
Uroguanylin inhibits H-ATPase activity and surface expression in renal distal tubules by a...
43
1 Uroguanylin inhibits H-ATPase activity and surface expression in 1 renal distal tubules by a PKG dependent pathway 2 3 1 Vanessa da Silva Lima, 2 Renato O. Crajoinas, 3 Luciene R. Carraro-Lacroix, 4 1 Alana N. Godinho, 1 João L. G. Dias, 2 Rafael Dariolli, 2 Adriana C. C. Girardi, 5 1,4 Manassés C. Fonteles, 3 Gerhard Malnic, 1 Lucília M. A. Lessa* 6 7 1 Superior Institute of Biomedical Sciences, State University of Ceará, Fortaleza, 8 CE, Brazil; 2 Heart Institute (InCor), University of São Paulo Medical School, São 9 Paulo, SP, Brazil; 3 Department of Physiology and Biophysics, Biomedical Sciences 10 Institute, University of São Paulo, São Paulo, SP, Brazil; 4 Mackenzie University, 11 São Paulo, SP, Brazil 12 13 Running title: Modulation of H-ATPase by uroguanylin 14 15 *Address correspondence to: 16 Lucília M. A. Lessa 17 Superior Institute of Biomedical Sciences 18 Av. Paranjana, 1700 19 60.714.903 Fortaleza - Ceará, Brazil 20 Phone: 55-85-3101-9836 21 e-mail: [email protected] 22 23 Articles in PresS. Am J Physiol Cell Physiol (July 16, 2014). doi:10.1152/ajpcell.00392.2013 Copyright © 2014 by the American Physiological Society.
Transcript of Uroguanylin inhibits H-ATPase activity and surface expression in renal distal tubules by a...

1
Uroguanylin inhibits H-ATPase activity and surface expression in 1
renal distal tubules by a PKG dependent pathway 2
3
1Vanessa da Silva Lima, 2Renato O. Crajoinas, 3Luciene R. Carraro-Lacroix, 4
1Alana N. Godinho, 1João L. G. Dias, 2Rafael Dariolli, 2Adriana C. C. Girardi, 5
1,4Manassés C. Fonteles, 3Gerhard Malnic, 1Lucília M. A. Lessa* 6
7
1Superior Institute of Biomedical Sciences, State University of Ceará, Fortaleza, 8
CE, Brazil; 2Heart Institute (InCor), University of São Paulo Medical School, São 9
Paulo, SP, Brazil; 3Department of Physiology and Biophysics, Biomedical Sciences 10
Institute, University of São Paulo, São Paulo, SP, Brazil; 4Mackenzie University, 11
São Paulo, SP, Brazil 12
13
Running title: Modulation of H-ATPase by uroguanylin 14
15
*Address correspondence to: 16
Lucília M. A. Lessa 17
Superior Institute of Biomedical Sciences 18
Av. Paranjana, 1700 19
60.714.903 Fortaleza - Ceará, Brazil 20
Phone: 55-85-3101-9836 21
e-mail: [email protected] 22
23
Articles in PresS. Am J Physiol Cell Physiol (July 16, 2014). doi:10.1152/ajpcell.00392.2013
Copyright © 2014 by the American Physiological Society.

2
Abstract 24
Cumulative evidence suggests that guanylin peptides play an important role 25
on electrolyte homeostasis. We have previously reported that uroguanylin (UGN) 26
inhibits bicarbonate reabsorption in renal distal tubule. In the present study, we 27
tested the hypothesis that the bicarbonaturic effect of UGN is at least in part due to 28
inhibition of H+-ATPase-mediated hydrogen secretionin the distal nephron. By in 29
vivo stationary microperfusion experiments we were able to show that UGN inhibits 30
H+-ATPase activity by a PKG dependent pathway, since KT5823 (PKG inhibitor) 31
abolished the UGN effect on distal bicarbonate reabsorption and H89 (PKA 32
inhibitor) was unable to prevent it. The in vivo results were confirmed by the in vitro 33
experiments, where we used fluorescence microscopy to measure intracellular pH 34
(pHi) recovery after an acid pulse with NH4Cl. By this technique we observed that 35
UGN and 8Br-cGMP inhibited H+-ATPase-dependent pHi recovery, and that the 36
UGN inhibitory effect was abolished in the presence of the PKG inhibitor. In 37
addition, by using RT-PCR technique we verified that MDCK-C11 cells express 38
guanylate cyclase-C. Besides, UGN stimulated an increase of both cGMP content 39
and PKG activity, but was unable to increase the production of cellular cAMP 40
content and PKA activity. Furthermore, we found that UGN reduced cell surface 41
abundance of H+-ATPase B1 subunit in MDCK-C11 and that this effect was 42
abolished by the PKG inhibitor. Taken together, our data suggest that UGN inhibits 43
H+-ATPase activity and surface expression in renal distal cells by a cGMP/PKG 44
dependent pathway. 45
46
Keywords: H+-ATPase; Uroguanylin; Renal Microperfusion; Distal tubule; 47
PKG; cGMP 48
49

3
Introduction 50
51
Guanylins comprise a family of peptides that play an important role in the 52
regulation of salt balance, pH regulation, appetite and gut health (28). Among 53
these peptides, uroguanylin (UGN) is the one with more pronounced renal actions 54
(8). 55
In the kidney, UGN modulates the excretion of sodium, potassium, 56
bicarbonate, chloride and water (1, 8, 21, 29). These actions occur by altered 57
tubular reabsorption and tubular secretion of these electrolytes along the nephron 58
without changes in glomerular filtration rate. Besides, most of these effects are 59
related to increase of cGMP levels in urine (8, 11). 60
In a previous study, our group has demonstrated that UGN inhibits 61
bicarbonate reabsorption in proximal and distal tubules (1). In proximal tubules, the 62
UGN effect was attributed to inhibition of NHE3 activity by activation of both PKG 63
and PKA pathways (21). However, in distal tubules, an additional mechanism, 64
other than inhibition of the Na+/H+ exchanger, was suggested for UGN action on 65
bicarbonate reabsorption (1). 66
The vacuolar H+-ATPase (V-ATPase) is the major cellular mechanism of H+ 67
secretion/bicarbonate reabsorption in the alpha-intercalated cells of the distal 68
segments of the nephron, playing an important role in lumen acidification (38). The 69
V-ATPase is a large multi-subunit protein that mediates ATP-driven vectorial H+ 70
transport across cell membranes. Its activity is controlled by a number of different 71
mechanisms, including regulation of the assembly of the V-ATPase via complex or 72
dynamic regulation of its subunit expression in the cell membrane surface (10). 73

4
Among the multiple subunits, B1 seems to occupy a prominent role in the 74
regulation of H+-ATPase. B1 subunit knockout mice develop tubular acidosis, 75
metabolic acidosis, dehydration, and growth retardation (7). In addition, the 76
inactivation of the B1 subunit in intercalated cells leads to type 1 distal renal tubular 77
acidosis (dRTA), a disease associated with salt- and potassium-wasting 78
nephropathy (14). 79
Considering that H+-ATPase in distal segments is constantly submitted to 80
hormonal regulation, such as the renin-angiotensin-aldosterone system (RAS) and 81
natriuretic peptides (25, 26, 40) and that we have previously demonstrated that 82
UGN inhibits hydrogen secretion in renal distal tubule, the purpose of the present 83
study was to test the hypothesis that the UGN inhibitory effect on distal bicarbonate 84
reabsorption might be dependent at least in part on H+-ATPase inhibition. The 85
signaling mechanisms involved in the inhibitory effect of UGN on hydrogen 86
secretion in renal distal tubules were also addressed. The findings of the present 87
study show that UGN inhibits H+-ATPase activity in both rat renal distal tubule and 88
in MDCK-C11 cells. This effect seems to be dependent on a reduction of the 89
surface expression of B1 H+-ATPase subunit. In addition, the current data show 90
that the cGMP/PKG signaling pathway and not cAMP/PKA is involved in the UGN 91
regulation of distal bicarbonate reabsorption. 92
93
94
95

5
Materials and methods 96
Reagents and Antibodies: All chemicals were obtained from Sigma 97
Chemical Co. (St. Louis, MO) unless otherwise noted. Uroguanylin was purchased 98
from Bachem (Philadelphia, PA). KT5823, a specific, inhibitor of protein kinase G 99
(PKG) was purchased from Calbiochem (San Diego, CA). EZ-LinkTM Sulfo-NHS-100
SS-Biotin as well as immunopure immobilized streptavidin were purchased from 101
Thermo Fisher Scientific Inc (Rockford, IL). A monoclonal antibody (mAb) to H+-102
ATPase subunit B1 (V-ATPase B1) was purchased from Santa Cruz Biotechnology 103
Inc. (Santa Cruz, CA). The mAb to actin (JLA20) was purchased from Merck 104
Millipore (Billerica, MA). Horseradish peroxidase-conjugated secondary antibodies 105
were purchased from Life Technologies Corporation (Carlsbad, CA). 106
Animals: Animal procedures and protocols were followed in accordance with 107
the ethical principles in animal research of the Brazilian College of Animal 108
Experimentation and were approved by the Committees of the State University of 109
Ceará and São Paulo. Experiments were performed using male Wistar rats (250 to 110
300g) housed under standardized conditions (constant temperature of 22°C, 12-h 111
dark-light cycle, and relative humidity of 60%) at the Superior Institute of 112
Biomedical Sciences of State University of Ceará and of São Paulo. In order to 113
perform stationary in vivo microperfusion, the animals were anesthetized with 114
intramuscular ketamine (75mg/Kg) and xylazine (8mg/Kg). The left jugular vein 115
was cannulated for infusion of 3% mannitol in isotonic saline at a rate of 116
0.1mL/min. The kidney was exposed by a lumbar approach and prepared for “in 117
vivo” micropuncture. 118

6
Stationary “in vivo” microperfusion: The microperfusion procedure was 119
performed as described previously (1). A proximal tubule was punctured by means 120
of a double-barrelled micropipette, one barrel being used to inject FDC-green 121
colored Ringer perfusion solution (in mM, NaCl, 80; KCl, 5; NaHCO3, 25; CaCl2, 1; 122
MgSO4, 1.2; and raffinose to reach isotonicity, at 0 PCO2), and the other to inject 123
Sudan-black colored castor oil used to block the injected fluid columns in the 124
lumen. A distal segment of the same nephron, recognized by the colored perfusion 125
and by its transepithelial PD (higher than 20 mV, lumen negative), was impaled by 126
a double barrelled asymmetric microelectrode to measure intratubular pH, the 127
larger barrel containing at its tip the H+-ion sensitive ion-exchange resin (Fluka, 128
Buchs, Switzerland) and the smaller, 1 M KCl reference solution colored by FDC-129
green. Properties of the microelectrode were described previously (13). The pH 130
microelectrodes were calibrated before and after every impalement on the kidney’s 131
surface by superfusion with 20mM Phosphate-Ringer buffer solutions containing 132
130mM NaCl, at 37°C. The pH values were adjusted to 6.5, 7.0 and 7.5 with 0.1N 133
NaOH or HCl. A luminal oil block was split by perfusions so that the solution was 134
isolated and blocked by oil. The perfusion rate was sufficient to increase luminal 135
pH to values near those of the perfusion fluid, i.e. 25 mM NaHCO3 or pH ~8. After 136
blocking the luminal solution with oil, the increase in luminal H+ activities, 137
representing bicarbonate reabsorption, were followed until a stable level was 138
reached. 139
By this technique several curves (about 3 to 7) control and experimental 140
were obtained per tubule, the mean of control or experimental curves constituting 141
the values for this tubule. The value of N given for an experimental condition 142

