld. Ze overwinteren vaak in ondergrondse of met grond overdekte constructies zoals bunkers, forten, en (ijs)kelders. Het jachtgebied is voornamelijk beschut gelegen water, maar ook boven land, boven bospaden of op beschutte, open plekken in het bos. De vliegroutes liggen langs lijnvormige landschapselementen (Lange et al., 1994; Limpens et al., 1997; Schober, 2001; Zukal & Ŕehák, 2006; Twisk et al., 2010). De afstand van kolonieplaats tot jachtgebied bedraagt gemiddeld 2 tot 4 km (tot 15 km). De afstand tussen zomer- en winterverblijven is meestal klein (15-25 km). In zomer leven vrouwtjes in kraamkolonies, mannetjes leven ‘s zomers apart van de vrouwtjes. De seizoenactiviteit van watervleermuis vertoont een bimodaal karakter met de hoogste piek in april en een kleinere piek in augustus-september (Ciechanowksi et al., 2009). Encarnação et al. (2006) nam een duidelijke piek waar van (jacht)activiteit van mannelijke watervleermuizen van eind juni tot midden augustus. Watervleermuis vertoonde in het studiegebied vliegactiviteit van begin mei tot einde augustus. De vliegactiviteit was onvoldoende groot om kwantitatieve analyses op uit te voeren. De activiteit was beperkt tot een aantal waterrijke locaties langsheen de Kleine Struisbeek. Er werden slechts een beperkt aantal vangstmomenten gedetecteerd. In de omgeving van het Klaverblad kon watervleermuis enkel als overwinterende soort met zekerheid vastgesteld worden.
Baardvleermuis is een soort van bosrijke gebieden, structuurrijke parken en cultuurlandschap met veel houtige landschapselementen en bij voorkeur weiden (Lange et al., 1994; Limpens et al., 1997; Schober, 2001; Zukal & Ŕehák, 2006; Twisk et al., 2010; Dietz et al., 2011; Buckley et al., 2012). De zomerverblijven zijn meestal gebouwen (huizen, kerken) en boomholten. Er wordt een netwerk van verblijfplaatsen bewoond, die om de 10 à 14 dagen worden gewisseld. Ze overwinteren vaak in ondergrondse of met grond overdekte constructies zoals bunkers, forten, en (ijs)kelders. De baardvleermuis leeft jaarrond in kolonies. Baardvleermuis jaagt langs bospaden, bosranden en open plekken in bos, soms boven water en volgen lijnvormige landschapselementen als vliegroutes. Het jachtgebied ligt vaak binnen een straal van 1 km rondom kolonieplaats (Buckley et al., 2012). In de zomer leven vrouwtjes in kraamkolonies (20-70 dieren). Mannetjes leven zomers apart van de vrouwtjes. Baardvleermuis is een standsoort. De afstand tussen winter- en zomerverblijven kan 10 tot 80 km bedragen. Baardvleermuis vertoonde in het studiegebied vliegactiviteit gedurende de ganse inventarisatieperiode. De vliegactiviteit was onvoldoende groot om kwantitatieve analyses op uit te voeren. De activiteit was beperkt tot een aantal locaties in de nabijheid van dreven of oud bosgebied. Myotis-soorten gebruiken uitsluitend FM-signalen. FM-signalen worden minder sterk opgevangen dan QCF-signalen door batdetectors (Brigham et al., 2004). Het aandeel Myotis-soorten is mogelijks onderschat.
Gewone grootoorvleermuis is een typische soort van bosrijk gebied en parken. De zomerverblijven zijn gesitueerd in (kerk)zolders en holle bomen. Er wordt een netwerk van verblijfplaatsen bewoond. De winterverblijven en paarverblijven zijn gesitueerd in (ijs)kelders, bunkers, forten, etc. De jachtgebieden zijn bos- en parkachtige gebieden en plantsoenen. Grootoorvleermuizen volgen vliegroutes langs lijnvormige landschapselementen (Lange et al., 1994; Limpens et al., 1997; Schober, 2001; Zukal & Ŕehák, 2006; Twisk et al., 2010). Het jachtgebied is gelegen binnen een straal van 4 km rondom de kolonieplaats. In de zomer leven vrouwtjes in kraamkolonies (gem. 5-30 dieren). Mannetjes verblijven in kleine groepjes in de omgeving van de kraamkolonie. De kraamtijd is van half juni tot half augustus. De afstand tussen winter- en zomerverblijven doorgaans beperkt tot enkele kilometers. Grootoorvleermuis

Pagina | 25
vertoonde in het studiegebied vliegactiviteit van einde mei tot einde september. De vliegactiviteit was sterk geconcentreerd rond de campus Drie Eiken en het gebied langs de Kleine Struisbeek. Grootoorvleermuizen jagen vaak in en tussen boomkruinen. De signalen van in vegetatie jagende soorten worden minder sterk opgevangen dan soorten die jagen in open terrein (Brigham et al., 2004). Het aandeel grootoorvleermuis is waarschijnlijk onderschat. Bij verder onderzoek van de Klaverbladdreef in 2013 werd door de onderzoekers bij elk bezoek grootoorvleermuis waargenomen (De Ridder, J & Sanders, D., 2014 in voorbereiding).
6.2. Landschapsgebruik Klaverblad
De waargenomen jachtgebieden en vliegroutes in de omgeving van kasteeldomein Klaverblad worden weergegeven op kaart 36. Het betreft een synthese van de observaties uit 2011 en 2012. De omgeving van de Moerelei, tussen kruispunt Klaverblad en Leemweg werd door vleermuizen gebruikt als vliegroute en jachtgebied. Gewone dwergvleermuis kon, bij elk bezoek aan het inventarisatiegebied, op route via de Moerelei naar de jachtterreinen worden waargenomen. De vliegactiviteit was vaak al bezig voor de aanvang van de effectieve tellingen. De vleermuizen volgden daarbij de rijen knotbomen, opgaande bomen en de bosrand. Het meetpunt KL01 ligt op deze vliegroute en in geschikt jachtgebied, waardoor de hoge vlieg- en jachtactiviteit verklaard kan worden. Gewone dwergvleermuizen maken zowel in de zomer als in de winter gebruik van gebouwen als verblijfplaats. Woonhuizen in de Varenvelden (gemeente Hemiksem) doen vermoedelijk dienst als dagverblijfplaatsen. Jenkins et al. (1998) toonden aan dat verblijfplaatsen van gewone dwergvleermuizen binnen 0,5 km van grotere loofboombossen liggen, en dat lijnvormige elementen vaak leiden van verblijfplaats naar jachtgebied. Verboom en Huitema (2010) toonden aan dat de voorkeur van gewone dwergvleermuis voor vliegroutes afhankelijk is van de aanwezigheid van goede jachtgebieden dichtbij de verblijfplaats. Laatvlieger, baardvleermuis en andere Myotis-soorten maakten ook in beperkte mate gebruik van de Moerelei als vliegroute.
Gewone dwergvleermuis foerageerde langs de bomenrijen en bosranden langs de Moerelei. De weilanden ten noorden en ten zuiden van de Moerelei werden gebruikt als jachtgebied door gewone dwergvleermuis en laatvlieger. Dwergvleermuis maakte hierbij gebruik van de kleine landschapselementen (knotwilgen en opgaande wilgen). Laatvlieger volgde kleine landschapselementen en joeg eveneens boven open stukken. Rosse vleermuis werd eveneens foeragerend waargenomen ter hoogte van de weilanden ten noorden van de Moerelei. In 2011 werden hier eveneens jagende rosse vleermuizen waargenomen (Johan De Ridder & Daniel Sanders; ongepubliceerd). De vleermuizen kwamen aanvliegen vanuit noordoostelijke richting, mogelijks vanuit Fort 7 of Schoonselhof.
Het noordelijke deel van de Klaverbladdreef werd door baardvleermuis en andere Myotis-soorten gebruikt als vliegroute. Opmerkelijk is dat de waarnemingen gebeurden in het verlichtte deel van de dreef. Aangezien de Myotis-soorten ook in de Moerelei werden waargenomen, wordt vermoed dat de vliegroute zich situeert van Moerelei naar noorden van Klaverbladdreef. Baardvleermuis vertoonde vliegactiviteit gedurende de ganse inventarisatieperiode. De activiteit was tot en met augustus beperkt tot de omgeving van de Klaverbladdreef (KL05 en KL06). Nadien werden de waarnemingen verder van deze punten gedaan. Dit komt overeen met de periode van het uiteenvallen van de kraamkolonies van baardvleermuis. Het is niet uit te sluiten dat er een kraamkolonie van baardvleermuis gesitueerd is in (de (nabijheid van) de Klaverbladdreef.

Pagina | 26
In het zuidelijk deel van de Klaverbladdreef werden vnl. foeragerende dwergvleermuizen en occasioneel laatvlieger op route waargenomen. De vleermuizen foerageerden vnl. boven de weilanden aan weerszijden van de dreef. Het meetpunt KL11 met zeer hoge vlieg- en jachtactiviteit bevindt zich op het einde van deze dreef. Het meetpunt KL07 met hoge jachtactiviteit bevindt zich in de dreef. De dreefbomen werden als richtinggevend gebruikt bij de jacht. De vleermuizen hielden daarbij twee tot drie meter afstand van de bomen. Er werd ook jacht tussen de bomen waargenomen. De binnenkant van de dreef werd eerder gebruikt als vliegroute. Dit is in overeenstemming met waarnemingen van Verboom en Huitema (2010) in Nederland. In 2011 werd vanaf september jachtactiviteit en vanaf oktober zwermactiviteit waargenomen van ruige dwergvleermuis in dit deel van de Klaverbladdreef. De vleermuizen foerageerden langs de buitenkant van de dreef en tussen de stammen van de dreefbomen (De Ridder & Sanders, ongepubliceerd). In 2012 was deze vlieg- en jachtactiviteit van ruige dwergvleermuis niet aanwezig. In 2012 werd in mei en juni vliegactiviteit van rosse vleermuis waargenomen in de Klaverbladdreef. In 2011 en 2012 werd in de maand augustus activiteit van rosse vleermuis waargenomen in de Klaverbladdreef. Het ging over slecht enkele passages. De paartijd van rosse vleermuis situeert zich in augustus. Er werden echter geen locaties met roepende mannetjes (roepbomen) vastgesteld. Ter hoogte van de twee toegangsdreven naar het kasteeldomein werd slechts beperkte activiteit waargenomen (vnl. dwergvleermuis). Op 25 oktober 2012 werd op het segment van de Klaverbladdreef tussen de twee toegangsdreven (KL09) een waarneming gedaan van een baardvleermuis. Op het eind van de Klaverbladdreef werd hoge jachtactiviteit waargenomen van gewone dwergvleermuis tussen de dreefbomen en op de rand van het akkerlandperceel. In 2011 werd hier ook jachtactiviteit van rosse vleermuis en ruige dwergvleermuis waargenomen (De Ridder & Sanders, ongepubliceerd). Grootoorvleermuis werd op 25 juli 2013 waargenomen in de Klaverbladdreef ter hoogte van dit punt.
De zuidelijke uitloper van de Klaverbladdreef en de straat Groenenhoek werd eveneens gedurende het ganse voorjaar en zomer gebruikt als vliegroute door gewone dwergvleermuis. De vleermuizen volgden hierbij de populierenrij langs de weg. De vleermuizen vlogen parallel met de bomenrij maar hielden hierbij een afstand van een 5-tal meter. In 2011 werd hier bijna het hele seizoen vlieg- en jachtactiviteit van laatvlieger vastgesteld (Johan De Ridder & Daniel Sanders, ongepubliceerd). Dit was in 2012 niet het geval.
Ter hoogte van de oude hoeve op de Groenenhoek werd bij elke ronde jagende gewone dwergvleermuis, soms meerdere exemplaren, waargenomen. Vermoedelijk is een zomerverblijfplaats of kraamkolonie van gewone dwergvleermuis gesitueerd in de oudere hoevegebouwen. Aanwezigheid van (kraam)verblijfplaats van laatvlieger is eveneens niet uit te sluiten. Ter hoogte van de oude hoeve aan meetpunt 15 werd eveneens vlieg- en jachtactiviteit waargenomen van gewone dwergvleermuis. Op 25 oktober was hier ook activiteit van ruige dwergvleermuis.
In de Valkenisseweg en het verlengde van de Groenenhoek werd gedurende het hele seizoen vlieg- en jachtactiviteit van gewone dwergvleermuis en laatvlieger waargenomen. Einde juli was de vliegactiviteit van gewone dwergvleermuis hoog op de punten KL18 en KL19. De jachtactiviteit was op dat moment eveneens hoog op punt KL18. Dit punt is gesitueerd ter hoogte van een lantaarnpaal. De hoge activiteit kan verklaard worden door een dwergvleermuis die aan de verlichting jaagt op insecten. De hoge vliegactiviteit in juli kan ook samenvallen met het uiteenvallen van kraamkolonies in kleine groepen. Sociale geluiden werden op dit segment vanaf einde augustus sporadisch waargenomen, en op alle locaties vanaf begin oktober. Op het punt KL17 werden einde augustus en begin oktober duidelijke baltsvluchten uitgevoerd door gewone dwergvleermuis. Dit wijst op territoriumvestiging door

