
Pollen vacuoles and their significance
11
REVIEW Pollen vacuoles and their significance Ettore Pacini • Ce ´dric Jacquard • Christophe Cle ´ment Received: 5 April 2011 / Accepted: 8 June 2011 / Published online: 26 June 2011 Ó Springer-Verlag 2011 Abstract Vacuoles of several types can be observed in pollen throughout its development. Their physiological significance reflects the complexity of the biological pro- cess leading to functional pollen grains. Vacuolisation always occurs during pollen development but when ripe pollen is shed the extensive translucent vacuoles present in the vegetative parts in previous stages are absent. Vacuole functions vary according to developmental stage but in ripe pollen they are mainly storage sites for reserves. Vacuoles cause pollen to increase in size by water accumulation and therefore confer some degree of resistance to water stress. Modalities of vacuolisation occur in pollen in the same manner as in other tissues. In most cases, autophagic vacuoles degrade organelles, as in the microspore after meiosis, and can be regarded as cytoplasm clean-up fol- lowing the transition from the diploid sporophytic to the haploid gametophytic state. This also occurs in the gener- ative cell but not in sperm cells. Finally, vacuoles have a function when microspores are used for pollen embryo- genesis in biotechnology being targets for stress induction and afterwards contributing to cytoplasmic rearrangement in competent microspores. Keywords ER Á Lysosomes Á Pollen Á Reserves Á Vacuoles Á Volume increase Introduction Vacuoles are absent in procaryotic cells but present in algae, fungi and terrestrial plants (Margulis and Schwartz 1982). In algae and fungi they are always present except in resting spores (Dottge 1973). Mosses show vacuoles in all cell types, even in the dispersing spores (Neidhart 1979; Brown and Lemmon 1981). In Pteridophyta, such as Matteuccia and Equisetum, ripe spores contain vacuoles; in others such as Polypodium vulgare vacuoles are absent and water content is around 5% (Cran 1979). In other terrestrial plants, vacuoles are reduced in meristems and practically absent in ripe pollen and seeds, namely in reproductive structures dispersed with a low water content in an arrested developmental state (Heslop-Harrison 1979; Footitt and Cohn 2001). Nevertheless vacuoles are present in seed and pollen at various stages of development as well as during their germination (Heslop-Harrison 1987; Hicks et al. 2004). Vacuole functions vary according to cell differentiation stage. The most common and widespread is regulation of water potential in the cytoplasm under normal conditions and stress (Maurel 1997; Kjellbom et al. 1999). Other recognised functions are (i) storage of different types of substances, such as carbohydrates and proteins or catabo- lites (John 1991); (ii) turnover of cytoplasm parts by lysosomes (Marty 1999); (iii) to drive plant cell growth during differentiation (Maurel 1997). The mode of vacu- olisation and the function of vacuoles in germinating pollen were revealed by Hicks et al. (2004). Despite the importance of vacuoles in plant cell physi- ology, the process of vacuolisation during pollen devel- opment has been described only incidentally by a few authors (Cass and Karas 1975; Pacini and Juniper 1984; Evans et al. 1992; Kuang and Musgrave 1996; Pacini et al. E. Pacini Dipartamento di Scienze Ambientali « Giacomino Sarfatti », Universita degli Studi di Siena, via PA Mattioli 4, 53100 Siena, Italy C. Jacquard Á C. Cle ´ment (&) URVVC, UPRES EA 2069, Laboratoire de Stress, De ´fenses et Reproduction des Plantes, Universite ´ de Reims Champagne Ardenne, BP 1039, 51687 Reims Cedex 2, France e-mail: [email protected] 123 Planta (2011) 234:217–227 DOI 10.1007/s00425-011-1462-4
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Pollen vacuoles and their significance

REVIEW
Pollen vacuoles and their significance
Ettore Pacini • Cedric Jacquard • Christophe Clement
Received: 5 April 2011 / Accepted: 8 June 2011 / Published online: 26 June 2011
� Springer-Verlag 2011
Abstract Vacuoles of several types can be observed in
pollen throughout its development. Their physiological
significance reflects the complexity of the biological pro-
cess leading to functional pollen grains. Vacuolisation
always occurs during pollen development but when ripe
pollen is shed the extensive translucent vacuoles present in
the vegetative parts in previous stages are absent. Vacuole
functions vary according to developmental stage but in ripe
pollen they are mainly storage sites for reserves. Vacuoles
cause pollen to increase in size by water accumulation and
therefore confer some degree of resistance to water stress.
Modalities of vacuolisation occur in pollen in the same
manner as in other tissues. In most cases, autophagic
vacuoles degrade organelles, as in the microspore after
meiosis, and can be regarded as cytoplasm clean-up fol-
lowing the transition from the diploid sporophytic to the
haploid gametophytic state. This also occurs in the gener-
ative cell but not in sperm cells. Finally, vacuoles have a
function when microspores are used for pollen embryo-
genesis in biotechnology being targets for stress induction
and afterwards contributing to cytoplasmic rearrangement
in competent microspores.
Keywords ER � Lysosomes � Pollen � Reserves �Vacuoles � Volume increase
Introduction
Vacuoles are absent in procaryotic cells but present in
algae, fungi and terrestrial plants (Margulis and Schwartz
1982). In algae and fungi they are always present except in
resting spores (Dottge 1973). Mosses show vacuoles in all
cell types, even in the dispersing spores (Neidhart 1979;
Brown and Lemmon 1981). In Pteridophyta, such as
Matteuccia and Equisetum, ripe spores contain vacuoles; in
others such as Polypodium vulgare vacuoles are absent and
water content is around 5% (Cran 1979). In other terrestrial
plants, vacuoles are reduced in meristems and practically
absent in ripe pollen and seeds, namely in reproductive
structures dispersed with a low water content in an arrested
developmental state (Heslop-Harrison 1979; Footitt and
Cohn 2001). Nevertheless vacuoles are present in seed and
pollen at various stages of development as well as during
their germination (Heslop-Harrison 1987; Hicks et al.
2004).
Vacuole functions vary according to cell differentiation
stage. The most common and widespread is regulation of
water potential in the cytoplasm under normal conditions
and stress (Maurel 1997; Kjellbom et al. 1999). Other
recognised functions are (i) storage of different types of
substances, such as carbohydrates and proteins or catabo-
lites (John 1991); (ii) turnover of cytoplasm parts by
lysosomes (Marty 1999); (iii) to drive plant cell growth
during differentiation (Maurel 1997). The mode of vacu-
olisation and the function of vacuoles in germinating pollen
were revealed by Hicks et al. (2004).
Despite the importance of vacuoles in plant cell physi-
ology, the process of vacuolisation during pollen devel-
opment has been described only incidentally by a few
authors (Cass and Karas 1975; Pacini and Juniper 1984;
Evans et al. 1992; Kuang and Musgrave 1996; Pacini et al.
E. Pacini
Dipartamento di Scienze Ambientali « Giacomino Sarfatti »,
Universita degli Studi di Siena, via PA Mattioli 4,
53100 Siena, Italy
C. Jacquard � C. Clement (&)
URVVC, UPRES EA 2069, Laboratoire de Stress, Defenses et
Reproduction des Plantes, Universite de Reims Champagne
Ardenne, BP 1039, 51687 Reims Cedex 2, France
e-mail: [email protected]
123
Planta (2011) 234:217–227
DOI 10.1007/s00425-011-1462-4

1992; McCormick 2004), while studying other aspects.