7
corresponds to the number of perfused tubules, approximately 1 to 3 being 143
perfused in one rat, and for each group at least 4 rats were used. Luminal HCO3- 144
activity, initially starting with 25 mM was then progressively reduced to a stationary 145
level (HCO3-s) by H+ secretion. The voltage between the microelectrode barrels, 146
representing the luminal H+ activity, was sampled every 0.5 second by an AD 147
converter (Lynx, São Paulo, Brazil) in a microcomputer. At the same time, the 148
potential difference between the reference barrel and ground (the rat tail) was 149
recorded, giving the evolution of transepithelial PD with time during the perfusion. 150
Luminal bicarbonate was calculated from luminal pH and blood PCO2 measured by 151
a Severinghaus electrode. The rate of tubular acidification was expressed as the 152
half-time of the exponential reduction of the injected HCO3- concentration to its 153
stationary level (t1/2). Net HCO3- reabsorption (JHCO3
-) per cm2 of tubule 154
epithelium was calculated from the equation: 155
2)(2ln
332
13
rHCOHCOt
J soHCO ∗−= −−− 156
where t/2 is the half-time of bicarbonate reabsorption, r is the tubule internal radius 157
measured by an ocular micrometer, and [HCO3-]o and [HCO3
-]s are the 158
concentrations of the injected HCO3- and HCO3
- at the stationary level, 159
respectively. The tubules were perfused with control solution, 10-6 M Uroguanylin 160
(UGN) and/or the specific NHE inhibitor, 10-4M HMA; H+-ATPase inhibitor, 10-7M 161
Concanamycin A. In the experiments designed to investigate the signaling 162
mechanisms of UGN action we used 10-6 M KT5823 (PKG inhibitor); 10-5 M H89 163
(PKA inhibitor). 164

8
Cell culture: MDCK-C11 cells were obtained from Dr. Hans Oberleitner and 165
used from passages (80-90). Serial cultures were maintained in DMEM low 166
glucose supplemented with 5% L-glutamine, 5% sodium piruvate, 10% (vol/vol) 167
heat-inactivated fetal bovine serum, 100 IU/ml penicillin, and 100 µg/ml 168
streptomycin. Cells were grown at 37°C, 95% humidified air-5% CO2 (pH 7.4) in a 169
CO2 incubator (Lab-Line Instruments, Melrose Park, IL). The cells were 170
subcultured with trypsin-EGTA (0.02%) and seeded on tissue culture plates 171
containing sterile glass coverslips (for pH measurements) to become confluent. For 172
all experiments, cells were placed in serum-free medium 24 h before the 173
experiments. 174
Gene expression of GC-C receptor and MDCK-C11 cells: RNA was 175
extracted from MDCK-C11 cells using TRIzol Reagent (Invitrogen, Carlsbad, CA) 176
following the manufacturer's protocol. cDNA synthesis from total RNA (1 µg) was 177
produced by reverse transcription (RT) using the superscript III kit according to the 178
manufacturer’s protocol (Invitrogen). Polymerase chain reaction (PCR) was 179
performed using Taq-polymerase manufacturing protocol (Promega). Briefly, 180
thermal cycling for initial GC-C analysis included a denaturation step at 95 °C for 5 181
min followed by 36 cycles of 95°C for 30 seconds, annealing temperature of 64.5 182
°C for 30 seconds and 72°C for 1 min. 28S gene was used as a housekeeping 183
gene. The initial 28S analysis included a denaturation step at 95 °C for 5 min 184
followed by 36 cycles of 95°C for 30 seconds, annealing temperature of 60 °C for 185
30 seconds and 72°C for 1 min. Primer sequences and expected product lengths 186
were: GC-C receptor (XM_543798.2) primer: forward 5'– 187
AACCATTGGCGATGCCTACA -3' and reverse 5'- AGTTGGCGAGCATGTCAGAA 188

9
-3'; 491bp and 28S primer: forward 5'– TCATCAGACCCCAGAAAAGG -3' and 189
reverse 5'- GATTCGGCAGGTGAGTTGTT -3'; 102bp. PCR products were 190
resolved by electrophoresis at 100 V through 2% agarose gel and visualized with 191
GelRed™ (Biotium). 192
Fluorescence microscopy: Intracellular pH was measured 193
spectrofluorimetrically at 37°C with the fluorescent pH-sensitive probe BCECF-AM. 194
Cells grown to confluence on glass coverslips were loaded with the dye by 195
exposure for 5 min to 12 μM BCECF-AM in the control solution (Table 1). The 196
acetoxymethyl ester form of BCECF enters the cell and is rapidly converted to the 197
anionic-free acid form by intracellular esterases. Following the loading period, the 198
glass coverslips were rinsed with the control solution to remove the BCECF-199
containing solution and placed in a thermoregulated chamber mounted on an 200
inverted epifluorescence microscope (Nikon, TMD). The measured area under the 201
microscope had a diameter of 260 μm and contained on the order of 40 cells. 202
Bathing solutions were rapidly exchanged without disturbing the position of the 203
coverslips. Fluorescence was monitored using 440 (pH insensitive) or 495 nm (pH 204
sensitive) alternately as excitation wavelengths, utilizing a xenon light source. 205
Emission was measured at 530 nm by a photomultiplier-based fluorescence 206
system (Georgia Instruments), at time intervals of 5 s. pHi was calculated from the 207
fluorescence emission ratio of the two excitation wavelengths using a standard 208
calibration procedure based on the use of 10 μM nigericin in high potassium Ringer 209
(Table 1), at pH 6, 7, and 8 (36). 210
Cell pH recovery: Cell pH recovery was evaluated following the acidification 211
of pHi with the NH4Cl pulse technique (2) after a 2-min exposure to 20 mM NH4Cl 212

10
(Table 1), in the absence of external Na+ (Table 1), to inhibit the Na+/H+ exchanger, 213
with or without several inhibitors, as described later. In all the experiments, we 214
calculated the initial rate of pH recovery (dpHi/dt, pH units/min) from the first 2 min 215
after the start of the pHi recovery curve by linear regression analysis. 216
cGMP and cAMP Assay: MDCK-C11 cells were cultured to 100% 217
confluence in 96-well plates. Cells were incubated for 10 min with culture medium 218
containing 1 mM 3-isobutyl-1-methylxanthine (IBMX) or IBMX plus 10-6 M 219
Uroguanilyn or 10-6 M ANP (used as positive control in cGMP assay) or 10-4 M 220
Forskolin (used as positive control in cAMP assay). cGMP and cAMP were 221
measured by using the AmershamTM Enzyme immunoassay BiotrakTM (EIA) 222
System (GE Healthcare) according to specifications of the manufacturer. 223
Determination of PKA Activity in Cell Lysates - MDCK-C11 grown to 224
confluence in 24 well-plates were treated or not with UGN and subsequently 225
solubilized in lysis buffer containing 20 mM MOPS, 50 mM β-glycerol phosphate, 226
50 mM NaF, 1 mM sodium vanadate, 5 mM EGTA, 2 mM EDTA, 1% NP40, 1 mM 227
DTT, 1 mM benzamidine, 1 mM PMSF, 10 µg/mL aprotinin and 10 µg/mL 228
leupeptin. PKA activity was measured in this cell lysate using a non-radioactive 229
PKA Kinase Activity Assay (Enzo Life Sciences, Inc., Farmingdale, NY), according 230
to the manufacturer's instructions. 231
Determination of PKG Activity in Cell Lysates - MDCK-C11 grown to 232
confluency in 24 well-plates were treated or not with UGN and subsequently 233
solubilized in extraction buffer containing 20 mM Tris, pH 7.4, 150 mM NaCl, 1 mM 234
EDTA, 1 mM EGTA, 0.2 mM PMSF, 1 µg/mL pepstatin, 0.5 µg/mL leupeptin, 2 mM 235
NaF, 0.2 mM Na3VO4, 5 mM β-mercaptoethanol. PKG activity was measured in 236

11
this cell lysate using a single-site and semi-quantative CycLex cGK Assay Kit 237
(CycLex Co., Ltd., Nagano, Japan) according to the manufacturer's instructions. 238
Cell surface biotinylation: The assay was performed as described previously 239
(3). Cells were rinsed twice in ice-cold PBS-Ca-Mg (PBS with 0.1 mM CaCl2, 1.0 240
mM MgCl2). Surface membrane proteins were then biotinylated by incubating the 241
cells twice for 25 min with 2mL of ice-cold biotinylation buffer (150 mM NaCl, 10 242
mM triethanolamine, 2 mM CaCl2, and 1.5 mg/mL EZ-Link sulfo-NHS-SS-biotin). 243
Cells were then rinsed twice for 20 min with a quenching buffer (PBS-Ca-Mg, 100 244
mM glycine), washed twice with ice-cold PBS-Ca-Mg and strapped into ice-cold 245
solubilization buffer (50 mM Tris, 150 mM NaCl, 5 mM EDTA, 0.5% sodium 246
deoxycholate, 1% Triton X-100, pH 7.4) containing protease inhibitors (0.7 µg/mL 247
pepstatin A, 0.5 µg/mL leupeptin, and 40 µg/mL PMSF). After lysis on ice for 60 248
min, extracts were centrifuged for 10 min at 14,000 g and 4°C. The protein 249
concentration of the supernatants was measured according to Lowry (22) and 250
equal protein amounts of cell lysate (500 µg) were equilibrated with streptavidin-251
agarose beads at 4°C. Prior to the addition of streptavidin, an aliquot of the 252
supernatant was saved for analysis of total B1- subunit H-ATPase protein 253
expression by immunoblotting. The beads were then washed three times in ice-254
cold solubilization buffer. Biotinylated proteins were released by incubation in 255
Laemmli buffer and subjected to SDS-PAGE and immunoblotting. 256
SDS-PAGE and immunoblotting - Protein samples were solubilized in 257
Laemmli sample buffer and separated by SDS-PAGE using 7.5% polyacrylamide 258
gels. For immunoblotting, proteins were transferred to PVDF (Millipore Immobilon-259
P, Millipore, Bedford, MA) at 350 mA for 8-10 hours at 4ºC with a TE 62 transfer 260