Pagina | 27
een mannetje. De dominante mannetjes beginnen baltsvluchten uit te voeren vanaf de kraamkolonies zijn uiteengevallen. De baltsvluchten starten midden juli en kunnen doorgaan tot in oktober, met een piek in september. De territoria hebben een gemiddelde diameter van 200 meter (Sachteleben & von Helversen, 2006).
6.3. Landschapsgebruik Kleine Struisbeek
De waargenomen jachtgebieden en vliegroutes in de omgeving van de Kleine Struisbeek en de campus Drie Eiken worden weergegeven op kaart 37. Het betreft een synthese van de observaties uit 2011 en 2012. Het Geitenpad tussen Kleine Doornstraat en Doornstraat werd door vleermuizen gebruikt als vliegroute. De waarnemingen waren voornamelijk gewone dwergvleermuis, maar ook laatvlieger, grootoorvleermuizen, baardvleermuis en watervleermuis. De vliegroute werd zuidelijk verder gezet langs de bomenrij in de Kleine Doornstraat, en noordelijk over de Doornstraat verder langs het Geitenpad richting de campus Drie Eiken van de Universiteit Antwerpen.
De uitloper van het bosgebied Mariënborgh langs de Kleine Struisbeek werd intensief gebruikt als jachtgebied door gewone dwergvleermuis. De jachtactiviteit was gelokaliseerd in het bos en langs de bosrand. Ter hoogte van meetpunt 14 werden meermaals meerdere exemplaren gewone dwergvleermuis foeragerend aangetroffen. Er werden bovendien verdedigingsvluchten waargenomen en sociale geluiden. Dit wijst er op dat meetpunt 14 mogelijks in een paarterritorium ligt. Mannetjes van gewone dwergvleermuis zijn territoriaal. Paarterritoria liggen vaak rond bomen. Mannetjes dwergvleermuizen nemen territoria in met een oppervlakte van 1,2 tot 10 ha. Deze territoria liggen bij voorkeur langs vliegroutes (Sachteleben en Von Helversen, 2006). Aan de bosrand (meetpunt GS13) werden 4 soorten waargenomen: gewone dwergvleermuis, ruige dwergvleermuis, grootoorvleermuis en watervleermuis (determinatieniveau: waarschijnlijk). Gewone dwergvleermuis werd buiten het beboste gedeelte ook foeragerend boven de Kleine Struisbeek waargenomen. Op 25 oktober 2012 werden twee exemplaren baardvleermuis op route vanuit het bos Mariënborgh richting de Kleine Doornstraat waargenomen. De winterverblijfplaats is vermoedelijk in de omgeving van het bos gesitueerd.
De vijvers en de extensieve graslanden langs de Kleine Struisbeek tussen Kleine Doornstraat en Doornstraat werden slechts beperkt gebruikt als jachtgebied. Vermoedelijk is er onvoldoende dekking van kleine landschapselementen, of is het voedselaanbod boven de vijvers nog van onvoldoende kwaliteit. Gewone dwergvleermuis joeg langs opgaande elementen zoals houtkanten, bomenrijen en boven wateroppervlakten (Verboom & Huitema, 1997; Jenkins et al., 1998; Kusch et al., 2004; Lesiński et al., 2011). Laatvlieger werd op deze locaties waargenomen, maar het betreft eerder dieren op route dan foeragerende exemplaren.
Het Geitenpad tussen Doornstraat en de campus Drie Eiken werd door vleermuizen gebruikt als vliegroute. De waarnemingen waren hoofdzakelijk gewone dwergvleermuizen, maar ook laatvlieger, ruige dwergvleermuis en grootoorvleermuizen. Gewone dwergvleermuis en laatvlieger foerageerden langs de structurerende houtkanten, knotbomen- en populierenrijen en boven de loop van de Kleine Struisbeek. De jachtactiviteit was echter beperkt.
De campus Drie Eiken is een jachtgebied voor verschillende vleermuissoorten. De vijver ter hoogte van GS07 werd door gewone dwergvleermuis als jachtgebied gebruikt. De vlieg- en

Pagina | 28
jachtactiviteit was er het grootst in augustus. Er werden dan ook sociale geluiden waargenomen. Jachtactiviteit en sociale geluiden werden eveneens begin mei waargenomen. Het ging voornamelijk over sociale geluiden van het type D. Dit zijn sociale geluiden met een complexe structuur. De sociale geluiden worden gebruikt door mannetjes als paringslokroep, maar ook om andere vleermuizen weg te jagen van een jachtgebied (Pfalzer & Kusch, 2003; Barlow & Jones, 1997). Grootoorvleermuis, watervleermuis en rosse vleermuis werden eveneens waargenomen op dit punt. In 2011 werden op dit meetpunt 5 soorten waargenomen: gewone dwergvleermuis, ruige dwergvleermuis, laatvlieger, rosse vleermuis en watervleermuis.
Op de parking ter hoogte van het Q-gebouw van de campus en de Universiteitsbaan werd voornamelijk vlieg- en jachtactiviteit van gewone dwergvleermuis waargenomen. Het meetpunt GS06 is hier gelegen op het bruggetje aan de parking. Vanaf einde juli tot begin oktober werd een hoge vliegactiviteit en jachtactiviteit van gewone dwergvleermuis waargenomen. Sociale geluiden en mogelijke zangvluchten werden waargenomen vanaf einde augustus. Watervleermuis op route werd op verschillende tijdstippen waargenomen ter hoogte van meetpunt GS06. De verbindingsgracht met houtkant werd telkens gevolgd door de watervleermuis. In 2011 werd op dit meetpunt ook activiteit van ruige dwergvleermuis, laatvlieger en rosse vleermuis waargenomen (Johan De Ridder & Daniel Sanders, ongepubliceerd).
De vijver aan de cafetaria van de Universiteit Antwerpen werd gedurende het hele seizoen gebruikt als jachtgebied door gewone dwergvleermuis. De dwergvleermuizen joegen zowel boven het wateroppervlakte als boven de kronen van de oeverbegroeiing. De activiteitsgraad lag er hoog ten gevolge van continue aanwezigheid van meerdere jagende exemplaren (minimum 2 of 3). Vanaf augustus werden op deze locatie sociale geluiden van gewone dwergvleermuis waargenomen. In mei en van augustus tot oktober foerageerden er op deze locatie eveneens ruige dwergvleermuis. Er werden sociale geluiden van het type C waargenomen. Type C sociale geluiden zijn karakteristiek voor interacties tussen vrouwtjes en jongen en zijn belangrijk tijdens tandemvluchten (Pfalzer en Kusch, 2003).
De vlieg- en jachtactiviteit van watervleermuis op de vijvers van de universiteitscampus was relatief beperkt. Het betreft toevallige waarnemingen en activiteitstellingen ter hoogte van de meetpunten GS03, GS05, GS07. Het punt GS06 ligt vermoedelijk langs een vliegroute. In 2011 werden langs dit punt eveneens vaak passages van watervleermuis waargenomen (Johan De Ridder & Daniel Sanders, ongepubliceerd). Het is niet duidelijk waarom er slechts beperkte activiteit is op de vijvers.
Grootoorvleermuis werd meermaals waargenomen op verschillende locaties op de campus Drie Eiken. Grootoorvleermuis is vaak moeilijk te detecteren met een batdetector. Er wordt daarom uitgegaan dat de campus een belangrijk leefgebied is voor deze soort. De activiteit situeerde zich voornamelijk langs het beboste gedeelte van de Kleine Struisbeek en in de buurt van de vijvers (GS03, GS04, GS05 en GS07). Grootoorvleermuis foerageert gemiddeld slechts 500 meter van de kraamplaats. Er wordt verwacht dat een kraamkolonie grootoorvleermuis aanwezig is op de campus Drie Eiken of in de omgeving ervan. Deze situeert zich mogelijks in één van de dreven die naar Fort 6 leidt (Ben Vanderwijden, mondelinge mededeling). Ben Vanderwijden (mondeling mededeling) nam ook een hoge activiteit van grootoorvleermuis waar op de campus Drie Eiken, en zwermgedrag aan het reduit van Fort 6.
Het segment van het Geitenpad tussen de campus en de Edegemsesteenweg loopt langs de Kleine Struisbeek en door een bebost gebied. Deze zone werd door gewone dwergvleermuis als