Special attention was recently paid to vacuoles throughout
pollen development and the results suggest that vacuoles
are plastic compartments and their aspects, origin and
functions vary during microspore/vegetative cell develop-
ment (Yamamoto et al. 2003).
The aim of this paper is to review current knowledge of
this process and to focus on the functions of vacuoles at
various steps of pollen ontogenesis in different groups.
Vacuole functions, activities and their manner of formation
are not only important for basic knowledge but also for
applied research, such as pollen embryogenesis (Touraev
et al. 1997), in vitro pollen development (Benito-Moreno
et al. 1988) and ripe pollen conservation (Yamamoto et al.
2003).
Vacuoles and pollen development
The process of vacuole formation in Angiosperm species
was followed from tetrad to ripe pollen stage (Fig. 1).
Except in orchids, a huge vacuole develops in the micro-
spore between tetrad release and the first pollen mitosis.
More precisely, the vacuolisation process concerns the
cytoplasm of meiocytes, microspores and vegetative cells
but not sperm cells. Some differences, for example the
number of waves of and peaks of vacuolisation are evident
in the species studied. Lycopersicum peruvianum and Olea
europaea have two peaks of vacuolisation, one in late
microspore stage when many small vacuoles forms, and the
other in middle bicellular stage when only one vacuole
forms (Fig. 1). In Prunus avium, Smilax aspera and Lilium,
the vacuolisation peak occurs at first haploid mitosis with
one big vacuole (Pacini and Franchi 1983; Pacini et al.
1986). Vice versa in Arabidopsis thaliana, Lolium perenne,
Zea mays and all grasses, it occurs in late microspore stage
with one big vacuole (Pacini and Keijzer 1989; Pacini et al.
1992).
In Lilium the vacuole develops from autophagic lyso-
somes after meiosis and occupies most of the microspore
volume just before mitosis (Clement et al. 1994). Soon
after mitosis, the vacuole is restricted to the vegetative cell
and reduces as starch accumulates. In the above species,
vacuoles, as extensive translucent areas, are not present in
ripe pollen though small vacuoles containing phytate can
be detected in some cases (Clement and Audran 1996).
Orchids are a totally different case for two reasons:
(i) vacuoles always destroy small portions of cytoplasm
and extensive vacuolisation is not observed; (ii) small
vacuoles with translucent content are present throughout
development and also in ripe pollen (Schlag and Hesse
1992; Pacini and Hesse 2002 and references therein).
Fig. 1 Scheme (not to scale) showing the extension of vacuolisation
(black areas) during pollen development of some representative
angiosperm species. Data of the authors and the literature cited
vacuolisation may occur in microspore, vegetative and generative
cells. A process of vacuolisation occurs in all species but peaks at
different stages of development. Vacuoles (extensive translucentareas) are absent from ripe pollen except that of orchids. Orchids are
a peculiar type because vacuoles are always small. Lycopersicumperuvianum and Olea europaea have two peaks of vacuolisation, one
during late microspore stage and the second during middle bicellular
stage. Prunus avium, Smilax aspera, Arabidopsis, Lilium, Loliumperenne, Zea mays and all the grasses only have one. Its apex is at first
haploid mitosis in Prunus and Smilax and in late microspore stage in
Lolium. GC generative cell, VC vegetative cell, SC sperm cell
218 Planta (2011) 234:217–227
123

Pollen development inside the anther is generally quite
synchronous, but pollen mitosis and the peak of vacuoli-
sation are asynchronous. Large differences were observed
in an aneuploid cultivar of olive, leading cytoplasmic
degeneration in microspores and bicellular pollen (Pacini,
unpublished data). On the other hand, the synchrony of
vacuolisation has a maximum in orchids with pollinia,
probably because of the presence of cytomictic channels up
to early bicellular stage (Pacini and Hesse 2002).
Vacuole origins
Plant vacuoles are essentially produced in the following not
mutually exclusive ways (Fig. 2): by endoplasmic reticulum
(ER) and Golgi vesicles, by different activities of ER and by
lysosomes (Marty 1999). Vacuoles observed during pollen
development have different origins: (A) ER cisterns dilate to
form small vacuoles with clear contents which may fuse to
create larger vacuoles; (B) ER cisterns sequester portions of
cytoplasm and the content is subsequently destroyed;
(C) lysosomes, either simple or complex, combine cases
(A) and (B); (D) dictyosome vesicles fuse, increase in size,
or fuse with ER cisternae in case (B) and (C).
These modalities commonly destroy ribosomes, and
may destroy other organelles such as mitochondria and
plastids. This has been documented sporadically in
microspore cytoplasm, during the phase from sporophyte to
gametophyte. It was described in further detail in the
generative cytoplasm of Prunus avium (Pacini et al. 1986)
and Lolium perenne (Pacini et al. 1992). It has been
interpreted as elimination of plastids enabling maternal
inheritance, but parts of cytoplasm containing ribosomes or
other organelles may be destroyed. Vacuoles are always
small in orchids even if all the modalities previously
described above can be recognised.
When vacuoles originate according to the modalities
(B) and (C) the destroyed part is evident for a while inside
it and at different stages of destruction. Ripe vacuoles, i.e.
when the destruction of cytoplasm is over, sometimes have
granular contents and in some cases PAS (periodic acid
Schiff) and PATAg (periodic acid–silver proteinate) posi-
tive contents which later become translucent (Fig. 3).
The term ‘‘vacuolated stage’’ was used as a term for a
stage development when vacuolisation is extensive
(Christensen and Horner 1974; Blackmore and Barnes
1990), which must not be considered correct because the
bulk of the process may occur at different stages of
development; besides, at least in Olea europaea and
Lycopersicum peruvianum there are two vacuolisation
peaks (Fig. 1).
