
Application of Tubular Reactor Technologies for the ... - MDPI
Upload
independentCategory
view
0download
0
Muscles that are stretched during contraction (eccentric
contractions) are susceptible to damage (Hough, 1902),
particularly when the eccentric exercise is prolonged and
unaccustomed. For example, when walking down a
mountain, the quadriceps group of muscles is used to
deaccelerate the body and undergo eccentric contraction.
Characteristically the muscles involved are weak but pain-
free immediately after the exercise, but in the subsequent
days the muscle becomes stiff, tender and may become
even weaker. This latter collection of symptoms is called
delayed onset muscle soreness (DOMS). Full recovery may
require several weeks and there is a pronounced training
effect; thus if the same exercise is repeated the symptoms
are substantially reduced (Balnave & Thompson, 1993).
The mechanisms involved in this damage are complex and
multifactorial (for recent reviews see Morgan & Allen,
1999; Proske & Morgan, 2001; Warren et al. 2001).
Early ultrastructural studies established that there are
characteristic changes in the sarcomere patterns including
localized regions of overstretched sarcomeres and irregular
and distorted Z lines (Fridén et al. 1981). Morgan
(1990) proposed a mechanism to explain the sarcomere
inhomogeneities on the basis of the mechanical properties
of eccentrically contracting muscle. He pointed out that
when the force on a muscle exceeded about 1.6 Po (Po is the
maximum isometric force), the stretching velocity increased
uncontrollably (Katz, 1939). Furthermore, sarcomeres
on the descending limb of the force–length curve are
intrinsically unstable (Huxley, 1980); this arises when a
sarcomere on the descending limb becomes longer than its
neighbour; it is then weaker than the neighbour and
therefore inclined to stretch further. As a consequence
during stretch on the descending limb of the force–length
curve, individual half-sarcomeres can stretch suddenly
and dramatically (the popping sarcomere hypothesis) and
much of the increase in muscle length can occur in a small
number of abnormally stretched sarcomeres (Brown &
Hill, 1991; Talbot & Morgan, 1996). Most of these
overstretched sarcomeres reinterdigitate when the muscle
relaxes (Talbot & Morgan, 1996), but during repeated
eccentric contractions it would be expected that the
regions of overstretched sarcomeres can extend laterally
and longitudinally through the muscle leading to the
characteristic structural changes (Allen, 2001).
Development of T-tubular vacuoles in eccentrically damagedmouse muscle fibresElla W. Yeung *, Christopher D. Balnave †, Heather J. Ballard‡, J.-P. Bourreau ‡ and David G. Allen †
† Institute for Biomedical Research and Department of Physiology, University of Sydney F13, NSW 2006, Australia, *Department of RehabilitationSciences, Hong Kong Polytechnic University, Hung Hom, Kowloon, Hong Kong and ‡ Department of Physiology, University of Hong Kong, Li Shu FanBuilding, 5 Sassoon Road, Hong Kong
Single fibres were dissected from mouse flexor digitorum brevis muscles and subjected to a protocol
of eccentric stretches consisting of ten tetani each with a 40 % stretch. Ten minutes later the fibres
showed a reduced force, a shift in the peak of the force–length relation and a steepening of the
force–frequency relation. Addition of the fluorescent dye sulforhodamine B to the extracellular
space enabled the T-tubular system to be visualized. In unstimulated fibres and fibres subjected to
10 isometric tetani, the T-tubules were clearly delineated. Sulforhodamine B diffused out of the
T-tubules with a half-time of 18 ± 1 s. Following the eccentric protocol, vacuoles connected to the
T-tubules were detected in six out of seven fibres. Sulforhodamine B diffused out of the vacuoles of
eccentrically damaged fibres extremely slowly with a half-time of 6.3 ± 2.4 min and diffused out of
the T-tubules with a half-time of 39 ± 4 s. Vacuole production was eliminated by application of
1 m ouabain to the muscle during the eccentric protocol. On removal of the ouabain, vacuoles
appeared over a period of 1 h and were more numerous and more widely distributed than in the
absence of ouabain. We propose that T-tubules are liable to rupture during eccentric contraction
probably because of the relative movement associated with the inhomogeneity of sarcomere
lengths. Such rupture raises intracellular sodium and when the sodium is pumped from the cell by
the sodium pump, the volume load of Na+ and water exceeds the capacity of the T-tubules and
causes vacuole production. The damage to the T-tubules may underlie a number of the functional
changes that occur in eccentrically damaged muscle fibres.
(Received 14 November 2001; accepted after revision 17 January 2002)
Corresponding author D. G. Allen: Institute for Biomedical Research and Department of Physiology, University of Sydney F13,NSW 2006, Australia. Email: [email protected]
Journal of Physiology (2002), 540.2, pp. 581–592 DOI: 10.1113/jphysiol.2001.013839
© The Physiological Society 2002 www.jphysiol.org

It is less clear how these early structural changes lead to the
reductions of force and to the inflammation and even cell
death that are apparent after several days (McCully &
Faulkner, 1985). Sarcomeres are connected in series in a
muscle so that damage to some sarcomeres would not
necessarily affect the maximum force. Instead one might
predict that they would act as increased compliance and
cause the force–length curve to shift to longer lengths as
has been observed (Katz, 1939; Talbot & Morgan, 1998).
However even when muscles are stretched to the peak of
the new force–length relation, force is often reduced.
Warren et al. (1993) suggested that damage to excitation–
contraction (E–C) coupling occurred in eccentrically
damaged mammalian muscle and this was subsequently
confirmed by direct measurements of intracellular calcium
(Balnave & Allen, 1995; Ingalls et al. 1998). It would be
expected that T-tubules and sarcoplasmic reticulum (SR)
would be susceptible to damage in the overstretched
regions and this may be the basis for the E–C coupling
damage. It is already known that T-tubular vacuoles can
occur in a range of muscle interventions involving osmotic
loads (Krolenko et al. 1998; Khan et al. 2000; Lännergren etal. 2000). Of particular interest is a report that gross
structural damage to a muscle by cutting it in half leads to
rapid and profuse development of vacuoles associated
with the T-tubules near the site of damage (Casademont etal. 1988). Very recently it has been shown in electron
microscopy (EM) studies that the T-tubules are abnormal
in eccentrically stretched muscles (Takekura et al. 2001).