Pagina | 29
vliegroute gebruikt en als foerageergebied. Opvallend is dat de activiteit hoog lag bij de start van de inventarisatie (soms meer dan 4 exemplaren), maar dat bij eigenlijke telling er slechts beperkte activiteit werd waargenomen. Dit kan er op wijzen dat de meetpunten dichtbij een dagverblijfplaats gelegen zijn, en daardoor vroeg op de avond als vliegroute worden gebruikt. Grootoorvleermuis werd op dit segment in de periode mei tot augustus verschillende malen waargenomen.
7. Aanbevelingen voor verder onderzoek
De omgeving van het domein Klaverblad, de Kleine Struisbeek en de campus Drie Eiken zijn een belangrijk leefgebied binnen de levenscyclus voor vleermuizen. Er wordt gedurende het hele voorjaar, zomer en nazomerseizoen activiteit van vleermuizen waargenomen. Alle vleermuisfuncties van het gebied zijn echter nog onvoldoende in kaart gebracht. Onderzoek met batdetectors geeft een beeld van de soortenrijkdom van een gebied, maar zou aangevuld moeten worden met o.a. vangsttechnieken, telemetrisch onderzoek en onderzoek naar kraamkolonies en verblijfplaatsen. Aandachtspunten bij verder onderzoek is dan ook het uitbreiden van de onderzoeksmethoden en het verder in kaart brengen van alle functies van het gebied voor vleermuizen. De volgende aanbevelingen voor verder onderzoek worden gedaan:
- Maken van hoogwaardige opnamen in de Klaverbladdreef of het gebruiken van vangsttechnieken om tot een zekere soortdeterminatie van Myotis-soorten te komen.
- Uitgebreid onderzoek van de vleermuisfuncties van de Klaverbladdreef voor o.a. Myotis-soorten en rosse vleermuis.
- Onderzoek van de vleermuizenfuncties van gebieden die momenteel onvoldoende onderzocht zijn in het kader van deze studie, oa. Kasteeldomein Klaverblad, Ferrarisbos langs de Fotografielaan, bosgebied Mariënborgh, en de Put van De Meyvis. Dit zijn overwegend privé-eigendommen waarvoor eerst toestemming van de eigenaar dient bekomen te worden voor verder onderzoek.
- Inspectie van de historische gebouwen van het kasteeldomein Klaverblad op aanwezigheid van kraamkolonies en winterverblijfplaatsen.
- Inventarisatie van kraamverblijfplaatsen met o.a. locatiestudie van een mogelijke kraamkolonie van grootoorvleermuis op de campus Drie Eiken.
- Uitvoeren van een jaarlijkse wintertelling in Schans 16. Opname van Schans 17 bij de wintertelling.
- Uitvoeren van controles op de cafetariavijver van de Universiteit Antwerpen in de periode februari – maart voor controle op aanwezigheid van ruige dwergvleermuis.

Pagina | 30
8. Aanbevelingen voor beheer
De aanbevelingen zijn algemeen geformuleerd, omdat het onderzoek verkennend opgevat was. Op basis van verder onderzoek van vleermuizenfuncties kunnen voorgestelde maatregelen verder uitgediept, of gelokaliseerd worden. Het voorkomen van vleermuizen wordt beperkt door de aanwezigheid van geschikt leefgebied, namelijk voldoende winter- en zomerverblijfplaatsen, een netwerk van jachtgebieden en de verbindingen tussen verblijfplaatsen en jachtgebieden (Jenkins et al., 1998). Bij het beheer van de jachtgebieden moet voldoende aandacht besteed worden aan het verkrijgen van een hoge insectenrijkdom. Dit is vaak het geval in gevarieerde gebieden met afwisseling van open en dichte structuren. Veel vleermuissoorten gebruiken holten in bomen als verblijfplaats. Vleermuizen maken deze holten niet zelf, maar zijn afhankelijk van diverse holtevormende processen. Vleermuizen leven in groepen en verhuizen regelmatig. Daarom zijn op lokale schaal gebieden nodig met een voldoende groot aanbod aan boomholten. De sterke afhankelijkheid van zeer specifieke holten in bomen is voor vleermuissoorten vaak beperkend voor voorkomen en verspreiding. Een afname van bomen met holten kan leiden tot een afname of het geheel verdwijnen van vleermuissoorten uit een bepaald gebied. Er kunnen specifieke tot algemene maatregelen getroffen worden om de waarde als vleermuizenhabitat van de onderzochte gebieden in stand te houden, of indien nodig te verhogen.
8.1. Voorzorgsmaatregelen
Belangrijke bedreigingen voor vleermuizen in Vlaanderen zijn vernietiging van leefomgeving en verstoring tijdens belangrijke fasen in de levenscyclus (Verkem, 1998; Paelinckx et al., 2009). De uitbreiding van industrie in de omgeving van het kasteeldomein Klaverblad is een bedreiging en verstoring van het leefgebied van vleermuizen. Tijdens en na de onderzoeksfase van deze studie werd dit duidelijk door o.a. de kap van het zogenaamde Ferrarisbos (september 2013) en de uitbreiding van het industrieterrein richting Klaverbladdreef. Hiermee verdwijnt niet enkel jachthabitat en potentiele zomer- en kraamverblijfplaatsen, maar er treedt eveneens verstoring op door toenemende lichtvervuiling binnen de invloedssfeer van een jachtgebied en vliegroute.
Bij uitvoering van beheermaatregelen in oude bosbestanden en dreven moet voldoende aandacht uitgaan naar mogelijke vleermuisbomen. Het beheer gebeurt dan het best onder begeleiding van een vleermuisdeskundige. De volgende krachtlijnen voor het beheer kunnen meegegeven worden:
- Huidige verblijfplaatsen in bomen zo veel mogelijk veilig stellen; - Herstel of kappen in de dreven of bossen gefaseerd uitvoeren met prioriteit aan niet
vleermuisbomen; - Bij uitvoering van verjongingen of onderhoudskappen in dreven, specifieke controle laten
uitvoeren van de geviseerde bomen door een vleermuisdeskundige op aanwezigheid van vleermuizen om kapslachtoffers te vermijden;
- Kappingen van oude loofbomen uitvoeren in voor vleermuizen minst gevoelige periode. Dit is het vroege voorjaar (april) of najaar (half september-eind oktober), maar alleen als er geen paarverblijven aanwezig zijn.
8.2. Beheermaatregelen voor de omgeving van het kasteeldomein Klaverblad
De omgeving van het kasteeldomein Klaverblad is een belangrijk leefgebied voor gewone dwergvleermuis, laatvlieger, ruige dwergvleermuis en een aantal Myotis-soorten. Gewone

Pagina | 31
grootoorvleermuis is vermoedelijk een belangrijke doelsoort. Beheervisies en beheer- en inrichtingsmaatregelen zouden deze soorten als doelsoort moeten meenemen. Het kasteeldomein Klaverblad wordt gekenmerkt door een afwisseling van weilanden, akkers en bossen. De verschillende functies zijn verweven via een netwerk van kleine landschapselementen. Dit maakt het een waardevol leefgebied voor vleermuizen. Er werden in het gebied zowel jachthabitat als vliegroutes vastgesteld. Maatregelen die gericht zijn op het versterken van de huidige functies, en het verhogen van de ecologische en milieukwaliteit zullen bijdragen aan de geschiktheid van vleermuizen in het gebied. Vele andere dieren en plantensoorten zullen eveneens profiteren van de voorgestelde maatregelen. De voorgestelde maatregelen zijn suggesties naar eigenaars en beheerders van het gebied, en worden bij voorkeur opgenomen in een geïntegreerd beheerplan waarbij rekening gehouden wordt met ecologische en erfgoedwaarden, maar ook met private belangen.
Alle vleermuizen in Europa zijn insectenetende zoogdieren. Een leefgebied is interessant als er jachtgebieden met voldoende insectendensiteit beschikbaar zijn. Het vermijden van het gebruik van insecticiden als algemene maatregel is daarom ten alle tijden aanbevolen.
Een deel van de Klaverbladdreef kan aangeduid worden als een ecologische groep (12-24 bomen), waarbij uitgegaan wordt van vooral die bomen waar nu al holten zijn. De bomen in deze groep worden niet gekapt bij verjonging en het beheer is gericht op hun vleermuisfunctie. Indien dit niet mogelijk is omwille van veiligheidsredenen of esthetische redenen dan kan het ophangen van vleermuiskasten een oplossing bieden.
Oude bosbestanden met oude bomen (>100 jaar oud) in de buurt van het Klaverbladdomein moeten in stand gehouden worden als mogelijks dagverblijf en kraamverblijfplaats voor vleermuizen. Dit is bijvoorbeeld van toepassing voor het Ferrarisbos nabij de Fotografielaan. Binnen bestaande jonge bossen kunnen delen aangeduid worden die oud mogen worden. Er kan een oud-hout eiland (3-5 ha) op het Klaverbladdomein aangeduid worden op <500m van de Klaverbladdreef, of op <500m van een dwarsdreef op de Klaverbladdreef. Het beheer in het oud-hout eiland wordt gericht naar vleermuizen en de ontwikkeling van een holterijk perceel. Streefdoel is 10 (potentiële of effectieve) vleermuizenbomen per hectare. Zolang er niet voldoende oude bomen beschikbaar zijn, kan tijdelijk gewerkt worden met vleermuizenkasten.
In de omgeving van het Klaverblad liggen heel wat extensief beheerde weilanden of paardenweiden. Deze vormen een jachtgebied van laatvlieger, rosse vleermuis en dwergvleermuizen. Simon et al. (2004) stelden bij telemetrisch onderzoek in Duitsland vast dat weilanden tot het voorkeursjachthabitat van laatvlieger behoren. Ze stelden bovendien een positieve correlatie vast tussen de aanwezigheid van kraamkolonies van laatvlieger en (door runderen) begraasde weilanden. Een beheer en beleid gericht op het verder ontwikkelen van een kleinschalig landschap met het behoud van weilandfunctie, versterken van kleine landschapselementen en een extensief beheer gericht op de ontwikkeling van soortenrijke graslanden of perceelranden met een hoge insectendiversiteit is aanbevolen. Verdere uitbreiding van industriële activiteiten (op landbouwgronden) richting het domein Klaverblad moet vermeden worden. Houtige kleine landschapselementen en perceelranden die aansluiten of deel uitmaken van een bosrijk landschap verruimen het jachtgebied van de o.a. baardvleermuis, gewone dwergvleermuis en laatvlieger en zorgen voor de verbinding tussen de verschillende foerageergebieden enerzijds en de verschillende verblijfplaatsen anderzijds. Verboom en Huitema (1997) argumenteren dat landschappen met een hoge densiteit aan lineaire elementen een meerwaarde hebben voor laatvlieger. Het behouden en versterken van het netwerk aan kleine landschapselementen in het gebied Klaverblad wordt daarom aanbevolen om het leef- en jachtgebied van laatvlieger te versterken. Bomen en bomenrijen