Pollen is internally and often externally polarised
because of the presence of apertures. It is not clear whether
vacuolisation in a given species always has the same pat-
tern or whether it varies from grain to grain. This question
arises from certain images of Lilium microspores showing
two extremely symmetrical vacuoles with respect to the
external geometry of the grain (Fig. 4). The peak of
a
b
c
d
Fig. 2 Vacuole biogenesis in plant cells. a ER cistern dilates at the
extremity to form small vacuoles (stars) with a clear content. b ER
vesicles sequestrate portions of cytoplasm and the content is
subsequently destroyed. C cytoplasm fraction following encapsulation
by lysosomes. c Secondary lysosomes, with internalised portion of
cytoplasm (stars) that is progressively digested. d Dictyosome
vesicles fuse between themselves increasing in size or fuse with ER
cisternae in the case b and c
Fig. 3 TEM section of a young microspore in Lilium showing
PATAg stained material (arrowheads) accumulated within the
developing vacuole. M microspore, L locular fluid, MW microspore
wall. Bar 20 lm
Planta (2011) 234:217–227 219
123
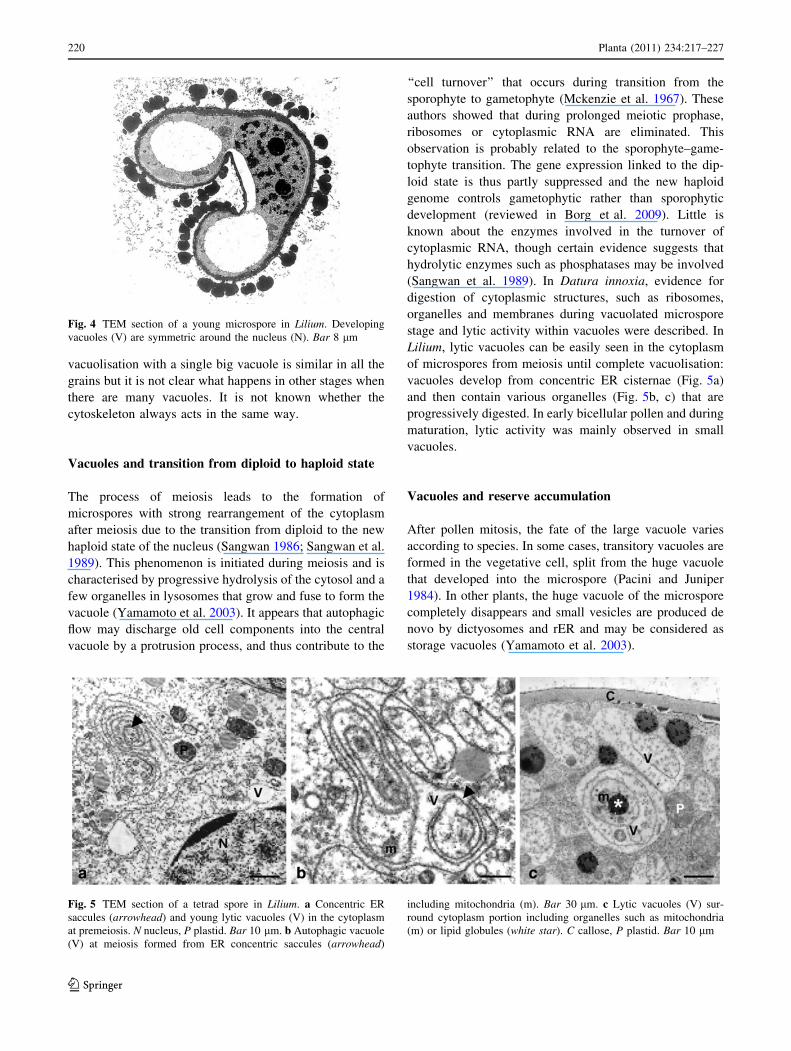
vacuolisation with a single big vacuole is similar in all the
grains but it is not clear what happens in other stages when
there are many vacuoles. It is not known whether the
cytoskeleton always acts in the same way.
Vacuoles and transition from diploid to haploid state
The process of meiosis leads to the formation of
microspores with strong rearrangement of the cytoplasm
after meiosis due to the transition from diploid to the new
haploid state of the nucleus (Sangwan 1986; Sangwan et al.
1989). This phenomenon is initiated during meiosis and is
characterised by progressive hydrolysis of the cytosol and a
few organelles in lysosomes that grow and fuse to form the
vacuole (Yamamoto et al. 2003). It appears that autophagic
flow may discharge old cell components into the central
vacuole by a protrusion process, and thus contribute to the
‘‘cell turnover’’ that occurs during transition from the
sporophyte to gametophyte (Mckenzie et al. 1967). These
authors showed that during prolonged meiotic prophase,
ribosomes or cytoplasmic RNA are eliminated. This
observation is probably related to the sporophyte–game-
tophyte transition. The gene expression linked to the dip-
loid state is thus partly suppressed and the new haploid
genome controls gametophytic rather than sporophytic
development (reviewed in Borg et al. 2009). Little is
known about the enzymes involved in the turnover of
cytoplasmic RNA, though certain evidence suggests that
hydrolytic enzymes such as phosphatases may be involved
(Sangwan et al. 1989). In Datura innoxia, evidence for
digestion of cytoplasmic structures, such as ribosomes,
organelles and membranes during vacuolated microspore
stage and lytic activity within vacuoles were described. In
Lilium, lytic vacuoles can be easily seen in the cytoplasm
of microspores from meiosis until complete vacuolisation:
vacuoles develop from concentric ER cisternae (Fig. 5a)
and then contain various organelles (Fig. 5b, c) that are
progressively digested. In early bicellular pollen and during
maturation, lytic activity was mainly observed in small
vacuoles.
Vacuoles and reserve accumulation
After pollen mitosis, the fate of the large vacuole varies
according to species. In some cases, transitory vacuoles are
formed in the vegetative cell, split from the huge vacuole
that developed into the microspore (Pacini and Juniper
1984). In other plants, the huge vacuole of the microspore
completely disappears and small vesicles are produced de
novo by dictyosomes and rER and may be considered as
storage vacuoles (Yamamoto et al. 2003).
Fig. 4 TEM section of a young microspore in Lilium. Developing
vacuoles (V) are symmetric around the nucleus (N). Bar 8 lm
Fig. 5 TEM section of a tetrad spore in Lilium. a Concentric ER
saccules (arrowhead) and young lytic vacuoles (V) in the cytoplasm
at premeiosis. N nucleus, P plastid. Bar 10 lm. b Autophagic vacuole
(V) at meiosis formed from ER concentric saccules (arrowhead)
including mitochondria (m). Bar 30 lm. c Lytic vacuoles (V) sur-
round cytoplasm portion including organelles such as mitochondria
(m) or lipid globules (white star). C callose, P plastid. Bar 10 lm
220 Planta (2011) 234:217–227
123

Some small translucent vesicles with electron opaque
cores are often observed in mature pollen (Fig. 6). Helsper
et al. (1984) and Butow et al. (1997) found phytate inside
them in Lilium and Chlorophyton pollen. They are involved
in providing inorganic reserves for germination but they do
not seem to give rise to vacuoles at germination (Baldi
et al. 1987). In Eryngium campestre, P particles including
polysaccharides have been described in the vegetative cell
during pollen maturation (Weber 1988). Using cryo-fixa-
tion/freeze substitution, Yamamoto et al. (2003) described
membrane-bound structures containing fibrillar substances
(MBFs) in mature pollen of Arabidopsis thaliana. These
structures are regarded as protein storage vacuoles for
further pollen germination. These special vacuoles origi-
nate from the activity of rough endoplasmic reticulum
(rER) that synthesizes large quantities of proteins that are
stored in the cytoplasm after formation of sperm cells. If
pollen is not dispersed after anther opening, these vacuoles
accumulate lytic enzymes and contribute to degradation of
the vegetative cell.
Various types of vacuoles/vesicles may be found in the
vegetative cell during pollen maturation. Hence they differ
in content and subsequently in function. For example, three
types of vesicles/vacuoles coexist in mature pollen of
Arabidopsis thaliana (Yamamoto et al. 2003): (i) P parti-
cles including polysaccharides that may be used for pollen
tube growth, (ii) lysosomes containing enzymes that take
part in pollen tube penetration of the female part and, (iii)
rER cisternae containing proteins that are also involved in
pollen tube elaboration.
At the onset of prophase, plastids are commonly undif-
ferentiated but accumulate starch and become amyloplasts
once or twice during development (Clement and Pacini
2001). In most species, there is only one phase of amylo-
genesis and this starts just after the peak of vacuolisation,
suggesting that sugars of vacuoles may be translocated to
plastids until needed for microspore/pollen metabolism.
Vacuoles are commonly absent in ripe pollen, and this is
very well documented when quantitative cytology is cou-
pled with three-dimensional reconstruction of organelles,
as in Plumbago zeylanica (Russell 1984) and Zea mays
(McConchie et al. 1987). Another clear demonstration of
the absence of vacuoles in ripe pollen is that of Evans et al.