However, there are no previous reports of vacuole
development in association with eccentric muscle damage.
There is also considerable evidence of surface membrane
damage following eccentric contractions. For instance,
some studies have shown reductions in the membrane
potential immediately after eccentric damage (McBride etal. 2000), although others have not (Warren et al. 1993).
We have recently shown that pH regulation is abnormal
immediately after eccentric damage; since transporters in
the surface membrane, such as the Na+–H+ exchanger and
the lactate transporter, dominate pH regulation, this
suggests some loss of membrane function (Yeung et al.2002). Substantial increases in the plasma protein levels of
soluble muscle proteins such as creatine kinase may be
observed several days after eccentric damage (Jones et al.1986), although this may be more indicative of the start of
cell breakdown than early membrane damage.
In this study, T-tubule morphology and function were
compared between fibres which had not contracted or had
undergone only isometric contractions, and fibres which
had undergone eccentric contractions. The morphology of
the T-tubules was examined using confocal microscopy
and an extracellular fluorescent dye, sulforhodamine B.
In addition the accessibility of the T-tubules from the
extracellular space was evaluated from the rate of washout
of sulforhodamine B from the T-tubules. Finally, the role
of the Na+,K+-ATPase in the development of T-tubule
vacuoles was evaluated by performing eccentric contractions
in the presence of ouabain.
METHODSSingle fibre dissection and mountingAdult, male mice were killed by rapid neck disarticulation andsingle muscle fibres were dissected from the flexor brevis musclesas previously described (Lännergren & Westerblad, 1987). Theseprocedures were approved by the Animal Ethical Committee ofthe University of Sydney. The isolated fibres were mountedbetween an Akers AE801 force transducer (SensoNor, Horten,Norway) and the arm of a motor (Model 300H, CambridgeTechnology, Cambridge, MA, USA). The motor allowed knownlength changes to be imposed on the muscle fibre. The fibres werestimulated with platinum-plate electrodes using pulses of 0.5 msduration at an intensity of ~1.2 w threshold. All tetanic contractionswere 400 ms in duration and the standard stimulation frequencywas 100 Hz.
SolutionsThe dissection was performed in a solution of the followingcomposition (m): 136.5 NaCl, 5 KCl, 1.8 CaCl2, 0.5 MgCl2, 0.4NaH2PO4 and 11.9 NaHCO3 (pH 8.0). During the experiment, thefibres were superfused at 2.2 ml min_1 in the following standardsolution (m): 121 NaCl, 5 KCl, 1.8 CaCl2, 0.5 MgCl2, 0.4 NaH2PO4,24 NaHCO3 and 5.5 glucose, with 0.2 % (v/v) fetal calf serum(Gibco). The solution was bubbled with 95 % O2–5 % CO2 (pH 7.4).In some experiments, 1 m ouabain (Sigma-Aldrich) was addedto inhibit the activity of Na+,K+-ATPase. Ouabain was dissolved indimethyl sulfoxide (DMSO) to give a concentration of 100 m.This stock solution was stored at a temperature of _20 °C,protected from light, and diluted into the standard solutionimmediately before use. All experiments were performed at roomtemperature (22–24 °C).
Eccentric contraction protocolThe experimental protocol used was similar to those describedpreviously (Balnave & Allen, 1995; Yeung et al. 2002). The basicprotocol (eccentric series) involved superimposing a stretch oneach of a series of 10 tetani. It is important to distinguish betweenthe force deficit caused by fatigue and that associated witheccentric contractions (Morgan & Allen, 1999) and for this reasonan isometric (control) series of tetani was also performed. We alsowished to establish that the force–frequency behaviour and theforce–length properties of the fibre were unaffected by the controlseries, but modified by the eccentric series. Thus the fullexperimental protocol was as follows. (i) The force–lengthrelation was established using tetani separated by 1 min andchanging length in 40 mm steps. The fibre was adjusted to Lo (thelength giving maximum force). (ii) Tetani at 10, 20, 30, 50, 70 and100 Hz were produced with 1 min of rest between each tetanus(force–frequency relation). (iii) Ten isometric tetani at 4 sintervals were given (the control series). (iv) After 10 min, theforce–length relation was redetermined. (v) The force–frequencyrelation was redetermined. (vi) Ten eccentric tetani were given at4 s intervals. The length change was either 25 % or 40 % of Lo. Thestretch was imposed over 100 ms starting after 200 ms of thetetanus. The fibre was shortened back to the Lo over 100 msstarting 200 ms after the end of the tetanus. (vii) After 10 min theforce–length relation was re-established and the fibre length
E. W. Yeung and others582 J. Physiol. 540.2

kept at the original Lo. (viii) The force–frequency relation wasredetermined. (ix) The fibre was then stretched to the newoptimum length (post eccentric Lo). (x) The force–frequencyrelation was redetermined. (xi) Finally, the fibre was removedfrom the motor and positioned on the coverslip of a new chamberused for confocal microscopy.
Confocal microscopyTo visualize the morphology of the T-tubular system, the fibre wasexamined with an inverted confocal microscope (Leica TCS SL,Heidelberg, Germany), using a w 63, NA 1.2 water immersionobjective lens. After the contraction protocol, the muscle fibre wastransferred into a perfusion chamber. The two tendons of the fibrewere fixed to the coverslip with silicone grease. The perfusionchamber was narrow so that the washout of extracellular spaceafter a solution change was reasonably fast (see Fig. 4A). T-tubularmorphology was examined by adding 0.5 m sulforhodamine B(Molecular Probes, OR, USA) to the perfusate which diffusedinto the T-system (Endo, 1966; Lännergren et al. 2000).Sulforhodamine B fluorescence was excited using the 543 nm lineof a helium–neon laser operated at 50 % maximum power andemitted light was collected between 570 and 610 nm. Theresolution of the microscope in this configuration was estimatedusing 170 nm diameter green fluorescent beads (PS-Speck,Molecular Probes). The half-maximal width of the resultingfluorescence signal was 0.35 mm in the horizontal direction and0.9 mm in the vertical direction.