Pagina | 32
worden net als houtkanten gevolgd als vliegroute tussen foerageergebied en verblijfplaats. Een aaneensluiting van dergelijke landschapselementen verhoogt de geleiding naar geschikte leefgebieden. Dit is relevant voor gewone en ruige dwergvleermuis, laatvlieger, baardvleermuis en watervleermuis. Vleermuizen zijn gevoelig voor lichtverstoring bij verblijfplaatsen, op vliegroutes en in het jachtgebied. De klaverbladdreef tussen Sint-Bernardsesteenweg en Moerelei is verlicht. Overschakelen naar een vleermuisvriendelijke verlichting voor dit segment kan op termijn een positieve maatregel zijn voor de aanwezige vleermuizenpopulatie. Het achterste deel van de Klaverbladdreef is niet verlicht door straatverlichting. Tussen meetpunt KL11 en KL09 wordt er echter storende verlichting waargenomen van het naastgelegen bedrijf. Het is aanbevolen om op termijn over te schakelen naar vleermuisvriendelijke verlichting. De aanleg van een bufferstrook van o.a. houtkanten of bomenrijen tussen het domein klaverblad en de naastgelegen industrie wordt aanbevolen. Dit zal een positief effect hebben op de landschappelijke beleving en het leefgebied van de aanwezige vleermuispopulatie. De Klaverbladdreef vormt een mogelijke verbinding tussen het domein Klaverblad en Fort 8. De verbinding kan versterkt worden door een inrichting van de oversteek van de Sint-Bernardsesteenweg met een middenberm met bomen (‘hop over’ voor vleermuizen), en het plaatsen van vleermuisvriendelijke verlichting in de aanpalende woonwijk.
In de Moerelei dient het beheer gericht te worden op het behouden en versterken van de kleine landschapselementen zoals (knot)bomenrijen en houtkanten. De bosranden aan de Moerelei dienen beheerd te worden in functie van ontwikkeling van mantelzoomvegetaties als geleidingselement en als jachtgebied. Een bosrandbeheer gericht op het verhogen van de insectendensiteit is aanbevolen. In de omgeving van het domein Klaverblad zijn heel wat dreven en opgaande bomenrijen (Klaverbladdreef, Groenenhoek, …). Deze worden gebruikt als jachtgebied en als geleidend element. Grote onderbrekingen van deze dreven en opgaande bomenrijen moeten vermeden worden en waar nodig kan worden aangeplant om het “gat” te dichten. Als bomenrijen alsnog moeten worden gekapt vanwege hun ouderdom of slechte staat, dan gebeurt de vervanging gefaseerd. De Moerelei vormt de enige verbinding voor vleermuizen richting Fort 7 en het Schoonselhof. Het oostelijk gedeelte van de Moerelei heeft een meer urbaan karakter. Een meer vleermuisvriendelijke inrichting van de Moerelei, o.a. door het plaatsen van aangepaste straatverlichting en door de aanplant van geleidende beplantingen zal de connectiviteit tussen het Klaverblad en het complex Fort 7 – Schoonselhof versterken.
De Groenenhoek is een landbouwgebied bij uitstek. Heel wat vleermuizen zoals gewone dwergvleermuis, laatvlieger en baardvleermuis vinden een geschikt jachtgebied rond veehoudende boerenerven maar evenzeer op plaatsen met veel houtige elementen zoals hoogstamboomgaarden, heggen en houtkanten of bomenrijke tuinen. Wickramasinghe et al. (2003) toonde aan dat vleermuizenactiviteit hoger is op biologische boerderijen dan op traditionele boerderijen. Buiten het behoud van bovenstaande structuurelementen, moet aandacht worden gegeven aan kleine structuurelementen waar rottend organisch materiaal of mest achterblijft. Composthopen, kippenrennen, brandhoutstapels, (val)fruitweiden, uitloopplekken voor vee aan stallen en paardenweiden lokken vaak veel insecten. Ook drenkplaatsen en natuurlijke tuinvijvers evenals structuurrijke beken vormen een geschikte jachtplek. Het vermijden van insecticiden is een belangrijke randvoorwaarde.
Schans 16 wordt momenteel gebruikt als overwinteringsplaats door vleermuizen. Er kunnen een aantal maatregelen getroffen worden om de vleermuizenfunctie van de schans te verhogen. Een belangrijke randvoorwaarde is hierbij het akkoord van de eigenaar(s). Schans 16 is bovendien uniek erfgoed. De maatregelen moeten gebeuren in afstemming met de

Pagina | 33
erfgoedwaarde van de schans en moeten omkeerbaar zijn. De volgende maatregelen kunnen genomen worden:
- verwijderen stortmateriaal, afval en puin uit de schans - traditore batterij ontoegankelijk maken van september tot april door het plaatsen van
deuren met horizontale vliegopening - tocht in traditore batterij vermijden door het plaatsen van gechicaneerd portaal - plaatsen van aanhechtingsmogelijkheden voor vleermuizen door het aanbrengen van
verruwingen aan de muren of het voorzien van voorwerpen met holten en spleten.
De functie van Schans 17 als vleermuizenoverwinteringsplaats is onbekend. De toestand van de schans is echter vergelijkbaar met Schans 16. Een inrichting kan zinvol zijn, maar een kleinveestal in gebruik is moeilijk combineerbaar met overwinterende vleermuizen. De betonnen fundering van de geschutskoepel kan mogelijks wel ingericht worden.
De verbinding van Schans 16, Schans 17 en het domein Klaverblad met Fort 8 kan gerealiseerd worden door een inrichting van de Sint-Bernardsesteenweg nabij de kruising met de Moerelei. Hier kunnen bomen en/of een bosje worden aangeplant om een geschikte ‘hop over’ voor vleermuizen te creëren.
8.3. Beheermaatregelen voor de omgeving van de Kleine Struisbeek
De omgeving van de Kleine Struisbeek vormt een jachtgebied en verbindingscorridor voor gewone dwergvleermuis, ruige dwergvleermuis en een aantal Myotis-soorten. Maatregelen die deze functies versterken, zullen bijdragen aan de bescherming van vleermuizen. Het versterken van de functionele verbinding met andere (potentiële) leefgebieden zoals Groen Neerland, bosgebied Mariënborgh, Fort 6 en de Put van de Meyvis is een belangrijk aandachtspunt.
Het bosgebied Mariënborgh is al een oud-hout eiland in de onmiddellijke nabijheid van de bedding van de Kleine Struisbeek. Er zijn al verschillende oude bomen met loshangende schors, scheuren, spleten en holten aanwezig. In de onmiddellijke omgeving van Mariënborgh werden typische bos- en boombewonende vleermuizen zoals gewone grootoorvleermuis, baardvleermuis en rosse vleermuis waargenomen. In het bosgebied Mariënborgh kunnen maatregelen in functie van vleermuizen genomen worden. Het bosbeheer dient zich in te eerste plaats te richten naar de ontwikkeling van een natuurlijk gevarieerd bos met een inheemse ondergroei o.a. door het verwijderen van rododendron en het verwijderen van groenafval en huishoudelijk afval uit het bosgebied. In een natuurlijker bos komt een hogere insectendensiteit voor en dat komt de vleermuizen ten goede. In de tweede plaats dient het beheer van het bos en de beukendreven zich te richten op het behoud van voldoende oude bomen (120 jaar of ouder) met voor vleermuizen geschikte holten. Het bos kan zich natuurlijk ontwikkelen, zonder dunnen of zuivering, waardoor bomen met elkaar concurreren. Er komen dan ook kwijnende bomen voor waar spechten en schimmels sneller kansen krijgen om holtes te ontwikkelen. Per hectare moet er minstens één tot drie dikke staande dode bomen met loshangende schors en holten behouden blijven. Streefbeeld is behoud en ontstaan van minstens tien vleermuisbomen (potentiële of werkelijke) per hectare. Indien dit niet haalbaar is, bijvoorbeeld omwille van veiligheidsredenen, dan kan het ophangen van vleermuiskasten uitkomst bieden.
Vleermuizen zijn gevoelig voor lichtverstoring bij verblijfplaatsen, op vliegroutes en in het jachtgebied. De sportterreinen ter hoogte van het bosgebied Mariënborgh worden ’s avonds tijdelijk verlicht, vaak op momenten waarop vleermuizen jagen. Het plaatsen van een

Pagina | 34
vleermuisvriendelijke verlichting kan hier wenselijk zijn. Type verlichting en kleur is hierbij belangrijk.
De Kleine Struisbeek en het Geitenpad vormen een vliegroute voor vleermuizen. De route leidt langs verschillende jachtgebieden. De aanwezige oude knotwilgen, maar ook houtkanten vormen een geleidingselement. Het behouden en versterken van kleine landschapselementen, o.a. door het heraanplanten van knotwilgen, is een belangrijk element om de route te versterken. De route wordt doorkruist door de Doornstraat. Het kruispunt van het Geitenpad met de Doornstraat vormt een open ruimte van meer dan 100 meter. De zone is bovendien verlicht. Dit vormt voor vele vleermuissoorten een barrière. De inrichting van een vleermuisvriendelijk kruispunt, o.a. door het plaatsen van aangepaste straatverlichting en door de aanplant van enkele geleidende beplantingen en een klein bosje is aanbevolen. Belangrijk aandachtspunt is dat de straatverlichting het baanvak verlicht en niet de houtige geleidingselementen.
De vijvers langsheen de Kleine Struisbeek in het segment tussen Kleine Doornstraat en Doornstraat worden slechts beperkt gebruikt als jachtgebied van vleermuizen. Ze liggen wel langs een vliegroute en zijn potentieel interessant als jachtgebied. Een aangepast beheer van de oevervegetatie en waterkwaliteit gericht op het verhogen van de insectendensiteit kan bijdragen aan de versterking als jachthabitat. De vijvers zijn verbonden via een centrale gracht. De afwateringsgrachten van een nabijgelegen landouwperceel zijn eveneens verbonden met deze gracht. Het risico van vermesting van de poelen ten gevolge van landbouwactiviteit op nabijgelegen percelen is bijgevolg groot.
De vijvers op de campus Drie Eiken worden momenteel gebruikt als foerageergebied van gewone dwergvleermuis. Watervleermuis foerageert hier slechts beperkt. De vijvers liggen wel op een vliegroute tussen de Campus Drie Eiken en o.a. de put van de Meyvis en Fort 6. Het behoud en versterken van deze vliegroute is aangewezen, net zoals het meer geschikt maken van de vijvers als jachthabitat voor watervleermuis. Grootoorvleermuis wordt regelmatig waargenomen op de campus Drie Eiken. Het voorkomen van de soort in relatie tot de campus en omliggende omgeving (o.a. Fort 6) is nog onvoldoende duidelijk om maatregelen te kunnen voorstellen.
Fort 6 is een zwermplaats en winterverblijfplaats voor vleermuizen. De landschappelijke verbinding tussen de campus Drie Eiken en Fort 6 moet behouden blijven en indien mogelijk versterkt. Op de campus Drie Eiken kunnen vleermuizenkasten opgehangen worden om het aantal geschikte dag- en kraamverblijfplaatsen te verhogen.