12
electrophoresis unit (GE HealthCare). Sheets of PVDF containing transferred 261
proteins were incubated first in Blotto (5% non-fat dry milk and 0.1% Tween 20 in 262
PBS, pH 7.4) for 1 hour to block nonspecific binding of antibody, followed by 263
overnight incubation in primary antibody diluted in Blotto (1:1000). The sheets were 264
then washed in Blotto and incubated for 1 hour with an appropriate HRP-265
conjugated secondary antibody diluted 1:2000 in Blotto. After washing 5X in Blotto 266
and 2X in PBS (pH 7.4), the sheets were incubated in an enhanced 267
chemiluminescense (ECL) reagent for 1 minute and then placed in a digital 268
imaging system (ImageQuant LAS 4000 mini, GE HealthCare) to visualize the 269
bands. The quantification was realized using ImageJ densitometry software. 270
Statistical analysis: The data were analysed by a Visual Basic program in 271
Excel software. Statistical comparisons were made by the unpaired t-test, taking 272
the probability of 0.05 (5%) as the limit of significance. When more than two groups 273
were compared, one way ANOVA followed by Tukey’s post hoc test, taking 0,05 274
(5%) as limit of significance, was performed. In microperfusion experiments a 275
minimum of 6 tubules were used (n = number of perfused tubules). 276
277
278
279
280
281
282
283
284

13
Results 285
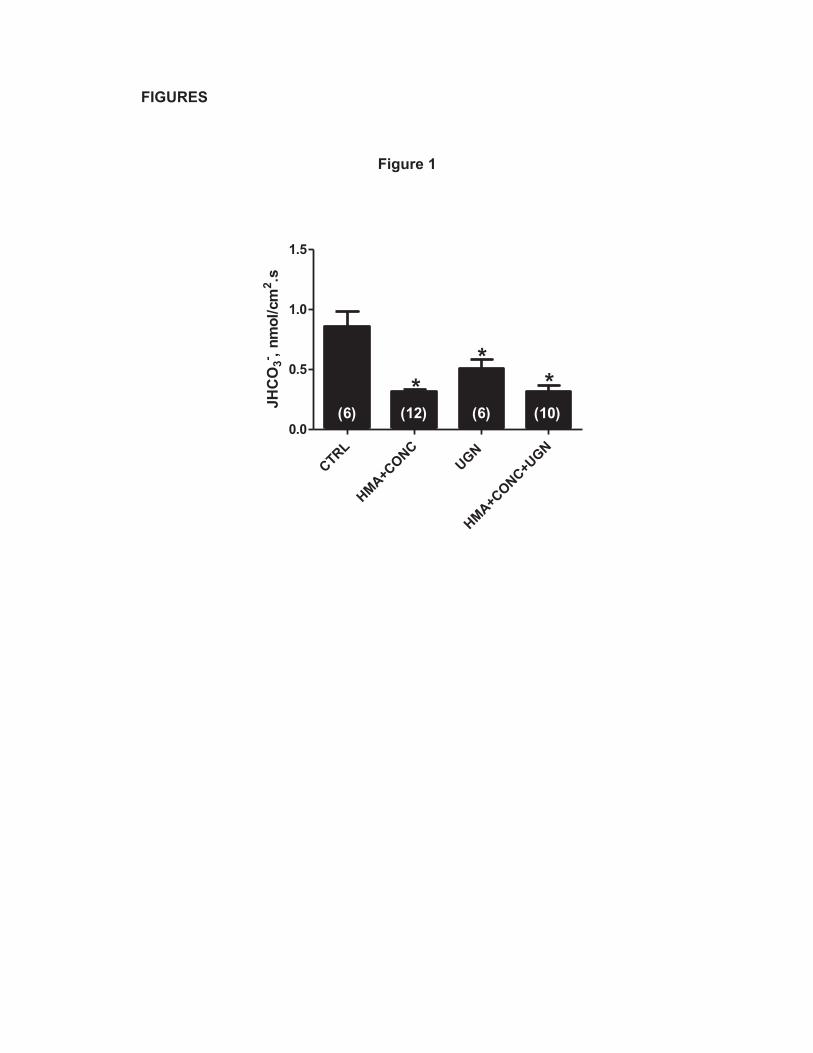
UGN inhibits H+-ATPase-mediated proton secretion in the rat distal tubule. 286
We have previously demonstrated that UGN inhibits distal bicarbonate 287
reabsorption (1) in the presence or absence of hexamethylene amiloride (HMA), a 288
Na+/H+ exchanger (NHE) inhibitor. These previous results suggest that UGN 289
inhibits NHE-dependent and H+-ATPase dependent proton secretion in this 290
nephron segment. To test the hypothesis that UGN inhibits H+-ATPase activity in 291
the distal nephron, we performed “in vivo” stationary microperfusion experiments in 292
which we perfused distal segments of the nephron with UGN alone or together with 293
concanamycin A (CONC), an H+-ATPase inhibitor and/or HMA. The perfusion of 294
CONC together with HMA significantly inhibited the secretion of H+ in rat distal 295
tubules compared to the control solution (Figure 1). Addition of UGN to CONC + 296
HMA also caused a significant inhibition when compared to the control and this 297
inhibition was similar to the one found when we perfused the distal tubules with 298
HMA + CONC (Figure 1). This finding demonstrates that the effect of UGN on 299
distal H+ secretion involves inhibition of no mechanism other than NHE or H+-300
ATPase. 301
302
Signaling mechanisms mediating the effects of UGN on bicarbonate reabsorption 303
in rat renal distal segments 304
Considering that the guanylate cyclase/cGMP/PKG pathway has been 305
described as the classical signaling mechanism for guanylin actions (8), we then 306
examined whether the activation of this signaling pathway mediates the effect of 307
UGN in the distal tubule. We first evaluated the effect of a cGMP analogue, 8Br-308

14
cGMP, on distal bicarbonate reabsorption. As shown in Figure 2A, the perfusion of 309
distal segments with 1µM 8Br-cGMP significantly inhibited the distal hydrogen 310
secretion similarly to UGN. Besides, in the presence of HMA (NHE inhibitor) there 311
was a slight tendency of an additional inhibition of hydrogen secretion by 8Br-312
cGMP, compared to HMA alone, but it was not significant, probably due to the 313
reduced permeability of the cGMP analogue (11). Nevertheless, as illustrated in 314
Figure 3A, there was no significant change in bicarbonate reabsorption when the 315
distal segments were perfused with UGN and the PKG inhibitor, KT5823, when 316
compared to the control or to the group perfused only with KT5823. However, 317
these groups were significantly different from the group perfused only with UGN 318
(Figure 3A). These results indicate that PKG inhibition prevents the UGN inhibitory 319
effect on hydrogen secretion in rat distal tubules, suggesting that the inhibition of 320
bicarbonate reabsorption by UGN is mediated, at least in part, by activation of the 321
cGMP/PKG signaling pathway. 322
In a previous study we showed that the UGN inhibitory effect on proximal 323
bicarbonate reabsorption was dependent on both PKG and PKA activation (21). 324
Thus, we evaluated the possible involvement of PKA on UGN action in distal 325
tubule. As seen in Figure 2B, the cAMP analogue, 8Br-cAMP, produced a 326
tendency to inhibit hydrogen secretion, although this trend did not reach 327
statistical significance. Besides, as illustrated in Figure 3B, the PKA inhibitor, H89, 328
associated with UGN was unable to reverse the inhibitory effect of this peptide on 329
distal hydrogen secretion, suggesting that PKA activation is not involved in the 330
UGN distal effect. 331
UGN inhibits H+-ATPase-dependent pHi recovery in MDCK-C11 cells 332

15
With the purpose to further investigate the inhibitory effect of UGN on H+-333
ATPase activity we performed fluorescence microscopy experiments to investigate 334
the UGN effect on H+-ATPase-dependent intracellular pH recovery in the distal 335
tubule cell line, MDCK-C11, a cell line with H+-secretory alpha intercalated cell 336
characteristics (12). In Figure 4, we can observe the original representative records 337
of pHi recovery experiments after an acid pulse with NH4Cl. As depicted in the 338
figure, there was an important inhibition of the rate of pHi recovery in 0Na+ solution 339
(Figure 4B) comparing to the original record of an experiment in the presence of 340
Na+ (Figure 4A). Under incubation with UGN 1µM, the inhibition of the Na+-341
independent pHi recovery was even more pronouced (Figure 4C). In Figure 5A, the 342
mean values of the pHi recovery rates and hydrogen flux of the groups discussed 343
above are plotted. The hydrogen flux (JH+) was calculated from the following 344
equation: 345
idtdpHiJH β⋅=+ / 346
where dpHi/dt is the rate of pHi recovery after the acid load and βi is the cytosolic 347
buffering capacity averaged for the respective pH range. We compared the rates of 348
H+ extrusion at the average pHi of all experimental groups and the respective 349
average βi of MDCK-C11 cells was used according to Fernandez et al (2000) (6). 350
The mean pHi used in the calculation of H fluxes corresponds to mean values 351
obtained during the pHi recovery period after the ammonium pulse. These data 352
show the significant inhibition of Na+-independent pHi recovery rates promoted by 353
UGN (Figure 5A and B). 354

16
MDCK-C11 cells express three mechanisms related to intracellular pH 355
regulation: NHE1 (isoform 1 of the Na+/H+ exchanger), H+/K+-ATPase and H+-356
ATPase (6, 12). Therefore, to further investigate whether the effect of UGN on Na+-357
independent pHi recovery in MDCK-C11 cells involved the inhibition of H+-ATPase, 358
fluorescence microscopy experiments were performed in the presence of 0Na+ 359
solution, to inhibit Na+-dependent pHi recovery; experiments with EIPA, a NHE1 360
inhibitor and Schering, a H+/K+-ATPase inhibitor, were also performed. In these 361
conditions, incubation of UGN promoted a significant additional inhibition of pHi 362
recovery and hydrogen flux, compared to the group treated with the inhibitors only. 363
These data indicate that UGN inhibits the pHi recovery and H+ flux dependent on 364
H+-ATPase activity (Figure 6A and B). 365
In order to investigate the involvement of cGMP/PKG pathway in this 366
inhibitory effect of UGN, we observed the effect of a cGMP analogue, 8Br-cGMP, 367
on H+-ATPase dependent pHi recovery. Incubation of the cGMP analogue caused 368
a significant inhibition of H+-ATPase activity, in a similar magnitude as did UGN 369
(Figure 5). Moreover, administration of KT5823, a PKG inhibitor, prevented the 370
effect promoted by UGN (Figure 6), demonstrating that inhibition of H+-ATPase by 371
UGN involves PKG activation. 372
373
Effect of UGN on intracellular contents of cGMP and cAMP, PKG and PKA 374
activities and GC-C expression in MDCK–C11 cells 375
Considering the possible involvement of the cGMP/PKG pathway in the 376
inhibitory effect of UGN on H+-ATPase activity (Figure 3A and 6), we evaluated the 377
effect of UGN on the intracellular cGMP concentration and PKG activity in distal 378