Confocal microscopy was performed on four unstimulated fibresand on three fibres that underwent only the control series ofisometric tetani. Seven fibres underwent the full protocoldescribed above. The entire length of the muscle fibre andmultiple depths were examined for morphological alterationsand/or presence of vacuolation, but because of dead fibres andconnective tissue the ends of the fibre were harder to visualize.Images are presented in two formats. In some figures, e.g. Fig. 2,sulforhodamine B was present in the extracellular space andproduced a very large fluorescent signal that caused the detectorto saturate (indicated by a blue colour). Even in a confocalmicroscope some fractions of the fluorescence above and belowthe fibre contributed to fibre fluorescence and this reduced theresolution of fluorescent structures within the fibre. For thisreason we also show figures, e.g. Fig. 5, collected 1 min afterextracellular sulforhodamine B had been washed out of theextracellular space. Images were scanned at a resolution of512 w 512 pixels.
In addition to studying the morphological alterations of theT-tubules, a second objective of our study was to examine thefunction of the T-tubules following eccentric contractions.Water-soluble dyes, such as sulforhodamine B (MW 559), diffusein and out of the T-tubular system with a time course in the rangeof 4–10 s (Endo, 1966). It is thus possible to examine theaccessibility of the T-tubules by measuring the rate of exit of thefluorescent dye. In order to achieve this, fibres were superfusedwith the standard solution containing sulforhodamine B for15–20 min before observations. The dye-containing solution wasthen removed and washed out with standard solution for at least20 min. Confocal images of the fibres were obtained every 10 s forthe first minute, every minute for the next 4 min and then at 5 minintervals over 20 min. The rate of loss of sulforhodamine B fromthe T-system was estimated from the decline of fluorescence,which was analysed using NIH Image (Scion Corporation, MD,USA). The average fluorescence intensity was determined in a
window of roughly half the diameter of the fibre placed overa representative region of the fibre. Before exposure tosulforhodamine B the fibres exhibited no significant fluorescence.Contributions to the fluorescence from dye outside the fibreprofile were judged to be small because the intensity of vacuoleschanged little as the extracellular dye was washed away (comparevacuoles in Fig. 3Ba and b). This procedure was repeated forseveral regions of each fibre to ensure that fluorescence intensitymeasurements were representative. When vacuoles were present amuch smaller box comparable to the size of a few vacuoles wasused and placed over the same group of vacuoles in each of a seriesof images.
Na+,K+-ATPase inhibitorWe further examined the morphology of the T-tubules aftereccentric contractions (n = 3) during exposure to 1 m ouabain,an inhibitor of Na+,K+-ATPase. The fibre was exposed to ouabainfrom 15 min before the eccentric protocol and the ouabainremained present throughout the eccentric protocol and duringthe examination of the fibre on the confocal microscope.Subsequently ouabain was washed out using standard solution forthe next 2 h and we again examined the fibre on the confocalmicroscope. Two fibres were treated in an identical manner exceptthat the experimental protocol involved isometric tetani only.
StatisticsData are quoted as means ± standard error of means (...) withthe number of experiments denoted as n. Statistical significancewas determined with Student’s paired t test for force recordingsand optimal length. The unpaired t test was used to compare therate of dye exit between fibres following isometric and eccentrictetani. The significance level was set at P < 0.05.
RESULTSEffect of the eccentric series on mechanicalperformanceThe force developed by muscle fibres following the eccentric
series (40 % stretch, n = 7) showed a large reduction to
34 ± 4 % of control (P < 0.0001), measured in a 100 Hz
isometric tetanus 10 min after the eccentric series and at
the original Lo (Fig. 1A). Force was not significantly reduced
after the isometric series (Fig. 1A). As a consequence of
eccentric contractions, there was a shift in optimum length
for force to longer muscle lengths which amounted to an
increase of Lo by a factor of 1.24 ± 0.02. There was no
significant shift in Lo after the isometric series (dashed line
in Fig. 1A). This shift in Lo following eccentric contraction
confirms earlier studies (Katz, 1939; Talbot & Morgan,
1998; Yeung et al. 2002). If the muscle was stretched to the
post-eccentric Lo the decline in force was reduced and the
developed force was 47 ± 3 % of the control conditions.
Figure 1B illustrates the force–frequency relations of the
fibres normalized to 100 Hz stimulation under each
condition. As previously demonstrated, eccentrically
damaged muscles have a steeper force–frequency relation
(Jones et al. 1989; Balnave & Allen, 1995; Yeung et al.2002). It can be seen that the pattern of force reduction was
similar regardless of whether the fibre was at the original Lo
or the post-eccentric Lo. As expected, the isometric series
Vacuoles in eccentric muscle damageJ. Physiol. 540.2 583

had no significant effect on the force–frequency relation
(dashed line in Fig. 1B). These data establish that the
muscle fibres show the previously established mechanical
criteria of eccentric muscle damage and that fatigue due to
the 10 isometric tetani was not a significant factor.
Confocal images of dye-loaded muscle fibresUnder confocal microscopy, control experiments with no
stimulation (n = 4) and those with isometric tetani series
(n = 3) show regular, well-aligned fluorescent lines running
transversely across the fibre (Fig. 2A). Only in the best
resolution images (e.g. Figs 2A and 3Aa) were we able to
partly visualize two T-tubules per sarcomere as has
previously been established in mammalian muscles
(Lännergren et al. 2000). This inability to resolve two
T-tubules per sarcomere probably results from the difficulty
of dissecting the fibres sufficiently cleanly that the fibre lies
very close to the coverslip (see discussion in Lännergren etal. 2000). In occasional images, longitudinal connections
were seen spanning two to five transverse lines.
