
Complete_thesis.pdf - the University of Groningen research ...
Upload
khangminh22Category
view
3download
0
University of Groningen
Bacillus mycoides: novel tools for studying the mechanisms of its interaction with plantsYi, Yanglei
IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite fromit. Please check the document version below.
Document VersionPublisher's PDF, also known as Version of record
Publication date:2018
Link to publication in University of Groningen/UMCG research database
Citation for published version (APA):Yi, Y. (2018). Bacillus mycoides: novel tools for studying the mechanisms of its interaction with plants.University of Groningen.
CopyrightOther than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of theauthor(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons).
The publication may also be distributed here under the terms of Article 25fa of the Dutch Copyright Act, indicated by the “Taverne” license.More information can be found on the University of Groningen website: https://www.rug.nl/library/open-access/self-archiving-pure/taverne-amendment.
Take-down policyIf you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediatelyand investigate your claim.
Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons thenumber of authors shown on this cover page is limited to 10 maximum.
Download date: 26-01-2022

BACILLUS MYCOIDES: novel tools for studying the mechanisms of its interaction with plants
YANGLEI YI

Bacillus mycoides: novel tools for studying the mechanisms of its interaction with plants
Academic Thesis, University of Groningen, the Netherlands
ISBN: 978-94-034-0644-2 978-94-034-0643-5 (ebook)Printing: Eikon +Cover & layout: Lovebird design. www.lovebird-design.com
© Y. Yi, Groningen, the Netherlands, 2018All rights reserved. No part of this publication may be reproduced or transmitted in any form or by any means, without written permission of the author.
Bacillus mycoides: novel tools for studying the mechanisms of
its interaction with plants
PhD thesis
to obtain the degree of PhD at the
University of Groningen
on the authority of the
Rector Magnificus Prof. E. Sterken
and in accordance with
the decision by the College of Deans.
This thesis will be defended in public on
Friday 25 May 2018 at 12.45 hours
by
Yanglei Yi
born on 24 January 1988
in Shaanxi, China

TABLE OF CONTENTS
CHAPTER 1 Introduction 7
CHAPTER 2 Draft genome sequences of seven Bacillus mycoides
strains, isolated from potato endosphere and soil 31
CHAPTER 3 Comparative Transcriptomics of Bacillus mycoides Strains
in Response to Potato-Root Exudates Reveals Different Genetic Adaptation of Endophytic and Soil Isolates 37
CHAPTER 4 Development of an efficient electroporation method for
rhizobacterial Bacillus mycoides strains 65
CHAPTER 5 Optimized fluorescent proteins for the rhizosphere-as-
sociated bacterium Bacillus mycoides with endophytic and biocontrol agent potential 81
CHAPTER 6 Exploring plant-microbe interactions of the rhizobac-
teria Bacillus subtilis and Bacillus mycoides by use of the CRISPR-Cas9 system 117
CHAPTER 7 General Discussion 151
Samenvatting 161 Acknowledgements 165
SupervisorsProf. O.P. Kuipers
Prof. J.W. Veening
Assessment Committee Prof. J. Falcao Salles Prof. A.J.M. Driessen Prof. J. Raaijmakers

1Introduction

1
9
Intr
oduc
tion
PLANT-MICROBE INTERACTION
Plants naturally harbor diverse species of microorganisms, because they offer a wide range of habitats, supporting microbial growth. These mi-croorganisms form a complex biological community interacting with the plant host, e.g. being pathogenic, or employing mutualism (symbi-onts), and commensalism (Pini et al., 2012). Bacteria that associate with plants are diverse in their ability to affect plant health, their genotypes, and their phenotypic characteristics. Although the majority of research on plant-associated microorganisms has focused on phytopathogens and diazotrophic (nitrogen-fixing) phytosymbionts, it is clear that many plant-associated microbes, even those that comprise only a small pro-portion of a community, have functions that are of agricultural or envi-ronmental importance (Beattie, 2006). Bacteria could reside in/on seeds, roots, leaves, and fruits of plants. The rhizosphere, the narrow zone of soil influenced by the root, is much richer in bacteria than the surround-ing bulk soil and even more densely populated than other parts of a plant (Lugtenberg and Kamilova, 2009) (Figure 1).
The rhizosphere contains an enormous range of compounds excreted by roots. The roots exudate a range of inorganic compounds like ions,
Figure 1. Plant-associated microorganisms, plant-microbe and microbe-microbe inter-action. Soil inhibiting bacteria are attracted by root secreted signals to become plant- associated bacteria. They may live in the rhizosphere, rhizoplane or endosphere. Some of them are aiding plant health by fighting against pathogens through niche competi-tion, antimicrobial production, and induced systematic resistance.

11
10 11
Intr
oduc
tion
Plan
t-mic
robe
inte
ract
ion
inorganic acids, oxygen and water. These compounds may directly affect the biogeochemistry of the soil (Jones et al., 2009). However, the major-ity of the root exudates are formed by organic materials, which can be di-vided into low-molecular-weight compounds and high-molecular-weight compounds. The low-molecular-weight compounds include amino acids, organic acids (citric, malic, succinic, oxalic and pyruvic), sugars (glucose, xylose, fructose, maltose, sucrose, ribose), phenolics, fatty acids and an array of secondary metabolites. The high-molecular weight compounds consist of mucilage and proteins (Badri and Vivanco, 2009; Rohrbacher and St-Arnaud, 2016), and rhizosphere microorganisms can use some of these compounds as an energy source for growth and development or as signaling molecules (Carvalhais et al., 2015). The composition of root ex-udates varies by plant species, developmental stage and plant growth sub-strate (Doornbos et al., 2011). For example, the organic acids and sugars in root exudates of tomato and cucumber increase during plant growth on stone wool (Kamilova et al., 2006). A systematic proteomic analysis of root exudates showed that the defense-related proteins such as chitinases, glucanases, and myrosinases showed more secretion during the flower-ing stage of Arabidopsis thaliana (De-la-Peña et al., 2010), while the release of amino acids and phenolics increased at later stages of life of Arabidopsis (Chaparro et al., 2013).
It is believed that the dynamic nature of root exudates is one of the driv-ing forces for the plant to select associated microbial communities that are favorable for the plant. The best-known plant-microbe symbiosis is the nodulation of legumes by rhizobia. This interaction is very specific, allow-ing certain rhizobial strains to nodulate with specific host legumes (Bais et al., 2006). In soil, a Rhizobium spp. finds its host legume plant from a distance by chemotaxis, being attracted to the root exudates due to the presence of flavonoids (Sugiyama and Yazaki, 2012). Flavonoids also reg-ulate the expression of nod genes that play important roles in nodulation establishment (Abdel-Lateif et al., 2012). Another widely-occurring plant- associated microbiome part is formed by arbuscular mycorrhizal fungi (AMF), which form a symbiotic relationship with more than 80% of terres-trial plants, from bryophytes to tracheophytes (Lee et al., 2013). Although AMF have low host specificity in their symbiosis with plants, they may also recognize their host by signals released by plant roots, similar to the rec-ognition by rhizobia. It has been shown that mycorrhizal fungi are me-diated by root-secreted compounds, such as strigolactone 5-deoxystrigol ( Yoneyama et al., 2008), sugars, and carbohydrates (Kiers et al., 2011).
Some specific root-released chemical signals have been identified, that play a role in recruiting specific bacteria to build mutualistic interactions with the plant. For example, the chemotaxis to amino acids of root exudates
has an important role in root colonization by Pseudomonas fluorescens Pf0-1 (Oku et al., 2012). In Azospirillum brasilense, the chemoreceptor- like pro-tein Tlp1 serves as an energy taxis transducer and affects the coloniza-tion on roots (Greer-Phillips et al., 2004). Chemotaxis towards A. thaliana root exudates was mediated by two well-characterized chemoreceptors, McpB and McpC, as well as by the orphan receptor TlpC in Bacillus subtilis (Allard-Massicotte et al., 2016). Moreover, plant roots also secrete com-pounds that mimic quorum-sensing (QS) signals of bacteria to stimulate QS-regulated responses of associated bacteria (Huang et al., 2014). The most common QS system occurs via N-acyl homoserine lactones (AHLs) binding to members of LuxR-family transcriptional regulators. Thus, they regulate the expression of downstream genes with the lux box in the pro-moter region (Hartmann et al., 2014). However, the LuxR-family regu-lator (Lux-R solo) found in plant-associated bacteria lacks the LuxI pro-tein when compared with other LuxR families. These regulators do not bind AHLs but to plant-produced compounds (González et al., 2013; Ven-turi and Fuqua, 2013). For instance, a LuxR solo regulator of P. fluorescens, PsoR, was identified to have a function in responding to plant compounds and regulating genes involved in the biosynthesis of various antimicro-bial compounds including 2,4-diacetylphloroglucinol, which has antago-nistic activity against the damping-off disease pathogen Pythium ultimum (Subramoni et al., 2011).
Following recognition and recruitment, bacteria physically interact with different parts of the roots from the tip to the elongation zone to form complex multicellular and often multispecies assemblies, including bio-films and smaller aggregates (Danhorn and Fuqua, 2007). The rhizobac-teria form biofilms in a heterogeneous way along the root surface, where the root tips usually have the least bacterial biofilm formed (Rudrappa et al., 2008). In general, the zone immediately behind the root tip is con-sidered to be a major site of colonization (Badri and Vivanco, 2009). This is partially due to the pH difference from the root tip to basal regions ( Peters, 2004), which might affect the bacterial growth and biofilm devel-opment. The nutrient availability is also quite different since particular cell types in the root system might be more important than others in the secretion of particular compounds. The biofilm formation of rhizobacte-ria on root surfaces is a mutualistic interaction. In B. subtilis-A. thaliana interaction, plant polysaccharides act as an environmental cue that trig-gers biofilm formation by the bacterium (Beauregard et al., 2013). In turn, plants benefit from the biofilm of rhizobacteria. The inhibitory effect of Lysobacter sp. strain SB-K88 against pathogenic Aphanomyces cochlioides is due to a combination of antibiosis and biofilm formation at the rhizo-plane of the host plant (Islam et al., 2005).

11
12 13
Intr
oduc
tion
Plan
t gro
wth
pro
mot
ing
rhiz
obac
teri
a (P
GPR
)
Although extensive studies have been performed to investigate plant- associated bacteria, we are far from understanding the plant- microbe in-teraction mechanisms, especially when indigenous multispecies bacterial communities are considered. So far, the studies on plant-growth promot-ing rhizobacteria (PGPR) and their beneficial effects on host plants have drawn considerable attention, but more research is needed to increase our understanding of the underlying processes.
PLANT GROWTH PROMOTING RHIZOBACTERIA (PGPR)
PGPR form a group of bacteria that colonize the host plant roots and pro-mote plant growth due to certain traits of the rhizobacteria. A diverse ar-ray of bacterial species including Azospirillum, Azotobacter, Bacillus, Burk-holderia, Enterobacter, Klebsiella, Pseudomonas, Serratia, and Xanthomonas were identified as PGPR of which Bacillus and Pseudomonas spp. are pre-dominant (Podile and Kishore, 2007). PGPR promote plant growth di-rectly by either facilitating resource acquisition (nitrogen, phosphorus, and essential minerals) or modulating plant hormone levels, or indirectly, by decreasing the inhibitory effects of various pathogens on plant growth and development in the form of biocontrol agents (Ahemad and Kibret, 2014) (Figure 2).
Of all the essential nutrients, nitrogen (N) is required by plants in the largest quantity and is most frequently the limiting factor in crop pro-ductivity. However, 78% of N is in the atmosphere and thus is unavaila-ble for growing plants. Biological N2 fixation by nitrogen-fixing microor-ganisms is widely distributed in nature. Rhizobia such as Rhizobium and Bradyrhizobium associated with legume plants are well known for their N2 fixing activity and are defined as symbiotic N2-fixing bacteria (Zah-ran, 2001). Free-living PGPR with N2-fixing capacity are defined as diaz-otrophs (Kennedy et al., 2004), which can form a non-obligate interaction with host plants. The second most important essential nutrient, phos-phate (P), is ubiquitously present in soil but has low solubility or can eas-ily be converted to insoluble forms (Hanif et al., 2015). Some PGPR pro-duce organic acids or enzymes including phosphatases and phytases to release soluble phosphorus from organic compounds in soil (Lugten-berg and Kamilova, 2009). Also, balanced and sufficient potassium and/or iron availability is essential to maintain a healthy active root system. PGPR that contribute to the uptake of these nutrients have been reported and offer a novel solution for sustainable agriculture (Sheng, 2005; Gupta et al., 2015). Hormones, also known as plant growth regulators, can reg-ulate plant growth and development by stimulating or inhibiting plant
growth. PGPR produce various phytohomones including auxin, gibber-ellins, and cytokinins to enhance plant growth (Ruzzi and Aroca, 2015). Some PGPR possessing 1-aminocyclopropane-1-carboxylate (ACC) deam-inase activity are able to reduce the ethylene level to relieve plants from several forms of stress (Bal et al., 2013). Rhizobacteria, such as B. subtilis, B. megaterium, and P. chlororaphis, promote plant growth by producing vol-atiles like 2,3-butanediol, dimethylhexadecylamine, and 2-pentylfuran (Chung et al., 2010; Zou et al., 2010; Velázquez-Becerra et al., 2011).
The indirect plant growth stimulation by PGPR basically involves the ability of PGPR to reduce the deleterious effects of plant pathogens on crop yield. Competition for the rhizosphere nutrients and niches is a fundamental mechanism by which PGPR protect plants from pathogens ( Compant et al., 2005). Prior to infection by a plant pathogen, PGPR col-onization could trigger plant defense mechanisms to prevent or reduce the severity of the disease. This phenomenon is known as the induction of systemic resistance (ISR), which is activated by PGPR and functions throughout the plant. On the other hand, PGPR produce a wide variety of compounds with antimicrobial properties to defend itself against other mi-croorganisms including phytopathogens. Siderophores, bacteriocins and antibiotics are three of the most effective and well-known mechanisms that PGPR employ to minimize or prevent phytopathogenic proliferation (Beneduzi et al., 2012). Under iron-limiting conditions, PGPR produce low-molecular- weight chelating compounds called siderophores. The produc-tion of siderophores plays an important role in reducing phytopathogen proliferation by iron deprivation and enhancing plant development by in-creased uptake of iron (Masalha et al., 2000; Hibbing et al., 2010). A vari-ety of microorganisms also exhibit antagonistic activity against pathogens by excreting cell wall hydrolytic enzymes including chitinases, proteases,
Figure 2. Schematic diagram showing that plant growth-promoting bacteria affect plant growth directly and indirectly. ISR: induced systemic resistance.

11
14 15
Intr
oduc
tion
Rhiz
osph
ere
Baci
llus f
unct
ion
and
phyl
ogen
y
and β-1,3-glucanase to destroy the cell wall integrity of pathogens (Com-pant et al., 2005). It has long been known that bacteriocins and antibiotics are involved in shielding host against pathogens and mediating local pop-ulation dynamics in the rhizosphere (Thomashow et al., 2008; Subrama-nian and Smith, 2015). B. clausii GM17 isolated from the rhizosphere of On-onis angustissima have been found to produce bacteriocin Bac-GM17 with bactericidal effect on Agrobacterium tumefaciens and fungistatic effect on Candida tropicalis (Mouloud et al., 2013). The bacteriocin thuricin 17, iso-lated from B. thuringiensis NEB17, shows plant growth stimulation effects on plants (Lee et al., 2009). Another mechanism involved in disease sup-pression of PGPR is the production of antibiotics. Pseudomonads produce lipopeptides (LPs) such as 2,4-diacetyl phloroglucinol, hydrogen cyanide, pyrrolnitrin, tensin, pederin, tropolone, phenazine, that have been exten-sively studied (Gross and Loper, 2009). Bacillus is well-known to produce fengycin and surfactin family LPs. In addition, the production of zwitter-micin A and bacillomycin, has also been reported (Arguelles-Arias et al., 2009; Kinsella et al., 2009; Pérez-García et al., 2011).
RHIZOSPHERE BACILLUS FUNCTION AND PHYLOGENY
Bacillus spp. are well-known rhizosphere residents of many plants and may directly or indirectly contribute to crop productivity. By standard isolation on nutrition-rich medium, isolates phylogenetically related to B. subtilis and B. cereus have been frequently recovered (Bai et al., 2002; Cavaglieri et al., 2005; Cazorla et al., 2007; Egamberdieva et al., 2008). Pandey and Palni (1997) reported that Bacillus species are the dominant bacteria of the rhizosphere of established tea bushes. B. subtilis and B. my-coides comprise a major part of the bacterial population, even during un-favorable periods, also due to their ability to sporulate. Moreover, culture- independent rhizosphere microbiome studies have indicated the presence of a large uncultured diversity in Bacillus (Felske et al., 2003; Choudhary and Johri, 2009; Pereira et al., 2011). In a disease- suppressive soil, Bacillus was identified as the dominant bacterial species. The strain B. amylolique-faciens NJN-6 isolated from this suppressive soil decreased Panama dis-ease of banana by 68.5% (Xue et al., 2015). The rhizobacteria B. subtilis and B. amyloliquefaciens FZB42 have 4–5% and 8.5% of their genome, re-spectively, devoted to antibiotic synthesis (Stein, 2005; Chen et al., 2007). Among all the synthesized antimicrobial compounds by Bacillus, cyclic li-popeptides (LPs) of the surfactin, iturin and fengycin families have been well- characterized, with potential uses in biotechnology and agriculture (Ongena and Jacques, 2008). It has been shown that specific strains of
the species B. amyloliquefaciens, B. subtilis, B. pasteurii, B. cereus, B. pum-ilus, and B. mycoides elicit a significant reduction in the incidence or se-verity of various diseases on a diversity of plant hosts via ISR mechanisms ( Kloepper et al., 2004; Ryu et al., 2004).
Several studies have shown that Bacillus species are highly diverse, not only among different species but also between strains of the same spe-cies. Analyses of whole bacterial communities showed a significant dis-tinction between soil and rhizosphere communities (Smalla et al., 2001). A significant difference was observed in the total and relative abundance of Bacillus- like sequences amplified from bulk soil and crop roots (Choudhary and Johri, 2009). Mavingui et al. (1992) studied the genetic and phenotypic diversity of 130 strains of B. polymyxa isolated from non- rhizosphere soil (32 strains), rhizosphere soil (38 strains), and the rhizoplane (60 strains). Their results showed that diversity within populations of B. polymyxa iso-lated from non-rhizosphere and rhizosphere soil is higher than that of B. polymyxa isolated from the rhizoplane. This phenomenon can be ex-plained by the selection effects of plant roots on particular strains of a spe-cies. On the other hand, the strains living in association with plants are well adapted to the specific niche and did evolve some beneficial features for the host. Comparative genomics revealed a core set of genes that might be found in beneficial B. amyloliquefaciens strain. The phylogenetic tree based on concatenated sequences of 11 conserved genes of various other Bacillus strain genomes, results in a plant- associated clade (Figure 3A) (Magno-Perez-Bryan et al., 2015). Phylogenomic analysis of B. subtilis and B. amyloliquefaciens indicated that strains isolated from plant-associated (PA) habitats could be clearly distinguished from those from non-plant- associated (nPA) niches in both species (Figure 3B). Furthermore, the core genomes of PA strains are more abundant in genes relevant to interme-diate metabolism and secondary metabolite biosynthesis, as compared to those of nPA strains. Moreover, they possess specific additional genes in-volved in the utilization of plant-derived substrates and the synthesis of antibiotics (Zhang et al., 2016).
ENDOPHYTIC BACILLUS SPP.
As soil-borne microorganism, the species of Bacillus is not only found in the rhizosphere but also in the endosphere of plants. Endophytic bacte-ria are defined as bacteria colonizing the internal tissues of plants with-out causing symptoms of infection or negative effects on their host (Col-lins et al., 2004). According to their life strategy and the dependency on the host, endophytic bacteria can be classified as obligate endophytes,

11
16 17
Intr
oduc
tion
Endo
phyt
ic B
acill
us sp
p.
facultative endophytes or opportunistic endophytes. Obligate endophytes depend entirely on their host for all of their needs and spend all of their life within the host. In contrast, facultative and opportunistic endophytes have a free-living stage outside the plant host (Hardoim et al., 2008). Once the mutualistic symbiosis has been established, the plants provide a uniform, nutrient-rich, and non-competitive niche for endophytes. In turn, the endophytic bacteria bring beneficial attributes to their hosts, as they enhance plant growth and health in similar ways as PGPR. For exam-ple, direct plant growth promotion can be achieved by producing plant growth regulators including auxins, cytokinins, and gibberellings, reduc-ing ethylene stress by 1-aminocyclopropane-1-carboxylate (ACC) deami-nase; or fixing nitrogen or solubilizing phosphorus to aid plant nutrient acquisition(Rosenblueth and Martínez-Romero, 2006; Reinhold-Hurek and Hurek, 2011). Endophytic bacteria also indirectly benefit plant growth by the production of antimicrobial compounds and the induction of plant defense mechanisms (Weyens et al., 2009). Moreover, endophytic bacteria can help the host to cope with extreme biotic (pathogen, pest) and abiotic (temperature, drought, and heavy metals contaminations) stresses, which can severely reduce crop production. When applied as plant growth pro-moting or biocontrol agents, endophytic bacteria have advantages over rhizosphere bacteria due to their close interaction with the plant. For the seed-transmitted endophytes, there is an extra advantage for commer-cialization. Seed treatment is an efficient delivery technique to place mi-crobial inocula into the soil, where they will be well positioned to colonize seedling roots and protect against soil-borne diseases (Card et al., 2016).
The dominant endophytic bacteria belong to three major phyla i) Prote-obacteria, including Azorhizobium, Rhizobium, Enterobacter, Pseudomonas, and Burkholderia (Rosenblueth and Martínez-Romero, 2006; Taghavi et al., 2010; Dudeja et al., 2012; Ali et al., 2014); ii) Actinobacteria e.g., Microbac-terium, Streptomyces, Curtobacterium, and Arthrobacter (Conn and Franco, 2004; Chung et al., 2010; Verma et al., 2011); and iii) Firmicutes, including Bacillus and Staphylococcus (Luo et al., 2012; Phukon et al., 2013). Species of these genera are ubiquitous in the soil/rhizosphere, which represents the main source of endophytic colonizers. Among them, the genus Ba-cillus stands out as one of most reported endophytic bacteria from var-ious plant species. The spore-forming property with additional features of being natural soil dwellers makes Bacillus suitable for further devel-opment into commercial biocontrol agents. A recent metagenomics study revealed the dominance of Bacillus as a major endophytic genus in rice roots, probably playing a key role in nitrogen fixation (Sengupta et al., 2017). Bodhankar et al. (2017) isolated 80 seed endophytic bacte-ria from 30 maize genotypes. Their results showed that Bacillus was the
Figure 3. Phylogenetic analysis of several Bacillus strains. (A) Phylogenetic tree of iso-lates closely related to species of B. amyloliquefaciens, B. subtilis, B. atrophaeus, and B. li-cheniformis. 11 concatenated genes (nusA, rpoA, dnaA, rpoB, gyrA, gyrB, rpoC, spoVG, sigW, sigH, and sigB) were handled to build a neighbor-joining tree, using MEGA 5 bootstrap values (10,000 repetitions), which are shown on the branches. (B) The maximum-likeli-hood phylogeny derived from the alignment of 1835 concatenation core genes of B. subtilis and B. amyloliquefaciens. Leaf clip art indicates an association with plants. Colors indicate the biogeographic origin of the strains (PA: plant-associated; nPA: non-plant-associated). (Images modified from Magno-Perez-Bryan et al. (2015); Zhang et al. (2016))

11
18 19
Intr
oduc
tion
A g
ener
al in
trod
uctio
n on
B. m
ycoi
des
most dominant encountered genus affiliated with the Phylum Firmicutes. To date, the species that have been demonstrated to be endophytes are mainly distributed among B. subtilis, B. amyloliquefaciens, B. mojavensis, B. cereus, B. thuringiensis, B. mycoides, B. velezensis, B. pumilus, B. megaterium, and Paenibacillus, with a few reports on B. altitudinis, B. capparidis, and B. endoradicis. Most of the endophytes exhibit at least one beneficial func-tion on plants, including growth promotion (Falcao et al., 2014), ISR (Yi et al., 2013), antifungal activity (Gond et al., 2015), antibacterial activity (Jasim et al., 2016), phytomediation (Bisht et al., 2014), and stress toler-ance (Gagne-Bourque et al., 2016).
A GENERAL INTRODUCTION ON B. MYCOIDES
Bacillus mycoides is a Gram positive, rod-shaped, and spore-forming bac-terium. The growth of B. mycoides on agar plates has a particular rhizoid shape, resulting from cells linked end to end, and grouped in filaments bundles (Figure 4). It belongs to the B. cereus species-group, which com-prises several closely related species including B. anthracis, B. cereus, B. thuringiensis, B. mycoides, B. weihenstephanensis, B. pseudomycoides and some recently identified species (Fiedoruk et al., 2017). However, B. my-coides received far less attention compared to other members, since it is neither pathogenic, as B. cereus and B. anthracis, nor pesticidal as B. thur-ingiensis. To date, there are more than twenty B. mycoides strains that have been sequenced, with genome sizes ranging from 5.17 megabases (mb) to 6.55 mb. All the sequenced strains contain several plasmids of various sizes. The type strain ATCC6462 has one chromosome of 5.25 mb, one big plasmid, pBMX_1, of 360 kilobases (kb), and two other plasmids with sizes of about 10 kb (Figure 5). It is psychrotolerant with optimal growth be-tween 25 °C and 30 °C (Fiedoruk et al., 2017).
The economic and biological significance of B. mycoides is being gradu-ally recognized. For example, B. mycoides TKU038 isolated from soil is able to produce a novel chitosanase converting chitinous into chito-oligomers with antioxidant and anti-inflammatory potential (Liang et al., 2016). A B. mycoides strain isolated from an oil field produced a high level of biosur-factant (Najafi et al., 2010). Most of the studies on B. mycoides are focusing on its biocontrol and PGPR activity. B. mycoides Bac J (BmJ) isolated from sugar beet leaves reduces Cercospora leaf-spot disease of sugar beet up to 91% (Bargabus et al., 2002). The induced systemic-acquired resistance via the induction of pathogenesis-related proteins and oxidative burst was the main mechanism (Bargabus et al., 2003). This strain delayed the onset of anthracnose disease and reduced the total and live spore production
of the causing pathogen Glomerella cingulata var. orbiculare, when used to induce SAR in cucumber (Neher et al., 2009). Bacillus mycoides SU-23 could completely suppress the damping-off symptoms caused by Pythium mamillatum on cucumber and has growth promoting activity on seedlings (Paul et al., 1995). Application of a surfactin producing B. mycoides culture completely suppressed the formation of water-soaked lesions on cucum-ber leaves and reduced Pythium damping-off by 35% in the greenhouse (Peng et al., 2017). Nitrogen fixation activity of B. mycoides was reported (Ambrosini et al.), which makes this a strain with biofertilizer potential.
METHODS USED IN STUDYING PLANT-MICROBE INTERACTIONS
Genome sequencing and comparative genomics have a major impact on our understanding of the genetic potential, ecology, and evolution of microorganisms. Thanks to new methods developed over the past two decades, genome sequencing is now much faster and less expensive than it was before. There is an increasing number of rhizobacteria and endo-phytic bacteria genome sequences being published, and it is now pos-sible to get a detailed insight into the evolution and genetic adaptation of rhizobacteria by comparative genomics approaches. The genomes of plant- associated bacteria are relatively large and versatile, often compris-ing more than one chromosome and/or multiple plasmids. Endophytes and rhizobacteria can display a range of different life-styles associated with plants, differing in their time spent free-living in the soil (Hardoim et al., 2008). Such differences in life-style should be reflected in their ge-nomes. The presence of genes involved in motility and chemotaxis in the genome is implicating the strategy that rhizobacteria use to move towards the site of colonization. Many of the PGPR that have been sequenced are of interest because of their role in plant growth promotion. The genes
Figure 4. The growth of B. mycoides in (A) LB liquid medium & (B) LB agar plate; and its cell morphology observed by a phase contrast microscope (C).

11
20 21
Intr
oduc
tion
Met
hods
use
d in
stud
ying
pla
nt-m
icro
be in
tera
ctio
ns
for synthesis of indole-3-acetic acid (IAA), the major naturally occurring auxin, are found in various species (Kaneko et al., 2010; Wu et al., 2011). Other important PGPR-related genes are encoding ACC deaminase, vola-tile compounds, siderophores, as well as antimicrobials.
Transcriptome analysis is an efficient approach to study gene expres-sion during plant-microbe interactions at a genome-wide scale. Plant roots produce and secrete numerous small molecular weight compounds into the rhizosphere. These plant-derived extracellular metabolites and signals can influence the behavior of rhizosphere-associated bacteria via
the activation or suppression of specific gene expression. The expression of several genes from P. aeruginosa involved in metabolism, chemotaxis and type II secretion was regulated by sugar-beet root exudates (Mark et al., 2005). It has been suggested that the availability of particular metab-olites in root exudates, especially amino acids and aromatic compounds, support P. putida to colonize the rhizosphere, and the expression of several genes implicated in plant-bacteria interactions was upregulated upon con-tact with those compounds (Matilla et al., 2007). Fan et al. (2012) studied the B. amyloliquefaciens FZB42 transcriptomic profiles in response to maize root exudates. Their results showed that several groups of genes that were strongly induced by root exudates are involved in metabolic pathways relating to nutrient utilization, bacterial chemotaxis and motility, and non-ribosomal synthesis of antimicrobial peptides and polyketides.
Fluorescent proteins (FPs) are powerful tools for studying gene ex-pression, protein localization and cell motility. In the merging fields of microbial ecology and plant physiology, visualization of FP-labeled rhizo-bacteria is a key prerequisite to gain detailed insights into colonization behavior and plant-bacteria interaction mechanisms. One advantage of using FP marker systems for in situ studies is that they allow direct visuali-zation of the tagged bacteria at the single cell level, without the addition of exogenous substrates (unlike luciferase-based systems), in a nondestruc-tive way (Larrainzar et al., 2005). By using FP-labeled cells and confocal laser scanning microscopy, it is possible to visualize bacteria colonizing and growing along with a host plant. Such technology has been applied to monitor GFP-Rhizobium in association with the root at early stages of nodulation (Gage et al., 1996). Until now, the engineering and screening of FPs has resulted in more color variants, expanded from blue at 448 nm to far-red at 630 nm for multicolor imaging. When mixed populations of dif-ferent FP-labeled bacteria are used, inter-bacteria interactions can be ob-served. Four FPs including enhanced cyan (ECFP), enhanced green (EGFP), enhanced yellow (EYFP), and the red fluorescent protein (DsRed) can be used to simultaneously visualize different populations of pseudomonads in the rhizosphere (Bloemberg et al., 2000). The dual-labeling of wild-type versus mutant strains of bacteria will provide important insights whether a particular mutant is compromised for rhizosphere competence.
Genetic manipulation is a crucial prerequisite for detailed analysis of the genetic basis of the physiological and ecological properties of rhizo-bacteria. The development of efficient genetic manipulation tools has al-lowed to identify bacterial mechanisms and genes involved in the PGPR effects. Such tools offer possibilities to improve the expression of useful traits into a certain bacterium or to remove one or more genes of undesir-able traits. In laboratory conditions, many plant-associated bacteria have
Figure 5. Circular representations of the B. mycoides ATCC6462 genome displaying rel-evant genome features. From the inner to the outer concentric circle: circle 1, genomic position; circle 2, GC skew. the origin of replication was clearly detectable by a bias of G toward the leading strand; circles 3, GC content; circles 4 and 5, predicted protein- coding sequences (CDS) on the forward and the reverse strand.

11
22 23
Intr
oduc
tion
Scop
e of
this
thes
is
been identified with PGPR or biocontrol properties. However, the effec-tiveness varied greatly from test to test when used as inoculants in field conditions, due to the difficulty in maintaining them in the rhizosphere. Thus, improving bacterial root colonization through genetic modifica-tion should be considered. The introduction of a DNA fragment contain-ing the sss (site-specific recombinase) gene into strain P. fluorescens F113 and WCS307 greatly increased its root tip colonization ability (Dekkers et al., 2000). It is well-known that many lepidopteran insects are suscep-tible to endotoxins produced by B. thuringiensis. However, this bacterium has a short field-life. When the Bt toxin is produced in P. fluorescens, it can be encapsulated and retain its effectiveness for two- to three times longer than other Bt formulations (Peng et al., 2003). Molecular genetic studies also enable researchers to analyze the relevance of antibiotic production by plant-associated bacteria in the biological control of deleterious bac-teria and fungi (Lindow et al., 1989). Antimicrobial deficient mutants of several bacteria showed a reduced biocontrol activity (Girard et al., 2006; Diego Romero et al., 2007).
SCOPE OF THIS THESIS
In this thesis, we use various approaches to investigate the role of B. my-coides in the rhizosphere and the molecular basis of its plant-associated life style. Preliminary studies by our collaborators have shown that there is an inter-strain variation among B. mycoides species. A large number of strains were isolated from either the endosphere of healthy potato roots or from bulk soil. In chapter 2, we sequenced the genomes of seven B. my-coides strains with four isolated from endosphere and three isolated from bulk soil. Their plant-associated or non-plant-associated phenotype was confirmed by plant inoculation assays. Comparative genomics showed that there is a putative “endophytic clade” in the species of B. mycoides. In chapter 3, we subsequently selected one representative of the endophytic strains and one of the soil strains and compared their plant colonization ability and transcriptome profile in response to potato root exudates. The results showed that the endophytic strain has a more profound transcrip-tional response to root exudates than the soil strain, which might explain its good rhizosphere fitness.
For environmental isolates, the lack of genetic manipulation tools is the bottleneck for deep molecular genetics studies. In chapter 4, we devel-oped an efficient electroporation method for B. mycoides to facilitate fur-ther molecular genetics studies on this species. Strong fluorescent proteins are needed for in-planta visualization of rhizobacteria. In chapter 5, we
optimized GFP and RFP for B. mycoides with highly improved brightness. In chapter 6, we developed a CRISPR/Cas9 genome editing system for rhizos-phere isolated B. mycoides and B. subtilis. Gene deletion and insertion were performed to explore their plant-microbe interactions. These tools will fa-cilitate future mechanistic studies on bacteria-plant interactions.
REFERENCES
Abdel-Lateif, K., Bogusz, D., and Hocher, V.
(2012) The role of flavonoids in the estab-
lishment of plant roots endosymbioses
with arbuscular mycorrhiza fungi, rhizo-
bia and Frankia bacteria. Plant Signal. Be-
hav. 7: 636–641.
Ahemad, M., and Kibret, M. (2014) Mecha-
nisms and applications of plant growth
promoting rhizobacteria: current perspec-
tive. J. King. Saud. Univ. Sci. 26: 1–20.
Ali, S., Duan, J., Charles, T.C., and Glick, B.R.
(2014) A bioinformatics approach to the
determination of genes involved in en-
dophytic behavior in Burkholderia spp. J.
Theor. Biol. 343: 193–198.
Allard-Massicotte, R., Tessier, L., Lécuyer, F.,
Lakshmanan, V., Lucier, J.-F., Garneau, D.
et al. (2016) Bacillus subtilis early coloniza-
tion of arabidopsis thaliana roots involves
multiple chemotaxis receptors. MBio 7:
e01664–01616.
Ambrosini, A., Stefanski, T., Lisboa, B.B., Bene-
duzi, A., Vargas, L.K., and Passaglia, L.M.P.
(2016) Diazotrophic bacilli isolated from
the sunflower rhizosphere and the poten-
tial of Bacillus mycoides B38V as biofer-
tiliser. Ann. Appl. Biol. 168: 93–110.
Arguelles-Arias, A., Ongena, M., Halimi, B.,
Lara, Y., Brans, A., Joris, B., and Fickers, P.
(2009) Bacillus amyloliquefaciens GA1 as a
source of potent antibiotics and other sec-
ondary metabolites for biocontrol of plant
pathogens. Microb. Cell Fact. 8: 63.
Badri, D.V., and Vivanco, J.M. (2009) Regula-
tion and function of root exudates. Plant
Cell Environ. 32: 666–681.
Bai, Y., D’Aoust, F., Smith, D.L., and Driscoll, B.T.
(2002) Isolation of plant-growth-promot-
ing Bacillus strains from soybean root nod-
ules. Can. J. Microbiol. 48: 230–238.
Bais, H.P., Weir, T.L., Perry, L.G., Gilroy, S., and
Vivanco, J.M. (2006) The role of root ex-
udates in rhizosphere interactions with
plants and other organisms. Annu. Rev.
Plant Biol. 57: 233–266.
Bal, H.B., Nayak, L., Das, S., and Adhya, T.K.
(2013) Isolation of ACC deaminase produc-
ing PGPR from rice rhizosphere and evalu-
ating their plant growth promoting activ-
ity under salt stress. Plant Soil 366: 93–105.
Bargabus, R., Zidack, N., Sherwood, J., and Ja-
cobsen, B. (2002) Characterisation of sys-
temic resistance in sugar beet elicited by a
non-pathogenic, phyllosphere-colonizing
Bacillus mycoides, biological control agent.
Physiol. Mol. Plant Pathol. 61: 289–298.
Bargabus, R.L., Zidack, N.K., Sherwood, J.E., and
Jacobsen, B.J. (2003) Oxidative burst elic-
ited by Bacillus mycoides isolate Bac J, a bio-
logical control agent, occurs independently
of hypersensitive cell death in sugar beet.
Mol. Plant Microbe Interact. 16 1145–1153.
Beattie, G. (2006) Plant-associated bacteria:
survey, molecular phylogeny, genomics
and recent advances. Plant-associated bac-
teria: 1–56. Springer, Dordrecht.

11
24 25
Intr
oduc
tion
Refe
renc
es
Beauregard, P.B., Chai, Y., Vlamakis, H., Losick, R.,
and Kolter, R. (2013) Bacillus subtilis biofilm
induction by plant polysaccharides. Proc.
Natl. Acad. Sci. U. S. A. 110: E1621–E1630.
Beneduzi, A., Ambrosini, A., and Passaglia,
L.M.P. (2012) Plant growth-promoting
rhizobacteria (PGPR): Their potential as
antagonists and biocontrol agents. Genet.
Mol. Biol. 35: 1044–1051.
Bloemberg, G.V., Wijfjes, A.H., Lamers, G.E.,
Stuurman, N., and Lugtenberg, B.J. (2000)
Simultaneous imaging of Pseudomonas
fluorescens WCS365 populations express-
ing three different autofluorescent pro-
teins in the rhizosphere: new perspectives
for studying microbial communities. Mol.
Plant Microbe Interact. 13: 1170–1176.
Bodhankar, S., Grover, M., Hemanth, S., Reddy,
G., Rasul, S., Yadav, S.K. et al. (2017) Maize
seed endophytic bacteria: dominance of
antagonistic, lytic enzyme-producing Ba-
cillus spp. 3. Biotech. 7: 232.
Card, S., Johnson, L., Teasdale, S., and Cara-
dus, J. (2016) Deciphering endophyte be-
haviour: the link between endophyte bi-
ology and efficacious biological control
agents. FEMS Microbiol. Ecol. 92.
Carvalhais, L.C., Dennis, P.G., Badri, D.V., Kidd,
B.N., Vivanco, J.M., and Schenk, P.M.
(2015) Linking jasmonic acid signaling,
root exudates, and rhizosphere micro-
biomes. Mol. Plant Microbe Interact. 28:
1049–1058.
Cavaglieri, L., Orlando, J., Rodriguez, M.,
Chulze, S., and Etcheverry, M. (2005) Bio-
control of Bacillus subtilis against Fusar-
ium verticillioides in vitro and at the maize
root level. Res. Microbiol. 156: 748–754.
Cazorla, F., Romero, D., Pérez-García, A., Lugten-
berg, B., Vicente, A.d., and Bloemberg, G.
(2007) Isolation and characterization of an-
tagonistic Bacillus subtilis strains from the
avocado rhizoplane displaying biocontrol
activity. J. Appl. Microbiol. 103: 1950–1959.
Chaparro, J.M., Badri, D.V., Bakker, M.G., Sugi-
yama, A., Manter, D.K., and Vivanco, J.M.
(2013) Root exudation of phytochemicals
in Arabidopsis follows specific patterns
that are developmentally programmed
and correlate with soil microbial func-
tions. PloS ONE 8: e55731.
Chen, X.H., Koumoutsi, A., Scholz, R., Eisenreich,
A., Schneider, K., Heinemeyer, I. et al. (2007)
Comparative analysis of the complete ge-
nome sequence of the plant growth-pro-
moting bacterium Bacillus amyloliquefaciens
FZB42. Nat. Biotechnol. 25: 1007–1014.
Choudhary, D.K., and Johri, B.N. (2009) Inter-
actions of Bacillus spp. and plants—with
special reference to induced systemic re-
sistance (ISR). Microbiol. Res. 164: 493–513.
Chung, E.-J., Park, J.-H., Park, T.-S., Ahn, J.-W.,
and Chung, Y.-R. (2010) Production of a
phytotoxic compound, 3-phenylpropionic
acid by a bacterial endophyte, Arthrobacter
humicola YC6002 isolated from the root of
Zoysia japonica. Plant Pathol. J. 26: 245–252.
Collins, M.D., Hoyles, L., Foster, G., and Falsen,
E. (2004) Corynebacterium caspium sp. nov.,
from a Caspian seal (Phoca caspica). Int. J.
Syst. Evol. Microbiol. 54: 925–928.
Compant, S., Duffy, B., Nowak, J., Clement,
C., and Barka, E.A. (2005) Use of plant
growth-promoting bacteria for biocontrol
of plant diseases: principles, mechanisms
of action, and future prospects. Appl. Envi-
ron. Microbiol. 71: 4951–4959.
Conn, V.M., and Franco, C.M. (2004) Analysis
of the endophytic actinobacterial popu-
lation in the roots of wheat (Triticum aes-
tivum L.) by terminal restriction fragment
length polymorphism and sequencing of
16S rRNA clones. Appl. Environ. Microbiol.
70: 1787–1794.
Danhorn, T., and Fuqua, C. (2007) Biofilm for-
mation by plant-associated bacteria. Annu.
Rev. Microbiol. 61: 401–422.
De-la-Peña, C., Badri, D.V., Lei, Z., Watson, B.S.,
Brandão, M.M., Silva-Filho, M.C. et al.
(2010) Root secretion of defense-related
proteins is development-dependent and
correlated with flowering time. J. Biol.
Chem. 285: 30654–30665.
Dekkers, L.C., Mulders, I.H., Phoelich, C.C.,
Chin-A-Woeng, T.F., Wijfjes, A.H., and
Lugtenberg, B.J. (2000) The sss coloniza-
tion gene of the tomato-Fusarium oxyspo-
rum f. sp. radicis-lycopersici biocontrol
strain Pseudomonas fluorescens WCS365
can improve root colonization of other
wild-type Pseudomonas spp. bacteria. Mol.
Plant Microbe Interact. 13: 1177–1183.
Diego Romero, Eva Arrebola, Antonio de Vi-
cente, Francisco M. Cazorla, Rivo H. Ra-
kotoaly, Oscar P. Kuipers et al. (2007) The
iturin and fengycin families of lipopep-
tides are key factors in antagonism of Ba-
cillus subtilis toward Podosphaera fusca.
Mol. Plant Microbe Interact. 20: 430–440.
Doornbos, R.F., van Loon, L.C., and Bakker,
P.A.H.M. (2011) Impact of root exudates
and plant defense signaling on bacterial
communities in the rhizosphere. A review.
Agron. Sustain. Dev. 32: 227–243.
Dudeja, S., Giri, R., Saini, R., Suneja-Madan, P.,
and Kothe, E. (2012) Interaction of endo-
phytic microbes with legumes. J. Basic Mi-
crobiol. 52: 248–260.
Egamberdieva, D., Kamilova, F., Validov, S., Ga-
furova, L., Kucharova, Z., and Lugtenberg, B.
(2008) High incidence of plant growth-stim-
ulating bacteria associated with the rhizo-
sphere of wheat grown on salinated soil in
Uzbekistan. Environ. Microbiol. 10: 1–9.
Falcao, L.L., Silva-Werneck, J.O., Vilarinho, B.R.,
da Silva, J.P., Pomella, A.W., and Marcellino,
L.H. (2014) Antimicrobial and plant
growth-promoting properties of the cacao
endophyte Bacillus subtilis ALB629. J. Appl.
Microbiol. 116: 1584–1592.
Fan, B., Carvalhais, L.C., Becker, A., Fedosey-
enko, D., von Wirén, N., and Borriss, R.
(2012) Transcriptomic profiling of Bacil-
lus amyloliquefaciens FZB42 in response to
maize root exudates. Bmc Microbiol. 12: 116.
Felske, A.D., Heyrman, J., Balcaen, A., and De
Vos, P. (2003) Multiplex PCR screening of
soil isolates for novel Bacillus-related lin-
eages. J. Microbiol. Methods 55: 447–458.
Fiedoruk, K., Drewnowska, J.M., Daniluk,
T., Leszczynska, K., Iwaniuk, P., and
Swiecicka, I. (2017) Ribosomal back-
ground of the Bacillus cereus group ther-
motypes. Sci Rep 7: 46430.
Gage, D.J., Bobo, T., and Long, S.R. (1996) Use of
green fluorescent protein to visualize the
early events of symbiosis between Rhizo-
bium meliloti and alfalfa (Medicago sativa).
J. Bacteriol. 178: 7159–7166.
Girard, G., Barends, S., Rigali, S., van Rij, E.T.,
Lugtenberg, B.J., and Bloemberg, G.V.
(2006) Pip, a novel activator of phenazine
biosynthesis in Pseudomonas chlororaphis
PCL1391. J. Bacteriol. 188: 8283–8293.
Gond, S.K., Bergen, M.S., Torres, M.S., and
White Jr, J.F. (2015) Endophytic Bacillus
spp. produce antifungal lipopeptides and
induce host defence gene expression in
maize. Microbiol. Res. 172: 79–87.
González, J.F., Myers, M.P., and Venturi, V.
(2013) The inter-kingdom solo OryR reg-
ulator of Xanthomonas oryzae is important
for motility. Mol. Plant Pathol. 14: 211–221.
Greer-Phillips, S.E., Stephens, B.B., and Alex-
andre, G. (2004) An energy taxis trans-
ducer promotes root colonization by
Azospirillum brasilense. J. Bacteriol. 186:
6595–6604.

