
Assessing jaguar abundance using remotely triggered cameras Table of Contents

S58 Journal of Basic Microbiology 2010, 50, S58–S67
© 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jbm-journal.com
Research Paper
Ultrastructure of Aphanomyces cochlioides zoospores and changes during their developmental transitions triggered by the host-specific flavone cochliophilin A
M. Tofazzal Islam
Graduate School of Agriculture, Hokkaido University, Kita-Ku, Sapporo, Japan
Aphanomyces cochlioides is a serious damping-off causing pathogen of sugar beet, spinach and some other members of Chenopodiaceae and Amaranthaceae. The biflagellated motile zoo-spores of the pathogen locate their host roots by perceiving the host-specific flavone cochlio-philin A (5-hydroxy-6,7-methylenedioxyflavone), transiently modify into cystospores that ger-minate prior to penetration. This study for the first time illustrated ultrastructure of the zoospores and morphological modification during their developmental transitions triggered by cochliophilin A using transmission electron microscopy (TEM). TEM revealed that zoospores had two heterokont flagella inserted laterally into a ventral groove of their body where each is attached to a kinetosome. In the cross sections of flagellar axonemes, two single and nine peripheral microtubules in doublets were clearly observed. Mitochondria, the Golgi complexes, finger print vesicles, and vesicles with striated electron opaque inclusion and vesicles con-taining a granular cortex and center were also detected. The latter vesicles disappeared and two flagella were shed when zoospores converted to spherical cystsopores. The shape, size and number of mitochondria were dynamically changed during the encystment of zoospores presumably through fission and fusion processes. The dynamics of mitochondria observed in this study indicated its distinct role in the signal transduction pathway of the zoospore encystment. This study also revealed the transformation of shape of nuclei from pyriform in zoospores to spherical in cystospores and lanceolate in the hyphae.
Keywords: Aphanomyces cochlioides / Zoospore encystment / Mitochondrial dynamics / Host-specific signal / Oomycetes / Flagella / TEM
Received: September 13, 2009; accepted: December 19, 2009
DOI 10.1002/jobm.200900301
Introduction*
Peronosporomycetes (Oömycetes in the old classifica-tion) belong to the kingdom Straminipila, are distinct from fungi and more closely related to brown algae and diatoms phylogenetically [1–3]. They cause many de-structive diseases of plants, animals, fishes and hu-mans. One of the members of peronosporomycetes, Aphanomyces cochlioides Drechs. in the family Leptoleg-niaceae (order Saprolegniales), is a serious pathogen of damping-off diseases in sugar beet, spinach and some other members of Chenopodiaceae and Amaranthaceae [4]. This fungus-like soilborne stramenopile reproduces Correspondence: Tofazzal Islam, School of Agriculture and Rural De-velopment, Bangladesh Open University, Gazipur-1705, Bangladesh E-mail: [email protected]
asexually via zoosporangia on the vegetative thallus or sexually via antheridia and oogonia on infected host. The non-motile primary zoospores, released from the cylindrical zoosporangia, immediately encyst and at-tach in clusters on the tip of the sporangia [4]. Laterally biflagellate, uninucleate, naked (no cell wall) and mo-tile secondary zoospores arising from the primary cystospores are thought to be the main infective propagules of seedlings [5–8]. The reniform-ovate sec-ondary zoospore of A. cochlioides has two heterokont (dissimilar) flagella, both inserted into a ventral groove [6]. The anterior flagellum possesses two rows of tripar-tite tubular hairs (TTHs) distributed throughout its length, whereas the posterior flagellum is ornamented with two rows of dense fine hairs except on its tapered terminal part [2, 6]. Once released from the primary

Journal of Basic Microbiology 2010, 50, S58–S67 Ultrastructure of asexual spores of Aphanomyces S59
© 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jbm-journal.com
cysts, the motile secondary zoospores precisely locate infection sites of the host roots by perceiving cochlio-philin A (5-hydroxy-6,7-methylenedioxyflavone), a host-specific flavone released from the roots [6, 7, 9]. This flavone signal triggers rapid differentiation (encyst-ment of zoospores followed by germination of cysto-spores) of the accumulated zoospores (within 30–40 min) required for the initiation of infection [10–12]. Changes in the morphological features of zoospores and the developmental transitions that occur during the interactions with host, were studied by light and scanning electron microscopy [6, 7, 12]. Islam [12] dem-onstrated that these rapid morphological changes of zoospores triggered by the host signal cochliophilin A were linked to the dynamic rearrangement of cy-toskeletal filamentous actin organization in A. coch-lioides cells [12]. Host-specific plant signals that trigger chemotaxis and differentiation of the zoospores were reported for some other soilborne Peronosporomycetes. For exam-ple, prunetin (5,4′-dihydroxy-7-methoxyisoflavone) iso-lated from the garden pea for A. euteiches [13], and daidzein and genistein from the soybean for Phyto-phthora sojae zoospores [14]. However, no attempts were made towards understanding the mode of action of these host signals on the developmental modifications of pathogen spores at an ultrastructural level. A clear understanding of the cellular and molecular mecha-nism involved with chemotaxis and zoospore differen-tiation should be helpful in designing biorational man-agement strategies against these pathogens [15, 16]. Zoosporogesis in A. cochlioides occur through a set of cellular processes which is distinctly different from other Peronosporomycetes such as Phytophthora, Py-thium, Plasmopara Saprolegnia, Isoachlya, Leptolegnia, and Dictyuchus [2, 15, 17]. The ultrastructural details of zoo-spores of some peronosporomycete genera like Phy-tophthora, Pythium, Sclerophthora, and Saprolegnia have been studied by transmission electron microscopy [18–25]. A detail ultrastructural study of zoospores, cysto-spores and germinated cystospores of a soilborne pea root pathogen, A. euteiches has been done by Hoch and Mitchell in early eighties [26, 27]. Cerenius and co-workers conducted untrastructural studies on secon-dary zoospores of A. astaci and A. laevis and reported differences between these two species [28, 29]. How-ever, no study was conducted on ultrastructure of A. cochlioides zoospores and their dynamic changes that occur during interactions with the host-specific coch-liophilin A. The objectives of this study were to illus-trate ultrastructure of A. cochlioides zoospores and their dynamic changes during interactions with the host-
specific flavone cochliophilin A by transmission elec-tron microscopy.