The most striking feature after 40 % stretch was the
presence of vacuoles in six of the seven fibres examined
(Fig. 2B and C). These vacuoles were present at our earliest
observation, which was 10–20 min after the eccentric
contractions. Vacuolation did not appear throughout the
whole preparation, but was confined to a focal region of
the fibre, so it is possible that they were present in the fibre
in which we failed to identify vacuoles, but in a very
restricted region. In the seven fibres, we counted the
number of vacuoles in a 50 mm length of the fibre and a
single plane of focus in the region where vacuoles were
most prevalent. The number of vacuoles varied between 32
and 0 (11 ± 4). Because the vacuoles were filled with dye
from the extracellular space, they must have had access to
the extracellular space presumably through the T-system.
The vacuoles varied in size and shape with some ovoid in
cross-section running parallel to the length of the muscle
fibre (Fig. 2B). In other experiments, the vacuoles were
larger (up to 2 mm) and roughly spherical in shape
(Fig. 2C). Sulforhodamine B was often applied several
times and the vacuoles persisted for more than 2 h. They
showed no sign of disappearing in contrast to vacuoles
after fatiguing stimulation, which had largely resolved after
1 h (Lännergren et al. 2000). In the eccentrically damaged
fibres, the T-tubules formed a wavy line across the fibre,
the edges of the T-tubules were less clearly defined, and
in some regions increased numbers of longitudinal
connections were apparent. These features were also
present in the one fibre in which no vacuoles could be
located. There was no obvious correlation between of the
presence or number of vacuoles and the magnitude of the
force deficit; for instance the fibre in which no vacuoles were
located had a roughly similar decline in force. In an attempt
to examine whether vacuoles could develop during the
eccentric stretches, but then seal over (Fraser et al. 1998),
we performed the eccentric protocol in two fibres in the
presence of sulforhodamine B. In these experiments the
vacuoles appeared similar to other fibres; furthermore the
dye was gradually lost from all vacuoles when extracellular
dye was removed. Thus we found no evidence of sealed
vacuoles that developed during the eccentric contraction.
Milder stretches of 25 % of Lo did not lead to the
production of vacuoles though force was reduced to
65 ± 10 % of control (n = 2).
E. W. Yeung and others584 J. Physiol. 540.2
Figure 1. Changes in mechanical properties of musclefollowing eccentric contractionsA, the length–force relations at various stages of the experimentalprotocol (n = 8). 0, control; 9, after the isometric series; and 1,after the eccentric series. The curves fitted to each set of data pointsare Gaussian distributions. B, force–frequency relations at variousstages in the protocol. Each plot shows relative force normalized to100 Hz force vs. different frequencies of stimulation (n = 8). 9,control; 0, post-isometric contractions; 1, post eccentriccontractions at the original Lo; and 6, after adjusting to newoptimal length. * Statistically significant difference from controlsituation (P < 0.05).

Rate of fluorescence washout from muscle fibresThe time course of sulforhodamine B washout gives an
estimate of the accessibility of the T-system to the
extracellular space (Endo, 1966). Figure 3 illustrates
representative images obtained during such experiments.
Figure 3Aa is a control image in the presence of
sulforhodamine B in a fibre that had the isometric series of
contractions only. Figure 3Ab and c show images after 1
and 20 min washout. Note that the major part of the dye
had left the T-tubules at 1 min, but faint T-tubules of
roughly similar intensity are present in the images both at
1 min and 20 min. Figure 3Ba–d shows an equivalent
series from an eccentrically damaged fibre. Note the
prominent vacuoles in the image in the presence of
sulforhodamine B (Fig. 3Ba) and that the T-tubules are
less clear than in the equivalent control image. After a
1 min washout, the intensity of dye in the vacuoles was
little changed, but most dye had left the T-tubules. After
20 min and even 2 h, vacuoles still contained some dye and
faintly stained T-tubules were still detectable.
These issues are explored more quantitatively in the graphs
shown in Fig. 4. In Fig. 4A the filled circles show the
washout of the extracellular space, which was reduced to
~5 % at 1 min. The open circles show the rate of decline of
fluorescence in T-tubules in fibres following isometric
tetani, which was roughly exponential with a half-time of
18 ± 1 s. As expected from the work of Endo (1966), the
main decline lags behind the extracellular space by 10–20 s
only, but there is an irreducible tail of fluorescence, which
is about 20 % of the maximum and shows only a minor
decline over 20 min. We are uncertain of the origin of this
fluorescence, but the most likely possibility is that of a
small fraction the dye, possibly caused by a small degree of
lipid solubility, remains bound to the membranes of the
T-system and dissociates very slowly. Following eccentric
contractions, the T-system (vacuoles + T-tubules) (n = 7)
showed significant slowing of washout rate when compared
with those fibres after isometric contractions (P < 0.01).
Note that in these data, which include both T-tubules and
vacuoles, there seems to be two phases in the decline, with
a rapid phase over the first minute and then a slower phase
over at least 20 min.
To determine if the efflux rate of the dye differs from
various structures, we first examined the decline of
fluorescence in vacuoles (Fig. 4B, filled circles, n = 5). It is
clear that the efflux rate is very slow and seems to follow
a roughly exponential time course with a half-time of
6.3 ± 2.4 min. We also examined the efflux from T-tubules
of fibres that had contracted eccentrically, but did not
show vacuoles in the field of view (Fig. 4B, filled triangles,
n = 4). These four fibres included the one eccentrically
damaged fibre that did not show vacuoles, two fibres from
experiments with 25 % stretch that also failed to exhibit
vacuoles, and one fibre that showed vacuoles, but the
measurements were made from an area that did not
exhibit vacuoles. None of these data were significantly
different from each other and they have been combined
in Fig. 4B. The dye leaves the eccentrically damaged
T-tubules much faster than vacuoles and the decline is
roughly exponential with a half-time of 39 ± 4 s. Finally, in
Vacuoles in eccentric muscle damageJ. Physiol. 540.2 585
Figure 2. Confocal images of the muscle fibres in thepresence of extracellular sulforhodamine BA, confocal image of a fibre following isometric contractionshowing well-aligned fluorescent patterns of the T-tubules. In allthe confocal images shown, increasing fluorescence intensity isindicated by the following colour order: black, dark red, yellow,white. Saturation of the detector occurs with dye outside the fibreand is indicated by the blue colour. B and C, fibre that had beenstretched by 40 % Lo showing vacuolation of the T-system. B showsan example of the ovoid-shaped vacuoles appearing longitudinallyover the fibre and C shows roughly spherical vacuoles locatedrandomly throughout the fibre. Scale bar, 10 mm.