(1991) who used pollen collected directly from the open
anther with a pollen collector. In many other cases, pollen
was fixed for observation while still in the anther where the
process of dehydration of pollen and anther is not com-
plete. This is probably why Theunis et al. (1991) found
vacuoles with translucent content in the vegetative cyto-
plasm of Spinacia oleracea pollen. Similarly, Chang and
Neuffer (1989) stated that in ripe Zea mays pollen, the
contents consists of a large centrally located vacuole and
the cytoplasm is pressed thinly between the vacuole and the
inner wall. These misinterpretations probably spring from
the fact that only shed pollen can be considered ripe; when
pollen is still in the anther it is difficult to determine
whether it is ripe or not. This means that real mature pollen
is only found when anthers are open and the pollen is in
equilibrium with the environment.
Vacuole and pollen volume increase
The growth of pollen during development is closely corre-
lated with vacuole development but occurs at different rates
in different plants (Fig. 1). Especially in some species, the
volume increase is greater in the final stages of development.
For example, in Cucurbita, the higher pollen volume the
grater the increase in vacuole volume during development
(Pacini 2000). In orchids, the increase in pollen volume and
development of vacuoles are limited (Pandolfi et al. 1993).
In most plant cells, the increase in vacuole volume is
correlated with water accumulation under the control of
aquaporins on plasma membranes and vacuoles (Maurel
1997; Kjellbom et al. 1999). This entry of water into vacu-
oles is linked to an increase in osmotic pressure in the vac-
uolar sap, due to accumulation of osmolites through the
tonoplast. In the case of pollen grains, the main growth of the
vacuole occurs when the microspores develop locular fluid
with large amounts of soluble and insoluble polysaccharides
arising from dissolution of callosic tetrad walls and from
tapetum secretions (Clement et al. 1998; Aouali et al. 2001).
It is therefore likely that the formation of the huge vacuole
leading to microspore and pollen size increase corresponds
to water and osmolite accumulation in vacuolar sap between
tetrad release and the first pollen mitosis.
Vacuolisation is followed by an increase in pollen grain
volume, but during ripening this volume decreases due to
Fig. 6 TEM section of vegetative cell in the mature pollen grain of
Lilium. Vacuoles (arrowheads) are small and include dense deposits.
L lipid, M mitochondrion, P plastid, PW pollen wall. Bar 20 lm
Planta (2011) 234:217–227 221
123

water loss (Pacini 2000), raising the question of what fills the
space left by vacuolisation. Destroyed cytoplasm is probably
replaced by formation of new cytoplasm. No quantitative
data is available on cytosol increase during pollen devel-
opment, but we presume that this increase is largely due to
storage of materials such as lipids (spherosomes) and car-
bohydrates, present in plastids or the cytoplasm (Franchi
et al. 1996). Unlike in seeds, extensive protein storage does
not seem common during pollen ripening (Pacini 1996).
Vacuoles and pollen water content during maturation
Pollen develops inside the loculus, in a fluid that conveys
nutrients from the sporophyte to the developing gameto-
phyte. The flow of these substances decreases and ceases
after tapetum degeneration. The locular fluid thus disap-
pears before anther opening, allowing pollen exposure to
dispersing agents (Pacini and Hesse 2004). Pollen may lose
or gain some water during exposure depending on the
environment (Pacini 2000).
It was recently shown that it is possible to distinguish two
pollen types according to water content at shedding: pollen
partially dehydrated (PDP) has a water content less than
30%, partially hydrated pollen (PHP) has a water content
over 30% (Nepi et al. 2001; Franchi et al. 2002). PDP loses
much water before or as soon as it is exposed and resists
better to environmental stresses such as temperature and low
relative humidity better that PHP. However it takes longer to
rehydrate and germinate on the stigma. PHP loses little water
before exposure and is more vulnerable to environmental
stresses; on the other hand it germinates more quickly on the
stigma because little water needs to be absorbed (Pacini and
Hesse 2004). Specific genes are expressed during pollen
hydration (Wilson et al. 1997; McCormick et al. 1994),
indicating that pollen water balance is under genetic control.
Both classes of pollen are devoid of extensive vacuoles.
In wheat (PHP), cyclosis occurs in ripe pollen (Heslop-
Harrison et al. 1997). The absence of vacuoles means that
PHP stores surplus water in the cytoplasm as free molecules
or associated with certain polymers, such as carbohydrates
and proteins. Since water is the best known fluid stored in
vacuoles of plant cells (Maurel 1997; Kjellbom et al. 1999),
aquaporins are probably involved in accumulation of water
in the microspore vacuole during development under opti-
mal conditions and during drought stress.
Vacuoles, variations in osmotic potential and water
stress during pollen development
The osmotic pressure in vacuoles of microspore/pollen is
regulated during pollen development. It fluctuates between
350 and 450 mosm/l during microspore vacuolisation
(Hoekstra et al. 1993), showing that (i) the process of
vacuolisation is linked to maintenance of strong osmotic
pressure in vacuolar sap and that (ii) the microspore vac-
uole presumably adapts to external variations that would
modify pressure in the immediate environment.
Male gametophytes have low tolerance to environmental
variations. For example, plant cells react to drought by
physiological modifications of the vacuole content and
osmotic pressure. In suspension cells there is a rapid
increase in vacuolar volume in response to salt stress
(Mimura et al. 2003). Pollen may be affected under water
stress but the vacuole does not seem to be the main case of
water injury during pollen development. The most sensitive
step of development is meiosis, upsets though pollen
remains sensitive through development (Saini 1997). In
fact, water stress upsets carbohydrate metabolism in the
whole anther and inhibits key enzymes of sugar metabo-
lism. Developing microspores/pollen grains are thus
deprived of nutrients, which affects their vigour and ability
to germinate.
In ripe pollen, vacuoles are reduced in size when pres-
ent. Pollen undergoes stress, especially during presentation
and dispersal, but the role of vacuoles has not been clearly
demonstrated. Depending on the capacity of pollen to
retain water, the following categories can be identified:
(i) pollen not dehydrated when anther opens and dispersed
in the sea (McConchie and Knox 1989); (ii) pollen that has
completed the process of dehydration prior to anther
opening; (iii) pollen that has not completed the process of
dehydration when the anther opens but completes it during
exposure or dispersal; (iv) pollen that is not dehydrated
when the anther opens and has no mechanism for retaining
water (Pacini 2000). Only type (i) and those of orchids
seem to have vacuoles at dispersal (McConchie and Knox
1989; Pacini and Hesse 2002).
Vacuoles and cell polarisation
Before the discovery that the cytoskeleton is responsible
for cyclosis and movement of the nucleus and organelles,
the formation of a big vacuole in late microspore stage was
interpreted as a means for nuclear displacement and
asymmetric mitosis (Knox and Ducker 1991). This does
not seem completely true because extensive vacuolisation
may occur at different stages, either earlier or later with
respect to pollen mitosis. Besides, in orchids extensive
vacuolisation does not occur but the microspore nucleus
polarizes before pollen mitosis.
On the other hand, the location of the asymmetric
division plane in the microspore also depends on the
location of microtubules close to the plasma membranes
222 Planta (2011) 234:217–227
123

(Twell et al. 1998). In species with huge vacuoles, the
position of the nucleus in the microspore before the first
pollen mitosis is therefore presumably due to the size of the
vacuole in the microspore and the position of the micro-
tubules generating the pre-prophase band. Analysis of
mutant Arabidopsis thaliana showed the crucial role of
microtubules orientation and the vacuole seemed to have a
secondary role (Twell et al. 1998).