Materials and methods
Culture of Aphanomyces cochlioides and production of zoospores Aphanomyces cochlioides (AC-5), a gift from Prof. R. Yoko-sawa, was isolated from the soil of a sugar beet field. It was routinely grown on corn meal agar (Difco) in the plastic Petri dishes (9 cm i.d.) at 20 °C [9]. High density A. cochlioides zoospore suspension was obtained follow-ing the protocols described previously [8, 15]. Dynamic developmental transitions of zoospores were achieved by applying cochliophilin A (1 × 10–6 M) coated Chro-mosorb W AW particles into zoospore suspension as described previously [10, 12]. To get a specific stage of the developing spores, the interactions between zoo-spores and the host signal cochliophilin A was stopped by adding 2% glutaraldehyde (TAAB, Birkshire, U.K.) to the spore suspension at a given time (1, 5, 10, 15, 20, 25, and 30 min) of interactions. The glutaraldehyde- fixed cell suspension of the specific life stage was used for further microscopic studies following the protocol described earlier [10, 12].
Transmission electron microscopy Exactly 100 μl of glutaraldehyde fixed (30 min) spore suspension (ca. 1 × 105 zoospores/ml) was taken in an Eppendorf tube and then post-fixed with osmium tetraoxide (10 g/l) for 2 h at 4 °C [12]. Then the samples were dehydrated in a graded acetone series, embedded with Epon 812 (electron microscopy grade, TAAB, U.K.) and polymerized at 60 °C for 24 h. Ultrathin sections (100 nm thickness) were cut with a diamond knife (Su-mitomo Electric Ind., SK1045, Tokyo) and stained with 2% uranyl acetate (5 min). Sections were washed with phosphate buffer saline and briefly stained with lead citrate (3 min), and observed under a Hitachi H-800 transmission electron microscope with an accelerating voltage of 75 kV [12, 30]. Each experiment was repli-cated three times and the representative images are presented.
Confocal laser scanning microscopy Hoechst 33258 (bisbenzamide) (Sigma-Aldrich H6024, St. Louis, MO, USA) was used for visualization of nuclei in the zoospores, cystospores and germinated cysto-spores of A. cochlioides following the protocols described by Deora et al. [31]. The stained spores were observed with a Zeiss confocal laser scanning microscope LSM410
S60 M. T. Islam Journal of Basic Microbiology 2010, 50, S58–S67
© 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jbm-journal.com
(Carl Zeiss, Oberkochen, Germany) in order to detect the nuclei [31].
Results
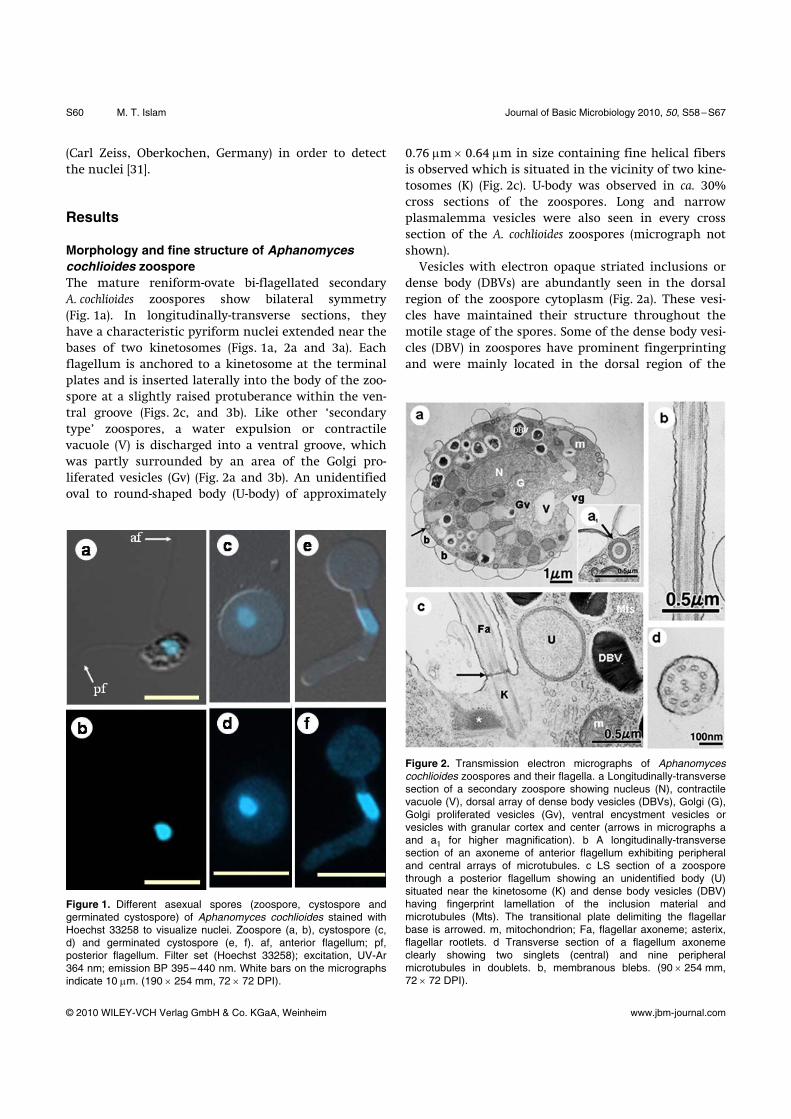
Morphology and fine structure of Aphanomyces cochlioides zoospore The mature reniform-ovate bi-flagellated secondary A. cochlioides zoospores show bilateral symmetry (Fig. 1a). In longitudinally-transverse sections, they have a characteristic pyriform nuclei extended near the bases of two kinetosomes (Figs. 1a, 2a and 3a). Each flagellum is anchored to a kinetosome at the terminal plates and is inserted laterally into the body of the zoo-spore at a slightly raised protuberance within the ven-tral groove (Figs. 2c, and 3b). Like other ‘secondary type’ zoospores, a water expulsion or contractile vacuole (V) is discharged into a ventral groove, which was partly surrounded by an area of the Golgi pro-liferated vesicles (Gv) (Fig. 2a and 3b). An unidentified oval to round-shaped body (U-body) of approximately
Figure 1. Different asexual spores (zoospore, cystospore and germinated cystospore) of Aphanomyces cochlioides stained with Hoechst 33258 to visualize nuclei. Zoospore (a, b), cystospore (c, d) and germinated cystospore (e, f). af, anterior flagellum; pf, posterior flagellum. Filter set (Hoechst 33258); excitation, UV-Ar 364 nm; emission BP 395–440 nm. White bars on the micrographs indicate 10 μm. (190 × 254 mm, 72 × 72 DPI).