E. W. Yeung and others586 J. Physiol. 540.2
Figure 3. Confocal images showing washout of sulforhodamine B Aa, fibre that had been subjected to 10 isometric tetani and was being perfused with sulforhodamine B. Aband c are after 1 and 20 min washout with standard solution. After the removal of the sulforhodamine B, thefluorescence density of the fibre reduces rapidly in the first minute. Ba, fibre that has been subjected toeccentric tetani of 40 % Lo in the presence of extracellular sulforhodamine B. Bright fluorescent vacuolesappeared longitudinally along the muscle fibre. Bb, c and d correspond to the time course of washout periodof 1 min, 20 min and 2 h, respectively. Scale bars, 10 mm.
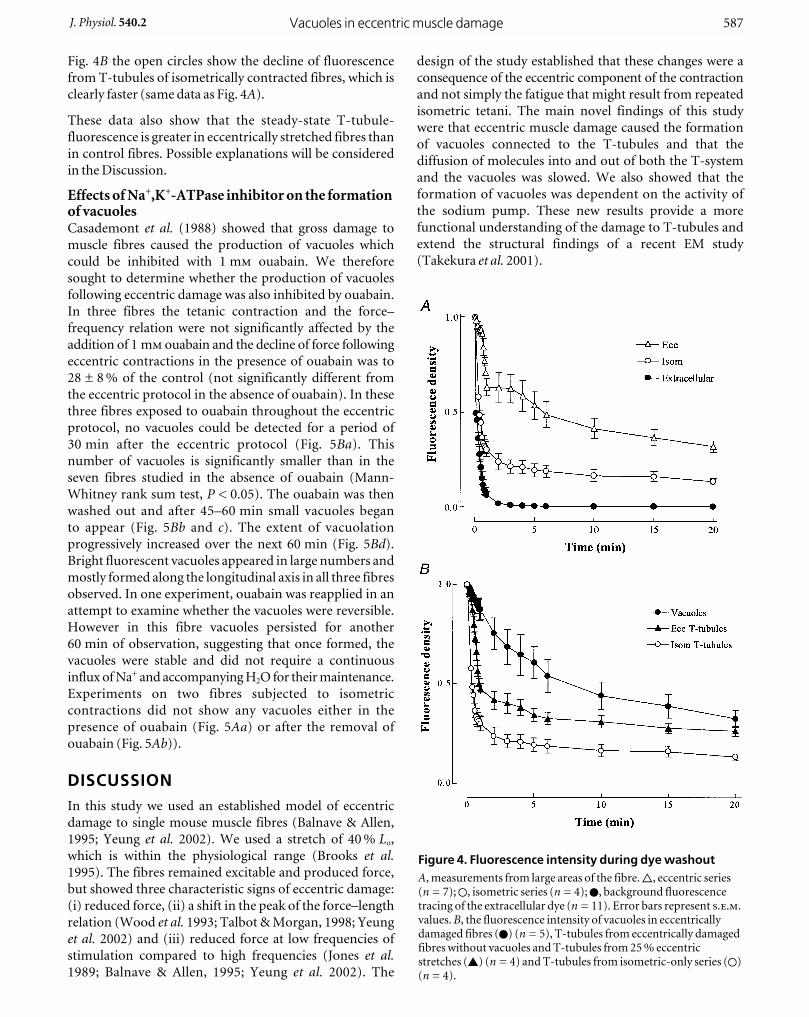
Fig. 4B the open circles show the decline of fluorescence
from T-tubules of isometrically contracted fibres, which is
clearly faster (same data as Fig. 4A).
These data also show that the steady-state T-tubule-
fluorescence is greater in eccentrically stretched fibres than
in control fibres. Possible explanations will be considered
in the Discussion.
Effects of Na+,K+-ATPase inhibitor on the formationof vacuolesCasademont et al. (1988) showed that gross damage to
muscle fibres caused the production of vacuoles which
could be inhibited with 1 m ouabain. We therefore
sought to determine whether the production of vacuoles
following eccentric damage was also inhibited by ouabain.
In three fibres the tetanic contraction and the force–
frequency relation were not significantly affected by the
addition of 1 m ouabain and the decline of force following
eccentric contractions in the presence of ouabain was to
28 ± 8 % of the control (not significantly different from
the eccentric protocol in the absence of ouabain). In these
three fibres exposed to ouabain throughout the eccentric
protocol, no vacuoles could be detected for a period of
30 min after the eccentric protocol (Fig. 5Ba). This
number of vacuoles is significantly smaller than in the
seven fibres studied in the absence of ouabain (Mann-
Whitney rank sum test, P < 0.05). The ouabain was then
washed out and after 45–60 min small vacuoles began
to appear (Fig. 5Bb and c). The extent of vacuolation
progressively increased over the next 60 min (Fig. 5Bd).
Bright fluorescent vacuoles appeared in large numbers and
mostly formed along the longitudinal axis in all three fibres
observed. In one experiment, ouabain was reapplied in an
attempt to examine whether the vacuoles were reversible.
However in this fibre vacuoles persisted for another
60 min of observation, suggesting that once formed, the
vacuoles were stable and did not require a continuous
influx of Na+ and accompanying H2O for their maintenance.
Experiments on two fibres subjected to isometric
contractions did not show any vacuoles either in the
presence of ouabain (Fig. 5Aa) or after the removal of
ouabain (Fig. 5Ab)).
