
The ultrastructure of transplanted rabbit retinal epithelium
Upload
independentCategory
view
2download
0
This article was downloaded by: [175.177.159.153]On: 21 March 2014, At: 06:02Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH,UK
Caryologia: InternationalJournal of Cytology,Cytosystematics andCytogeneticsPublication details, including instructions forauthors and subscription information:http://www.tandfonline.com/loi/tcar20
The Ultrastructure of PollenGrain Development inLycopersicum PeruvianumEttore Pacinia & Barrie Juniperb
a Istituto di Botanica, Università di Siena, Siena,Italyb Botany School, Oxford University, Oxford, GreatBritain.Published online: 31 Jan 2014.
To cite this article: Ettore Pacini & Barrie Juniper (1984) The Ultrastructure ofPollen Grain Development in Lycopersicum Peruvianum, Caryologia: InternationalJournal of Cytology, Cytosystematics and Cytogenetics, 37:1-2, 21-50, DOI:10.1080/00087114.1984.10797684
To link to this article: http://dx.doi.org/10.1080/00087114.1984.10797684
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all theinformation (the “Content”) contained in the publications on our platform.However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness,or suitability for any purpose of the Content. Any opinions and viewsexpressed in this publication are the opinions and views of the authors, andare not the views of or endorsed by Taylor & Francis. The accuracy of theContent should not be relied upon and should be independently verified with

primary sources of information. Taylor and Francis shall not be liable for anylosses, actions, claims, proceedings, demands, costs, expenses, damages,and other liabilities whatsoever or howsoever caused arising directly orindirectly in connection with, in relation to or arising out of the use of theContent.
This article may be used for research, teaching, and private study purposes.Any substantial or systematic reproduction, redistribution, reselling, loan,sub-licensing, systematic supply, or distribution in any form to anyone isexpressly forbidden. Terms & Conditions of access and use can be found athttp://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

CARYOLOGIA Vol. 37, n. 1-2: 21-50, 1984
THE ULTRASTRUCTURE OF POLLEN GRAIN DEVELOPMENT IN LYCOPERSICUM PER UVIANUM
ETTORE PACINI and BARRIE JUNIPER* Istituto di Botanica, Universita di Siena, Siena, Italy, and *Botany School, Oxford University, Oxford, Great Britain.
SUMMARY- The ultrastructure of the development of the pollen grain is described from the first meiotic division to the binucleate phase. The first meiotic division is synchronous in all the microspore mother cells and cytomictic channels still join their protoplasts together. The tapetal cells, their individual walls now degraded, almost completely surround but do not touch the microspore mother cells. In the latter the cytoplasm can be divided into two zones. The central zone, the division zone, is occupied primarily by ribosomes, smoothER and dictyosomes. In the outer zone lie all the other usual organelles and small vacuoles. The plastids are partly surrounded by ER. In interphase the cytomictic channels close and the second division is not synchronous. Autophagic vacuoles form, apparently destroying some of the ribosomes. The callose walls form after meiosis and the tapetum degenerates. The primexine, which appears to have a polysaccharide skeleton, begins to form between the plasmalemma and the callose wall. The callose wall then dissolves, the pores begin to form, sporopollenin begin to accumulate and the exine is completed. The spores then pass through a stage of vacuolation and of amylogenesis and loss of starch. At mitosis the new wall separating the vegetative and generative cells is formed, and it is not perforated by plasmodesmata. The pore structure is now completed and autophagic vacuoles destroy further regions both of the vegetative cell's cytoplasm and to a lesser extent that of the generative cell. New cytoplasm is successively formed, together with another wave of amylogenesis and loss of starch; these two processes occur simultaneously with the dehydration of the grain as the pollen structure matures.
INTRODUCTION
The basic ultrastructural features of the first phase (the meiotic prophase) and the final phase (the ripe pollen phase) of Lycopersicum peruvianum have already been described (PACINI and CRESTI 1978; CRESTI et al. 1975).
PACINI and CREST! (1976) have also observed an association between the plastids and endoplasmic reticulum (ER) in the same species from the end of meiosis to the first haploid mitosis.
In addition a complete reproductive calendar from flower differentiation to seed ripening has been drawn up (PACINI and SARFATTI 1978). In this paper
Research work supported by CNR, Italy. Special grant I.P.R.A.- Sub-project 1. Paper N. 130.
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

22 PACINI and JUNIPER
we complete the missing sections of the ultrastructural description of pollen grain development in L. peruvianum and compare both the new and the old findings.
MATERIALS AND METHODS
PACINI and SARFATTI (1978) showed that there was a precise correlation between bud length on the one hand and microspore and pollen development on the other. Anthers could therefore easily be collected at the required stages. A further check was made of smears in acetic orcein under the light microscope. The material was then collected and treated as previously described (PACINI and CREST! 1976, 1978).
RESULTS
The period of development has been arbitrarily divided into three to aid description namely meiosis, the spore stage and the binucleate pollen grain stage. These phases are not equal in time and the details of the stages of pollen grain development are summarized in Table 1.
Meiosis.
Prophase. - The details of the me10t1c prophase have already been reported (PACINI and CRESTI 1978) and are only summarised here. At prophase the anther loculus has already formed. All the plasmodesmata between the microspore mother cells (mMc), and the tapetal cells have been occluded, but not all of those between the mMCs themselves. Some of the latter become cytomictic channels. The mMCs become isolated and the callosic wall gradually develops between their primary walls and the plasmalemma. In the mMC cytoplasm small vesicles, probably derived from the dictyosomes, can be seen near the plasmalemma together with ER profiles and microtubules. The latter are sometimes single, sometimes in bundles, running from the nuclear membrane to the plasmalemma and orthogonal to the cell wall. Other cytoplasmic features are roundish mitochondria, undifferentiated plastids, some rough ER (RER) with their profiles sometimes dilated at their tips, numerous apparently not very active dictysomes, and a few small round vacuoles.
First meiotic division. - After prophase the rest of the meiotic process is very rapid. The floral bud is about 4.5 mm long at this point, just when the corolla is beginning to outgrow the calyx. The first metiotic division (Figs. 1 and 2) is synchronous in all the mMCs and the cytomictic channels are still present. The tapetal cells, although free from the mMC, remain in close proximity by becoming progressively cup shaped and partly surrounding the neighbouring mMC (Fig. 3).
During the first meiotic division two distinct cellular areas can be seen; a
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