0.76 μm × 0.64 μm in size containing fine helical fibers is observed which is situated in the vicinity of two kine-tosomes (K) (Fig. 2c). U-body was observed in ca. 30% cross sections of the zoospores. Long and narrow plasmalemma vesicles were also seen in every cross section of the A. cochlioides zoospores (micrograph not shown). Vesicles with electron opaque striated inclusions or dense body (DBVs) are abundantly seen in the dorsal region of the zoospore cytoplasm (Fig. 2a). These vesi-cles have maintained their structure throughout the motile stage of the spores. Some of the dense body vesi-cles (DBV) in zoospores have prominent fingerprinting and were mainly located in the dorsal region of the
Figure 2. Transmission electron micrographs of Aphanomyces cochlioides zoospores and their flagella. a Longitudinally-transverse section of a secondary zoospore showing nucleus (N), contractile vacuole (V), dorsal array of dense body vesicles (DBVs), Golgi (G), Golgi proliferated vesicles (Gv), ventral encystment vesicles or vesicles with granular cortex and center (arrows in micrographs a and a1 for higher magnification). b A longitudinally-transverse section of an axoneme of anterior flagellum exhibiting peripheral and central arrays of microtubules. c LS section of a zoospore through a posterior flagellum showing an unidentified body (U) situated near the kinetosome (K) and dense body vesicles (DBV) having fingerprint lamellation of the inclusion material and microtubules (Mts). The transitional plate delimiting the flagellar base is arrowed. m, mitochondrion; Fa, flagellar axoneme; asterix, flagellar rootlets. d Transverse section of a flagellum axoneme clearly showing two singlets (central) and nine peripheral microtubules in doublets. b, membranous blebs. (90 × 254 mm, 72 × 72 DPI).

Journal of Basic Microbiology 2010, 50, S58–S67 Ultrastructure of asexual spores of Aphanomyces S61
© 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jbm-journal.com
zoospores (Fig. 2c). Vesicles with an electron opaque cortex and center also present in the zoospores (arrows in Fig. 2a and a1). These vesicles were almost equal in size and observed only at the periphery of the zoo-spores, which were completely disappeared during the encystment of zoospores (Fig. 4c). Large membranous blebs or blisters (Figs. 2a and 4a) synchronously an-chored to the vesicles of electron opaque cortex and center surrounding the zoospores. Mitochondrial (m) ultrastructure of A. cochlioides zoospores is typical of that for other peronosporomycetes exhibiting tubular cristae (Fig. 2a). However, many of the mitochondria observed in zoospores contain concentric and ribosome-like inclusions (Fig. 2a and arrow in Fig. 4b). Variable size and shapes of mitochondria were generally present at the ventral regions particularly near the Golgi prolif-erated area of the zoospores (Fig. 2a). The flagellar base of the peronosporomycete zoo-spores is composed of the kinetosome and attached roots of microtubules [2]. In the present study, two
Figure 3. Transmission electron micrographs of Aphanomyces cochlioides zoospores and flagella. a. TS section of a zoospore through anterior flagellum showing nucleus (N), microtubules (Mts), kinetosome (K), Golgi bodies (G) and basal rootlets containing microtubules (asterix) of the flagellum. Fa, cross section of a flagellum axoneme indicating arrangement of microtubules. b. Longitudinal section through the posterior (Pf) and anterior flagella (Af) and associate kinetosomes (K), of a zoospore. Mitochonrion (m), contractile vacuole (V), and rough Golgi vesicles (Gv) are seen. The transitional plates delimiting the flagellar bases are arrowed but the transitional helices are not clearly resolved. c. LS section through a posterior flagellum up to the tapered tip. Central and peripheral microtubules arrays are extended throughout the flagellum except the tapered tip of the posterior flagellum (ttPf) are seen. (190 × 254 mm, 72 × 72 DPI).
kinetosomes (K) of A. cochlioides zoospores are oriented in a V configuration approximately 135° angle to each other and appear to be interconnected by an electron opaque striate fiber bundle (Fig. 3a and b). Other fiber bundles or ‘rootlets’ (asterix) that partially surrounding the kinetosomes and radiate into the cytoplasm just below the plasmalemma of the zoospore body were also observed (Figs. 2c, 3a and b). There was a well-defined terminal plate (0.31 μm i.d.) in the flagellar base of the zoospore (arrows in Figs. 2c and 3b), but a transitional helix structure was not clearly observed. Longitudinal sections of the flagella display the presence of periph-eral and central microtubules throughout the flagellar axoneme except the tapered tip of the posterior flagel-lum (Fig. 3c). A cross section of a flagellar axoneme (275–300 nm i.d.) clearly shows that the microtubules are organized as two single (central) and nine periph-eral microtubules in doublets, which is typical in the flagella of peronosporomycete zoospores (Fig. 2d). The diameter of each microtubule was 17–21 μm. Figure 3c shows a longitudinal section of a posterior flagellum with a 1.5–1.6 μm long and 75–78 nm diameter ta-pered tip. The tapered tip of the posterior flagellum (ttPf) is seen free from any traces of microtubules (Fig. 3c).