Pagina | 35
9. Literatuurlijst
Adriaens D., Adriaens T., & Ameeuw G. (red.) (2008). Ontwikkeling van criteria voor de beoordeling van de lokale staat van instandhouding van de habitatrichtlijnsoorten. Rapporten van het Instituut voor Natuur- en Bosonderzoek 2008 (35). Instituut voor Natuur- en Bosonderzoek, Brussel.
Ahlén I. & Baagøe H.J., (1999). Use of ultrasound detectors for bat studies in Europe: experiences from field identification, surveys, and monitoring. Acta Chiropterologica 1 (2): 137-150.
Baetens, J., De Ridder, J., Gyselings, R., Backx, H., Waeterschoot, H. & Meire, P. (2007). Open advies inrichtings- en beheerplan BPA30/Neerland.
Baetens, J., Beers, P., De Ridder, J. & Versweyveld, S. (2003). Fort 7: Antwerpen: E-022: eerste monitoringsrapport en conformverklaring: december 2003: dossier administratie. Monitoringrapport. Natuurpunt: Mechelen. 18 pp.
Bates, D., Maechler, M., Bolker, B. & Walker, S. (2013) lme4: linear mixed-effects models using S4 classes. R Package version 1.0.5.
Beers, P., De Ridder, J. & Versweyveld, S. (2009). Fort 7 E-022 (Wilrijk) : 2e monitoringrapport januari 2009 : dossier ANB Monitoringrapport, E-022. Natuurpunt: Mechelen. 14 pp.
Barataud, M. (2012). Ecologie acoustique des chiroptères d’Europe, identification des espèces, étude de leurs habitats et comportements de chasse. Biotope, Mèze; Muséum national d’Histoire naturelle, Paris (collection Inventaires et biodiversité). 344p.
Barlow K.E. & Jones G., (1997). Function of pipistrelle social calls: field data and a playback experiment. Animal Behaviour 53: 991-999.
Bartonička, T. (2002). Habitat use of four bat species in Jablonec n.N. revealed by bat detector. Przyrode Sudetów Zachodnich Suplement 2: 79 – 87.
Bartonička, T. & Zukal, J. (2003). Flight activity and habitat use of four bat species in a small town revealed by bat detectors. Folia Zoologica 52(2): 155–166.
Bat Conservation Trust (2007). Bat Surveys – Good Practice Guidelines. Bat Conservation Trust, London. 82 p.
Beckers K., Vermeersch G., Maes D., Adriaens T., De Beer D., De Knijf G., Bosmans R., Hendrickx F., Jooris R., Maelfait J.P., Van Den Berge K., Van Keer K., Van Landuyt W. & Van Thuyne G. (2009). Een gericht natuurbeleid voor de prioritaire soorten in de provincie Antwerpen. Rapport INBO.R.2009.7. Instituut voor Natuur- en Bosonderzoek, Brussel, i.o. van de Provincie Antwerpen
Boeckx K. & Lefèvre A., 2002. Chiropterologisch onderzoek in de ruilverkaveling Herenthout-Bouwel. Natuurpunt Studie, Mechelen.
Boeckx K. & Lefèvre A., 2002. Chiropterologisch onderzoek in de ruilverkaveling Herenthout-Bouwel. Natuurpunt Studie, Mechelen. Antwerpse koepel voor natuurstudie jaarboek 2002. 43 - 49
Boers, K., Verkem S., & Willems, W. (2010). Verslag fortentelweekend 5-6-7 februari 2010. Natuurpunt Werkgroep Vleermuizen. 12p.
Boers, K., Verkem S., & Willems, W. (2013). Verslag fortentelweekend 1-2-3 februari 2013. Natuurpunt Werkgroep Vleermuizen. 9p.
Boers K., Lefevre A. & Verkem S. (2012). Vleermuisinventarisatie in het kader van het provinciaal RUP voor de “Poort Vrieselhof – Fort van Oelegem” te Ranst. Rapport Natuurpunt Studie 2012/3, Natuurpunt Studie, Mechelen, België. 26p.
Brigham, R. M., et al., (2004). Bat Echolocation Research: tools, techniques and analysis. Bat Conservation International. Austin, Texas. 167p.

Pagina | 36
Buckley, D.J., et al. (2012) The spatial ecology of the whiskered bat (Myotis mystacinus) at the western extreme of its range provides evidence of regional adaptation. Mammal. Biol. http://dx.doi.org/10.1016/j.mambio.2012.06.007
Catto C.M.C., Hutson A.M., Racey P.A. & Stephenson P.J. (1996). Foraging behaviour and habitat use of the serotine bat (Eptesicus serotinus) in southern England. Journal of Zoology 238: 623-633.
Catto, C. M. C., Racey, P. A. & Stephenson P. J. (1995). Activity patterns of the serotine bat (Eptesicus serotinus) at a roost in southern England. Journal of Zoology 235(4): 635–644.
Cel’uch, M. & Kaňuch, P. (2004). Foraging and flight activity of bats in beech-oak forests (Western Carpathians). Folia Oecologica 31(1):8-16.
Ciechanowski M. & Jarzembowski, T. (2009). Dynamics of social organization and phenology of Nathusius’ pipistrelle Pipistrellus nathusii (Keyserling and Blasius, 1839) (Chiroptera: Vespertilionidae) occupying bird and bat boxes – interseasonal approach. Le Rhinolophe 18: 1-6
Ciechanowski, M., Zasjac, T., Zielinska, A. & Dunajski, R. (2010). Seasonal activity patterns of seven vespertilionid bat species in Polish lowlands. Acta Theriologica 55 (4):301-314.
Criel, D., Lefevre, A., Van Den Berge, K., Van Gompel, J. & Verhagen, R. (1994) Rode lijst van de zoogdieren in Vlaanderen. AMINAL, Brussel.
de Jong, J. & Ahlén I. (1991). Factors affecting the distribution pattern of bats in Uppland, central Sweden. Holarctic Ecology 14: 92–96.
De Ridder, J., Sanders D., Van Deynze, W. & Waeterschoot, H. (2014). Vleermuizen op Fort 7: synthese van de beschikbare gegevens en aanzet tot een visie en zoneringsplan. Rapport Natuurpunt Zuidrand Antwerpen werkgroep Studie 2014/1. In voorbereiding
Demolder H. & Peymen, J. (2012). Natuurindicatoren 2012. Toestand van de natuur in Vlaanderen: cijfers voor het beleid. Mededeling van het Instituut voor Natuur- en Bosonderzoek, INBO.M. 2012.2, Brussel.
Dekeukeleire, D., Janssen, R., Boers, K. & Willems, W. (2011). Verkennend onderzoek van zwermende vleermuizen bij de forten van Antwerpen in 2010. Rapport Natuurpunt Studie 2011/9, Natuurpunt Studie, Mechelen, België.
Dienst Duurzaam Milieu- en Natuurbeleid (2010). Provinciale Prioritaire Soorten Provincie Antwerpen. Provincie Antwerpen.
Dietz, C., Helversen O. von, & D. Nill. (2011). Vleermuizen. Alle soorten van Europa en Noord-West Afrika. Tirion Natuur. 400 p.
Dijkstra, V. & Korsten, E. (2005). Handleiding wintertellingen van vleermuizen. Voor het monitoren van vleermuizen in de winter. Zoogdierenvereniging VZZ. Arnhem. 20p.
Encarnação, J. A., Kierdorf, U. & Wolters, V. (2006). Seasonal variation in nocturnal activity of male Daubenton’s bats, Myotis daubentonii (Chiroptera: Vespertilionidae). Folia Zoologica 55(3): 237–246
Flaquer, C., Torre I. & Arrizabalaga A. (2007). Comparison of sampling methods for inventory of bat communities. Journal of Mammalogy. 88(2):526-533.
Gaisler J., Zukal J., Rehak Z. & Homolka M., (1998). Habitat preference and flight activity of bats in a city. Journal of Zoology 244: 439-445.
Gegevensautoriteit Natuur. (2009). Vleermuisprotocol. http://www.gegevensautoriteit-natuur.nl/files/vleermuisprotocol-2-april-2009-pdf.pdf(07-10-2011)
Gegevensautoriteit Natuur. (2012). Vleermuisprotocol. http://www.gegevensautoriteitnatuur.nl/items/vleermuisprotocol-bijgesteld.aspx
Gelhaus, M. & Zahn, A. (2010). Roosting ecology, phenology and foraging habitats of a nursery colony of Pipistrellus nathusii in the southwestern part of its reproduction range. Vespertilio 13–14: 93–102

Pagina | 37
Gils, R. (1998). Vesting Antwerpen deel 2: De Pantservesting 1885 - 1914. België onder de wapens 7. 80p.
Jahelková, H. (2010). Unusual social calls of Nathusius’ pipistrelle (Vespertilionidae, Chiroptera) recorded outside the mating season. Folia Zoologica. 60(1): 25–30.
Jenkins, E.V., Laine T., Morgan, S.E., Cole K.R. & Speakman J.R. (1998). Roost selection in the pipistrelle bat, Pipistrellus pipistrellus (Chiroptera: Vespertilionidae), in northeast Scotland. Animal Behaviour 56:909-917.
Jonker, M. N., De Boer, W.F., Kurvers, R. & Dekker, J. (2010) Foraging and public information use in common pipistrelle bats (Pipistrellus pipistrellus): a field experiment. Acta Chiropterologica, 12(1): 197–203.
Kapfer, G. & Aron, S. (2007). Temporal variation in flight activity, foraging activity and social interactions by bats around a suburban pond. Lutra 50(1): 9 – 18.
Kervyn T. & Libois R. (2007). The Diet of the serotine bat: A Comparison between rural and urban environments. Belgian Journal of Zoology 138 (1) : 41-49
Kurta A. & Teramino J.A. (1992). Bat community structure in an urban park. Ecography 15: 257-261.
Kusch J., Weber C., Idelberger S. & Koob T., (2004). Foraging habitat preferences of bats in relation to food supply and spatial vegetation structures in a western European low mountain range forest. Folia Zoologica 53 (2): 113-128
Kusch J. & Idelberger S. (2005). — Spatial and temporal variability of bat foraging in a western European low mountain range forest. Mammalia 69 (1): 21-33.
Lange R., Twisk, P., van Winden, A., & van Diepenbeek, A. (1994). Zoogdieren van West-Europa (deel vleermuizen). Uitgave KNNV-uitgeverij.
Lee, Y. & McCracken, G. F. (2004). Flight Activity and Food Habits of Three Species of Myotis Bats (Chiroptera: Vespertilionidae) in Sympatry. Zoological Studies 43(3): 589-597.
Lesiński, G., Olszewski, A. & Popczyk, B. (2011). Forest roads used by commuting and foraging bats in edge and interior zones. Polish Journal of Ecology 59(3):611-616.
Limpens, H., Mostert, K. & Bongers, W. (red.) (1997). Atlas van de Nederlandse vleermuizen. Onderzoek naar verspreiding en ecologie. Stichting Uitgeverij KNNV, Utrecht.
Meschede, A. & Heller, K. (200). Ökologie und schutz von Fledermäusen in Wäldern. Bundesamt für Naturschutz: Schriftenreihe für landschaftsplege und Naturschuz Heft 77. 374 p.
Mickleburgh S.P., Hutson A.M. & Racey P.A., (2002). A review of the global conservation status of bats. Oryx 36 (1): 18-34.
Middleton, N. E. (2006). A study of the emission of social calls by Pipistrellus spp. within central Scotland; including a description of their typical social call structure. BaTML Publications, Vol 3, 23-28.
NARA 2005: Dumortier M., De Bruyn L., Hens M., Peymen J., Schneiders A., Van Daele T., Van Reeth W., Weyembergh G. & Kuijken E., (2005). Natuurrapport 2005. Toestand van de natuur in Vlaanderen: cijfers voor het beleid. Mededeling van het Instituut voor Natuurbehoud nr. 24, Brussel.
Nieuwborg H. (red.) (2001). Antwerpse Koepel voor Natuurstudie (ANKONA). Jaarboek 2000. Provinciebestuur Antwerpen.
Nyssen, P. (2012). Natura 2000 inventarisaties in Wallonië: overzicht van 5 jaar vleermuizenstudie. Presentatie Vleermuizenstudiedag KBIN – 24 maart 2012
Paelinckx D., et al. (red.) (2009). Gewestelijke doelstellingen voor de habitats en soorten van de Europese Habitat- en Vogelrichtlijn voor Vlaanderen. Mededelingen van het Instituut voor Natuur- en Bosonderzoek INBO.M.2009.6, Brussel, 669 p.