Plastid function depends on information originating in
the nucleus. The extension of the vacuole in the microspore
after meiosis concentrates the organelles around the
nucleus favouring exchange between the nucleus and the
semi-autonomous organelles (Nagata et al. 2000).
Vacuolisation and pollen embryogenesis
Pollen embryogenesis is a biotechnological process leading
to the formation of double haploid plants from microsp-
ores, which would normally become pollen grains. The
process is characterised by a switch in microspore physi-
ology from a gametophytic programme to an embryogenic
sporophytic programme. The switch is triggered by stress.
The phase of microspore vacuolisation is the most
suitable time to collect microspores or early bicellular
pollen grain. In this stage, a group of specific so-called
‘‘early’’ genes are expressed in the microspore (Honys and
Twell 2003). Later on after vacuole regression in maturing
pollen it becomes almost impossible for the developing
gametophyte to change its developmental programme.
During vacuolisation the microspore is still plastic and can
take another development pathway, though the implica-
tions of the vacuole in this trait are still unclear.
Using the microspore as a tool for pollen embryogene-
sis, attention must be paid to osmotic potential during
microspore isolation and pre-treatment (Hoekstra et al.
1993; Jacquard et al. 2009). Most pre-treatment media are
liquid, which requires that the osmotic pressure of the
medium be properly adjusted. If the osmotic pressure of
the medium is lower than that of the vacuolar sap, the
microspores may burst by massive water uptake. Con-
versely, a high osmotic pressure in the pre-treatment
medium stresses the microspore and temporarily plasmo-
lyses it. This stress triggers microspore reorientation in
many cases (Wojnarowiez et al. 2004). Though the influ-
ence of the osmotic stress on microspore physiology is
unknown, vacuoles are the first target when inducing pollen
embryogenesis.
Observations of the vacuole by transmission electron
microscopy using the tonoplast feature as marker can be
used to determine microspore competence for embryo-
genesis at a very early stage of the process. Indeed, sen-
sitive microspores are characterised by deposition of a
tannin layer inside tonoplasts (Sangwan and Camefort
1983). The relation between the presence of tannin in
tonoplasts and microspore competence for haploid
embryogenesis is unclear.
The vacuole of the microspore is also affected during
the embryogenic process (Clement et al. 2005). To induce
pollen embryogenesis, microspore stressing pre-treatment
is done during microspore vacuolisation. The behaviour of
stressed microspores has been studied in detail using cell
tracking in tobacco, barley and wheat (Touraev et al. 1997;
Kumlehn and Lorz 1999; Indrianto et al. 2001). The
nucleus gradually moves to the centre of the microspore,
possibly guided by the cytoskeleton. At the same time, the
vacuole breaks up and sub-vacuoles become regularly
distributed around the central nucleus to form a typical
‘‘star like’’ microspore (Fig. 7). These events are regarded
as the first signs of microspore reorientation and are a
necessary step in the process of pollen embryogenesis
(Indrianto et al. 2001). The ‘‘star like microspore’’ devel-
ops an embryogenic programme. Maraschin et al. (2005)
demonstrated in barley that androgenesis is not only linked
to a peculiar type of vacuole, but also to the presence of
intine.
In the days after pre-treatment, organelle degradation and
lytic activity of the vacuole related to change in nuclear
ploidy can be observed (Sangwan and Sangwan-Norreel
1987; Garrido et al. 1995). This indicates that the pro-
gramme of cytoplasm reorganisation initiated in the
microspore in vivo after meiosis is maintained under in vitro
conditions, despite the drastic treatment of microspores.
Reisen et al. (2005) stressed the vacuole of suspension
culture cells, osmotically or with PEG, demonstrating an
increase in tonoplast size and ‘‘double’’ membrane struc-
ture. They postulated that this response creates a reserve
membrane for when original conditions are restored.
During embryogenesis the central vacuole does not
disappear, but divides simultaneously with the cell, pro-
viding one vacuole per cell at each stage of early
embryogenesis (Sangwan and Sangwan-Norreel 1987).
A similar situation occurs in somatic cells of carrot and
Cichorium during de-differentiation and embryogenesis
(Sangwan and Sangwan-Norreel 1987). This suggests
fragmentation of the central vacuole during these processes
and regression of the central vacuole during differentiation
and maturation. Tonoplasts seem to play an active but
unknown role during in vitro embryogenesis.
Molecular biology of pollen vacuoles
Rojo et al. (2003) showed that the protein complex local-
ised in tonoplasts is common to many vegetative cell types
and conserved by virtue of homology with yeast tonoplast
Planta (2011) 234:217–227 223
123

proteins. The role of vacuolar ATPase in male gameto-
phytes was revealed by Dettmer et al. (2005) and indeed a
large central vacuole where osmotically active substances
are transported is necessary for cell expansion. Besides,
plant cells and especially ripe pollen must maintain their
intracellular ion homeostasis throughout adverse conditions
that may occur during pollination. Several vacuolar ion
channels, such as H?-ATPase or calcium channels, appear
to be expressed at all stages of pollen development
(Jakobsen et al. 2005; Peiter et al. 2005; Bock et al. 2006)
and during germination and pollen tube growth (Pertl et al.
2009; Sheoran et al. 2009). Strompen et al. (2005) found
that vacuolar H? ATPase, was present during somatic
development but absent during development of both male
and female gametophytes. It was also shown that vacuolar
acidic invertase involved in wall metabolism and sugar
metabolism is up-regulated during pollen development and
tube growth (Holmes-Davis et al. 2005; Dai et al. 2006).
Plastid metabolism interferes with that of vacuoles.
Using mutants, Niewiadomski et al. (2005) demonstrated
that plastidial glucose 6 phosphate is essential for pollen
maturation and embryo sac development. Mutant pollen
does not form lipid bodies and pollen structure disinte-
grates when vacuole organisation is disrupted.
In recent years, the function of aquaporins, a class of
membrane proteins facilitating water diffusion across
membranes, has been characterised (Luu and Maurel
2005). The presence and/or significance of these molecules
has not been well studied during pollen ripening, presen-
tation and dispersal. They may have a role in anther and
pollen dehydration before dehiscence. They have been
detected in most anther tissues (Ruiter et al. 1997; O’Brien
et al. 2002; Alexandersson et al. 2005; Bots et al. 2005).
Some aquaporin genes are pollen specific (Bock et al.
2006; Maeshima and Ishikawa 2008; Soto et al. 2008),
suggesting that expression of aquaporins in pollen is under
strict regulation by the gametophytic programme. Although
they are likely involved in the process of water accumu-
lation within developing vacuoles during vacuolation, the
involvement of aquaporins has not yet been clearly
reported in pollen development but rather in pollen ger-
mination upon contact with the stigma (Ikeda et al. 1997).
Thus, their role is probably crucial when pollen is exposed
to the environment. We can suppose that the actual func-
tion of the aquaporins may be in the pollen tube elongation,
in the stigma-style transition and in the pollen rehydration.
Conclusions and perspectives
Structural and chemical research into pollen vacuoles are
both afflicted by methodological problems. Structural
observations are mainly done with material rehydrated
during fixation because glutaraldehyde and osmium
tetroxide are dissolved in water. This means that pollen
dispersed with low water content rehydrate during fixation.
We do not know if the small vacuoles sometimes present in
ripe pollen images are due to this rehydration. Even if we
assume that small vacuoles are part of pollen structure, it is
impossible to separate the vacuoles to analyse their
contents.