11
26 27
Intr
oduc
tion
Refe
renc
es
Gross, H., and Loper, J.E. (2009) Genomics of
secondary metabolite production by Pseu-
domonas spp. Nat. Prod. Rep. 26: 1408–1446.
Gupta, G., Parihar, S.S., Ahirwar, N.K., Snehi,
S.K., and Singh, V. (2015) Plant growth
promoting rhizobacteria (PGPR): current
and future prospects for development of
sustainable agriculture. J. Microb. Biochem.
Technol. 7: 96–102.
Hanif, M.K., Hameed, S., Imran, A., Naqqash, T.,
Shahid, M., and Van Elsas, J.D. (2015) Iso-
lation and characterization of a beta-pro-
peller gene containing phosphobacterium
Bacillus subtilis strain KPS-11 for growth
promotion of potato (Solanum tuberosum
L.). Front. Microbiol. 6: 583.
Hardoim, P.R., van Overbeek, L.S., and Elsas,
J.D. (2008) Properties of bacterial endo-
phytes and their proposed role in plant
growth. Trends Microbiol. 16: 463–471.
Hartmann, A., Rothballer, M., Hense, B.A., and
Schröder, P. (2014) Bacterial quorum sensing
compounds are important modulators of mi-
crobe-plant interactions. Front. Plant Sci. 5.131
Hibbing, M.E., Fuqua, C., Parsek, M.R., and Pe-
terson, S.B. (2010) Bacterial competition:
surviving and thriving in the microbial
jungle. Nat. Rev. Microbiol. 8: 15–25.
Huang, X.-F., Chaparro, J.M., Reardon, K.F.,
Zhang, R., Shen, Q., and Vivanco, J.M.
(2014) Rhizosphere interactions: root exu-
dates, microbes, and microbial communi-
ties. Botany 92: 267–275.
Islam, M.T., Hashidoko, Y., Deora, A., Ito, T., and
Tahara, S. (2005) Suppression of damp-
ing-off disease in host plants by the rhizo-
plane bacterium Lysobacter sp. strain SB-K88
is linked to plant colonization and antibiosis
against soilborne peronosporomycetes. Appl.
Environ. Microbiol. 71: 3786–3796.
Jasim, B., Sreelakshmi, S., Mathew, J., and Rad-
hakrishnan, E.K. (2016) Identification of
endophytic Bacillus mojavensis with highly
specialized broad spectrum antibacterial
activity. Biotech. 6: 187.
Jones, D.L., Nguyen, C., and Finlay, R.D. (2009)
Carbon flow in the rhizosphere: carbon
trading at the soil–root interface. Plant
Soil 321: 5–33.
Kamilova, F., Kravchenko, L.V., Shaposh-
nikov, A.I., Azarova, T., Makarova, N., and
Lugtenberg, B. (2006) Organic acids, sug-
ars, and L-tryptophane in exudates of veg-
etables growing on stonewool and their ef-
fects on activities of rhizosphere bacteria.
Mol. Plant Microbe Interact. 19: 250–256.
Kaneko, T., Minamisawa, K., Isawa, T., Nakat-
sukasa, H., Mitsui, H., Kawaharada, Y. et al.
(2010) Complete genomic structure of the
cultivated rice endophyte Azospirillum sp.
B510. DNA Res. 17: 37–50.
Kennedy, I.R., Choudhury, A., and Kecskés,
M.L. (2004) Non-symbiotic bacterial di-
azotrophs in crop-farming systems: can
their potential for plant growth promo-
tion be better exploited? Soil Biol. Biochem.
36: 1229–1244.
Kiers, E.T., Duhamel, M., Beesetty, Y., Men-
sah, J.A., Franken, O., Verbruggen, E. et al.
(2011) Reciprocal rewards stabilize coop-
eration in the mycorrhizal symbiosis. Sci-
ence 333: 880–882.
Kinsella, K., Schulthess, C.P., Morris, T.F., and
Stuart, J.D. (2009) Rapid quantification
of Bacillus subtilis antibiotics in the rhizo-
sphere. Soil Biol. Biochem. 41: 374–379.
Kloepper, J.W., Ryu, C.-M., and Zhang, S. (2004)
Induced systemic resistance and promo-
tion of plant growth by Bacillus spp. Phyto-
pathol. 94: 1259–1266.
Larrainzar, E., O’Gara, F., and Morrissey, J.P.
(2005) Applications of autofluorescent
proteins for in situ studies in microbial
ecology. Annu. Rev. Microbiol. 59: 257–277.
Lee, E.-H., Eo, J.-K., Ka, K.-H., and Eom, A.-H.
(2013) Diversity of arbuscular mycorrhizal
fungi and their roles in ecosystems. Myco-
biol. 41: 121–125.
Lee, K.D., Gray, E.J., Mabood, F., Jung, W.-J.,
Charles, T., Clark, S.R. et al. (2009) The
class IId bacteriocin thuricin-17 increases
plant growth. Planta 229: 747–755.
Liang, T.W., Chen, W.T., Lin, Z.H., Kuo, Y.H.,
Nguyen, A.D., Pan, P.S., and Wang, S.L.
(2016) An amphiprotic novel chitosanase
from Bacillus mycoides and its application
in the production of chitooligomers with
their antioxidant and anti-inflammatory
evaluation. Int. J. Mol. Sci. 17.
Lindow, S.E., Panopoulos, N.J., and McFarland,
B.L. (1989) Genetic engineering of bacte-
ria from managed and natural habitats.
Science 244: 1300–1307.
Lugtenberg, B., and Kamilova, F. (2009) Plant-
growth-promoting rhizobacteria Annu. Rev.
Microbiol. 63: 541–556.
Luo, S., Xu, T., Chen, L., Chen, J., Rao, C., Xiao,
X. et al. (2012) Endophyte-assisted promo-
tion of biomass production and metal-up-
take of energy crop sweet sorghum by
plant-growth-promoting endophyte Ba-
cillus sp. SLS18. Appl. Microbiol. Biotechnol.
93: 1745–1753.
Magno-Perez-Bryan, M.C., Martinez-Garcia, P.M.,
Hierrezuelo, J., Rodriguez-Palenzuela, P., Ar-
rebola, E., Ramos, C. et al. (2015) Compara-
tive genomics within the Bacillus genus re-
veal the singularities of two robust Bacillus
amyloliquefaciens biocontrol strains. Mol.
Plant Microbe Interact. 28: 1102–1116.
Mark, G.L., Dow, J.M., Kiely, P.D., Higgins, H.,
Haynes, J., Baysse, C. et al. (2005) Tran-
scriptome profiling of bacterial responses
to root exudates identifies genes involved
in microbe-plant interactions. Proc. Natl.
Acad. Sci. U. S. A. 102: 17454–17459.
Masalha, J., Kosegarten, H., Elmaci, Ö., and Men-
gel, K. (2000) The central role of microbial
activity for iron acquisition in maize and
sunflower. Biol. Fertil. Soil. 30: 433–439.
Matilla, M.A., Espinosa-Urgel, M., Rodriguez-
Herva, J.J., Ramos, J.L., and Ramos-
Gonzalez, M.I. (2007) Genomic analysis
reveals the major driving forces of bacterial
life in the rhizosphere. Genome Biol. 8: R179.
Mavingui, P., Laguerre, G., Berge, O., and Heu-
lin, T. (1992) Genetic and phenotypic diver-
sity of Bacillus polymyxa in soil and in the
wheat rhizosphere. Appl. Environ. Micro-
biol. 58: 1894–1903.
Mouloud, G., Daoud, H., Bassem, J., Atef, I.L.,
and Hani, B. (2013) New bacteriocin from
Bacillus clausii strain GM17: purification,
characterization, and biological activity.
Appl. Biochem. Biotechnol. 171: 2186–2200.
Najafi, A.R., Rahimpour, M.R., Jahanmiri, A.H.,
Roostaazad, R., Arabian, D., and Ghobadi,
Z. (2010) Enhancing biosurfactant produc-
tion from an indigenous strain of Bacillus
mycoides by optimizing the growth condi-
tions using a response surface methodol-
ogy. Chem. Eng. J. 163: 188–194.
Neher, O.T., Johnston, M.R., Zidack, N.K., and
Jacobsen, B.J. (2009) Evaluation of Bacillus
mycoides isolate BmJ and B. mojavensis iso-
late 203-7 for the control of anthracnose of
cucurbits caused by Glomerella cingulata
var. orbiculare. Biol. Control 48: 140–146.
Oku, S., Komatsu, A., Tajima, T., Nakashimada,
Y., and Kato, J. (2012) Identification of che-
motaxis sensory proteins for amino acids
in Pseudomonas fluorescens Pf0-1 and their
involvement in chemotaxis to tomato root
exudate and root colonization. Microbes
Environ. 27: 462–469.
Ongena, M., and Jacques, P. (2008) Bacillus lipo-
peptides: versatile weapons for plant disease
biocontrol. Trends Microbiol. 16: 115–125.

11
28 29
Intr
oduc
tion
Refe
renc
es
Pandey, A., and Palni, L.M.S. (1997) Bacillus
species: The dominant bacteria of the rhi-
zosphere of established tea bushes. Micro-
biol. Res. 152: 359–365.
Paul, B., Charles, R., and Bhatnagar, T. (1995)
Biological control of Pythium mamilla-
tum causing damping-off of cucumber
seedlings by a soil bacterium, Bacillus my-
coides. Microbiol. Res. 150: 71–75.
Peng, R., Xiong, A., Li, X., Fuan, H., and Yao, Q.
(2003) A delta-endotoxin encoded in Pseu-
domonas fluorescens displays a high degree
of insecticidal activity. Appl. Microbiol. Bio-
technol. 63: 300–306.
Peng, Y.-H., Chou, Y.-J., Liu, Y.-C., Jen, J.-F.,
Chung, K.-R., and Huang, J.-W. (2017) Inhi-
bition of cucumber Pythium damping-off
pathogen with zoosporicidal biosurfac-
tants produced by Bacillus mycoides. J.
Plant Dis. Protect. 124: 481–491.
Pereira, P., Ibáñez, F., Rosenblueth, M., Etch-
everry, M., and Martínez-Romero, E. (2011)
Analysis of the bacterial diversity associ-
ated with the roots of maize (Zea mays L.)
through culture-dependent and culture-in-
dependent methods. ISRN Ecology 2011.
Pérez-García, A., Romero, D., and de Vicente, A.
(2011) Plant protection and growth stimu-
lation by microorganisms: biotechnolog-
ical applications of Bacilli in agriculture.
Curr. Opin. Biotechnol. 22: 187–193.
Peters, W.S. (2004) Growth rate gradients and
extracellular pH in roots: how to control
an explosion. New Phytologist 162: 571–574.
Phukon, M., Sahu, P., Srinath, R., Nithya, A.,
and Babu, S. (2013) Unusual occurrence
of Staphylococcus warneri as endophyte in
fresh fruits along with usual Bacillus spp. J.
Food Safety 33: 102–106.
Pini, F., Frascella, A., Santopolo, L., Bazzicalupo,
M., Biondi, E.G., Scotti, C., and Mengoni,
A. (2012) Exploring the plant-associated
bacterial communities in Medicago sativa
L. BMC Microbiol. 12: 78.
Podile, A.R., and Kishore, G.K. (2007) Plant
growth-promoting rhizobacteria. In Plant-as-
sociated bacteria: Springer, pp. 195–230.
Reinhold-Hurek, B., and Hurek, T. (2011) Living
inside plants: bacterial endophytes. Curr.
Opin. Plant Biol. 14: 435–443.
Rohrbacher, F., and St-Arnaud, M. (2016) Root
exudation: the ecological driver of hydro-
carbon rhizoremediation. Agronomy 6: 19.
Rosenblueth, M., and Martínez-Romero, E.
(2006) Bacterial endophytes and their in-
teractions with hosts. Mol. Plant Microbe
Interact. 19: 827–837.
Rudrappa, T., Biedrzycki, M.L., and Bais, H.P.
(2008) Causes and consequences of plant-
associated biofilms. FEMS Microbiol. Ecol.
64: 153–166.
Ruzzi, M., and Aroca, R. (2015) Plant growth-pro-
moting rhizobacteria act as biostimulants
in horticulture. Sci. Hortic. 196: 124–134.
Ryu, C.M., Farag, M.A., Hu, C.H., Reddy, M.S.,
Kloepper, J.W., and Pare, P.W. (2004) Bacte-
rial volatiles induce systemic resistance in
Arabidopsis. Plant Physiol. 134: 1017–1026.
Sengupta, S., Ganguli, S., and Singh, P.K. (2017)
Metagenome analysis of the root endo-
phytic microbial community of Indian rice
(O. sativa L.). Genomics Data 12: 41–43.
Sheng, X. (2005) Growth promotion and in-
creased potassium uptake of cotton and
rape by a potassium releasing strain of
Bacillus edaphicus. Soil Biol. Biochem. 37:
1918–1922.
Smalla, K., Wieland, G., Buchner, A., Zock, A.,
Parzy, J., Kaiser, S. et al. (2001) Bulk and
rhizosphere soil bacterial communities
studied by denaturing gradient gel elec-
trophoresis: plant-dependent enrichment
and seasonal shifts revealed. Appl. Environ.
Microbiol. 67: 4742–4751.
Stein, T. (2005) Bacillus subtilis antibiotics:
structures, syntheses and specific func-
tions. Mol. Microbiol. 56: 845–857.
Subramanian, S., and Smith, D.L. (2015) Bac-
teriocins from the rhizosphere microbi-
ome — from an agriculture perspective.
Front. Plant Sci. 6: 909.
Subramoni, S., Gonzalez, J.F., Johnson, A.,
Pechy-Tarr, M., Rochat, L., Paulsen, I. et al.
(2011) Bacterial subfamily of LuxR regu-
lators that respond to plant compounds.
Appl. Environ. Microbiol. 77: 4579–4588.
Sugiyama, A., and Yazaki, K. (2012) Root exu-
dates of legume plants and their involve-
ment in interactions with soil microbes.
In Secretions and exudates in biological sys-
tems: Springer, pp. 27–8.
Taghavi, S., van der Lelie, D., Hoffman, A., Zhang,
Y.B., Walla, M.D., Vangronsveld, J. et al. (2010)
Genome sequence of the plant growth pro-
moting endophytic bacterium Enterobacter
sp. 638. PLoS Genet. 6: e1000943.
Thomashow, L.S., Bonsall, R.F., and Weller,
D.M. (2008) Detection of antibiotics pro-
duced by soil and rhizosphere microbes in
situ. Secondary metabolites in soil ecology:
23–36. Springer, Berlin, Heidelberg
Velázquez-Becerra, C., Macías-Rodríguez, L.I.,
López-Bucio, J., Altamirano-Hernández,
J., Flores-Cortez, I., and Valencia-Cantero,
E. (2011) A volatile organic compound
analysis from Arthrobacter agilis identi-
fies dimethylhexadecylamine, an amino-
containing lipid modulating bacterial
growth and Medicago sativa morphogene-
sis in vitro. Plant Soil 339: 329–340.
Venturi, V., and Fuqua, C. (2013) Chemical signal-
ing between plants and plant-pathogenic
bacteria. Annu. Rev. Phytopathol. 51: 17–37.
Verma, V., Singh, S., and Prakash, S. (2011)
Bio-control and plant growth promo-
tion potential of siderophore producing
endophytic Streptomyces from Azadirachta
indica. J. Basic Microbiol. 51: 550–556.
Weyens, N., van der Lelie, D., Taghavi, S., and
Vangronsveld, J. (2009) Phytoremediation:
plant-endophyte partnerships take the chal-
lenge. Curr. Opin. Biotechnol. 20: 248–254.
Wu, X., Monchy, S., Taghavi, S., Zhu, W., Ramos,
J., and van der Lelie, D. (2011) Compara-
tive genomics and functional analysis of
niche-specific adaptation in Pseudomonas
putida. FEMS Microbiol. Rev. 35: 299–323.
Xue, C., Penton, C.R., Shen, Z., Zhang, R.,
Huang, Q., Li, R. et al. (2015) Manipulating
the banana rhizosphere microbiome for
biological control of Panama disease. Sci.
Rep. 5: 11124
Yi, H.-S., Yang, J.W., and Ryu, C.-M. (2013) ISR
meets SAR outside: additive action of the
endophyte Bacillus pumilus INR7 and the
chemical inducer, benzothiadiazole, on in-
duced resistance against bacterial spot in
field-grown pepper. Front. Plant Sci. 4: 122.
Yoneyama, K., Xie, X., Sekimoto, H., Takeuchi,
Y., Ogasawara, S., Akiyama, K. et al. (2008)
Strigolactones, host recognition signals
for root parasitic plants and arbuscular
mycorrhizal fungi, from Fabaceae plants.
New Phytologist 179: 484–494.
Zahran, H.H. (2001) Rhizobia from wild le-
gumes: diversity, taxonomy, ecology, ni-
trogen fixation and biotechnology. J. Bio-
technol. 91: 143–153.
Zhang, N., Yang, D., Kendall, J.R., Borriss, R., Dru-
zhinina, I.S., Kubicek, C.P. et al. (2016) Com-
parative genomic analysis of Bacillus am-
yloliquefaciens and Bacillus subtilis reveals
evolutional traits for adaptation to plant-as-
sociated habitats. Front. Microbiol. 7: 2039.
Zou, C., Li, Z., and Yu, D. (2010) Bacillus me-
gaterium strain XTBG34 promotes plant
growth by producing 2-pentylfuran. J. Mi-
crobiol. 48: 460–466.

2Draft genome sequences of seven Bacillus mycoides strains, isolated from potato endosphere and soil
Yanglei Yia, Anne de Jonga, Jan Spoelderb,d, J. Theo M. Elzengab, Jan Dirk van Elsasc, Oscar P. Kuipersa
Molecular Genetics, Groningen Biomolecular Sciences and Biotechnology Institute, University of Groningen, Groningen, the Netherlandsa. Plant Physiology, Groningen Institute for Evolutionary Life Sciences, University of Groningen, Groningen, the Netherlandsb. Microbial Ecology, Groningen Institute for Evolutionary Life Sciences, University of Groningen, Groningen, the Netherlandsc.HLB Research & Consultancy in Agriculture, Kampsweg 27, 9418 PD Wijster, the Netherlandsd.
This chapter is partly based on:Yi, Y., de Jong, A., Spoelder, J., Elzenga, J. T. M., van Elsas, J. D., & Kuipers, O. P. (2016). Draft genome sequence of Bacillus mycoides M2E15, a strain isolated from the endosphere of potato. Genome announcements, 4(1), e00031–16.

2
33
Dra
ft g
enom
e se
quen
ces o
f sev
en B
acill
us m
ycoi
des s
trai
ns,
isol
ated
from
pot
ato
endo
sphe
re a
nd so
il
Bacillus mycoides is a rod-shaped and spore-forming gram-positive bac-terium which is associated with the Bacillus cereus group. On agar plates, it forms unique rhizoid colonies resulting from cells that are linked end-to-end. The strong intra-cellular connections yield a stable network that favors interactions with other soil inhabitants, such as microbes and plant roots (Turchi et al., 2012). There are several studies showing that B. mycoides is able to promote the plant growth (Karagöz et al., 2012), fix N2 (Ambrosini et al., 2016), and control several plant diseases (Paul et al., 1995; Bargabus et al., 2002; Neher et al., 2009). We found some B. mycoides strains to be present in the endosphere of potato without causing visible signs of disease. In order to understand the genetic make-up of this bac-terium, we present the draft genome of several B. mycoides strains, which was isolated from potato endosphere or soil (Wijster, the Netherlands).
Strains were grown overnight in Luria-Bertani (LB) broth at 30 °C, 200 rpm. Bacterial cells were harvested at the exponential growth phase and lysed with lysozyme. After RNase treatment, proteins were removed by proteinase K digestion. DNA was extracted from the lysate by phe-nol-chloroform treatment and recovered by isopropanol precipitation. The extracted DNA was checked for purity by Nanodrop (Thermo Fisher Scientific) and 1% agarose gel, then quantified by Quantus™ Fluorom-eter (Promega). The high-quality DNA was sequenced using the MiSeq sequencing system of Illumina, yielding 250 bp paired-end reads with a mean library size of 400 bp. De novo assembly was performed using Vel-vet (Zerbino and Birney, 2008). Prediction of protein-encoding regions and automatic functional annotation was performed using the Rapid Annotations using Subsystem Technology (RAST) server (Aziz et al., 2008). Moreover, the bacteriocin identification tool Bagel3 was used (van Heel et al., 2013).
The whole genome alignment of the seven strains was performed with GEGENEES tool (Ågren et al. 2012). Phylogram was generated with Neigh-bour joining method and is shown in Figure 1. The resulting phylogenetic tree has two main branches that group the 29 genomes into two species clades: B. mycoides and other B. cereus-group members. For B. mycoides, we selected the 7 strains based on their origin of isolation and plant col-onization ability: four strains isolated from the endosphere that could be endophytic again (EC18, M2E15, S2E19, and S3E15), and three strains iso-lated from soil which could not be endophytic (SB4, SB8, and SB13). The phylogenetic tree showed that the four endophytic strains are clustered together, indicating that the endophytic strains have some common fea-tures. Intriguingly, the soil strain SB8 was also grouped in this putative endophytic clade.

22
34 35
Dra
ft g
enom
e se
quen
ces o
f sev
en B
acill
us m
ycoi
des s
trai
ns,
isol
ated
from
pot
ato
endo
sphe
re a
nd so
il
ACK
NO
WLE
DG
MEN
TS
Table 1. Genome features and GenBank accession numbers of the strains.
Species Strain Source of isolation Accession No.Bacillus mycoides EC18 Endosphere MRWW00000000.1Bacillus mycoides S2E19 Endosphere MRWV00000000.1Bacillus mycoides M2E15 Endosphere LLWA00000000.1Bacillus mycoides S3E15 Endosphere MRWU00000000.1Bacillus mycoides SB4 Soil MRZX00000000.1Bacillus mycoides SB8 Soil MRWS00000000.1Bacillus mycoides SB13 Soil MRWT00000000.1
Nucleotide sequence accession number. The genome sequences of the 7 B. mycoides strains have been deposited as whole genome shotgun projects at GenBank under the accession numbers listed in Table 1.
ACKNOWLEDGMENTS
Y.Yi was supported by a scholarship to from the China Scholarship Council (201306300040).
REFERENCES
Ågren, J., Sundström, A., Håfström, T., &
Segerman, B. (2012). Gegenees: frag-
mented alignment of multiple genomes
for determining phylogenomic distances
and genetic signatures unique for speci-
fied target groups. Plos One 7(6): e39107.
Ambrosini, A., Stefanski, T., Lisboa, B.B.,
Beneduzi, A., Vargas, L.K., and Passaglia,
L.M.P. (2016) Diazotrophic Bacilli isolated
from the sunflower rhizosphere and the
potential of Bacillus mycoides B38V as bio-
fertiliser. Ann Appl Biol 168: 93-110.
Aziz, R.K., Bartels, D., Best, A.A., DeJongh, M.,
Disz, T., Edwards, R.A. et al. (2008) The
RAST Server: rapid annotations using sub-
systems technology. BMC Genomics 9: 75.
Bargabus, R., Zidack, N., Sherwood, J., and Ja-
cobsen, B. (2002) Characterisation of sys-
temic resistance in sugar beet elicited by a
non-pathogenic, phyllosphere-colonizing
Bacillus mycoides, biological control agent.
Physiol Mol Plant Pathol 61: 289-298.
Karagöz, K., Ateş, F., Karagöz, H., Kotan, R., and
Çakmakçı, R. (2012) Characterization of
plant growth-promoting traits of bacteria
isolated from the rhizosphere of grape-
vine grown in alkaline and acidic soils. Eur
J Soil Biol 50: 144-150.
Neher, O.T., Johnston, M.R., Zidack, N.K., and
Jacobsen, B.J. (2009) Evaluation of Bacillus
mycoides isolate BmJ and B. mojavensis iso-
late 203-7 for the control of anthracnose
of cucurbits caused by Glomerella cingu-
lata var. orbiculare. Biological Control 48:
140-146.
Paul, B., Charles, R., and Bhatnagar, T. (1995)
Biological control of Pythium mamilla-
tum causing damping-off of cucumber
seedlings by a soil bacterium, Bacillus my-
coides. Microbiol Res 150: 71-75.
Figure 1. Phylogenetic analysis based on the whole genome sequences of the 7 B. my-coides isolates and other representatives of the B. cereus-group: B. cereus, B. anthracis, and B. thuringensis.
Turchi, L., Santini, T., Beccari, E., and Di Franco,
C. (2012) Localization of new peptidogly-
can at poles in Bacillus mycoides, a mem-
ber of the Bacillus cereus group. Arch Mi-
crobiol 194: 887-892.
van Heel, A.J., de Jong, A., Montalbán-López,
M., Kok, J., and Kuipers, O.P. (2013) BA-
GEL3: automated identification of genes
encoding bacteriocins and (non-) bacte-
ricidal posttranslationally modified pep-
tides. Nucleic Acids Res 41: W448-W453.
Zerbino, D.R., and Birney, E. (2008) Velvet: al-
gorithms for de novo short read assembly
using de Bruijn graphs. Genome Res 18:
821-829.

3Comparative transcriptomics of Bacillus mycoides strains in response to potato-root exudates reveals different genetic adaptation of endophytic and soil isolates
Yanglei Yi, Anne de Jong, Elrike Frenzel and Oscar P. Kuipers
Molecular Genetics Department, Groningen Biomolecular Sciences and Biotechnology Institute, University of Groningen, Groningen, Netherlands
This chapter was published as:Yi, Y., de Jong, A., Frenzel, E., & Kuipers, O. P. (2017). Comparative transcriptomics of Bacillus mycoides strains in response to potato-root exudates reveals different genetic adaptation of endophytic and soil isolates. Frontiers in Microbiology, 8: 1487.

3
39
Com
para
tive
tran
scri
ptom
ics o
f Bac
illus
myc
oide
s str
ains
in re
spon
se to
pot
ato-
root
ex
udat
es re
veal
s diff
eren
t gen
etic
ada
ptat
ion
of e
ndop
hytic
and
soil
isol
ates
ABSTRACT
Plant root secreted compounds alter the gene expression of associated microorganisms by acting as signal molecules that either stimulate or re-pel the interaction with beneficial or harmful species, respectively. How-ever, it is still unclear whether two distinct groups of beneficial bacte-ria, non-plant-associated (soil) strains and plant associated (endophytic) strains, respond uniformly or variably to the exposure with root exu-dates. Therefore, Bacillus mycoides, a potential biocontrol agent and plant growth-promoting bacterium, was isolated from the endosphere of pota-toes and from soil of the same geographical region. Confocal fluorescence microscopy of plants inoculated with GFP-tagged B. mycoides strains showed that the endosphere isolate EC18 had a stronger plant coloniza-tion ability and competed more successfully for the colonization sites than the soil isolate SB8. To dissect these phenotypic differences, the ge-nomes of the two strains were sequenced and the transcriptome response to potato root exudates was compared. The global transcriptome profiles evidenced that the endophytic isolate responded more pronounced than the soil-derived isolate and a higher number of significant differentially expressed genes were detected. Both isolates responded with the altera-tion of expression of an overlapping set of genes, which had previously been reported to be involved in plant–microbe interactions; including or-ganic substance metabolism, oxidative reduction, and transmembrane transport. Notably, several genes were specifically upregulated in the en-dosphere isolate EC18, while being oppositely downregulated in the soil isolate SB8. These genes mainly encoded membrane proteins, transcrip-tional regulators or were involved in amino acid metabolism and biosyn-thesis. By contrast, several genes upregulated in the soil isolate SB8 and downregulated in the endosphere isolate EC18 were related to sugar trans-port, which might coincide with the different nutrient availability in the two environments. Altogether, the presented transcriptome profiles pro-vide highly improved insights into the life strategies of plant-associated endophytes and soil isolates of B. mycoides.
INTRODUCTION
The rhizosphere harbors an enormous pool of soil microorganisms and is considered as the ‘hot spot’ for microbial colonization and activity (Prashar et al., 2013). Some of the rhizobacteria even have the capacity of multiplying inside roots and begin an endophytic lifestyle (Hardoim et al., 2008). Rhizobacteria and plants naturally interact in various ways. Ben-eficial plant– microbial interactions in the rhizosphere can result in the promotion of plant health and development (Market al., 2005). A number of plant growth-promoting rhizobacteria (PGPR) have been commercially used as adjuncts to agricultural practice and show great promise (Glick, 2012). The mechanisms of PGPR and plant root interaction have drawn considerable attention (Kuiper et al., 2004; Santos et al., 2014; Pangesti et al.,2015). However, only a few studies on plants and their interaction with endophytic bacteria have been reported so far.
Endophytic bacteria are defined as bacteria capable of colonizing the in-ternal tissue of the plant without harming the host or causing any disease symptoms (Mónica and Esperanza,2006). It has been shown that many endophytic bacteria can promote plant growth with a similar mechanism as rhizobacteria, e.g., phosphate solubilisation (Wakelin et al., 2004), in-dole acetic acid production (Lee et al., 2004; Jasim et al., 2014), and si-derophore production (Rosconi et al., 2013). A number of endophytic bac-teria are able to prevent the deleterious effects of certain pathogens by producing antimicrobial compounds or inducing systematic resistance in the host plant (Gómez-Lama Cabanás et al., 2014; Kandel et al., 2017). The beneficial effects of endophytes are often more pronounced than those of many rhizosphere-colonizing bacteria (Hardoim et al., 2008). A large diversity of bacterial species have the ability to adopt an endophytic lifestyle. These comprise more than 200 bacterial genera from 16 phyla, with Actinobacteria, Proteobacteria, and Firmicutes being the most pre-dominant genera (Malfanova et al.,2013; Jin et al., 2014). The rhizosphere- competent bacteria, including Azospirillum, Bacillus, and Pseudomonas genera, are most frequently reported as plant internal tissue residents. In fact, the endophytic bacterial diversity can be considered as a subset of the rhizosphere and/or the root-associated bacterial population ( Santoyo et al., 2016). This implies that only a limited number of members of the rhizosphere-competent bacteria can succeed in becoming endophytic. This is due to several factors: (1) plants can selectively recruit certain ben-eficial rhizobacteria (Lakshmanan et al., 2013); (2) only those rhizobacte-ria that are well-adapted for living inside plants and simultaneously show an aggressive colonization ability can become endophytes. It has been re-ported that several Rhizobium etli strains were preferentially encountered

33
40 41
Com
para
tive
tran
scri
ptom
ics o
f Bac
illus
myc
oide
s str
ains
in re
spon
se to
pot
ato-
root
ex
udat
es re
veal
s diff
eren
t gen
etic
ada
ptat
ion
of e
ndop
hytic
and
soil
isol
ates
INTR
OD
UCT
ION
as maize endophytes and the highly competitive strain Ch24-10 was the most tolerant to 6-methoxy-2-benzoxazolinone, a maize antimicrobial compound that is inhibitory to some bacteria and fungi (Rosenblueth and Martínez-Romero, 2004). Pseudomonas strains that were isolated from the endosphereare phenotypically distinct from Pseudomonas isolates ob-tained from the root surface as identified by their lipopolysaccharide pat-tern, cell envelope protein pattern, and other biochemical characteristics (van Peer et al., 1990). A comparative genomics study by Timm et al. (2015) showed that endosphere isolates of P. fluorescens have significantly more metabolic pathways for utilization of plant signaling compounds than rhizosphere isolates.
During initiation of root surface colonization by rhizosphere microor-ganisms, the first event in the interaction with plants is the encountering of root exudates (Shidore et al., 2012). Plant roots can exude up to 20% of fixed carbon into the soil. Amino acids, organic acids, sugars, aromat-ics, and various other secondary metabolites comprise the majority of the low molecular-weight root exudates, whereas high-molecular-weight exudates primarily include polysaccharides and proteins (Brencic and Winans, 2005). Microbes can use these compounds as carbon- and energy sources for growth and development. Apart from being an exogenous nu-trition, some molecules from exudates may act as signals to alter specific gene expression patterns in the microbe, which might influence its inter-action with the host (Mark et al., 2005). For example, the plant-released molecules, including flavonoids, stachydrine, and trigonelline are recog-nized as signals for induction of nodulation genes in various legume-as-sociated bacteria (Phillips et al., 1992; Janczareket al., 2015). The citric acid in cucumber exudates was shown to recruit B. amyloliquefaciens SQR9 and to induce its biofilm formation (Zhang et al., 2014). Root colonization by endophytic bacteria is hypothesized to follow a similar process as de-scribed for rhizosphere bacteria. It has been shown that plant compounds from leaf macerates can activate a LuxR family transcription factor and its regulated genes of the cottonwood tree endophyte Pseudomonas sp. GM79 (Schaefer et al., 2016). Thus, molecules released from plants might play an important role in the plant–endophyte interaction.
Bacillus mycoides is a spore-forming, gram-positive bacterium belong-ing to the B. cereus sensu lato group. Among this group, B. mycoides is prob-ably the least recognized and un-researched species, since it is in contrast to B. cereus and B. anthracis not pathogenic and does not have insecticidal activity as for instance B. thuringiensis. On agar plates, B. mycoides forms unique rhizoid colonies with chains of cells forming filaments projecting radially and curving to the left or the right (Di Franco et al.,2002). B. mycoides is ubiquitous in the soil and the rhizosphere. Some isolates have beneficial
plant growth promoting (PGP) and biocontrol activity in various plants, in-cluding sunflower (Ambrosini et al., 2016), cucumber (Neher et al., 2009), and sugar beet (Bargabus et al., 2002). Our research showed that certain B. mycoides strains were able to endophytically colonize potato roots with po-tential PGP activity (unpublished data). However, the mechanisms of the beneficial interaction between the host and this endophyte are not known.
Transcriptomics is a powerful approach to investigate plant–microbe interactions, but in situ transcriptomics of plant–microbe interaction models has limitations due to the imbalances of biomass ratios. As an al-ternative, root exudates or similar simplified systems are therefore used to study the bacterial response in the symbiotic interaction models. Root exudates were shown to have profound effects on gene expression pat-terns of rhizosphere Bacilli including B. amyloliquefaciens FZB42 (Fan et al., 2012) and B. amyloliquefaciens SQR9 (Zhang et al., 2015). A transcriptome analysis conducted with the endophytic bacterium Azoarcus sp. strain BH72 and Oryza sativa root exudates, identified several genes required for rhizosphere competence (Shidore et al., 2012). Taghavi et al. (2015) used sucrose as sole carbon source to mimic the plant-associated lifestyle of Enterobacter sp. 638.Their transcriptome study revealed that the small RNA csrABCD regulon plays a role in the physiological adaptation and possibly mediates the shift between the free-living and a plant-associated endophytic lifestyle. In this study, we compared the transcriptome pro-files of an endophytic strain and a soil strain of B. mycoides in response to potato root exudates. The aim was to investigate differences in the ge-netic responses of the endophytic and the soil isolate and to decipher the endophytic determinants of B. mycoides. The colonization ability of the endophyte strain EC18 and the soil strain SB8 was tested by plant inocu-lation assays and in situ fluorescence microscopy monitoring. The whole genome of the two strains was sequenced and used as a reference for the transcriptome analysis. B. mycoides strain EC18 and SB8 were grown in the presence or absence of root exudates of potato plants, and differences in their gene expression profiles were analyzed by RNA-seq. Some genes with specific functionality showed a distinct trend between endophyte and soil strain and may be considered as key factors determining the en-dophytic lifestyle of B. mycoides.
MATERIALS AND METHODS
Strains and Culture Condition Bacillus mycoides EC18 was isolated from internal root tissue of the potato cultivar Seresta during the early flowering stage, grown on a practice field

33
42 43
Com
para
tive
tran
scri
ptom
ics o
f Bac
illus
myc
oide
s str
ains
in re
spon
se to
pot
ato-
root
ex
udat
es re
veal
s diff
eren
t gen
etic
ada
ptat
ion
of e
ndop
hytic
and
soil
isol
ates
MAT
ERIA
LS A
ND
MET
HO
DS
in a sandy soil at Wijster, the Netherlands. The strain SB8 was isolated from bulk soil of the same area. For isolation, samples were spread on Nu-trient Broth agar plates (NB, 0.5% peptone, 0.3% yeast extract, 0.5% NaCl, 1.5% agar) supplemented with 0.1 mg/ml cycloheximide to inhibit fungal growth and incubated at 28 °C for 24 h. The strains were identified by par-ticular rhizoid colony morphology and 16S rRNA gene sequence compar-ison. After isolation, B. mycoides strains were grown in Luria-Bertani (LB, 1% tryptone, 0.5% yeast extract, 0.5%NaCl) or Brain Heart Infusion (BHI, BactoTM, BD Biosciences, France) medium at 30 °C with 200 rpm agita-tion. E. coli strain MC1061 was used for cloning and was grown in LB at 37 °C with 220 rpm agitation. When needed, 50 mg/ml of kanamycin or 100 mg/ml of ampicillin (E. coli) or 4 mg/ml of chloramphenicol (B. my-coides) were added to the growth media.
Construction of GFP- and RFP-tagged B. mycoides The vector PYB was constructed by replacing the pAMβ1 origin of repli-cation (ori) of plasmid PATS28 (Namy et al., 1999) with the gram- positive temperature sensitive ori PWV01 from PAW068 plasmid (Wilson et al., 2007). Thus, the new vector PYB contains a PUC ori, a spectinomycin re-sistance gene, and a gram-positive temperature sensitive ori. A 1 kb frag-ment of the α-amylase gene was amplified from the genome of B. mycoides and digested with the restriction enzymes SacI and KpnI and ligated into the corresponding site of PYB resulting in the plasmid PYB_amy. The green fluorescent protein (GFP) and red fluorescent protein (RFP) genes were cloned into PYB_amy to obtain the vectors PYB_amyGFP and PYB_amyRFP. The plasmid PYB_amyGFP was transformed into B. my-coides EC18 and PYB_amyRFP was transformed into strain SB8 by electro-poration as described previously (Yi and Kuipers, 2017) and colonies were selected on 100 μg/ml spectinomycin LB plates at 30 °C. Transformants expressing GFP or RFP were selected by an Olympus MVX10 MacroView fluorescence microscope. Several bright colonies were inoculated in BHI liquid medium and grown at 37 °C to achieve a single crossover recom-bination. The cultures were diluted and plated on a BHI agar plate with 100 μg/ml spectinomycin and incubated at 37 °C. Successful GFP and RFP integration was verified by colony PCR. The expression of the fluores-cent protein was analyzed by fluorescence microscopy (Nikon Eclipse Ti, Japan) equipped with a CoolsnapHQ2 CCD camera. Images with GFP- or RFP-fluorescent cells were taken using 450–490 or 560–600 nm excita-tion for fluorescence channels and an Intensilight light as phase contrast channel. Final pictures were generated by ImageJ software by merging the two channels.
Root Colonization of the Endophyte and Soil Strains Observed by Confocal Laser Scanning Microscopy (CLSM) The overnight culture of GFP-tagged EC18 and RFP-tagged SB8 were di-luted 50 times and the strains were grown in BHI liquid medium at 37 °C to the exponential growth phase. Cells were collected by centrifuging at 3,000 × g and cell pellets were washed by 10 mM MgSO4 twice and re-suspended in 10 mM MgSO4 to an OD600 of 0.65. The viable cells were counted by counting colonies on LB agar plate containing 100 μg/ml spectinomycin. The densities of EC18 and SB8 suspension were 1.2 × 108 to 1.4 × 108 cfu ml−1. The plant root inoculation protocol was modified from Prieto et al. (2011). Briefly, Chinese cabbage seeds were surface sterilized by soaking in 2–3% sodium hypochlorite and 70% ethanol for 2 min, re-spectively. Seeds were washed with sterilized water three times to remove the remaining disinfectant and germinated on 25% Hoagland medium solidified with 0.6% agar. After 7 days growing in a climate chamber at 25 °C and 16/8 h light/dark photo-period, plant root was dipped for 45 min in bacterial suspensions of GFP-tagged B. mycoides EC18, RFP-tagged B. mycoides SB8, and a 1:1 mixture of the two strains. Plants were then trans-ferred to solid Hoagland medium plates and incubated in a climate cham-ber at 25 °C and 16/8 h light/dark.
For the visualization of the plant colonization by B. mycoides, fresh roots of cabbage plants were collected at 2 and 3 days after bacterial inoculation. The whole root was mounted on a glass slide and imaged using a confo-cal laser scanning microscope (Zeiss LSM 800, Carl Zeiss, Germany). La-sers at the wavelength of 488 and 561 nm were used to excite GFP and RFP, respectively. In order to visualize the endophytic colonization, a three- dimensional model that was rendered from about 60 optical sections at 1 μm intervals, was obtained with the Zen lite software (Carl Zeiss, Germany).
Genomic DNA Extraction, Sequencing, and Annotation Bacillus mycoides strains were grown overnight in BHI broth at 30 °C, 200 rpm. One ml of overnight culture was centrifuged and the pellet was resuspended in 250 μl SET buffer (75 mM NaCl, 25 mM EDTA, and 20 mM Tris-HCl at pH 7.5). The cells were lysed with lysozyme at 37 °C for 30 min. Then 10 μl of 10 mg/ml RNase was added and incubated 10 min at room temperature to remove RNA. After that, 25 μl SDS (10%) and 10 μl pro-teinase K (20 mg/ml) were added and the cells were incubated at 55 °C for 30 min. The lysate was treated with phenol–chloroform and the water phase was used to recover DNA by isopropanol precipitation. Precipitated DNA was washed two times with cold 70% ethanol and finally dissolved in TE buffer (10 mM Tris-HCl, 1 mM EDTA, pH 7.5). The quality and quan-tity of the genomic DNA was checked with a Nanodrop (ND-1000, Thermo

33
44 45
Com
para
tive
tran
scri
ptom
ics o
f Bac
illus
myc
oide
s str
ains
in re
spon
se to
pot
ato-
root
ex
udat
es re
veal
s diff
eren
t gen
etic
ada
ptat
ion
of e
ndop
hytic
and
soil
isol
ates
MAT
ERIA
LS A
ND
MET
HO
DS
Scientific, United States) and a Promega fluorometer (Quantus, Promega, United States).
Purified genomic DNA was sequenced using the MiSeq sequencing sys-tem of Illumina (Zerbino and Birney, 2008), yielding 250 bp paired-end reads with a mean library size of 400 bp. De novo assembly was performed using Velvet (Zerbino and Birney, 2008). Prediction of protein-encoding re-gions and automatic functional annotation was performed using the Rapid Annotations using Subsystem Technology (RAST) server (Aziz et al., 2008). Genes for tRNAs and rRNAs were predicted with tRNAscan-SE (Lowe and Eddy, 1997) and RNAmmer server (Lagesen et al., 2007) respectively, with the prokaryotic default setting. Insertion sequences (ISs) were identified by BLASTN searches against the ISfinder database (Siguier et al., 2006). Prophage regions were identified using the PHAST web server (Zhou et al., 2011). Orthologous protein coding sequences in B. mycoides ATCC 6462, B. mycoides EC18, and B. mycoides SB8 were analyzed by the OMA (Ortholo-gous Matrix) orthology database (Altenhoff et al., 2014). In addition, the functionality analysis of all the genes was performed by searching against protein domain databases, including GO (Harris et al., 2004) and COG (clusters of orthologous groups) (Tatusov et al., 2003).
Root Exudate Preparation and Bacterial Treatment For the collection of potato root exudates, tubers were rinsed with water to remove surface particles. Clean tubers were bathed in 2–3% sodium hy-pochlorite and 70% ethanol for 2–3 min, followed by five washing steps with sterile distilled water. The tubers were then sowed in autoclaved pots containing wet vermiculite and kept in a climate chamber at 26 °C, with a photoperiod of 16 h of light and 8 h of dark for 2 weeks. When the shoots sprouted, the seedlings were transferred into beakers filled with 20 ml of autoclaved water. The potato tubers were fitted in small plastic baskets, which support the tubers being just above the water surface and the roots hanging in the water. The water containing the exudates was collected and the beakers were refilled with sterile water. Sample collection was performed for 7 days after transferring the seedlings. A 100 μl aliquot of each collection was taken and spread on an LB plate to check for contami-nation and the contaminated samples were discarded. The other samples were stored at −20 °C until use. An overnight culture of B. mycoides was diluted to an initial OD600 of 0.05 in 90 ml LB medium supplemented with 10 ml of root exudates collected from potato plants. The same amount of Mili-Q water served as control. Ten milliliter of root exudates were se-lected, since no discernible effect on bacterial growth was observed at this level. The cultures were grown at 30 °C, 200 rpm for 1 h and cells were harvested for RNA isolation.
RNA Extraction The total RNA was isolated with the Roche RNA isolation kit with some modifications. The cell pellets were flash-frozen in liquid nitrogen and resuspend in 400 μl TE buffer in 2 ml screw-cap tube. Then 50 μl of 10% SDS, 250 μl chloroform, 250 μl phenol and 0.5 g glass beads (0.5 μm) were added. The samples were homogenized using a Mini-BeadBeater (607, BioSpec, United States) for three 45 s cycles and were chilled on ice be-tween cycles. Following the disruption, samples were centrifuged at 4 °C and the supernatant was mixed with chloroform. After centrifugation at 4 °C for 5 min, 500 μl of the upper phase were transferred to a fresh tube and 1 ml of lysis/binding buffer from the Roche RNA isolation kit were added. The mixture was transferred to filter and collection tubes and cen-trifuged 15 s at 8,600 rpm. The filter tube was dried by an additional cen-trifugation step for 15 s at 8,600 rpm. DNaseI solution was added and DNA was removed by incubation at 15–25 °C for 20–30 min. After two washing steps with the wash buffer (Roche RNA isolation kit), the RNA was eluted with 50 μl elution buffer (Roche RNA isolation kit) and stored at −80 °C before sequencing. The RNA quality was analyzed with the Bioanalyzer (Agilent Technologies, Wilmington, DE, United States).
RNA Sequencing and Data Analyses RNA sequencing was conducted at PrimBio Research Institute (Exton, PA, United States). The raw RNA-Seq reads were trimmed from the adapter sequences and mapped against the reference genome sequence of each strain. The RPKM (Reads Per Kilobase Million) table of the core genome of the three strains (4,743 genes) was generated. Differential gene expression analyses were carried out with the web server pipeline T-REx (de Jong et al., 2015), in which the RPKM values of the B. mycoides strain grown in the pres-ence of root exudates was compare with that grown in the absence of root exudates. The significance threshold was p-value < 0.05 and fold-change > 2 (“TopHits” in T-Rex). Functional categorization including GO, InterPro, KEGG and PFAM domains, was performed on the Genome2D web server2. The RNA-seq data from this study have been submitted to the NCBI Gene Expression Omnibus (GEO3) under the accession number GSE98148.
RESULTS AND DISCUSSION
Root Colonization of Endophytic and Soil-Associated B. mycoides Strains In order to generate traceable B. mycoides strains for plant-interaction im-aging experiments, the plasmid PYB_amyGFP was transformed into the

33
46 47
Com
para
tive
tran
scri
ptom
ics o
f Bac
illus
myc
oide
s str
ains
in re
spon
se to
pot
ato-
root
ex
udat
es re
veal
s diff
eren
t gen
etic
ada
ptat
ion
of e
ndop
hytic
and
soil
isol
ates
RESU
LTS
AN
D D
ISCU
SSIO
N
endophytic B. mycoides strain EC18 by electroporation. A GFP gene con-trolled by a constitutive promoter was integrated in the α-amylase locus by curing the thermo-sensitive plasmid backbone at 37 °C. A RFP gene was introduced into the strain SB8 with the plasmid PYB_amyRFP by ap-pplying the same procedure. In order to compare the rhizosphere compe-tence of the B. mycoides EC18 cells expressing GFP (Figure 1A) and of the SB8 cells expressing RFP (Figure 1B), a 1:1 mixture of the strains was inocu-lated into the hydroponic plant interaction model, and strain competition experiments were performed (Figure 1C). Inoculated cabbage plants were grown in a climate chamber and root samples were taken 1 and 3 days af-ter inoculation (DAI). Subsequently, samples were examined for the pres-ence of bacterial cells using confocal laser scanning microscopy (CLSM). When the strains were inoculated separately, both the endophytic strain B. mycoides EC18 (Figure 1D) and the soil strain B. mycoides SB8 (Figure 1E) were detected in the rhizosphere 1 DAI. Remarkably, when a 1:1 mixture of the EC18 and SB8 cells was inoculated, the roots were predominantly colonized by EC18 (Figure 1F). After 3 days of inoculation, colonization of the roots was progressing and the attachment of cells was significantly in-creased in the EC18 single inoculation group (Figure 1G). A similar trend was also observed for the SB8 population, albeit less significantly (Fig-ure 1H). In the mixed inoculation group, the two strains shared a few col-onization sites, however, the EC18 micro-populations predominated over the SB8 micro-populations (Figure 1I). A three-dimensional model showed that B. mycoides EC18 was able to colonize cortical tissues and xylem ves-sels 3 DAI (Figure 1J). In the mixture inoculation group, both strains were detected on the root surface, but only EC18 was detected inside the root tissues (Figure 1L). In the B. mycoides SB8 single inoculation experiment, endophytic colonization was not detected (Figure 1K).
As soil-borne microorganisms, several rhizosphere Bacillus species can enter plants and colonize internal tissues and many of them have shown PGP effects (Ji et al., 2008; Yi et al., 2013; Harun-Or-Rashid et al., 2017). The ability to colonize the rhizoplane is required for a successful endophytic colonization. As shown in Figures 1D, E, G, H, although both the EC18 and the SB8 isolates can establish a rhizoplane colonization, EC18 showed a more successful, faster and aggressive attachment. Following root attach-ment, the endophytic B. mycoides strain used a penetration process to enter the root interior and initiate the endophytic cell development. It has been discussed that some rhizosphere bacteria can occasionally and temporar-ily become endophytic, since passive penetration can take place at areas with damaged tissue, such as those occurring at root emergence sites or root tips (Compant et al., 2010). However, our data indicate that specific cell adaptations might be involved in active penetration of the root system
by B. mycoides, because EC18 and SB8 colonized the same sites of the root, but only EC18 was able to enter the root tissue (Figure 1L).
Figure 1. Root colonized by Bacillus mycoides observed by CLSM. GFP-tagged B. my-coides EC18 (A), RFP-tagged B. mycoides SB8 (B), and a 1:1 mixture of the two strains (C) were inoculated to the seedlings. Root samples were inspected 1 and 3 days after inoc-ulation (DAI). (D, G) Root inoculated with B. mycoides EC18 at 1 and 3 DAI. (E, H) Root inoculated with B. mycoides SB8 at 1 and 3 DAI. (F, I) Root inoculated with a 1:1 mixture of B. mycoides EC18 and B. mycoides SB8 at 1 and 3 DAI. (J–L) Three-dimensional model of roots inoculated with EC18, SB8, and a 1:1 mixture of the two strains 3 DAI.