Pagina | 38
Pfalzer, G. & Kusch, J. (2003). Structure and variability of bat social calls: implications for specificity and individual recognition. Journal of Zoology 261: 21-33.
Philippart F. (2011). Antwerpen: De commandobunkers van de vliegvelden van Deurne & Wilrijk, Dossier Simon Stevin Vlaams Vestingbouwkundig Centrum.
Prims, Fl. (1952). Geschiedenis van Wilrijk. Uitgave van het gemeentebestuur van Wilrijk. 410 p.
Robinson M.F., & Stebbings R.E. (1997). Home range and habitat use by the serotine bat, Eptesicus serotinus, in England. Journal of Zoology 243: 117-136.
Russ J. (2012). British bat calls: a guide to species identification. Pelagic Publishing, UK. 192p.
Russ J.M. & Montgomery W.I. (2002). Habitat associations of bats in Northern Ireland: implications for conservation. Biological Conservation 108, 49-58.
Russ, J.M., O’Neill, J.K. & Montgomery, W.I. (1998). Nathusius’ bats (Pipistrellus nathusii, Keyserling & Blasius 1839) breeding in Ireland. Journal of Zoology 245: 345 – 349.
Russo D. & Jones G., (2003). Use of foraging habitats by bats in a Mediterranean area determined by acoustic surveys: conservation implications. Ecography 26: 197-209.
Rydell, J., Entwistle, A. & Racey, P.A. (1996). Timing of Foraging Flights of Three Species of Bats in Relation to Insect Activity and Predation Risk. Oikos Vol. 76, No. 2 pp. 243-252
Sachteleben J. & von Helversen, O. (2006). Songflight behaviour and mating system of the pipistrelle bat (Pipistrellus pipistrellus) in an urban habitat. Acta Chiropterologica 8(2):391-401
Schober, W., Grimmberger, E. & Lina, P. (vert. en bewerk.). (2001). Gids van de vleermuizen van Europa, Azoren en Canarische Eilanden: met specifieke informatie over de vleermuizen in Nederland en België. Tirion, 263 p.
Sendor, T. (2002). Population ecology of the pipistrelle bat (Pipistrellus pipistrellus Schreber, 1774): the significance of the year-round use of hibernacula for life histories. Dissertation zur Erlangung des Doktorgrades der Naturwissenschaften. 146p.
Simon, M., Hüttenbügel, S. & Smit-Viergutz, Janna (2004). Ecology and conservation of Bats in villages and towns. Bundesamt für Naturschutz: Schriftenreihe für landschaftsplege und Naturschuz Heft 77. Bonn – Bad Godesberg. 263p.
Thomaes, A. & Hofman, M. (2009). Landschapsgebruik door vleermuizen in de omgeving van het Fort van Haasdonk en de Barbierbeek. Zwijndrecht, Natuurpunt-WAL en Natuurpunt-Studie.
Twisk, P., van Diepenbeek, A. & Bekker, J. P. (2010). Veldgids Europese Zoogdieren (deel vleermuizen en redactie). Uitgave KNNV-uitgeverij.
Van der Wijden, B. & Verkem, S. (2004). Chiropterologisch onderzoek in het kader van het onderzoek naar de haalbaarheid van het natuurinrichtingsproject Zoerselbos. Bestek nr. AMINAL/NATA/VL01 (26/03/2003). A.B.Consultancy g.c.v., Dendermonde, 117p.
Vaughan N., Jones G. & Harris S., (1997). Habitat use by bats (Chiroptera) assessed by means of a broad-band acoustic method. Journal of Applied Ecology 34, 716-730
Verbeylen G. & Lefevre A. (2010). Onderzoek naar foerageergebieden, vliegroutes, kraamkolonies en zomerverblijfplaatsen van vleermuizen in Fort 8 en omgeving. Rapport Natuur.studie 2010/6, Natuurpunt Studie (Vleermuizenwerkgroep) i.o.v. Soresma i.k.v. Masterplan Stad Antwerpen, Mechelen, België.
Verboom B., & Huitema H. (1997). The importance of linear landscape elements for the pipistrelle Pipistrellus pipistrellus and the serotine Eptesicus serotinus. Landscape Ecology 12 (2): 117-125.
Verboom B., & Huitema H. (2010). The Influence of treeline structure and wind protection on commuting and foraging pipistrelles (Pipistrellus pipistrellus). Lutra 53 (2): 63 - 80

Pagina | 39
Verkem, S. & Verhagen, R. (1998). Bescherming vleermuizen. Rapport AMINAL/afdeling Natuur/ 1995/nr 8.
Verkem, S., De Maeseneer, J. Vandendriessche, B., Verbeylen, G. & Yskout, S. (2003). Zoogdieren in Vlaanderen. Ecologie en verspreiding van 1987 tot 2002. Natuurpunt Studie & JNM-zoogdierenwerkgroep, Mechelen & Gent, België.
Verkem S. (2008). Verslag fortentelweekend 1-2-3 februari 2008. Natuurpunt Werkgroep Vleermuizen. 9p.
Verkem S. (2009). Verslag fortentelweekend 6-7-8 februari 2009. Natuurpunt Werkgroep Vleermuizen. 10p
Wickramasinghe, L. P., Harris, S., Jones G. & Vaughan, N. (2003). Bat activity and species richness on organic and conventional farms: impact of agricultural intensification. Journal of Applied Ecology 40: 984–993
Willems, W., Boers, K., Vandendriesche, B., Vanreusel, W. & Lefevre, A. (2009). Monitoring handleiding Natuurpunt Module F10 – Wintertelling vleermuizen. 6 p.
Willems W., Verbeylen G. & Lefevre A. (2011). Onderzoek naar overwintering en zwermgedrag van vleermuizen in Fort 8. Rapport Natuur.studie 2011/5, Natuurpunt Studie (Vleermuizenwerkgroep) i.o.v. Soresma i.k.v. Masterplan Stad Antwerpen, Mechelen, België.
Wood, S. (2014) mgcv: Mixed GAM Computation Vehicle with GCV/AIC/REML smoothness estimation. R Package version 1.7.28.
Zoogdiervereniging & Probos. (2012). Laanbeheer en vleermuizen; met oog voor veiligheid en cultuurhistorie; met bijdragen van: E.A. Jansen, M.H.A. van Benthem, C. de Groot, P. Twisk & H.J.G.A. Limpens, Rapport 2012.10. Zoogdiervereniging & Probos, Nijmegen/Wageningen. 69p.
Zukal J., & Řehák, Z. (2006) Flight activity and habitat preference of bats in a karstic area, as revealed by bat detectors. Folia Zoologica 55(3): 273–281
Zuur A.F., I. E. N., Walker N.J., Saveliev A.A. & Smith G.M. (2009) Mixed effect models and extensions in ecology with R. Springer, New York, U.S.A.

Pagina | 40
10. Bijlagen
10.1. Overzicht van tel-data, tijdstippen en weersomstandigheden
Tabel 3. Overzicht van de waarnemingsdata, locatie, start- en einduur, start-temperatuur en weersomstandigheden in 2011.
Datum Locatie Start Einde Start
temp. Weer
Kraamperiode
18/05/2011 Klaverblad 22:00 0:15 15 °C Gedeeltelijk bewolkt
23/05/2011 Kleine Struisbeek 22:00 23:24 16 °C Gedeeltelijk bewolkt
13/06/2011 Klaverblad 22:34 0:19 18 °C bewolkt, winderig
15/06/2011 Kleine Struisbeek 22:55 00:23 18 °C open, drukkend,
maansverduistering
Lactatie 25/07/2011 Klaverblad 22:05 0:20 19,5 °C bewolkt, winderig
27/07/2011 Kleine Struisbeek 22:00 23:38 21 °C bewolkt, lichte wind
Post-lactatie
22/08/2011 Klaverblad 21:23 23:30 19 °C bewolkt, winderig, regen (laatste 10 minuten)
24/08/2011 Kleine Struisbeek 21:24 23:16 18 °C licht bewolkt tot open, licht wind
14/09/2011 Klaverblad 20:30 23:19 22 °C bewolkt en wisselvallig, kort en hevige regenbuien, lichte wind
21/09/2011 Kleine Struisbeek 20:30 22:12 17 °C bewolkt, winderig
10/10/2011 Klaverblad 19:23 21:39 21 °C bewolkt, winderig
12/10/2011 Kleine Struisbeek 19:45 22:15 20 °C licht bewolkt, windstil tot matige wind, later op de avond regen

Pagina | 41
Tabel 4. Overzicht van de waarnemingsdata, locatie, start- en einduur, start-temperatuur en weersomstandigheden in 2012.
Datum Locatie Start Einde Start
temp. Weer
Kraamperiode
3/05/2012 Klaverblad 21:40 0:03 13 °C bewolkt, lichte wind
4/05/2012 Kleine Struisbeek 21:46 23:10 11 °C bewolkt, koude wind
23/05/2012 Klaverblad 22:15 0:30 20 °C half open, half sluierbewolking
24/05/2012 Kleine Struisbeek 22:19 23:53 25 °C open, zwoel, hoge luchtvochtigheid
Lactatie
27/06/2012 Klaverblad 22:32 0:42 20,5 °C drukkend, bewolkt, af en toe een druppel
28/06/2012 Kleine Struisbeek 22:37 0:24 27 °C
afwisselend open en bewolkt, drukkend, later op de avond wind opkomend
25/07/2012 Klaverblad 22:20 0:38 23,5 °C open hemel, zachte wind, halve maan
26/07/2012 Kleine Struisbeek 22:31 0:12 26 °C open hemel, drukkend, windstil, halve maan
Post-lactatie
23/08/2012 Klaverblad 21:40 23:00 20,5 °C bewolkt, matige tot sterke wind, regen
25/08/2012 Kleine Struisbeek 22:07 23:38 19,5 °C bewolkt, winderig, halve maan
26/08/2012 Klaverblad 21:20 23:34 16,5 °C bewolkt, half open, windstil, lichte mist
26/09/2012 Kleine Struisbeek 20:58 22:37 14,5 °C open en windering, bewolkt tegen einde
1/10/2012 Klaverblad 19:48 22:05 18 °C bewolkt en winderig, af en toe regen
25/10/2012 Klaverblad 19:24 21:54 12,5 °C bewolkt, koude, felle wind
26/10/2012 Kleine Struisbeek 19:25 20:43 7,5°C bewolkt, koude, felle wind