DISCUSSIONIn this study we used an established model of eccentric
damage to single mouse muscle fibres (Balnave & Allen,
1995; Yeung et al. 2002). We used a stretch of 40 % Lo,
which is within the physiological range (Brooks et al.1995). The fibres remained excitable and produced force,
but showed three characteristic signs of eccentric damage:
(i) reduced force, (ii) a shift in the peak of the force–length
relation (Wood et al. 1993; Talbot & Morgan, 1998; Yeung
et al. 2002) and (iii) reduced force at low frequencies of
stimulation compared to high frequencies (Jones et al.1989; Balnave & Allen, 1995; Yeung et al. 2002). The
design of the study established that these changes were a
consequence of the eccentric component of the contraction
and not simply the fatigue that might result from repeated
isometric tetani. The main novel findings of this study
were that eccentric muscle damage caused the formation
of vacuoles connected to the T-tubules and that the
diffusion of molecules into and out of both the T-system
and the vacuoles was slowed. We also showed that the
formation of vacuoles was dependent on the activity of
the sodium pump. These new results provide a more
functional understanding of the damage to T-tubules and
extend the structural findings of a recent EM study
(Takekura et al. 2001).
Vacuoles in eccentric muscle damageJ. Physiol. 540.2 587
Figure 4. Fluorescence intensity during dye washout A, measurements from large areas of the fibre. 9, eccentric series(n = 7); 1, isometric series (n = 4); 0, background fluorescencetracing of the extracellular dye (n = 11). Error bars represent ...values. B, the fluorescence intensity of vacuoles in eccentricallydamaged fibres (0) (n = 5), T-tubules from eccentrically damagedfibres without vacuoles and T-tubules from 25 % eccentricstretches (8) (n = 4) and T-tubules from isometric-only series (1)(n = 4).

E. W. Yeung and others588 J. Physiol. 540.2
Figure 5. Confocal images of muscle fibres in ouabain experimentsA, fluorescence images following isometric (control) series. Aa, 20 min after isometric contractions, ouabainpresent throughout. Ab, after ouabain had been washed out for 60 min. No vacuoles were observed at anyperiod. B, fluorescent images following the eccentric series. Ba, 20 min after the eccentric series, ouabainpresent throughout, no vacuoles observed. Bb, after 40 min washout, hints of vacuoles began to appear. Bcand d correspond to 60 and 80 min after the removal of ouabain. Extensive vacuoles appear over this period.Scale bars, 10 mm. All images in this figure were collected 1 min after washout of sulforhodamine B, whichwas present for 10–20 min before washout.

The study by Takekura et al. (2001) involved rats that
performed a series of downhill runs over 90 min. The
animals were killed at intervals up to 10 days after the
termination of exercise. Identified fast or slow fibres were
studied with high voltage EM and a staining technique that
identified T-tubules or SR. Four changes in the T-system
were identified: (i) increase in the longitudinal elements of
the T-tubules, (ii) changes in the organization of triads,
(iii) the development of calveolar clusters and (iv) the
appearance of multiple connections between two and three
T-tubules and three and four terminal cisternae (pentads
and heptads). The increase in longitudinal T-tubules was
greatest at 3 days and, although we used objective methods
to search for them in our fibres, we could not detect
increased longitudinal T-tubules in our time frame
(0.5–2 h). The calveolar clusters are interesting and were
apparent immediately after exercise but only in slow fibres.
Nevertheless it seems possible that some vacuoles and
caveolar clusters might be similar or identical structures.
Mechanism of production of vacuoles in skeletalmuscleVacuoles have previously been described in a range of
situations including glycerol removal (Krolenko et al.1998; Khan et al. 2000), muscle fatigue (Lännergren etal. 2000) and gross membrane damage (Casademont et al.1988). An extensive review of the mechanism of production
and the significance of skeletal muscle vacuoles has recently
been published (Krolenko & Lucy, 2001). However vacuoles
have not previously been described as part of eccentric
damage, although with the benefit of hindsight, structures
that might be vacuoles are visible in the study by Warrren etal. (1995) (see their Fig. 6).
Lännergren et al. (2000) showed that vacuoles could
develop during the recovery from repeated isometric
tetani in amphibian muscle fibres. Thus it is important to
establish that the vacuoles we observed in the present
experiments were a consequence of the eccentric damage
and not caused by some aspect of the repeated tetani.
Firstly, like Lännergren et al. (2000), we did not observe
vacuoles in mouse fibres after repeated isometric tetani.
Secondly, vacuoles were localized following eccentric
damage, whereas they were widespread in fatigued
amphibian fibres. Thirdly, vacuoles in association with
fatigue disappeared over the course of an hour or so,
whereas those associated with eccentric damage seemed
quite stable over several hours. Thus the vacuoles described
in the present study have different characteristics to those
observed after repeated contractions. It seems that the
vacuoles we observed are specific to eccentric damage.
In all situations where vacuoles have been described, the
muscle is subject to an abnormal osmotic load (Krolenko
& Lucy, 2001). For instance when glycerol is removed from
the extracellular space after a period of exposure, the muscle
Vacuoles in eccentric muscle damageJ. Physiol. 540.2 589
Figure 6. Hypothesis for mechanism and consequences of T-tubular rupture in eccentricmuscle damageA, diagram illustrating sarcomere inhomogeneities in an eccentrically damaged fibre. Sarcomeres 1 and 4 areof normal length, sarcomere 2 is shorter than average while sarcomere 3 is longer. T-tubules in mammalianmuscle are at the overlap of thick and thin filaments and are assumed to be subject to shearing stress(indicated by dashed lines). Where the shear stress is greatest (between sarcomeres 2 and 3), T-tubules areassumed to rupture. B, illustration of some hypotheses of the consequences of eccentric damage followingrupture of T-tubules; AP, action potential. See text for further discussion.

swells as water enters to equilibrate intracellular and
extracellular osmolarity. Subsequently the fibre slowly
returns to its normal volume as glycerol and accompanying
water leave the cell. Vacuoles (and detubulation) occur
during this latter phase and it is thought that the volume
load of water and glycerol leaving the cell via the T-tubules
exceeds their capacity causing them to swell and form
vacuoles. Similarly, during fatigue it is postulated that the
load of lactic acid and accompanying water leaving the cell
in the recovery period causes vacuole development
(Lännergren et al. 2000). It is less clear whether the volume
load simply exceeds the normal flow of the T-system or
whether the osmotic forces also lead to compression of
some T-tubules and reduce their fluid transport capability.