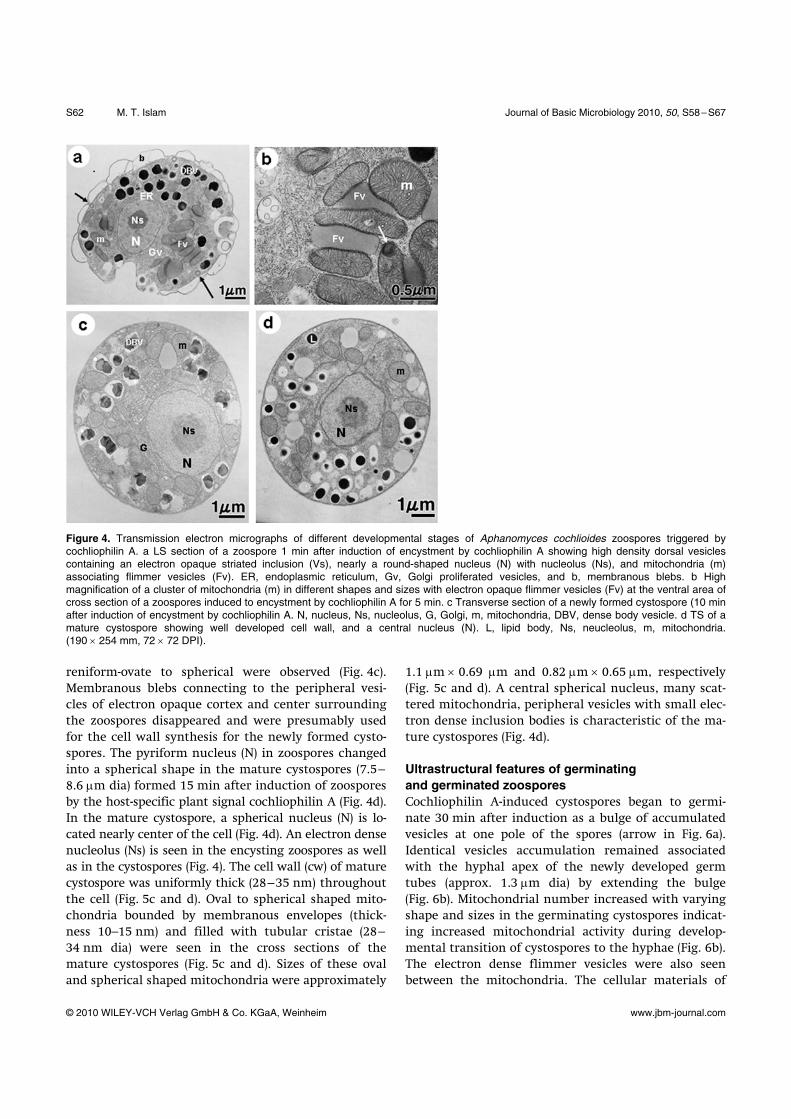
Ultrastructural changes of zoospores during encystment by cochliophilin A Both flagella of the zoospores were shed approximately 1 min after induction of encystment by the cochlio-philin A (Fig. 4a). Soon after the detachment of flagella, the contractile vacuole disorganized and completely disappeared. The pyriform nucleus (N) changed into spherical and surrounded by varying shapes and sizes of abundant mitochondria (m) (Fig. 5a and b). The shape, size and density of mitochondria were dynami-cally changed especially in the Golgi proliferated area during the process of encystment as shown in Figs. 4a, b, 5a and b. Sometimes clusters of mitochondria with variable shape and sizes interconnected by electron dense flimmer vesicles (Fv) were seen in the zoospores fixed after 1 min treatment with cochliophilin A (Fig. 4b). Some of the mitochondria contain concentric inclusions and ribosome-like particles (arrows in Fig. 4b and 5a). The center of some mitochondria also contains electron dense granular structures (Fig. 4b). Ultrastructural changes of A. cochlioides zoospores during first 5 min of interactions with cochliophilin A are characterized by the rapid and drastic reorganiza-tion of cellular constituents. Concurrent with flagellar detachment, rapid changes in shape of the spores from
S62 M. T. Islam Journal of Basic Microbiology 2010, 50, S58–S67
© 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jbm-journal.com
Figure 4. Transmission electron micrographs of different developmental stages of Aphanomyces cochlioides zoospores triggered by cochliophilin A. a LS section of a zoospore 1 min after induction of encystment by cochliophilin A showing high density dorsal vesicles containing an electron opaque striated inclusion (Vs), nearly a round-shaped nucleus (N) with nucleolus (Ns), and mitochondria (m) associating flimmer vesicles (Fv). ER, endoplasmic reticulum, Gv, Golgi proliferated vesicles, and b, membranous blebs. b High magnification of a cluster of mitochondria (m) in different shapes and sizes with electron opaque flimmer vesicles (Fv) at the ventral area of cross section of a zoospores induced to encystment by cochliophilin A for 5 min. c Transverse section of a newly formed cystospore (10 min after induction of encystment by cochliophilin A. N, nucleus, Ns, nucleolus, G, Golgi, m, mitochondria, DBV, dense body vesicle. d TS of a mature cystospore showing well developed cell wall, and a central nucleus (N). L, lipid body, Ns, neucleolus, m, mitochondria. (190 × 254 mm, 72 × 72 DPI).
reniform-ovate to spherical were observed (Fig. 4c). Membranous blebs connecting to the peripheral vesi-cles of electron opaque cortex and center surrounding the zoospores disappeared and were presumably used for the cell wall synthesis for the newly formed cysto-spores. The pyriform nucleus (N) in zoospores changed into a spherical shape in the mature cystospores (7.5–8.6 μm dia) formed 15 min after induction of zoospores by the host-specific plant signal cochliophilin A (Fig. 4d). In the mature cystospore, a spherical nucleus (N) is lo-cated nearly center of the cell (Fig. 4d). An electron dense nucleolus (Ns) is seen in the encysting zoospores as well as in the cystospores (Fig. 4). The cell wall (cw) of mature cystospore was uniformly thick (28–35 nm) throughout the cell (Fig. 5c and d). Oval to spherical shaped mito-chondria bounded by membranous envelopes (thick-ness 10–15 nm) and filled with tubular cristae (28–34 nm dia) were seen in the cross sections of the mature cystospores (Fig. 5c and d). Sizes of these oval and spherical shaped mitochondria were approximately
1.1 μm × 0.69 μm and 0.82 μm × 0.65 μm, respectively (Fig. 5c and d). A central spherical nucleus, many scat-tered mitochondria, peripheral vesicles with small elec-tron dense inclusion bodies is characteristic of the ma-ture cystospores (Fig. 4d).