Pagina | 42
Het jaar 2011 was het warmste jaar te Brussel-Ukkel sinds het begin van de regelmatige meteorologische waarnemingen in 1833. De gemiddelde jaartemperatuur bereikte 11,6°C, dit was 1,1°C boven de normale waarde (10,5°C). De gemiddelde maandtemperaturen waren hoger dan de normale behalve tijdens de maand juli en augustus. De gemiddelde temperatuur van de zomer was lager dan de zomerse normale. De gemiddelde temperaturen van de winter, de lente en de herfst waren respectievelijk 1,5°C, 2,1°C en 1,5°C hoger dan de seizoensnormalen, terwijl de zomertemperatuur 0,8°C lager was dan de normale. De lente 2011 (maart tot mei) was bijzonder opvallend. Te Ukkel waren de temperaturen zeer uitzonderlijk hoog; de gemiddelde temperatuur bereikte 12,2°C (normale: 10,1°C). Tijdens het gehele lenteseizoen waren de neerslaghoeveelheden alsook het aantal neerslagdagen bijzonder laag. Het neerslagtotaal zorgde ervoor dat dit de 3de droogste lente was te Brussel – Ukkel sinds 1833. De zomer 2011 (juni tot augustus) kende te Ukkel een zeer uitzonderlijk aantal neerslagdagen (61 neerslagdagen). De hoge frequentie van het aantal neerslagdagen is gecorreleerd met een zeer abnormaal hoge waarde van het neerslagtotaal tijdens de zomer: 317,2 mm werd te Ukkel gemeten (normale: 224,6 mm). De hoge frequentie van het aantal neerslagdagen verklaart dat de zonneschijnduur abnormaal laag was over de ganse zomer met een zonneschijnduur van 465 h 40 min (normale: 578 h 20 min). Deze omstandigheden verklaren eveneens dat de gemiddelde zomertemperatuur lager was dan normaal; hij bedroeg 16,7°C, voor een normale van 17,5°C. De herfst 2011 (september tot november) was tegengesteld aan de zomer. Te Ukkel was het aantal neerslagdagen zeer uitzonderlijk laag, slechts 37 dagen (normale: 51 d.). Gelijkaardig gecorreleerd, observeerden wij een abnormaal lage neerslaghoeveelheid; wij hebben een seizoenstotaal van 140,4 mm (normale: 219,9 mm) gemeten. Het teveel aan zonneschijn was ook opvallend met een totaal van 450 h 02 zonneschijn (normale: 322 h 00 min), waardoor deze herfst de 5de zonnigste werd sinds 1887. De herfst was uitzonderlijk warm, met een gemiddelde temperatuur van 12,4 °C (normale: 10,9 °C).
Het jaar 2012 was in Ukkel abnormaal nat terwijl het aantal neerslagdagen normaal was. Ook de temperaturen en de zonneschijnduur waren normaal. De gemiddelde luchttemperatuur te Ukkel bedroeg in 2012 10,6 °C (ref. periode 1981-2010: 10,5 °C). De gemiddelde temperatuur van de meeste kalendermaanden afzonderlijk was vrijwel normaal. De maanden februari en april waren zeer abnormaal koud terwijl maart zeer abnormaal warm was. De lente was thermisch vrijwel normaal: de gemiddelde temperatuur bedroeg 10,5 °C (ref. periode 1981-2010: 10,1 °C). De lente begon warm, in de eerste lentemaand kregen we een lang stabiel, zonnig, droog en vrij warm tijdvak. In de tweede maandhelft overschreed het maximum reeds 3 keer 20 °C te Ukkel: de maxima van 15, 22 en 23 maart bedroegen respectievelijk 20,3 °C, 20,6 °C en 20,6 °C. Vooral de laatste decade was zeer zacht met een gemiddelde van 11,4 °C (normaal: 7,9 °C). Geen enkele decade in april gaf een hoger gemiddelde dan dat van de laatste decade van maart. April verliep immers nogal fris en erg wisselvallig dit jaar, zeker gedurende de eerste 20 dagen waren de temperaturen ondermaats. Het gemiddelde van de eerste 2 decades bedroeg te Ukkel 7,2 °C (normaal: 9,0 °C). Het zonnig karakter van het weer in een groot deel van de maand maart zorgde ervoor dat de dagtemperaturen soms al behoorlijk konden oplopen, de wisselvalligheid en het relatief sombere weer in april hielden de dagtemperaturen aan de lage kant, dit had als gevolg dat april frisser verliep dan maart, de gemiddelde temperatuur voor maart en april was respectievelijk 8,9 °C en 8,4 °C. Het gemiddeld maximum van de maand maart was met 13,3 °C zeer uitzonderlijk hoog (ref. periode 1981-2010: 10,4 °C), het was de hoogste waarde sinds tenminste 1901, het vorig record dateert uit maart 1991 met 13,1 °C. Het gemiddeld maximum van april 2012 bedroeg 12,5 °C, het gemiddeld minimum van maart en april was respectievelijk 4,6 °C en 4,4 °C. De maand mei was thermisch vrijwel normaal, in het begin van de maand was het nog erg

Pagina | 43
wisselvallig en dit weerspiegelde zich tevens in het temperatuurverloop. Doordat we afwisselend in een frissere en warme luchtsoort terechtkwamen, kregen we enkele keren belangrijke temperatuurschommelingen in relatief korte periodes. Aan het einde van de maand werd het een aantal dagen stabiel en warm weer met de eerste zomerse dagen in het binnenland.
Ook de zomer was normaal met een gemiddelde temperatuur van 17,3 °C (ref. periode 1981-2010: 17,5 °C). Juni en de eerste 20 dagen van juli verliepen wat frisser dan gemiddeld, nadien brak een stabielere en warmere periode aan, de warme augustusmaand heeft de wat frisse juni- en julimaand gecompenseerd, in de tweede helft van augustus werd het gedurende een paar dagen zelfs zeer warm.
De herfstmaanden kenden geen thermische uitschieters. Het begin van de maand september was zonnig en vrij warm, terwijl de laatste 10 dagen ietwat aan de koele kant waren. 9 September was de warmste dag van de herfst met waarden op de meeste plaatsen ruim boven 25 °C. Tussen 10 september en 15 oktober lagen de temperaturen meestal op een normaal niveau voor de tijd van het jaar. Vermeldenswaardig is wel het temperatuurverloop in de laatste decade van de maand oktober. Tussen 20 en 25 oktober kregen wij zeer warme zuidelijke luchtstromingen met temperaturen die overdag ruim boven 20 °C uitkwamen, op enkele dagen haalden we zelfs lokaal 23 °C of 24 °C (Ukkel op 22 oktober: 24,5 °C, een nooit gezien maximum in die periode van het jaar). Op 26 oktober kwamen we terecht in koele polaire lucht zodat de temperaturen een stuk onder 10 °C bleven, in de nacht van 27 op 28 oktober vroor het in een groot deel van het land, in Ukkel bereikte het minimum -2,1 °C. (bron www.meteobelgie.be)

Pagina | 44
10.2. Analyse geluidsopnamen
Fig 6. Sonogram grootoorvleermuis op meetpunt KL11 (25 juli 2012)

Pagina | 45
Fig 7. Sonogram ruige dwergvleermuis op meetpunt GS05 (26 september 2012)

Pagina | 46
Fig 8. Sonogram laatvlieger tussen meetpunt KL03 en KL04 (23 augustus 2012)

Pagina | 47
Fig 9. Sonogram gewone dwergvleermuis (inclusief sociaal geluid), ruige dwergvleermuis en watervleermuis op meetpunt GS05 (25 augustus 2012)

Pagina | 48
Fig 10. Sonogram baardvleermuis op meetpunt GS15 (25 oktober 2012)

Pagina | 49
10.3. Kaarten
Waargenomen vlieg- en jachtactiviteit worden per datum weergegeven op de topografische kaart NGI 1/10.000. Vliegactiviteit van gewone dwergvleermuis op de meetpunten wordt op afzonderlijke kaart weergegeven omwille van leesbaarheid.
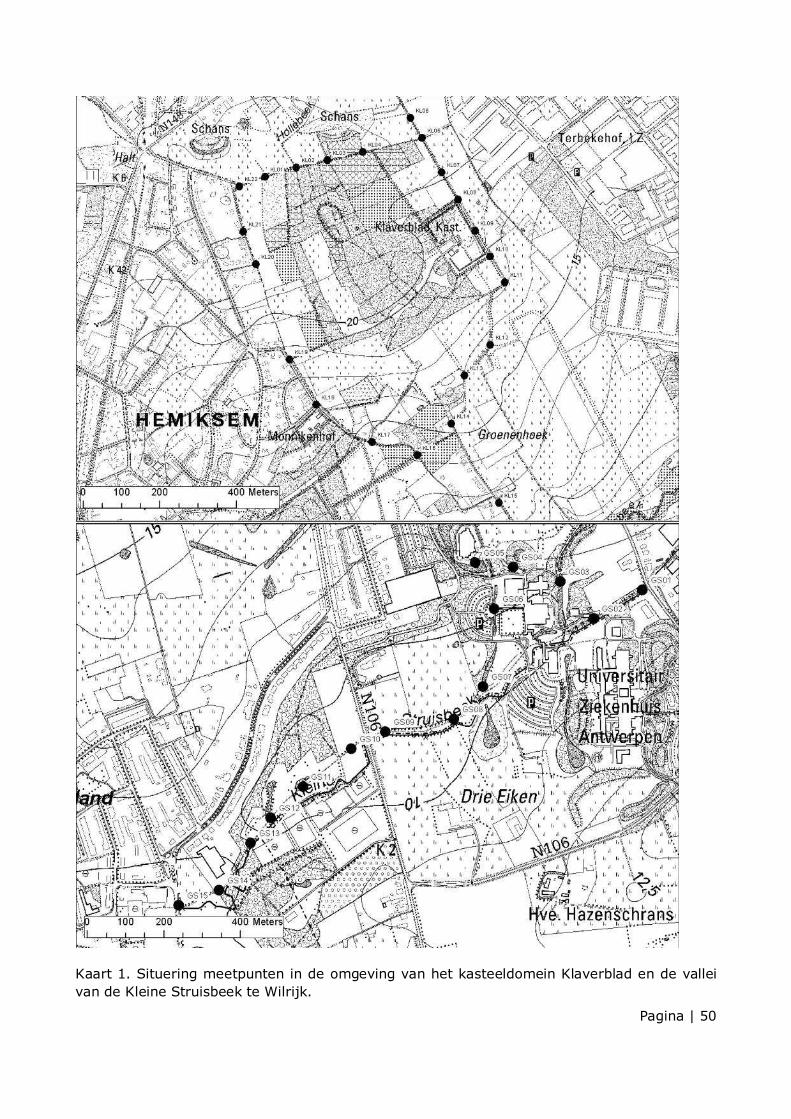
Pagina | 50
Kaart 1. Situering meetpunten in de omgeving van het kasteeldomein Klaverblad en de vallei van de Kleine Struisbeek te Wilrijk.

Pagina | 51
Kaart 2. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van het kasteeldomein Klaverblad op 18 mei 2011.

Pagina | 52
Kaart 3. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van het kasteeldomein Klaverblad op 13 juni 2011.

Pagina | 53
Kaart 4. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van het kasteeldomein Klaverblad op 25 juli 2011.

Pagina | 54
Kaart 5. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van het kasteeldomein Klaverblad op 22 augustus 2011.

Pagina | 55
Kaart 6. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van het kasteeldomein Klaverblad op 14 september 2011.

Pagina | 56
Kaart 7. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van het kasteeldomein Klaverblad op 10 oktober 2011.

Pagina | 57
Kaart 8. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van het kasteeldomein Klaverblad op 3 mei 2012.

Pagina | 58
Kaart 9. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van het kasteeldomein Klaverblad op 23 mei 2012.

Pagina | 59
Kaart 10. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van het kasteeldomein Klaverblad op 27 juni 2012.