What might cause an osmotic load in association with
eccentric muscle damage? Key observations were made by
Casademont et al. (1988) who showed that when a muscle
was cut in half, very large numbers of vacuoles developed
that were attached to the T-tubules close to the cut region.
They showed that vacuole production could be limited by
a range of procedures including removing Na+ from the
extracellular fluid and addition of a sodium pump
inhibitor. They argued that following surface membrane
damage, Na+ entered the myoplasm from the extracellular
space. This stimulated the Na+ pump which would then
pump the excess Na+ into the extracellular space of the
T-tubules. The volume flow of H2O which accompanies
the Na+ was proposed to exceed the flow capability of the
T-tubules and to cause the vacuole production. We propose
that in eccentric damage, T-tubules suffer shearing damage
when adjacent myofilaments show disparate degrees of
stretch (Fig. 6A). Once disruption of a T-tubule occurs
there will be leakage of extracellular contents into the
myoplasm and vice versa. One important consequence of
this process will be an increase in [Na+]i and [Ca2+]i close to
the end of the disrupted T-tubule. Presumably disrupted
T-tubules either reconnect or seal over and remain
disconnected; however, we have no information on the time
course or frequency of these two processes. We propose
that this localized increase in [Na+]i and [Ca2+]i initiates
two consequences of the initial T-tubular disruption as
indicated in Fig. 6B.
The main evidence in favour of our proposal is extensive
earlier work by Casademont et al. (1988) in a situation of
indisputable membrane damage and our demonstration
that ouabain can completely prevent the development of
vacuoles. It might be argued that the failure to observe
vacuoles in ouabain is simply due to chance, since not all
eccentrically stretched fibres exhibited vacuoles, but this
explanation is eliminated by the subsequent observation of
extensive vacuole development as the ouabain was washed
out of the preparation. The increased frequency of vacuole
development after ouabain is consistent with an increase
in intracellular sodium when the Na+ pump is inhibited.
The slow time course of the appearance of vacuoles as
ouabain is washed off (which is slower than the appearance
of vacuoles after eccentric contractions) is consistent with
the slow washout of ouabain from muscles (Nielsen &
Clausen, 1996). The gradual increase in background fibre
fluorescence as ouabain is washed off (compare Fig. 5Bb to
d) probably represents a slow increase in the number of
vacuoles and in the number of blocked T-tubules so that
sulforhodamine leaves the fibre more slowly with each
subsequent washout.
Our hypothesis for the formation of vacuoles requires the
presence of Na+ pumps in the T-tubules. Early studies
showed that Na+ pumps are present at a reduced density in
the T-tubules (Venosa & Horowicz, 1981). More recent
studies have confirmed the original observation and, in
addition, demonstrate that the distribution of Na+ pump
isoforms is different in the T-tubule compared to the
surface membrane (Williams et al. 2001).
Slowed diffusion in and out of the T-systemIt is established that molecules enter and leave the
T-system with a time course that is only moderately
greater than that expected for free diffusion (Hodgkin &
Horowicz, 1960). In keeping with this, sulforhodamine B
enters and leaves the T-system with a half-time of between
5 and 20 s depending on the diameter of the fibre (Endo,
1996); we confirmed this finding in normal fibres which
had a half-time for removal of sulforhodamine B of
18 ± 1 s. In contrast, removal of sulforhodamine B from
the vacuoles is greatly slowed with a half-time of around
6 min. It is clear from these studies that a substantial
barrier to diffusion is located at the point of connection of
the vacuoles with the T-system.
Of special interest is our observation that, in eccentrically
damaged fibres that did not exhibit vacuoles, the rate of
diffusion of sulforhodamine from the preparation was
significantly slowed to 39 ± 4 s. This is important because
it establishes that there is some change in T-tubular
properties in eccentrically damaged fibres even in the
absence of vacuoles. Furthermore it suggests that at least
some of the barrier to diffusion lies distributed in the
T-system. One possibility is that the T-system is compressed
by the osmotic changes reducing both volume flow and
diffusion. Another possibility is that the rupture of T-tubules
and subsequent sealing over make the diffusion pathway
more restricted and/or more tortuous.
Another interesting observation is the presence of
fluorescence in the T-tubules of normal fibres after 20 min
and the increase in this intensity in eccentrically damaged
fibres. We argued earlier that the fluorescence in normal
fibres could represent binding of the dye to the membrane.
The increased fluorescence following eccentric damage
could arise (i) because the surface area of T-system has
increased, (ii) because the diffusion pathway is more
E. W. Yeung and others590 J. Physiol. 540.2

tortuous, particularly if many T-tubules near the surface
membrane were disrupted, (iii) because some T-tubules
have formed sealed sections from which the dye cannot
escape or (iv) because some dye has entered the myoplasm
and is trapped there. Some of these possibilities could be
distinguished if higher resolution images of T-tubular
staining were available or EM studies of the T-tubules at
the appropriate time were made.
Consequences of T-tubular damage for eccentricmuscle damageThe observation of vacuoles is the most compelling
evidence of T-tubular damage, but do vacuoles necessarily
affect muscle function? In muscle fatigue, vacuoles are a
prominent feature of the recovery period in amphibian
fibres, but they correlate poorly with the degree of
functional recovery (Lännergren et al. 2000). Similarly in
our experiments all the features of eccentric damage were
present in 1/7 fibres subjected to 40 % stretch and 2/2
fibres subjected to 25 % stretch and yet no vacuoles could
be found. In addition, the distribution of vacuoles was
often quite localized while the sarcomere inhomogeneities
and ionic changes appeared to be much more widely
distributed (Balnave et al. 1997). Furthermore, the degree
of reduction of force was similar in the experiments in the
presence of ouabain despite the fact that no vacuoles were
detected. These features lead us to the view that the
fundamental T-tubular defect is not the presence of
vacuoles, but a more minor degree of damage to the
T-tubules that is not detectable with our present images.