TA
BL
E
1 -
Eve
nt a
nd S
tage
s du
ring
th
e de
velo
pmen
t of
«Ly
cope
rsic
um
peru
vian
um».
Rad
ially
arr
ange
d m
icro
tubu
les
Cal
losi
c w
all
form
atio
n C
ytom
icti
c ch
anne
ls p
rese
nt
Pri
mex
ine
form
atio
n E
xine
com
plet
ion
PE
R
.... -~ 81J
::l
....
"§ ·1
3 o
..c
"' "'
"' ....
.<::
"'
u:::::
<:.e pr
opha
se
c .8 ~ ·~
·~ 8
inte
rpha
se
ME
IOC
YT
E
c -~
:~
"0
.!:l 0 . ., 8 - - te
trad
s
c .g "" ~ 0.. "' "' [ cs ~
mon
onuc
leat
e ga
met
ophy
te
MIC
RO
S P
OR
E
t--
--
"' cs ·§ "0
·a ~
.<:: - bi
nucl
eate
gam
etop
hyte
PO
LL
EN
G
RA
IN
1-
- --
"' u c ~ ~ .... .fi c <
Vac
uola
tion
t-~------
-~ --
-A
myl
ogen
esis
--
--
--
-In
tine
for
mat
ion
--
-E
R s
tack
s --
Deh
ydra
tion
---
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

24 PACINI and JUNIPER
central zone where the divisions occur and an outer region. The central zone is rich in ribosomes, most of them aggregated into spirally arranged polyribosomes, together with dilated smooth ER cisternae. Many apparently very active dictyosomes are located in the peripheral part of the central zone (Fig. 1).
The outer zone of the cytoplasm contains numerous roundish mitochondria, short profiles of ER, small vacuoles, and proplastids. Short cisternae of ER begin to surround some of the proplastids. These associated structures (PER) have already been described (PACINI and CRESTI 1976). They are present from the first meiotic division to the first haploid mitosis and may, at certain stages, involve all the microspore plastids. At telophase of this division no wall is formed in the meiocyte.
Interphase. - During this interphase the cytomictic channels, which had formed during the prophase, become occluded. The organelles which could be seen outside the division zone during the first meiosis now appear in the centre of the cell between the two nuclei. Only ribosomes can now be seen in the peripheral cytoplasm (Fig. 3).
Second meiotic division. - This division, unlike the first, is not synchronous in all the meiocytes. Even within one meiocyte the two free nuclei seem to divide independently so that one can find, side by side, one nucleus completing telophase and one just starting prophase. However, in 30-40% of the cases the two divisions do occur synchronously (Fig. 4). The axes of the mitotic spindles are always at right angles to one another (Fig. 4). The main changes observed in the cytoplasmic organelles appear to be an increase in the number of the plastids and the development to what seem to be autophagic vacuoles. These vacuoles seem to be formed when an ER cisterna wraps around a spherical cytoplasmic region. In section this cisternal profile appears as two circular concentric sections, the inner one then disaggregated and disappeared. The enclosed cytoplasm consists mainly of ribosomes which too disappear giving rise to a vacuole (Fig. 5). During the second meiotic division the cup-shaped tapetal cells are still seen to be closely surrounding the neighbouring meiocytes and tetrads.
Tetrad stage. The succeeding events are here again arbitrarily subdivided, but some of
the phenomena described here as if sequential may sometimes occur at the same time.
Immediately after meiosis the callose walls start to form which will separate the microspores (Fig. 6). At first small vesicles, which probably originate from the nearby active dictyosomes, seem to unite. Then small, apparently callosic accumulations appear surrounded by numerous polyribosomes (Fig. 7). These accumulations increase in size and unite to form a callosic wall. This new wall is about one third of the thickness of that of the microspore
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