Ultrastructural features of germinating and germinated zoospores Cochliophilin A-induced cystospores began to germi-nate 30 min after induction as a bulge of accumulated vesicles at one pole of the spores (arrow in Fig. 6a). Identical vesicles accumulation remained associated with the hyphal apex of the newly developed germ tubes (approx. 1.3 μm dia) by extending the bulge (Fig. 6b). Mitochondrial number increased with varying shape and sizes in the germinating cystospores indicat-ing increased mitochondrial activity during develop-mental transition of cystospores to the hyphae (Fig. 6b). The electron dense flimmer vesicles were also seen between the mitochondria. The cellular materials of

Journal of Basic Microbiology 2010, 50, S58–S67 Ultrastructure of asexual spores of Aphanomyces S63
© 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jbm-journal.com
Figure 5. Transmission electron micrographs of mitochondria in cochliophilin A-induced zoospores and in mature cystospore of Aphanomyces cochlioides. a LS section of a zoospore through a flagellum 1 min after induction of encystment by cochliophilin A. Some mitochondria (m) in varying shapes and sizes with concentric inclusions and ribosome-like particles (arrows). Gv, Golgi proliferated vesicles, Fa, flagellum axoneme, N, nucleus. b TS of an encysting zoospore (30 s after induction by cochliophilin A) through ventral region showing contractile vacuole (V), mitochondria (m), nucleus (N), and Golgi proliferated vesicles (Gv). High density variable shapes and sizes of mitochondria (m) (white arrows) are seen. Some of the mitochondria contain concentric inclusion and ribosome-like particles (arrows). c TS of an oval-shaped mitochondrion with conspicuous tubular cristae (tc) in a mature cystospore (20 min induction). mm, mitochondrial membrane. d TS of a spherical mitochondrion with dense conspicuous tubular cristae in a mature cystospore (20 min induction). cw, cell wall; tc, tubular cristae. (254 × 190 mm, 72 × 72 DPI).
cystospores gradually moved to the growing hyphal germ tubes as a result a vacuole (V) is developed oppo-site to the germ tube (Fig. 6b, b’). Fig. 6d shows a longi-tudinal section of a mature hypha. The nucleus (N) in the mature hypha is lanceolate (Fig. 6c).
Discussion
This study for the first time illustrated the ultrastruc-tures of the motile secondary zoospores of A. cochlioides including flagella and their rapid changes occurred during the interaction with the host-specific signal cochliophilin A with very high resolution (Figs. 1–6). Many interesting features of A. cochlioides such as fine structure of flagella, cellular organelles in zoospores and the dynamic changes of vesicles, mitochondria and
nuclei that associated with the dramatic morphological changes of zoospores triggered by the host-specific signal cochliophilin A were revealed. These changes presumably occur during the early events of plant-peronosporomycete interactions [6, 7]. The basic ultrastructural organization of the secon-dary zoospores of A. cochlioides is quite similar to those observed in the secondary zoospores of other species of Aphanomyces studied such as A. euteiches, A. astaci and A. laevis [26–28]. However, this study revealed that some organelles of A. cochlioides zoospores are slightly different from the ultrastructure of other related spe-cies. For example, the water expulsion apparatus of A. cochlioides is well differentiated and quite similar to A. euteiches and A. astaci but slightly different from A. laevis that possess less differentiated water expulsion apparatus [28]. Likewise, the structure of an unknown

S64 M. T. Islam Journal of Basic Microbiology 2010, 50, S58–S67
© 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jbm-journal.com
Figure 6. Ultrastructure of germinating and germinated cystospo-res and hyphae of Aphanomyces cochlioides. a TS section of a cystospore showing nucleus (N), nucleolus (Ns), mitochondria (m), and accumulation of high density vesicles (arrows) at one pole of the cell as a sign of germination. cw, cell wall. b LS section of a germinated cystospores (60 min of induction) showing accumulation of high density vesicles at the apical region of the germ tube (arrow), and inset a cross section of the tip of a germ tube full of vesicles (b’). c Longitudinal section of a mature A. cochlioides hypha showing mitochondria (m), lanceolate nucleus (N), and a large vacuole (V). (190 × 254 mm, 72 × 72 DPI).