Vacuoles seem practically absent in ripe pollen, because
extensive translucent areas as in the vegetative part of the
plant, are not observed. Only small areas with electron
opaque deposits can be seen and are interpreted as phytate
deposits. They were not observed when pollen was col-
lected from open anthers or if the fixative was devoid of
water. However, the cytoplasm was full of small vesicles
containing polysaccharides content (Weber 1988; Franchi
et al. 1996; Yamamoto et al. 2003) and the cytosol con-
tained sucrose, glucose, fructose (Speranza et al. 1997) and
lipid deposits of spherosoms, but these can be considered
small vacuoles created by ER dilation.
In this respect, mature pollen is similar to seeds: con-
ventional vacuoles are absent from endosperm and embryo
cells of mature seeds. Though ripe seed vacuoles differ
from those of pollen because they store proteins (Marty
1999), in both pollen and seeds water content dramatically
decreases before dispersal, with some exceptions such as
PHP and recalcitrant seeds (Baskin and Baskin 1998).
Fig. 7 Dynamics of vacuoles in
the early steps of microspore
embryogenesis
224 Planta (2011) 234:217–227
123

Partially dehydrated pollen (Nepi et al. 2001; Franchi et al.
2002) and orthodox seeds (Baskin and Baskin 1998) resist
to severe environmental stresses better. The behaviour of
male and female gametophytes is different because the
latter always have a big central vacuole (Howden et al.
1998).
The behaviour of vacuoles is not uniform throughout the
angiosperms during pollen development. There seem to be
two main types: extensive vacuolisation during develop-
ment and its virtual absence in ripe pollen, as in almost all
angiosperms; reduced vacuolisation during development
and presence even in the ripe stage as in orchids. This may
be because pollen of orchids with pollinia as pollen dis-
persal unit is PHP, not totally exposed to the environment,
but protected inside the anther until pollinator arrival
(Pacini and Hesse 2002, 2004). In fact, it stays inside the
anther until loaded by insects (Pacini and Hesse 2002).
Under such sheltered conditions even pollen dehydration is
not necessary and pollen vacuoles are present, and resist
well out of the anther because of their high sucrose content
(Nepi et al. 2001; Franchi et al. 2002).
If vacuoles as water reserves are absent in ripe pollen,
even in PHP, new vacuoles are formed during rehydration
and early germination (Southworth and Dickinson 1981;
Hicks et al. 2004). The content of reserve containing ves-
icles is mobilised for activation and germination. There is
little data on this topic, despite its great interest, especially
if structural observations are linked or proceed with
molecular biology observations on gene action.
In the last decade, many genes have been shown to be
specifically expressed at special steps of pollen develop-
ment that coincide with the formation or degradation of
vacuoles in the microspore or pollen grain (Oldenhof et al.
1996). Unfortunately the physiological function of many of
these genes is unknown (Honys and Twell 2003) despite
correlations with vacuolisation.
Several pathways remain to be investigated related to
pollen vacuole and its role in pollen biology/biotechnology.
Regarding pollen behaviour, it would be of interest to
determine the role of vacuole in pollen tolerance to dehy-
dration during pollination. Pollen survival is a key step of
sexual reproduction and its capacity to tolerate water stress
conditions the achievement of the reproductive process.
Furthermore, small vacuoles in mature pollen grain are
likely involved in the process of pollen rehydration pre-
ceding pollen tube germination. There is currently only
poor information on this process. Regarding pollen
embryogenesis, vacuole in microspore/pollen at the time of
pre-treatment plays a central role since it conditions the
position of the nucleus which determines the symmetry of
the first division following microspore/pollen reorientation.
Here again, data are missing about mechanisms regulating
this process.
References
Alexandersson E, Fraysse L, Sjovall-Larsen S, Gustavsson S, Fellert
M, Karlsson M, Johanson U, Kjellbom P (2005) Whole gene
family expression and drought stress regulation of aquaporins.
Plant Mol Biol 59:469–484
Aouali N, Laporte P, Clement C (2001) Pectin secretion and
distribution in the anther during pollen development in Lilium.
Planta 213:71–79
Baldi BG, Franceschi VR, Loewus FA (1987) Localization of
phosphorus and cation reserves in Lilium longiflorum pollen.
Plant Physiol 83:1018–1021
Baskin CC, Baskin JM (1998) Seeds: ecology biogeography and
evolution of dormancy and germination. Academic Press, San
Diego
Benito-Moreno RM, Macke F, Hause MT, Alwen A, Heberle-Bors E
(1988) Sporophytes and male gametophytes from in vitro
cultured, immature tobacco pollen. In: Cresti M, Gori P, Pacini
E (eds) Sexual reproduction in higher plants. Springer, Berlin,
pp 137–142
Blackmore S, Barnes S (1990) Pollen wall development in angio-
sperms. In: Blackmore S, Knox RB (eds) Microspore: evolution
and ontogeny. Academic Press, London, pp 173–192
Bock KW, Honys D, Ward JM, Padmanaban S, Nawrocki EP, Hirschi
KD, Twell D, Sze H (2006) Integrating membrane transport with
male gametophyte development and function through transcri-
ptomics. Plant Physiol 140:1151–1168
Borg M, Brownfield L, Twell D (2009) Male gametophyte develop-
ment: a molecular perspective. J Exp Bot 60:1465–1478
Bots M, Feron R, Uehlein N, Weterings K, Kaldenhoff R, Mariani T
(2005) PIP1 and PIP2 aquaporins are differentially expressed
during tobacco anther and stigma development. J Exp Bot
56:113–121
Brown RC, Lemmon BE (1981) Aperture development in spore of the
moss, Trematodon longicollis Mx. Protoplasma 106:273–287
Butow R, Rodriguez-Garcia MI, Alche JD, Gorska-Brylass A (1997)
Calcium in electron-dense globoids during pollen grain matura-
tion in Chlorophytum elatum R.Br. Planta 203:413–421
Cass DD, Karas I (1975) Development of sperm cells in barley. Can J
Bot 53:1051–1062
Chang T, Neuffer MG (1989) Maize microsporogenesis. Genome
32:232–244
Christensen JE, Horner HT (1974) Pollen pore development and its
spatial orientation during microsporogenesis in the grass Sor-ghum bicolor. Am J Bot 61:604–623
Clement C, Audran JC (1996) In vivo and in vitro pollen maturation of
Lilium. Influence of carbohydrates. Acta Soc Bot Pol 65:73–82
Clement C, Pacini E (2001) Anther plastids in angiosperms. Bot Rev
67:54–73
Clement C, Chavant L, Burrus M, Audran JC (1994) Anther starch
variations in Lilium during pollen development. Sex Plant
Reprod 7:347–356
Clement C, Laporte P, Audran JC (1998) The loculus content and
tapetum during pollen development in Lilium. Sex Plant Reprod
11:94–106
Clement C, Sangwan RS, Sangwan BS (2005) Microspore embryo
induction and development in higher plants: cytological and
ultrastructural aspects. In: Palmer CE, Keller WA, Kasha KJ
(eds) Haploids in crop improvement II. Springer, Berlin,
pp 53–72
Cran DR (1979) The ultrastructure of fern gametophyte cells. In: Dyer
AF (ed) The experimental biology of ferns. Academic Press,
London, pp 171–213
Dai S, Li L, Chen T, Chong K, Xue Y, Wang T (2006) Proteomic
analyses of Oryza sativa mature pollen reveal novel proteins
Planta (2011) 234:217–227 225
123

associated with pollen germination and tube growth. Proteomics
6:2504–2529
Dettmer J, Schubert D, Calvo-Weimar O, Stierhof Y-D, Schmidt R,
Schumacher K (2005) Essential role of V-ATPase in male
gametophyte development. Plant J 41:117–124
Dottge JD (1973) The fine structure of algal cells. Academic Press,
London
Evans DE, Taylor PE, Singh MB, Knox RB (1991) Quantitative
analysis of lipids and protein from the pollen of Brassica napusL. Plant Sci 73:117–126
Evans DE, Taylor PE, Singh MB, Knox RB (1992) The interrela-
tionship between the accumulation of lipids, protein and level of
acyl carrier protein during development of Brassica napus L.
pollen. Planta 186:343–354
Footitt S, Cohn MA (2001) Developmental arrest: from sea urchin to
seeds. Seed Sci Res 11:2–16
Franchi GG, Bellani LM, Nepi M, Pacini E (1996) Types of
carbohydrate reserves in pollen: localization, systematic distri-
bution and ecophysiological significance. Flora 191:143–159
Franchi GG, Nepi M, Pacini E (2002) Partially hydrated pollen:
taxonomic distribution, ecological and evolutionary significance.