POLLEN GRAIN DEVELOPMENT IN LYCOPERSICUM PERUVIANUM 25
mother cell wall from which it is now indistinguishable (Fig. 8). Plastids now appear to be more numerous, increase in size and keep dividing. They are lobed and all appear to have PER. Almost all the ER at this stage appears to be in PER type associations. Mitochondria also appear to be more numerous than during the meiosis.
Soon after the completion of this callosic wall the pectocellulosic microspore mother cell wall, which is still present on the surface of the tetrads, disappears and in its place the primexine is formed. Primexine is a structure formed between the callosic wall and the plasmalemma and is so called because it is the spatial precursor of exine (HESLOP-HARRISON 1963). This structure appears to have a polysaccharide skeleton acting as a template for the other components of the protosporopollenin. The polysaccharide skeleton is Thiery positive (Fig. 9). The primexine (except in the poral region) forms a slightly electron dense layer above the plasmalemma, covered with short electron dense rods (the future bacula). The outer ends of these rods spread laterally to touch one another forming a layer which will give rise to the tectum of the mature pollen (Figs. 10 and 11). Between the bacula there is a granular, electron dense, Thiery positive material which appears to act as a template (Fig. 12).
The spore.
The length of this stage varies according to when the first haploid mitosis occurs. While the microspore is maturing the tapetum appears to continue its 'nurse' role and finally degenerates.
Microspore release. - During primexine formation (tetrad stage) the exterior of the mMC callosic wall becomes spongy (Fig. 10). Then gradually all the callosic walls become less electron dense and granular (Figs. 11 and 13) and in the end dissolve freeing the microspores. At this point the germination pores of the pollen become apparent as dome-shaped regions crossed by electrondense filaments extending from the inner layer of the primexine (Fig. 14). Only the inner layer of the primexine is preent under the pollen furrows (Fig. 15). During the gradual dissolution of the callose wall the shape of the microspores changes from tetrahedal to round. Many microtubules both single and grouped can be seen at this stage (Figs. 16 and 17).
The completion of the exine.- The remains of the callose wall persist for a while on the surface of the microspores (Figs. 18 and 19) together with the Thiery positive material between the bacula (Figs. 18 and 19). As soon as the microspores are released the thickening of the basal layer and tectum begins. Polyribosomes are conspicuous in the cytoplasm near to the plasmalemma and often correspond in their groupings to the bacula (Fig. 18). The basal layer thickens by the development of high and low electron-dense lamina. A granular zone develops between the newly formed basal layer and the slightly convoluted plasmalemma (Fig. 20). The tectum too becomes thicker and on its outside appear clumps of not yet completely polymerized sporopollenin (Figs. 20 and
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

26 PACINI and JUNIPER
21) which progressively become organized into the familiar spiny surface of the pollen grain (Figs. 22 and 28). The basal layer is complete before the tectum (Fig. 21) and very often some helicoidal polyribosomes can be seen with their axis parallel to and close to the plasmalemma (Fig. 21). As the tapetum starts to degenerate the exine is completed (Fig. 22). Orbicles (arrowed} have already begun to appear on the surface of the tapetal cells (Fig. 22).
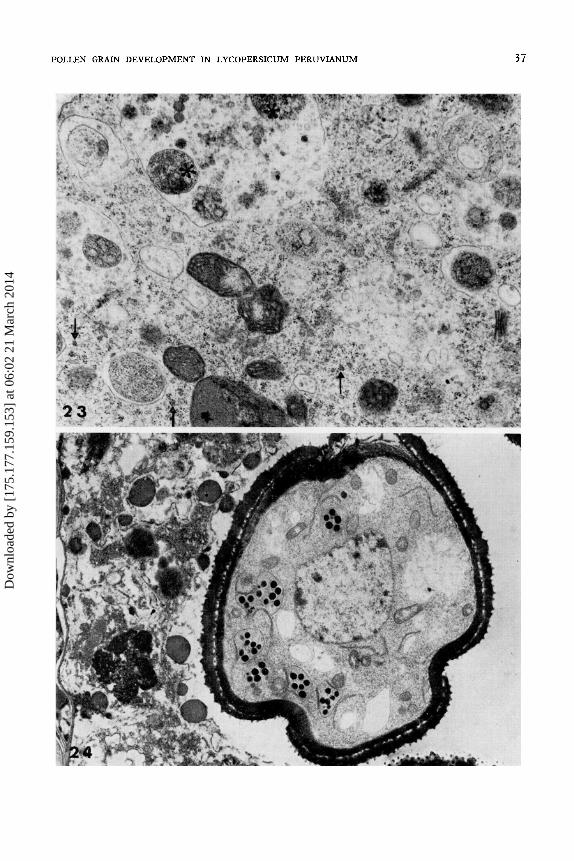
First vacuolation of the microspore. - This period of vacuolation starts during the second meiotic division (Fig. 5) and lasts through the whole of the tetrad stage. The process becomes more pronounced as the microspores are released from the callosic wall. The separate vacuoles displace larger and larger areas of cytoplasm and some of them coalesce (Fig. 23). But within the same microspore several different phases of cytoplasmic degeneration can be observed (Fig. 23). During the first vacuolation period about 20% of the cytoplasm seems to degenerate (Fig. 24). The cytoplasmic area not ivolved in vacuolation contains both dictyosomes and polyribosomes which, unlike the free ribosomes, do not seem to be affected by the general degeneration (Fig. 23).
First amy/agenesis. - Plastids start to accumulate starch during the late vacuolation phase and seem to occupy about 10% of the microspore volume (Figs. 24 and 25). The amyloplasts contain lamellae and 8-12 electron dense starch grains. They are no longer eliptical, but more or less round. PER is now less frequently seen and the ER cisternae are only adjacent to about 15% of the plastid envelope. At the same time profiles of free ER cisternae are abundant. The mitochondria elongate and electron dens globules, possibly lipid droplets, appear in the cytoplasm (Figs. 24 and 25). Amylogenesis is not synchronous in all the micros pores of the same anther. The starch grains may disappear just before or sometimes during the first haploid mitosis. The degeneration of the tapetal cells proceeds all through this first starch formation and during the amylolysis (Fig. 24). By the first mitosis the organelles are no longer recognisable and the nuclei are pycnotic.
Intine production and the initial formation of the pore. -Towards the end of microspore development, while the exine is being formed and vacuolation and loss of the starch grains continue, both the intine and the polysaccharide part of the pore develop. The plasmalemma detaches from the basal layer starting at the poral zone border and this detachment proceeds both inwards from the poral zone and sideways to the interporal zone. At this stage single microtubules can be seen parallel to the cell wall (Fig. 28). In the interporal zone, between the plasmalemma and the exine, Thiery positive material accumulates. The intine as well as the polysaccharide part of the pore seem to originate from this material. lntine formation stops shortly after the first haploid mitosis. The mature microspore now has two walls; the intine Thiery positive and the exine Thiery negative except for the primexine residues and the very extensive layer of wall sculpturing (Fig. 29).
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