33
48 49
Com
para
tive
tran
scri
ptom
ics o
f Bac
illus
myc
oide
s str
ains
in re
spon
se to
pot
ato-
root
ex
udat
es re
veal
s diff
eren
t gen
etic
ada
ptat
ion
of e
ndop
hytic
and
soil
isol
ates
RESU
LTS
AN
D D
ISCU
SSIO
N
Genomic Analyses of B. mycoides EC18 and SB8The Whole Genome Shotgun sequence of strains EC18 and SB8 has been deposited at GenBank under the accession MRWW00000000 and MRWS00000000, respectively. The general features of the genomes of EC18, SB8 and the reference strain ATCC 6462 are listed in Table 1. EC18 has a genome size of 5.75 Mb (Mega Base pairs) and SB8 has a genome size of 5.98 Mb, both of which are bigger than the type strain ATCC 6462 (5.64 Mb). The GC content of all three strains is around 35%. EC18 encodes 6,014 pro-teins, 77 RNA genes (7 rRNA and 70 tRNA genes) and 71 prophage- associated genes, while the SB8 genome comprises 6,250 protein- coding genes, 47 RNA genes (3 rRNA and 44 tRNA genes), and 177 prophage- associated genes. As shown in Figure 2, the core genome of the three strains consists of 4,743 or-thologous groups and a pan-genome size of 7,234 genes, among which 357 genes are unique to EC18 and 646 genes are unique to SB8. 810 ortholo-gous genes are shared by EC18 and SB8, 104 genes are shared between EC18 and ATCC 6462, while only 51 genes are shared between SB8 and ATCC 6462. The fact that SB8 and EC18 share 810 orthologous genes distinct from ATCC 6462, suggests that the latter strains are more closely related to each other than to the type strain isolate.
Global Transcriptional Profiles in Response to Root Exudates About 10 million reads were generated by RNA-Seq for each sample, of which 60–70% passed the quality filtering (Phred quality scores of > 20) using the FASTX-Toolkit version 0.0.13.2. Subsequently, the filtered reads were mapped on the genome of type strain B. mycoides ATCC 6462 to de-duce the RPKM value of each gene or RNA. The RPKM values of the core genes (4,743 genes) were used to compare the transcriptomes of the strains under study using the T-REx pipeline (de Jong et al., 2015). Significant dif-ferentially expressed genes (DEGs) were shown in the TopHits genes close to cutoff values (p-value 0.05 and fold-change of 2) (Figures 3A&B).
In the endophytic strain EC18, 715 genes were significantly changed by the exposure to root exudates. Of those, 408 genes were upregulated and
Table 1. A comparison of the genomic features of Bacillus mycoides strain ATCC6462, EC18, and SB8.
ATCC6462 EC18 SB8Genome size 5.64 Mb 5.75 Mb 5.98 MbGC content 35.4 % 35.1% 35.1%Protein 5421 6014 6250RNA genes 179 77 50IS elements 22 42 47Phage-associated genes 26 71 177
307 genes were downregulated (Figure 3C). 193 out of the 715 altered genes were represented in the COG database and hence used to perform ortolo-ganalyses. As shown in Figure 4, a large part of the genes is poorly charac-terized and belong to the group [R] (General function prediction only) or [S] (Function unknown). Apart from this, the upregulated genes were mainly implicated in amino acid metabolism and signal transduction, while many of the significantly downregulated genes were involved in cell cycle control, amino acid transport and amino acid metabolism, cell wall and membrane biogenesis, transcription, and carbohydrate transport and metabolism. The soil strain SB8 responded to the root exudates with an upregulated of 261 genes, whereas 219 genes were significantly downregulated (Figure 3D). 109 of the in total 480 altered genes were found in the COG database. The largest group of altered genes were again clustered in [R] and [S], thereby leaving their function unknown or only providing an insight into a gener-ally assumed function. The genes represented by COG classes with known function were mostly categorized in amino acid transport and metabolism, cell cycle control, and energy production and conversion. Taken together, the results showed clear differences in gene expression changes between EC18 and SB8 in response to potato root exudates.
Figure 2. Venn diagram showing the core genome and the genes specific of the B. my-coides strain ATCC 6462, EC18, and SB8.
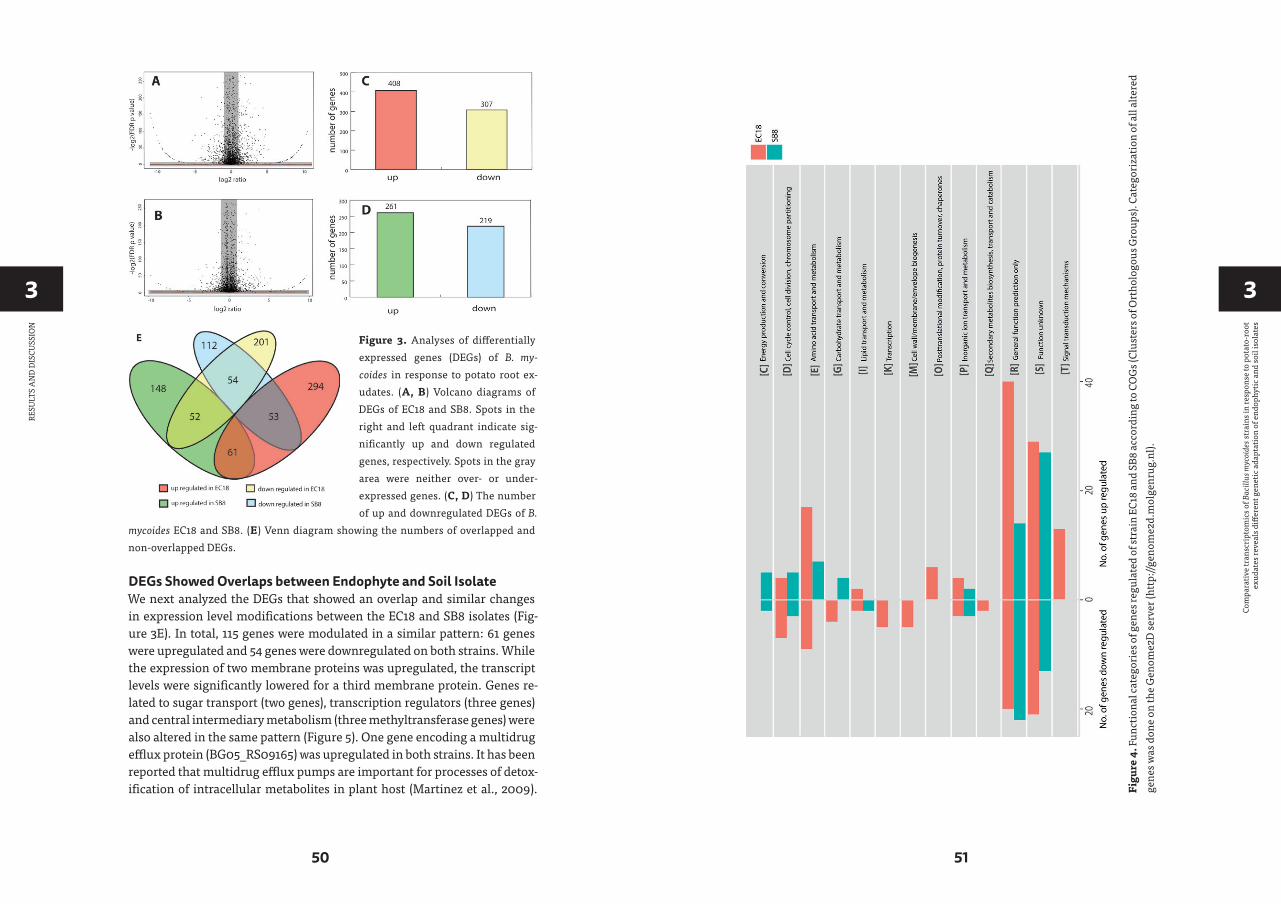
33
50 51
Com
para
tive
tran
scri
ptom
ics o
f Bac
illus
myc
oide
s str
ains
in re
spon
se to
pot
ato-
root
ex
udat
es re
veal
s diff
eren
t gen
etic
ada
ptat
ion
of e
ndop
hytic
and
soil
isol
ates
RESU
LTS
AN
D D
ISCU
SSIO
N
DEGs Showed Overlaps between Endophyte and Soil Isolate We next analyzed the DEGs that showed an overlap and similar changes in expression level modifications between the EC18 and SB8 isolates (Fig-ure 3E). In total, 115 genes were modulated in a similar pattern: 61 genes were upregulated and 54 genes were downregulated on both strains. While the expression of two membrane proteins was upregulated, the transcript levels were significantly lowered for a third membrane protein. Genes re-lated to sugar transport (two genes), transcription regulators (three genes) and central intermediary metabolism (three methyltransferase genes) were also altered in the same pattern (Figure 5). One gene encoding a multidrug efflux protein (BG05_RS09165) was upregulated in both strains. It has been reported that multidrug efflux pumps are important for processes of detox-ification of intracellular metabolites in plant host (Martinez et al., 2009).
Figure 3. Analyses of differentially expressed genes (DEGs) of B. my-coides in response to potato root ex-udates. (A, B) Volcano diagrams of DEGs of EC18 and SB8. Spots in the right and left quadrant indicate sig-nificantly up and down regulated genes, respectively. Spots in the gray area were neither over- or under- expressed genes. (C, D) The number of up and downregulated DEGs of B.
mycoides EC18 and SB8. (E) Venn diagram showing the numbers of overlapped and non-overlapped DEGs.
Figu
re 4
. Fun
ctio
nal c
ateg
orie
s of g
enes
regu
late
d of
stra
in E
C18
and
SB8
acco
rdin
g to
CO
Gs (
Clus
ters
of O
rtho
logo
us G
roup
s). C
ateg
oriz
atio
n of
all
alte
red
gene
s was
don
e on
the
Gen
ome2
D se
rver
(htt
p://g
enom
e2d.
mol
genr
ug.n
l).

33
52 53
Com
para
tive
tran
scri
ptom
ics o
f Bac
illus
myc
oide
s str
ains
in re
spon
se to
pot
ato-
root
ex
udat
es re
veal
s diff
eren
t gen
etic
ada
ptat
ion
of e
ndop
hytic
and
soil
isol
ates
RESU
LTS
AN
D D
ISCU
SSIO
N
Figu
re 5
. Ove
rlap
ping
DEG
s of B
. myc
oide
s str
ain
EC18
and
SB8
in re
spon
se to
root
exud
ates
. Red
repr
esen
ts si
gnifi
cant
upr
egul
atio
n an
d bl
ue in
dica
tes
sign
ifica
nt d
ownr
egul
atio
n of
gen
e ex
pres
sion
.
Similar studies with other rhizosphere-associated Bacillus species have shown that the sporulation and germination process can be affected by root exudates (Fan et al., 2012; Zhang et al., 2015). In line with these findings, nine sporulation and two germination genes were altered in the two strains in this study. This indicates that modulation of sporulation and germina-tion pathways in response to root exudates is a response of Bacillus species independent from their ecological niche as a rhizosphere, endosphere, or soil strain. Furthermore, five genes with oxidoreductase activity were both downregulated by root exudates in the two strains.
DEGs Showed Opposite Trends between Endophyte and Soil Isolate A total of 105 DEGs showed a different pattern between EC18 and SB8. 53 genes were upregulated in EC18 but downregulated in SB8. On the contrary, 52 genes were downregulated in EC18 but upregulated in SB8 (Figure 3E). The different expression profiles suggest that endophytic and soil- associated B. mycoides apply different genetic adaptation strategies to recognize or re-spond to plant-released signals. Presumably, the endophytic strain is able to change its gene expression pattern and thus adapt the metabolism toward a physiological state that enables an optimal nutrient acquisition, com-petition with species from the same niche and colonization of the plant.
As shown in Figure 6, the genes being upregulated in EC18 and down-regulated in SB8 included transcriptional regulators (five genes) among which IclR is related to multidrug resistance and degradation of aromatic compounds in soil bacteria (Song et al., 2004; Molina-Henares et al., 2006) and the sigma-28 factor is reported to transcribe the flagellin gene and con-trol the transcription of a regulon specifying flagellar, chemotaxis, and motility functions in B. subtilis (Helmann and Chamberlin, 1987; Mirel and Chamberlin, 1989). Additionally, genes involved in the metabolism of amino acids showed an oppositely regulated expression pattern between the two strains. These included two genes involved in tryptophan metab-olism (BG05_24905 and BG05_24920), three genes involved in branched-chain amino acid metabolism, e.g., BG05_22255 (ilvB), BG05_22250 (ilvC), and BG05_22265 (ilvE), and two genes involved in cysteine/methionine me-tabolism (BG05_11740 and BG05_23955). Xie et al. (2015) reported that sev-eral genes related to amino acids metabolism were stimulated when B. subti-lis OKB105 was incubated with rice seedlings for 2 h. This finding is perhaps not surprising for rhizobacteria and endophytic bacteria, since amino acids are one of the major constituents of plant root exudates (Lugtenberg et al., 2001; Mark et al., 2005). However, it is interesting to see that those genes are downregulated in the soil strain when it encounters the root exudates.
Another group of genes was upregulated in the soil-derived strain SB8 and downregulated in endophytic strain EC18. These included two

33
54 55
Com
para
tive
tran
scri
ptom
ics o
f Bac
illus
myc
oide
s str
ains
in re
spon
se to
pot
ato-
root
ex
udat
es re
veal
s diff
eren
t gen
etic
ada
ptat
ion
of e
ndop
hytic
and
soil
isol
ates
RESU
LTS
AN
D D
ISCU
SSIO
N
Figu
re 6
. Diff
eren
tially
exp
ress
ed g
enes
show
opp
osite
tren
ds in
B. m
ycoi
des s
trai
n EC
18 a
nd S
B8 in
resp
onse
to ro
ot e
xuda
tes.
Red
repr
esen
ts si
gnifi
cant
up
regu
latio
n an
d bl
ue in
dica
tes s
igni
fican
t dow
nreg
ulat
ion
of g
ene
expr
essi
on.
transporters involved in multidrug resistance proteins (BG05_26905 and BG05_26390) and several transporters including a major facilita-tor superfamily (MFS) transporter (BG05_17370), a citrate transporter (BG05_28360), a cellobiose PTS transporter (BG05_24345), and a putres-cine importer (BG05_24345). Notably, a previous study has shown that the increased rate of putrescine uptake can decrease the competitive root col-onization ability of Pseudomonas fluorescens WCS365 (Kuiper et al., 2001). The upregulation of this gene might thus decrease the rhizosphere colo-nization competence of SB8.
DEGs Exclusively Changed in the Endophyte or the Soil Isolate In addition to the overlapping transcriptional responses of the two strains, there is a large number of genes that were specifically modulated in one of the two strains studied. As shown in Figure 4, 495 DEGs were identified as endophytic strain-specific responses (294 up- and 201 downregulated) and 260 DEGs were identified as soil strain-specific responses (148 up- and 112 downregulated). Some of these genes have unknown functions and are currently annotated as hypothetical proteins (141 for EC18 and 91 for SB8). Altogether, this large number of specialized transcriptional changes underpins the different and specialized responses of endophytic and soil B. mycoides strain to the plant signals. We selectively discuss some genes that may participate in B. mycoides–plant interactions. In gen-eral, the endophytic strain showed a more complex response than the soil isolate. The DEGs are categorized based on GO, COG, and Pfam databases.
The endophytic strain responded to the root exudate with a significant upregulation of genes related to amino acid transmembrane transport, while genes involved in sugar transport including BG05_RS27120 (cel-lobiose PTS transporter IIA) and BG05_RS26300 (arabinose ABC trans-porter permease) were downregulated (Figure 7). However, as shown in Figure 8, several genes related to the PTS (phosphohistone-sugar phos-photransferase system) were significantly upregulated in the soil isolate SB8, i.e., those genes including sugar ABC transporter (BG05_RS11820), PTS lactose transporter subunit (BG05_RS06025), PTS cellobiose trans-porter subunit (BG05_RS27110 and BG05_RS27115), and PTS sugar trans-porter (BG05_RS27275). It was reported that some rhizosphere bacteria se-lectively and preferentially consume amino acids instead of sugars. The catabolite repression of glucose assimilation in the presence of amino ac-ids was observed among the rhizosphere genera Pseudomonas and Micro-monospora (Hoskisson et al., 2003; Moreno et al., 2009). Moreover, it has been shown that the enhanced ability to catabolize amino acids is respon-sible for the fitness gain during nutrient-limiting conditions (Zinser and Kolter, 1999). This suggests that bacteria, which are capable of modulating

33
56 57
Com
para
tive
tran
scri
ptom
ics o
f Bac
illus
myc
oide
s str
ains
in re
spon
se to
pot
ato-
root
ex
udat
es re
veal
s diff
eren
t gen
etic
ada
ptat
ion
of e
ndop
hytic
and
soil
isol
ates
RESU
LTS
AN
D D
ISCU
SSIO
N
the amino acid uptake and preferring their utilization may have a selec-tive advantage under such conditions. In this study, we observed that even within the same species, an endophytic strain and a soil strain show dif-ferent metabolic preferences for amino acids or sugars due to the adapta-tion of the endosphere or soil niche.
A group of genes related to metabolic processes were specifically altered in the endophytic strain EC18 (Figure 7). These included four upregulated hydrolase genes (BG05_RS07285, BG05_RS13815, BG05_RS18105, and BG05_RS20350) and one downregulated hydrolase gene (BG05_RS16815). Additionally, four genes of the alpha/beta hydrolase fold protein family were upregulated and three downregulated in EC18. In comparison, only three alpha/beta hydrolase genes are altered in SB8 (two upregulated and one downregulated) (Figure 8). Proteins of the alpha/beta hydrolase fold family are involved in the degradation of plant cell wall polymers in Bur-kholderia spp. (Ali et al., 2014). Similarly, the expression of pullulanase
Figure 7. Differentially expressed genes specifically present in endophytic strain B. my-coides EC18 in response to root exudates. Red represents significant upregulation and blue indicates significant downregulation of gene expression.
Figu
re 8
. Diff
eren
tially
ex-
pres
sed
gene
s spe
cific
ally
pr
esen
t in
the
soil
stra
in B
. m
ycoi
des S
B8 in
resp
onse
to
root
exu
date
s. R
ed re
p-re
sent
s sig
nific
ant u
preg
-ul
atio
n an
d bl
ue in
dica
tes
sign
ifica
nt d
ownr
egul
atio
n of
gen
e ex
pres
sion
.

33
58 59
Com
para
tive
tran
scri
ptom
ics o
f Bac
illus
myc
oide
s str
ains
in re
spon
se to
pot
ato-
root
ex
udat
es re
veal
s diff
eren
t gen
etic
ada
ptat
ion
of e
ndop
hytic
and
soil
isol
ates
CON
CLU
SIO
N
(BG05_RS18715) and of the chitin-binding protein (BG05_RS19070) was upregulated in EC18. These two genes belong to the O-glycosyl hydrolase family and it might be possible that they are involved in the metabolism of plant-derived compounds. Three proteolysis genes including pyrro-lidone-carboxylate peptidase (BG05_RS16435), D-alanyl-D-alanine car-boxypeptidase (BG05_RS18945), and a serine protease (BG05_RS21540) were upregulated in EC18 (Figure 7), while all three proteolysis genes were repressed in SB8. Moreover, five glyoxalase genes were specifically upreg-ulated in EC18 (Figure 7). Five genes encoding glucose/ribitol dehydroge-nase (BG05_RS12980, BG05_RS14070, BG05_RS17890, BG05_RS24540, and BG05_RS07590) were upregulated in EC18. The overall metabolic gene expression profile suggests that the endophytic strain consumes limited numbers of sugars (glucose/ribitol), while also using more types of other carbon substrates.
Among the genes related to stress response, three genes involved in multidrug resistance are downregulated in two strains, with two genes (BG05_RS22665 and BG05_RS26910) being more significantly al-tered in EC18 (Figures 7, 8). A stress-induced protein (BG05_RS01935), a stress response protein (BG05_RS28400), and a general stress protein (BG05_RS26065) were specifically upregulated in endophytic strain. Genes encoding a methanol dehydrogenase (BG05_RS02305) and a strep-tomycin biosynthesis protein (BG05_RS25055) were also upregulated.
CONCLUSION
Endophytic bacteria have emerged as a valuable source of biocontrol agents and metabolites, which has great potential in sustainable agricul-tural application. However, many aspects of the interaction between en-dophytic bacteria and plants are unclear, e.g., the endophytic colonization process and its differences compared to non-endophytic microorganisms. This study was designed to attribute the properties specific for an endo-phytic B. mycoides strain or its soil counterpart. Indeed, the CLSM obser-vation showed that the endophytic strain is a more aggressive colonizer than the soil strain at an early stage of interaction with the plant root. The endophyte competes for the colonization site with the soil strain on the root surface and eventually penetrates the internal tissue of the root. It is presumed that the response of bacteria to environmental cues enables the gene expression pattern necessary for the endophytic life cycle. Our results suggest that root exudates of potato plants have a profound impact on gene expression levels in B. mycoides and that the endophytic strain showed more active or directed responses than the soil strain. The two
strains showed a different metabolic preference when they encountered root exudates. The upregulation of genes related to amino acids metab-olism, several proteolytic enzymes and O-glycosyl hydrolases points to-ward a specific adaptation to the ecological niche and a good rhizosphere fitness of the endophytic strain. In comparison, the nutrition source for the soil isolate seems to be constrained to sugars, which might hamper its colonization ability and proliferation rate in the rhizosphere. In con-clusion, this study provides new insights into the different transcriptomic adaptations between an endophytic bacterium and its soil counterpart in response to potato root exudates and provides new knowledge on endo-phytic bacteria-plant interactions.
AUTHOR CONTRIBUTIONS
Conceived and designed experiments: YY, OK. Performed the experiments: YY. Performed GFP and RFP selection experiments: EF and YY. Performed bioinformatic analysis: YY and AJ. Wrote the paper: YY, OK, and EF.
ACKNOWLEDGMENTS
We thank Prof. Jan Dirk van Elsas, Prof. Theo Elzinga, and Jan Spoelder at the University of Groningen for providing B. mycoides strains. YY was sup-ported by a scholarship from China Scholarship Council.
REFERENCES
Ali, S., Duan, J., Charles, T. C., and Glick, B. R.
(2014). A bioinformatics approach to the
determination of genes involved in en-
dophytic behavior in Burkholderia spp. J.
Theor. Biol. 343: 193–198.
Altenhoff, A. M., Škunca, N., Glover, N., Train,
C.-M., Sueki, A., Piližota, I., et al. (2014).
The OMA orthology database in 2015: func-
tion predictions, better plant support, syn-
teny view and other improvements. Nucleic
Acids Res. 43: D240–D249.
Ambrosini, A., Stefanski, T., Lisboa, B. B., Ben-
eduzi, A., Vargas, L. K., and Passaglia, L.
M. P. (2016). Diazotrophic bacilli isolated
from the sunflower rhizosphere and the
potential of Bacillus mycoides B38V as bio-
fertiliser. Ann. Appl. Biol. 168: 93–110.
Aziz, R. K., Bartels, D., Best, A. A., DeJongh,
M., Disz, T., Edwards, R. A., et al. (2008).
The RAST Server: rapid annotations us-
ing subsystems technology. BMC Genom-
ics 9: 75.
Bargabus, R., Zidack, N., Sherwood, J., and Ja-
cobsen, B. (2002). Characterisation of sys-
temic resistance in sugar beet elicited by a
non-pathogenic, phyllosphere-colonizing

33
60 61
Com
para
tive
tran
scri
ptom
ics o
f Bac
illus
myc
oide
s str
ains
in re
spon
se to
pot
ato-
root
ex
udat
es re
veal
s diff
eren
t gen
etic
ada
ptat
ion
of e
ndop
hytic
and
soil
isol
ates
REFE
REN
CES
Bacillus mycoides, biological control agent.
Physiol. Mol. Plant Pathol. 61: 289–298.
Brencic, A., and Winans, S. C. (2005). Detec-
tion of and response to signals involved
in host-microbe interactions by plant-
associated bacteria. Microbiol. Mol. Biol.
Rev. 69: 155–194.
Compant, S., Clément, C., and Sessitsch, A.
(2010). Plant growth-promoting bacteria in
the rhizo- and endosphere of plants: their
role, colonization, mechanisms involved
and prospects for utilization. Soil Biol. Bio-
chem. 42: 669–678.
de Jong, A., van der Meulen, S., Kuipers, O. P.,
and Kok, J. (2015). T-REx: transcriptome
analysis webserver for RNA-seq expres-
sion data. BMC Genomics 16: 663.
Di Franco, C., Beccari, E., Santini, T., Pisaneschi,
G., and Tecce, G. (2002). Colony shape as a
genetic trait in the pattern-forming Bacil-
lus mycoides. BMC Microbiol. 2: 33.
Fan, B., Carvalhais, L. C., Becker, A., Fedosey-
enko, D., von Wirén, N., and Borriss, R.
(2012). Transcriptomic profiling of Bacil-
lus amyloliquefaciens FZB42 in response to
maize root exudates. BMC Microbiol. 12: 116.
Glick, B. R. (2012). Plant growth-promoting
bacteria: mechanisms and applications.
Scientifica 2012: 963401.
Gómez-Lama Cabanás, C., Schilirò, E., Valverde-
Corredor, A., and Mercado-Blanco, J. (2014).
The biocontrol endophytic bacterium Pseu-
domonas fluorescens PICF7 induces systemic
defense responses in aerial tissues upon col-
onization of olive roots. Front. Microbiol. 5: 427.
Hardoim, P. R., van Overbeek, L. S., and Elsas,
J. D. (2008). Properties of bacterial endo-
phytes and their proposed role in plant
growth. Trends. Microbiol. 16: 463–471.
Harris, M. A., Clark, J., Ireland, A., Lomax, J.,
Ashburner, M., Foulger, R., et al. (2004).
The gene ontology (GO) database and
informatics resource. Nucleic Acids Res.
32(Suppl. 1): D258–D261.
Harun-Or-Rashid, M., Khan, A., Hossain, M. T.,
and Chung, Y. R. (2017). Induction of sys-
temic resistance against aphids by endo-
phytic Bacillus velezensis YC7010 via ex-
pressing PHYTOALEXIN DEFICIENT4 in
Arabidopsis. Front. Plant Sci. 8: 211.
Helmann, J. D., and Chamberlin, M. J. (1987).
DNA sequence analysis suggests that
expression of flagellar and chemotaxis
genes in Escherichia coli and Salmonella ty-
phimurium is controlled by an alternative
sigma factor. Proc. Natl. Acad. Sci. U.S.A.
84: 6422–6424.
Hoskisson, P., Sharples, G., and Hobbs, G.
(2003). The importance of amino acids as
carbon sources for Micromonospora echi-
nospora (ATCC 15837). Lett. Appl. Microbiol.
36: 268–271.
Janczarek, M., Rachwał, K., Marzec, A.,
Grządziel, J., and Palusińska-Szysz, M.
(2015). Signal molecules and cell-surface
components involved in early stages of the
legume–rhizobium interactions. Appl. Soil
Ecol. 85: 94–113.
Jasim, B., Jimtha John, C., Shimil, V., Jyothis,
M., and Radhakrishnan, E. (2014). Studies
on the factors modulating indole-3-acetic
acid production in endophytic bacterial
isolates from Piper nigrum and molecu-
lar analysis of ipdc gene. J. Appl. Microbiol.
117: 786–799.
Ji, X., Lu, G., Gai, Y., Zheng, C., and Mu, Z.
(2008). Biological control against bac-
terial wilt and colonization of mulberry
by an endophytic Bacillus subtilis strain.
FEMS Microbiol. Ecol. 65: 565–573.
Jin, H., Yang, X., Yan, Z., Liu, Q., Li, X., Chen,
J., et al. (2014). Characterization of rhi-
zosphere and endophytic bacterial com-
munities from leaves, stems and roots of
medicinal Stellera chamaejasme L. Syst.
Appl. Microbiol. 37: 376–385.
Kandel, S. L., Firrincieli, A., Joubert, P. M.,
Okubara, P., Leston, N. D., McGeorge,
K. M., et al. (2017). An in vitro study of
bio-control and plant growth promotion
potential of Salicaceae endophytes. Front.
Microbiol. 8: 386.
Kuiper, I., Bloemberg, G. V., Noreen, S., Thom-
as-Oates, J. E., and Lugtenberg, B. J. (2001).
Increased uptake of putrescine in the rhi-
zosphere inhibits competitive root coloni-
zation by Pseudomonas fluorescens strain
WCS365. Mol. Plant Microbe Interact. 14:
1096–1104.
Kuiper, I., Lagendijk, E. L., Bloemberg, G. V.,
and Lugtenberg, B. J. (2004). Rhizoreme-
diation: a beneficial plant-microbe inter-
action. Mol. Plant Microbe Interact. 17: 6–15.
Lagesen, K., Hallin, P., Rødland, E. A., Stærfeldt,
H.-H., Rognes, T., and Ussery, D. W. (2007).
RNAmmer: consistent and rapid annota-
tion of ribosomal RNA genes. Nucleic Ac-
ids Res. 35: 3100–3108.
Lakshmanan, V., Castaneda, R., Rudrappa, T.,
and Bais, H. P. (2013). Root transcriptome
analysis of Arabidopsis thaliana exposed to
beneficial Bacillus subtilis FB17 rhizobac-
teria revealed genes for bacterial recruit-
ment and plant defense independent of
malate efflux. Planta 238: 657–668.
Lee, S., Flores-Encarnacion, M., Contreras-Ze-
ntella, M., Garcia-Flores, L., Escamilla, J.,
and Kennedy, C. (2004). Indole-3-acetic
acid biosynthesis is deficient in Gluconac-
etobacter diazotrophicus strains with muta-
tions in cytochrome c biogenesis genes. J.
Bacteriol. 186: 5384–5391.
Lowe, T. M., and Eddy, S. R. (1997). tRNA-
scan-SE: a program for improved detec-
tion of transfer RNA genes in genomic se-
quence. Nucleic Acids Res. 25: 955–964.
Lugtenberg, B. J., Dekkers, L., and Bloemberg,
G. V. (2001). Molecular determinants of
rhizosphere colonization by Pseudomonas.
Annu. Rev. Phytopathol. 39: 461–490.
Malfanova, N., Lugtenberg, B., and Berg, G.
(2013). Bacterial Endophytes: Who and
Where, and What are they Doing there?
Hoboken, NJ: Wiley-Blackwell.
Mark, G. L., Dow, J. M., Kiely, P. D., Higgins, H.,
Haynes, J., Baysse, C., et al. (2005). Tran-
scriptome profiling of bacterial responses
to root exudates identifies genes involved
in microbe-plant interactions. Proc. Natl.
Acad. Sci. U.S.A. 102: 17454–17459.
Martinez, J. L., Sanchez, M. B., Martinez-Solano,
L., Hernandez, A., Garmendia, L., Fajardo, A.,
et al. (2009). Functional role of bacterial mul-
tidrug efflux pumps in microbial natural eco-
systems. FEMS Microbiol. Rev. 33: 430–449.
Mirel, D. B., and Chamberlin, M. J. (1989). The
Bacillus subtilis flagellin gene (hag) is tran-
scribed by the sigma 28 form of RNA poly-
merase. J. Bacteriol. 171: 3095–3101.
Molina-Henares, A. J., Krell, T., Eugenia Guaz-
zaroni, M., Segura, A., and Ramos, J. L.
(2006). Members of the IclR family of bac-
terial transcriptional regulators function
as activators and/or repressors. FEMS Mi-
crobiol. Rev. 30: 157–186.
Mónica, R., and Esperanza, M.-R. (2006). Bacte-
rial endophytes and their interactions with
Hosts. Mol. Plant Microbe Interact. 19: 827–837.
Moreno, R., Martínez-Gomariz, M., Yuste, L.,
Gil, C., and Rojo, F. (2009). The Pseudomo-
nas putida Crc global regulator controls
the hierarchical assimilation of amino ac-
ids in a complete medium: evidence from
proteomic and genomic analyses. Pro-
teomics 9: 2910–2928.
Namy, O., Mock, M., and Fouet, A. (1999).
Co-existence of clpB and clpC in the Bacil-
laceae. FEMS Microbiol. Lett. 173: 297–302.

33
62 63
Com
para
tive
tran
scri
ptom
ics o
f Bac
illus
myc
oide
s str
ains
in re
spon
se to
pot
ato-
root
ex
udat
es re
veal
s diff
eren
t gen
etic
ada
ptat
ion
of e
ndop
hytic
and
soil
isol
ates
REFE
REN
CES
Neher, O. T., Johnston, M. R., Zidack, N. K., and
Jacobsen, B. J. (2009). Evaluation of Bacillus
mycoides isolate BmJ and B. mojavensis iso-
late 203-7 for the control of anthracnose
of cucurbits caused by Glomerella cingulata
var. orbiculare. Biol. Control 48: 140–146.
Pangesti, N., Pineda, A., Dicke, M., and Loon, J.
(2015). Variation in plant-mediated inter-
actions between rhizobacteria and cater-
pillars: potential role of soil composition.
Plant Biol. 17: 474–483.
Phillips, D. A., Joseph, C. M., and Maxwell, C.
A. (1992). Trigonelline and stachydrine re-
leased from alfalfa seeds activate NodD2
protein in Rhizobium meliloti. Plant Physiol.
99: 1526–1531.
Prashar, P., Kapoor, N., and Sachdeva, S. (2013).
Rhizosphere: its structure, bacterial diver-
sity and significance. Rev. Environ. Sci. Bio/
Technol. 13: 63–77.
Prieto, P., Schiliro, E., Maldonado-Gonzalez, M.
M., Valderrama, R., Barroso-Albarracin,
J. B., and Mercado-Blanco, J. (2011). Root
hairs play a key role in the endophytic col-
onization of olive roots by Pseudomonas
spp. with biocontrol activity. Microb. Ecol.
62: 435–445.
Rosconi, F., Davyt, D., Martínez, V., Martínez, M.,
Abin-Carriquiry, J. A., Zane, H., et al. (2013).
Identification and structural characteriza-
tion of serobactins, a suite of lipopeptide
siderophores produced by the grass endo-
phyte Herbaspirillum seropedicae. Environ.
Microbiol. 15: 916–927.
Rosenblueth, M., and Martínez-Romero, E.
(2004). Rhizobium etli maize populations
and their competitiveness for root coloni-
zation. Arch. Microbiol. 181: 337–344.
Santos, F., Peñaflor, M. F. G., Paré, P. W.,
Sanches, P. A., Kamiya, A. C., Tonelli, M.,
et al. (2014). A novel interaction between
plant-beneficial rhizobacteria and roots:
colonization induces corn resistance
against the root herbivore Diabrotica spe-
ciosa. PLoS ONE 9: e113280.
Santoyo, G., Moreno-Hagelsieb, G., Orozco-
Mosqueda Mdel, C., and Glick, B. R. (2016).
Plant growth-promoting bacterial endo-
phytes. Microbiol. Res. 183: 92–99.
Schaefer, A. L., Oda, Y., Coutinho, B. G., Pel-
letier, D. A., Weiburg, J., Venturi, V., et al.
(2016). A LuxR homolog in a cottonwood
tree endophyte that activates gene expres-
sion in response to a plant signal or spe-
cific peptides. mBio 7: e01101–16.
Shidore, T., Dinse, T., Ohrlein, J., Becker, A., and
Reinhold-Hurek, B. (2012). Transcriptomic
analysis of responses to exudates reveal
genes required for rhizosphere compe-
tence of the endophyte Azoarcus sp. strain
BH72. Environ. Microbiol. 14: 2775–2787.
Siguier, P., Pérochon, J., Lestrade, L., Mahillon, J.,
and Chandler, M. (2006). ISfinder: the refer-
ence centre for bacterial insertion sequences.
Nucleic Acids Res. 34 (Suppl. 1): D32–D36.
Song, Y., Tong, Z., Wang, J., Wang, L., Guo, Z., Han,
Y., et al. (2004). Complete genome sequence
of Yersinia pestis strain 91001, an isolate avir-
ulent to humans. DNA Res. 11: 179–197.
Taghavi, S., Wu, X., Ouyang, L., Zhang, Y. B.,
Stadler, A., McCorkle, S., et al. (2015). Tran-
scriptional responses to sucrose mimic
the plant-associated lifestyle of the plant
growth promoting endophyte Enterobacter
sp. 638. PLoS ONE 10: e0115455.
Tatusov, R. L., Fedorova, N. D., Jackson, J. D., Ja-
cobs, A. R., Kiryutin, B., Koonin, E. V., et al.
(2003). The COG database: an updated
version includes eukaryotes. BMC Bioin-
formatics 4: 41.
Timm, C. M., Campbell, A. G., Utturkar, S. M.,
Jun, S. R., Parales, R. E., Tan, W. A., et al.
(2015). Metabolic functions of Pseudo-
monas fluorescens strains from Populus
deltoides depend on rhizosphere or endo-
sphere isolation compartment. Front. Mi-
crobiol. 6: 1118.
van Peer, R., Punte, H. L., de Weger, L. A., and
Schippers, B. (1990). Characterization of
root surface and endorhizosphere pseu-
domonads in relation to their coloniza-
tion of roots. Appl. Environ. Microbiol. 56:
2462–2470.
Wakelin, S. A., Warren, R. A., Harvey, P. R., and
Ryder, M. H. (2004). Phosphate solubiliza-
tion by Penicillium spp. closely associated
with wheat roots. Biol. Fertil. Soils 40: 36–43.
Wilson, A. C., Perego, M., and Hoch, J. A. (2007).
New transposon delivery plasmids for in-
sertional mutagenesis in Bacillus an-
thracis. J. Microbiol. Methods 71, 332–335.
Xie, S., Wu, H., Chen, L., Zang, H., Xie, Y., and
Gao, X. (2015). Transcriptome profiling of
Bacillus subtilis OKB105 in response to rice
seedlings. BMC Microbiol. 15: 21.
Yi, H.-S., Yang, J. W., and Ryu, C.-M. (2013). ISR
meets SAR outside: additive action of the
endophyte Bacillus pumilus INR7 and the
chemical inducer, benzothiadiazole, on in-
duced resistance against bacterial spot in
field-grown pepper. Front. Plant Sci. 4: 122.
Yi, Y., and Kuipers, O. P. (2017). Development
of an efficient electroporation method for
rhizobacterial Bacillus mycoides strains. J.
Microbiol. Methods 133: 82–86.
Zerbino, D. R., and Birney, E. (2008). Velvet: al-
gorithms for de novo short read assembly
using de Bruijn graphs. Genome Res. 18:
821–829.
Zhang, N., Wang, D., Liu, Y., Li, S., Shen, Q., and
Zhang, R. (2014). Effects of different plant
root exudates and their organic acid com-
ponents on chemotaxis, biofilm forma-
tion and colonization by beneficial rhizo-
sphere-associated bacterial strains. Plant
Soil 374: 689–700.
Zhang, N., Yang, D., Wang, D., Miao, Y., Shao, J.,
Zhou, X., et al. (2015). Whole transcrip-
tomic analysis of the plant-beneficial rhizo-
bacterium Bacillus amyloliquefaciens SQR9
during enhanced biofilm formation regu-
lated by maize root exudates. BMC Genom-
ics 16: 685.
Zhou, Y., Liang, Y., Lynch, K. H., Dennis, J. J., and
Wishart, D. S. (2011). PHAST: a fast phage
search tool. Nucleic Acids Res. 39(Suppl. 2):
W347–W352.
Zinser, E. R., and Kolter, R. (1999). Mutations
enhancing amino acid catabolism confer
a growth advantage in stationary phase. J.
Bacteriol. 181: 5800–5807.

4Development of an efficient electroporation method for rhizobacterial Bacillus mycoides strains
Yanglei Yi, Oscar P. Kuipers
Molecular Genetics, Groningen Biomolecular Sciences and Biotechnology Institute, University of Groningen, Groningen, The Netherlands
This chapter was published as:Yi, Y., & Kuipers, O. P. (2017). Development of an efficient electroporation method for rhizobacterial Bacillus mycoides strains. Journal of microbiological methods, 133: 82–86.

4
67
Dev
elop
men
t of a
n effi
cien
t ele
ctro
pora
tion
met
hod
for r
hizo
bact
eria
l Ba
cillu
s myc
oide
s str
ains
ABSTRACT
In order to develop a method for electroporation of environmental Bacil-lus mycoides strains, we optimized several conditions that affect the elec-troporation efficiency of this bacterium. By combining the optimized conditions, the electroporation efficiency of strain EC18 was improved to (1.3 ± 0.6) × 105 cfu/μg DNA, which is about 103-fold increase in com-parison with previously reported value. The method was further vali-dated on various B. mycoides strains, yielding reasonable transformation efficiencies. Furthermore, we confirmed that restriction/modification is the main barrier for electroporation of this bacterium. To the best of our knowledge, this is the first systematic investigation of various parameters of electroporation of B. mycoides. The electroporation method reported will allow for efficient genetic manipulation of this bacterium.
INTRODUCTION
Bacillus mycoides is a spore-forming Gram-positive bacterium that is commonly found in soil and the rhizosphere. It belongs to the B. cereus sensu lato group, which includes B. cereus, B. thuringiensis, and B. anthracis. B. mycoides has received the least attention among this group because it is not a human pathogen as B. anthracis and B. cereus. Moreover, there are several reports on the insecticidal effect of B. thuringiensis, which stimu-lated further studies using genetic modification (Turchi et al., 2012). The B. mycoides species features a unique filamentous growth pattern, either being rotated clockwise or counterclockwise (Di Franco et al., 2002). Now-adays, more and more studies on B. mycoides are focusing on their plant-growth promoting activities (Ambrosini et al., 2016; Bargabus et al., 2002; Neher et al., 2009).
In order to study the plant interaction mechanism of B. mycoides, it is necessary to be able to perform molecular genetics studies on this bac-terial species. A high transformation efficiency is required to establish the genetic manipulation systems, e.g., enabling gene deletion and mu-tation within the genome of this organism. Several techniques, includ-ing phage transduction, protoplast transformation, natural competence, and electroporation have been applied to incorporate exogenous DNA into Bacillus cells (Barlass et al., 2002; Lu et al., 2012; Romero et al., 2006). Among these methods, electroporation is usually the quickest and most reproducible method. Di Franco et al. (2002) modified the electrotrans-formation protocol of Macaluso and Mettus (1991) for B. thuringiensis, but the implementation on B. mycoides resulted in a very low efficiency (less than 200 cfu/μg plasmid DNA). Our preliminary experiments show that by applying the method reported by Ehling-Schulz et al. (2005), which was originally developed for B. cereus, the B. mycoides strain EC18 could be transformed, albeit at a low efficiency. Apart from this, no other pro-tocols have been developed on electroporation methods for B. mycoides. This work aimed at developing an efficient electroporation method for several environmental B. mycoides strains. B. mycoides EC18 which was isolated from the endosphere of potato, and displaying potential plant growth-promoting activity (data not shown), was used as a first testing strain. Factors including growth media, growth phase, electroporation buffer, pulse strength, and incubation time that can affect electroporation efficiency were optimized. As a result, a high electroporation efficiency of (1.3 ± 0.6) × 105 cfu/μg DNA) was obtained. Furthermore, we evalu-ated our optimized protocol on other B. mycoides strains as well, which resulted in electroporation efficiencies ranging from (7.3 ± 2.1) × 102 to (1.3 ± 0.6) × 105 cfu/μg DNA.