body called, U-body located near the kinetosomes of A. cochlioides zoospore (Fig. 2c) is similar to A. laevis and A. euteiches zoospore but different from that of A. astaci [28, 29]. The contents of U-body in A. astaci appeared as paracrystalline compared to helical fibers observed in the U-body of other Aphanomyces [28, 29]. In a previous study, Cerenius and co-workers ob-served vesicles with flocculent contents in the periph-eral cytoplasm of the zoospores of two aquatic species of Aphanomyces, A. astaci and A. laevis, which were emp-tied during encystment [28]. In the present study, no
traces of flocculent vesicles were observed in A. coch-lioides zoospores. This result is consistent with the ob-servation made by Hoch and Mitchell [26] on zoospores of the pea pathogen A. euteiches lacking flocculent vesi-cles [28, 29]. Thus, absence and presence of this kind of unstable type flocculent vesicles in the phytopatho-genic soilborne Aphanomyces species (A. cochlioides and A. euteiches) in aquatic Aphanomyces species, respectively, could draw an evolutionary significance for these two groups of Aphanomyces [28, 29]. Peripheral vesicles with granular contents in A. coch-lioides zoospores, are mostly located at the end of pe-ripheral cysternae in close proximity to plasmalemma (Fig. 2a). These vesicles were interconnected by large blebs or blister like membranous structures (b in Fig. 2a). The interior of these vesicles was organized into two clearly visible layers under TEM, an outer zone with distinct periodicity and a homogeneous lumen and inner zone (Fig. 2a1). No traces of these vesicles were observed in the serial TEM sections of the cysto-spores (Fig. 4c). These observations indicated that these vesicles might be involved in the formation of the cell wall of newly formed cystospores [20, 21], and can be termed as encystment vesicles. Identical peripheral vesicles were observed in the secondary zoospores of all species of the Aphanomyces studied [26–29]. They are highly ordered and distinct from the microbodies with electron dense crystalline inclusions observed in the zoospores of Pythium proliferum [22]. Eukaryotic flagella and cilia are important motile organelles for locomotion and transportation of mate-rials in many organisms, including animals, plants, and protists. The current study clearly demonstrated that the axoneme of an A. cochlioides flagellum contained nine doublet peripheral microtubules (MTs) and one central pair of singlet MTs, comprising the ‘9 + 2’ struc-ture commonly observed in the flagella of perono-sporomycete zoospores (Fig. 2d) [2]. The detail ultra-structure of flagella with high resolution illustrated in this report by a series of TEM sections (Figs. 2c, d and 3a–c) has not been demonstrated so far for any perono-sporomycete zoospores. Although electron microscopy has been commonly used for structural analysis of flagellar axoneme since 1950s [32], currently X-ray dif-fraction method is considered as an efficient tool for precise molecular analysis of the architecture of flagel-lar axonemes [33]. Further studies on architecture of flagellar axonemes of A. cochlioides should utilize this advance tool. Zoospore of A. cochlioides has a large vacuole known as contractile or water expulsion vacuole at the ventral groove region where two flagella are inserted into the

Journal of Basic Microbiology 2010, 50, S58–S67 Ultrastructure of asexual spores of Aphanomyces S65
© 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jbm-journal.com
spore body. A contractile vacuole or water expulsion vacuole is present in a wide range of unicellular micro-organisms including the peronosporomycete zoospores [26, 34]. In A. euteiches zoospores, Hoch and Mitchell observed a rhythmic contraction and relaxation of this vacuole at about 5 sec intervals in the living zoospores [26]. Similar contraction vacuoles were also observed in the zoospores of A. astaci, A. laevis [28] and in some Saprolegnia species [2, 34]. The A. cochlioides zoospores contain dense body vesi-cles (DBV) with prominent finger printing (Fig. 2c). As in other Peronosporomycetes, the DBVs with electron-opaque contents have been known as part of the cells vacuolar storage system [35, 36]. A similar organization of vacuolar granules was observed in A. euteiches zoo-spores and in chlamydospores of Phytophthora [26, 37]. The non-vacuolar, cytoplasmic DBV fraction is eventu-ally incorporated into the zoospore initials, where the inclusions acquire their characteristic fingerprint sub-structure as described in many Peronosporomycetes (Fig. 2c) [26, 37]. Very little is known about the molecu-lar details of these DBVs in zoospores. However, in Phytophthora these vesicles contain phosphorylated my-colaminarins at the lamellate stage [38]. Due to the prominence and presumed importance of DBVs, Chil-vers et al. [39] related it to the phosphate or polyphos-phate storage differences between Eumycota and the Peronosporomycetes. The most interesting feature observed in the current TEM study was the dynamic changes of size, shape and number of mitochondria in the cochliophilin A-induced developing zoospores. The presence of a cluster of mi-tochondria with concentric inclusions and ribosome-like particles that connected by electron dense flimmer vesicles was seen in the sections of zoospores 1 min after induction of encystment by cochliophilin A (Figs. 4b and 5a). The shape and size of mitochondria were found relatively stable and almost uniformly dis-tributed in the mature cystospores which is considered a relatively stationary stage (Fig. 5c and d). These obser-vations suggested that a dynamic mitochondrial fission and fusion might take place during the asexual devel-opment of zoospores triggered by the host signal [40, 41]. This study also clearly demonstrated the ultra-structural details of a typical mitochondrion of a per-onosporomycete phytopathogen with very high resolu-tion (Fig. 5c and d). Encystment of zoospores is critical for successful infection of plants by the zoosporic peronosporomy-cetes. Although the signal transduction pathway of the encystment of zoospores has not been completely elu-cidated, experimental evidence suggest that chemotaxis
and subsequent differentiation (encystment) of A. coch-lioides zoospores triggered by the host-specific cochlio-philin A is initiated by a G protein-coupled receptor, which activate phosphoinositide and Ca2+ second mes-senger pathways [10, 15]. Mitochondria are known to be involved with a range of cellular processes such as sig-naling and differentiation of the cells in addition to their primary role of supplying energy [40, 41]. The dynamic changes of size, shape and number of mito-chondria during encystment of A. cochlioides zoospores triggered by the host-specific cochliophilin A observed in the current study indicated a distinct role of mito-chondria in the signal transduction pathway of zoo-spore encystment. The dynamic fission and fusion of mitochondria during encystment of zoospores shown in the current study was not reported earlier. However, the dynamics of fission and fusion in mammalian mi-tochondria have been demonstrated [41]. Germination of A. cochlioides cystospore initiates ap-proximately 30 min after induction of encystment by cochliophilin A as a localized bulge in the cyst wall (Fig. 6a–c). The apex of A. cochlioides germ tubes con-tained numerous vesicles with dense fibriller contents similar to those reported in other peronosporomycetes (arrow in Fig. 6b) [26, 35]. However, no deposition of electron opaque amorphic material was visible on the outer wall surface of the bulge and germ tube as ob-served in A. euteiches [26]. These vesicles might contain materials for cell wall synthesis for the growing hyphae as well as enzymes to aid host root penetration. Results from my recent study showed that cytoskeletal actin microfilaments remained as a cap at the apex of the newly formed germ tubes of cochliophilin A-induced germinated cystospores [12]. These observations sug-gested that vesicles were likely transported to the site of germ tube emergence or apex of the germ tube through the actin cables [12]. The morphology and dynamic changes of nuclei in different asexual life stages of A. cochlioides were also clearly demonstrated in this paper by parallel studies using TEM and Hoechst 33258 staining followed by CLSM (Figs. 1–6). The shape of nuclei in zoospore, cystospore and germinated cysto-spore/hypha were pyriform, spherical and lanceolate, respectively. In conclusion, this study illustrated the fine struc-ture of A. cochlioides zoospores and their ultrastructural changes occurred during the interaction with the host-specific plant signal cochliophilin A. Further study is needed to elucidate the precise role of mitochon- dria during encystment of the peronosporomycete zoospores triggered by the host-specific plant sig-nal.