17
tubule cells. Figure 7A illustrates that the incubation of MDCK-C11 cells with UGN 379
for 10 minutes promoted a significant increase of intracellular cGMP content. In this 380
experiment, ANP (atrial natriuretic peptide), an agonist of guanylate cyclase – A 381
(GC-A), was used as a positive control, which demonstrated an effect similar to 382
UGN. Furthermore, both UGN and ANP were also able to stimulate a significant 383
increase of PKG activity in MDCK-C11 cells (Figure 7B). Considering these 384
findings we next examined and confirmed that MDCK-C11 cells express the 385
classical guanylin receptor, guanylate cyclase C (GC-C) (Figure 7C). It is worth 386
mentioning that the GC-C-mRNA could only be detected when higher amounts 387
(give the used concentration) of cDNA template were used for amplification 388
(Figure 7C), suggesting that MDCK cells display very low endogenous expression 389
of this receptor. 390
We also evaluated the effect of UGN on intracellular cAMP concentration 391
and PKA activity. Incubation of MDCK-C11 cells with UGN did not change either 392
intracellular cAMP content or PKA activity (Figure 8A and B). In these 393
experiments, forskolin was used as a positive control, and in contrast to the UGN 394
effect, forskolin incubation promoted a significant increase of both intracellular 395
cAMP concentration and PKA activity. 396
397
Effect of UGN on H+-ATPase surface expression in distal tubule cells 398
In order to investigate the mechanism by which UGN inhibits H+-ATPase activity, 399
we next examined whether UGN was capable of decreasing the B1 H+-ATPase 400
subunit cell surface expression in MDCK-C11 cells (Figure 9). The total levels of 401
B1 H+-ATPase subunit (Figure 9A), the β-subunit of H+/K+-ATPase (Figure 9B) and 402

18
actin in 15 min- vehicle, UGN and UGN + KT5823-treated cells were examined to 403
ensure that the yield of protein extracted was constant in each of the experimental 404
conditions. As observed in the representative immunoblotting shown in Figure 9C, 405
the inhibition of H+-ATPase by UGN was accompanied by a reduction of the 406
surface amount of V-ATPase. The peptide induced an inhibition of surface H+-407
ATPase B1 subunit by 54 ± 8%, relative to control. In addition, in the presence of 408
KT5823, a PKG inhibitor, this effect of UGN was abolished. Consistent with the 409
functional findings shown in Figures 1 and 5, UGN did not affect the cell surface 410
expression of the β-subunit of H+/K+-ATPase in MDCK-C11 cells (Figure 9D), 411
suggesting that this peptide does not modulate H/K-ATPase-mediated hydrogen 412
secretion in the distal tubule. 413
414
415

19
Discussion 416
It is now recognized the diuretic, natriuretic and kaliuretic properties of UGN. 417
In addition a potential effect of this peptide on renal tubular acidification has been 418
demonstrated (1, 21, 29). In a previous study, we demonstrated that UGN inhibits 419
bicarbonate reabsorption / hydrogen secretion in rat renal proximal and distal 420
segments (1). Moreover, an additional mechanism, besides inhibition of NHE2, 421
was proposed for the effect of UGN on hydrogen secretion in distal segments, 422
since in the presence of the NHE inhibitor HMA, UGN promoted an additional 423
inhibitory effect on hydrogen secretion (1). 424
Our present findings show that UGN inhibits H+-ATPase activity but not 425
H+/K+-ATPase-mediated hydrogen secretion in the distal nephron cells, both in vivo 426
and in vitro. In microperfusion experiments conducted in the presence of the 427
Na+/H+ exchanger (HMA) and H+-ATPase (concanamicin) inhibitors, no additional 428
inhibitory effect was promoted by UGN. These findings reinforce our previous 429
study, which suggested that the remaining inhibitory effect promoted by UGN on 430
distal bicarbonate reabsorption, in the presence of HMA, involves inhibition of H+-431
ATPase (1). 432
MDCK-C11 cells have properties of intercalated cells which secrete protons 433
and chloride in the collecting duct of mammals (6, 12). Since we have proposed to 434
evaluate H+-ATPase activity, our pHi recovery experiments were undertaken in the 435
presence of NHE1 and H/K-ATPase inhibitors, since these transporters are also 436
constitutively expressed in MDCK-C11 cells (6). Thus, in the presence of their 437
respective inhibitors and 0Na+ solution the remaining hydrogen secretion in MDCK 438
- C11 cells would be performed by H+-ATPase. In these conditions, our findings 439

20
clearly showed that UGN inhibits the Na+- independent pHi recovery exerted by H+-440
ATPase. 441
Several studies indicate an important role for guanylins in pancreatic and 442
intestinal secretion of HCO3- (17, 18, 19, 32). In intestinal cells, guanylin,
443
uroguanylin and the thermo-stable toxin of Escherichia coli (Sta) stimulate anion 444
secretion into the intestinal lumen. This mechanism involves the increase of 445
intracellular cGMP by activating a guanylate cyclase C (GC-C). The activation of 446
this pathway leads to phosphorylation of the cystic fibrosis transmembrane 447
regulator (CFTR) by PKG (18, 32). Activation of CFTR stimulates the activity of a 448
Cl/HCO3- exchanger, which promotes an increase in secretion of HCO3
- into the 449
intestinal lumen (19). 450
In the kidney, it has been demonstrated that pendrin, an anion exchanger 451
found in β-intercalated cells, is downregulated at the transcriptional level by UGN 452
(29). Recently, our group has also demonstrated a role for UGN in the control of 453
luminal acidification (1, 21). Our findings suggest that guanylin peptides inhibit 454
bicarbonate reabsorption in proximal and distal segments of the nephron (1,20,21). 455
In contrast, recently, Rozenfeld and coworkers have demonstrated that UGN 456
downregulates the pendrin gene in β-intercalated cells (29) which in turns would 457
inhibit bicarbonate secretion. It is possible that this effect of UGN in β-intercalated 458
cells could be sort of a compensatory mechanism for the peptide inhibitory effect 459
on distal hydrogen secretion through α-intercalated cells of UGN. 460
The inhibition of H+-ATPase by a cGMP-dependent mechanism has been 461
described in plants (35) and in rat cortical collecting ducts (37). In the current 462
study we demonstrated that a permeable cGMP analogue inhibits distal hydrogen 463

21
secretion. In addition, a previous study from our laboratory has also demonstrated 464
the inhibition of distal hydrogen secretion by renoguanylin, a new member of the 465
guanylin family, through a mechanism dependent on PKG activation (20). 466
Moreover, the expression of guanilate cyclase C receptor has been demonstrated 467
in rat renal cortex (21) and, also, in MDCK-C11 cells, favoring the activation of the 468
classical guanylin receptor by UGN in these studies. In fact, our data show that 469
UGN incubation was able to stimulate the increment of the intracellular level of 470
cGMP and PKG activity in MDCK-C11 cells, which indicates the activation of the 471
classical pathway by UGN in this study. 472
In addition to the undeniable importance of the GC-C pathway on guanylin 473
action, especially during salt overload, when this receptor is upregulated (9, 27), 474
the presence of an additional receptor, other than GC-C, for UGN effect in the 475
kidney has been suggested (33). The mentioned studies have shown that GC-C 476
knockout mice still exhibit UGN-induced natriuresis (5), and also that UGN could 477
activate a G-protein receptor sensitive to pertussis toxin, in proximal tubule cells 478
(34). Moreover, we have previously demonstrated that UGN inhibits bicarbonate 479
reabsorption in proximal tubules through inhibition of NHE3 activity, by a 480
mechanism dependent on the activation of not only PKG but also the PKA 481
pathway. However, this same study showed that the activation of adenylyl cyclase 482
was not involved in the proximal effect of UGN (21). 483
In contrast to our previous work in proximal tubules (21), the present 484
observations showed that the PKA pathway was not involved in the distal effect of 485
UGN on hydrogen secretion, since the inhibition of PKA did not prevent the 486
inhibitory effect of UGN on distal hydrogen secretion. Besides, neither the 487

22
intracellular content of cAMP nor PKA activity were changed by UGN incubation of 488
MDCK-C11. Accordingly, the perfusion of distal segments with a cAMP analogue 489
did not promote a significant inhibitory effect of distal hydrogen secretion. 490
The activity of H+-ATPase may be regulated by trafficking, domain 491
assembly/disassembly, and changes in the ratio of ATP hydrolysis/H+-pumping as 492
well as by other means (39). Subcellular localization, regulation, and functional 493
differences of H+-ATPase populations may at least in part be regulated by the 494
presence of specific subunit isoforms. The B1, α4 and d2 isoforms have been 495
labeled as intercalated cell specific (38). Holliday et al. reported that the amino-496
terminal domains of both isoforms of the B subunit, B1 and B2, contain binding 497
sites to F-actin which may be responsible for the interaction between V-ATPase 498
and actin filaments in vivo and could allow for the observed trafficking (15). 499
The importance of the B1 subunit for the regulation of the proton pump has 500
been evidenced in patients with the inherited form of type 1 dRTA (distal renal 501
tubular acidosis). In humans, mutations of the gene encoding for the B1 subunit 502
(Atp6v1b1) in intercalated cells of distal tubule has been involved in the 503
pathogenesis of this disease (16). The characteristics of dRTA are not limited to 504
abnormal acid-base balance; hence it causes metabolic acidosis, but often 505
includes a salt-and potassium- losing nephropathy that may lead to hypokalemia 506
and dehydration (14, 30, 31). In the current study, we demonstrate that the 507
inhibitory effect of UGN on H+-ATPase activity in distal tubule involves reduction of 508
H+-ATPase B1 subunit abundance in the plasma membrane of distal tubule cells. 509
Besides, this effect was abolished in the presence of the PKG inhibitor, reinforcing 510
the involvement of the GC-C/cGMP/PKG pathway in the distal effect of UGN. 511

23
In summary, our findings indicate a role for UGN, a peptide known for its 512
effect in the regulation of sodium homeostasis, also in acid-base balance, by 513
inhibiting distal bicarbonate reabsorption. We suggest that UGN activates GC-C 514
receptors in distal segments of the nephron by increasing intracellular cGMP 515
content leading to PKG activation and decreasing insertion of H-ATPase B1 516
subunit in the apical membrane of α-intercalated cells. The data presented 517
herewith, also corroborate with previous studies that have postulated a role for 518
intercalated cells in the maintenance of body fluid and electrolyte balance, 519
involving inhibition of the B1 subunit of H+-ATPase (14). Furthermore, our current 520
and previous findings (21) and those of others (23, 26, 40) suggest UGN as a 521
possible counterregulator for the actions of the renin-angiotensin-aldosterone 522
system, a well-known promoter of sodium retention and acid extrusion by the 523
kidneys. 524
525
526