Plant Syst Evol 234:211–227
Garrido D, Vicente O, Heberle-Bors E, Rodriguez-Garcia I (1995)
Cellular changes during the acquisition of embryogenic potential
in isolated pollen grains of Nicotiana tabacum. Protoplasma
186:220–230
Helsper JPFG, Linskens HF, Jackson JF (1984) Phytate metabolism in
Petunia pollen. Phytochemistry 23:1841–1845
Heslop-Harrison J (1979) An interpretation of the hydrodynamics of
pollen. Am J Bot 66:737–743
Heslop-Harrison J (1987) Pollen germination and pollen-tube growth.
Int Rev Cytol 107:1–78
Heslop-Harrison J, Heslop-Harrison Y, Heslop-Harrison JS (1997)
Motility in ungerminated grass pollen: association of myosin with
polysaccharide-containing wall-precursors bodies (P bodies). Sex
Plant Reprod 10:65–66
Hicks GR, Rojo E, Hong S, Carter DG, Raikhel N (2004)
Germinating pollen has tubular vacuoles, displays highly
dynamic vacuole biogenesis, and requires VACUOLESS1 for
proper function. Plant Physiol 134:1227–1239
Hoekstra FA, van Zijderweld MH, Heidekamp F, van der Mark F
(1993) Microspore culture of Hordeum vulgare L.: the influence
of density and osmolality. Plant Cell Rep 12:661–665
Holmes-Davis R, Tanaka CK, Vensel WH, Hurkman WJ, McCormick
S (2005) Proteome mapping of mature pollen of Arabidopsisthaliana. Proteomics 5:4864–4884
Honys D, Twell D (2003) Comparative analysis of the Arabidopsispollen transcriptome. Plant Physiol 132:640–652
Howden R, Park SK, Moore JM, Orme J, Grossniklaus U, Twell D
(1998) Selection of T-DNA tagged male and female gameto-
phytic mutants by segregation distortion in Arabidopsis. Genet-
ics 149:621–631
Ikeda S, Nasrallah JB, Dixit R, Preiss S, Nasrallah ME (1997) An
aquaporin-like gene required for the Brassica self-incompatibil-
ity response. Science 276:1564–1566
Indrianto A, Barinova I, Touraev A, Heberle-Bors E (2001) Tracking
individual wheat microspores in vitro: identification of embryo-
genic microspores and body axis formation in the embryo. Planta
212:163–174
Jacquard C, Mazeyrat-Gourbeyre F, Devaux P, Boutilier K, Baillieul
F, Clement C (2009) Microspore embryogenesis in barley:
anther pre-treatment stimulates plant defence gene expression.
Planta 229:393–402
Jakobsen MK, Poulsen LR, Schulz A, Fleurat-Lessard P, Moller A,
Husted S, Schiott M, Amtmann A, Palmgren MG (2005) Pollen
development and fertilization in Arabidopsis is dependent on the
male gametogenesis impaired anthers gene encoding a type V
P-type ATPase. Genes Dev 19:2757–2769
John P (1991) Fructan quality and fructan synthesis. Biochem Soc T
19:569–572
Kjellbom P, Larsson C, Johansson I, Karlsson M, Johanson U (1999)
Aquaporins and water homeostasis in plants. Trends Plant Sci
4:308–314
Knox RB, Ducker SC (1991) The evolution of gametes—from motility
to double fertilization. In: Blackmore S, Barnes SH (eds) Pollen and
spores-pattern of diversification. Systematic Association Special
Volume n�44. Clarendon Press, Oxford, pp 345–361
Kuang A, Musgrave ME (1996) Dynamics of vegetative cytoplasm
during generative cell formation and pollen maturation in
Arabidopsis thaliana. Protoplasma 194:81–90
Kumlehn J, Lorz H (1999) Monitoing sporophytic development of
individual microspores of barley (Hordeum vulgare L.). In:
Clement C, Pacini E, Audran JC (eds) Anther and pollen: from
biology to biotechnology. Springer, Berlin, pp 183–190
Luu D-T, Maurel C (2005) Acquaporins in a challenging environ-
ment: molecular gears for adjusting plant water status. Plant Cell
Environ 28:85–96
Maeshima M, Ishikawa F (2008) ER membrane aquaporins in plants.
Eur J Physiol 456:709–716
Maraschin SF, Vennik M, Lamers GEM, Spaink HP, Wang M (2005)
Time-lapse tracking of barley androgenesis reveals position-
determined cell death within pro-embryos. Planta 220:531–540
Margulis L, Schwartz KV (1982) Five Kingdoms: an illustrated guide
to the phyla of life on earth. Freeman and Co, San Francisco
Marty F (1999) Plant vacuoles. Plant Cell 11:587–599
Maurel C (1997) Aquaporins and water permeability of plant
membranes. Annu Rev Plant Physiol Plant Mol Biol 48:399–429
McConchie CA, Knox RB (1989) Pollination and reproductive
biology of seagrasses. In: Larkum AWD, McComb AJ, Sheperd
SA (eds) Biology of seagrasses. Elsevier, Amsterdam, pp 74–111
McConchie CA, Hough T, Knox RB (1987) Ultrastructural analysis
of the sperm cells of mature pollen of maize, Zea mays.
Protoplasma 139:9–19
McCormick S (2004) Control of male gametophyte development.
Plant Cell 16:142–153
McCormick S, Curie C, Eyal Y, Muschietti J, Dirks L, Kulikauskas R
(1994) Molecular biology of male gametogenesis. Euphytica
79:245–250
Mckenzie A, Heslop-Harrison J, Dickinson HG (1967) Elimination of
ribosomes during meiotic prophase. Nature 215:997–999
Mimura T, Kura-Hotta M, Tsujimura T, Ohnishi M, Miura M, Okazaki Y,
Miura M, Maeshima M, Washitani-Nemoto S (2003) Rapid increase
of vacuolar volume in response to salt stress. Planta 216:397–402
Nagata N, Saito C, Sakai A, Kuroiwa H, Kuroiwa T (2000) Unique
positioning of mitochondria in developing microspores and
pollen grains in Pharbitis nil: mitochondria cover the nuclear
surface at specific developmental stages. Protoplasma 213:74–82
Neidhart HV (1979) Comparative studies of sporogenesis in Bryo-
phytes. In: Clarke CGS, Duckett JS (eds) Bryophyte systematics.