POLLEN GRAIN DEVELOPMENT IN LYCOPERSICUM PERUVIANUM 27
The pore is now in two parts; ~ polysaccharide inner zone which is a continuation of the intine. The polysaccharide part is thicker than the intine layer and appears in cross-section as a crescent. In contrast the exine layers are slender and irregular and the tectum is not even continuous. There are no bacula over this zone and the basal layer is represented by a few lamina which form a fringe to the normal interporal foot layer (Fig. 26). At the beginning of the pore development, while the crescent-shaped polysaccharide layer is forming but is still electron translucent and weakly Thiery negative, the plasmalemma evaginates and forms numeros, sometimes branched tubules penetrating into the crescent-shaped layer as far as the exine (Fig. 27). Many ER cisternae can be seen in the cytoplasm facing these plasmalemma tubules (Fig. 27).
The binucleate pollen grain.
The longest stage of pollen grain development comprises both its ripening and complete tapetal degeneration. The main features are a second vacuolation and amylogenesis.The latter is again followed by an amylolysis, the disappearence of the characteristic PER, the pore completion, formation of ER stacks and the final dehydration.
First haploid mitosis. - This mitosis, which is not synchronous in all the grains, can be found when the length of the floral bud is about 5.25-7.5 mm. After the first vacuolation, at the end of the preceding stage, a newly formed cytoplasm appears while vacuoles are much less numerous. Free cisternae apparently increase in number while PER decreases and only a few plastids still contain starch grains. Just before the first haploid mitosis the nucleus moves to one side of the microspore, long ER cisternae surround the division zone during metaphase, anaphase and early telophase (Fig. 30). The spindle axis is always at right angles to the cell wall. At telophase a new wall separating the vegetative and generative cells is formed by the fusion of vesicles probably coming from the dictyosomes (Fig. 31). This wall has no plasmodesmata and is relatively thin; nervetheless it contains polysaccharides (Fig. 32). In longitudinal section the new generative cell is crescent-shaped whilst in cross section it is more or less round (Fig. 32) like the nucleus inside it.
The completion of the pore. - The pore is finally completed after the first haploid mitosis. Thiery positive polysaccharide material accumulates in the main pore region between the fringe-like exine lamellae and the tubules forming the plasmalemma. Seen in cross section the pore now has the shape of a ben spindle with its convex side facing the cytoplasm (Fig. 33). In the fully mature pore, the polysaccharide material is slightly granular and the convex side is now turned outwards (Fig. 35) in the shape of an umbo; the same shape that can be observed in the hydrated mature pollen grain.
Second vacuolation. - This process is similar to that of the first vacuolation; disaggregagion takes place via what appear to be autophagic vacuoles
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

28 PACINI and JUNIPER
sequestering the ribosomes and lipid globules (Fig. 34). These autophagic vacuoles coalesce into one single large vacuole which fills 40-50% of the grain volume. This extensive vacuolation pushes the vegetative nucleus and the generative cell towards the cell wall, along with the remaining cytoplasm. Some granular material can be seen inside this large central vacuole (Fig. 35).
Second amylogenesis. - This phase is preceded by an increase in the volume of cytoplasm following the second vacuolation and by an apparent increase in the number of plastids. Each amyloplast in section has 3 or 4 moderately electron dense starch grains (Fig. 36). The most starch seems to be present when the floral bud is 10.5 mm long and at this point the amyloplasts appear to occupy 30-40% of the grain volume. At the same time the mitochondria are all located near the cell wall and the ER cisternae appear to become more numerous and are arranged parallel to each other. Then the amylolysis starts lasting until the floral bud is 11.5 mm long.
The changes in the generative cell. - When first formed the generative cell adheres to the grain wall. After the second vacuolation the wall separating it from the vegetative cell is completed by a thinner and less Thiery positive region. The generative cell now detaches from the intine and moves to the centre of the grain. It is now spindle-shaped and its cytoplasm has mitochondria, dictyosomes, ER cisternae, but apparently no plastids. Its cytoplasm is vetigial and a small amount of it is apparently destroyed by the autophagic vacuoles (Fig. 32).
The dehydration stage. - This occurs during the last stage of pollen development. The tapetal cells have now completely collapsed, leaving only some small electron dense globules, probably lipid, deposited on the exine. The orbicles, formed in the previous stage, also persist, but are now in direct contact with the inner sides of the anther wall layer. Pollen dehydration begins during the second vacuolation and continues through the second stage of starch deposition. The cytoplasm inside the grain darkens, the polyribosomes start to dissociate and the other organelles become less distinct (Figs. 35, 36 and 37) except for the ER cisternae which gather together into stacks (Figs. 36-37).
The mature grain. - The various changes occuring synchronously with
Fig. 1. - Metaphase of the first meiotic division. All the organelles are located between the callose wall (CW) and the zone of division. In this division zone virtually only ribosomes are visible, whilst on the zone's periphery are some dilated cisternae (asterisks) and dictyosomes (D). Some spindle fibres are indicated by arrows. X 12,000. Fig. 2. - Detail of the microspore cytoplasm during the first meiotic division. A plastid (P), mitochondria (M) and small vacuoles (V) are discernible. Two ER cisternae, (arrowed) are dilated at their extremities and appear to be producing vesicles. X 32,400. Fig. 3. -Interphase at the end of the first meiotic division. The organelles are now in the centre of the meiocyte whilst the ribosomes are clustered around the periphery. The tapetal cells are concave where they juxtapose a meiocyte. X 3,200.
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