24
References 527
1. Amorim JB, Musa-Aziz R, Lessa LM, Malnic G, Fonteles MC. Effect of 528
uroguanylin on potassium and bicarbonate transport in rat renal tubules. 529
Can J Physiol Pharmacol 84: 1003-1010, 2006. 530
2. Boron WF, De Weer P. Intracellular pH transients in squid giant axons 531
caused by CO2, NH3, and metabolic inhibitors. J Gen Physiol 67: 91-532
112, 1976. 533
3. Carraro-Lacroix LR, Girardi AC, Malnic G. Long-term regulation of 534
vacuolar H(+)-ATPase by angiotensin II in proximal tubule cells. Pflugers 535
Arch 458: 969-79, 2009. 536
4. Carraro-Lacroix LR, Malnic G. Acid-base transport by the renal distal 537
nephron. J Nephrol.23 Suppl 16: S19-27, 2010. 538
5. Carrithers SL, Ott CE, Hill MJ, Johnson BR, Cai W, Chang JJ, Shah 539
RG, Sun C, Mann EA, Fonteles MC, Forte LR, Jackson BA, Giannella 540
RA, Greenberg RN. Guanylin and uroguanylin induce natriuresis in mice 541
lacking guanylyl cyclase-C receptor. Kidney Int 65: 40-53, 2004. 542
6. Fernandez R, Oliveira-Souza M, Malnic G. Na+-independent proton 543
secretion in MDCK-C11 cells. Pflugers Arch 441: 287-93, 2000. 544
7. Finberg KE, Wagner CA, Bailey MA, Wang T, Mentone SA, 545
Kashgarian M, Geibel JP, Lifton, RS. Loss of plasma membrane H-546
ATPase activity from cortical collecting duct intercalated cells of H-547
ATPase B1 subunit deficient mice: a mouse model of distal renal tubular 548
acidosis. J Am Soc Nephrol 13:4, 2002. 549
8. Fonteles MC, do Nascimento NR. Guanylin peptide family: history, 550
interactions with ANP, and new pharmacological perspectives. Can J 551
Physiol Pharmacol 89: 575-85, 2011. 552
9. Fonteles MC, Havt A, Prata RB, Prata PH, Monteiro HS, Lima AA, 553
Jorge AR, Santos CF, Greenberg RN, Nascimento NR. High-salt 554
intake primes the rat kidney to respond to a subthreshold uroguanylin 555
dose during ex vivo renal perfusion. Regul Pept 158: 6-13, 2009. 556
10. Forgac M. Vacuolar ATPases: rotary proton pumps in physiology and 557
pathophysiology. Nat Rev Mol Cell Biol 8: 917-29, 2007. 558

25
11. Forte LR, London RM, Freeman RH and Krause WJ. Guanylin 559
peptides: renal actions mediated by cyclic GMP. Am J Physiol Renal 560
Physiol 278: F180-F191, 2000. 561
12. Gekle M, Wuensch S, Oberleithner H, Silbernagl S. Characterization 562
of two MDCK cell subtypes as a model system to study principal cell and 563
intercalated cell properties. Pfluegers Arch 428:157-62, 1994. 564
13. Gil FZ, Malnic G. Effect of amphotericin B on renal tubular acidification 565
in the rat. Pfluegers Arch 413: 280-286, 1989. 566
14. Gueutin V, Vallet M, Jayat M, Peti-Peterdi J, Cornière N, Leviel F, 567
Sohet F, Wagner CA, Eladari D, Chambrey R. Renal β-intercalated 568
cells maintain body fluid and electrolyte balance. J Clin Invest 123:4219-569
31, 2013. 570
15. Holliday LS, Lu M, Lee BS, Nelson RD, Solivan S, Zhang L, Gluck 571
SL. The amino-terminal domain of the B subunit of vacuolar H+-ATPase 572
contains a filamentous actinbinding site. J Biol Chem 275: 32331-7, 573
2000. 574
16. Karet FE, Finberg KE, Nelson RD, Nayir A, Mocan H, Sanjad SA, 575
Rodriguez-Soriano J, Santos F, Cremers CW, Di Pietro A, Hoffbrand 576
BI, Winiarski J, Bakkaloglu A, Ozen S, Dusunsel R, Goodyer P, 577
Hulton SA, Wu DK, Skvorak AB, Morton CC, Cunningham MJ, Jha V, 578
Lifton RP. Mutations in the gene encoding B1 subunit of H+-ATPase 579
cause renal tubular acidosis with sensorineural deafness. Nat Genet 21: 580
84-90, 1999. 581
17. Kulaksiz H, Cetin Y. The electrolyte/fluid secretion stimulatory peptides 582
guanylin and uroguanylin and their common functional coupling proteins 583
in the rat pancreas: a correlative study of expression and cell-specific 584
localization. Pancreas. Am J Pathol 25:170-5, 2002. 585
18. Kulaksiz H, Schmid A, Hönscheid M, Eissele R, Klempnauer J, Cetin 586
Y. Guanylin in the human pancreas: a novel luminocrine regulatory 587
pathway of electrolyte secretion via cGMP and CFTR in the ductal 588
system. Histochem Cell Biol 115:131-45, 2001. 589

26
19. Lee MG, Choi JY, Luo X, Strickland E, Thomas PJ, Muallem S. Cystic 590
fibrosis transmembrane conductance regulator regulates luminal Cl-591
/HCO3- exchange in mouse submandibular and pancreatic ducts. J Biol 592
Chem 274: 14670-7, 1999. 593
20. Lessa LM, Amorim JB, Fonteles M, Malnic G. Effect of renoguanylin 594
on hydrogen/bicarbonate ion transport in rat renal tubules. Regul Pept 595
157: 37-43, 2009. 596
21. Lessa LM, Carraro-Lacroix LR, Crajoinas RO, Bezerra CN, Dariolli R, 597
Girardi AC, Fonteles MC, Malnic G. Mechanisms underlying the 598
inhibitory effects of uroguanylin on NHE3 transport activity in renal 599
proximal tubule. Am J Physiol Renal Physiol 303: F1399-408, 2012. 600
22. Lowry OH, Rosebrough NJ, Farr AL, Randall RJ. Protein 601
measurement with the Folin phenol reagent. J Biol Chem 193: 265-275, 602
1951. 603
23. Michell AR, Debnam ES, Unwin, RJ. Regulation of Renal Function by 604
the Gastrointestinal Tract: Potential Role of Gut-Derived Peptides and 605
Hormones. Annu Rev Physiol 70:379-403, 2008. 606
24. Moss NG, Riguera DA, Fellner RC, Cazzolla C, Goy MF, Moss NG, 607
Goy MF. Natriuretic and antikaliuretic effects of uroguanylin and 608
prouroguanylin in the rat. Am J Physiol Renal Physiol 299: F1433-42, 609
2010. 610
25. Oliveira-Souza M, Malnic G, Mello-Aires M. Atrial natriuretic peptide 611
impairs the stimulatory effect of angiotensin II on H+-ATPase. Kidney 612
international 62: 1693–9, 2002. 613
26. Pech V, Zheng W, Pham TD, Verlander JW, Wall SM. Angiotensin II 614
activates H+-ATPase in type A intercalated cells. J Am Soc Nephrol 615
19:84-91, 2008. 616
27. Potthast R, Ehler E, Scheving LA, Sindic A, Schlatter E, Kuhn M. 617
High salt intake increases uroguanylin expression in mouse kidney. 618
Endocrinology 142: 3087-3097, 2001. 619
28. Rahbi H, Narayan H, Jones DJL, Ng LL. The uroguanylin system and 620
human disease. Clinical science 123: 659–68, 2012. 621

27
29. Rozenfeld J, Tal O, Kladnitsky O, Adler L, Efrati E, Carrithers SL, 622
Alper SL, Zelikovic I. The pendrin anion exchanger gene is 623
transcriptionally regulated by uroguanylin: a novel enterorenal link. Am J 624
Physiol Renal Physiol 302: F614-24, 2012. 625
30. Sebastian A, McSherry E, Morris RC Jr. Renal potassium wasting in 626
renal tubular acidosis (RTA): its occurrence in types 1 and 2 RTA despite 627
sustained correction of systemic acidosis. J Clin Invest 50: 667-78, 1971. 628
31. Sebastian A, McSherry E, Morris RC Jr. Impaired renal conservation of 629
sodium and chloride during sustained correction of systemic acidosis in 630
patients with type 1, classic renal tubular acidosis. J Clin Invest 58:454-631
69, 1976. 632
32. Seidler U, Blumenstein I, Kretz A, Viellard-Baron D, Rossmann H, 633
Colledge WH, Evans M, Ratcliff R, Gregor M. A functional CFTR 634
protein is required for mouse intestinal cAMP-, cGMP- and Ca(2+)-635
dependent HCO3- secretion. J Physiol 505:411-23, 1997. 636
33. Sindic A, Basoglu C, Cerci A, Hirsch JR, Potthast R, Kuhn M, 637
Ghanekar, Viswerswariah SS, Schlatter E. Guanylin, uroguanylin, and 638
heat-stable euterotoxin activate guanylate cyclase C and/or a pertussis 639
toxin-sensive G protein in human proximal tubule cells. J Biol Chem 640
277:17758-64, 2002. 641
34. Sindic A, Velic A, Basoglu C, Hirsch JR, Edemir B, Kuhn M, 642
Schlatter E. Uroguanylin and guanylin regulate transport of mouse 643
cortical collecting duct independent of guanylate cyclase C. Kidney Int 644
68: 1008-1017, 2005. 645
35. Suwastika IN, Gehring CA. The plasma membrane H+-ATPase from 646
Tradescantia stem and leaf tissue is modulated in vitro by cGMP. Arch 647
Biochem Biophys 367:137-9, 1999. 648
36. Thomas J, Buchsbaum R, Zimniak A, Racher E. Intracellular pH 649
measurements in Ehrlich ascites tumor cells utilizing spectroscopic 650
probes generated in situ. Biochemistry 18: 2210-2218, 1979. 651