44
68 69
Dev
elop
men
t of a
n effi
cien
t ele
ctro
pora
tion
met
hod
for r
hizo
bact
eria
l Ba
cillu
s myc
oide
s str
ains
Mat
eria
ls a
nd m
etho
ds
MATERIALS AND METHODS
Bacterial strains, plasmids, and mediaEscherichia coli strains were grown in Luria broth at 37 °C, 220 rpm. B. my-coides strains were isolated from a potato field in Wijster (the Nether-lands) (Table 3) and were grown at 30 °C, 200 rpm in LB. When necessary, 100 μg/ml of ampicillin or 4 μg/ml of chloramphenicol was added to the culture medium. Plasmid DNA from E. coli was purified using the Nucle-oSpin plasmid isolation kit (Macherey-Nagel GmbH & Co. Duren, Ger-many) according to the manufacturer’s instructions. A Geobacillus–E. coli shuttle vector pNW33N (Bacillus Genetic Stock Centre) isolated from E. coli MC1061was used as the plasmid for electroporation protocol opti-mization. pNW33N isolated from E. coli JM110 was used to test the plasmid methylation effects on transformation efficiency.
Screening for growth mediumThe B. mycoides strain was streaked on LB agar plate and grown at 30 °C overnight. One single colony was inoculated into various media including LB (10 g tryptone, 5 g yeast extract, 10 g NaCl in 1 L deionized water, pH 7.2), LBS (10 g tryptone, 5 g yeast extract, 10 g NaCl, and 91.1 g sorbitol in 1 L deionized water, pH 7.2), LBSP (10 g tryptone, 5 g yeast extract, 10 g NaCl, 50 mMKH2PO4 and K2HPO4, and 91.1 g sorbitol in 1 L deionizedwater, pH 7.2), 2×YT (16 g tryptone, 10 g yeast extract and 5 g NaCl in 1 L deionized water, pH 7.2), and BHIS (34 g BHI, and 91.1 g sorbitol in 1 L deionized water, pH 7.2).
Preparation of electro-competent cellsB. mycoides cells were grown overnight in an appropriate medium at 30 °C, 200 rpm. The culture was diluted with the same medium to obtain an initial optical density at 600 nm (OD600) of about 0.05 then continu-ally grown. The OD600 was measured by a Genesys 20 spectrophotome-ter (ThermoSpectonic, USA) during bacterial growth. When the OD600 reached the appropriate value, the cell culture was transferred into a 50 mL centrifuge tube and cooled on ice for 10 min. Cells were then col-lected by centrifugation at 4 °C, 3,000 × g for 10 min. After being washed four times in corresponding electroporation buffer (pre-chilled), the cell pellets were re-suspended in 1 mL of the electroporation buffer. The re-sulting electro-competent cells were flash-frozen in liquid nitrogen and stored at −80 °C prior to electroporation.
Cell wall-weakening treatmentSerial concentrations of Glycine or DL-threonine were added when bacterial cultures reached an OD600 = 0.85 for the cell-wall weakening
treatment. These treated bacterial cultures were continued to shake for 1 h. After that, electro-competent cells were prepared with the methods mentioned above.
ElectroporationThe electroporation was performed as previously reported method (Peng et al., 2009; Turgeon et al., 2006) with slight modification. 100 μl of fro-zen electro-competent cells was thawed on ice and mixed with 2–3 μl of plasmid DNA (1 μg). Cells mixed with the same amount of deionized wa-ter served as negative control. The mixture was loaded into an electropora-tion cuvette (2 mm electrode gap, pre-chilled) and exposed to a single pulse in a Gene Pulser System (Bio-Rad, USA) with the settings of 25 μF capac-itance, 100, 200, or 400 Ω resistance, and voltage ranges between 5 and 12.5 kV cm−1. After pulse-shock, cells were immediately added with 1 ml of pre-warmed corresponding growth medium and transferred to a 2 mL Eppendorf tube. After 2 h incubation at 30 °C, 180 rpm, the dilutions of the recovered cell culture were plated on LB agar with 4 μg/ml of chloramphen-icol. The plates were incubated at 30 °C overnight and transformation effi-ciencies (cfu/μg DNA) were calculated by counting the colonies on plates.
RESULTS AND DISCUSSION
Optimization of growth conditionsFor determining the optimal growth medium, we transformed B. my-coides according to the protocol published by Ehling-Schulz et al. (2005). Strain EC18 was grown in different media before preparing the electro- competent cells. Media including LB, LBS, LBSP, 2×YT and BHIS were cho-sen according to reported electroporation methods for Bacillus species (Zhang et al., 2011a, 2015). When the OD600 reached a value of 0.6, the competent cells were prepared by washing the cell pellet with increasing concentrations of ice-cold glycerol (2.5, 5, and 10%). Electroporation was performed with 1 μg pNW33N plasmid DNA and recovered for 2 h with the corresponding growth media. The efficiencies were calculated af-ter 1 day (Table 1) on 4 μg/ml chloramphenicol selection media. Among the five tested media, the super-rich medium BHIS showed the highest efficiency of (6.2 ± 1.4) × 103 cfu/μg plasmid DNA. Zhang et al. (2011a) re-ported that the transformation efficiency of B. amyloliquefaciens was pos-itively correlated to concentrations of salts, whereas negatively related to the nutritional ingredient concentration. Surprisingly, we observed an opposite effect of the nutritional and salt concentrations on transforma-tion efficiencies in our study. The transformation efficiency by culturing

44
70 71
Dev
elop
men
t of a
n effi
cien
t ele
ctro
pora
tion
met
hod
for r
hizo
bact
eria
l Ba
cillu
s myc
oide
s str
ains
Resu
lts a
nd d
iscu
ssio
n
in the LBSP medium is lower than culturing in LBS medium. Moreover, B. mycoides EC18 could not grow in the hypertonic media NCM and M9YE (data not shown). We hypothesized that B. mycoides EC18 is sensitive to salt concentrations.
Table 1. Effect of growth media on the electroporation efficiency of B. mycoides EC18.
Medium Transformation efficiency ( cfu/μg DNA)LB (3.4 ± 1.0) ×102
LBS (8.3 ± 2.5) ×102
LBSP (2.0 ± 1.1) ×102
2x YT (5.4 ± 1.3) ×102
BHIS (6.2 ± 1.4) ×103
Cells were grown in different media to OD600 ~ 0.6 to prepare the electro-competent cells using series concentration of glycerol solution as electroporation buffer. 1 μg of pNW33N plasmid were used for electroporation with the settings 25 μF, 10 kV cm-1, 200 Ohms. Data are shown as mean ± standard deviation based on 3 replications.
According to previous reports, cells of B. cereus and B. thuringiensis col-lected at early growth-stage show better electroporation efficiency than late growth-stage cultures (Peng et al., 2009; Turgeon et al., 2006), while B. subtilis WB800 has high electroporation efficiency of 1.88 × 10 5cfu/μg DNA at late growth-stage (OD600 = 2.2–2.3) (Lu et al., 2012). To investigate the effects of the growth phase of B. mycoides on the electroporation ef-ficiency, B. mycoides cells were cultured in BHIS medium to OD600 from 0.3 to 1.8 for competent cell preparations. Our results indicated thatwhen OD600 is between 0.9 and 1, the highest electroporation efficiency was ob-tained (Figure 1A). According to the growth curve (Figure 1B), cells are in the early stage of exponential growth. A similar phenomenonwas also reported for B. subtilis ZK by Zhang et al. (2015). This indicates that cells from the early exponential phase result in higher electroporation effi-ciency of B. mycoides.
Optimization of cell wall-weakening agentsThe cell wall-weakening agents, glycine and DL-threonine, are widely used to improve the electroporation efficiency of environmental strains. These amino acids can reduce the peptidoglycan bonds and loosen up the cell wall by replacing the L- and D-alanine bridges (Hammes et al., 1973). In this study, EC18 cells were first grown in BHIS medium, and when the OD600 reached about 0.85, glycine and threonine were added at different concentrations (0%, 1%, 2%, 3%, 4% and 5%). After 1 h of additional in-cubation, the cells were collected and concentrated to obtain competent cells. Electroporation treatments were performed and the greatest trans-formation efficiency was obtained in the 2% glycine treatment group. The same concentration of threonine also improved the transformation efficiency, which was however lower than that of glycine (Figure 2). Cell growth rate was slightly reduced in the presence of both glycine and DL-threonine, and high concentrations of glycine treatments resulted in
Figure 1. Effect of growth phase on the electroporation efficiency of B. mycoides EC18. (A) The transformation efficiency at different growth phases of B. mycoides EC18. Cells were grown in BHIS medium to various OD600 density for the electro-competent cells preparation, using serial concentrations of a glycerol solution as electroporation buffer. 1 μg of pNW33N plasmid was used for electroporation with the settings 25 μF, 10 kV cm-1, 200 Ohms. (B) The growth of B. mycoides EC18 in BHIS medium at 30°C, 200 rpm. At different time points, the optical density of the cell cultures was measured spectropho-tometrically (OD600). (N = 3, bar = standard deviation)

44
72 73
Dev
elop
men
t of a
n effi
cien
t ele
ctro
pora
tion
met
hod
for r
hizo
bact
eria
l Ba
cillu
s myc
oide
s str
ains
Resu
lts a
nd d
iscu
ssio
n
cellular lysis in the samples (data not shown). These results demonstrate that the transformation efficiency of B. mycoides has been significantly enhanced by a cell wall-weakening treatment. The optimal results were obtained at incubations with 2% glycine.
Effects of buffer on the electroporation efficiencyAccording to previous studies, the composition of the electroporation buffer has a tremendous impact on transformation efficiency. Xue et al. (1998) improved the transformation efficiency of B. subtilis and B. licheni-formis by applying electroporation buffers with high osmolarity. It has also been reported that with the addition of trehalose, the transformation efficiency of B. subtilis can be enhanced dramatically to about 100-fold (Lu et al., 2012). In this study, EC18 cells growing in BHIS medium was added with 2% glycine when the cell density reached an OD600 = 0.85. After 1 h of additional incubation, cells were collected (OD600 ~0.95) and washed four times in different electroporation buffers (Table 2) to obtain compe-tent cells. The transformation efficiency is shown in Table 2. The buffers consisting of glycerol exhibited better transformation efficiencies than those with sucrose. The highest transformation efficiency was obtained
with buffer G containing 10% glycerol, 0.25 M sorbitol, and 0.25 M treha-lose. However, fewer transformants were obtained when the sorbitol and trehalose concentration increased to 0.5 M. The addition of salts (buffers K, L, and M) showed no improvement on transformation efficiency. Based on these results, buffer G was used throughout.
Effects of the electric field on the electroporation efficiencyA relatively high electrical field is necessary to create pores in the cells and thus provide a temporary pathway for exogenous DNA. After the electric pulse, they gradually reseal and most cells recover. However, if the electrical field is above a certain level, most pores either do not re-seal or reseal too slowly to preserve cell viability (Kotnik et al., 2015). To
Figure 2. Effect of wall-weakening agents on the electroporation efficiency of B. my-coides EC18. Cells were grown in BHIS medium to OD600 ~0.85, then incubated 1 h with different concentration of cell wall-weakening agents. Electro-competent cells were prepared using glycerol solutions as electroporation buffer. 1 μg of pNW33N plasmid were used for electroporation with the settings 25 μF, 10 kV cm-1, 200 Ohms. (N = 3, bar = standard deviation)
Table 2. Effect of electroporation buffers on transformation efficiencies in B. mycoides EC18.
Buffer Base component Salts component Transformation efficiency
(cfu/μg DNA)
Reference
A Glycerol solution (2.5, 5, and 10%)
- (4.6±1. 7) × 104 (Ehling-Schulz, et al., 2005)
B 10% glycerol - (4.0 ± 2.0) × 104 (Zhang et al., 2011)C 10% glycerol,
0.25 M sorbitol- (6.2 ± 2.8) × 104 (Zhang, et al., 2011)
D 10% glycerol, 0.5 M sorbitol
- (5.8 ± 1.8)× 104 (Zhang, et al., 2011)
E 10% glycerol, 0.25 M sorbitol, 0.25 M mannitol
- (4.2 ± 1.7)× 104 This study
F 10% glycerol, 0.5 M sorbitol, 0.5 M mannitol
- (4.0 ± 0.8)× 102 (Xue, et al., 1998)
G 10% glycerol, 0.25 M sorbitol, 0.25 M trehalose
- (6.8 ± 1.6)× 104 This study
H 10% glycerol, 0.5 M sorbitol, 0.5 M trehalose
- (4.0 ± 0.8)× 102 This study
I 10% glycerol, 0.25 M sorbitol, 0.25 M mannitol, 0.25 M trehalose
- (1.2 ± 0.8)× 102 This study
J 0.5 M sorbitol, 0.5 M mannitol, 0.5 M trehalose
- 44 ± 22 (Zhang, et al., 2015)
K 0.5 M sorbitol, 0.5 M mannitol, 0.5 M trehalose
0.5 mM MgCl2, 0.5 mM K2HPO4/KH2PO4
No transfor-mants
(Zhang, et al., 2015)
L 272 mM sucrose 0.5 mM MgCl2, 0.5 mM K2HPO4/KH2PO4
(5.2 ± 2.0)× 102 (Silo-Suh et al., 1994)
M 250 mM sucrose, 10% glycerol
1 mM Hepes; 1 mM MgCl2 No transfor-mants
(Turgeon, et al., 2006)
Cells were grown in BHIS medium to OD600 ~0.85, then incubated 1 h with 2% glycine. Electro-competent cells were prepared using different electroporation buffers. 1 μg of pNW33N plasmid was used for electroporation with the settings 25 μF, 10 kV cm-1, 200 Ohms. Data are shown as mean ± standard deviation of 3 replications.

44
74 75
Dev
elop
men
t of a
n effi
cien
t ele
ctro
pora
tion
met
hod
for r
hizo
bact
eria
l Ba
cillu
s myc
oide
s str
ains
Resu
lts a
nd d
iscu
ssio
n
optimize the electric field for B. mycoides EC18, a gradient of field strength (7.5–12.5 kV cm−1) and resistance values (100, 200, and 400 Ω) at 25 μF was tested. The results are shown in Figure 3, an electric field of 10 kV cm−1 and resistance of 200 Ω led to the optimal transformation efficiency. This electric field is lower than that of B. subtilis (Lu et al., 2012; Zhang et al., 2015) and closely related species, e.g., B. cereus and B. thuringiensis (Peng et al., 2009; Turgeon et al., 2006). These results indicate that B. mycoides is more vulnerable to high electric fields compared to other Bacillus species.
Effects of recovery timeAfter an electric pulse, the injured bacterial cells need several hours to re-tain viability and to allow phenotypic expression. To optimize the recovery time, the pulse-shocked cells were immediately transferred to BHIS me-dium and incubated at 30 °C, 180 rpm for 2–7 h. As shown in Figure 4, the transformation efficiency was increased as the incubation time extended. Remarkably, the transformation efficiency reached 1.2 × 105 cfu/μg DNA after 5 h of incubation. Further elongation of the incubation time only led to a slight increase in transformation efficiency. Hence, we used 5 h
of recovery time for B. mycoides. Xue et al. (1998) pointed out that the in-creased transformation efficiency at the longer recovery times might be due to the division of transformed cells. Zhang et al. (2011b) also used a long recovery time of 5 hrs for electroporation of Arthrobacter.
Application of the optimized transformation protocol on other B. mycoides strains and the effects of plasmid methylationIn order to validate the optimized protocol in other environmental B. my-coides isolates, we combined all the improved factors and applied the method on 5 other strains in addition to strain EC18. Since the restriction of methylated DNA in several Bacillus species is known as a major bar-rier to transformation and genetic manipulation (Sitaraman and Leppla, 2012), we also compared the transformation efficiency of plasmids that were isolated from E. coli methylation strain MC1061 (dam+/dcm+) and methylation-deficient strain JM110 (dam−/dcm−). As shown in Table 3, for the plasmid isolated from E. coli MC1061, only strain EC18 showed high transformation efficiency of (1.1 ± 0.6) × 105 cfu/μg DNA. Notably, strain SB4 was transformed at a much lower level of efficiency and the other
Figure 3. Effect of electric field on the electroporation efficiency of B. mycoides EC18. Cells were grown in BHIS medium to OD600 ~0.85, then incubated 1 h with 2% glycine. Electro-competent cells were prepared using buffer G. 1 μg of pNW33N plasmid were used for electroporation with a gradient of field strength (7.5–12.5 kV·cm-1) and resis-tance values (100, 200, and 400 Ohms) at 25 μF. (N = 3, bar = standard deviation)
Figure 4. Effect of incubation time on the electroporation efficiency of B. mycoides EC18. Cells were grown in BHIS medium to OD600 ~0.85, then incubated 1 h with 2% glycine. Electro-competent cells were prepared using buffer G. 1 μg of pNW33N plasmid were used for electroporation with the settings 25 μF, 10 kV cm-1, 200 Ohms. Cells were then incubated at 30 °C, 180 rpm for 2–7 hrs. (N = 3, bar = standard deviation)

44
76 77
Dev
elop
men
t of a
n effi
cien
t ele
ctro
pora
tion
met
hod
for r
hizo
bact
eria
l Ba
cillu
s myc
oide
s str
ains
Conc
lusi
on
strains were not transformable at all. However, when non- methylated plasmid DNA was used, this protocol can efficiently be applied for all the tested B. mycoides strains. The transformation efficiencies are ranging from (7.3 ± 2.1) × 102 to (1.3 ± 0.6) × 105 cfu/μg DNA. These results indi-cate that our optimized protocol can be widely used in B. mycoides strains. Methylation of transmitted DNA can clearly affect the efficiency of trans-formation, with the exception of strain EC18, which only showed a slight difference between methylated and non-methylated DNA.
CONCLUSION
This paper describes an efficient electroporation method for several en-vironmental B. mycoides strains, which is reproducible and conven-ient. By using this method, we obtained transformation efficiency up to (1.3 ± 0.6) × 105 cfu/μg DNA. We confirmed that our protocol can be applied to several B. mycoides strains. The method described here facilitates ad-vanced molecular genetics studies in this important biocontrol bacterium.
ACKNOWLEDGEMENT
We are grateful to Prof. J.D. van Elsas and J. Spoelder (University of Gronin-gen, the Netherland) for providing the B. mycoides strains. We thank Dr. C. Song in our group for critical reading and suggestions. Y. Yi was supported by a scholarship from the China Scholarship Council (201306300040).
REFERENCES
Ambrosini, A., Stefanski, T., Lisboa, B.B., Ben-
eduzi, A., Vargas, L.K., Passaglia, L.M.P.,
2016. Diazotrophic bacilli isolated from
the sunflower rhizosphere and the poten-
tial of Bacillus mycoides B38V as biofer-
tiliser. Ann. Appl. Biol. 168: 93–110.
Bargabus, R., Zidack, N., Sherwood, J., Jacob-
sen, B., 2002. Characterisation of sys-
temic resistance in sugar beet elicited by a
non-pathogenic, phyllosphere-colonizing
Bacillus mycoides, biological control agent.
Physiol. Mol. Plant Pathol. 61: 289–298.
Barlass, P.J., Houston, C.W., Clements, M.O.,
Moir, A., 2002. Germination of Bacillus ce-
reus spores in response to L-alanine and to
inosine: the roles of gerL and gerQ oper-
onsc. Microbiology 148: 2089–2095.
Di Franco, C., Beccari, E., Santini, T., Pisaneschi,
G., Tecce, G., 2002. Colony shape as a ge-
netic trait in the pattern-forming Bacillus
mycoides. BMC Microbiol. 2: 33.
Ehling-Schulz, M., Vukov, N., Schulz, A., Sha-
heen, R., Andersson, M., Märtlbauer, E.,
et al., 2005. Identification and partial
characterization of the nonribosomal pep-
tide synthetase gene responsible for cere-
ulide production in emetic Bacillus cereus.
Appl. Environ. Microbiol. 71: 105–113.
Hammes, W., Schleifer, K., Kandler, O., 1973.Mode
of action of glycine on the biosynthesis of
peptidoglycan. J. Bacteriol. 116: 1029–1053.
Kotnik, T., Frey,W., Sack, M., Haberl Meglic, S.,
Peterka, M., Miklavcic, D., 2015. Electropo-
ration-based applications in biotechnol-
ogy. Trends Biotechnol. 33: 480–488.
Lu, Y.P., Zhang, C., Lv, F.X., Bie, X.M., Lu, Z.X.,
2012. Study on the electro-transformation
conditions of improving transformation
efficiency for Bacillus subtilis. Lett. Appl.
Microbiol. 55: 9–14.
Table 3. Electroporation of wild type B. mycoides strains with methylated and non- methylated plasmid DNA.
Strain Electroporation efficiency of different plasmid source(cfu/μg DNA) E.coli MC 1061 (dam+/dcm+) E. coli JM110 (dam-/dcm-)
B. mycoides EC18 (1.1 ± 0.6) ×105 (1.3 ± 0.6) × 105
B. mycoides M2E15 - (3.7 ± 1.2) × 103
B. mycoides S2E19 - (1.7 ± 0.4) × 104
B. mycoides S3E15 - (7.3 ± 2.1) × 102
B. mycoides SB4 (1.8 ± 0.6) × 103 (6.4 ± 1.2) × 104
B. mycoides SB8 - (5.7 ± 1.6) × 104
Cells were grown in BHIS medium to OD600 ~0.85, then incubated 1 h with 2% glycine. Electro-competent cells were prepared using buffer G. 1 μg of PNW33N plasmid were used for electroporation with the settings as: 25 μF, 10 kV cm-1, 200 Ohms. Cells were then incubated at 30 °C,180 rpm for 5 hrs. Data are shown as mean ± standard deviation of 3 replications.
Macaluso, A., Mettus, A.M., 1991. Efficient
transformation of Bacillus thuringiensis
requires nonmethylated plasmid DNA. J.
Bacteriol. 173: 1353–1356.
Neher, O.T., Johnston, M.R., Zidack, N.K., Ja-
cobsen, B.J., 2009. Evaluation of Bacillus
mycoides isolate BmJ and B. mojavensis iso-
late 203-7 for the control of anthracnose of
cucurbits caused by Glomerella cingulata
var. orbiculare. Biol. Control 48: 140–146.
Peng, D., Luo, Y., Guo, S., Zeng, H., Ju, S., Yu,
Z., et al., 2009. Elaboration of an electro-
poration protocol for large plasmids and
wild-type strains of Bacillus thuringiensis. J.
Appl. Microbiol. 106: 1849–1858.
Romero, D., Perez-Garcia, A., Veening, J.W., de
Vicente, A., Kuipers, O.P., 2006. Transfor-
mation of undomesticated strains of Bacil-
lus subtilis by protoplast electroporation. J.
Microbiol. Methods 66: 556–559.
Silo-Suh, L.A., Lethbridge, B.J., Raffel, S.J., He,
H., Clardy, J., Handelsman, J., 1994. Biolog-
ical activities of two fungistatic antibiotics
produced by Bacillus cereus UW85. Appl.
Environ. Microbiol. 60: 2023–2030.
Sitaraman, R., Leppla, S.H., 2012. Methyla-
tion-dependent DNA restriction in Bacil-
lus anthracis. Gene 494: 44–50.
Turchi, L., Santini, T., Beccari, E., Di Franco, C.,
2012. Localization of new peptidoglycan at
poles in Bacillus mycoides, a member of the
Bacillus cereus group. Arch.Microbiol. 194:
887–892.
Turgeon, N., Laflamme, C., Ho, J., Duchaine, C.,
2006. Elaboration of an electroporation
protocol for Bacillus cereus ATCC 14579. J.
Microbiol. Methods 67: 543–548.
Xue, G.-P., Johnson, J.S., Dalrymple, B.P.,
1998. High osmolarity improves the
electrotransformation efficiency of the

4
78
Refe
renc
es
gram-positive bacteria Bacillus subtilis and
Bacillus licheniformis. J. Microbiol. Methods
34: 183–191.
Zhang, G.Q., Bao, P., Zhang, Y., Deng, A.H.,
Chen, N., Wen, T.Y., 2011a. Enhancing
electro-transformation competency of
recalcitrant Bacillus amyloliquefaciens
by combining cell-wall weakening and
cell-membrane fluidity disturbing. Anal.
Biochem. 409: 130–137.
Zhang, H., Li, Y., Chen, X., Sheng, H., An, L.,
2011b. Optimization of electroporation
conditions for Arthrobacter with plasmid
PART2. J. Microbiol. Methods 84: 114–120.
Zhang, Z., Ding, Z.T., Shu, D., Luo, D., Tan, H.,
2015. Development of an efficient electro-
poration method for iturin A- producing
Bacillus subtilis ZK. Int. J. Mol. Sci. 16:
7334–7351.

5Optimized fluorescent proteins for the rhizosphere-associated bacterium Bacillus mycoides with endophytic and biocontrol agent potential
Yanglei Yi1†, Elrike Frenzel1†, Jan Spoelder2, J. Theo M. Elzenga2, Jan Dirk van Elsas3 and Oscar P. Kuipers1
¹Department of Molecular Genetics, Groningen Biomolecular Sciences and Biotechnology Institute, University of Groningen, Groningen, the Netherlands.
²Plant Physiology, Groningen Institute for Evolutionary Life Sciences, University of Groningen, Groningen, the Netherlands.
³Microbial Ecology, Groningen Institute for Evolutionary Life Sciences, University of Groningen, Groningen, the Netherlands. †equal contribution to this work
This chapter was published as:Yi, Y., Frenzel, E., Spoelder, J., Elzenga, J. T. M., van Elsas, J. D., & Kuipers, O. P. (2018). Optimized fluorescent proteins for the rhizosphere-associated bacterium Bacillus mycoides with endophytic and biocontrol agent potential. Environmental microbiology reports. 10(1):57–74.

5
83
Opt
imiz
ed fl
uore
scen
t pro
tein
s for
the
rhiz
osph
ere-
asso
ciat
ed
bact
eriu
m B
acill
us m
ycoi
des w
ith e
ndop
hytic
and
bio
cont
rol a
gent
pot
entia
l
ABSTRACT
Tracking of fluorescent protein (FP)-labelled rhizobacteria is a key prereq-uisite to gain insights into plant-bacteria interaction mechanisms. How-ever, the performance of FPs mostly has to be optimized for the bacterial host and for the environment of intended application. We report on the construction of mutational libraries of the superfolder green fluorescent protein sfGFP and the red fluorescent protein mKate2 in the bacterium B. mycoides, which next to its potential as plant-biocontrol agent occasion-ally enters an endophytic lifestyle. By fluorescence-activated cell sorting and comparison of signal intensities at the colony and single-cell level, the variants sfGFP (SPS6) and mKate (KPS12) with significantly increased brightness were isolated. Their high applicability for plant-bacteria inter-action studies was shown by confocal laser scanning microscopy track-ing of FP-tagged B. mycoides strains after inoculation to Chinese cabbage plants in a hydroponic system. During the process of colonization, strain EC18 rapidly attached to plant roots and formed a multicellular matrix, especially at the branching regions of the root hair, which probably con-stitute entrance sites to establish an endophytic lifestyle. The universal applicability of the novels FPs was proven by expression from a weak pro-moter, dual-labelling of B. mycoides, and by excellent expression and de-tectability in additional soil- and rhizosphere-associated Bacillus species.
INTRODUCTION
Fluorescent proteins (FPs) are widely used in living prokaryotic and eukar-yotic cells as genetically encoded fluorescent labels to study cell motility, changes in gene activity and protein localization and dynamics (Chudakov et al., 2010; Kremers et al., 2011). After discovery of the first FP, the green flu-orescent protein (GFP) of Aequoria victoria (Shimomura et al., 1962), cloning of its structural gene paved the way for extensive protein engineering stud-ies (Prasher et al., 1992). These resulted in a plethora of diverse FPs with emis-sion light wavelengths ranging from blue at 448 nm to yellow at 526 nm. Un-til now, the palette of colour variants for multicolour imaging is constantly expanded (Day and Davidson, 2009). Extensive efforts have also been made to identify and engineer red FPs (RFP) that emit in the yellow-orange to far-red regions of the visible light spectrum (Piatkevich et al., 2010; Shemiakina et al., 2012; Rodriguez et al., 2016). The usage of RFPs is especially advanta-geous in mammalian cells or plant tissues because these are more transpar-ent to red light (Rizzo et al., 2009). This enables high-contrast imaging due to a low autofluorescence background, as RFPs are highly compatible with existing confocal-microscope lasers and the respective filter sets.
In environmental microbiology, ecophysiology and particularly in plant-microorganism interaction studies, the application of FP markers has recently become a powerful approach for exploring microbial func-tions in situ in natural ecosystems, e.g., the rhizosphere and plant en-dosphere. Applying biosensor strains for analysing the microbial function in symbiotic or competing communities has led to significant advances in these areas (Larrainzar et al., 2005). However, an on-site and widespread application of FPs in plant ecophysiology is still restricted by the complex-ity of the rhizosphere and endosphere environments and matrices. Unlike the laboratory-based in vitro systems, rhizosphere samples are usually as-sociated with complex organic and inorganic materials, which show high background levels of autofluorescence when studied with visualization tools like fluorescence microscopy. Moreover, plant roots are able to ac-tively alter the rhizospheric oxygen content and pH (Blossfeld et al., 2011), which in turn can affect the brightness of FPs by impeding chromophore maturation (Heim et al., 1994; Shu et al., 2006), altering the chromo-phore protonation state (Das et al., 2003) or causing misfolding of the FP (Craggs, 2009). Additionally, the functionality of FPs is highly dependent on the bacterial expression host and often needs to be validated or even optimized to fulfil the desired experimental requirements ( Hebisch et al., 2013). For in situ studies using FP-tagged organisms, it is, therefore, of spe-cial importance to consider that extrinsic and intrinsic cellular factors im-pact or modulate the performance of FPs (Shaner et al., 2005).

55
84 85
Opt
imiz
ed fl
uore
scen
t pro
tein
s for
the
rhiz
osph
ere-
asso
ciat
ed
bact
eriu
m B
acill
us m
ycoi
des w
ith e
ndop
hytic
and
bio
cont
rol a
gent
pot
entia
l
Intr
oduc
tion
B. mycoides is a chain-forming bacterium, which is associated with the Bacillus cereus sensu lato group. This species has a particular asymmetric ‘hairy’ shape on agar plates. The bundles of filaments resulting from ex-tensive chaining and linkage of cells show either a clockwise or counter-clockwise growth pattern (Di Franco et al., 2002). B. mycoides is ubiquitous and abundant in soils and the rhizosphere of plants, its natural niches (Neher et al., 2009; Ambrosini et al., 2016). Although B. mycoides was oc-casionally isolated from food cross-contaminated from soil ( Samapundo et al., 2014), it is widely recognized as a non-pathogenic bacterium (Na-kamura and Jackson, 1995). It has a low thermo tolerance of 37 °C with an optimal growth temperature between 25 °C and 30 °C (Guinebretière et al., 2008). Various B. mycoides strains isolated from the rhizosphere show plant growth-promoting effects on several crops. It was for instance shown that the B. mycoides isolate S4 promotes phosphorous solubilisa-tion and iron release by its siderophore production activity, which in-creases the photosynthesis and chlorophyll content of the runner bean Phaseolus coccineus L. (Stefan et al., 2013). Moreover, elicitation of an in-duced systematic resistance (ISR) by Bacillus strains led to a significant re-duction in the severity or incidence rates of various diseases on a diversity of plant hosts (Kloepper et al., 2004). Elicitation of ISR on sugar-beet was found to be associated with increased peroxidase activity coupled to an enhanced production of chitinase by the B. mycoides strain BmJ ( Bargabus et al., 2002; Bargabus et al., 2004). This strain was furthermore able to control anthracnose of cucurbits through the induction of systemic ac-quired resistance (SAR) (Neher et al., 2009). The biocontrol potential of B. mycoides against the plant pathogens Sclerotinia sclerotiorum or Botry-tis cinerea is based on the bacterial production of antimicrobial products such as bacillomycin D, fengycin, zwittermicin A or volatiles (Guetsky et al., 2002; Athukorala et al., 2009).
We isolated B. mycoides strains from the endosphere of healthy potato plants (Yi et al., 2017), which indicates that the interaction could be com-mensal or mutualistic. However, the ecological relationship between dif-ferent B. mycoides strains and plants has not been exhaustively studied. The observation of various stages of the colonization processes is critical to understand the physiological and molecular mechanisms of bacteria- plant interaction. Although there are many fluorescent tools available for low-GC Gram-positive spore formers, they are not optimized for use in en-dophytic B. mycoides. The lack of optimized fluorescent proteins to label this rhizobacterium for tracking and visualizing its development in planta or in hydroponic culture systems, and for labelling of promoters to ana-lyse factors that contribute to its endophytic lifestyle is currently a bottle-neck and necessitates further tool development.
In this study, we applied a random mutagenesis approach to generate mutational libraries of the green fluorescent protein sfGFP and the red fluorescent protein mKate2. After in vivo isolation of single mutants by fluorescence-activated cell sorting (FACS) and screening of fluorescence intensities during B. mycoides colony development by stereo fluores-cence microscopy, three brightly expressed candidates for each FP were obtained. Intriguingly, variants selected under pH shifted conditions showed the highest improvement in fluorescence signal intensity. We further demonstrate that these FP variants are suitable for B. mycoides-in planta localization studies with the advantage of showing an improved background signal-to-noise ratio. The universal applicability of the nov-els FPs was further proven by their detectability even when expressed from a weak promoter in B. mycoides and by excellent expression and de-tectability in additional soil- and rhizosphere-associated Bacillus species. The successful double-labelling and dual-colour imaging of B. mycoides indicates that the improved FPs reported here can be further applied for advanced molecular genetic studies, such as gene expression and protein localization in B. mycoides.
METEARIALS AND METHODS
Strains and growth conditions All strains, plasmids and primers used in this study are listed in Table 1. B. mycoides EC18 was isolated from the endosphere of a potato plant (Wijster, the Netherlands). B. subtilis HS3 and B. amyloliquefaciens HS9 were isolated from grass rhizosphere (Groningen, the Netherlands). The Bacillus strains were routinely cultured in Luria-Bertani (LB) medium at 30 °C with aeration at 200 rpm. All Escherichia coli strains were cultured in LB broth at 37 °C with aeration at 220 rpm. For cloning and selection purposes, ampicillin was added at a concentration of 100 μg/ml for E. coli and chloramphenicol and spectinomycin at a concentration of 4 μg/ml (LB-Cm4) and 100 μg/ml (LB-Spc100) for Bacillus strains respectively.
Random mutagenesis of fluorescent protein genes for E. coli library construction E. coli libraries of randomly mutagenized sfGFP(Sp) or mKate2 proteins (Table 1) were generated with the GeneMorph II Random mutagenesis kit according to manufacturer’s instructions (Agilent Technologies) as described elsewhere (Frenzel et al., 2017). In brief, the sfgfp(Sp) gene en-coded on plasmid pKB01-sfGFP(Sp) was mutagenized by error-prone PCR using the primer pair pKBO1derMut_F and pKBO1derMut_R (Table 1). The

55
86 87
Opt
imiz
ed fl
uore
scen
t pro
tein
s for
the
rhiz
osph
ere-
asso
ciat
ed
bact
eriu
m B
acill
us m
ycoi
des w
ith e
ndop
hytic
and
bio
cont
rol a
gent
pot
entia
l
MET
EARI
ALS
AN
D M
ETH
OD
S
XbaI/SphI digested PCR products were ligated into the equally cut replica-tive Bacillus/E. coli-shuttle plasmid pNW-Ppta-3TER (Table 1), which con-tains a Parageobacillus thermoglucosidans-derived constitutive promoter of the housekeeping pta gene and a threefold transcriptional terminator. In the same manner, the primer mKa-te2Mut_F and mKate2Mut_R (Ta-ble 1) were used to amplify and mutagenize mKate2 from 0.1 ng of target DNA residing on plasmid pAD651 (Table 1). The XbaI/ SphI cut fragments were cloned into the same restriction sites of the replicative Bacillus–E. coli shuttle vector pAD43-25 (Table 1), thereby releasing the gfpmut3A gene and placing mKate2mut expression under the control of the constitutive upp promoter from B. cereus UW85.
E. coli Top10 cells were transformed with the method as described by Sambrook et al. (Sambrook et al., 1989). From these, 20 randomly chosen colonies were grown separately in LB-Cm15 medium, and the mutation fre-quency of the FPs was estimated after plasmid isolation by double- stranded sequencing using the primer pairs pAD_for/pAD_rev for mKate2 and pNW33N_for/ pNW33N_rev for sfgfp(Sp) respectively (Table 1). Whole plas-mid libraries were generated as described previously (Frenzel et al., 2017). In brief, approximately 100,000 E. coli colonies were pooled after 24–30 h of growth at 37 °C from plates by resuspension in LB medium and the vector mixture was extracted with the JETSTAR Plasmid Purification Kit according to the manufacturer’s instruction (GENOMED, Löhne, Germany).
Preparation of competent B. mycoides cells, electroporation and library setup B. mycoides EC18 aliquots were prepared for electroporation according to a protocol previously established for B. cereus (Ehling-Schulz et al., 2005). Library vector DNA was added in an amount of 1–2 μg to the cells, and elec-troporation was performed applying settings of 2.0 kV, 25 μF and 200 Ω in a 2-mm cuvette using a Bio Rad Gen Pulser II electroporation system (Bio-Rad). After addition of 1 ml LB medium, cells were grown for 2 h at 30 °C and 150 rpm for recovery and then plated on LB-Cm4. After 16–24 h of growth at 30 °C, colonies were harvested from the plates and pooled in LB medium. The libraries were stored at −80 °C as 15% glycerol stocks.
Fluorescence-activated cell sorting of B. mycoides FP libraries B. mycoides EC18 sfGFP(Sp)mut or mKate2mut libraries were inoculated in 50 ml of LB-Cm4 and grown at pH 7.0 or pH 6.0 to an OD600 of 0.3–0.6, representing the exponential phase of growth. Since B. mycoides shows extensive cell-chaining, a mild sonication step of 4 rounds of 3 × 10 pulses of 1 s with an amplitude of 30% (Vibra CellTM, model VCX 130, Sonics and Materials, Newtown, CT, USA) was applied to disassemble the aggregated Ta
ble
1. S
trai
ns, p
lasm
ids a
nd p
rim
ers u
sed
in th
is st
udy.
Stra
ins
Rele
vant
char
acte
ristic
Refe
renc
eB.
myc
oide
s EC1
8W
ild ty
pe. I
sola
ted
from
pot
ato
endo
sphe
re.
Yi et
al.,
201
7 B.
subt
ilis H
S3W
ild ty
pe. I
sola
ted
from
gra
ss rh
izos
pher
e.Z.
Li. U
npub
lishe
dB.
am
ylol
iqef
acie
ns H
S9W
ild ty
pe. I
sola
ted
from
gra
ss rh
izos
pher
e.Z.
Li. U
npub
lishe
dB.
cere
us A
TCC
1098
7B.
cere
us ty
pe st
rain
Ra
sko
et a
l., 2
004
E. co
li TO
P10
F- m
crA
Δ(m
rr-h
sdRM
S-m
crBC
) φ80
lacZ
ΔM
15 Δ
lacX
74 re
cA1 a
raD1
39 Δ
(ara
-leu)
7697
gal
U g
alK
rpsL
endA
1 nup
GLa
bora
tory
stoc
k U
nive
rsity
of G
roni
ngen
Plas
mid
spK
B01-
sfG
FP(S
p)E.
coli-
Stre
ptoc
occu
s shu
ttle
vect
or e
ncod
ing s
uper
fold
er G
FP co
don-
optim
ized
for S
trept
ococ
cus p
neum
onia
e; A
pR ; Tet
R(O
verk
amp
et a
l., 2
013)
pNW
-Ppt
a-3T
ERE.
coli-
Baci
llus s
pp. s
hutt
le ve
ctor
for F
P lib
rary
cons
truc
tion.
Con
stitu
tive
Ppta
pro
mot
er fr
om P
arag
eoba
cillu
s the
rmo-
gluc
osid
asiu
s DSM
254
2 an
d th
ree-
fold
tran
scrip
tiona
l ter
min
ator
der
ived
from
pKB
01; r
eplic
ates
in B
acill
us st
rain
s w
ith p
BC1 o
rigin
Cm
R
(Fre
nzel
et a
l. 20
17)
pAD
651
E. co
li-Ba
cillu
s spp
. shu
ttle
vect
or e
ncod
ing
mKa
te2;
repl
icat
es in
Bac
illus
stra
ins w
ith p
TA10
60 o
rigin
; ApR ; C
mR
(Eijl
ande
r and
Kui
pers
, 201
3)pA
D43
-25
E. co
li-Ba
cillu
s spp
. shu
ttle
vect
or fo
r FP
libra
ry co
nstr
uctio
n.; c
onst
itutiv
e Pu
pp p
rom
oter
from
B. c
ereu
s UW
85; r
epli-
cate
s in
Baci
llus s
trai
ns w
ith p
TA10
60 o
rigin
ApR ; C
mR
(Dun
n an
d H
ande
lsm
an, 1
999)
pNW
-sfG
FP(S
618)
Der
ivat
ive
of p
NW
-Ppt
a-3T
ER e
ncod
ing
sfG
FP(S
618)
for o
ptim
ized
exp
ress
ion
in B
. myc
oide
sTh
is st
udy
pNW
-sfG
FP(S
709)
Der
ivat
ive
of p
NW
-Ppt
a-3T
ER e
ncod
ing
sfG
FP(S
709)
for o
ptim
ized
exp
ress
ion
in B
. myc
oide
sTh
is st
udy
pNW
-sfG
FP(S
PS6)
Der
ivat
ive
of p
NW
-Ppt
a-3T
ER e
ncod
ing
sfG
FP(S
PS6)
for o
ptim
ized
exp
ress
ion
in B
. myc
oide
sTh
is st
udy
pAD
-mKa
te(K
603)
Der
ivat
ive
of p
AD43
-25
enco
ding
mKa
te(K
603)
for o
ptim
ized
exp
ress
ion
in B
. myc
oide
sTh
is st
udy
pAD
-mKa
te(K
713)
Der
ivat
ive
of p
AD43
-25
enco
ding
mKa
te(K
713)
for o
ptim
ized
exp
ress
ion
in B
. myc
oide
sTh
is st
udy
pAD
-mKa
te(K
PS12
)D
eriv
ativ
e of
pAD
43-2
5 en
codi
ng m
Kate
(KPS
12) f
or o
ptim
ized
exp
ress
ion
in B
. myc
oide
sTh
is st
udy
PYB_
amyG
FPD
eriv
ativ
e of
PAT
∆S28
. Rep
licat
ive
in E
. col
i and
cond
ition
ally
repl
icat
ive
in B
acill
us sp
ecie
s. Ex
pres
sion
of t
he sf
GF-
P(SP
S6) g
ene
is d
riven
by t
he co
nstit
utiv
e Pp
ta p
rom
oter
from
P. th
erm
oglu
cosid
asiu
s DSM
254
2; S
pecR .
This
stud
y
pAD
-KPS
12-P
man
Der
ivat
ive
of p
AD-m
Kate
(KPS
12).
The
upp
prom
oter
was
repl
aced
by m
anno
se in
duci
ble
prom
oter
.Th
is st
udy
pAD
-mKa
te2-
Pman
Der
ivat
ive
of p
AD-m
Kate
2. T
he u
pp p
rom
oter
was
repl
aced
by m
anno
se in
duci
ble
prom
oter
.Th
is st
udy
pAD
-sfG
FP(S
p)-P
man
Der
ivat
ive
of p
AD-K
PS12
-Pm
an. T
he R
FP g
ene
was
repl
aced
by g
fp(S
p)Th
is st
udy
pAD
-SPS
6-Pm
anD
eriv
ativ
e of
pAD
-KPS
12-P
man
. The
RFP
gen
e w
as re
plac
ed b
y gfp
(SPS
6)Th
is st
udy
Prim
ers
pKBO
1der
Mut
_F
TCCT
CTAG
AACT
AATA
AGGA
GGA
CAAA
CATG
(Xba
I)(F
renz
el et
al.
2017
)pK
BO1d
erM
ut_R
CCTG
CATG
CCCT
TGAC
TAGT
GCT
CATT
ATTA
(Sph
I)(F
renz
el et
al.
2017
)pN
W33
N_f
orTC
GG
GATT
CGTT
TTAC
TTTC
C(F
renz
el et
al.
2017
)pN
W33
N_r
evCA
ATTT
CACA
CAG
GAAA
CAG
C(F
renz
el et
al.
2017
)m
Kate
2Mut
_FGA
CTCT
AGAG
GAAA
ATTA
ATGT
CAGA
ACTT
(Xba
I)Th
is st
udy
mKa
te2M
ut_R
ACTG
CATG
CCTA
CTCA
AGCT
TTTA
TTAT
TA (S
phI)
This
stud
ypA
D_fo
rAT
TGAA
GAAG
ACTG
CCGA
GACT
This
stud
ypA
D_re
vCG
AAG
CTCG
GCG
GATT
TGTC
This
stud
yam
yFGA
GCG
GTAC
CTCA
TTTT
GACG
GAAC
GGA
TTG
(Kpn
I)Th
is st
udy
amyR
GAG
CGAG
CTCT
TCAT
CACC
AATT
TACC
GCT
G (S
acI)
This
stud
yPm
anF
GAG
CGAA
TTCG
CAG
GGA
TTAT
TCCT
TGCT
TT (E
coRI
)Th
is st
udy
Pman
RGA
GCT
CTAG
ATGT
CTCA
ACTG
TATA
CCGA
AATC
(Xba
I)Th
is st
udy