POllEN GRAIN DEVELOPMENT IN LYCOPERSICUM PERUVIANUM 29
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

Fig. 4.- The second meiotic division. The organelles are now localized in the space between the two divisions. The meiotic spindles are orientated at right angles to one another. The tapetum as in Fig. 3 is concave around the surface of the meiocyte. X 3,200. Fig. 5. - A detail of the cytoplasm during the second meiotic division. The plastids (P) are undifferentiated. The matrix of each mitochondrion appears more electron dense than that of the plastids. Inside a vacuole a small portion of cytoplasm is apparently dissolving. X 36,000. Figs. 6, 7 and 8. - Formation of the callose wall between the microspores. Fig. 6. - The formation of the callose wall is centripetal. Numerous vesicles, probably the product of dictyosomes (arrows) appear to be fusing. There are also a few callose masses (arrow heads), polyribosomes are present in the cytoplasm near the growing wall. X 18,000. Fig. 7. - The wall is not quite complete. Many polyribosomes are visible adjacent to it. X 18,000. Fig. 8. - The callose wall separating the microspores has just formed. It is about one third of the thickness of the microspore mother cell wall from which it cannot be distinguished. X 18,000.
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

POLLEN GRAIN DEVELOPMENT IN LYCOPERSICUM PERUVIANUM 31
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

Figs. 9, 10, 11 and 12. - Primexine formation. Fig. 9. -The formation of primexine begins. Thiery stained section. The space between the callosic wall (CW) and the plasmalemma of each microspore is irregularly positive to the Thiery stain. Outside the callosic wall, which does not react, the primary pectocellulosic wall (arrow) is still present and reacts positively. A tapetal cell (T) is visible in the upper part of the figure. X 10,800. Fig. 10. -Mature primexine. Note the difference between the primexine in the interporal zone and that in the pore. The primary microspore mother cell wall has disappeared on the tetrad surface and the underlying callosic wall is now spongy. X 10,800. Fig. 11. - Primexine detail. Some of the detail of the mature exine can already be seen: tectum (long arrows), bacula (short arrows). Between one bacula and another an electrodense granular material has developed. Note the spongy nature of the callosic wall (CW). X 43,000. Fig. 12. - The primexine sectioned tangentially and at the same stage as Fig. 11. Stained with Thiery's reagent. The bacula do not react to the Thiery test (arrow head); the granular material between the bacula is positive. (CW) callosic wall. X 45,000. Fig. 13. - Microspore release. The callosic wall has almost disappeared; only a few weakly electrondense traces remain. X 7,500. Fig. 14. - Detail of pore after separation of the microspores. The development of the exine is beginning to be apparent in the foot layer which forms by flat extensions from the pore margin. X 28,000. Fig. 15.- Exine cross section at the furrow level after the separation of the microspores. The exine in the furrow consists only of the foot layer. X 28,000. Figs. 16 and 17. - Microtubules during the young microspore stage. Fig. 16.- Bundles of microtubules almost normal to the plane of the plasmamembrane (arrow heads). One of the microtubules ends against the plasmamembrane (arrow). X 28,000. Fig. 17. - Bundles of micro tubules (arrow heads) near to and approximately normal to a glancing section of the nuclear membrane. Many pores are visible in the nuclear membrane. X 32,000.
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

POLLEN GRAIN DEVELOPMENT IN LYCOPERSICUM PERUVIANUM 33
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

Figs. 18, 19, 20 and 21. - Stages in the completion of the exine. Fig. 18. - The foot layer is thicker now than in Fig. 11 and the electrondense granular material between the bacula is still present. Polyribosomes are frequent in the cytoplasm near the plasmamembrane. Traces of the original callosic wall are still present outside the exine. X 22,000. Fig. 19. - Thiery stained section. Microspore at the same stage as in the preceding figure. Thiery positive polysaccharide material between the bacula is still present. X 22,000. Fig. 20. -The foot layer now becoming thicker. Weak electron staining and strong electron staining layers are now being deposited alternately. Plasmamembrane is undulant. X 22,600. Fig. 21. - The thickness of the exine, but not its final architecture, is now apparent. The tectum is sculptured, but not yet complete. A helical polyribosome (arrow) and a microtubule more or less normal to the plasmamembrane (arrow head) can be seen. X 23,000. Fig. 22. - A microspore and tapetal cells with the exine now complete. The degeneration of the tapetal cells has already begun. On the plasmamembrane surface facing the loculus many orbicles can now be seen (arrows). The plastids of the microspores have a characteristic curved shape. X 7,200.
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

POLLEN GRAIN DEVELOPMENT IN LYCOPERSICUM PERUVIANUM 35
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

Fig. 23. - First vacuolation and the formation of autophagic vacuoles. Many autophagic vacuoles in different stages of development can be seen. Occasionally vacuoles in a different state of development can be seen joining one another (asterisks). Polyribosomes (arrows) can clearly be seen in the cytoplasm not affected by the action of the autophagic vacuoles. X 20,000. Fig. 24. -The end of the first vacuolation and the first amylogenesis. The light areas show that the autophagic vacuoles appear to have destroyed about 20% of the young rnicrospore cytoplasm. ER cisternae, amyloplasts with strongly electron dense starch grains and elongated mitochondria can clearly be seen. The tapetal cells are now at an advanced stage of degeneration. X 7,500.
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014
POLLEN GRAIN DEVELOPMENT IN LYCOPERSICUM PERUVIANUM 37
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