28
37. Tojo A, Guzman NJ, Garg LC, Tisher CC, Madsen KM. Nitric oxide 652
inhibits bafilomycin-sensitive H+-ATPase activity in rat cortical collecting 653
duct. Am J Physiol Renal,Fluid Electrolyte Physiol 267:F509-F515, 1994. 654
38. Wagner CA, Devuyst O, Bourgeois S, Mohebbi N. Regulated acid-655
base transport in the collecting duct. Pflugers Arch 458: 137-56, 2009. 656
39. Wagner CA, Finberg KE, Breton S, Marshansky V, Brown D, Geibel 657
JP. Renal vacuolar H+-ATPase. Physiol Rev 84: 1263-314,2004. 658
40. Winter C, Kampik NB, Vedovelli L, Rothenberger F, Paunescu TG, 659
Stehberger PA, Brown D, John H, Wagner CA. Aldosterone stimulates 660
vacuolar H(+)-ATPase activity in renal acid-secretory intercalated cells 661
mainly via a protein kinase C-dependent pathway. Am J Physiol Cell 662
Physiol 301: C1251-61, 2011. 663
664
665
666
667
668
669
670
671
29
672
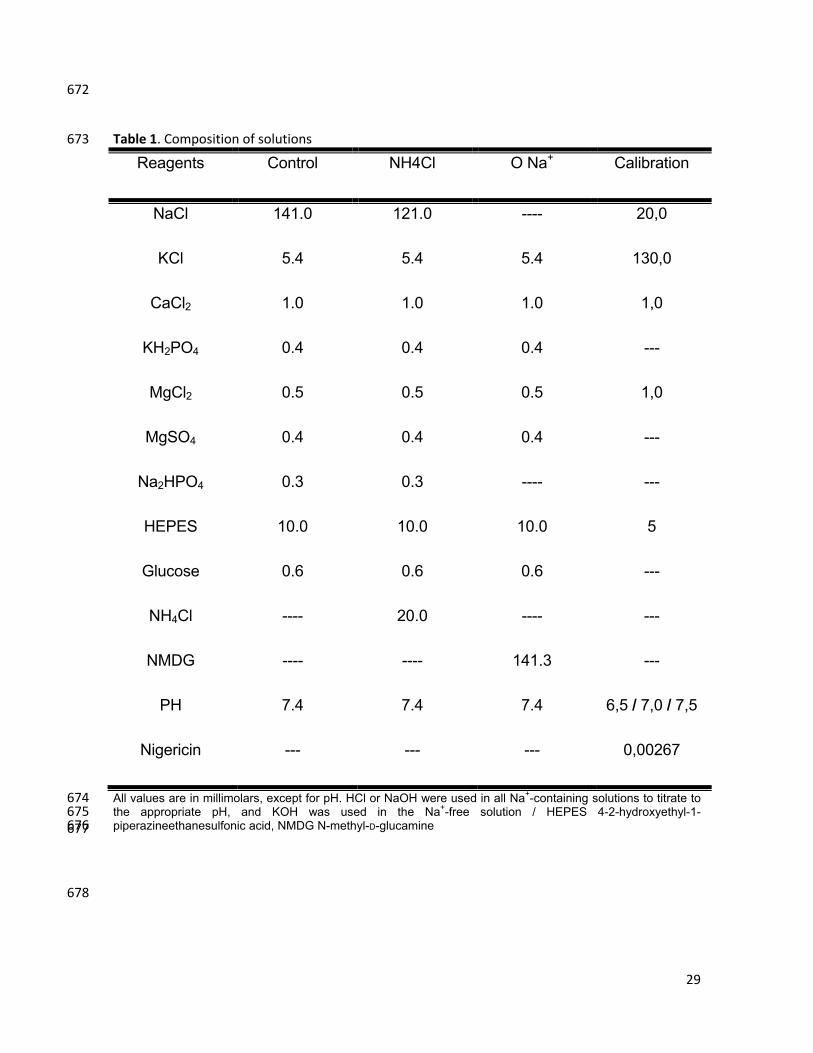
Table 1. Composition of solutions 673
Reagents Control NH4Cl O Na+ Calibration
NaCl 141.0 121.0 ---- 20,0
KCl 5.4 5.4 5.4 130,0
CaCl2 1.0 1.0 1.0 1,0
KH2PO4 0.4 0.4 0.4 ---
MgCl2 0.5 0.5 0.5 1,0
MgSO4 0.4 0.4 0.4 ---
Na2HPO4 0.3 0.3 ---- ---
HEPES 10.0 10.0 10.0 5
Glucose 0.6 0.6 0.6 ---
NH4Cl ---- 20.0 ---- ---
NMDG ---- ---- 141.3 ---
PH 7.4 7.4 7.4 6,5 / 7,0 / 7,5
Nigericin --- --- --- 0,00267
All values are in millimolars, except for pH. HCl or NaOH were used in all Na+-containing solutions to titrate to 674 the appropriate pH, and KOH was used in the Na+-free solution / HEPES 4-2-hydroxyethyl-1-675 piperazineethanesulfonic acid, NMDG N-methyl-D-glucamine 676
677
678

30
FIGURE LEGENDS 679
680
Figure 1: UGN effect on distal bicarbonate reabsorption involves H+-681
ATPase and NHE inhibition. JHCO3- was evaluated by means of stationary 682
microperfusion according to the protocol described in “Methods”. Kidney distal 683
tubules were perfused with control solution alone or together with uroguanylin 684
(UGN) (10-6 M) or the NHE inhibitor HMA (10-4 M) or/and the H-ATPase inhibitor 685
Concanamycin A (10-7M). Data are means ± SE. *p < 0.05 vs Control. 686
687
Figure 2 - 8Br-cGMP but not 8Br-cAMP mimicks the UGN effect on distal 688
bicarbonate reabsorption A, Experiments were performed in the presence of 689
8br-cGMP (10-6 M) and/or HMA (10-4 M), a selective inhibitor of NHE. B. Tubules 690
were perfused with 8br-cAMP (10-6 M) and/or HMA (10-4 M). Numbers of perfused 691
tubules are indicated in the bars. Data are means ± SE. *p < 0.05 vs Control; &p < 692
0,05 vs KT5823+UGN. 693
694
Figure 3: The inhibitory effect of UGN on H+ secretion in rat renal distal 695
tubule is mediated by PKG. A, Experiments were performed in the presence or 696
absence of uroguanylin (UGN) (10-6 M) and KT5823 (10-6 M), a selective inhibitor 697
of cGMP-dependent protein kinase (PKG), which prevented the effect promoted by 698
the peptide. B. Tubules were perfused with control solution with or without 699
uroguanylin and H89 (10-5 M), a PKA inhibitor, which was unable to prevent the 700
inhibitory effect of Uroguanylin. Numbers of perfused tubules are indicated in the 701
bars. Data are means ± SE. *p < 0.05 vs Control; #p < 0,05 vs KT5823+UGN. 702

31
Figura 4: Original Traces of Fluorescence Microscopy Experiments with 703
MDCK-C11 cells. The records show the recovery of pHi along 2 minutes after an 704
acid pulse with NH4Cl. The Figure shows three different conditions. A, control 705
solution with 135 mM Na+; B, 0Na+ solution and C, 1 µM uroguanylin (UGN) added 706
to the 0NA+ solution. 707
708
Figure 5: Uroguanylin inhibits Na+-independent pH recovery and hydrogen 709
flux (JH+) in MDCK C11 cells. MDCK-C11 cells were treated with control solution 710
(135mM Na), 0NA solution or 1µM Uroguanylin diluted in 0Na solution. After 711
intracellular acidification by means of the ammonium pulse technique in MDCK-712
C11 cells, the initial rates of pH recovery (pH units/min) were calculated from the 713
curves by linear regression analysis (A). In B are plotted the rates of hydrogen 714
extrusion which were obtained from the product of dpHi/dt and intracellular buffer 715
capacity βi of the avarege pHi of the experimental groups (pHi 6,5). Uroguanylin 716
(UGN) significantly inhibits Na+-independent pHi recovery and Hydrogen extrusion 717
in MDCK-C11.Number of experiments is indicated in the bars.*p<0,05 vs control, # 718
p<0.05 vs 0 Na. 719
720
Figure 6: Uroguanylin inhibits H+-ATPase-dependent pH recovery in MDCK 721
C11 cells by a PKG dependent pathway. MDCK-C11 cells were treated with 1µM 722
uroguanylin (UGN) or 8br-cGMP in the presence of EIPA (NHE1 inhibitor) and 723
Schering (H/K-ATPase inhibitor) and/or KT5823 (PKG inhibitor) diluted in 0Na 724
solution, after intracellular acidification by means of the ammonium pulse 725
technique. The initial rates of pHi recovery (pH units/min) were calculated from the 726

32
curves by linear regression analysis (A). In B are plotted the rates of hydrogen 727
extrusion which was obtained from the product of dpHi/dt and intracellular buffer 728
capacity βi of the average pHi of the experimental groups (pHi 6,5).Uroguanylin 729
(UGN) significantly inhibits H+-ATPase-dependent pHi recovery and hydrogen 730
extrusion in MDCK-C11 cells. Number of experiments is indicated in the 731
bars.*p<0.05 vs EIPA + SCH. 732
733
Figure 7. UGN stimulates increase of intracellular cGMP content and PKG 734
activity in MDCK-C11 cells, which express GC-C. A, Cells were incubated for 10 735
min with culture medium containing 1 mM 3-isobutyl-1-methylxanthine (IBMX) or 736
IBMX and 10-6 M Uroguanilyn or 10-6 M ANP during 10 minutes prior to lysis. The 737
generation of intracellular cGMP was estimated by enzyme immunoassay (cGMP 738
Direct Biotrak EIA, GE Healthcare) according to manufacturer’s protocol B, PKG 739
activity of UGN-treated cells was measured by the single-site and semi-quantative 740
CycLex cGK Assay Kit (CycLex Co., Ltd., Nagano, Japan) according to the 741
manufacturer’s protocols. UGN promoted an increase of PKG activities, as shown 742
in the bar graph. *p < 0.05 vs. Control.; C, Expression of GC-C receptor in MDCK-743
C11 cells. Total RNA was extracted, reverse transcribed and amplified as 744
described in “Methods”. PCR amplification, using specific primers, gave rise to one 745
band with the predicted size of 491 bp. MDCK-C11 cell cDNA was also tested for 746
the housekeeping gene 28S-rRNA. 747
748
Figure 8: UGN does not stimulate increase of intracellular cAMP content and 749
PKA activity in MDCK-C11 cells. A, MDCK- C11 cells were incubated for 10 min 750

33
with culture medium containing 1 mM 3-isobutyl-1-methylxanthine (IBMX) or IBMX 751
with uroguanylin (10-6 M) or forskolin (10-4 M). cAMP levels were measured by 752
enzyme immunoassay (cAMP Direct Biotrak EIA, GE Healthcare). Each assay was 753
performed in triplicate, and the mean values of 3 assays were calculated. B, PKA 754
activity was measured in MDCK-C11 lysates after treatment with UGN by the PKA 755
Kinase Activity Assay (Enzo Life Sciences, Inc., Farmingdale, NY) according to the 756
manufacturer's instructions. UGN does not promote a significant increase in the 757
activity of PKA, as shown in the bar graph. ***p < 0.05 vs. Control. 758
759
Figure 9. UGN decreases the surface expression of H+-ATPase B1 subunit in 760
MDCK-C11 cells by a PKG dependent mechanism. A and B. Equivalent 761
quantities (50 µg) of total cellular lysates from MDCK-C11 cells treated during 15 762
min with vehicle (CTRL), UGN or UGN combined with the PKG inhibitor were 763
subjected to SDS-PAGE and immunoblotting. A. Representative immunoblotting 764
using the monoclonal antibody against the B1 subunit of H+-ATPase. Actin was 765
used as an internal control. B. Representative immunoblotting using the 766
monoclonal antibody against the β subunit of H+/K+-ATPase. Actin was used as an 767
internal control. C and D. Cell surface biotinylated proteins, after treatment with 768
uroguanylin (UGN) or UGN + KT5823 (PKG inhibitor) for 15 minutes, were 769
subjected to SDS-PAGE and immunoblotting. Immunoblot analyses were 770
performed using (C) a monoclonal antibody against the B1 subunit of H+-ATPase 771
or (D) a monoclonal antibody against the β subunit of H+/K+-ATPase. The amount 772
of H+-ATPase B1 subunit and H+/K+-ATPase β subunit were quantitated by 773

34
densitometry. Number of experiments are indicated within the parenthesis. 774
**P<0.01 vs. CTRL 775
FIGURES
Figure 1
CTRL
HMA+CONC
UGN
HMA+CONC+U
GN0.0
0.5
1.0
1.5
** *
(6) (6)(12) (10)JHC
O3- , n
mol
/cm
2 .s