S66 M. T. Islam Journal of Basic Microbiology 2010, 50, S58–S67
© 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jbm-journal.com
Acknowledgements
The author is very thankful to Prof. Satoshi Tahara, Graduate School of Agriculture of Hokkaido University, Japan for his enormous supports and encouragements during this study. Special thanks are due to Prof. R. Yokosawa (Health Science University of Hokkaido, Hokkaido, Japan) for his kind gift of A. cochlioides AC-5. Mr. Toshiaki Ito, Electron Microscopic Laboratory of Hokkaido University deserves thanks for his assistance during SEM and TEM studies. The author is very thank-ful to two anonymous reviewers for their scholarly comments and valuable suggestions for the improve-ment of the manuscript. Dr. M. Rahman (NCSU Dept. Plant Pathology, USA) deserves special thanks for lin-guistic editing. Sincere appreciations are also due to the Government of Japan for Monbukagakusho Scholarship during this study and Alexander von Humboldt Foun-dation for fellowship during preparation of this manu-script.
References
[1] Baldauf, S.L., Roger, A.J., Wenk-Siefert, I. and Doolittle, W.F., 2000. A kingdom-level phylogeny of eukaryotes based on combined protein data. Science, 290, 972–977.
[2] Dick, M.W., 2001. The Peronosporomycetes. In: The My-cota VII, Part A, Systematics and Evolution (McLaughlin, D.J., McLaughlin, E.G., Lemke, P.A., eds.). Springer-Verlag, Berlin, Heidelberg, pp. 39–72.
[3] Yoon, H.S., Hackett, J.D., Pinto, G. and Bhattacharya, D., 2002. The single, ancient origin of chromist plastids. Proc. Natl. Acad. Sci. USA, 99, 15507–15512.
[4] Papavizas, G.C. and Ayers, W.A., 1974. Aphanomyces spe-cies and their root diseases in pea and sugar beet. USDA Agric. Res. Serv. Tech. Bull., 1485, 1–158.
[5] Haverson, R.M. and Rush, C.M., 1993. An environmentally controlled experiment to monitor the effect of Aphanomy-ces root rot and rhizomania on sugar beet. Phytopathol-ogy, 83, 1220–1223.
[6] Islam, M.T., Ito, T. and Tahara, S., 2001. Morphological studies on zoospores of Aphanomyces cochlioides and changes during the interaction with host materials. J. Gen. Plant Pathol., 67, 255–261.
[7] Islam, M.T., Ito, T. and Tahara, S., 2002. Microscopic stu-dies on attachment and differentiation of zoospores of the phytopathogenic fungus Aphanomyces cochlioides. J. Gen. Plant Pathol., 68, 111–117.
[8] Islam, M.T., Sakasai, M., Hashidoko, Y., Deora, A., Saki-hama, Y. and Tahara, S., 2007. Composition of culture medium influences zoosporogenesis and differentiation of Aphanomyces cochlioides. J. Gen. Plant Pathol., 73, 324–329.
[9] Horio, T., Kawabata, Y., Takayama, T., Tahara, S., Kawa-bata, J., Fukushi, Y., Nishimura, H. and Mizutani, J., 1992.
A potent attractant of zoospores of Aphanomyces cochlioides isolated from its host, Spinacia oleracea. Experientia, 48, 410–414.
[10] Islam, M.T., Ito, T. and Tahara, S., 2003. Host-specific plant signal and G-protein activator, mastoparan, triggers differentiation of zoospores of the phytopathogenic oomy-cete Aphanomyces cochlioides. Plant Soil, 255, 131–142.
[11] Islam, M.T., Hashidoko, Y., Ito, T. and Tahara, S., 2004. Interruption of the homing events of phytopathogenic Aphanomyces cochlioides zoospores by secondary metabo-lites from nonhost Amaranthus gangeticus. J. Pestic. Sci., 29, 6–14.
[12] Islam, M.T., 2008. Dynamic rearrangement of F-actin organization triggered by host-specific plant signal is lin-ked to morphogenesis of Aphanomyces cochlioides zoospo-res. Cell Motil. Cytoskel., 65, 553–562.
[13] Yokosawa, R., Kuninaga, S. and Sekizaki, H., 1986. Aphanomyces euteiches zoospore attractant isolated from pea roots prunetic. Ann. Phytopathol. Soc. Jpn., 52, 809–816.
[14] Morris, P.F. and Ward, E.W.B., 1992. chemoattraction of zoospores of the phytopathogen Phytophthora sojae by isoflavones. Physiol. Mol. Plant Pathol., 40, 17–22.
[15] Islam, M.T. and Tahara, S., 2001. Chemotaxis of fungal zoospores, with special reference to Aphanomyces coch-lioides. Biosci. Biotechnol. Biochem., 65, 1933–1948.
[16] Islam, M.T. and Tahara, S., 2005. Bioactive secondary metabolites related to life-cycle development of oomycete phytopathogenis. In: Studies in Natural Products Che-mistry (Atta-ur-Rahman, ed.), Vol. 32 (part L). Elsevier B.V. the Netherlands, pp. 1053–1122.