Fig. 25. - Plastids at the first amylogenesis stage. Amyloplast sections show at least eight electron dense starch grains and some plastid membranes (arrow heads). ER cisternae are either free from cytoplasm or closely associated with the outer of the amyloplast membranes forming PER (arrow). What appears to be the remains of an autophagic vacuole (A V) and electron dense globules, lipid bodies (LB), are also present. X 20,400. Figs. 26 and 27. - Pore formation. Fig. 26. - The plasmamembrane starts to pull away from the exine beginning at the periphery of the pore. The foot layer penetrates into the cap of the pore through slender lamella ted extensions. Tectum and bacula are not present. X 18,000. Fig. 27. -The formation of the plasmamembrane evaginations at the pore zone. As the plasmamembrane retracts from the pore it forms extensive and sometimes branched tubules. The space between the plasmamembrane and exine can now be seen to be slightly granular and electron dense. ER cisternae can be seen in the cytoplasm paralleling the plaslamembrane. X 20,000. Figs. 28 and 29. - Intine formation. Fig. 28. - The plasmalemma can be seen retracting away from the foot layer in small areas, a thin and weakly electron dense granular deposit. A microtubule (arrow) can be seen parallel to the intine. X
30,600. Fig. 29. - At a similar stage to the preceding figure, but stained with Thiery's reagent. The thin intine is Thiery positive. Material, staining as if it were polysaccharide, is evident between the bacula and the surface of the exine sculptures. X 30,600.
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

POLLEN GRAIN DEVELOPMENT IN LYCOPERSICUM PERUVIANUM 39
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

Fig. 30. - Metaphase of the first haploid mitosis. The spindle is at right angles to the wall and the division area is surrounded by many long profiles of ER. X 12,600. Fig. 31. - At the end of the first haploid mitosis. The vegetative and the generative nuclei lie close to one another. The wall which will separate them is being formed centrifugally and is indicated by lines with dot extensions. X 16,200.
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

POLLEN GRAIN DEVELOPMENT IN LYCOPERSICUM PERUVIANUM 41
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

Fig. 32. - The vegetative cell and the generative cell after the wall separating them is complete. Stained with Thiery's reagent. The generative cell is seen in cross section. The new wall is slightly less responsive to the Thiery reagent than the intine (arrow). ER cisternae, mitochondria and what is probably an autophagic vacuole (asterisk) are present in the generative cell cytoplasm. X 10,200. Fig. 33. - Detail of the pollen pore at the same stage as the preceding figure. The pore at this stage is in the shape of a crescent with its convex side orientated towards the centre of the grain. X 13,200. Fig. 34.- Second vacuolation. Vacuoles contain material which is not yet disaggregated. The plastids (P) are starting to accumulate the starch of the second amylogenesis stage. ER cisternae are now more abundant. In the right hand bottom corner of the figure is the generative cell (GC). X 12,000. Fig. 35. - Detail of the vegetative nucleus (vn) and the pore during the second vacuolation. The cytoplasm at this stage is reduced to a peripheral strip. The large vacuole (V) has a granular content. The pore emerges from the surface of the grain as an umbo. X 10,200.
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

POLLEN GRAIN DEVELOPMENT IN LYCOPERSICUM PERUVIANUM 43
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

Fig. 36. - Detail of the cytoplasm of the vegetative cell during the second amylogenesis. (VN) the vegetative nucleus. Plastids have increased in size and apparently in number and all appear to have starch grains. Mitochondria are mostly located near the grain wall. ER cisternae have become more conspicuous and appear as parallel stacks. X 18,000. Fig. 37.- The cytoplasms of the vegetative and generative cells (GC) in the mature grain. Some ER cisternae can be seen to be gathered together in parallel stacks and some surround what appear to be lipid globules. (LB). The plastids (P) no longer contain starch. X 20,000.
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

POLLEN GRAIN DEVELOPMENT IN LYCOPERSICUM PERUVIANUM 45
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

46 PACINI and JUNIPER
dehydration contribute to the appearance of the mature grain, which has been described in a previous paper (CRESTI et al., 1975). The principal ultrastructural features of the mature grain are: a small number of plastids, peripherally arranged mitochondria, numerous ribosomes, dictyosomes mostly near the cell wall and a lobed vegetative nucleus lying alongside the generative cell in the centre of the grain. Numerous stacks of cisternae of RER are prominent and these often surround osmiophilic, presumably lipid, bodies. The generative cell is separated from the cytoplasm of the vegetative cell by an an indented wall. Its nucleus appears to have no nucleolus. Its cytoplasm contains many ribosomes, dictyosomes, a few mitochondria and some small ER cisternae always located near the nuclear envelope. About 1% of the pollen grains when shed appear to be empty; their degeneration occurs during the microspore or binucleate stage. The mature pollen grains are then released through the apex of the anther.
DISCUSSION
Meiosis.
During meiosis two different phenomena are influencing the state of the new cells. The fundamental genetic reorganisation brings about the new haploid condition. In the new haploid state no significant changes are noticeable in the shape of the mitochondria, but they increase in number during interphase. The plastids too increase and associate with the ER to form the characteristic PER (PACINI and CREST! 1976). PER seems to be correlated with the contribution of the microspore to the formation of the exine. The other major phenomena of this period of development are the predetermination of the shape of the microspores and the positioning of the germination pore. Both of these depend fundamentally on the spindle orientation of the second meiotic division. If the orientation of the second meiotic division spindle is mechanically distorted the morphology of the pollen grain and the position of the pore is changed (HESLOP-HARRISON 1971).
The walls separating the tetrads are formed like those of the mMCs by the union of vesicles probably derived from the dictyosomes (PACINI and CREST! 1978). At this stage the polyribosomes are in rosettes, whilst on the other hand during the formation of the callosic microspore mother cell wall they are spirally arranged. Another difference which can be observed is that, at first, inside the tetrads, the callose masses are small, but later on they gather together to form a continuous wall. In the microspore mother cell, on the other hand, the callosic wall is immediately continuous and later on just thickens. This distinction is probably based on the fact that the microspore mother cell callosic wall is an accessory wall formed beneath the primary pectocellulose wall, whilst the walls separating the tetrads are formed after a division.
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