Figure 2
Control
8br-c
GMPHMA
8br-c
GMP + HMA
0.0
0.5
1.0
1.5
* *
(6) (7) (8) (10)
A
JHC
O3- , n
mol
/cm
2 .s
Control
8br-c
AMPHMA
8br-c
AMP + HMA
0.0
0.5
1.0
1.5
(6) (9) (8) (9)
B
JHC
O3- , n
mol
/cm
2 .s
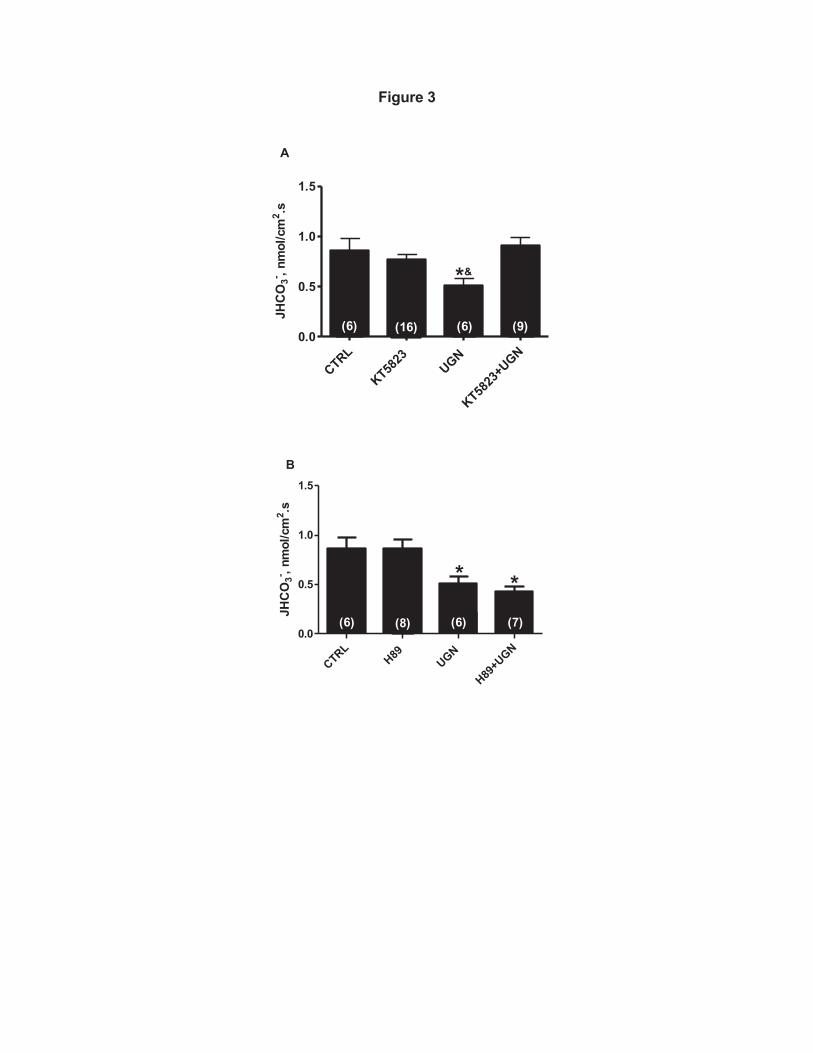
Figure 3
CTRL
KT5823
UGN
KT5823
+UGN
0.0
0.5
1.0
1.5
*
A
(6) (16) (9)(6)
&JH
CO
3- , nm
ol/c
m2 .s
CTRLH89
UGN
H89+U
GN0.0
0.5
1.0
1.5
* *(6) (8) (6) (7)
B
JHC
O3- , n
mol
/cm
2 .s
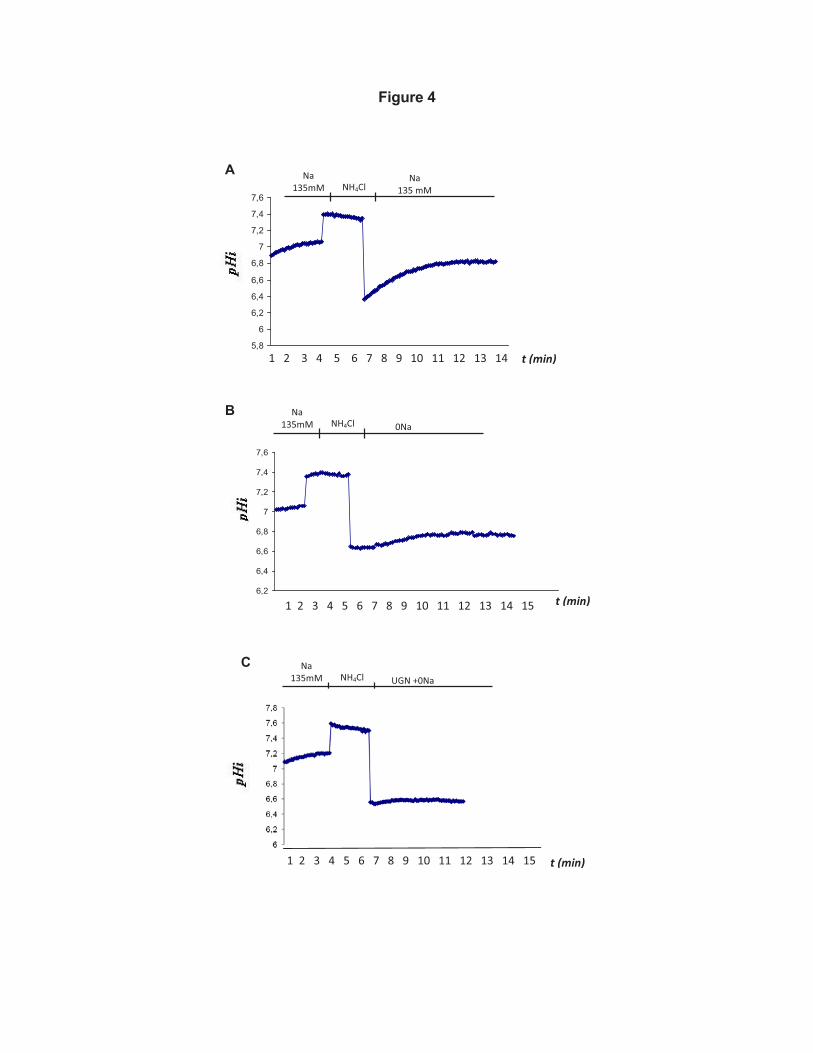
Figure 4
5,8
6
6,2
6,4
6,6
6,8
7
7,2
7,4
7,6
6,2
6,4
6,6
6,8
7
7,2
7,4
7,6
1 2 3 4 5 6 7 8 9 10 11 12 13 14 t (min)
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 t (min)
B
A Na 135mM NH4Cl
Na 135 mM
Na 135mM NH4Cl
0Na
C Na 135mM NH4Cl
UGN +0Na
t (min) 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
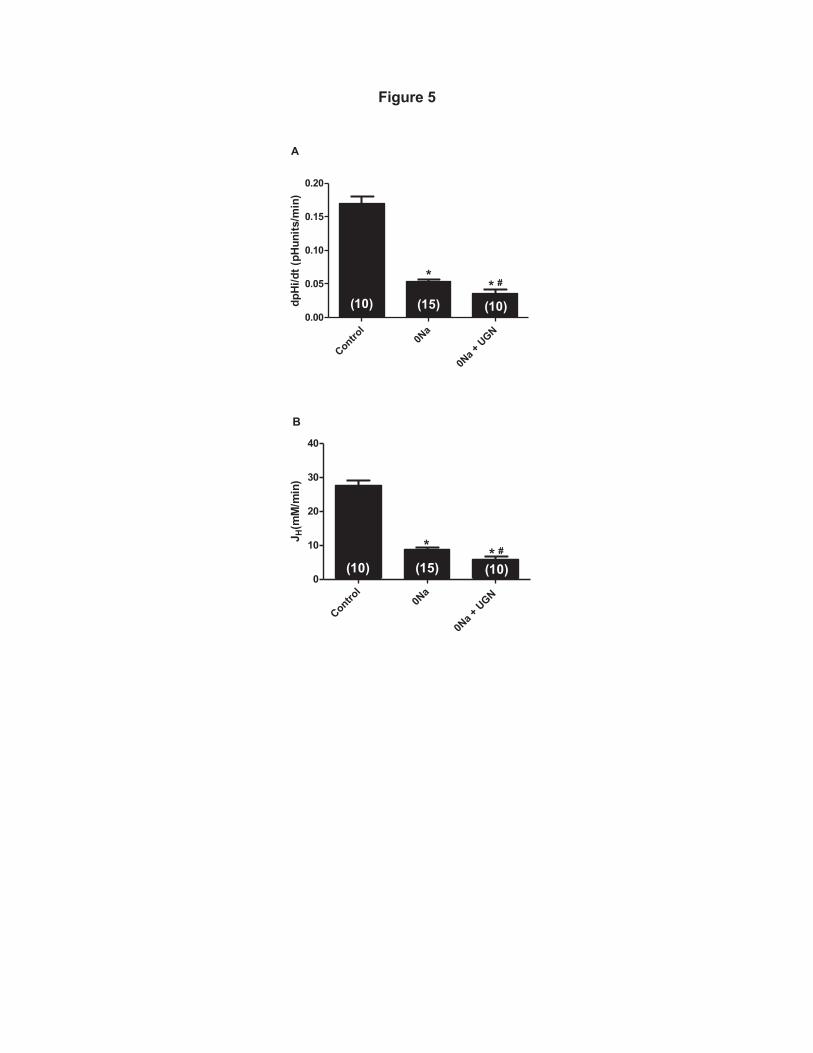
Figure 5
Control
0Na
0Na +
UGN
0.00
0.05
0.10
0.15
0.20
** #
A
(10) (15) (10)dpH