55
88 89
Opt
imiz
ed fl
uore
scen
t pro
tein
s for
the
rhiz
osph
ere-
asso
ciat
ed
bact
eriu
m B
acill
us m
ycoi
des w
ith e
ndop
hytic
and
bio
cont
rol a
gent
pot
entia
l
MET
EARI
ALS
AN
D M
ETH
OD
S
cells. Cells were sorted on a BD FACS Aria II (BD Biosciences) at 20 psi using a 70 μM nozzle at a flow rate of 1.0 with the highest sort precision mode (0-32-0 sort purity mask). Cellular debris and chained cells were ex-cluded using a sequential gating strategy with FCS height versus widths, followed by SCC height versus width. For separation of the brightest vari-ants, a cut-off of 3% of the brightest event in the first round of cell sorting and 0.3% of the brightest events in the second round of sorting with the light scatter parameters (ex: 488 nm, em: 525/50 nm, 505 LP filter for GFP; and ex: 592 nm, em 620/30 nm, 600 nm LP filter for RFP) was chosen. See Supporting Information Figure S1 for a work-flow scheme.
In total, 20,000 cells were isolated by sorting and aliquots were plated on LB-Cm4 and grown 16–24 h at 30 °C, while the remaining cells (ca. 104 CFU) were inoculated into fresh LB-Cm4 and grown either at the same pH (6.0 or 7.0) as the first cultures, or the pH was ‘swapped’ to sort bright variants functional at both pH 6.0 and pH 7.0. Cultures were incubated for 16 h at 30 °C and 200 rpm until the following round of cell sorting.
Screening of FP variants and flow cytometry measurements After FACS sorting, the final fluids containing bright cells were plated on LB-Cm4 plates and grown overnight at 30 °C. The colonies were observed by Olympus MVX10 macro zoom fluorescence microscope equipped with a PreciseExcite light-emitting diode (LED) for fluorescence illumination. The filter setting for GFP was excitation at 460/480 nm and emission at 495/540 nm with a 485-nm dichromatic mirror; and for RFP the filter set-ting was excitation at 545/580 nm and emission at 610 nm with a 600 nm dichromatic mirror. Pictures were acquired with an Olympus XM10 mon-ochrome camera (Olympus Corporation, Tokyo, Japan). Twenty of the brightest colonies in each screening group (pH 6.0, pH 7.0 or pH shift) were re-streaked and the fluorescence of individual cells was assessed by FC. The fluorescence of selected GFP variants was quantitatively de-termined with a FACS-Canto flow cytometer (BD Biosciences) equipped with a 15 mW, 488 nm argon ion laser. All samples were grown in LB-Cm4 liquid medium, re-suspended in PBS and sonicated as described above to disperse cell-clumps prior to analysis. GFP emission was detected at 525/50 nm with an excitation of 488 nm. The RFP signals were measured in a FACS Aria II with excitation at 592 nm and emission at 620/30 nm. Per sample, 50,000 cells were analysed. Data acquisition and analysis was performed using the FACSDiva software (BD Biosciences) and the FCSalyzer software (version 0.9.13-alpha).
Electroporation of B. amyloliquefaciens, B. subtilis and B. cereus For B. amyloliquefaciens and B. subtilis, one single colony was inoculated into 50 ml LBSP medium (LB supplemented with 0.5 M sorbitol and 50 mM KH2PO4 and K2HPO4) and grown to an OD600 of 0.65. Cells were collected by centrifugation at 5,000 g, 4 °C for 10 min. The supernatant was discarded and the pellet was washed with cold electroporation buffer (10% glycerol with 0.25 M sorbitol) for four times. Finally, the cells were suspended in 1 ml electroporation buffer. Aliquots of 100 μl were frozen in liquid nitrogen and stored at −80 °C until the electroporation was per-formed. For B. cereus, electro-competent cells were prepared as described before (Ehling-Schulz et al., 2005). For all Bacillus strains, the electropora-tion was performed as described for B. mycoides.
Strain construction for double-FP-labelling of B. mycoides The plasmid PYB was generated by replacing the pAMβ1 replication or-igin (ori) of PATDS28 (Namy et al., 1999) with a temperature sensitive ori from the PAW068 plasmid for Gram-positive bacteria (Wilson et al., 2007). Then a 1 kb-fragment of the α-amylase gene was amplified from the genome of B. mycoides EC18 with the primers amyF and amyR. This fragment was further digested with the KpnI and SacI enzymes and in-serted into the PYB plasmid at the same restriction site to give rise to the plasmid PYB_amy. The sfgfp(SPS6) gene together with the Ppta pro-moter was inserted into PYB_amy at the restriction sites EcoRI and Hin-dIII, which resulted in the plasmid PYB_amyGFP. This plasmid was then transformed into B. mycoides EC18 and plated on LB plates with 100 μg/ml spectinomycin. One colony was picked and grown in BHI liquid medium with 100 μg/ml spectinomycin over night at 30 °C. The culture was then diluted 100× with the same medium and grown at 37 °C to block the rep-lication of the plasmid. A serial dilution of the culture was plated on BHI-Spec100 agar and cultured at 37 °C overnight. The colonies were checked by PCR for successful single cross-over recombination. The EC18 strain carrying the chromosomally integrated sfGFP(SPS6) reporter was used to make electrocompetent cells and the plasmid pAD-mKate(KPS12) was transformed into the strain. The double-labelled cells were selected on LB-Cm4/Spec100 agar grown at 30 °C. Presence of the FP reporters was verified by doublestranded sequencing of the PCR products.
Strain construction for FP expression from mannose-inducible promoter A fragment of the promoter PmanP that is positively regulated by man-nose (Wenzel et al., 2011) was cloned from the plasmid pJOE8999 ( Altenbuchner, 2016) using the primers PmanF and PmanR. The plasmid

55
90 91
Opt
imiz
ed fl
uore
scen
t pro
tein
s for
the
rhiz
osph
ere-
asso
ciat
ed
bact
eriu
m B
acill
us m
ycoi
des w
ith e
ndop
hytic
and
bio
cont
rol a
gent
pot
entia
l
MET
EARI
ALS
AN
D M
ETH
OD
S
pAD-mKate(KPS12) was cleaved with EcoRI and XbaI, and the vector backbone was ligated with the PmanP fragment cleaved by the same en-zymes to give rise to pAD-KPS12-Pman. To construct the mannose con-trolled GFP vector, the gfp (SPS6) gene was cut with XbaI and SphI from pNW- sfGFP(SPS6) and then inserted into pAD-KPS12-Pman at the same restriction site to give the new plasmid pAD-SPS6-Pman. The mannose- inducible FP strain was obtained by transforming the final plasmid into the B. mycoides EC18 strain. The overnight culture of each strain was di-luted 50 times with LB-CM4 with different concentrations of mannose. After around 6 hours of growth, fluorescence signals of the strains were measured by FC and images were captures with fluorescence microscopy.
Fluorescence microscopy Single cell observation was performed with an Olympus IX71 micro-scope (Personal DV, Applied Precision; assembled by Imsol, Preston, UK) equipped with a Nomarski DIC and a 100 W mercury vapor lamp for FP excitation. A 10× eyepiece and a 100× phase-contrast objective was used to examine the cells. GFP variants were detected with excita-tion at 465–495 nm, 505 nm dichroic mirror and emission at 515–555 nm. RFP variants were detected with an excitation at 528–553 nm, 565 nm di-chroic mirror and emission at 590–650 nm. Images were captured with a CoolSNAP HQ2 camera (Princeton Instruments, Trenton, NJ, USA). The intensity of single cell was calculated with the ImageJ software ( https://imagej.nih.gov/ij/). The region of cells in which the fluorescence signal was quantified was selected manually. The total cell fluorescence was calculated by the following formula: corrected total cell fluorescence (CTCF) = Integrated Density – (Area of selected cell × Mean fluorescence of background readings) (Pozniak et al., 2016). At least 500 cells from three independent biological replicates were analyzed.
Growth curves of FP-labelled B. mycoides strains and plasmid stability assay B. mycoides strains transformed with the different FP variants were tested for their growth pattern and plasmid stability. The growth curve was de-termined by plotting the optical density values (OD600) in LB liquid me-dium versus time. For the plasmid stability assay, each strain was grown to stationary phase in LB medium at 25 °C (the same as plant culturing temperature) with 200 rpm aeration and then diluted by 50× in LB. The diluted culture was continued growing to stationary with the same condi-tions. Two more cycles of subsequent dilution were performed in the next two days. At day 3, the culture was serially diluted and plated on LB agar plates with or without chloramphenicol and the CFU/ml was calculated.
In situ observation of FP labelled strains by confocal laser scanning microscopy Chinese cabbage seeds were surface sterilized in 70% ethanol for 2 min, followed by a bath in 3% sodium hypochlorite for 2 min. After the ster-ilization treatment, seeds were washed four times in sterile deionized water. The excessive water on the seeds surface was removed with auto-claved filter paper. The seeds were inoculated into Petri dishes contain-ing 25% Hoagland solution (Hoagland and Arnon, 1950) solidified with 1% agar and incubated for germination and growth in a culture room at (25 ± 2 °C) with a 12-h photoperiod for six days. The seedlings were then transferred to 3-L hydroponic trays containing 25% Hoagland’s solution and continued to grow for 2 days. Hoagland solution was aerated using air stones connected to an aquarium air pump. The B. mycoides strains transformed with the different FP variants were grown to the exponential growth phase, and then 10 ml culture was collected and re-suspended in 25% Hoagland’s solution. The hydroponic system was inoculated with a final concentration of 2 × 104 CFU/ml B. mycoides cells.
After 2–3 days of inoculation, the colonization of B. mycoides on the roots of the cabbage seedlings was assessed using a ZEISS LSM 800 CLSM (Carl Zeiss, Germany) equipped with diode lasers and GaAsP detector. Im-ages for fluorescent light channels were taken simultaneously with im-ages of the bright field channel. To achieve the maximum brightness of each FP and low background auto-fluorescence of the plant tissue, the settings of the confocal microscope were adjusted as follows: For GFP ob-servation, 0.2% power of the 488 nm laser line was used for excitation and 509–546 nm was set as emission wavelength. For RFP detection, 1% power of the 561 nm laser line was used as excitation wavelength and 600–680 nm was set as emission wavelength. The pinhole size for GFP was 25 mm and for RFP was 30 μm, pixel scanning time was 2.06 μs and line scanning time was 2.47 ms with a line averaging of 2.
RESULTS AND DISCUSSION
Construction of GFP and RFP libraries and isolation of bright variants by B. mycoides cell sorting FPs are indispensable tools for molecular biology and microbial ecology. However, FPs are not equally well expressed in different bacterial species, presumably due to the codon-usage bias which affects the translation, folding and maturation efficiency of the proteins. Previous studies proved that different GFP variants display strongly variable fluorescence intensi-ties in low-GC Gram-positive organisms (Overkamp et al., 2013).

55
92 93
Opt
imiz
ed fl
uore
scen
t pro
tein
s for
the
rhiz
osph
ere-
asso
ciat
ed
bact
eriu
m B
acill
us m
ycoi
des w
ith e
ndop
hytic
and
bio
cont
rol a
gent
pot
entia
l
Resu
lts a
nd d
iscu
ssio
n
Our preliminary experiments showed that sfGFP(Sp), a robust, fast- folding and fast-maturing ‘superfolder’ GFP (Pedelacq et al., 2006; Overkamp et al., 2013) was functionally expressed in B. mycoides. However, the signal was too weak for studying bacteria-plant interactions and cell tracking when sfGFP(Sp) expression was driven by weak promoters (data not shown). To track different bacterial strains at the same time, or to simultaneously follow distinct promoter activities within one cell, it would be desirable that multiple fluorescent markers are well expressed and detectable in the same target organism. Since mKate2 has a fluorescence spectrum that substantially differs from GFP, with an excitation maximum of 588 nm and an emission maximum of 633 nm (Shemiakina et al., 2012), it is highly suitable for co-labelling experiments with GFP. However, initial bench-marking experiments with the red fluorescent protein variants mCherry and mKate2 cloned on multicopy plasmids revealed that the signal inten-sity of mKate2 was very low and close to the level of autofluorescence of B. mycoides cells. Moreover, the expression of mCherry could neither be detected by flow cytometry (FC) nor by fluorescence microscopy (data not shown). This is in line with the observation that mKate2 was better suited for promoter labelling studies than mCherry in the closely related species B. cereus (Eijlander and Kuipers, 2013).
By applying a random mutagenesis approach, we obtained a sfGFP li-brary that was cloned into the replicative E. coli-Bacillus shuttle plasmid pNW33N, resulting in a total of 115,000 E. coli TOP10 clones. The plas-mids carrying the mutated sfGFP(Sp) gene were isolated and transformed into B. mycoides EC18 by electroporation, resulting in a library size of 44,000 clones with a mutational frequency of one to four nucleotides per sfGFP(Sp) gene. However, mKate2 showed no fluorescence when being expressed under the control of the same pta promoter as sfGFP (data not shown). As a result, the mKate2 mutation library was constructed with the replicative plasmid pAD43-25, which carries the comparably stronger upp promoter. The plasmid library was transformed into E. coli TOP10, thereby yielding a size of 115,200 colonies. The transformation of the li-brary into B. mycoides EC18 resulted in a 43,820 clone-sized library with a mutation rate of one to three nucleotides per mKate2 gene.
To isolate the brightest sfGFP(Sp) and mKate2 mutants, the B. my-coides libraries were grown planktonically until the exponential growth phase was reached. We considered that the bacteria encounter environ-ments with different pH during the establishment of the endophytic life-style, since cabbage-related plants such as Arabodopsis thaliana induce a soil acidification in the rhizosphere region (for a recent publication, see Barbez et al., 2017). Therefore, the B. mycoides libraries were grown in three groups with different pH conditions: pH 6.0, pH 7.0 and pH shift
condition. For the latter, brightest cells were first enriched at pH 6.0, and then subcultured at pH 7.0 followed by a second round of FACS enrich-ment. As shown in the Supporting Information Figure S1, around 0.3% of
Figure 1. Fluorescence quantification of sfGFP variants in B. mycoides. (A) B. mycoides EC18 carrying pNW33N-Ppta-3TER plasmids with the respective fluorescent protein variants were grown overnight in LB and subjected to analysis by FC. (B) Single-cell flu-orescence was measured with fluorescence microscopy. The fluorescence intensities are normalized for background fluorescence and cell area (see Material and Methods). Mean fluorescence was calculated from at least 500 cells. Error bars indicate the stan-dard errors of the means obtained from three independent biological and technical rep-licates. Asterisks denote significant differences between the mean fluorescence inten-sity of sfGFP and the respective mutant variants (p<0.05).

55
94 95
Opt
imiz
ed fl
uore
scen
t pro
tein
s for
the
rhiz
osph
ere-
asso
ciat
ed
bact
eriu
m B
acill
us m
ycoi
des w
ith e
ndop
hytic
and
bio
cont
rol a
gent
pot
entia
l
Resu
lts a
nd d
iscu
ssio
n
the mildly sonicated cell population was sorted after a first FACS enrich-ment step from all pH conditions. After spreading and incubation at 30 °C on LB-Cm4 agar, 20 of the brightest colonies were selected by visual ap-pearance with a Olympus MVX10 macro zoom fluorescence microscope (Supporting Information Figure S1). To obtain pure colonies arising from single clones, the 20 preselected colonies from each condition were re-streaked twice on LB-Cm4. After quantification of signal intensities and the amplitude of fluorescence signals at the single-cell level from expo-nential phase cultures by FC, we selected three of the best performing GFP and RFP variants with high brightness and small fluorescence signal devi-ations for further analyses.
Improvement of GFP signal intensities The mean fluorescence intensity (MFI) of the sfGFP variants selected un-der pH 6.0, pH 7.0 and pH shift conditions (termed S618, S709 and SPS6) was measured by FC. As shown in Figure 1A, the MFI of all selected sf-GFP variants was increased by at least 50% in comparison to the orig-inal sfGFP reporter in B. mycoides. The variant selected under pH shift conditions, sfGFP(SPS6), exhibited the strongest mean fluorescence sig-nal. Signal intensities of single cells stemming from a colony grown on solid medium was examined by fluorescence microscopy (Figure 1B). In general, the differences in the average fluorescence levels analysed from microscopy images correlated well with the results obtained by FC. The highest improvement with regard to the mean brightness level was ob-served for the optimized sfGFP(SPS6) protein when compared to the orig-inal sfGFP(Sp) protein.
Although the signal intensity of all sfGFP variants was not evenly distributed within single cells, obvious differences in the mean bright-ness levels between the optimized and the original sfGFP were observed, clearly showing an improved detectability by visualization methods (Fig-ure 2A and B). Variation of the signals could be related to an uneven dis-tribution of FP proteins and/or differences in the plasmid copy numbers in the daughter cells, as it was previously discussed for the closely related bacterium B. cereus ( Eijlander and Kuipers, 2013). However, with a few ex-ceptions, the mechanism of cell division and separation (Di Franco et al., 2000; Turchi et al., 2012) and the possibility that the extensive cell chain-ing might be connected to multicellular cooperation (Shapiro, 1998), such as the exchange of DNA, nutrients or signals between the B. mycoides cells, has not been studied so far. We thus tested the possibility that the consti-tutive expression of the FPs poses a metabolic burden to the cells and that separating daughter cells might undergo a loss of the replicative plasmid. Comparison of the growth behaviour between B. mycoides wild type cells
and cells carrying the FP expression plasmids did not reveal any growth retardation or growth defects from early logarithmic to the late station-ary phase (Supporting Information Figure S2A). In addition, fluorescence imaging of complex B. mycoides colonies showed that the expression of sfGFP generally did not affect the development of colonies (Figure 2B). Due to the compactness and multilayered rhizoid growth, however, the difference in brightness between the optimized sfGFP and the original
Figure 2. (A) Visualization of sfGFP variant expression in planktonically grown, expo-nential-state cells of B. mycoides by fluorescence microscopy. For comparison of the sfGFP fluorescence intensities, the same imaging conditions were applied (ex: 465–495 nm, em: 515–555 nm; exposure: 0.15 s with 32% excitation xenon light (300 W); 100× phase-con-trast objective). The white bar represents 5 μm. (B) Comparison of fluorescence signal in-tensities of sfGFP variants in B. mycoides colonies. LB-Cm4 plates were spot-inoculated with equal amounts of B. mycoides cells and incubated for 18 h at 30 °C. Images were acquired with a microscope using the same imaging conditions (ex: 460/480 nm, em: 495/540 nm, 50% of excitation light, exposure time: 100 ms). The white scale bar represents 0.5 cm. Representative images from three independent biological replicates are shown.

55
96 97
Opt
imiz
ed fl
uore
scen
t pro
tein
s for
the
rhiz
osph
ere-
asso
ciat
ed
bact
eriu
m B
acill
us m
ycoi
des w
ith e
ndop
hytic
and
bio
cont
rol a
gent
pot
entia
l
Resu
lts a
nd d
iscu
ssio
n
sfGFP(Sp) variants was not as apparent as compared to the measurements of cells grown under planktonic conditions (Figure 2B). Tracing the plas-mid presence and inheritance without antibiotic pressure by sequential propagation over several days (approximately 150 generations) showed no indication of significant plasmid loss (Figure S2B). This indicates that the plasmid itself is stably inherited and that the reason for signal inten-sity variation between single cells is more complex and needs to be ad-dressed in greater detail in a separate study.
Improvement of RFP signal intensities Determination of the MFI of the in vivo selected RFP variants revealed that the signal intensities of the mutants K603 and K713 selected under pH 6.0 and pH 7.0 conditions was increased 7- and 6-fold in comparison to the original mKate2 protein (Piatkevich et al., 2010) respectively, while mKate2(KPS12), which was selected under pH shift conditions, showed a 10-fold improvement of fluorescence (Figure 3A). This was further corrob-orated by the quantification of fluorescence signals emitted from single cells by fluorescence microscopy showing that KPS12 was the best per-forming mKate2 variant in B. mycoides (Figure 3B). Cells carrying the orig-inal mKate2 emitted a very weak fluorescence signal, which was barely above the autofluorescence of B. mycoides cells at 528–553 nm excitation. In contrast, K603, K713 and KPS12 showed a significant improvement of fluorescence signal emission (Figure 4A). Colony imaging revealed that the signal-to-noise ratio was significantly improved for all three variants as compared to mKate2, resulting in clearly detectable colonies on solid growth media. Moreover, in the complex B. mycoides colonies, in which cells are less well aerated than in shaken planktonic cultures, KPS12 still gave the highest signals among all examined variants (Figure 4B).
As observed for the GFP variants, cells showed a variation in the in-tensity of fluorescence signals when expressing the RFP proteins, which was not caused by growth retardation effects or by a loss of the replicative plasmid encoding the RFP (Supporting Information Figure S2A and B). The mechanism causing these phenotypic differences thus needs to be addressed in a separate study.
Co-expression of optimized GFP and RFP variants in B. mycoides Based on the results obtained from protein optimization, we next tested the suitability of the best performing variants for dual-labelling and si-multaneous visualization in the B. mycoides background. To additionally address the question whether the signal variation could be reduced, we first integrated the sfGFP(SPS6) gene driven by the constitutive Ppta pro-moter into the chromosome of B. mycoides EC18 at the α-amylase gene
locus. In a next step, the pAD-mKate(KPS12) plasmid was transformed into the single-copy reporter strain to obtain the co-expression strain. The GFP and RFP signals were simultaneously detectable by FC and by fluores-cence microscopy and were clearly distinguishable from each other under
Figure 3. Fluorescence quantification of mKate2 variants in B. mycoides. (A) B. mycoides EC18 carrying pAD43-25 plasmid derivatives with the respective fluorescent protein variants were grown overnight in LB and subjected to analysis by FC. (B) Single-cell fluorescence was measured by fluorescence microscopy. Fluorescence intensities are normalized for background fluorescence and cell area and the mean fluorescence was calculated from at least 500 cells. Error bars indicate the standard errors of the means (n53). Asterisks denote significant differences between the mean fluorescence intensity of mKate2 and the respective mutant variants (p<0.05).

55
98 99
Opt
imiz
ed fl
uore
scen
t pro
tein
s for
the
rhiz
osph
ere-
asso
ciat
ed
bact
eriu
m B
acill
us m
ycoi
des w
ith e
ndop
hytic
and
bio
cont
rol a
gent
pot
entia
l
Resu
lts a
nd d
iscu
ssio
n
the given differential excitation and detection conditions, showing that a cross-talk caused by a spectral overlap between the FPs is negligible (Fig-ure 5). This indicates that neither the excitation nor the emission spectra are significantly changed from the original protein variants, which have been previously shown to be compatible in multicolour imaging studies in Streptococcus pneumonia (Kjos et al., 2015). To our knowledge, this is the
first report of a successful dual-FP-labelling approach in bacilli of the B. cereus sensu lato group. Notably, the GFP signal intensity distribution was more homogeneous within the cells due to the presence of a chromosom-ally integrated single copy of the gfp gene (Figure 5D). This in turn strongly indicates that the differences in plasmids copy numbers among daughter cells within a colony substantially impact the signal amplitude per cell.
FP expression from a mannose-inducible promoter For plant-interaction studies, it would further be desirable to confirm the expression of the optimized FP variants when driven by weak or condition- dependent promoters. This is currently hampered by the lack of systematically characterized promoter regions in B. mycoides. We ini-tially recognized that a mannose-inducible promoter (Pman) from B. sub-tilis (Altenbuchner, 2016) is ‘leaky’ in B. mycoides, thereby conferring a basal, low level of FP expression. To compare the performance of the
Figure 4. (A) Visualization of mKate2 variant expression in planktonically grown, exponential-state cells of B. mycoides by fluorescence microscopy. For comparison of the mKate2 fluorescence intensities, the same imaging conditions were applied (ex: 528/553 nm, em: 590/650 nm; exposure: 0.45 s with 50% excitation xenon light (300 W); 100× phase-contrast objective). The white bar represents 5 μm. (B) Com-parison of fluorescence signal intensities of mKate2 variants in B. mycoides colonies. LB-Cm4 plates were spot-inoculated with equal amounts of sporulated B. mycoides cells and incubated for 18 h at 30 °C. Images were acquired with a microscope using the same imaging conditions (ex: 545/580 nm, em: 610 nm, 100% of excitation light, exposure time: 3.5 s). The white scale bar represents 0.5 cm. Representative images from three independent biological replicates are shown.
Figure 5. Co-expression of optimized FPs in B. mycoides EC18. The strain was double-la-belled by chromosomal integration of a single copy of sfGFP(SPS6) into the amyE lo-cus and electroporation of the replicative plasmid pAD-mKate2(KPS12) into the reporter strain. (A and B) FC measurements of GFP and RFP channels. (C–E) Microscopic ob-servation from phase-contrast, GFP (ex: 465–495 nm, em: 515–555 nm; exposure: 2.62 s with 32% excitation xenon light), and RFP (ex: 528/553 nm, em: 590/650 nm; exposure: 0.637 s with 50% excitation xenon light) channels.

55
100 101
Opt
imiz
ed fl
uore
scen
t pro
tein
s for
the
rhiz
osph
ere-
asso
ciat
ed
bact
eriu
m B
acill
us m
ycoi
des w
ith e
ndop
hytic
and
bio
cont
rol a
gent
pot
entia
l
Resu
lts a
nd d
iscu
ssio
n
optimized and original FPs under the control of Pman, fusions were con-structed based on the replicative plasmid pAD (Table 1) and transformed into B. mycoides EC18. FC measurements revealed that both optimized variants sfGFP(SPS6) and mKate(KPS12) were significantly better detect-able than the parental FP versions (Supporting Information Figure S3). The MFI stemming from the optimized variants increased with increas-ing mannose concentrations, whereas no significant MFI increase could be observed for the original proteins. This shows that that the improved variants are better suited for the detection when fused to weak promoters, because the parental FPs are only expressed at levels close to the autoflu-orescence of B. mycoides cells under these conditions.
Expression of novel FPs in other rhizosphere-associated Bacillus strains Although this study mainly focused on the development of optimized FP variants to allow the tracing of root-associated and endophytic B. mycoides strains, we next tested the performance of the improved FPs in addtional Bacillus species that mainly thrive in the soil. The sfGFP variant SPS6 was transformed into B. cereus ATCC 10987, B. subtilis HS3 and B. amyloliquefa-ciens HS9. The strains HS3 and HS9 were isolated from grass rhizosphere and are potentially PGP-promoting (unpublished data). In comparison to the original sfGFP(Sp) protein, sfGFP(SPS6) showed a 3- to 5-fold, signif-icantly improved fluorescence intensity in all three strains, thereby fa-cilitating their detection by fluorescence microscopy (Figure 6A–C). The improvement of brightness of mKate(KPS12) is shown in Figure 6D–F. While expression of the original mKate2 variant was barely measurable in B. cereus ATCC 10987, as stated earlier (Eijlander and Kuipers, 2013), mKate(KSP12) showed a fivefold improved brightness and was detectable by both fluorescence microscope and FC (Figure 6D). A significant increase in brightness was also observed in the rhizosphere-derived B. subtilis HS3 and B. amyloliquefaciens HS9 hosts (Figure 6E and F). Altogether, the im-proved variants were well expressed in the species tested and considerably facilitated the detection of Bacillus strains by live-cell imaging methods.
In situ performance of improved FPs to localize B. mycoides during establishment of an endophytic lifestyle In the rhizosphere, bacteria-plant interactions play an important role in maintaining plant health. The possibility of visualizing these interactions in situ is a key step for understanding the ecophysiology and basic biology underlying the beneficial processes (Larrainzar et al., 2005). Germaine et al. (2004) studied the endophytic behaviour of three Pseudomonas spe-cies by tracing the GFP-labelled cells during colonization of poplar trees.
Figu
re 6
. Co
mpa
riso
n of
the
per
-fo
rman
ce o
f th
e or
igin
al a
nd o
pti-
miz
ed F
P va
rian
ts in
add
ition
al so
il an
d rh
izos
pher
e-de
rive
d Ba
cillu
s sp
ecie
s. T
he s
fGFP
imag
ing
cond
i-tio
ns w
ere
the
sam
e fo
r eac
h st
rain
(e
x: 4
65–4
95 n
m,
em:
515–
555
nm
with
32%
xen
on l
ight
exc
itatio
n),
but
diffe
rent
exp
osur
e tim
es w
ere
appl
ied.
Exp
osur
e tim
e (A
) B. c
ereu
s AT
CC 1
0987
is 0
.4 s;
(B
) B.
sub
tilis
H
S3 is
0.0
85 s
and
(C)
B. a
myl
oliq
-ue
faci
ens
HS9
is
0.05
0 s.
The
im
-ag
ing
cond
ition
s fo
r m
Kate
2 va
ri-
ants
wer
e th
e sa
me
for
each
str
ain
(ex:
528
/553
nm
, em
: 59
0/65
0 nm
w
ith 5
0% e
xcita
tion
xeno
n lig
ht),
but
diffe
rent
exp
osur
e tim
e w
ere
appl
ied.
The
exp
osur
e tim
e fo
r: (A
) B.
cer
eus
ATCC
109
87 is
30
s; (B
) B.
subt
ilis
HS3
is 0
.33
s an
d (C
) B. a
m-
ylol
ique
faci
ens
HS9
is 3
.75
s. R
epre
-se
ntat
ive
imag
es a
re s
how
n. T
he
whi
te b
ar re
pres
ents
5 μ
m.

55
102 103
Opt
imiz
ed fl
uore
scen
t pro
tein
s for
the
rhiz
osph
ere-
asso
ciat
ed
bact
eriu
m B
acill
us m
ycoi
des w
ith e
ndop
hytic
and
bio
cont
rol a
gent
pot
entia
l
Resu
lts a
nd d
iscu
ssio
n
Bloemberg et al. (2000) labelled P. fluorescens with the enhanced cyan FP, enhanced green FP, enhanced yellow FP and the DsRed RFP reporter pro-tein. After inoculation of tomato plant seedlings, mixed microcolonies as well as single populations could be simultaneously visualized, which re-vealed a dynamic behaviour of localizing to sites on the roots and in the root/soil interface.
To finally demonstrate the applicability of the in vivo selected GFP and RFP variants for in planta studies, the endophytic B. mycoides isolate EC18 expressing the different FP proteins from constitutive promoters was in-oculated on Chinese cabbage (Brassica rapa) roots in a hydroponic system. At day 2 and day 3 after inoculation, roots were sampled and analysed con-cerning fluorescence signals and in planta localization of B. mycoides by
confocal microscopy. For GFP, both the original sfGFP(Sp) (Figure 7A) and the variant sfGFP(SPS6) (Figure 7B and C) provided a well-detectable flu-orescence signal at very low excitation strength, which prevented the oc-currence of an autofluorescence background from the root cells. In con-trast to sfGFP(Sp), cells tagged with the SPS6 mutant were more readily detectable; indicating that the higher brightness observed in fluorescence microscopy and FC experiments is also advantageous for confocal micros-copy applications. At 2 days post inoculation (DPI), B. mycoides initiated the attachment to the epidermis of the primary roots (Figure 7B). After 3 DPI, the bacteria started to aggregate as microcolonies on the surface, especially at the emerging site of root hairs. Additionally, a few chaining cells that translocated into the endosphere were observed (Figure 7C). In-terestingly, junctions of primary and lateral roots seem to be a preferred niche for microcolony establishment of B. amyloliquefaciens FZB42 (Fan et al., 2011), which might indicate that these are preferred sites for feeding on nutrients or endophytic entry into the plant.
Similar bacteria-plant interaction patterns were observed when B. my-coides was labelled with the red spectrum reporter protein variant mKate2 (Figure 7D) and mKate2(KPS12) (Figure 7E and F). Due to the inherently low fluorescence intensity of the original mKate2 protein during expression in B. mycoides, the fluorescent signal is very weak and can hardly be detected (Figure 7D). Enhancement of the excitation power and digit gain settings also deteriorated the signal-to-noise ratio and thus induced a high auto-fluorescence background of the plant tissue (data not shown). In compari-son, the enhanced fluorescence intensity of the mutant KPS12 allowed the detection of B. mycoides above the autofluorescence of the root hairs with-out extensive adjustment of the excitation parameters. Two days after inoculation, B. mycoides cells were attached to root hair cells (Figure 7E). These interactions were shown to play a key role in the endophytic col-onization of olive plant roots by Pseudomonas species (Prieto et al., 2011). Three days after inoculation, a higher number of cells were aggregated on the root epidermis and some cells were growing in the endosphere of the root hair as well as in the main root. A massive amount of B. mycoides cells colonized the elongation region of the root hair, which might represent an entrance point for B. mycoides to establish an endophytic lifestyle (Fig-ure 7F). Similarly, Ji and colleagues (2008) observed that the endophytic B. subtilis strain Lu144 enters into mulberry seedlings through the cracks formed at the lateral root junctions and the zone of differentiation and elongation. We speculate that the junctions of root hairs and main roots are the preferred and specific colonization sites for endophytic Bacillus strains, since for Gram-negative bacteria such as Pseudomonas, coloni-zation was more evenly distributed and observed in older basal root parts
Figure 7. In planta observation of life B. mycoides EC18 cells in the rhizosphere of Chi-nese cabbage. Labelling of B. mycoides with a set of in vivo-selected GFP and RFP variants allows in situ tracking of cabbage root colonization in a hydroponic system. (A) Epider-mal colonization two DPI is visualized with the original sfGFP(Sp) reporter protein. (B) Cells labelled with the improved sfGFP(SPS6) variant aggregate on the root surface two DPI. (C) Cells labelled with the improved sfGFP(SPS6) interact with the root hair form-ing small microcolonies and establish endophytic colonization three DPI. (D) Tracking of B. mycoides cells labelled with the original mKate2 reporter is aggravated due to a low mKate2 brightness and high autofluorescence background of the plant tissue. (E) Detec-tion of B. mycoides expressing the improved mKate2 (KPS12) reporter protein reveals the interaction of cells with root hairs two DPI and the entry into the endophytic coloniza-tion lifestyle three DPI (F). The scale bar represents 10 μm.

55
104 105
Opt
imiz
ed fl
uore
scen
t pro
tein
s for
the
rhiz
osph
ere-
asso
ciat
ed
bact
eriu
m B
acill
us m
ycoi
des w
ith e
ndop
hytic
and
bio
cont
rol a
gent
pot
entia
l
Resu
lts a
nd d
iscu
ssio
n
or the root hair of barley (Buddrus-Schiemann et al., 2010). This might be linked to differences in chemoattractants and preferred metabolites asso-ciated with the different root regions and cell types, as has been indicated earlier (Brimecombe et al., 2007). However, the detailed mechanism of endophytic plant colonization by B. mycoides as well as a systematic com-parison to tackle differences in the colonization mechanisms between Gram-negative and Gram-positive bacteria needs further thorough inves-tigation.
Sequence analysis of in vivo selected FPs optimized for in planta studies Early attempts to optimize the heterologous expression of GFP revealed that the fluorescence properties can be modulated by mutations within the fluorophore region, resulting in altered excitation and emission spec-tra (Ehrig et al., 1995). The S65T substitution leads, for instance, to GFP de-rivatives with a red-shifted excitation maximum and strongly increased fluorescence (Heim et al., 1995; Chiu et al., 1996). Since the wild-type GFP is prone to misfolding and aggregation, which causes reduced chromo-phore maturation and low yields, a variety of studies aimed at improving the folding properties of GFP and other FPs (Hsu et al., 2009). The FP vari-ant Emerald contains the S65T and F64L mutations featured in enhanced GFP (eGFP), and has four additional point mutations that improve the effi-ciency of maturation and folding at 37 °C, and increase the intrinsic bright-ness (Day and Davidson, 2009). Another approach to obtain improved FP variants is the adaptation of FP genes to the typical codon usage of the host organism, which in some cases could improve the translation efficiency, resulting in higher FP expression and thus fluorescence signals (Sastalla et al., 2009; Leroch et al., 2011). Such codon optimized FPs have been de-veloped for the cyan fluorescent protein and a yellow fluorescent protein in B. anthracis (Sastalla et al., 2009), for GFP and RFP in Botrytis cinerea (Leroch et al., 2011) and for GFP in Zymoseptoria tritici (Kilaru et al., 2015).
To analyse the changes associated with the improved functionality of the FP variants that were in vivo isolated from B. mycoides, the nucleo-tide mutations, amino acid exchanges as well as codon usage frequencies were compared between the mutated and the originating genes (Table 2). In comparison to the original sfGFP(Sp), the variant sfGFP(S618) carries the exchanges K156E and V176I. These two sites are solvent-exposed and located at the α-helix region between two β-strands (Supporting Informa-tion Figure S4A). Pedelacq and colleagues (2006) reported that mutations at these flexible linker positions (e.g., Y145F and Y171V) are likely to elim-inate aggregation-prone or off-pathway folded proteins from the folding trajectory. The variant sfGFP(S709) contains the three mutations T59I,
P192P and Q204H; with the silent mutation P192P being distant from the chromophore, while the mutation T59I was close to the chromophore and buried in the centre of the β-barrel (Supporting Information Figure S4B). The mutation Q204H resides in the 10th β-strand closely located to the chromophore. The β-strand mutations F99S/M153T/V163A in GFPuv/cycle 3 variant were shown to change the surface hydrophobicity and, therefore, the aggregation propensity of the protein (Fukuda et al., 2000). The best performing GFP variant in this study, SPS6, contains the silent mutation A179A and a N39D exchange. The mutation Y39N was located between the 2nd and 3rd β-strand (Supporting Information Figure S4C) and was reported to increase folding rates and stability in sfGFP (Pedelacq et al., 2006). The substitution of asparagine to aspartic acid may further improve these effects.
The three in vivo-selected, optimized mKate2 mutants contained differ-ent nucleotide substitutions, which were either solely silent mutations [N186N in the case of mKate(K603)], solely amino acid exchanges [G152D in mKate(K713)] or a combination of both (K185K, R220R and D206Y in the brightest variant mKate(KPS12) obtained from pH shift experiments). The mutation sites are indicated on the 3D crystallographic structure of mKate2 in Supporting Information Figure S4D–F. Since the RFPs are gen-erally less well characterized than GFPs with regards to folding/unfold-ing kinetics and crystallization studies, the impact of these mutations on the improvement of fluorescence signal intensity and brightness or the
Table 2. Overview of mutations of sfGFP and mKate2 variants optimized for expression and in planta localization of B. mycoides.
FP variant
Nucle-otide posi-tiona)
Nucleo-tide ex-change
Codonmutation
Amino acid
positiona)
Amino acid
mutation
Codon usage frequency of
original amino acidb)
Codon usage frequency of introduced
amino acidb)
S618 466 A->G AAA->GAA 156 K156E 6.14 5.73526 G->A GTT->ATT 176 V176I 3.12 5.17
S709 176 C->T ACT->ATT 59 T59I 1.24 5.01576 A->G CCA->CCG 192 P192P 1.64 0.76612 A->T CAA->CAT 204 Q204H 3.03 1.63
SPS6 115 A->G AAC->GAC 39 N39D 1.08 1.45537 T->C GCT->GCC 179 A179A 2.98 1.21
K603 558 T->C AAT->AAC 186 N186N 2.04 1.77K713 455 G->A GGT->GAT 152 G152D 1.12 3.07KPS12 555 G->A AAG->AAA 185 K185K 1.93 5.1
616 G->T GAT->TAT 206 D206Y 3.07 2.1660 T->G CGT->CGG 220 R220R 0.69 0.99
a) relative distance from translation start of FPb) According to codon usage frequency table for B. mycoides EC18 (accessible at the Ge-nome 2D webserver http://server.molgenrug.nl/index.php)

55
106 107
Opt
imiz
ed fl
uore
scen
t pro
tein
s for
the
rhiz
osph
ere-
asso
ciat
ed
bact
eriu
m B
acill
us m
ycoi
des w
ith e
ndop
hytic
and
bio
cont
rol a
gent
pot
entia
l
Conc
lusi
on
folding and maturation efficacy is not readily explained. Although none of the mutations was located within the β-barrel and in close proximity to the chromophore, some mutations may increase the performance by altering the aggregation behaviour or translation and folding speed by exchanging less preferred to more frequently used codons in B. mycoides. Especially the mutation K185K in the KPS12 variant increased the codon usage preference from 1.93% to 5.1% (Table 2). This might be associated with an increase of the translation and/or the folding speed, which prob-ably prevents the accumulation of non-matured and non-functional pro-tein precursors. Interestingly, the mKate2 variant FusionRed also had a mutation at this position (Shemiakina et al., 2012), which is reported to alter kinetics and efficiency of protein maturation.
CONCLUSION
By applying random mutagenesis and fluorescence-assisted cell sorting on sfGFP and mKate2 mutational libraries in life B. mycoides cells, we were able to isolate three brighter and well expressed variants of each FP pro-tein for this bacterium. The improved performance of the FPs was con-firmed at the population level by monitoring colony development on solid growth medium and at the single-cell level by FC and fluorescence mi-croscopy of cells grown in liquid cultures. An extended applicability was proven by double-labelling of B. mycoides with the best performing vari-ants sfGFP(SPS6) and mKate(KPS12). This revealed that i) the fluorescence signals were simultaneously detectable and clearly distinguishable from each other and ii) that chromosomal integration of the reporter proteins reduces cell-to-cell signal variations. To our knowledge, this is the first report of a successful dual-FP-labelling approach in bacilli of the B. cereus sensu lato group. Constitutive expression of the FPs from replicative plas-mids did not affect the growth behaviour of B. mycoides and the plasmids were kept even without selection pressure by antibiotics over several cell generations. This indicates that FP expression does not represent a met-abolic burden to the cells and altogether shows that the novel variants are suitable visualization markers without causing a loss of plant colo-nization ability. Finally, the optimized variants proved to be highly suita-ble for confocal laser scanning microscopy (CLSM) observations to study plant-microbe interactions and endophytic processes of B. mycoides. As a case study, we visualized the early stages of endophytic colonization in a hydroponic system. In line with previous studies, the formation of a mul-ticellular matrix or microcolonies was revealed to be a prerequisite for en-dophytic colonization, in which the root hair and the elongation region
of root hairs constitute potential entry sites to establish an endophytic life-style. The variants reported can be also used to study the expression of genes with weak promoters and proved to be well expressed and de-tectable in additional soil- and rhizosphere-associated Bacillus species. Moreover, the brightest variant for both GFP and RFP were selected in the pH-shift group, which renders them especially suitable to study bacteria- plant interactions.
ACKNOWLEDGEMENTS
Conceived and designed experiments: EF, YY, OK. Performed the experi-ments: YY and EF. Performed isolation of B. mycoides and gave advice on plant cultures: JS, TME, JDvE. Wrote the manuscript: EF, YY, OK. All au-thors revised and approved the final manuscript. We thank Anne de Jong from the Molecular Genetics Department of the University of Groningen for programming the Gene Alignment Analysis Tool and Anita Kram from the Molecular Cell Biology department of the University of Gronin-gen for her excellent technical advice on cell sorting. Y. Yi was supported by a grant of the Chinese Scholarship Council (CSC). The authors declare no competing financial interest.

55
108 109
Opt
imiz
ed fl
uore
scen
t pro
tein
s for
the
rhiz
osph
ere-
asso
ciat
ed
bact
eriu
m B
acill
us m
ycoi
des w
ith e
ndop
hytic
and
bio
cont
rol a
gent
pot
entia
l
Supp
lem
anta
ry M
ater
ial
Figure S2. Analysis of metabolic burden of FP expression and plasmid stability in B. my-coides EC18. (A) OD 600 measurements in LB medium comparing the growth behaviour of wild-type EC 18 and its derivative strains carrying the reporter proteins on replicative plasmids. (B) Cultivation of subsequent culture dilutions and final plating assay to deter-mine the plasmid presence with and without antibiotic pressure in B. mycoides.
Figure S3. Comparison of the performance of original and optimized FP variants fused to a weak, mannose-inducible promoter. (A) Detection of the expression of sfGFP(Sp) and the optimized sfGFP(SPS6) protein in exponential phase cells of B. mycoides EC18 by FC (B) Detection of the expression of mKate2 and optimized mKate(KSP12) in exponen-tial phase cells of B. mycoides EC18 by FC.
SUPPLEMANTARY MATERIAL
Figure S1. Workflow for in vivo isolation of optimized GFP [sfGFP(Sp)] and RFP (mKate2) variants from B. mycoides. GFP and RFP mutant libraries were constructed by random mutagenesis and cloned into the E. coli-Bacillus shuttle plasmids pNW-Ppta-3TER and pAD43-25 respectively. After transformation of B. mycoides, cells were grown in different pH conditions and two rounds of fluorescence-activated cell sort-ing were performed with a subsequent enrichment of ~3% and ~0.3% of the bright-est single cells from the total population. After plating, colonies displaying the high-est fluorescence signals were selected by use of a fluorescence stereo microscope.