[17] Scott, W.W., 1961. A monograph of the genus Aphanomy-ces. Virginia Agric. Exp. Stn. Tech. Bull., 151, 1–95.
[18] Ho, H.H., Zachariah, K. and Hickman, C.J., 1968. The ultrastructure of zoospores of Phytophthora megasperma var. sojae. Can. J. Bot., 46, 37–41.
[19] Desjardins, P.R., Wang, M.C. and Bartnicki-Garcia, S., 1973. Electron microscopy of zoospores and cysts of Phy-tophthora palmivora: morphology and surface texture. Arch. Mikrobiol., 88, 61–70.
[20] Hardham, A.R., 1995. Polarity of vesicles distribution in oomycete zoospores: development of polarity and impor-tance for infection. Can. J. Bot., 73, S400–S407.
[21] Dearnaley, J.D.W., Maleszka, J. and Hardham, A.R., 1996. Synthesis of zoospore peripheral vesicles during sporula-tion of Phytophthora cinnamomi. Mycol. Res., 100, 39–48.
[22] Lunney, O.Z. and Bland, C.E., 1976. Ultrastructural obser-vations of mature and encysting zoospores of Pythium pro-liferum de Bary. Protoplasma, 90, 119–137.
[23] Fukutomi, M. and Akai, S., 1966. Fine structure of the zoospores, cystospores, and germ tubes of Sclerophthora macrospora. Trans. Mycol. Soc. Jpn., 7, 199–202.
[24] Holloway, S.A. and Heath, I.B., 1977. An ultrastructural analysis of the changes in organelle arrangement and structure between the various spore types of Saprolegnia. Can. J. Bot., 55, 1328–1339.
[25] Barr, D.J.S. and Désaulniers, N.L., 1987. Ultrastructure of the Lagena radicicola zoospore, including a comparison with the primary and secondary Saprolegnia zoospores. Can. J. Bot., 65, 2161–2176.

Journal of Basic Microbiology 2010, 50, S58–S67 Ultrastructure of asexual spores of Aphanomyces S67
© 2010 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.jbm-journal.com
[26] Hoch, H.C. and Mitchell, J.E., 1972. The ultrastructure of zoospores of Aphanomyces euteiches and of their encyst-ment and subsequent germination. Protoplasma, 75, 113–138.
[27] Hoch, H.C. and Mitchell, J.E., 1972. The ultrastructure of Aphanomyces euteiches during asexual spore formation. Phytopathology. 62, 149–160.
[28] Cerenius L, Olson L.W., Lange, L. and Söderhäll, K., 1984. Secondary zoospores of Aphanomyces astaci and Aphanomy-ces laevis. Nordic J. Bot., 4, 696–706.
[29] Olson, L.W., Cerenius, L., Lange L. and Söderhäll, K., 1984. The primary and secondary spore cyst of Aphanomy- ces (Oomycetes, Saprolegniales). Nord. J. Bot., 4, 681–696.
[30] Islam, M.T., Hashidoko, Y., Deora, A., Ito, T. and Tahara, S., 2005. Suppression of damping-off disease in host plants by the rhizoplane bacterium Lysobacter sp. strain SB-K88 is linked to plant colonization and antibiosis against soilborne Peronosporomycetes. Appl. Environ. Mi-crobiol., 71, 3786–3796.
[31] Deora, A., Hashidoko, Y., Islam, M.T., Aoyama, Y., Ito, T. and Tahara, S., 2006. An antagonistic rhizoplane bacte-rium Pseudomonas sp. strain EC-S101 physiologically stresses a spinach root rot pathogen Aphanomyces cochlioi-des. J. Gen. Plant Pathol., 72, 57–64.
[32] Afzelius, B., 1959. Electron microscopy of the sperm tail. Results obtained with a new fixative. J. Biophys. Biochem. Cytol., 5, 269–278.
[33] Iwamoto, H., 2008. Theory of diffraction from eukaryotic flagellar axonemes. Cell Motil. Cytoskel., 65, 563–571.
[34] Crump, E. and Branton, D., 1966. Behavior of primary and secondary zoospores of Saprolegnia sp. Can. J. Bot., 44, 1393–1400.
[35] Hemmes, D.E. and Hohl, H.R., 1971. Ultrastructural as-pects of encystations and cyst germination in Phytophthora parasitica. J. Cell Sci., 9, 175–191.
[36] Beakes, G.W., 1994. Sporulation of lower fungi. In: The Growing Fungus (Gow, N.A.R., Gadd, G.M., eds.). London: Chapman and Hall, pp. 339–366.
[37] Hemmes, D.R., 1983. Cytology of Phytophthora. In: Phy-tophthora: Its Biology, Taxonomy, Ecology and Pathology (Erwin DC, Bartnicki-Garcia S., Tsao P.H., eds.). St. Paul, Minnesota: American Phytopathological Society, pp. 9–40.
[38] Bartnicki-Garcia, S. and Wang, M.C., 1983. Biochemical aspects of morphogenesis in Phytophthora. In: Phytophthora: Its Biology, Taxonomy, Ecology and Pathology (Erwin DC, Bartnicki-Garcia S., Tsao P.H., eds.). St. Paul, Minnesota: American Phytopathological Society, pp. 121–137.
[39] Chilvers, G.A., Lapeyrie, F.F. and Douglass P.A., 1985. A contrast between Oomycetes and other taxa of mycelial fungi in regard to metachromatic granule formation. New Phytol., 99, 203–210.
[40] McBride, H.M., Neuspiel, M. and Wasiak, S., 2006. Mito-chondria: more than just a powerhouse. Curr. Biol., 16, R551.
[41] Liu, X., Weaver, D., Shirihai, O. and Hajnóczky, G., 2009. Mitochondrial ‘kiss-and-run’: interplay between mito-chondrial motility and fusion-fission dynamics. EMBO J., 28, 3074–3089.
Copyright © 2022 FDOKUMEN




