Academic Press, London, pp 251–280
Nepi M, Franchi GG, Pacini E (2001) Pollen hydration status at
dispersal: cytophysiological features and strategies. Protoplasma
216:171–180
Niewiadomski P, Knappe S, Geimer S, Fischer K, Schulz B, Unte US,
Rosso MG, Ache P, Flugge U-F, Schneider A (2005) The
Arabidopsis plastidic glucose 6-phosphate/phosphate translocator
GPT1 is essential for pollen maturation and embryo sac devel-
opment. Plant Cell 17:746–759
O’Brien M, Bertrand C, Matton DP (2002) Characterization of
a fertilization-induced and developmentally regulated plasma-
membrane aquaporin expressed in reproductive tissues, in the
wild potato Solanum chacoense Bitt. Planta 215:485–493
226 Planta (2011) 234:217–227
123

Oldenhof MT, Groot PFM, Visser JH, Schrauven JAM, Wullems GJ
(1996) Isolation and characterization of a microspore-specific
gene from tobacco. Plant Mol Biol 31:213–225
Pacini E (1996) Types and meaning of pollen carbohydrates reserves.
Sex Plant Reprod 9:362–366
Pacini E (2000) From anther and pollen ripening to pollen presen-
tation. Plant Syst Evol 22:19–43
Pacini E, Franchi GG (1983) Pollen grain development in Smilaxaspera and possible function of the loculus. In: Mulcahy DL,
Ottaviano E (eds) Pollen: biology and implications in plant
breeding. Elsevier Biomedical, New York, pp 183–190
Pacini E, Hesse M (2002) Types of pollen dispersal units in orchids,
and their consequences for germination and fertilisation. Ann
Bot 89:653–664
Pacini E, Hesse M (2004) Cytophysiology of pollen presentation and
dispersal. Flora 199:273–285
Pacini E, Juniper BJ (1984) The ultrastructure of pollen grain
development in Lycopersicum peruvianum. Caryologia 37:21–50
Pacini E, Keijzer CJ (1989) Ontogeny of intruding non periplasmodial
tapetum in the wild chicory (Cichorium intybus L. Compositae).
Plant Syst Evol 167:149–164
Pacini E, Bellani LM, Lozzi R (1986) Pollen, tapetum and anther
development in two cultivars of sweet cherry (Prunus avium).
Pytomorphology 36:197–210
Pacini E, Taylor PE, Singh MB, Knox RB (1992) Development of
plastids, including amyloplasts and starch grains, in pollen and
tapetum of rye-grass, Lolium perenne. Ann Bot 70:179–188
Pandolfi T, Pacini E, Calder DM (1993) Ontogenesis of monad pollen
in Pterostylis plumosa (Orchidaceae, Neottioideae). Plant Syst
Evol 186:175–185
Peiter E, Maathuis FJ, Mills LN, Knight H, Pelloux J, Hetherington
AM, Sanders D (2005) The vacuolar Ca2?-activated channel
TPC1 regulates germination and stomatal movement. Nature
434:404–408
Pertl H, Schulze WX, Obermeyer G (2009) The pollen organelle
membrane proteome reveals highly spatial-temporal dynamics
during germination and tube growth of lily pollen. J Proteome
Res 8:5142–5152
Reisen D, Marty M, Leborgne-Castel N (2005) New insights into the
tonoplast architecture of plant vacuoles and vacuolar dynamics
during osmotic stress. BMC Plant Biol 5:1–13
Rojo E, Zouhar J, Kovaleva V, Hong S, Raikhel NV (2003) The AtC-
VPS protein complex is localized to the tonoplast and the
prevacuolar compartment in Arabidopsis. Mol Biol Cell
14:361–369
Ruiter RK, van Eldik GJ, van Herpen MMA, Schrauwen JM,
Wullems GJ (1997) Expression in anthers of two genes encoding
Brassica oleracea transmembrane channel proteins. Plant Mol
Biol 34:163–168
Russell SD (1984) Ultrastructure of the sperm of Plumbago zeylanica:
II. Quantitative cytology and three-dimensional organization.
Planta 162:385–391
Saini HS (1997) Effects of water stress on male gametophyte
development in plants. Sex Plant Reprod 10:67–73
Sangwan RS (1986) Formation and cytochemistry of nuclear vacuoles
during meiosis in Datura. Eur J Cell Biol 40:210–218
Sangwan RS, Camefort H (1983) The tonoplast, a specific marker of
embryogenic microspores of Datura cultured in vitro. Histo-
chemistry 78:473–480
Sangwan RS, Sangwan-Norreel BS (1987) Ultrastructural cytology of
plastids in pollen grains of certain androgenic and nonandro-
genic plants. Protoplasma 138:11–22
Sangwan RS, Mathivet V, Vasseur G (1989) Ultrastructural locali-
zation of acid phosphatase during male meiosis and sporogenesis
in Datura: evidence for digestion of cytoplasmic structures in the
vacuoles. Protoplasma 149:38–46
Schlag M, Hesse M (1992) The formation of the generative cell in
Polystachya pubescens (Orchidaceae). Sex Plant Reprod
5:131–137
Sheoran IS, Pedersen EJ, Ross ARS, Sawhney VK (2009) Dynamics
of protein expression during pollen germination in canola
(Brassica napus). Planta 230:779–793
Soto G, Alleva K, Mazzella MA, Amodeo G, Muschiett JP (2008)
AtTIP1;3 and AtTIP5;1, the only highly expressed Arabidopsis
pollen-specific aquaporins, transport water and urea. FEBS Lett
582:4077–4082
Southworth D, Dickinson DB (1981) Ultrastructural changes in
germinating lily pollen. Grana 20:29–35
Speranza A, Calzoni GL, Pacini E (1997) Occurrence of mono- or
disaccharides and polysaccharide reserves in mature pollen
grains. Sex Plant Reprod 10:110–115
Strompen G, Dettmer J, Stierhof Y-D, Schumacher K, Jurgens G,
Mayer U (2005) Arabidopsis H?-ATPase subunit E isoform 1 is
required for Golgi organization and vacuole function in
embryogenesis. Plant J 41:125–132
Theunis CH, Cresti M, Milanesi C (1991) Studies of the mature
pollen of Spinacia oleracea after freeze substitution and
observed with confocal laser scanning fluorescence microscopy.
Bot Acta 104:324–329
Touraev A, Vicente O, Heberle-Bors E (1997) Initiation of micro-
spore embryogenesis by stress. Trends Plant Sci 2:297–302
Twell D, Ki Park S, Lalanne E (1998) Asymmetric division and cell-
fate determination in developing pollen. Trends Plant Sci
3:305–310
Weber M (1988) Metabolism of P-particles (polysaccharide particles)
in mature pollen grains of Eryngium campestre L. (Apiaceae).
Protoplasma 146:65–71
Wilson C, Voronin V, Touraev A, Vicente O, Heberle-Bors E (1997)
A developmentally regulated MAP kinase activated by hydration
in tobacco pollen. Plant Cell 9:2093–2100
Wojnarowiez G, Caredda S, Devaux P, Sangwan RS, Clement C
(2004) Effects of different osmotica on androgenesis and
albinism in barley (Hordeum vulgare L.). J Plant Physiol
161:747–755
Yamamoto Y, Nishimura M, Hara-Nishimura I, Noguchi T (2003)
Behavior of vacuoles during microspore and pollen development
in Arabidopsis thaliana. Plant Cell Physiol 44:1192–1201
Planta (2011) 234:217–227 227
123




