Pagina | 60
Kaart 11. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van het kasteeldomein Klaverblad op 25 juli 2012.

Pagina | 61
Kaart 12. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van het kasteeldomein Klaverblad op 23 augustus 2012.

Pagina | 62
Kaart 13. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van het kasteeldomein Klaverblad op 26 augustus 2012.

Pagina | 63
Kaart 14. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van het kasteeldomein Klaverblad op 1 oktober 2012.

Pagina | 64
Kaart 15. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van het kasteeldomein Klaverblad op 25 oktober 2012.

Pagina | 65
Kaart 16. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van de Kleine Struisbeek op 23 mei 2011.

Pagina | 66
Kaart 17. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van de Kleine Struisbeek op 15 juni 2011.

Pagina | 67
Kaart 18. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van de Kleine Struisbeek op 27 juli 2011.

Pagina | 68
Kaart 19. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van de Kleine Struisbeek op 24 augustus 2011.

Pagina | 69
Kaart 20. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van de Kleine Struisbeek op 21 september 2011.

Pagina | 70
Kaart 21: Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van de Kleine Struisbeek op 12 oktober 2011.
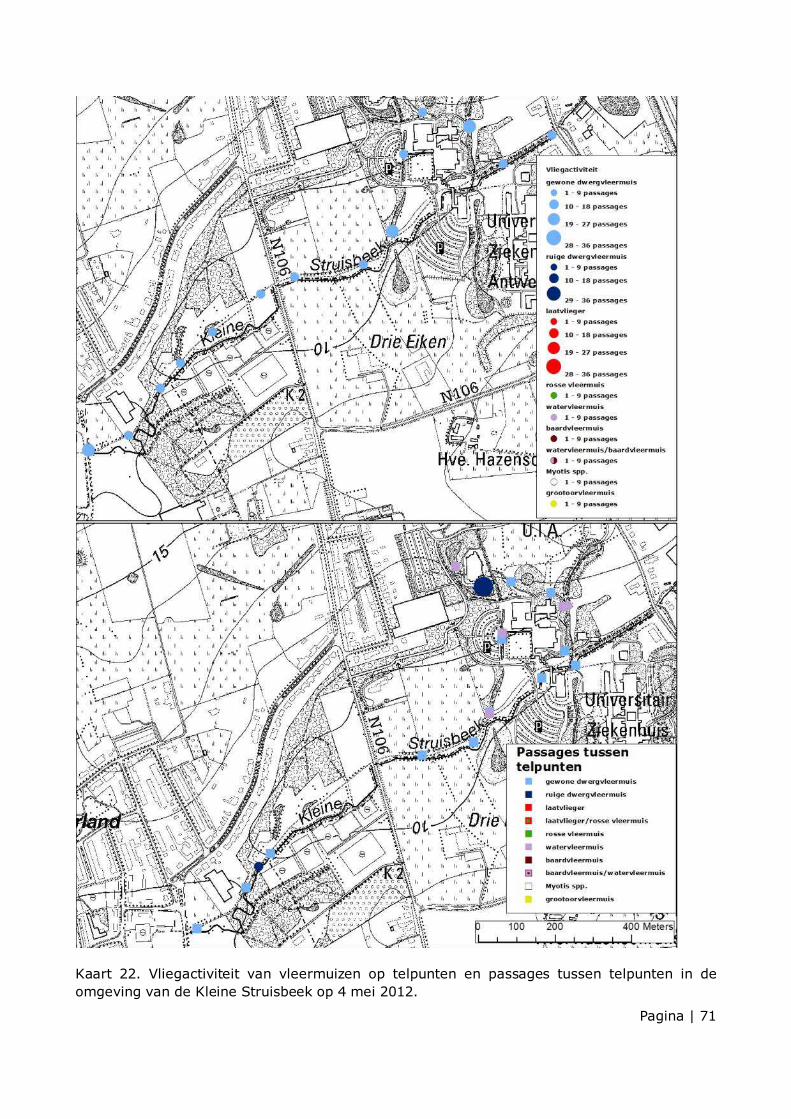
Pagina | 71
Kaart 22. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van de Kleine Struisbeek op 4 mei 2012.

Pagina | 72
Kaart 23. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van de Kleine Struisbeek op 24 mei 2012.

Pagina | 73
Kaart 24. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van de Kleine Struisbeek op 28 juni 2012.

Pagina | 74
Kaart 25. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van de Kleine Struisbeek op 26 juli 2012.

Pagina | 75
Kaart 26. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van de Kleine Struisbeek op 25 augustus 2012.

Pagina | 76
Kaart 27. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van de Kleine Struisbeek op 26 september 2012.

Pagina | 77
Kaart 28. Vliegactiviteit van vleermuizen op telpunten en passages tussen telpunten in de omgeving van de Kleine Struisbeek op 26 oktober 2012.

Pagina | 78
Kaart 29. Jachtactiviteit van gewone dwergvleermuis op telpunten in de omgeving van het kasteeldomein Klaverblad op 3 mei 2012 en langs de Kleine Struisbeek op 4 mei 2012.

Pagina | 79
Kaart 30. Jachtactiviteit van gewone dwergvleermuis op telpunten in de omgeving van het kasteeldomein Klaverblad op 23 mei 2012 en langs de Kleine Struisbeek op 24 mei 2012.

Pagina | 80
Kaart 31. Jachtactiviteit van gewone dwergvleermuis op telpunten in de omgeving van het kasteeldomein Klaverblad op 27 juni 2012 en langs de Kleine Struisbeek op 28 juni 2012.

Pagina | 81
Kaart 32. Jachtactiviteit van gewone dwergvleermuis op telpunten in de omgeving van het kasteeldomein Klaverblad op 25 juli 2012 en langs de Kleine Struisbeek op 26 juli 2012.

Pagina | 82
Kaart 33. Jachtactiviteit van gewone dwergvleermuis op telpunten in de omgeving van het kasteeldomein Klaverblad op 23 en 26 augustus 2012 en langs de Kleine Struisbeek op 25 augustus 2012.

Pagina | 83
Kaart 34. Jachtactiviteit van gewone dwergvleermuis op telpunten in de omgeving van het kasteeldomein Klaverblad op 1 oktober 2012 en langs de Kleine Struisbeek op 26 september 2012.

Pagina | 84

Pagina | 85
Kaart 35. Jachtactiviteit van gewone dwergvleermuis op telpunten in de omgeving van het kasteeldomein Klaverblad op 25 oktober 2012 en langs de Kleine Struisbeek op 26 oktober 2012.

Pagina | 86
Kaart 36. Overzicht vleermuizenfuncties in de omgeving van de het kasteeldomein Klaverblad (2011 – 2012).

Pagina | 87
Kaart 37. Overzicht vleermuizenfuncties in de omgeving van de Kleine Struisbeek (2011 – 2012

Pagina | 88
10.4. Beeldmateriaal
Foto 1. Moerelei ter hoogte van telpunt KL01 (Johan De Ridder, 12 januari 2013)
Foto 2. Moerelei ter hoogte van telpunt KL03 (Johan De Ridder, 10 november 2011)

Pagina | 89
Foto 3. Klaverbladdreef ter hoogte van KL05 (Johan De Ridder, 12 januari 2013)
Foto 4. Jachtgebied gewone dwergvleermuis ter hoogte van KL05 (Johan De Ridder, 12 januari 2013)

Pagina | 90
Foto 5. Klaverbladdreef met zicht op KL06 (Johan De Ridder, 12 januari 2013)
Foto 6. Kasteeldreef ter hoogte van KL08 (Johan De Ridder, 3 november 2007)

Pagina | 91
Foto 7. Kasteeldreef ter hoogte van KL10 (Johan De Ridder, 3 november 2007)
Foto 8. Groenenhoek ter hoogte van KL11 (Johan De Ridder, 3 november 2007)

Pagina | 92
Foto 9. Groenenhoek aan telpunt KL012 (Johan De Ridder, 10 november 2011)
Foto 10. Dreefrelict ter hoogte van telpunt KL013 (Johan De Ridder, 10 november 2011)

Pagina | 93
Foto 11. Groenenhoek aan telpunt KL014 (Johan De Ridder, 10 november 2011)
Foto 12. Groenenhoek aan telpunt KL16 (Johan De Ridder, 10 november 2011)

Pagina | 94
Foto 13. Groenenhoek tussen telpunt KL16 en KL17 (Johan De Ridder, 15 april 2006)
Foto 14. Jachtgebied laatvlieger ter hoogte van KL19 (Johan De Ridder, 10 november 2011)

Pagina | 95
Foto 15. Valkenisseweg aan telpunt KL019 (Johan De Ridder, 10 november 2011)
Foto 16. Leemweg met zicht op KL20 (Johan De Ridder, 10 november 2011)

Pagina | 96
Foto 17. Traditorebatterij schans 16 (Johan De Ridder, 10 november 2011)
Foto 18. Gallerij traditorebatterij schans 16 (Johan De Ridder, 3 januari 2006)

Pagina | 97
Foto 19. Traditorebatterij schans 17 (Johan De Ridder, 11 augustus 2004)
Foto 20. Controletoren schijnvliegveld van Wilrijk (Johan De Ridder, 3 november 2007)

Pagina | 98
Foto 21. Geitenpad ter hoogte van telpunt GS15 (Johan De Ridder, 2 april 2013)
Foto 22. Kleine Struisbeek ter hoogte van telpunt GS15 (Johan De Ridder, 2 april 2013)

Pagina | 99
Foto 23. Geitenpad ter hoogte van telpunt GS14 (Johan De Ridder, 2 april 2013)
Foto 24. Geitenpad ter hoogte van telpunt GS13 (Johan De Ridder, 2 april 2013)

Pagina | 100
Foto 25. Vijver langsheen het Geitenpad ter hoogte van telpunt GS12 (Johan De Ridder,
3 maart 2013)
Foto 26. Geitenpad ter hoogte van telpunt GS11 (Johan De Ridder, 2 april 2013)

Pagina | 101
Foto 27. Kleine Struisbeek ter hoogte van telpunt GS09 (Johan De Ridder, 28 december
2005)
Foto 28. Kleine Struisbeek segment tussen telpunt GS09 en GS08 (Johan De Ridder, 2
april 2013)

Pagina | 102
Foto 29. Geitenpad tussen telpunt GS09 en GS08 (Johan De Ridder, 2 april 2013)
Foto 30. Vijver op de campus Drie Eiken ter hoogte van telpunt GS07 (Johan De Ridder,
2 april 2013)

Pagina | 103
Foto 31: Geitenpad tussen telpunt GS07 en GS06 (Johan De Ridder, 2 april 2013)
Foto 32: Vliegroute watervleermuis ter hoogte van telpunt GS06 (Johan De Ridder, 2
april 2013)

Pagina | 104
Foto 33: Jachtgebied watervleermuis ter hoogte van telpunt GS03 (Johan De Ridder, 2
april 2013)
Foto 34: Geitenpad ter hoogte van telpunt GS02 (Johan De Ridder, 2 april 2013)

Pagina | 105
Foto 35: Geitenpad ter hoogte van telpunt GS01 (Johan De Ridder, 2 april 2013)
Copyright © 2022 FDOKUMEN




