55
110 111
Opt
imiz
ed fl
uore
scen
t pro
tein
s for
the
rhiz
osph
ere-
asso
ciat
ed
bact
eriu
m B
acill
us m
ycoi
des w
ith e
ndop
hytic
and
bio
cont
rol a
gent
pot
entia
l
Refe
renc
es
Figure S4. Three-dimensional structure of sfGFP(Sp) (PDB ID: 2B3P) and mKate2 (PDB ID: 3BXB) variants. The mutation sites of the improved FP variants are indicated in yel-low. The chromophore was highlighted in green for sfGFP (A–C) and red for mKate2 (D–F). A: S618; B: S709; C: SPS6; D: K603; E: K713; F: KPS12. The structure was visualized with the Cn3D software.
REFERENCES
Bloemberg, G.V., Wijfjes, A.H., Lamers, G.E.,
Stuurman, N., and Lugtenberg, B.J. (2000)
Simultaneous imaging of Pseudomonas
fluorescens WCS365 populations express-
ing three different autofluorescent pro-
teins in the rhizosphere: new perspectives
for studying microbial communities. Mol.
Plant Microbe Interact. 13: 1170–1176.
Blossfeld, S., Gansert, D., Thiele, B., Kuhn, A.J.,
and Lösch, R. (2011) The dynamics of oxy-
gen concentration, pH value, and organic
acids in the rhizosphere of Juncus spp. Soil
Biol. Biochem. 43: 1186–1197.
Buddrus-Schiemann, K., Schmid, M., Sch-
reiner, K., Welzl, G., and Hartmann, A.
(2010) Root colonization by Pseudomonas
sp. DSMZ 13134 and impact on the indige-
nous rhizosphere bacterial community of
barley. Microb. Ecol. 60: 381–393.
Brimecombe, M.J., De Leij, F.A.A.M., Lynch,
J.M. (2007) Rhizodeposition and Micro-
bial Populations. In The Rhizosphere-Bio-
chemistry and Organic Substances at the
Soil-Plant Interface. Pinton, R., Varanini,
Z., Nannipieri, P. (eds). Baco Raton, FL:
CRC Press,. pp. 73–89.
Chiu, W., Niwa, Y., Zeng, W., Hirano, T., Ko-
bayashi, H., and Sheen, J. (1996) Engi-
neered GFP as a vital reporter in plants.
Curr. Biol. 6: 325–330.
Chudakov, D.M., Matz, M.V., Lukyanov, S., and
Lukyanov, K.A. (2010) Fluorescent proteins
and their applications in imaging living
cells and tissues. Physiol. Rev. 90: 1103–1163.
Craggs, T.D. (2009) Green fluorescent protein:
structure, folding and chromophore mat-
uration. Chem. Soc. Rev. 38: 2865–2875.
Das, A.K., Hasegawa, J.Y., Miyahara, T., Ehara,
M., and Nakatsuji, H. (2003) Electronic ex-
citations of the green fluorescent protein
chromophore in its protonation states: SAC/
SAC-CI study. J. Comput. Chem. 24: 1421–1431.
Day, R.N., and Davidson, M.W. (2009) The flu-
orescent protein palette: tools for cellular
imaging. Chem. Soc. Rev. 38: 2887–2921.
Di Franco, C., Pisaneschi, G., and Beccari, E.
(2000) Molecular analysis of two roll-
ing-circle replicating cryptic plasmids, pB-
MYdx and pBMY1, from the soil gramposi-
tive Bacillus mycoides. Plasmid 44: 280–284.
Di Franco, C., Beccari, E., Santini, T., Pisaneschi,
G., and Tecce, G. (2002) Colony shape as a
genetic trait in the pattern-forming Bacil-
lus mycoides. BMC Microbiol. 2: 33.
Dunn, A.K., and Handelsman, J. (1999) A vec-
tor for promoter trapping in Bacillus cereus.
Gene 226: 297–305.
Ehling-Schulz, M., Vukov, N., Schulz, A., Sha-
heen, R., Andersson, M., Martlbauer, E.,
and Scherer, S. (2005) Identification and
partial characterization of the nonribo-
somal peptide synthetase gene responsi-
ble for cereulide production in emetic Ba-
cillus cereus. Appl. Environ. Microbiol. 71:
105–113.
Ehrig, T., O’Kane, D.J., and Prendergast, F.G.
(1995) Green-fluorescent protein mutants
with altered fluorescence excitation spec-
tra. FEBS Lett. 367: 163–166.
Eijlander, R.T., and Kuipers, O.P. (2013) Live-cell
imaging tool optimization to study gene
expression levels and dynamics in single
cells of Bacillus cereus. Appl. Environ. Mi-
crobiol. 79: 5643–5651.
Fan, B., Chen, X.H., Budiharjo, A., Bleiss, W.,
Vater, J., and Borriss, R. (2011) Efficient col-
onization of plant roots by the plant growth
promoting bacterium Bacillus amyloliquefa-
ciens FZB42, engineered to express green flu-
orescent protein. J. Biotechnol. 151: 303–311.
Frenzel, E., Legebeke, J., van Strahlen, A., van
Kranenburg, R., Kuipers, O.P. (2017) In vivo
selection of sfGFP variants with improved
and reliable functionality in industrially
Altenbuchner, J. (2016) Editing of the Bacillus
subtilis genome by the CRISPR-Cas9 system.
Appl. Environ. Microbiol. 82: 5421–5427.
Ambrosini, A., Stefanski, T., Lisboa, B.B., Bene-
duzi, A., Vargas, L.K., and Passaglia, L.M.P.
(2016) Diazotrophic bacilli isolated from
the sunflower rhizosphere and the poten-
tial of Bacillus mycoides B38V as biofer-
tiliser. Ann. Appl. Biol. 168: 93–110.
Athukorala, S.N., Fernando, W.D., and Rashid,
K.Y. (2009) Identification of antifungal
antibiotics of Bacillus species isolated
from different microhabitats using poly-
merase chain reaction and MALDI-TOF
mass spectrometry. Can. J. Microbiol. 55:
1021–1032.
Barbez, E., Dunser, K., Gaidora, A., Lendl, T.,
and Busch, W. (2017) Auxin steers root cell
expansion via apoplastic pH regulation in
Arabidopsis thaliana. Proc. Natl. Acad. Sci.
U. S. A. 114: E4884–E4893.
Bargabus, R.L., Zidack, N., Sherwood, J., and Ja-
cobsen, B. (2002) Characterisation of sys-
temic resistance in sugar beet elicited by a
non-pathogenic, phyllosphere-colonizing
Bacillus mycoides, biological control agent.
Physiol. Mol. Plant. Pathol. 61: 289–298.
Bargabus, R.L., Zidack, N.K., Sherwood, J.E., and
Jacobsen, B.J. (2004) Screening for the iden-
tification of potential biological control
agents that induce systemic acquired resis-
tance in sugar beet. Biol. Control 30: 342–350.

55
112 113
Opt
imiz
ed fl
uore
scen
t pro
tein
s for
the
rhiz
osph
ere-
asso
ciat
ed
bact
eriu
m B
acill
us m
ycoi
des w
ith e
ndop
hytic
and
bio
cont
rol a
gent
pot
entia
l
Refe
renc
es
important thermophilic bacteria. Biotech-
nol. Biofuels. (in press).
Fukuda, H., Arai, M., and Kuwajima, K. (2000)
Folding of green fluorescent protein
and the cycle3 mutant. Biochemistry 39:
12025–12032.
Germaine, K., Keogh, E., Garcia-Cabellos, G.,
Borremans, B., Lelie, D., Barac, T., et al.
(2004) Colonisation of poplar trees by gfp
expressing bacterial endophytes. FEMS
Microbiol. Ecol. 48: 109–118.
Guetsky, R., Shtienberg, D., Elad, Y., Fischer, E.,
and Dinoor, A. (2002) Improving biologi-
cal control by combining biocontrol agents
each with several mechanisms of disease
suppression. Phytopathology 92: 976–985.
Guinebretière, M.-H., Thompson, F.L., So-
rokin, A., Normand, P., Dawyndt, P., Eh-
ling-Schulz, M., et al. (2008) Ecological
diversification in the Bacillus cereus Group.
Environ. Microbiol. 10: 851–865.
Hebisch, E., Knebel, J., Landsberg, J., Frey, E.,
Leisner, M., and Hofmann, A. (2013) High
variation of fluorescence protein matu-
ration times in closely related Escherichia
coli strains. PLoS One 8: e75991.
Heim, R., Prasher, D.C., and Tsien, R.Y. (1994)
Wavelength mutations and posttransla-
tional autoxidation of green fluorescent
protein. Proc. Natl. Acad. Sci. U. S. A. 91:
12501–12504.
Heim, R., Cubitt, A.B., and Tsien, R.Y. (1995)
Improved green fluorescence. Nature 373:
663–664.
Hoagland, D.R., and Arnon, D.I. (1950) The
water-culture method for growing plants
without soil. Circular California Agric. Exp.
Station C347: 36–39.
Hsu, S.T., Blaser, G., and Jackson, S.E. (2009)
The folding, stability and conformational
dynamics of beta-barrel fluorescent pro-
teins. Chem. Soc. Rev. 38: 2951–2965.
mycoides isolate BmJ and B. mojavensis iso-
late 203-7 for the control of anthracnose of
cucurbits caused by Glomerella cingulata
var. orbiculare. Biol. Control 48: 140–146.
Overkamp, W., Beilharz, K., Detert Oude Weme,
R., Solopova, A., Karsens, H., Kovacs, A.T.,
et al. (2013) Benchmarking various green
fluorescent protein variants in Bacillus
subtilis, Streptococcus pneumoniae, and
Lactococcus lactis for live cell imaging.
Appl. Environ. Microbiol. 79: 6481–6490.
Pedelacq, J.D., Cabantous, S., Tran, T., Terwil-
liger, T.C., and Waldo, G.S. (2006) En-
gineering and characterization of a su-
perfolder green fluorescent protein. Nat.
Biotechnol. 24: 79–88.
Piatkevich, K.D., Hulit, J., Subach, O.M., Wu, B.,
Abdulla, A., Segall, J.E., and Verkhusha,
V.V. (2010) Monomeric red fluorescent
proteins with a large Stokes shift. Proc.
Natl. Acad. Sci. U. S. A. 107: 5369–5374.
Pozniak, P.D., Darbinyan, A., and Khalili, K.
(2016) TNF-α/ TNFR2 regulatory axis stim-
ulates EphB2-mediated neuroregenera-
tion via activation of NF-κB. J. Cell Physiol.
231: 1237–1248.
Prasher, D.C., Eckenrode, V.K., Ward, W.W., Pren-
dergast, F.G., and Cormier, M.J. (1992) Pri-
mary structure of the Aequorea victoria
green-fluorescent protein. Gene 111: 229–233.
Prieto, P., Schiliro, E., Maldonado-Gonzalez,
M.M., Valderrama, R., Barroso-Albarracin,
J.B., and Mercado-Blanco, J. (2011) Root hairs
play a key role in the endophytic coloniza-
tion of olive roots by Pseudomonas spp. with
biocontrol activity. Microb. Ecol. 62: 435–445.
Rasko, D.A., Ravel, J., Økstad, O.A., Helgason,
E., Cer, R.Z., Jiang, L., et al. (2004) The ge-
nome sequence of Bacillus cereus ATCC
10987 reveals metabolic adaptations and a
large plasmid related to Bacillus anthracis
pXO1. Nucleic Acids Res. 32: 977–988.
Ji, X., Lu, G., Gai, Y., Zheng, C., and Mu, Z.
(2008) Biological control against bacterial
wilt and colonization of mulberry by an
endophytic Bacillus subtilis strain. FEMS
Microbiol. Ecol. 65: 565–573.
Kilaru, S., Schuster, M., Studholme, D., Soanes,
D., Lin, C., Talbot, N.J., and Steinberg, G.
(2015) A codon-optimized green fluorescent
protein for live cell imaging in Zymoseptoria
tritici. Fungal Genet. Biol. 79: 125–131.
Kjos, M., Aprianto, R., Fernandes, V.E., Andrew,
P.W., van Strijp, J.A.G., Nijland, R., et al. (2015)
Bright fluorescent Streptococcus pneumo-
niae for live-cell imaging of host-pathogen
interactions. J. Bacteriol. 197: 807–818.
Kloepper, J.W., Ryu, C.-M., and Zhang, S. (2004)
Induced systemic resistance and promo-
tion of plant growth by Bacillus spp. Phyto-
pathology 94: 1259–1266.
Kremers, G.-J., Gilbert, S.G., Cranfill, P.J., Davidson,
M.W., and Piston, D.W. (2011) Fluorescent
proteins at a glance. J. Cell. Sci. 124: 157–160.
Larrainzar, E., O’gara, F., and Morrissey, J.P.
(2005) Applications of autofluorescent
proteins for in situ studies in microbial
ecology. Annu. Rev. Microbiol. 59: 257–277.
Leroch, M., Mernke, D., Koppenhoefer, D.,
Schneider, P., Mosbach, A., Doehlemann,
G., and Hahn, M. (2011) Living colors in
the gray mold pathogen Botrytis cinerea:
codon-optimized genes encoding green
fluorescent protein and mCherry, which
exhibit bright fluorescence. Appl. Environ.
Microbiol. 77: 2887–2897.
Nakamura, L., and Jackson, M. (1995) Clarifica-
tion of the taxonomy of Bacillus mycoides.
Int. J. Syst. Evol. Microbiol. 45: 46–49.
Namy, O., Mock, M., and Fouet, A. (1999) Co-ex-
istence of clpB and clpC in the Bacillaceae.
FEMS Microbiol. Lett. 173: 297–302.
Neher, O.T., Johnston, M.R., Zidack, N.K., and
Jacobsen, B.J. (2009) Evaluation of Bacillus
Rizzo, M.A., Davidson, M.W., and Piston, D.W.
(2009) Fluorescent protein tracking and
detection: fluorescent protein structure
and color variants. Cold Spring Harb. Pro-
toc. 2009: pdb. top63.
Rodriguez, E.A., Tran, G.N., Gross, L.A., Crisp,
J.L., Shu, X., Lin, J.Y., and Tsien, R.Y. (2016)
A far-red fluorescent protein evolved from
a cyanobacterial phycobiliprotein. Nat.
Methods 13: 763–769.
Samapundo, S., Devlieghere, F., Xhaferi, R.,
and Heyndrickx, M. (2014) Incidence, di-
versity and characteristics of spores of psy-
chrotolerant spore formers in various REP-
FEDS produced in Belgium. Food Microbiol.
44: 288–295.
Sambrook, J., Fritsch, E.F., and Maniatis, T.
(1989) Molecular Cloning: A Laboratory
Manual. New York: Cold Spring Harbor
Laboratory Press.
Sastalla, I., Chim, K., Cheung, G.Y., Pomerant-
sev, A.P., and Leppla, S.H. (2009) Codon-
optimized fluorescent proteins designed
for expression in low-GC gram-positive
bacteria. Appl. Environ. Microbiol. 75:
2099–2110.
Shaner, N.C., Steinbach, P.A., and Tsien, R.Y.
(2005) A guide to choosing fluorescent
proteins. Nat. Methods 2: 905–909.
Shapiro, J.A. (1998) Thinking about bacterial
populations as multicellular organisms.
Annu Rev Microbiol 52: 81–104. Shemiak-
ina, I.I., Ermakova, G.V., Cranfill, P.J., Baird,
M.A., Evans, R.A., Souslova, E.A., et al.
(2012) A monomeric red fluorescent pro-
tein with low cytotoxicity. Nat. Commun.
3: 1204.
Shimomura, O., Johnson, F.H., and Saiga, Y.
(1962) Extraction, purification and proper-
ties of aequorin, a bioluminescent protein
from the luminous hydromedusan, Ae-
quorea. J. Cell Physiol. 59: 223–239.

5
114
Refe
renc
es
Shu, X., Shaner, N.C., Yarbrough, C.A., Tsien, R.Y.,
and Remington, S.J. (2006) Novel chromo-
phores and buried charges control color in
mFruits. Biochemistry 45: 9639–9647.
Stefan, M., Munteanu, N., Stoleru, V., Mihasan,
M., and Hritcu, L. (2013) Seed inoculation
with plant growth promoting rhizobacte-
ria enhances photosynthesis and yield of
runner bean (Phaseolus coccineus L.). Sci.
Hortic. 151: 22–29.
Turchi, L., Santini, T., Beccari, E., and Di Franco,
C. (2012) Localization of new peptidogly-
can at poles in Bacillus mycoides, a member
of the Bacillus cereus group. Arch. Microbiol.
194: 887–892.
Wenzel, M., Muller, A., Siemann-Herzberg, M.,
and Altenbuchner, J. (2011) Self-inducible
Bacillus subtilis expression system for re-
liable and inexpensive protein production
by high-cell-density fermentation. Appl.
Environ. Microbiol. 77: 6419–6425.
Wilson, A.C., Perego, M., and Hoch, J.A. (2007)
New transposon delivery plasmids for
insertional mutagenesis in Bacillus an-
thracis. J. Microbiol. Methods 71: 332–335.
Yi, Y., de Jong, A., Frenzel, E., and Kuipers, O.P.
(2017) Comparative transcriptomics of Ba-
cillus mycoides strains in response to pota-
to-root exudates reveals different genetic
adaptation of endophytic and soil isolates.
Front. Microbiol 8: 1487.

6Exploring plant-microbe interactions of the rhizobacteria Bacillus subtilis and Bacillus mycoides by use of the CRISPR-Cas9 system
Yanglei Yi, Zhibo Li, Chunxu Song, Oscar P. Kuipers
Molecular Genetics, Groningen Biomolecular Sciences and Biotechnology Institute, University of Groningen, Groningen, The Netherlands
This chapter is in preparation for publication

6
119
Expl
orin
g pl
ant-m
icro
be in
tera
ctio
ns o
f the
rhiz
obac
teri
a Ba
cillu
s sub
tilis
and
Bac
illus
myc
oide
s by
use
of th
e CR
ISPR
-Cas
9 sy
stem
ABSTRACT
Bacillus subtilis HS3 and Bacillus mycoides EC18 are two rhizosphere-asso-ciated bacteria with plant growth-promoting activity. The CRISPR-Cas9 system was implemented to study various aspects of plant-microbe in-teraction mechanisms of these two environmental isolates. The results show that fengycin and surfactin are involved in the antifungal activity of B. subtilis HS3. Moreover, this strain emits several other volatile organic compounds (VOCs) than 2,3-butanediol, contributing to plant growth promotion. CLSM (Confocal laser scanning microscopy) observations of the GFP-labelled strain showed that HS3 selectively colonizes root hairs of grass (Lolium perenne) in a hydroponic system. For B. mycoides EC18, we found that the wild-type EC18 strain and a ∆asbA (petropectin-deficient) mutant, but not the ∆dhbB (bacillibactin-deficient) and ADKO (asbA and dhbB double knockout) mutants, can increase the plant biomass and to-tal chlorophyll. All the mutant strains have a reduced colonization ca-pability on Chinese cabbage (Brassica rapa) roots, at the root tip and root hair region compared to the wild-type strain. These results indicate that the siderophore, bacillibactin, is involved in the plant growth promot-ing activity and could affect the root colonization of B. mycoides. Collec-tively, the CRISPR/Cas9 system we developed for environmental isolates is broadly applicable and will facilitate deciphering the mechanisms of Bacillus-plant interactions.
INTRODUCTION
Plant growth-promoting rhizobacteria (PGPR) are soil bacteria that are able to colonize the surface of the root (and sometimes root inner tissues) and to stimulate plant growth and health (Vacheron et al., 2013). The use of PGPR as supplements to, or replacements of, chemical fertilizers or pesticides have steadily increased in the last decades (Adesemoye and Kloepper, 2009). A large diversity of bacterial species has been reported to have PGPR activity. Bacillus is one of the PGPR that received most exten-sive attention due to its specific metabolic and physiological traits includ-ing secretion of antimicrobial or signal peptides and formation of stress resistant-endospores (Kumar et al., 2011), which facilitate the formulation into commercial fertilizers or biocontrol agents.
B. subtilis is widely recognized as a model organism for studying plant-microbe interaction, because it was found in association with dif-ferent plants as a good colonizer, which is highly important for biocon-trol application. The colonization of B. subtilis on root surfaces involves chemotaxis and biofilm formation mechanisms (Beauregard et al., 2013; Allard-Massicotte et al., 2016). After colonization, B. subtilis benefits the plant growth in various ways, including suppressing plant pathogens by the secretion of antimicriobial compounds, such as fengycin (Romero et al., 2007) and/or the induction of systematic resistance (Ongena et al., 2007). Moreover, direct plant growth promotion effects can be achieved by the secretion of cytokine hormones and volatiles that modify plant hormone homeostasis. Apart from B. subtilis, other Bacillus species with biocontrol and plant growth promotion (PGP) potential have broadened the application capacity of this genus. For instance, the rhizosphere- associated B. mycoides can actively colonize root surfaces and form bio-film-like matrix, while some of the strains are even capable of entering root tissues and employ a endophytic life strategy (Yi et al., 2017a). It has been reported that B. mycoides increases disease resistance of sugar beet through the elicitation of systemic resistance (Bargabus et al., 2002) or oxidative burst (Bargabus et al., 2003) of the host. The biocontrol effects of B. mycoides against Botrytis cinerea and Pythium aphanidermatum were confirmed in greenhouse conditions (Guetsky et al., 2002; Peng et al., 2017). Moreover, B. mycoides strains promote sunflower growth due to their nitrogen fixation activity (Ambrosini et al., 2016).
Although the significance of applying Bacillus with PGPR properties is being appreciated, the difficulty to study plant interaction mechanisms is hampered by the absence of an efficient genome editing system, e.g. enabling gene deletion/inactivation/insertion in different Bacillus envi-ronmental isolates. Conventional methods for Bacillus gene inactivation

66
120 121
Expl
orin
g pl
ant-m
icro
be in
tera
ctio
ns o
f the
rhiz
obac
teri
a Ba
cillu
s sub
tilis
and
Bac
illus
myc
oide
s by
use
of th
e CR
ISPR
-Cas
9 sy
stem
Intr
oduc
tion
based on single or double crossover homologous recombination of a chro-mosomal gene by a mutated allele carried on a suicide vector require a relatively high transformation efficiency (Vagner et al., 1998; Zhang et al., 2015). This limitation hampers the application of this method since most of the undomesticated Bacillus strains are poorly naturally competent or otherwise hard to transform. To overcome this, a two-step system was de-veloped for non-transformable Bacillus, which involved a second counter selection step by an antibiotic marker or a toxic gene (Suzuki et al., 2012; Dong and Zhang, 2014). However, those methods are still time- consuming and tedious since large-scale screening is needed for selecting desired double crossover events. Apart from the homologous recombination mutagenesis methods, an insertion-based mutagenesis including group II intron and transposon insertion normally either have low frequency or show a preference for a particular insertion motif (Shevchenko et al., 2002; Green et al., 2012; Saldanha et al., 2013). For these reasons, there is a strong necessity for developing a more efficient method for directed mutagenesis in environmental Bacillus species.
Clustered regularly interspaced short palindromic repeat (CRISPR) and the associated system is an adaptive immune system of bacteria and ar-chaea against bacteriophages (Barrangou et al., 2007). The type II CRISPR system from Streptococcus pyogenes is the most compact CRISPR machin-ery by now and has been engineered to a precise genome editing tool (Jinek et al., 2012). This system has been adapted for targeted genome ed-iting in not only eukaryotes, but also prokaryotes. For example, it has been successfully applied in a wide variety of bacteria including Escherichia coli (Li et al., 2015; Pyne et al., 2015), S. pneumonia (Jiang et al., 2013), Lactoba-cillus reuteri (Oh and van Pijkeren, 2014), Streptomyces sp.(Tong et al., 2015), Clostridium spp. (Wang et al., 2016) etc. In Bacillus, such system has been developed for the model organism B. subtilis 168 by Altenbuchner (2016). The plasmid pJOE8999 contains a cas9 gene controlled by a mannose in-ducible promoter and a sgRNA driven by a constitutive promoter. A simi-lar system has been developed for B. subtilis ATCC6051 (Zhang et al., 2016). A year after, Park and co-workers described a two-plasmid CRISPR/Cas9 system to delete large fragments of in the B. subtilis chromosome (So et al., 2017). However, those methods are developed either for model strains or for undomesticated strains with natural transformability.
B. subtilis HS3 and B. mycoides EC18 were isolated from grass rhizos-phere and potato endosphere, respectively. B. subtilis strain HS3 displays excellent antifungal and plant growth promoting activity, while B. my-coides EC18 has good endophytic and plant growth promotion potential. Inspired by the high genome editing efficiency in prokaryotic organ-isms of the CRISPR/Cas9 system, we attempted to implement this system
in these two rhizosphere-associated Bacillus strains. In order to further investigate the molecular mechanisms of their plant-microbe interac-tions, we describe a high efficient genome editing method based on the CRISPR/Cas9 system. By applying this method, we generated three and two mutants in B. subtilis HS3 and B. mycoides EC18, respectively, with po-tential relevance for their biocontrol abilities. Genetic, phenotypic and microscopic analyses were conducted to assess functions of the mutants during plant-Bacillus interactions.
MATERIALS AND METHODS
Bacterial strains and growth conditionsStrains used in this study are shown in Table 1. E. coli MC1061 was em-ployed for plasmid construction. B. subtilis HS3 and B. mycoides EC18 were isolated from grass rhizosphere (Groningen, the Netherlands) and potato endosphere (Wijster, the Netherlands) respectively. E. coli and B. subti-lis were grown in Lysogeny Broth (LB-Lennox: 1% Bacto-Tryptone, 0.5% Bacto-yeast extract, 0.5% NaCl) at 37 °C, 220 rpm. B. mycoides was grown in brain heart infusion (BactoTM BHI, BD Bioscience, France) medium at 30 °C, 200 rpm. When required, antibiotics were added to the medium in the following final concentrations: 30 µg ml−1 or 10 µg ml−1 of kanamycin for E. coli and B. subtilis HS3 respectively. 100 µg ml−1 of spectinomycin for both E. coli, and B. mycoides EC18. 0.2% of mannose was added to the me-dium to induce the cas9 expression. Solid media were prepared by adding 1.0% (wt/vol) agar (Bacto Agar, BD Bioscience, France).
DNA manipulation and oligonucleotidesGenomic DNA was isolated using GenElute bacterial genomic DNA kit (Sigma-Aldrich, USA) according to the manufacture’s instruction. Plas-mid DNA isolation, PCR products clean-up, and gel extraction were per-formed using commercial NucleoSpin kits (Macherey-Nagel GmbH & Co., Germany). Restriction, ligation, agarose gel electrophoresis, and trans-formation of E. coli were performed as established protocols. Phusion (Thermo Scientific) polymerases were used for PCR to construct plasmid and DreamTaq polymerases DNA (Thermo Scientific) was used for colony PCR. Annealing reaction was carried out in annealing buffer containing 10 mM Tris (pH 8.0) and 50 mM NaCl with 10 µM of each oligonucleotide. Reaction tubes were heated to 95 °C for 5 min and cool to room tempera-ture slowly. Oligonucleotides are listed in Table S1.

66
122 123
Expl
orin
g pl
ant-m
icro
be in
tera
ctio
ns o
f the
rhiz
obac
teri
a Ba
cillu
s sub
tilis
and
Bac
illus
myc
oide
s by
use
of th
e CR
ISPR
-Cas
9 sy
stem
Mat
eria
ls a
nd m
etho
ds
Table 1. Plasmids and strains used in this study.
Strains or plasmids Relevant characteristic ReferenceStrains E. coli MC1061 F-, araD139, D(ara-leu)7696, D(lac)X74, galU,
galK, hsdR2, mcrA, mcrB1, rspLLab stock
B. subtilis HS3 isolated from grass rhizosphere This studyB. mycoides EC18 isolated from potato endosphere Yi et al. (2017)B. subtilis HS3ΔsigF HS3 derivative, ΔsigF This studyB. subtilis HS3ΔbdhA HS3 derivative, ΔbdhA This studyB. subtilis HS3Δsfp HS3 derivative, Δsfp This studyB. subtilis HS3ΔalsD HS3 derivative, ΔalsD This studyB. subtilis HS3GFP HS3 derivative, GFP inserted into the chro-
mosome. This study
B. mycoides EC18ΔsigF EC18 derivative, ΔsigF This studyB. mycoides EC18ΔasbA EC18 derivative, ΔasbA This studyB. mycoides EC18ΔdhbB EC18 derivative, ΔdhbB This studyB. mycoides EC18ADKO EC18 derivative, ΔasbA, ΔdhbB This studyB. mycoides EC18GFP EC18 derivative, GFP inserted into the chro-
mosome.This study
B. mycoides EC18ΔAGFP EC18 derivative, ΔasbA, GFP inserted into the chromosome.
This study
B. mycoides EC18ΔDGFP EC18 derivative, ΔdhbB, GFP inserted into the chromosome.
This study
B. mycoides EC18ADGFP EC18 derivative, ΔasbA, ΔdhbB, GFP in-serted into the chromosome.
This study
PlasmidspJOE8999 PUC ori, kanR, rep pE194ts, gRNA,
Pman-cas9. Altenbuchner (2016)
pYCR PUC ori, spec R, rep pWVO1ts, gRNA, Pman-cas9.
This study
PDR111_GFP(Sp) bla amyE’ Phyperspank, specR, lacI ‘amyE Overkamp et al. (2013)pNW-sfGFP(SPS6) E. coli-Bacillus shutter vector, Ppta-sfGF-
P(SPS6), cmRYi et al. (2018)
PAW068 PUC ori, C9 transposase, rep pWVO1ts, cmR, specR
Wilson et al. (2007)
pJOE8999_gsigFHR pJOE8999 derivative, containing 20-nt spacer targeting sigF and fused up- and down- homologous fragment.
This study
pJOE8999_gsfpHR pJOE8999 derivative, containing 20-nt spacer targeting sfp and fused up- and down- homologous fragment.
This study
pJOE8999_galsDHR pJOE8999 derivative, containing 20-nt spacer targeting alsD and fused up- and down- homologous fragment.
This study
pJOE8999_gbdhHR pJOE8999 derivative, containing 20-nt spacer targeting bdhA and fused up- and down- homologous fragment.
This study
pJOE8999_gamyGFP pJOE8999 derivative, containing 20-nt spacer targeting α-amylase gene, and a gfp gene between the fused up- and down- ho-mologous fragment.
This study
pYCR_gsigFHR pYCR derivative, containing 20-nt spacer targeting sigF and fused up- and down- ho-mologous fragment.
This study
pYCR_gasbHR pYCR derivative, containing 20-nt spacer targeting asbA and fused up- and down- ho-mologous fragment.
This study
pYCR_gdhbHR pYCR derivative, containing 20-nt spacer targeting dhbB and fused up- and down- ho-mologous fragment.
This study
pYCR_gamyGFP pYCR derivative, containing 20-nt spacer targeting α-amylase gene, and a gfp gene between the fused up- and down- homolo-gous fragment.
This study
Plasmid constructionThe plasmid pJOE8999 (Altenbuchner, 2016) was used for B. subtilis HS3 genome editing. For each mutant, the 20-nt protospacer sequence was de-signed by a web-based tool Benchling (https://www.benchling.com/) with B. subtilis 168 as reference genome and “NGG” as PAM (Protospacer adjacent motif) sequence. The sequence with high on-target and off-target scores was chosen. As a proof of principle, the sigF gene was chosen because it is conserved in Bacilli. sigF encodes the forespore specific transcription fac-tor σF that controls genes required for the early stages of prespore devel-opment (Hilbert and Piggot, 2004). To make a 121-bp deletion in the sigF gene, pJOE8999_gsigFHR was constructed in two steps. Plasmid pJOE8999 was first digested by BsaI resulting a 7,423 bp fragment, which was ligated with the annealing product of primers HS3_gsigF_F and HS3_gsigF_R to generate plasmid pJOE8999_gsigF. A fragment of ~1 kb upstream and downstream of the spacer sequence was amplified by HS3_sigFHR1_F & HS3_sigFHR1_R and HS3_sigFHR2_F & HS3_sigFHR2_R primer sets, respec-tively. The two fragments were connected by overlap PCR with the primers HS3_sigFHR1_F and HS3_sigFHR2_R, then digested with SfiI, purified, and ligated into pJOE8999_gsigF to give plasmid pJOE8999_gsigFHR. In a sim-ilar way, pJOE8999_gbdhHR, pJOE8999_galsDHR, pJOE8999_gsfpHR vec-tors were constructed with designed primers (see Table S1) to make bdhA, alsD, and sfp gene knockout mutant. To insert a GFP into the genome, an α-amylase gene was chosen as integration location. pJOE8999_gamy vec-tor was constructed by inserting HS3_gamy_F and HS3_gamy_R anneal-ing product into BsaI site. Then a 2,259 bp fragment including the whole α-amylase gene was amplified from the B. subtilis HS3 genome by primer set HS3_amy1F_sfiI & HS3_amy2R_sfiI, and ligated into the SfiI site of pJOE8999_gamy to obtain pJOE8999_gamyFL. The pJOE8999_gamyFL vector was used as a template to be amplified by primers HS3_amy1R and HS3_amy2F. The resulting PCR product was ligated with a gfp fragment ob-tained from pNW33N_sfGFP(SPS6) by EcoRI and HindIII cutting to give pJOE8999_gamyGFP.
Since B. mycoides has a low thermotolerance at 37 °C, the tempera-ture sensitive ori of pJOE8999 was changed from pET194ts to PWVO1, which is amplification permissive at 30 °C and non-permissive at 37 °C. The pJOE8999 backbone without pET194ts was amplified by primers pJOE-PET194_F and pJOE-PET194_R and the PWVO1 ori was amplified by primer PWV01_F and PWV01_R from plasmid PAW068. The two frag-ments were connected by Quick-fusion cloning to generate pJOE-PWVO1. Plasmid pYCR was constructed by isolating the SpecR cassete as a 1,025 bp XhoI/PstI fragment from PDR111_GFP(Sp) (Overkamp et al., 2013) and ligat-ing it with the XhoI/PstI-digested pJOE-PWVO1. The construction scheme

66
124 125
Expl
orin
g pl
ant-m
icro
be in
tera
ctio
ns o
f the
rhiz
obac
teri
a Ba
cillu
s sub
tilis
and
Bac
illus
myc
oide
s by
use
of th
e CR
ISPR
-Cas
9 sy
stem
Mat
eria
ls a
nd m
etho
ds
of the CRISPR/CAS plasmid pYCR used in B. mycoides is shown in Figure S1. When designing the 20-bp protospacer sequence in Benchling, the B. my-coides ATCC6462 genome was used as reference and “NGG” as PAM. As a proof-of-principle trial, a 60-bp deletion of sigF gene was achieved by us-ing plasmid pYCR_gsigFHR. The procedure to produce pYCR_gsigFHR was the same as that described for pJOE8999_gsigFHR. Plasmid pYCR was first digested with BsaI and ligated with the oligo annealing product of EC18_gsigF_F and EC18_gsigF_R to get the plasmid pYCR_gsigF. Then a
~2 kb fragment combined with an upstream and a downstream fragment of the sigF spacer sequence was inserted at the SfiI site of pYCR_gsigF to gener-ate pYCR_gsigFHR. In a similar way, vector pYCR_gasbHR, pYCR_gdhbHR, and pYCR_gamyGFP were constructed. For GFP insertion at the α-amylase gene locus, the vector pYCR_gamyGFP was generated with the same pro-cedure as for pJOE8999_gamyGFP. All the plasmids are listed in Table S1.
Electroporation and mutant generationThe electroporation factors including growth media, growth stage, elec-troporation buffer, pulse strength, and recovery incubation time were op-timized step-by-step for B. subtilis HS3. Briefly, one B. subtilis HS3 colony was inoculated in LBSP medium (LB medium supplemented with 50 mM KH2PO4 and K2HPO4, and 0. M sorbitol) overnight, then diluted 50 times in LBSP medium until the OD600 reached 0.65. The cell culture was centri-fuged 10 min at 4,000 g, 4 °C and the supernatant was discarded. The cell pellet was washed with pre-chilled electroporation buffer (0.25 M sorbi-tol in 10% glycerol solution) for 4 times and suspended in 1 ml electro-poration buffer. 100 μl aliquots were flash frozen in liquid nitrogen and stored at –80 °C until use. For B. mycoides EC18, the electroporation was performed as described previously (Yi and Kuipers, 2017).
About 1–2 µg plasmids were added into B. subtilis HS3 or B. mycoides EC18 electro-competent cells and exposed to a single pulse in a Gene Pul-ser System (Bio-Rad, USA) with the settings 25 μF, 10 kV cm−1, 200 Ω. Af-ter electroporation, 1 ml growth media were directly added to the cells and incubated for 16–18 h at 28 °C, 200 rpm. The whole cultures were then plated on LB plate with corresponding antibiotic (10 µg ml−1 kanamycin for HS3 and 100 µg ml−1 spectinomycin for EC18) and 0.2% mannose to activate the cas9 gene expression. After incubation at 28 °C for 24 h, po-tential mutants were randomly picked and subjected to colony PCR with primers binding the flanking region of deletion or insertion sites on the genome (Table S1). A control was included using the genomic DNA as a template for PCR. PCR products were purified and verified by DNA se-quencing. Colonies showing expected knockout band size were grown in LB liquid media with appropriate antibiotic and 0.2% mannose at 28 °C,
200 rpm overnight to remove any unedited cells. Edited bacterial cells carrying the knockout plasmid were inoculated into growth medium (LB for B. subtilis and BHI for B. mycoides) without antibiotic and grown at plasmid replication-defective temperatures (45 °C for B. subtilis and 37 °C for B. mycoides) to stationary phase. Two more rounds of growth were con-ducted by diluting the stationary phase culture into fresh medium. Then a serial dilution of the culture was plated on LB agar plates in the absence of antibiotics and incubated at plasmid replication-defective temperatures overnight to curing the plasmid. Plasmid-cured candidates were identi-fied by the loss of antibiotic resistance.
Microscopy For microscopic observation of sporulating cells of B. subtilis HS3 and B. mycoides EC18, the wildtype strains, as well as their sigF gene mutant HS3∆sigF and EC18∆sigF were first grown on LB agar plates for overnight at 30 °C. A single colony was inoculated into Schaeffer’s sporulation me-dium (Schaeffer et al., 1965) at 30 °C, 200 rpm. After growing for 30 h to a late stage of sporulation, 1 μl of the diluted culture was loaded on a 1.0% agarose microscopy slide. Samples were observed by a Nikon Ti-E micro-scope equipped with a CoolsnapHQ2 camera, a 40× phase contrast objec-tive, and an Intensilight light source. Pictures were taken with 0.35 s of exposure for phase contrast.
For the detection of fluorescent signals of the GFP insertion mutant, cells were grown in LB liquid medium overnight at 30 °C, 200 rpm. One microliter of the diluted cell cultures was observed as described before. Pictures were taken with the following settings: 0.35 s of exposure for phase contrast, 2 s exposure for fluorescence at 440–490 nm excitation via a dichroic mirror of 495 nm and an emission filter at 500–505 nm. The final pictures were generated by ImageJ software.
High-performance liquid chromatography (HPLC) and MALDI-TOFLipopeptides were extracted as previously described (Vater et al., 2002). Briefly, B. subtilis HS3 and its sfp mutant were grown in 200 ml LB broth at 28 °C, 220 rpm for overnight. The supernatant was collected by centrifuga-tion (10,000 × g, 10 min) and acidify by adding 6 M HCl to a final pH of 2.0. After overnight acidify at 4 °C, the precipitation was collected by centrifu-gation (5,000 × g, 20 min) and extracted with 10 ml methanol for 2 h. The crude extracts were filter sterilized with 0.45 µm DuraporeTM membrane.
To check the lipopeptide extraction, crude extracts were subjected to HPLC equipped with Aeris wide pore 3.6u XB-C18 250 × 4.6mm column. Eluent A was Mili-Q water with 0.1% trifluoroacetic acid (TFA); eluent B was HPLC-grade acetonitrile with 0.1% TFA. In each run, 100 μl crude

66
126 127
Expl
orin
g pl
ant-m
icro
be in
tera
ctio
ns o
f the
rhiz
obac
teri
a Ba
cillu
s sub
tilis
and
Bac
illus
myc
oide
s by
use
of th
e CR
ISPR
-Cas
9 sy
stem
Mat
eria
ls a
nd m
etho
ds
extract was injected and eluted by a two-step gradient from 5 to 45% elu-ent B in 20 min and from 45–95% eluent B in 75 min (20 to 95 min).
Peaks only existed in B subtilis HS3 wild-type strain and these were col-lected for MALDI-TOF analysis. Samples of 0.5 μl were spotted and dried on the target. Subsequently, 0.5 μl of matrix solution (5 mg/ml α- cyano-4-hydroxycinnamic acid dissolved in 50% acetonitrile containing 0.1% TFA) was spotted on top of the sample. Mass spectra were obtained using a Voyager DE PRO MALDI-TOF mass spectrometer (Applied Biosystems).
Antifungal testThe fungi were inoculated in PDA agar plate and incubated at 28 °C for 5 days. An agar plug (5 mm diameter) with fungal hyphae was then in-oculated into a new PDA agar plate. To prepare bacteria inoculums, an overnight culture of B. subtilis HS3 and its sfp mutant were pelleted and suspended in sterile MQ water to a final OD600 of 1.0. Subsequently, 5 μl of each inoculum were spotted 2 cm away from the fungi plug symmet-rically. Plates were sealed with parafilm and incubated at 28 °C for 5 days.
Effects of bacterial volatile organic compounds (VOCs) on plants and 2,3-butanediol detection To determine the effects of VOCs on the growth of perennial ryegrass (Lolium perenne), seedlings were exposed to the VOCs emitted by the wild-type B. subtilis HS3 or the mutants in a dual-dish plates system. The du-al-dish plates were prepared with a small petri dish containing LB (with 1% glucose) agar fixed in a large round petri dish containing ½ MS agar (Duchefa Biochemie). For surface sterilization, grass seeds were pre-treated with 0.3 M HCl for 6 h, followed by a 5 min bath in 2% sodium hypochlorite and then rinsed with sterile water for 10 times. The sterile seeds were sown on wet filter paper in a square plate, which was sealed with parafilm and then incubated at 25 °C in the dark. After sown and ger-mination for 5 days, the grass seedlings were then transferred to dual dish plates on ½ MS medium. After 2 days, 5 μl overnight bacterial culture (di-luted to OD600 = 1.0) was inoculated on LB medium in the dual-dish plates. Plates were sealed with parafilm and incubated in the climate chamber (25/21 °C of day/night, 16/8 h of day/night, 120 μmol m-2 s-1, 70% humid-ity). After 7 days, the plant dry weights were measured.
The production of 2,3-butanediol in the B. subtilis HS3 culture was de-tected by thin layer chromatography (TLC) as described before (Saran et al., 2014). Briefly, the bacteria were grown in 10 ml Erlenmeyer flasks contain-ing LB medium supplemented with 1% glucose at 30 °C, 150 rpm for 48 h. The culture was then collected and centrifuged at 10,000 × g for 10 min to spin down the cells. The supernatant was spotted on silica gel TLC plates
and use 70:30:1.5 of the hexane: ethyl acetate:glacial acetic acid as the mo-bile phase. Butanediol was visualized after the plates were sprayed with vanillin reagent and incubated at 100 °C until the spots were visible.
Siderophore detection Siderophore detection assay was performed using the overlay chrome az-urol S (O-CAS) method (Pérez-Miranda et al., 2007). Iron-deprived LB me-dium was made by stirring 5 g chelex 100 resin (Sigma, U.S.) with 100 mL LB for one hour. Strains were inoculated on iron-deprived LB agar plate at 30 °C overnight. The overlay CAS was applied to the plates and a change in color was observed after a period of one hour.
Effects of a siderophore-producing/-deficient strain on iron-starved plantsChinese cabbage seeds were first washed with sterilized water to remove dust and particles. The seeds were then immersed in 2–3% sodium hy-pochlorite for 5 min and rinsed with distilled water. Finally, seeds were dipped in 70% ethanol for 2–3 min followed by thorough rinsing with sterile water. The surface sterilized seeds were aseptically placed on 25% iron-deprived Hoagland medium solidified with 1% agar. The plates were incubated for germination and growth in a climate chamber (21 °C, 16/8 h of day/light, ~120 μmol m-2s-1, 72% humidity) for one week. The seedlings were transferred to a 3-L hydroponic system containing 25% Hoagland’s solution (without EDTA as chelate agent) and continued to grow for 2 days. Hoagland solution was aerated by air pump through a 0.45 μm fil-ter membrane. 10 mL of B. mycoides wild-type or mutant strain in expo-nential growth phase were collected by centrifugation at 10,000 × g for 1 min. The cell pellet was resuspended in sterile water and added to the hydroponic system with final concentration of 2–3 × 104 CFU/ml cells. Af-ter two weeks of co-culturing, plants were harvested and the fresh and dry weight of root and shoot were measured. The total chlorophyll content was determined according to the method of Arnon (1949).
Plant colonization observed by confocal laser scanning microscopeGrass seeds were germinated on wet filter paper under sterile conditions as described before. The seedlings were transferred to a hydroponic sys-tem containing 1 L of ½ MS liquid medium and grown for 2 days. Then the overnight culture of B. subtilis HS3_GFP was centrifuged at 10,000 rpm for 1 min. The cell pellet was suspended with ½ MS and inoculated to grass plants at a final concentration of 104 CFU/ml. GFP labeled B. my-coides EC18 wild-type and siderophore deficient mutants were visualized