POLLEN GRAIN DEVELOPMENT IN LYCOPERSICUM PERUVIANUM 47
The autophagic vacuoles form during the second meiosis and they reach their full development during the microspore phase. They will be discussed in detail later.
The tetrad stage. - The most important event of the tetrad stage is primexine formation. Primexine has two constituents, the first is a template on which the second constituent, protosporopollenin develops (HESLOP-HARRISON 1968). The latter is the special precursor of the mature exine which is composed of sporopollenin. The template, on the other hand, is apparently cellulose and its elimination by cellulase inhibits exine formation (HESLOPHARRISON 1971) . The primexine has a uniform pattern all over the microspore surface, except over the regions which are to become the generative pores and furrows. According to DICKINSON (1976) the «membranous shield laid down under the cytoplasm» seems to protect each pore against the formation of primexine. We have never seen this phenomenon in Lycopersicum pollen, but it is present in Olea europea (PACINI, unpublished data).
The microspore stage. - In addtion to completing the exine this phase also includes a change of shape, a vacuolation and an amylogenesis. Both vacuolation and amylogenesis will occur again during the development of the pollen grain.
Microtubules, radially arranged, i.e. orthogonal to the plasmamembrane, appear twice during the formation of Lycopersicum pollen grains. They first appear before meiosis, when microspore mother cells become isolated and form the callosic wall (PACINI and CREST! 1978), and again after the release of the microspore. Many authors have described microtubules arranged in different ways during the initial microspore phases and have interpreted them in a variety of ways (ECHLIN and GoDWIN 1969). However, we are of the opinion that the microtubules are connected with the alteration of the shape of the microspores and of the microspore mother cells.
Vacuolation also occurs twice and this phenomenon always appear to derive from autophagic vacuoles apparently generated from the ER cisternae, and enclosing parts of the cytoplasm. The enclosed cytoplasmic regions always contain ribosomes and after the first haploid mitosis a few lipid globules. MEPHAM and LANE (1970) in Tradescantia and EcHLIN (1972) in Helleborus considered that the ER was responsible for the vacuolation of microspores, but did not describe or interpret the autophagic vacuoles.
According to MATILE (1975) «autophagic activity is particularly conspicuous in cells which undergo extensive differentiation». In this instance only the ribosomes appear to be suffering a significant change whilst the organelles differentiate and dedifferentiate independently. The two environments in which the autophagic vacuoles develop during Lycopersicum pollen development are very different. The ribosomes coded by the microspore mother cell are probably destroyed during the second meiotic division and the tetrad phase. After microspore release, however, the destruction of ribosomes of
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

48 PACINI and JUNIPER
diploid origin may continue as well as the destruction of ribosomes of haploid origin from the new haploid nuclei. The transistion from the diploid phase to the haploid phase is different in Lycopersicum peruvianum to that in Lilium (DICKINSON and HESLOP-HARRISON 1977). In the latter no autophagic vacuoles are present and the production of new ribosomes is favoured by the disaggregagion of «nucleoloids» produced by the nucleolar organizer during meiotic prohase.
Two separate vacuolations are observed in the pollen grain after the first haploid mitosis namely in the vegetative cell and in the generative cell. Autophagic vacuoles of the vegetative cell are larger than those of the generative cell probably because the cytoplasm of the former is more extensive. We suspect that autophagic vacuoles are formed in both cells because the microspore cell, via the first haploid mitosis, gives rise to two highly differentiated cells with different functions. After such an irregular division the cytoplasm no longer corresponds to the vegetative and generative nuclei. It is therefore destroyed to allow the creation of a new environment to conform to the new nuclei. The two stages of amylogenesis occur slightly later than vacuolation and again are not identical. Both the size of the starch grains and their electron density and abundance are different. Amyloplasts formed in the microspore contain many highly electron dense starch grains and their morphology resembles that of amyloplasts in the sieve elements. In the binucleate grain, on the other hand, they resemble the amyloplasts of the anther wall and other floral parts. It is not clear why these amyloplasts should be morphologically different from one another, as their source material, when hydrolysed, apparently produces the same substrate. We can only speculate that the two types of starch grains have a different rate of metabolism.
The Pollen Grain. -The irst haploid mitosis is not synchronous in all the microspores of an anther. A thin dividing wall is always centrifugally formed as observed by ANGOLD (1965) in Endymion and by GoRSKA-BRYLASS (1970) in various species. In Helleborus EcHLIN (1972) noted that the formation of this new cell wall is centrifugal, but the thickening is centripetal. A striking feature of the cytoplasm during the first haploid mit~sis is the presence of ER cisternae at the periphery. Once the mitosis is complete, the ER cisternae are distributed to the new generative and vegetative cells. These ER cisternae may, during the first haploid mitosis, form a barrier preventing the plastids reaching the new generative cell. Recently CLAUHS and GRUN (1977) in Solanum have noticed plastids inside the generative cell as well as inside the vegetative cell. The plastids of the generative cell, however, degenerate and are not present in the ripe pollen grain.
After the first haploid mitosis and after the formation of the new cytoplasm the second stage of vacuolation and the second stage of amylogenesis take place. Since the divisions are not synchronous, the grains that are the first to become binucleate have more time to complete their maturation. The grains
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