i/dt (
pHun
its/m
in)
Control
0Na
0Na +
UGN
0
10
20
30
40
(10) (15) (10)* * #
B
J H(m
M/m
in)
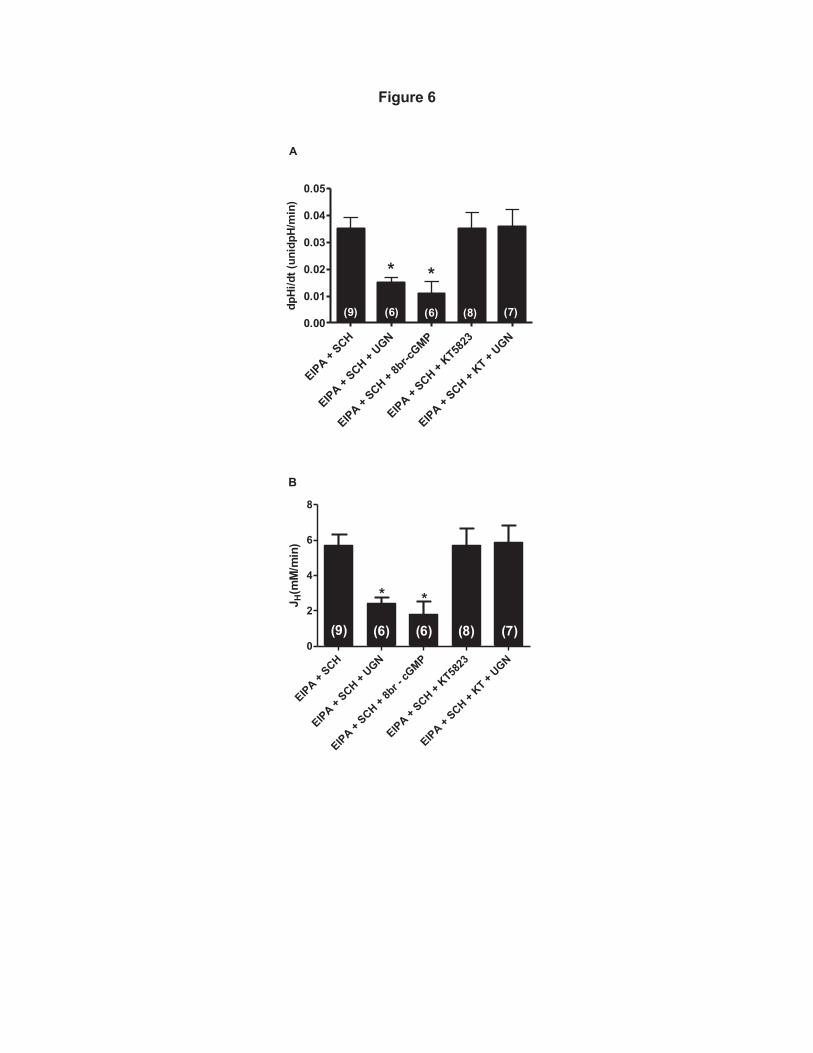
Figure 6
EIPA + SCH
EIPA + SCH +
UGN
EIPA +
SCH + 8b
r-cGMP
EIPA + SCH +
KT5823
EIPA + SCH +
KT + UGN
0.00
0.01
0.02
0.03
0.04
0.05
* *
(6) (6) (8) (7)(9)dpHi
/dt (
unid
pH/m
in)
A
EIPA + SCH
EIPA + SCH +
UGN
EIPA + SCH +
8br -
cGMP
EIPA + SCH +
KT5823
EIPA + SCH +
KT + UGN
0
2
4
6
8
* *
(9) (6) (6) (8) (7)
B
J H(m
M/m
in)
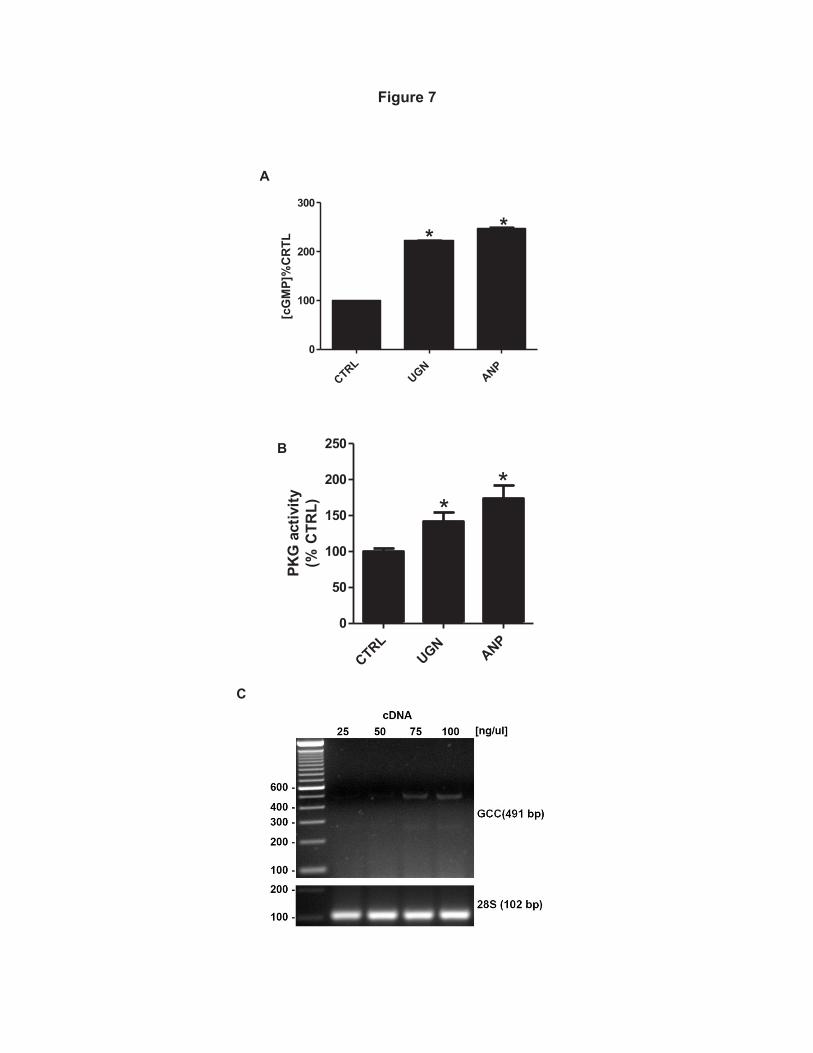
Figure 7
CTRLUGN
ANP 0
100
200
300
* *
[cG
MP]
%C
RTL
CTRLUGN
ANP0
50
100
150
200
250
**
PKG
act
ivity
(% C
TRL)
A
B
C
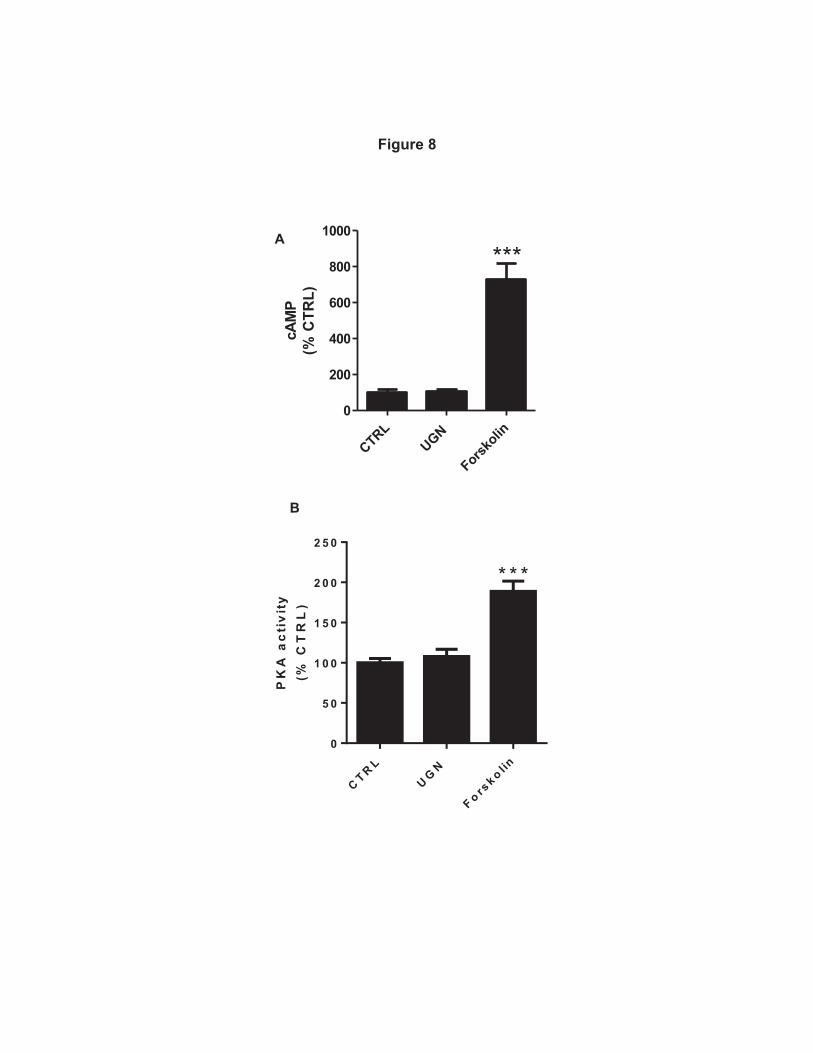
Figure 8
CTRLUGN
Forskolin
0
200
400
600
800
1000
***cA
MP
(% C
TRL)
C T R L
U GN
F o rsk o lin
0
5 0
1 0 0
1 5 0
2 0 0
2 5 0
* * *
PK
A a
cti
vit
y(%
CT
RL
)
A
B
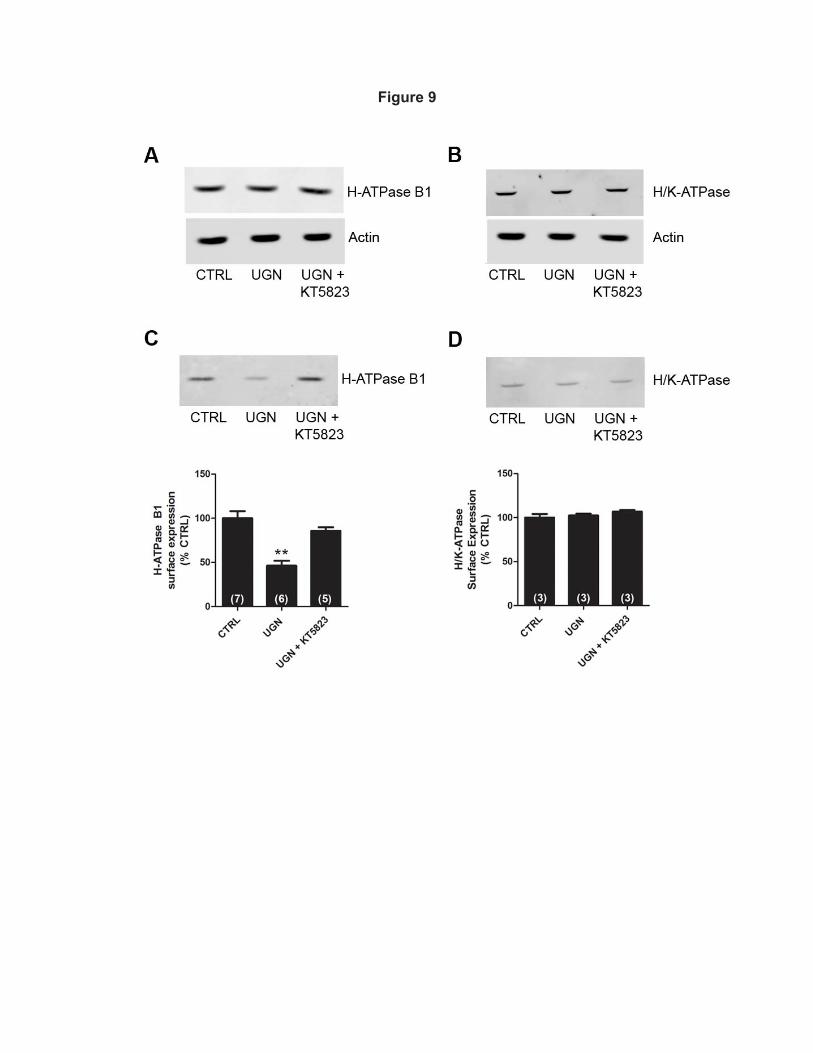
Figure 9




