66
128 129
Expl
orin
g pl
ant-m
icro
be in
tera
ctio
ns o
f the
rhiz
obac
teri
a Ba
cillu
s sub
tilis
and
Bac
illus
myc
oide
s by
use
of th
e CR
ISPR
-Cas
9 sy
stem
Resu
lts a
nd d
iscu
ssio
n
in the rhizosphere of Chinese cabbage. Seven days old iron-starved seed-lings were transferred to 25% Hoagland (EDTA was omitted) and grown for 2 days. B. mycoides EC18_GFP, EC18ΔasbA_GFP, EC18ΔdhbA_GFP, and EC18ADKO_GFP were grown overnight. Bacterial cells were collected at 10,000 rpm for 1 min and then inoculated to Chinese cabbage plants at a final concentration of 104 CFU/ml.
After 3 days of co-culturing with bacteria, root samples of grass and Chinese cabbage were used for observation by a confocal laser scanning microscope (CLSM, LSM 800, Carl Zeiss, Germany) equipped with diode lasers and GaAsP detector. Images for fluorescence and bright field light channels were taken simultaneously. The CLSM settings were adjusted as follows: 0.2% power of the 488-nm laser line was for excitation and 509–546 nm was set as emission wavelength. The pinhole size was 25 µm and pixel scanning time was 2.06 µs. The line scanning time was 2.47 ms with a line averaging of 2.
RESULTS AND DISCUSSION
Implementation of CRISPR/Cas9 systems in B. subtilis HS3 and B. mycoides EC18The all-in-one CRISPR/Cas9 genome editing system pJOE8999 has been established for the model strain of B. subtilis 168 previously ( Altenbuchner, 2016). The shuttle vector pJOE8999 has a pUC minimal origin of replica-tion (ori) to facilitate the cloning in E. coli and a gram-positive tempera-ture permissive ori pET194TS, which is permissive in B. subtilis below 37 °C and non-permissive above 42 °C. The cas9 gene is controlled by a man-nose inducible promoter and the sgRNA is driven by a strong constitu-tive promoter. Although this system is highly efficient in B. subtilis 168 via natural transformation, the implementation of such system in an envi-ronmental Bacillus strain has not been reported. The gene deletion of this system depends on a double-crossover allelic exchange event, which nor-mally happens at a low frequency (Leloup et al., 1997). Since many envi-ronmental strains lack a functional competence system, electroporation was chosen to incorporate the plasmid into bacterial strains. In B. subtilis HS3, the transformation efficiency reached to (4.2 ± 1.1) × 103 cfu µg-1 DNA when using pNW33n plasmid. We transformed pJOE8999_gsigFHR into HS3 to delete the sigF gene. In order to increase the probability of dou-ble-crossover events before the induction of cas9 gene, the cells were cul-tured overnight at 28 °C, 200 rpm directly after electroporation in 1 mL LBSP medium without antibiotic and mannose. After plating the trans-formed cultures on LB agar with 10 µg ml−1 kanamycin and 0.2% mannose,
approximately 200 colonies were obtained per µg plasmid DNA. Colony PCR with primer HS3_sigFcheckF and HS3_sigFcheckR showed a 100% editing efficiency (number of colonies with desired fragment size/total colony checked) (Figure S2). The mutant was cured from plasmids and confirmed by PCR (Figure 1A). Then it was subjected to microscope ob-servation for the sporulation ability. As shown in Figure 1C&D, almost all wild-type cells complete the sporulation process and contain phase-bright forespores. In comparison, the mutant B. subtilis HS3∆sigF cells have an abnormal morphology caused by a failure to form proper spores.
Genetic manipulation tools for B. mycoides have not been developed so far. Since the pJOE8999 system had high efficiency in the environmental isolate B. subtilis HS3, we developed a similar system, pYCR, for B. mycoides (Figure S1). In order to knockout the sigF gene, plasmid pYCR_gsigFHR was transformed into B. mycoides EC18 by electroporation as described previ-ously (Yi and Kuipers, 2017). The transformed cells were cultured in BHIS medium overnight and plated on LB agar plates with 100 µg ml−1 spectin-omycin and 0.2% mannose. After incubation for 24 h at 30 °C, the trans-formants showed heterogeneity in colony size (Figure S3). However, only the big colonies could grow when single colonies were inoculated into LB medium with 100 µg ml−1 spectinomycin. Thus, we speculated that the small colonies were possibly false positive colonies. Several “true” transformants (big colonies) were checked by colony PCR with primers EC18_sigFcheckF and EC18_sigFcheckR. Ten out of thirteen colonies showed the expected mutant fragment size, which resulted in an editing efficiency of 77% (Fig-ure S2). The plasmid-cured mutant was confirmed by PCR and then grown in sporulation medium for microscopic observation. As shown in Fig-ure 1D&E, the EC18 wild-type strain has entered the last stage of sporula-tion and phase-bright forespores can be observed, while the mutant strain EC18∆sigF showed an abnormal cell morphology and some cells lysed.
Fengycin and surfactin lipopeptides are involved in antifungal activity of B. subtilis HS3Production of antimicrobials by rhizobacteria is one of the major mecha-nisms for its antagonistic activity against phytopathogens, and they have great application potential being used as biocontrol agents. It has been reported that lipopeptides are the main antifungal compounds produced by Bacillus during the interactions with fungi (Zihalirwa Kulimushi et al., 2017). Antagonistic experiments showed that B. subtilis HS3 has signifi-cant antifungal activity against several phytopathogenic fungi (data not shown). In order to investigate if lipopeptides are (partially) responsible for the antifungal activity, we knocked out the 4’-phosphopantetheinyl transferase encoding gene (sfp) which is required for the production of
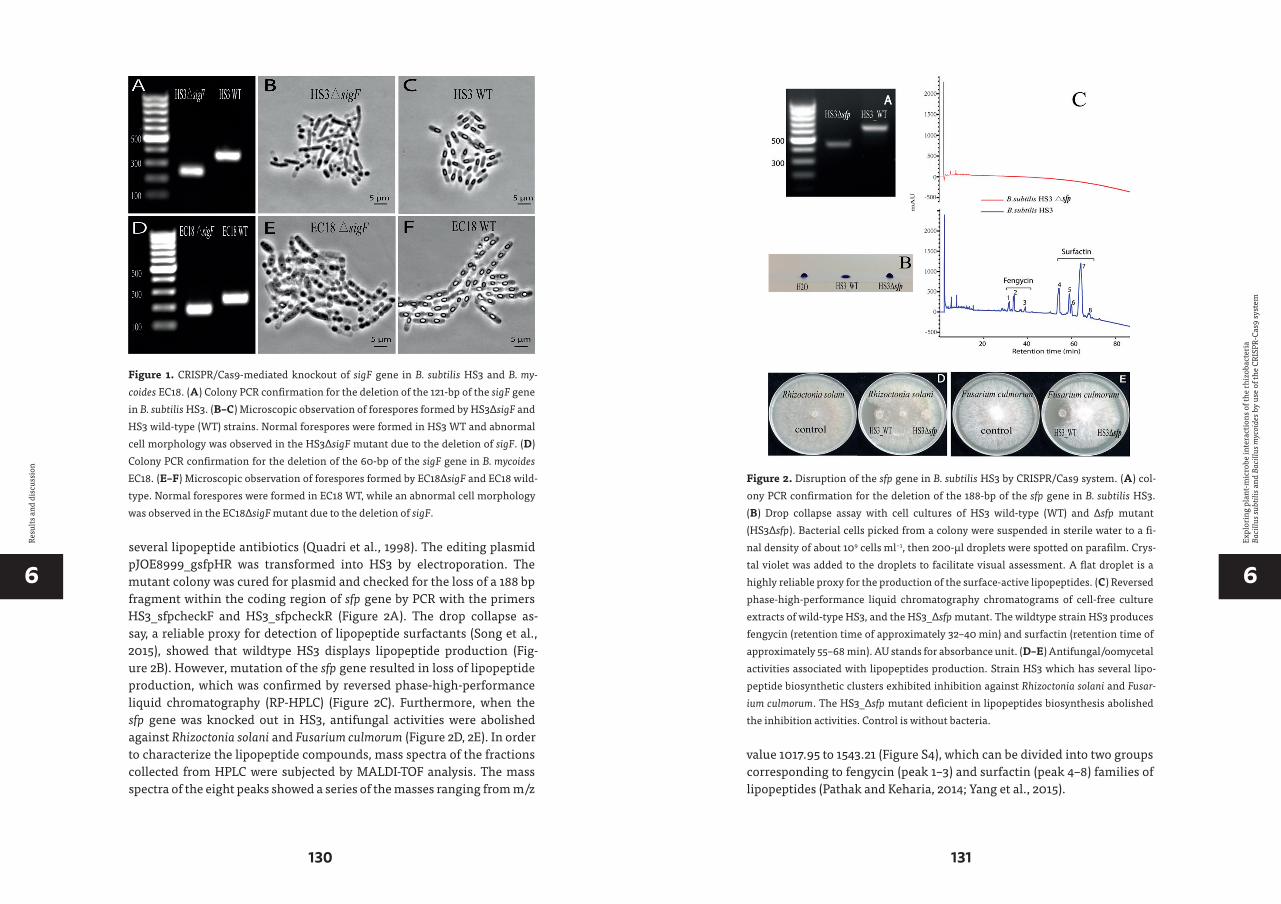
66
130 131
Expl
orin
g pl
ant-m
icro
be in
tera
ctio
ns o
f the
rhiz
obac
teri
a Ba
cillu
s sub
tilis
and
Bac
illus
myc
oide
s by
use
of th
e CR
ISPR
-Cas
9 sy
stem
Resu
lts a
nd d
iscu
ssio
n
several lipopeptide antibiotics (Quadri et al., 1998). The editing plasmid pJOE8999_gsfpHR was transformed into HS3 by electroporation. The mutant colony was cured for plasmid and checked for the loss of a 188 bp fragment within the coding region of sfp gene by PCR with the primers HS3_sfpcheckF and HS3_sfpcheckR (Figure 2A). The drop collapse as-say, a reliable proxy for detection of lipopeptide surfactants (Song et al., 2015), showed that wildtype HS3 displays lipopeptide production (Fig-ure 2B). However, mutation of the sfp gene resulted in loss of lipopeptide production, which was confirmed by reversed phase-high-performance liquid chromatography (RP-HPLC) (Figure 2C). Furthermore, when the sfp gene was knocked out in HS3, antifungal activities were abolished against Rhizoctonia solani and Fusarium culmorum (Figure 2D, 2E). In order to characterize the lipopeptide compounds, mass spectra of the fractions collected from HPLC were subjected by MALDI-TOF analysis. The mass spectra of the eight peaks showed a series of the masses ranging from m/z
Figure 1. CRISPR/Cas9-mediated knockout of sigF gene in B. subtilis HS3 and B. my-coides EC18. (A) Colony PCR confirmation for the deletion of the 121-bp of the sigF gene in B. subtilis HS3. (B–C) Microscopic observation of forespores formed by HS3∆sigF and HS3 wild-type (WT) strains. Normal forespores were formed in HS3 WT and abnormal cell morphology was observed in the HS3∆sigF mutant due to the deletion of sigF. (D) Colony PCR confirmation for the deletion of the 60-bp of the sigF gene in B. mycoides EC18. (E–F) Microscopic observation of forespores formed by EC18∆sigF and EC18 wild-type. Normal forespores were formed in EC18 WT, while an abnormal cell morphology was observed in the EC18∆sigF mutant due to the deletion of sigF.
value 1017.95 to 1543.21 (Figure S4), which can be divided into two groups corresponding to fengycin (peak 1–3) and surfactin (peak 4–8) families of lipopeptides (Pathak and Keharia, 2014; Yang et al., 2015).
Figure 2. Disruption of the sfp gene in B. subtilis HS3 by CRISPR/Cas9 system. (A) col-ony PCR confirmation for the deletion of the 188-bp of the sfp gene in B. subtilis HS3. (B) Drop collapse assay with cell cultures of HS3 wild-type (WT) and Δsfp mutant (HS3Δsfp). Bacterial cells picked from a colony were suspended in sterile water to a fi-nal density of about 109 cells ml−1, then 200-μl droplets were spotted on parafilm. Crys-tal violet was added to the droplets to facilitate visual assessment. A flat droplet is a highly reliable proxy for the production of the surface-active lipopeptides. (C) Reversed phase-high-performance liquid chromatography chromatograms of cell-free culture extracts of wild-type HS3, and the HS3_Δsfp mutant. The wildtype strain HS3 produces fengycin (retention time of approximately 32–40 min) and surfactin (retention time of approximately 55–68 min). AU stands for absorbance unit. (D–E) Antifungal/oomycetal activities associated with lipopeptides production. Strain HS3 which has several lipo-peptide biosynthetic clusters exhibited inhibition against Rhizoctonia solani and Fusar-ium culmorum. The HS3_Δsfp mutant deficient in lipopeptides biosynthesis abolished the inhibition activities. Control is without bacteria.

66
132 133
Expl
orin
g pl
ant-m
icro
be in
tera
ctio
ns o
f the
rhiz
obac
teri
a Ba
cillu
s sub
tilis
and
Bac
illus
myc
oide
s by
use
of th
e CR
ISPR
-Cas
9 sy
stem
Resu
lts a
nd d
iscu
ssio
n
Volatiles produced by B. subtilis HS3 promote grass growth2,3-butanediol is known to be a bacterial VOC that stimulate plant growth (Ryu et al., 2003). B. subtilis is capable of converting acetoin to 2,3-butane-diol by the enzyme 2,3-butanediol dehydrogenase (BDH), which is en-coded by the bdhA gene (Nicholson, 2008). The biosynthesis of acetoin in-volved the enzymes acetolactate synthase and acetolactate decarboxylase. The genes encoding these enzymes are alsS and alsD (Renna et al., 1993). In order to block the 2,3-butanediol production in B. subtilis HS3, we de-leted the bdhA and alsD gene by the editing vector pJOE8999_galsDHR and pJOE8999_gbdhHR, respectively. After transformation, the colonies on LB-Km plates were checked by colony PCR. The primers HS3_alsDcheckF and HS3_alsDcheckR were used for HS3_∆alsD mutant, and the primers HS3_bdhAcheck-F and HS3_bdhAcheck-R were used for HS3_∆bdhA mu-tant. As shown in Figure 3A, both mutants showed the expected smaller fragment size than the wild-type. In order to detect the 2,3-butanediol production, wild-type and mutant strains were grown in LB medium con-taining 2% glucose, and the culture supernatant was run on silica gel TLC. A standard 2,3-butanediol was included as a control. The TLC results showed that the HS3 wild-type strain produced 2,3-butanediol, while the two mutants abolished the production (Figure 3B). Grass seedlings (Lolium perenne) were exposed to VOCs produced by either wild-type HS3 or the mutants. Shoot and root biomass were increased approximately 2~3-fold compared to control plants for both wild-type HS3 and the mutants, and there was no significant difference between the wild-type strain and the mutants (Figure 3C&D). Collectively, these results indicate that B. subtilis HS3 may emit other VOCs than 2,3-butanediol to mediate the growth of plants, which was beyond the scope of this study, but worthwhile to be further investigated in future work.
B. subtilis HS3 colonization of grass root observed by CLSMThe colonization of roots by rhizobacteria is the initial step in the interac-tion between beneficial bacteria and the host plant (Ahmad et al., 2011). Investigating the colonization behavior and establishment of bacteria in the root system is of great ecological interest. Moreover, monitoring in-oculated bacteria is essential for the assessment of the effectiveness and consistent performance of inoculated PGPR. A prerequisite for such a study is the ability to track a specific microbe in the complex rhizosphere environment (Kluepfel, 1993). A classical approach is to introduce antibi-otic resistance to the bacterium, then plate and enumerate the introduced bacterium on selective medium. However, this method is indirect and has several drawbacks including marker instability and high background from antibiotic resistance microbial populations (Benizri et al., 2001).
In situ methods coupled with molecular techniques can better capture mi-crobial activity and interactions with the host (Nawy, 2016). Fluorescent protein (FP) labeling system for in situ studies allows direct visualization of the tagged bacteria at the single-cell level, without the addition of ex-ogenous substrates (Larrainzar et al., 2005). To study bacterial-host inter-actions, inserting the FP gene into bacterial chromosome provides an ad-vantageous approach, since plasmid-based FP labeling is mostly instable,
Figure 3. Disruption of the alsD and bhdA gene in B. subtilis HS3 by CRISPR/CAS9 sys-tem. (A) colony PCR confirmation for the deletion of the 249-bp of the alsD gene and 138-bp of the dhbA gene in B. subtilis HS3. (B) TLC detection of 2,3-butanediol from HS3 wild-type (WT), HS3_∆alsD, and HS3_∆bdhA. Standard, 2,3-butanediol was included as a positive control. (C–D) The biomass quantification and growth promotion of grass after 2 weeks of exposure to VOCs from HS3, HS3_∆alsD, and HS3_∆bdhA. The two mu-tants as well as the wild type HS3 increase the biomass of grass by emitting VOCs.

66
134 135
Expl
orin
g pl
ant-m
icro
be in
tera
ctio
ns o
f the
rhiz
obac
teri
a Ba
cillu
s sub
tilis
and
Bac
illus
myc
oide
s by
use
of th
e CR
ISPR
-Cas
9 sy
stem
Resu
lts a
nd d
iscu
ssio
n
and the fluorescence signal show heterogeneously among different cells (Rodriguez et al., 2006). The CRISPR/Cas9 system is a convenient and effi-cient method to perform marker-free genome insertions.
In a previous study, we selected an optimized gfp for rhizosphere Bacilli (Yi et al., 2018). The optimized gfp was introduced to the editing plasmid, pJOE8999_gamyGFP, which was transformed into HS3 by electroporation. The chromosomal GFP insertion was confirmed by colony PCR with the primers HS3_GFPchechF and HS3_GFPchechR. The anticipated fragment size of 3,399 bp was obtained (Figure 4 A). Fluorescent microscopy showed that HS3_GFP cells were expressing GFP homogeneously (Figure 4B). The GFP-labelled strain was inoculated with grass plants. After two and three
Figure 4. Insertion of gfp gene into the chromosome of B. subtilis HS3 by CRISPR/Cas9 system and the observation of GFP-tagged cells in the grass rhizosphere from a hydro-ponic system. (A) colony PCR confirmation for the insertion of the gfp gene into B. sub-tilis HS3 chromosome. (B) fluorescent microscopic observation of the GFP-tagged HS3 cells. The HS3 GFP cells showed homogeneously green auto-fluorescence. (C–H) In planta observation of B. subtilis HS3 cells in the rhizosphere of grass grown in a hydro-ponic system. Low amounts of colonization were found at the root tip, while strong col-onization was exclusively observed at root hairs.
days of inoculation, the intact grass roots were visualized by CLSM imme-diately after sampling. The gfp-tagged cells could be clearly detected on the root surface but not in all parts. No colonization was observed at the root tip (Figure 4C). Interestingly, low colonization was observed at the older basal root parts. But strong colonization was only found on the root hairs (Figure 4D-H). Similar bacteria colonization pattern was reported by Prieto et al. (2011) when Pseudomonas spp. was inoculated into olive roots.
Siderophores produced by B. mycoides promote the growth of iron-starved plants Siderophores are low-molecular-weight iron chelators produced by bac-teria and fungi under iron-limiting conditions and facilitate the solubi-lization and transport of iron into the cell by the cognate transport sys-tem (Schalk et al., 2011). Siderophores might also play an important role in plant-microbe interactions, since PGPR that produce siderophores combat the pathogenic microorganisms sequestering Fe3+ near the roots ( Shilev, 2013). Moreover, the bacterial siderophores are often used by plants as iron source contributing to plant nutrition. For example, sidero-phores from strain Chryseobacterium C138 are effective in supplying Fe to iron-starved tomato plants by the roots (Radzki et al., 2013). The genome comparison analysis revealed two gene clusters responsible for sidero-phore biosynthesis in B. mycoides EC18: the asb operon for petrobactin and the dhb operon for bacillibactin (Figure S5) and siderophore produc-tion was confirmed (Figure 5B). It has been shown that petrobactin was required for B. anthractis growth both in iron-depleted conditions and in macrophages (Cendrowski et al., 2004). The presence of petrobactin bio-synthesis gene (asbA) has been used as a marker gene for screening PGPR Bacillus (Lyngwi et al., 2016).
In order to determine whether if petrobactin and bacillibactin are in-volved in the B. mycoides-plant interaction, we generated mutants of these two operons. The 20-nt spacer sequence was designed within the open reading frame of asbA and dhbB gene (Figure S5). The spacer se-quences as well as 1 kb flanking the up- and downstream region were in-troduced into pYCR to generate the editing plasmid pYCR_gasbHR and pYCR_gdhbHR. The editing plasmids were transformed into B. mycoides EC18, and the transformants on LB-SPEC100 plates were checked by col-ony PCR to assess the successful knockout. The colonies with expected knockout size were cured from the plasmid, which resulted in the clean knockout strain B. mycoides EC18ΔasbA and B. mycoides EC18ΔdhbB. To make the asbA and dbhB double knockout strain B. mycoides ADKO, the plasmid pYCR_gdhbHR was transformed into the electro-competent cell of EC18ΔasbA, and the transformants were selected and confirmed as

66
136 137
Expl
orin
g pl
ant-m
icro
be in
tera
ctio
ns o
f the
rhiz
obac
teri
a Ba
cillu
s sub
tilis
and
Bac
illus
myc
oide
s by
use
of th
e CR
ISPR
-Cas
9 sy
stem
Resu
lts a
nd d
iscu
ssio
n
before. Colony PCR for ∆asbA, ∆dhbB, and the double knockout mutant ADKO showed that all the mutants have expected smaller fragment sizes compared with the wild-type strain: 463 bp for EC18ΔasbA and 650 for EC18 wild type using primers EC18_asb_checkF and EC18_asb_checkR, and 184 bp for EC18ΔdhbB and 299 bp for EC18 wild type using primers EC18_dhb_checkF and EC18_dhb_checkR (Figure 5A). The siderophore production of all the mutants and the wild type were inspected by O-CAS agar (Figure 5B). The production of siderophores by EC18ΔasbA is compa-rable to that of wild-type. In contrast, the EC18ΔdhbB had little production, and no siderophore production was detected from the double knockout EC18 ADKO. In the hydroponic system, the wild-type EC18 and the mutant EC18ΔasbA increased the total chlorophyll content of Chinese cabbage sig-nificantly. There were no significant differences of the total chlorophyll of the plants treated with EC18ΔdhbB and EC18 ADKO when compared to the control group (Figure 5C). Biomass was also increased in the plants treated with wild type EC18 and EC18ΔasbA, with no significant difference between them. On the other hand, ∆dhbB mutant and double mutant ∆ADKO abol-ished the plant growth promoting effect completely (Figure 5D). These re-sults indicate that bacillibactin is the main siderophore produced by B. my-coides and contributes to the plant growth promoting effects.
Siderophore-deficient strains show reduced plant colonization abilitySince the strain EC18ΔdhbB and EC18ADKO were unable to promote plant growth, we further compared their plant colonization capacity with the wild-type strain. The gfp gene with a strong constitutive promoter Ppta was inserted into the chromosome of the strain EC18, EC18ΔasbA, EC18ΔdhbA, and EC18ADKO to generate the GFP-tagged strain EC18GFP, EC18ΔAGFP, EC18ΔDGFP, and EC18ADGFP. All the strains were checked by PCR with the primers EC18_GFP_gcheckF and EC18_GFP_gcheckR binding the flanking region of the spacer sequence in the chromosome. The antic-ipated 3,434 bp amplicon size (Figure 6A) and homogeneous fluorescence of the cells (Figure 6B) indicate the gfp insertion was successful and func-tional. The GFP-tagged strains were inoculated with the Chinese cabbage roots in a hydroponic system. After 2 days, the plant roots were collected and directly observed by CLSM. The wild-type strain colonizes all plant root regions in a heterogeneous manner: a few cells colonize at the root tip, while most cells colonize the root hair initiation region, and a large number of cells aggregated at the junction region of main root and lat-eral root (Figure 6C). The petropectin mutant strain EC18ΔAGFP showed less colonization on the root tip and root hair region, compared to the wild-type control, while the lateral root emergence site was colonized
by large bacterial populations (Figure 6D). For the bacillibactin mutant EC18ΔDGFP and the double knockout strain EC18ADGFP, the colonization of root tip and root hair regions was only occasionally observed. The cells of the two strains were arranged in microcolonies at the junctions of lat-eral roots with the primary roots (Figure 6E &F).
In conclusion, we implemented the CRISPR/Cas9 system to perform gene knockout and chromosomal insertion in environmental B. subti-lis HS3 and B. mycoides EC18 strains. The high efficiency in the two phy-logenetically distant Bacillus species implies the great potential of the CRISPR/Cas9 system in genome editing of rhizosphere Bacilli. The gen-erated mutants were used to study various traits in plant-microbe inter-actions. By interrupting the sfp gene in B. subtilis HS3, we demonstrated that the surfactin and fengycin family lipopeptides are responsible for the antagonistic activity against two relevant fungal pathogens. Moreo-ver, we revealed that 2,3-butanediol is not the main VOC produced by HS3
Figure 5. Disruption of siderophore biosynthesis genes of B. mycoides EC18 by the CRISPR/Cas9 system. (A) colony PCR confirmation for the deletion of the 187-bp of asbA gene (ΔasbA), 115-bp of the dhbB gene (ΔdhbB), and the double deletion of these two genes in B. mycoides EC18 (ADKO). (B) Siderophore detection by the O-CAS method. The yellowish halo indicates siderophore production. (C) The total chlorophyll content of Chinese cabbage treated with B. mycoides EC18 wild-type and siderophore-deficient mutants, EC18ΔasbA, EC18ΔdhbB, and the double mutant EC18ADKO. (D) The biomass quantification of Chinese cabbage treated with B. mycoides EC18 wild-type and sidero-phore-deficient mutants.

66
138 139
Expl
orin
g pl
ant-m
icro
be in
tera
ctio
ns o
f the
rhiz
obac
teri
a Ba
cillu
s sub
tilis
and
Bac
illus
myc
oide
s by
use
of th
e CR
ISPR
-Cas
9 sy
stem
Resu
lts a
nd d
iscu
ssio
n
Figure 6. Insertion of GFP into the chromosome of B. mycoides EC18 by CRISPR/Cas9 system and the observation of GFP-tagged cells in the rhizosphere of Chinese cabbage from a hydroponic system. (A) colony PCR confirmation for the insertion of the gfp gene in the chromosome B. mycoides EC18 and mutants. (B) fluorescent microscopic observa-tion of the GFP-tagged B. mycoides wild-type and mutants cells. The EC18 WT as well as
to promote grass growth. By using a single-copy GFP-tagged mutant, we showed that HS3 selectively colonizes on root hairs. In B. mycoides EC18, by interrupting two siderophore biosynthesis gene via our CRISPR/Cas9 system, we highlighted the important role of petrobactin, in the plant growth promoting and root colonization.
ACKNOWLEDGEMENTS
We thank Dr. Auke van Heel in our group for his suggestions and com-ments on the manuscript. Y. Yi and Z. Li were financially supported by the Chinese Scholarship Council (CSC). The authors declare no conflict of interest.
all the mutants’ cells showed homogeneously green auto-fluorescence. (C–F) In planta observation of B. mycoides wild-type and mutant cells colonized in the rhizosphere of Chinese cabbage grown in a hydroponic system.

66
140 141
Expl
orin
g pl
ant-m
icro
be in
tera
ctio
ns o
f the
rhiz
obac
teri
a Ba
cillu
s sub
tilis
and
Bac
illus
myc
oide
s by
use
of th
e CR
ISPR
-Cas
9 sy
stem
Supp
lem
enta
ry ta
ble
and
figur
es:
SUPPLEMENTARY TABLE AND FIGURES:
Tabl
e S1
. Pri
mer
s use
d in
this
stud
y.
Olig
onuc
leot
ides
Sequ
ence
Feat
ure
and
purp
ose
HS3
_gsig
F_F
AAAC
ACAG
TGCA
GGAG
ATCG
CTGA
Prot
ospa
cer f
or ta
rget
ing s
igF
gene
of B
.subt
ilis H
S3H
S3_g
sigF_
RTA
CGTC
AGCG
ATCT
CCTG
CACT
GTPr
otos
pace
r for
targ
etin
g sig
F ge
ne o
f B.su
btili
s HS3
HS3
_sig
FHR1
_FAA
GG
CCAA
CGAG
GCC
GTGA
CTCA
ATCA
TTGG
AGAA
GGam
plify
1kb
fragm
ent u
pstr
eam
of s
igF
spac
er se
quen
ce o
f B.su
btili
s HS3
, with
sfiI s
iteH
S3_s
igFH
R1_R
CAGT
ACAA
CATT
ACCT
TTAC
GGTT
CCGT
CATC
ampl
ify 1k
b fra
gmen
t ups
trea
m o
f sig
F sp
acer
sequ
ence
of B
.subt
ilis H
S3H
S3_s
igFH
R2_F
CCGT
AAAG
GTAA
TGTT
GTAC
TGGC
CCAA
GAG
ampl
ify 1k
b fra
gmen
t dow
nstr
eam
of s
igF
spac
er se
quen
ce o
f B.su
btili
s HS3
HS3
_sig
FHR2
_RAA
GG
CCTT
ATTG
GCC
CATG
GCCA
CATA
TTGA
TCGA
G
ampl
ify 1k
b fra
gmen
t dow
nstr
eam
of s
igF
spac
er se
quen
ce o
f B.su
btili
s HS3
, with
sfiI s
iteH
S3_s
igFc
heck
FAA
ACAC
AGTG
CAGG
AGAT
CGCT
GAch
eck
sigF
mut
ant o
f B.su
btili
s HS3
HS3
_sig
Fche
ckR
TACG
TCAG
CGAT
CTCC
TGCA
CTGT
chec
k sig
F m
utan
t of B
.subt
ilis H
S3H
S3_g
bdhA
-FAA
ACCG
TCGT
AGTC
GAGC
CGAT
TTPr
otos
pace
r for
targ
etin
g bdh
gene
of B
.subt
ilis H
S3H
S3_g
bdhA
-RTA
CGAA
ATCG
GCTC
GACT
ACGA
CGPr
otos
pace
r for
targ
etin
g bdh
gene
of B
.subt
ilis H
S3H
S3_b
dhA-
HR1
-FAA
GG
CCTT
ATTG
GCC
CGAA
TGGC
GTGA
TTTC
GGam
plify
1kb
fragm
ent u
pstr
eam
of b
dh sp
acer
sequ
ence
of B
.subt
ilis H
S3, w
ith sfi
I site
HS3
_bdh
A-H
R1-R
CGGT
GAAG
TCGG
CGGT
TTCT
CTGA
ATAC
ampl
ify 1k
b fra
gmen
t ups
trea
m o
f bdh
spac
er se
quen
ce o
f B.su
btili
s HS3
HS3
_bdh
A-H
R2-F
AACC
GCCG
ACTT
CACC
GGAG
AATT
CATG
Cam
plify
1kb
fragm
ent d
owns
trea
m o
f bdh
spac
er se
quen
ce o
f B.su
btili
s HS3
HS3
_bdh
A-H
R2-R
AAG
GCC
AACG
AGG
CCCC
CCGC
TACT
TTAC
CTTT
GCam
plify
1kb
fragm
ent d
owns
trea
m o
f bdh
spac
er se
quen
ce o
f B.su
btili
s HS3
, with
sfiI s
iteH
S3_b
dhAc
heck
-FGC
CTTC
AGCG
CTTC
AATG
chec
k bd
hA m
utan
t of B
.subt
ilis H
S3H
S3_b
dhAc
heck
-RGG
CAGC
AAGA
TGGC
ATAA
Cch
eck
bdhA
mut
ant o
f B.su
btili
s HS3
HS3
_gsf
p-F
AAAC
CTGG
GTCA
TTGG
TGCG
TTTG
Prot
ospa
cer f
or ta
rget
ing s
fp ge
ne o
f B.su
btili
s HS3
HS3
_gsf
p-R
TACG
CAAA
CGCA
CCAA
TGAC
CCAG
Prot
ospa
cer f
or ta
rget
ing s
fp ge
ne o
f B.su
btili
s HS3
HS3
_sfp
-HR1
FAA
GG
CCTT
ATTG
GCC
CCGA
GGTT
GAGC
GGTT
ATTT
Cam
plify
1kb
fragm
ent u
pstr
eam
of s
fpsp
acer
seq
uenc
e o
f B.su
btili
s HS3
, with
sfiI s
iteH
S3-s
fp-H
R1R
CTTT
AGCA
CACA
AGAC
AAGG
ACGA
GCAG
ACam
plify
1kb
fragm
ent u
pstr
eam
of s
fpsp
acer
sequ
ence
of B
.subt
ilis H
S3H
S3-s
fp-H
R2F
GTCC
TTGT
CTTG
TGTG
CTAA
AGCG
GATA
TCG
ampl
ify 1k
b fra
gmen
t dow
nstr
eam
of s
fpsp
acer
sequ
ence
of B
.subt
ilis H
S3H
S3-s
fp-H
R2R
AAG
GCC
AACG
AGG
CCGT
TCTG
CGCT
GGAC
ATTT
ampl
ify 1k
b fra
gmen
t dow
nstr
eam
of s
fpsp
acer
sequ
ence
of B
.subt
ilis H
S3, w
ith sfi
I site
HS3
_sfp
chec
kFCA
TTTT
GTAG
CCGG
GATC
Gch
eck
sfp
mut
ant o
f B.su
btili
s HS3
HS3
_sfp
chec
kRGC
AGAC
GGAG
GATC
TAGA
ATch
eck
sfp
mut
ant o
f B.su
btili
s HS3
HS3
-gal
sD-F
AAAC
TTCT
GTTC
ATTT
ACGT
TCTT
Prot
ospa
cer f
or ta
rget
ing a
lsD ge
ne o
f B.su
btili
s HS3
HS3
-gal
sD-R
TACG
AAGA
ACGT
AAAT
GAAC
AGAA
Prot
ospa
cer f
or ta
rget
ing a
lsD ge
ne o
f B.su
btili
s HS3
HS3
-als
D-H
R1-F
AAG
GCC
TTAT
TGG
CCCT
GGAT
GTAT
TCGG
CTAA
CCam
plify
1kb
fragm
ent u
pstr
eam
of a
lsD sp
acer
sequ
ence
of B
.subt
ilis H
S3, w
ith sfi
I site
HS3
-als
D-H
R1-R
GGGT
TTGA
CTAC
GTGC
CAAT
GGTT
GAAG
ampl
ify 1k
b fra
gmen
t ups
trea
m o
f alsD
spac
er se
quen
ce o
f B.su
btili
s HS3
HS3
-als
D-H
R2-F
TGGC
ACGT
AGTC
AAAC
CCAA
TCAG
CTCT
Cam
plify
1kb
fragm
ent d
owns
trea
m o
f alsD
spac
er se
quen
ce o
f B.su
btili
s HS3
HS3
-als
D-H
R2-R
AAG
GCC
AACG
AGG
CCTG
CTTA
CCAG
CCTG
ATCT
TGam
plify
1kb
fragm
ent d
owns
trea
m o
f alsD
spac
er se
quen
ce o
f B.su
btili
s HS3
, with
sfiI s
iteH
S3_a
lsDc
heck
FAT
CGTA
ACCG
TGCA
ATCC
chec
k alsD
mut
ant o
f B.su
btili
s HS3
HS3
_als
Dche
ckR
TAGC
ACTC
TGCG
CATC
ACch
eck a
lsD m
utan
t of B
.subt
ilis H
S3H
S3_g
amy_
FTA
CGAG
AAAG
GGCT
TTGA
ATGA
CGPr
otos
pace
r for
targ
etin
g α-a
myl
ase
gene
of B
.subt
ilis H
S3H
S3_g
amy_
RAA
ACCG
TCAT
TCAA
AGCC
CTTT
CTPr
otos
pace
r for
targ
etin
g α-a
myl
ase
gene
of B
.subt
ilis H
S3H
S3_a
my1
F_sfi
IAA
GG
CCAA
CGAG
GCC
CCGA
GAAT
AGAC
ACCA
AAGA
AGam
plify
~2k
b fra
gmen
t flan
king
the
α-am
ylas
espa
cer s
eque
nce
of B
.subt
ilis H
S3, w
ith sfi
I site
HS3
_am
y2R_
sfiI
AAG
GCC
TTAT
TGG
CCAA
GCAT
TTAC
CTGG
CTCC
AAam
plify
~2k
b fra
gmen
t flan
king
the
α-am
ylas
espa
cer s
eque
nce
of B
.subt
ilis H
S3, w
ith sfi
I site
HS3
_am
y1R
TTCG
AGCG
TTGA
ATTC
TGCG
TGAC
ATCC
Cam
plify
pla
smid
bac
kbon
e of
pJO
E899
9_ga
myF
LH
S3_a
my2
FAG
CAAG
ATTC
AAGC
TTAG
TTAC
GGCA
GTCA
ATTT
TGG
ampl
ify p
lasm
id b
ackb
one
of p
JOE8
999_
gam
yFL
Olig
onuc
leot
ides
Sequ
ence
Feat
ure
and
purp
ose
HS3
_GFP
chec
hFGT
TTAA
AGAA
TGGG
CTGC
AAG
chec
k gf
p in
sert
ion
of B
.subt
ilis H
S3H
S3_G
FPch
echR
GCGT
CAGC
GTGT
AAAT
TCC
chec
k gf
p in
sert
ion
of B
.subt
ilis H
S3PW
V01_
F AA
TTCG
CATC
ACAC
GGAC
TCTA
GAGG
ATCA
TTTT
GTTT
ATTG
ampl
ify P
WVO
1 ori
from
pAW
068
PWV0
1_R
AGAC
ACCT
AAAT
TCAG
CAAT
CGGA
TGCG
ATTA
TTG
ampl
ify P
WVO
1 ori
from
pAW
068
pJO
E-PE
T194
_FTG
AATT
TAGG
TGTC
TGCA
GTTG
am
plify
PJO
E899
9 ba
ckbo
ne w
ithou
t PET
194
ori
pJO
E-PE
T194
_RCG
TGTG
ATGC
GAAT
TCTT
GA
ampl
ify P
JOE8
999
back
bone
with
out P
ET19
4 or
iSp
ecF
GAGC
CTG
CAG
GTAA
CGTG
ACTG
GCAA
GAG
ampl
ify sp
ectin
omyc
ine
resis
tenc
e ge
ne fr
om P
DR11
1 pla
smid
,with
Pst
I site
Spec
RGA
GCCT
CGAG
GAAA
AAAG
TGTT
TCCA
CCAT
TTTT
TCam
plify
spec
tinom
ycin
e re
siste
nce
gene
from
PDR
111 p
lasm
id,w
ith X
hoI s
iteEC
18_g
sigF_
FTA
CGAA
GGGC
TCCA
ACGA
TTAA
TGPr
otos
pace
r for
targ
etin
g sig
F ge
ne o
f B.m
ycoi
des E
C18
EC18
_gsig
F_R
AAAC
CATT
AATC
GTTG
GAGC
CCTT
Prot
ospa
cer f
or ta
rget
ing s
igF
gene
of B
.myc
oide
s EC1
8EC
18_s
igFH
R1_F
AAG
GCC
AACG
AGG
CCTG
GTTT
AGGC
GTTA
TATT
AGGC
ampl
ify 1k
b fra
gmen
t ups
trea
m o
f sig
F sp
acer
sequ
ence
of B
.myc
oide
EC1
8, w
ith sfi
I site
EC18
_sig
FHR1
_RAG
AGCC
TGTT
CTAG
ACGG
ACTT
TGTT
TCCT
GTTT
Cam
plify
1kb
fragm
ent u
pstr
eam
of s
igF
spac
er se
quen
ce o
f B.m
ycoi
de E
C18
EC18
_sig
FHR2
_FCA
AAGT
CCGT
CTAG
AACA
GGCT
CTTG
AACT
AACG
ampl
ify 1k
b fra
gmen
t dow
nstr
eam
of s
igF
spac
er se
quen
ce o
f B.m
ycoi
de E
C18
EC18
_sig
FHR2
_RAA
GG
CCTT
ATTG
GCC
ACAA
CCAT
ACAA
GTCC
GAAG
ampl
ify 1k
b fra
gmen
t dow
nstr
eam
of s
igF
spac
er se
quen
ce o
f B.m
ycoi
de E
C18,
with
sfiI s
iteEC
18_s
igFc
heck
FGT
GACG
ATGG
ATCA
GTGA
AAG
ch
eck
sigF
mut
ant o
f B.m
ycoi
des E
C18
EC18
_sig
Fche
ckR
TAAT
TGGA
TCTC
CGTC
GTTT
TCch
eck
sigF
mut
ant o
f B.m
ycoi
des E
C18
EC18
_gas
b_F
TACG
AAGT
GGAG
AATG
GATT
ACGG
Prot
ospa
cer f
or ta
rget
ing a
sbA
gene
of B
.myc
oide
s EC1
8EC
18_g
asb_
RAA
ACCC
GTAA
TCCA
TTCT
CCAC
TTPr
otos
pace
r for
targ
etin
g asb
A ge
ne o
f B.m
ycoi
des E
C18
EC18
_asb
HR1
_FAA
GG
CCAA
CGAG
GCC
CATC
TCAG
CAAT
TGGA
AATG
Gam
plify
1kb
fragm
ent u
pstr
eam
of s
igF
spac
er se
quen
ce o
f B.m
ycoi
de E
C18,
with
sfiI s
iteEC
18_a
sbH
R1_R
ACGG
TCCG
TTCA
TGTT
CCGC
GATT
TGTT
TC
am
plify
1kb
fragm
ent u
pstr
eam
of s
igF
spac
er se
quen
ce o
f B.m
ycoi
de E
C18
EC18
_asb
HR2
_FCG
GAAC
ATGA
ACGG
ACCG
TCAT
CTAT
TTG
ampl
ify 1k
b fra
gmen
t dow
nstr
eam
of s
igF
spac
er se
quen
ce o
f B.m
ycoi
de E
C18,
with
sfiI s
iteEC
18_a
sbH
R2_R
AAG
GCC
TTAT
TGG
CCCT
AAAT
CCGC
TAAG
CGAT
GG
ampl
ify 1k
b fra
gmen
t dow
nstr
eam
of s
igF
spac
er se
quen
ce o
f B.m
ycoi
de E
C18,
with
sfiI s
iteEC
18_a
sb_c
heck
FGC
TTGG
CGAG
ATAT
GGAT
TGch
eck
asbA
mut
ant o
f B.m
ycoi
des E
C18
EC18
_asb
_che
ckR
CAGG
ATTC
GTAC
CTTC
TCCA
TAG
chec
k as
bA m
utan
t of B
.myc
oide
s EC1
8EC
18_g
dhb_
FTA
CGCA
GCAA
CCTT
CTCA
CCAC
CAPr
otos
pace
r for
targ
etin
g dhb
A ge
ne o
f B.m
ycoi
des E
C18
EC18
_gdh
b_R
AAAC
TGGT
GGTG
AGAA
GGTT
GCTG
Prot
ospa
cer f
or ta
rget
ing d
hbA
gene
of B
.myc
oide
s EC1
8EC
18_d
hbH
R1_F
AA
GG
CCAA
CGAG
GCC
AGCA
ATGA
TTTG
GCTT
GATG
ampl
ify 1k
b fra
gmen
t ups
trea
m o
f dhb
A sp
acer
sequ
ence
of B
.myc
oide
EC1
8, w
ith sfi
I site
EC18
_dhb
HR1
_RGT
AAGC
TCAT
CGCA
AGTT
CCTT
GCAT
CTTT
CTC
am
plify
1kb
fragm
ent u
pstr
eam
of d
hbA
spac
er se
quen
ce o
f B.m
ycoi
de E
C18
EC18
_dhb
HR2
_F
CAAG
GAAC
TTGC
GATG
AGCT
TACT
CCAG
ATG
am
plify
1kb
fragm
ent d
owns
trea
m o
f dhb
A sp
acer
sequ
ence
of B
.myc
oide
EC1
8EC
18_d
hbH
R2_R
AAG
GCC
TTAT
TGG
CCAA
TCCA
TAGC
CATC
CATC
ACam
plify
1kb
fragm
ent d
owns
trea
m o
f dhb
A sp
acer
sequ
ence
of B
.myc
oide
EC1
8, w
ith sfi
I site
EC18
_dhb
_che
ckF
AAAC
GTGC
GGTT
CTTC
TAAT
Cch
eck
dhbA
mut
ant o
f B.m
ycoi
des E
C18
EC18
_dhb
_Che
ckR
GCAC
TATA
TCTC
CATT
TGGT
TAGG
chec
k dh
bA m
utan
t of B
.myc
oide
s EC1
8EC
18_g
amy_
FTA
CGAT
CATG
CCAG
ACAG
CAAA
CGPr
otos
pace
r for
targ
etin
g α-a
myl
ase
gene
of B
.myc
oide
EC1
8EC
18_g
amy_
RAA
ACCG
TTTG
CTGT
CTGG
CATG
ATPr
otos
pace
r for
targ
etin
g α-a
myl
ase
gene
of B
.myc
oide
EC1
8EC
18_a
my1
F_sfi
IAA
GG
CCTT
ATTG
GCC
ACTC
ATTA
GATT
CCAA
TGAG
TTCT
ACam
plify
~2k
b fra
gmen
t flan
king
the
α-am
ylas
espa
cer s
eque
nce
of B
.myc
oide
EC1
8, w
ith sfi
I site
EC18
_am
y2R_
sfiI
AAG
GCC
AACG
AGG
CCCA
TAGA
AATC
CCTC
CGTT
ATGC
am
plify
~2k
b fra
gmen
t flan
king
the
α-am
ylas
espa
cer s
eque
nce
of B
.myc
oide
EC1
8, w
ith sfi
I site
EC18
_am
y1R
AGCA
AGAT
TCAA
GCTA
TGAA
AAAT
CACC
CTGC
ACTC
GCam
plify
pla
smid
bac
kbon
e of
pYC
R_ga
myF
LEC
18_a
my2
FTT
CGAG
CGTT
GAAT
TGTC
CAAA
CGAA
AGCC
ATCT
AAA
ampl
ify p
lasm
id b
ackb
one
of p
YCR_
gam
yFL
EC18
_GFP
_gch
eckF
ACTA
TAGC
TCGT
GCTT
CTGT
C
chec
k gf
p in
sert
ion
of B
.myc
oide
s EC1
8EC
18_G
FP_g
chec
kRGC
CATT
GCGT
ACAT
AATG
TTAG
Gch
eck
gfp
inse
rtio
n of
B.m
ycoi
des E
C18
ITA
LIC
BOLD
: ste
aky
end
afte
r olig
o an
neal
ing;
UN
DER
LIN
E B
OLD
: res
tric
tion
enzy
me

66
142 143
Expl
orin
g pl
ant-m
icro
be in
tera
ctio
ns o
f the
rhiz
obac
teri
a Ba
cillu
s sub
tilis
and
Bac
illus
myc
oide
s by
use
of th
e CR
ISPR
-Cas
9 sy
stem
Supp
lem
enta
ry ta
ble
and
figur
es:
Figure S1. Construction scheme of the pYCR plasmid. The construction was started from pJOE8999 by replacing the pE194ts origin of replication (ori) with the pWVO1 ori. Then the kanamycin resistance gene was replaced by spectinomycin resistance gene.
Figure S2. Colony PCR results of the ΔsigF knockout in the two stains. (A) B. subtilis HS3 transformed with pJOE8999_gsigFHR plasmid and (B) B. mycoides EC18 transformed with pYCR_gsigFHR plasmid were grown on LB agar plate supplemented with 0.2% mannose and appropriate antibiotics. Colonies were subjected for PCR with the primers listed in Table S1. A smaller amplicon indicates the correct knockout.
Figure S3. B. mycoides EC18 colonies after being transformed with (A) pYCR and (B) pYCR_gsigFHR. The transformed EC18 cells were plated on LB plates contain-ing 100 µg ml−1 spectinomycin and 2% mannose. The plates were incubated at 30 °C overnight.