POLLEN GRAIN DEVELOPMENT IN LYCOPERSICUM PERUVIANUM 49
that have a longer binucleate phase accumulate more starch and this may explain their different behaviour at the time of germination, both on the stigma or on the artificial medium. Not all the grains germinate at the same time. Some do not germinate at all. These differences which occur between the grains can first be observed during the diad stage when the cytomictic channels, which apparently ensure the synchrony of the first meiotic division, are occluded (HESLOP-HARRISON 1968; PACINI and CRESTI 1977). Even in the same meiocyte the second meiotic division may occur out of phase.
ER cisternae are virtually absent during meiosis and in the tetrad stage. They become more abundant in the microspore stage and increase enormously in the binucleate grain, where, during amylogenesis, the ER stacks are formed. Such stacks are particularly noticeable in mature pollen grains of which they may form part of the reserve material (CRESTI et al., 1975). Other pollen grain reserves are lipid globules and, according to STANLEY and LINSKENS (1975), low molecular weight sugars derived from the second amylogenesis. However, it is unlikely that all the starch formed during the second amylogenesis, about 30-40% of the whole grain, is metabolized during the activity preceding the anthesis. In our opinion these low molecular weight sugars are used for both the metabolism of pollen grains and pollen tubes and also to form the pollen tube walls during germination, after the pollen grain activation is complete. This last phase, the pollen grain development, involving both the metabolism of nucleic (MASCARENHAS and BELL 1970) and the low molecular weight sugars, seems to be designed to prepare the pollen grain for germination.
REFERENCES
ANGOLD R.E.,1965.- The formation of the generative cell in the pollen grain of Endymion non-scriptus (L). J. Cell Sci., 3: 573-578.
CLAUHS R.,P. and GRUN P., 1977.- Changes in plastid and mitochondrion content during maturation of generative cells of Solanum (Solanaceae). Am. J. Bot., 64: 377-383.
CREsTI M., PACINI E., SARFATTI G. and SIMONCIOLI C., 1975. - Ultrastructural features and storage function of Lycopersicum peruvianum pollen. In: «Gamete competition in Plants and Animals» (ed. by Mulchay D.L., 1975). North-Holland Publishing Company-Amsterdam, the Netherlands.
DICKINSON H. G., 1976. - Common factors in exine deposition. In: «The evolutionary significance of the exine». (Ed. by Ferguson I.K. and Muller J.). Linnean Soc. Symp. Series, No. 1, 1976, pp. 67-90.
DICKINSON H.G. and HESLOP-HARRISON J., 1977. - Ribosomes, membranes and organelles during meiosis in angiosperms. Phil. Trans. R. Soc. London B., 277: 327-342.
EcHLIN P., 1972. - The ultrastructure and ontogeny of pollen in Helleborus foetidus L. IV. Pollen grain maturation. J. Cell Sci., 11: 111-129.
ECHLIN P. and GoDWIN H., 1969. - The ultrastructure and ontogeny of pollen in Helleborus foetidus L. III. The formation of the pollen grain wall. J. Cell Sci., 5: 459-477.
GoRSKA-BRYLASS A., 1970. - The «callose stage» of the generative cells in pollen grains. Grana, 10: 21-30.
HESLOP-HARRISON J., 1968. - Wall development within the microspore tetrad of Lilium longiflorum. Can. J. Bot., 46: 1185-1192.
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014

50 PACINI and JUNIPER
1971. - Wall pattern formation in Angiosperm microsporogenesis. In: «Control mechanisms of growth and differentiation». Symp. Soc. Exp. Bioi., 25: 277-300.
MAscARENHAs].P. and BELL E., 1970.- RNA synthesis during development of the male gametophyte of Tradescantia. Devl. Bioi., 21: 475-490.
MATILE P.H., 1975. - The lytic compartment of plant cells. Springer-Verlag, Vienna-New York. MEPHAM R.H. and LANE J.R., 1970. - Obseroations on the fine structure of developing microspores of
Tradescantia bracteata. Protoplasma, 70: 1-20. PACINI E. and CRESTI M., 1976. -Close association between plastids and endoplasmic reticulum cisterns
during pollen grain develop'ent in Lycopersicum peruvianum. J. Ultrastruct. Res., 57: 260-265. -, 1978. - Ultrastructural characteristics of the tapetum and microspore mother cells in Lycopersicum
peruvianum during meiotic prophase. Bull. Soc. Bot. Fr., Actualites botaniques, Number 1-2: 121-128.
PACINI E. and SARFATTI G., 1978. -The reproductive calendar of Lycopersicum peruvianum Mill. Bull. Soc. Bot. Fr., Actualites botaniques, Number 1-2: 259-299.
STANLEY R.G. and LINSKENS H.F., 1975. - Pollen: biology, biochemistry, management. SpringerVerlag, Berlin, Heidelberg, New York.
Received 28 September 1983; revision accepted 7 February 1984
Dow
nloa
ded
by [
175.
177.
159.
153]
at 0
6:02
21
Mar
ch 2
014
Copyright © 2022 FDOKUMEN




