66
144 145
Expl
orin
g pl
ant-m
icro
be in
tera
ctio
ns o
f the
rhiz
obac
teri
a Ba
cillu
s sub
tilis
and
Bac
illus
myc
oide
s by
use
of th
e CR
ISPR
-Cas
9 sy
stem
Supp
lem
enta
ry ta
ble
and
figur
es:
Figure S4. MALDI-TOF of the peaks collected from the crude extracts of B. subtilis HS3 culture. The main mass numbers of m/z were indicated.
Figu
re S
5. C
ompa
riso
n of
the
pect
roba
ctin
and
bac
illib
actin
bi
osyn
thes
is g
ene
clus
ter o
f B.
myc
oide
s EC1
8 w
ith cl
ose
re-
late
d sp
ecie
s. T
he d
esig
n of
the
20-n
t spa
cer a
nd h
omol
ogou
s re
pair
ing
frag
men
t is s
how
n.

66
146 147
Expl
orin
g pl
ant-m
icro
be in
tera
ctio
ns o
f the
rhiz
obac
teri
a Ba
cillu
s sub
tilis
and
Bac
illus
myc
oide
s by
use
of th
e CR
ISPR
-Cas
9 sy
stem
Refe
renc
es:
REFERENCES:
Adesemoye, A.O., and Kloepper, J.W. (2009)
Plant-microbes interactions in enhanced
fertilizer-use efficiency. Appl. Microbiol.
Biotechnol. 85: 1–12.
Ahmad, F., Husain, F.M., and Ahmad, I. (2011)
Rhizosphere and root colonization by
bacterial inoculants and their monitor-
ing methods: a critical area in PGPR re-
search. In Microbes and Microbial technol-
ogy: Springer, pp. 363–391.
Allard-Massicotte, R., Tessier, L., Lecuyer, F.,
Lakshmanan, V., Lucier, J.F., Garneau, D.
et al. (2016) Bacillus subtilis Early Coloniza-
tion of Arabidopsis thaliana Roots Involves
Multiple Chemotaxis Receptors. mBio 7:
e01664–16.
Altenbuchner, J. (2016) Editing of the Bacillus
subtilis Genome by the CRISPR-Cas9 Sys-
tem. Appl. Environ. Microbiol. 82: 5421–5427.
Ambrosini, A., Stefanski, T., Lisboa, B.B., Bene-
duzi, A., Vargas, L.K., and Passaglia, L.M.P.
(2016) Diazotrophic bacilli isolated from
the sunflower rhizosphere and the poten-
tial of Bacillus mycoides B38V as biofer-
tiliser. Ann. Appl. Biol. 168: 93–110.
Arnon, D.I. (1949) Copper enzymes in isolated
chloroplasts. Polyphenoloxidase in Beta
vulgaris. Plant Physiol. 24: 1.
Bargabus, R., Zidack, N., Sherwood, J., and Ja-
cobsen, B. (2002) Characterisation of sys-
temic resistance in sugar beet elicited by a
non-pathogenic, phyllosphere-colonizing
Bacillus mycoides, biological control agent.
Physiol. Mol. Plant Pathol. 61: 289–298.
Bargabus, R.L., Zidack, N.K., Sherwood, J.E., and
Jacobsen, B.J. (2003) Oxidative burst elic-
ited by Bacillus mycoides isolate Bac J, a bio-
logical control agent, occurs independently
of hypersensitive cell death in sugar beet.
Mol. Plant Microbe Interact. 16: 1145–1153.
Barrangou, R., Fremaux, C., Deveau, H., Rich-
ards, M., Boyaval, P., Moineau, S. et al.
(2007) CRISPR provides acquired resis-
tance against viruses in prokaryotes. Sci-
ence 315: 1709–1712.
Beauregard, P.B., Chai, Y., Vlamakis, H., Los-
ick, R., and Kolter, R. (2013) Bacillus sub-
tilis biofilm induction by plant polysac-
charides. Proc. Natl. Acad. Sci. U. S. A. 110:
E1621–1630.
Benizri, E., Baudoin, E., and Guckert, A. (2001)
Root Colonization by Inoculated Plant
Growth-Promoting Rhizobacteria. Biocon-
trol Sci. Technol. 11: 557–574.
Cendrowski, S., MacArthur, W., and Hanna, P.
(2004) Bacillus anthracis requires sidero-
phore biosynthesis for growth in macro-
phages and mouse virulence. Mol. Micro-
biol. 51: 407–417.
Dong, H., and Zhang, D. (2014) Current devel-
opment in genetic engineering strategies
of Bacillus species. Microb. Cell Fact 13: 63.
Green, B., Bouchier, C., Fairhead, C., Craig,
N.L., and Cormack, B.P. (2012) Insertion
site preference of Mu, Tn5, and Tn7 trans-
posons. Mobile DNA 3: 3.
Guetsky, R., Elad, Y., Shtienberg, D., and Dinoor,
A. (2002) Improved Biocontrol of Botrytis
cinerea on Detached Strawberry Leaves by
Adding Nutritional Supplements to a Mix-
ture of Pichia guilermondii and Bacillus my-
coides. Biocontrol Sci. Technol. 12: 625–630.
Hilbert, D.W., and Piggot, P.J. (2004) Compart-
mentalization of gene expression during
Bacillus subtilis spore formation. Microbiol.
Mol. Biol. Rev. 68: 234–262.
Jiang, W., Bikard, D., Cox, D., Zhang, F., and Mar-
raffini, L.A. (2013) RNA-guided editing of
bacterial genomes using CRISPR-Cas sys-
tems. Nat. Biotechnol. 31: 233–239.
Jinek, M., Chylinski, K., Fonfara, I., Hauer, M.,
Doudna, J.A., and Charpentier, E. (2012)
A programmable dual-RNA–guided DNA
endonuclease in adaptive bacterial immu-
nity. Science 337: 816–821.
Kluepfel, D.A. (1993) The behavior and track-
ing of bacteria in the rhizosphere. Annu
Rev Phytopathol 31: 441–472.
Kumar, A., Prakash, A., and Johri, B. (2011) Ba-
cillus as PGPR in crop ecosystem. In Bacte-
ria in agrobiology: crop ecosystems: Springer,
pp. 37–59.
Larrainzar, E., O’Gara, F., and Morrissey, J.P.
(2005) Applications of Autofluorescent
Proteins for In Situ studies in Microbial
Ecology. Annu. Rev. Microbiol. 59: 257–277.
Leloup, L., Ehrlich, S.D., Zagorec, M., and
Morel- Deville, F. (1997) Single-crossover
integration in the Lactobacillus sake chro-
mosome and insertional inactivation of
the ptsI and lacL genes. Appl. Environ. Mi-
crobiol. 63: 2117–2123.
Li, Y., Lin, Z., Huang, C., Zhang, Y., Wang, Z., Tang,
Y.-j. et al. (2015) Metabolic engineering of
Escherichia coli using CRISPR–Cas9 medi-
tated genome editing. Metab. Eng. 31: 13–21.
Lyngwi, N.A., Nongkhlaw, M., Kalita, D., and
Joshi, S.R. (2016) Bioprospecting of Plant
Growth Promoting Bacilli and Related
Genera Prevalent in Soils of Pristine Sa-
cred Groves: Biochemical and Molecular
Approach. PloS ONE 11: e0152951.
Nawy, T. (2016) Capturing microbial interac-
tions. Nat Methods 14: 35–35.
Nicholson, W.L. (2008) The Bacillus subtilis
ydjL (bdhA) gene encodes acetoin reduc-
tase/2, 3-butanediol dehydrogenase. Appl.
Environ. Microbiol. 74: 6832–6838.
Oh, J.-H., and van Pijkeren, J.-P. (2014)
CRISPR- Cas9-assisted recombineering in
Lactobacillus reuteri. Nucleic Acids Res.
42: e131–e131.
Ongena, M., Jourdan, E., Adam, A., Paquot, M.,
Brans, A., Joris, B. et al. (2007) Surfactin and
fengycin lipopeptides of Bacillus subtilis as
elicitors of induced systemic resistance in
plants. Environ. Microbiol. 9: 1084–1090.
Overkamp, W., Beilharz, K., Detert Oude Weme,
R., Solopova, A., Karsens, H., Kovacs, A.T.
et al. (2013) Benchmarking Various Green
Fluorescent Protein Variants in Bacillus
subtilis, Streptococcus pneumoniae, and
Lactococcus lactis for Live Cell Imaging.
Appl. Environ. Microbiol. 79: 6481–6490.
Pathak, K.V., and Keharia, H. (2014) Identifica-
tion of surfactins and iturins produced by
potent fungal antagonist, Bacillus subti-
lis K1 isolated from aerial roots of banyan
(Ficus benghalensis) tree using mass spec-
trometry. 3 Biotech. 4: 283–295.
Peng, Y.-H., Chou, Y.-J., Liu, Y.-C., Jen, J.-F.,
Chung, K.-R., and Huang, J.-W. (2017) Inhi-
bition of cucumber Pythium damping-off
pathogen with zoosporicidal biosurfac-
tants produced by Bacillus mycoides. J.
Plant Dis. Protection 124: 481–491.
Pérez-Miranda, S., Cabirol, N., George-Téllez,
R., Zamudio-Rivera, L., and Fernández, F.
(2007) O-CAS, a fast and universal method
for siderophore detection. J. Microbiol.
Methods 70: 127–131.
Prieto, P., Schiliro, E., Maldonado-Gonzalez,
M.M., Valderrama, R., Barroso-Albarracin,
J.B., and Mercado-Blanco, J. (2011) Root
hairs play a key role in the endophytic col-
onization of olive roots by Pseudomonas
spp. with biocontrol activity. Microb. Ecol.
62: 435–445.
Pyne, M.E., Moo-Young, M., Chung, D.A., and
Chou, C.P. (2015) Coupling the CRISPR/

66
148 149
Expl
orin
g pl
ant-m
icro
be in
tera
ctio
ns o
f the
rhiz
obac
teri
a Ba
cillu
s sub
tilis
and
Bac
illus
myc
oide
s by
use
of th
e CR
ISPR
-Cas
9 sy
stem
Refe
renc
es:
Cas9 System with Lambda Red Recombi-
neering Enables Simplified Chromosomal
Gene Replacement in Escherichia coli. Appl.
Environ. Microbiol. 81: 5103–5114.
Quadri, L.E., Weinreb, P.H., Lei, M., Nakano,
M.M., Zuber, P., and Walsh, C.T. (1998)
Characterization of Sfp, a Bacillus subtilis
phosphopantetheinyl transferase for pep-
tidyl carrier protein domains in peptide
synthetases. Biochemistry 37: 1585–1595.
Radzki, W., Gutierrez Manero, F.J., Algar, E., Lu-
cas Garcia, J.A., Garcia-Villaraco, A., and
Ramos Solano, B. (2013) Bacterial sidero-
phores efficiently provide iron to iron-
starved tomato plants in hydroponics
culture. Antonie Van Leeuwenhoek 104:
321–330.
Renna, M.C., Najimudin, N., Winik, L., and
Zahler, S. (1993) Regulation of the Bacillus
subtilis alsS, alsD, and alsR genes involved
in post-exponential-phase production of
acetoin. J. Bacteriol. 175: 3863–3875.
Rodriguez, H., Mendoza, A., Antonia Cruz, M.,
Holguin, G., Glick, B.R., and Bashan, Y.
(2006) Pleiotropic physiological effects
in the plant growth-promoting bacterium
Azospirillum brasilense following chromo-
somal labeling in the clpX gene. FEMS Mi-
crobiol. Ecol. 57: 217–225.
Romero, D., de Vicente, A., Rakotoaly, R.H., Du-
four, S.E., Veening, J.-W., Arrebola, E. et al.
(2007) The iturin and fengycin families of
lipopeptides are key factors in antagonism
of Bacillus subtilis toward Podosphaera
fusca. Mol. Plant Microbe Int. 20: 430–440.
Ryu, C.-M., Farag, M.A., Hu, C.-H., Reddy, M.S.,
Wei, H.-X., Paré, P.W., and Kloepper, J.W.
(2003) Bacterial volatiles promote growth
in Arabidopsis. Proc. Natl. Acad. Sci. U. S. A.
100: 4927–4932.
Saldanha, R.J., Pemberton, A., Shiflett, P., Pe-
rutka, J., Whitt, J.T., Ellington, A. et al.
(2013) Rapid targeted gene disruption in
Bacillus anthracis. BMC Biotechnol. 13: 72.
Saran, S., Yadav, S., and Saxena, R. (2014) Devel-
opment of a highly sensitive, fast and effi-
cient screening technique for the detection
of 2, 3-butanediol by thin layer chromatog-
raphy. J. Chromatogr. Sep. Tech. 5: 1.
Schaeffer, P., Millet, J., and Aubert, J.-P. (1965)
Catabolic repression of bacterial sporula-
tion. Proc Natl Acad Sci U S A 54: 704–711.
Schalk, I.J., Hannauer, M., and Braud, A. (2011)
New roles for bacterial siderophores in
metal transport and tolerance. Environ.
Microbiol. 13: 2844–2854.
Shevchenko, Y., Bouffard, G.G., Butterfield,
Y.S., Blakesley, R.W., Hartley, J.L., Young,
A.C. et al. (2002) Systematic sequencing
of cDNA clones using the transposon Tn5.
Nucleic Acids Res. 30: 2469–2477.
Shilev, S. (2013) Soil Rhizobacteria Regulating the
Uptake of Nutrients and Undesirable Ele-
ments by Plants. In Plant Microbe Symbiosis:
Fundamentals and Advances. Arora, N.K. (ed).
New Delhi: Springer India, pp. 147–167.
So, Y., Park, S.-Y., Park, E.-H., Park, S.-H., Kim,
E.-J., Pan, J.-G., and Choi, S.-K. (2017) A
Highly Efficient CRISPR-Cas9-Mediated
Large Genomic Deletion in Bacillus subti-
lis. Front. Microbiol. 8: 1167.
Song, C., Voort, M., Mortel, J., Hassan, K.A., El-
bourne, L.D., Paulsen, I.T. et al. (2015) The
Rsm regulon of plant growth- promoting
Pseudomonas fluorescens SS101: role of
small RNAs in regulation of lipopeptide
biosynthesis. Microb. Biotechnol. 8: 296–310.
Suzuki, H., Murakami, A., and Yoshida, K.-I.
(2012) Counterselection system for Geo-
bacillus kaustophilus HTA426 through dis-
ruption of pyrF and pyrR. Appl. Environ.
Microbiol. 78: 7376–7383.
Tong, Y., Charusanti, P., Zhang, L., Weber, T.,
and Lee, S.Y. (2015) CRISPR-Cas9 based
engineering of actinomycetal genomes.
ACS Synth. Biol. 4: 1020–1029.
Vacheron, J., Desbrosses, G., Bouffaud, M.-L.,
Touraine, B., Moënne-Loccoz, Y., Muller,
D. et al. (2013) Plant growth-promoting
rhizobacteria and root system function-
ing. Front. Plant Sci. 4: 356.
Vagner, V., Dervyn, E., and Ehrlich, S.D. (1998)
A vector for systematic gene inactiva-
tion in Bacillus subtilis. Microbiology 144:
3097–3104.
Vater, J., Kablitz, B., Wilde, C., Franke, P., Me-
hta, N., and Cameotra, S.S. (2002) Ma-
trix-assisted laser desorption ioniza-
tion-time of flight mass spectrometry of
lipopeptide biosurfactants in whole cells
and culture filtrates of Bacillus subtilis
C-1 isolated from petroleum sludge. Appl.
Environ. Microbiol. 68: 6210–6219.
Wang, Y., Zhang, Z.T., Seo, S.O., Lynn, P., Lu, T.,
Jin, Y.S., and Blaschek, H.P. (2016) Bacte-
rial Genome Editing with CRISPR-Cas9:
Deletion, Integration, Single Nucleotide
Modification, and Desirable “Clean” Mu-
tant Selection in Clostridium beijerinckii
as an Example. ACS Synth. Biol. 5: 721–732.
Wilson, A.C., Perego, M., and Hoch, J.A. (2007)
New transposon delivery plasmids for
insertional mutagenesis in Bacillus an-
thracis. J. Microbiol. Methods 71: 332–335.
Yang, H., Li, X., Li, X., Yu, H., and Shen, Z.
(2015) Identification of lipopeptide iso-
forms by MALDI-TOF-MS/MS based on
the simultaneous purification of iturin,
fengycin, and surfactin by RP-HPLC.
Anal Bioanal Chem 407: 2529–2542.
Yi, Y., and Kuipers, O.P. (2017) Development of
an efficient electroporation method for
rhizobacterial Bacillus mycoides strains. J.
Microbiol. Methods 133: 82–86.
Yi, Y., de Jong, A., Frenzel, E., and Kuipers, O.P.
(2017a) Comparative Transcriptomics of
Bacillus mycoides Strains in Response to
Potato-Root Exudates Reveals Different
Genetic Adaptation of Endophytic and
Soil Isolates. Front. Microbiol. 8: 1487.
Yi, Y., Frenzel, E., Spoelder, J., Elzenga, J.T.M., El-
sas, J.D.v., and Kuipers, O.P. (2018) Optimized
fluorescent proteins for the rhizosphere-as-
sociated bacterium Bacillus mycoides with
endophytic and biocontrol agent potential.
Environ. Microbiol. Rep. 10: 57–74.
Zhang, K., Duan, X., and Wu, J. (2016) Multi-
gene disruption in undomesticated Bacil-
lus subtilis ATCC 6051a using the CRISPR/
Cas9 system. Sci. Rep. 6: 27943.
Zhang, Z., Ding, Z.T., Shu, D., Luo, D., and Tan,
H. (2015) Development of an efficient elec-
troporation method for iturin A- producing
Bacillus subtilis ZK. Int. J. Mol. Sci. 16:
7334–7351.
Zihalirwa Kulimushi, P., Arguelles Arias, A.,
Franzil, L., Steels, S., and Ongena, M.
(2017) Stimulation of Fengycin-Type An-
tifungal Lipopeptides in Bacillus amyloliq-
uefaciens in the Presence of the Maize Fun-
gal Pathogen Rhizomucor variabilis. Front.
Microbiol. 8: 850.

7General discussion

7
153
Gen
eral
dis
cuss
ion
The world population is expected to reach 8.5 billion by 2030, 9.7 bil-lion in 2050 and 11.2 billion in 2100, according to a report by the United Nations Department of Economic and Social Affairs (www.un.org/en/ development/desa/news/population/2015-report.html). To produce suf-ficient food remains a major task for modern societies. This means there will be an increasing demand for agricultural productivity of food crops, as plants form the basis of every food chain (Morrissey et al., 2004). Over the last decades, the agriculture and horticulture productivity has largely depended on the use of chemical fertilizers, pesticides, and herbicides. These chemicals can help in boosting the growth of plants, but they also have devastating side effects in the long run for the environment. How to increase crop yields without causing environmental damage is a great challenge globally. Great efforts have been made by scientists to increase the productivity, disease resistance and stress tolerance of crops through breeding or genetic modification methods. However, these efforts have mainly focused on plant phenotypes. The effects of the beneficial plant- associated microbial community, also referred to as the second genome of the plants, on the host health and development have been a topic of in-creasing attention by the scientific community in the last years.
The root-associated microbiome plays a primary role in controlling phytopathogens. One well-known example is the occurrence of disease- suppressive soils, which are exceptional ecosystems in which little or no disease occurs under conditions that are favorable for disease devel-opment (Bulgarelli et al., 2013). The suppressive soil can be induced af-ter a severe disease outbreak of a monoculture of the same crop species for several years. This is because plants can exploit microbial consortia from soil for protection against infections when attacked by pathogens (Mendes et al., 2011). Meanwhile, the legume–rhizobium symbiosis is a classic model of mutualistic plant-microbe interaction in which the bac-teria bring fixed nitrogen to the plant, receiving, in turn, the sanctuary and sugars that the plant cell can provide (Long, 2001). The root-nodule bacteria can be manipulated ecologically, agronomically, edaphically and genetically to improve legume productivity and, as a consequence, soil fertility (Brockwell et al., 1995). Apart from disease prevention and nitro-gen fixation, rhizosphere dwelling microbes have various plant promot-ing effects including plant hormone production (Idris et al., 2007), host stress tolerance enhancement (Barka et al., 2006), and systematic resist-ance induction (Schuhegger et al., 2006). An understanding of the funda-mental mechanisms of how plants and soil microbes co-exist and benefit each other can therefore provide new strategies to improve plant produc-tivity and reduce the use of chemicals, which destruct the biological com-munity and destabilize the agro-ecosystem. In this study, we showed that

77
154 155
Gen
eral
dis
cuss
ion
Gen
eral
dis
cuss
ion
Bacillus mycoides is abundantly present in healthy potato roots, and lives inside root tissue as an endophytic bacterium. Moreover, there is a large diversity among the rhizosphere- and endosphere-isolated strains. Other researchers also reported the biocontrol and nitrogen fixation activity of B. mycoides isolated from various plant species. This thesis work aimed to understand the molecular mechanism of B. mycoides-plant interactions. To this end, a multidisciplinary approach was adopted and novel molecu-lar genetic tools were developed (Figure 1).
During the adaptation to the rhizosphere niche, the rhizosphere-associ-ated strains evolved specific properties different from the non-rhizosphere strains. Comparative genomics of plant-associated Pseudomonas spp. re-vealed that most of the biological control genes are in the variable regions of the genome (Loper et al., 2012), indicating that horizontal gene transfer played a key role in the adaptation process. The strains isolated from dif-ferent niches also have different metabolic preferences. Timm et al. (2015) compared the genome of rhizosphere and endosphere isolated Pseudomonas fluorescens strains. Their results showed that multiple pathways relevant for
UP_EC18UP_SB8
61
5352
54148 294
112 201
DOWN_EC18DOWN_SB8
Chapter 3 Chapter 4
AB marker
HR fragment
cas 9
gRNA
CAS9-gRNA complex
?
Chapter 6Chapter 5
Figure 1. A multidisciplinary approach was adopted to study the plant-microbe inter-actions of B. mycoides. Novel molecular genetics tools and methods were developed that can also be useful in other rhizosphere dwelling Bacilli.
plant-bacterial interactions are enriched in endosphere-isolated genomes. In the case of B. mycoides, only some strains have the ability to penetrate plant tissue and start an endophytic life stage. Since the plant beneficial ef-fects of endophytes are greater than those of many rhizosphere-colonizing bacteria (Hardoim et al., 2008), we were interested in exploring the genetic difference of the endophytic and soil strains of B. mycoides. In chapter 2, we selected 7 strains based on their origin of isolation and plant colonization ability: four strains isolated from the endosphere that could be endophytic again (plant-associated), and three strains isolated from soil which could not be endophytic (non-plant-associated). The whole genome phylogenetic tree showed that the endophytic strains are clustered together, indicating that the endophytic strains have some common features.
It has been hypothesized that rhizosphere bacteria start root coloniza-tion when encountering root exudates. Understanding the bacterial re-sponse to root exudates is a key step in deciphering plant-microbe interac-tion mechanisms. In chapter 3, we compared the transcriptome profiles of an endophytic strain and a soil strain of B. mycoides in response to po-tato root exudates by RNA-seq. We first used confocal laser scanning mi-croscope (CLSM) to confirm that the endophytic strain has better root col-onization ability than the soil strain. Our transcriptome results revealed that the endophytic strain showed a more active response than the soil strain to potato root exudates (endophyte strain has more genes altered by root exudates). The upregulation of genes related to amino acid metab-olism, several proteolytic enzymes, and O-glycosyl hydrolases points to-ward a specific adaptation to the ecological niche and a good rhizosphere fitness of the endophytic strain. In comparison, the upregulation of sugar transport and metabolite genes of the soil isolate indicates a narrow nu-trition source, which might hamper its colonization ability and prolifera-tion rate in the rhizosphere.
For environmental isolated bacteria, the lack of efficient transforma-tion methods is a bottleneck to establish the genetic manipulation sys-tems, e.g., enabling gene deletion and mutation within their genome. It has been found that many environmental isolated bacteria are quite resist-ant to natural transformation due to their low genetic competence levels (Duitman et al., 2007). So far, different methodologies have been proposed for transforming exogenous DNA into bacterial cells. Phage-mediated transduction has high efficiency in some bacteria (Winstel et al., 2015). However, bacteriophages have a limited host range and homologous re-combination is required to perform such method. Although protoplast transformation (Li et al., 2016) and protoplast electroporation (Romero et al., 2006) have also been described, a possible disadvantage with the protoplast is that some bacteria are vulnerable to cell wall degrading

77
156 157
Gen
eral
dis
cuss
ion
Gen
eral
dis
cuss
ion
enzymes and very precise manipulation is needed. In chapter 4, we de-veloped a method for electroporation of environmental Bacillus mycoides strains by optimizing several conditions that affect the electroporation efficiency. We found that the transformation efficiency positively corre-lated to the media richness (LB<2XTY<BHIS). The special chaining prop-erty of B. mycoides might play a role in determining the transformation ef-ficiency, because the cells were less aggregated in rich medium. We found that cells collected from the early exponential phase (OD600 = 0.9~1) re-sult in a higher electroporation efficiency. This is different with B. subti-lis WB800, which has high electroporation efficiency at late growth-stage (OD600 = 2.2–2.3) (Lu et al., 2012). The addition of 2% glycine to the culture for 1 hour significantly improved the electroporation efficiency because glycine can reduce the peptidoglycan bonds and loosen up the cell wall by replacing the L- and D-alanine bridges. Moreover, most B. mycoides strains show a higher transformation efficiency when using non- methylated plasmids isolated from the methylation-defect strain E. coli JM110, which indicates that restriction/modification is the main barrier for electropo-ration of this bacterium.
Visualization of fluorescent protein (FP)-labeled rhizobacteria in the rhizosphere and endosphere is a key prerequisite to gain detailed insights into endophytic behavior and plant-bacterial interaction mechanisms. However, the performance of fluorescent proteins has to be optimized for the bacterial host. A widely used approach to obtain improved FP variants is the adaptation of FP genes to the typical codon usage of the host organ-ism. Such codon optimized FPs have been developed for the cyan fluores-cent protein and a yellow fluorescent protein in Bacillus anthracis (Sastalla et al., 2009), for GFP and RFP in Botrytis cinerea (Leroch et al., 2011) and for GFP in Zymoseptoria tritici (Kilaru et al., 2015). In chapter 5, we used a different in vivo cell sorting approach to select the best performing FP variants in B. mycoides from mutational libraries of the superfolder green fluorescent protein sfGFP and the red fluorescent protein mKate2. After selection, the sfGFP variant SPS6 and the mKate2 variant KPS12, with sig-nificantly increased brightness, were isolated. Both optimized variants were highly suitable for in situ localization studies. The strain EC18 rap-idly attached to plant roots and formed a multicellular matrix on the sur-face. During the process of colonization, the root hairs and their branch-ing regions were hot spots for plant-microbe interactions and probably constitute entrance sites for B. mycoides to establish an endophytic life-style, although both the endophytic strain B. mycoides EC18 and the soil strain B. mycoides SB8 were detected in the rhizosphere. When a 1:1 mix-ture of the EC18 and SB8 cells was inoculated to plants, the roots were pre-dominantly colonized by EC18 (Chapter 2).
The CRISPR-Cas9 system is a powerful and revolutionary genome- editing tool for eukaryotic genomes, but its use in undomesticated envi-ronmental isolated Bacillus strains is still underdeveloped. In Chapter 6, we implemented the CRISPR-Cas9 system in rhizosphere-isolated B. sub-tilis HS3 and B. mycoides EC18 to study their plant-microbe interactions. All required genetic elements were incorporated into one plasmid, which was transformed into the bacterial cells with the electroporation method developed in chapter 3. It was found that the B. mycoides produced bacil-libactin (siderophore) aid the growth of iron-starved plants. The advan-tage of the CRISPR-Cas9 system is that the plasmid can be cured after the genome has been edited. The marker-free mutant can be subjected to the next round of mutagenesis with the same procedure. Based on this prin-ciple, we further inserted a gfp gene into the genome of the mutants and the wild type strain. The GFP-tagged strains were inoculated with cab-bage plants and the roots were observed by confocal laser scanning micro-scope. The results showed that the siderophore-deficient mutants have a reduced plant colonization ability compared with wild type.
In summary, different strategies were applied to investigate the plant-microbe interactions of B. mycodies. Some of the tools described here are also useful in other Bacillus species, e.g., the FP variants and the CRISPR-Cas9 system. However, there are still many questions that are open and more research is needed to fully understand the relationship of B. mycoides and its host plant.
REFERENCES
Barka, E.A., Nowak, J., and Clément, C. (2006)
Enhancement of chilling resistance of in-
oculated grapevine plantlets with a plant
growth-promoting rhizobacterium, Burk-
holderia phytofirmans strain PsJN. Appl. En-
viron. Microbiol. 72: 7246–7252.
Brockwell, J., Bottomley, P.J., and Thies, J.E.
(1995) Manipulation of rhizobia micro-
flora for improving legume productivity
and soil fertility: A critical assessment.
Plant and Soil 174: 143–180.
Bulgarelli, D., Schlaeppi, K., Spaepen, S., Ver
Loren van Themaat, E., and Schulze-Lefert,
P. (2013) Structure and functions of the
bacterial microbiota of plants. Annu Rev
Plant Biol. 64: 807–838.
Duitman, E.H., Wyczawski, D., Boven, L.G.,
Venema, G., Kuipers, O.P., and Hamoen,
L.W. (2007) Novel methods for genetic
transformation of natural Bacillus subti-
lis isolates used to study the regulation
of the mycosubtilin and surfactin syn-
thetases. Appl. Environ. Microbiol. 73:
3490–3496.
Hardoim, P.R., van Overbeek, L.S., and Elsas,
J.D. (2008) Properties of bacterial endo-
phytes and their proposed role in plant
growth. Trends Microbiol. 16: 463–471.

7
158
Gen
eral
dis
cuss
ion
Idris, E.E., Iglesias, D.J., Talon, M., and Borriss,
R. (2007) Tryptophan-dependent produc-
tion of indole-3-acetic acid (IAA) affects
the level of plant growth promotion by Ba-
cillus amyloliquefaciens FZB42. Mol. Plant
Microbe Interact. 20: 619–626.
Kilaru, S., Schuster, M., Studholme, D., Soanes,
D., Lin, C., Talbot, N.J., and Steinberg, G.
(2015) A codon-optimized green fluores-
cent protein for live cell imaging in Zy-
moseptoria tritici. Fungal Genet. Biol. 79:
125–131.
Leroch, M., Mernke, D., Koppenhoefer, D.,
Schneider, P., Mosbach, A., Doehlemann,
G., and Hahn, M. (2011) Living colors in
the gray mold pathogen Botrytis cinerea:
codon-optimized genes encoding green
fluorescent protein and mCherry, which
exhibit bright fluorescence. Appl. Environ.
Microbiol. 77: 2887–2897.
Li, H., Zhang, L., Guo, W., and Xu, D. (2016)
Development of a genetically engineered
Escherichia coli strain for plasmid transfor-
mation in Corynebacterium glutamicum. J.
Microbiol. Methods 131: 156–160.
Long, S.R. (2001) Genes and signals in the Rhi-
zobium-legume symbiosis. Plant Physiol.
125: 69–72.
Loper, J.E., Hassan, K.A., Mavrodi, D.V., Da-
vis, E.W., 2nd, Lim, C.K., Shaffer, B.T. et al.
(2012) Comparative genomics of plant-as-
sociated Pseudomonas spp.: insights into
diversity and inheritance of traits involved
in multitrophic interactions. PLoS Genet.
8: e1002784.
Lu, Y.P., Zhang, C., Lv, F.X., Bie, X.M., and Lu,
Z.X. (2012) Study on the electro-transfor-
mation conditions of improving transfor-
mation efficiency for Bacillus subtilis. Lett.
Appl. Microbiol. 55: 9–14.
Mendes, R., Kruijt, M., de Bruijn, I., Dekkers,
E., van der Voort, M., Schneider, J.H. et al.
(2011) Deciphering the rhizosphere mi-
crobiome for disease-suppressive bacteria.
Science 332: 1097–1100.
Morrissey, J.P., Dow, J.M., Mark, G.L., and
O’Gara, F. (2004) Are microbes at the root
of a solution to world food production?
EMBO Reports 5: 922–926.
Romero, D., Perez-Garcia, A., Veening, J.W., de
Vicente, A., and Kuipers, O.P. (2006) Trans-
formation of undomesticated strains of
Bacillus subtilis by protoplast electropora-
tion. J. Microbiol. Methods 66: 556–559.
Sastalla, I., Chim, K., Cheung, G.Y., Pomer-
antsev, A.P., and Leppla, S.H. (2009) Co-
don-optimized fluorescent proteins
designed for expression in low-GC
gram-positive bacteria. Appl. Environ. Mi-
crobiol. 75: 2099–2110.
Schuhegger, R., Ihring, A., Gantner, S., Bahnweg,
G., Knappe, C., Vogg, G. et al. (2006) Induc-
tion of systemic resistance in tomato by
N-acyl-L-homoserine lactone- producing
rhi zosphere bacteria. Plant, Cell and Envi-
ronment 29: 909–918.
Timm, C.M., Campbell, A.G., Utturkar, S.M.,
Jun, S.R., Parales, R.E., Tan, W.A. et al.
(2015) Metabolic functions of Pseudo-
monas fluorescens strains from Populus
deltoides depend on rhizosphere or endo-
sphere isolation compartment. Front. Mi-
crobiol. 6: 1118.
Winstel, V., Kuhner, P., Krismer, B., Peschel, A.,
and Rohde, H. (2015) Transfer of plasmid
DNA to clinical coagulase-negative staph-
ylococcal pathogens by using a unique
bacteriophage. Appl. Environmental Micro-
biol. 81: 2481–2488.

SSamenvatting

S
163
Sam
enva
ttin
g
Bacteriën die plantenwortels koloniseren en de plantengroei bevorderen worden plantengroei-bevorderende rhizobacteriën (PGPR) genoemd. Ver-schillende studies hebben een toename in gezondheid en productiviteit van verschillende plantensoorten aangetoond door de toepassing van PGPR onder zowel normale als stressvolle omstandigheden. De toepassing van PGPR kan de wereldwijde afhankelijkheid van gevaarlijke landbouwche-micaliën verminderen die biologische levensgemeenschappen vernietigen en het agro-ecosysteem destabiliseren. Van alle PGPR is Bacillus een van de meest bestudeerde soorten vanwege zijn competitieve kolonisatie van de wortels en grote weerstand tegen zware omstandigheden. De mechanis-men voor plantengroeibevordering door Bacillus betreffen directe effecten (toename in de beschikbaarheid van nutriënten voor de plant) en / of indi-recte effecten (antagonistische effecten tegen pathogenen). Initiële experi-menten door onze collega’s toonden aan dat Bacillus mycoides overvloedig aanwezig is in gezonde aardappelwortels, en zelfs verblijft in wortelweef-sel als endofytische bacterie. Bovendien is er een grote diversiteit tussen de stammen afkomstig van de rhizosfeer en endosfeer. Andere onderzoekers hebben ook de biocontrole en stikstoffixatie-activiteit laten zien van B. my-coides geïsoleerd uit verschillende plantensoorten. Dit proefschrift was be-doeld om de moleculaire mechanismen onderliggende de B. mycoides-plant interacties te begrijpen. Hiervoor werd een multidisciplinaire aanpak toe-gepast en werden nieuwe moleculair genetische hulpmiddelen ontwikkeld.
B. mycoides is alomtegenwoordig in aarde en rhizosfeer. Sommige stam-men hebben het vermogen om plantenweefsel binnen te dringen en een endofytische levensfase te starten. In hoofdstuk 2 & 3 gebruikten we ver-gelijkende genoom en transcriptoom benaderingen om de genetische de-terminanten van de endosfeer (plant-geassocieerde) stammen ten opzichte van bodem (niet plant-geassocieerde) stammen te identificeren. De fylo-genetische stamboom die gebaseerd was op de genoom sequenties van drie endofytische stammen en drie bodem stammen toonde aan dat de endofy-tische stammen geclusterd zijn, wat aangeeft dat de endofytische stammen enkele gemeenschappelijke kenmerken hebben. Bovendien vertoonde de endofytische stam meer actieve of gerichte reacties tegen exsudaat van aardappelwortel dan de bodemstam. De verhoogde expressie van genen gerelateerd aan het aminozuurmetabolisme, verschillende proteolytische enzymen en O-glycosylhydrolasen wijzen op een specifieke aanpassing aan de ecologische niche en een goede rhizosfeer fitheid van de endofytische stam. Ter vergelijking: de voedingsbron voor de bodem stam lijkt beperkt te zijn tot suikers, wat het kolonisatievermogen en de proliferatie snelheid in de rhizosfeer zou kunnen belemmeren.
Voor milieu-geïsoleerde bacteriën is het gebrek aan een efficiënte trans-formatiewerkwijze een knelpunt om genetische manipulatiesystemen op

S
164 165
Sam
enva
ttin
g
te zetten, bijvoorbeeld die het verwijderen en veranderen van genen in hun genoom mogelijk maken. In hoofdstuk 4 hebben we een methode ontwik-keld voor elektroporatie van milieu-afkomstige Bacillus mycoides stammen door factoren te optimaliseren die de elektroporatie efficiëntie beïnvloeden. Deze methode is reproduceerbaar en makkelijk te gebruiken. De hoge effi-ciëntie vergemakkelijkt geavanceerde genetische studies in deze bacterie.
Visualisatie van fluorescent eiwit-gelabelde rhizobacteriën in de rhizo-sfeer en de endosfeer is een belangrijke vereiste om gedetailleerd inzicht te krijgen in endofytisch gedrag en interacties tussen planten en bacteriën. Echter, moet de werking van fluorescerende eiwitten meestal worden geop-timaliseerd voor de bacteriële gastheer. In hoofdstuk 5 construeerden we een reeks mutanten van het ‘’superfolder’’ groene fluorescerende eiwit sf-GFP en het rood fluorescerende eiwit mKate2, in de bacterie B. mycoides. Na in vivo selectie van de best presterende fluorescente eiwit varianten door flu-orescentie-geactiveerde celsortering, werden de sfGFP-variant SPS6 en de mKate2-variant KPS12 met significant verhoogde helderheid geïdentificeerd. Beide geoptimaliseerde varianten bleken zeer geschikt voor in situ lokalisatie. De stam EC18 hechtte zich snel aan plantenwortels en vormde een multicellu-laire matrix op het oppervlak. Tijdens het kolonisatieproces zijn de wortelha-ren en de vertakkende gebieden hotspots voor plant-microbe interacties en vormen ze waarschijnlijk toegangsplaatsen voor B. mycoides om een endofy-tische levensstijl te beginnen. Hoewel zowel de endofytische stam B. mycoides EC18 en de bodem stam B. mycoides SB8 werden gedetecteerd in de rhizosfeer, werden de wortels voornamelijk gekoloniseerd door EC18 nadat de plant be-ent was met een 1: 1 mengsel van de EC18- en SB8-cellen (hoofdstuk 3).
Een gebrek aan efficiënte hulpmiddelen voor genetische manipulatie heeft het onderzoek naar de plant-microbe interacties van het geslacht Bacillus belemmerd. Het CRISPR-Cas9-systeem is een krachtige en revo-lutionaire bewerkingsmethode voor eukaryotische genomen, maar het gebruik ervan in wilde Bacillus stammen is zeer beperkt. In hoofdstuk 6 hebben we het CRISPR-Cas9-systeem geïmplementeerd in rhizosfeer- geïsoleerde B. subtilis en B. mycoides om vervolgens hun plant-microbe interacties te bestuderen. We toonden aan dat de door B. mycoides gepro-duceerde sideroforen de groei van ijzer-gelimiteerde planten bevorderde. Mutanten deficient in siderofoor productie vertonen een verminderd ver-mogen tot kolonisatie van planten in vergelijking met het wildtype.
Samengevat werden er verschillende strategieën toegepast om de plant- microbe interacties van B. mycoides te onderzoeken. Sommige van de hier bes-chreven hulpmiddelen zijn ook bruikbaar in andere Bacillus soorten, bijvoor-beeld de fluorescente eiwit varianten en het CRISPR-Cas9 systeem. Er zijn echter nog steeds veel onbeantwoorde vragen en meer onderzoek is nodig om de relatie tussen B. mycoides en de gastheerplant volledig te begrijpen.
Ack
now
ledg
emen
ts
A
ACKNOWLEDGEMENTS
This thesis is the outcome of a multi-year journey with many ups and downs, I am taking this opportunity to express my gratitude to everyone who have supported me throughout my Ph.D. First and foremost, I want to thank my supervisor Prof. Oscar P. Kuipers. Without your assistance and dedicated guidance in every step of the way, this thesis would have never been accomplished. I appreciate all your contributions of time and ideas to make my Ph.D productive and stimulating. The joy and enthusiasm you have for scientific research was contagious and motivational, even during tough times. You give me freedom in research and always have patience on me. Your trust and encouragement is the initiation for pursuing my scientific goal and motivation for writing of this thesis.
I would like to express my sincere gratitude to Prof. Jan Kok and Prof. Jan-Willem Veening for all kind suggestions and discussions during group meetings. My thanks also go to my reading committee, Prof. Arnold J.M. Driessen, Prof. Joana Falcao Salles and Prof. Jos Raaijmakers, for reading the draft and providing valuable comments.
The work on plant-associated Bacillus mycoides was initiated by Prof. Jan Dirk van Elsas, Prof. Theo Elzenga, and Jan Spoelder. After I came to Groningen in 2013, Oscar and I decided to participate in this interesting project. Back then, this project was extremely challenging for me as I had not work on Bacillus before and nobody in our group have related expe-rience. Thanks to our excellent collaborators for sharing the knowledge and expertise in the study of plant physiology and microbial ecology. I have learnt a lot from our B. mycoides meetings.
I have obtained lots of help from so many intelligent colleagues in the group. Siger Holsappel, Anne Hesseling, and Harma Karsens, thank you all for keeping the group running smoothly. Marielle, many thanks for in-troducing me to the lab. You have taught me a lot of lab skills required particularly in the beginning. Good luck with your defense later on! Anne de Jong, I am very appreciated for the kind help that you provided for the genome and transcriptome analysis in chapter 2 and 3. Ruud and Manolo, thank you for making me integrated to the group. Elrike, many thanks for the collaboration on the FP-optimization experiment. My gratefulness also goes to my students Evenlien and Vytenis. Thank you guys for work-ing with me. You guys definitely have a great future ahead.
In the last two years of my Ph.D, I feel less “lonely” at work because we gradually build up a plant-microbe team. Chunxu, it was always an enjoy-able experience for all the discussions with you. Zhibo, thank you for your generous help in chapter 5 and 6. Lu and Claudia, thanks for your company during conference traveling. Good luck with your Ph.D project. I must thank

166 167
Ack
now
ledg
emen
ts
Ack
now
ledg
emen
ts
AA
the Bacillus lab members for making the lab a pleasant place to work. Mirjam, thanks you for all the efforts in maintaining the lab crystal clean, and also for sharing your idears on various topics. Amanda, you are such a nice per-son who is always willing to help others. Thanks for translating the Dutch summary for me. Auke, thank you for bringing your cheerfulness to the lab and being my paranymph, as well as all the help you offered for my defense. Ard Jan, Barbara, and Luiza, thank you for all the help you offered to the lab.
I am also grateful to all the former and present colleagues in Molgen including Jeroen, Rybon, Katrin, Morten, Martijn, Angel, Afzal, Irfan, Taketo, Yoshi, Xin, Tonia, Lieke, Dongdong, Liang, Sjoerd, Ruben, Rieza, Jakob, Jhonatan, Xinghong, Jingqi, Zhenhua, Fangfang, Anne-Marie, Patricia, Eduardo, Anna, I also thank the group members that have moved to Lausanne: Clement, Renske, Dimitra, Arnau, Lance, Stefano, and Xue. Thank you for all your support and help during these years.
Qian and Haojie, you deserve a special thank for many reasons. We came to Groningen at the same time and started our Ph.D project in Molgen on the same day. We share opinions, discuss scientific or personal- related topics, helping each other to overcome problems and difficulties. These memories will never be erased from my memory.
What’s more, I was so lucky to have amazing officemates: Maike, Marielle, Jingjing, and the previous member Sornchai. Thank you guys for creating a pleasant atmosphere. And thank you Maike for reading the chapter 1 and providing useful suggestions.
Getting through my dissertation required more than academic sup-port, and I have many people to appreciate for spending the leisure time with me. My first thankfulness goes to the table tennis club member: Jelle, Robin, Chenxi, and Zhibo, we had such enjoyable time. Secondly, the weekend basketball team member: Yuanze, Qihui, Hang He, Hengxu, Ling Liu, Lei Lv, I enjoyed a lot when we play together. My gratefulness also goes to all my friends outside the lab for your warmhearted company during weekends and holidays. Alberto, we met each other in an English course in the beginning Ph.D. Thank you for inviting me to your house and meet so many nice friends: Ajren, Jalmer, Roccio, and Simone. We had great time together. The fusion dinners we had, the barbecue and the paintball game we played, especially the escape room, will always be one of my best memories in Goningen. Martinus and Anna, what a fabulous time we had in Brazil! Most importantly, we had the “Yanglei-try” contin-ued in Amsterdam, Leiden, Groningen, and I believe it will be continued on in the future. Bin Jiang, Lingyang, Boqun, I could not express my entire gratitude to your guys in a few lines here. The unforgettable cakes, and all the drinks and dinners has dramatically strengthened our friendship. Thank you for all the help at work and in daily life.
My gratitude also goes to all my Chinese friends. Youtao Liu, thank you for showing me around when I started my Ph.D, I am sure you will have a successful career after your post-doc in Canada. Jie, Chao, and Bing, we were in the same lab in China and now study in different countries, so nice that we are still able to catch up in Europe. Good luck with your Ph.D study. I also thank all the friends I met in Goningen. Nana Yu, Pu Yang, Miao Wang, Yingruo, Xiangfeng, Yuxiang, Jinfeng, Peiliang & Lijuan, Jing Wu, Yanjun & Peipei, Peng Wang, Min & Xuewen, Huifang, Gang Huang, Lianghui, Wei Qin, your company and presence has become an important part of my life in this city.
Last but not least, nobody has been more important to me in the pur-suit of this project than the members of my family. I would like to thank my dear parents, your love and guidance are with me wherever I go. My dear brother Yanghui, thank you for all the encouragement you provided by phone calls and messages. Most importantly, I would like to thank my loving and supportive wife, Fan, for providing me un-ended inspirations. It was very difficult for us as we were working in different countries, that you have to travel so many times between China and Netherlands. The holidays we had together in those beautiful European cities has been the best time in my life. I look forward to creating our new life together!
Copyright © 2022 FDOKUMEN




