However we do have evidence for this abnormality based
on the slower diffusion of sulforhodamine B from the
T-tubules of eccentrically damaged fibres that did not
exhibit vacuoles. Thus we propose that shearing damage to
T-tubules at multiple sites in the muscle, triggered by the
sarcomere inhomogeneities, is the primary T-tubular
defect (Fig. 6A). Vacuoles presumably develop in sites
where this primary damage is so great that the osmotic load
of pumping out the consequent rise in Na+ is sufficiently
great to produce vacuoles (central column of Fig. 6B).
Figure 6B illustrates how we propose that the shearing
damage to T-tubules contributes to other aspects of
eccentric damage. The left hand pathway illustrates some
of the consequences of Ca2+ influx from the extracellular
space. A rise in resting [Ca2+]i has been noted in several
studies (Balnave & Allen, 1995; Ingalls et al. 1998) though
focal rises, which would be predicted by our hypothesis,
have not been detected (Balnave et al. 1997), perhaps
because they are very transient. Several studies have shown
that a rise of the time-averaged [Ca2+]i in a muscle can
inhibit SR Ca2+ release (Lamb et al. 1995; Bruton et al.1996; Chin & Allen, 1996) so the rise in resting [Ca2+]i
could conceivably cause the reduced Ca2+ transients that
have been observed in eccentrically damaged muscles
(Balnave & Allen, 1995; Ingalls et al. 1998). It is also widely
thought that rises in [Ca2+]i may contribute to activation of
proteases and phospholipases and to the ensuing cell
damage and inflammation (Belcastro et al. 1998).
The right hand pathway illustrates some of the possible
consequences of the ruptured or compressed T-tubules. It
is possible that inward conduction of the action potential
would be affected and this is another possible mechanism
for the reduced SR Ca2+ release, which is characteristic
of eccentrically damaged mammalian muscle (references
above). This component of damage could be the cause of
the slowed diffusion of sulforhodamine B, and if removal of
protons is also slowed, it may explain the impaired pH
regulation that we have recently described (Yeung et al. 2002).
ConclusionThese observations of damage and functional changes in
the T-tubular system offer new insights into some of the
early changes in eccentric damage. The formation of
vacuoles, which can be inhibited by ouabain, is strong
evidence of intracellular Na+ loading, presumably through
damaged T-tubules. The ionic changes secondary to
T-tubular damage and the reduced exchange of ions,
metabolites and fluid across the T-tubular network are
capable of explaining a range of phenomena that occur in
eccentrically damaged muscle.
REFERENCESA, D. G. (2001). Eccentric muscle damage: mechanisms of early
reduction of force. Acta Physiologica Scandinavica 171, 311–319.
B, C. D. & A, D. G. (1995). Intracellular calcium and
force in single mouse muscle fibres following repeated
contractions with stretch. Journal of Physiology 488, 25–36.
B, C. D., D, D. F. & A, D. G. (1997). Distribution of
sarcomere length and [Ca2+]i in single fibres from mouse skeletal
muscle following stretch-induced injury. Journal of Physiology 502,
649–659.
B, C. D. & T, M. W. (1993). Effect of training on
eccentric exercise-induced muscle damage. Journal of AppliedPhysiology 75, 1545–1551.
B, A. N., S, L. D. & R, D. A. (1998). Exercise-
induced muscle injury: a calpain hypothesis. Molecular andCellular Biochemistry 179, 135–145.
B, S. V., Z, E. & F, J. A. (1995). Injury to muscle
fibres after single stretches of passive and maximally stimulated
muscles in mice. Journal of Physiology 488, 459–469.
B, L. M. & H, L. (1991). Some observations on variations in
filament overlap in tetanized muscle fibres and fibres stretched
during a tetanus, detected in the electron microscope after rapid
fixation. Journal of Molecular and Cellular Cardiology 12, 171–182.
B, J. D., L, J. & W, H. (1996). Effects of
repetitive tetanic stimulation at long intervals on
excitation–contraction coupling in frog skeletal muscle. Journal ofPhysiology 495, 15–22.
C, J., C, S. & K, G. (1988). Vacuolation
of muscle fibers near sarcolemmal breaks represents T-tubule
dilatation secondary to enhanced sodium pump activity. Journal ofNeuropathology and Experimental Neurology 47, 618–628.
Vacuoles in eccentric muscle damageJ. Physiol. 540.2 591

C, E. R. & A, D. G. (1996). The role of elevations in
intracellular Ca2+ concentration in the development of low
frequency fatigue in mouse single muscle fibres. Journal ofPhysiology 491, 813–824.
E, M. (1966). Entry of fluorescent dyes into the sarcotubular
system of frog skeletal muscle. Journal of Physiology 185, 224–238.
F, J. A., S, J. N., H, A. R. & H, C. L.
(1998). The tubular vacuolation process in amphibian skeletal
muscle. Journal of Muscle Research and Cell Motility 19, 613–629.
F, J., S, M. & E, B. (1981). A morphological
study of delayed muscle soreness. Experientia 37, 506–507.
H, A. L. & H, P. (1960). The effect of sudden
changes in ionic concentrations on the membrane potential of
single muscle fibres. Journal of Physiology 153, 370–385.
H, T. (1902). Ergographic studies in muscular soreness.
American Journal of Physiology 7, 76–92.
H, A. F. (1980). Reflections on Muscle. Liverpool University
Press, Liverpool.
I, C. P., W, G. L., W, J. H., W, C. W. &
A, R. B. (1998). E–C coupling failure in mouse EDL
muscle after in vivo eccentric contractions. Journal of AppliedPhysiology 85, 58–67.
J, D. A., N, D. J., R, J. M. & T, S. E. (1986).
Experimental human muscle damage: morphological changes in
relation to other indices of damage. Journal of Physiology 375,
435–448.
J, D. A., N, D. J. & T, C. (1989). Mechanical
influences on long-lasting human muscle fatigue and delayed-
onset pain. Journal of Physiology 412, 415–427.
K, B. (1939). The relation between force and speed in muscular
contraction. Journal of Physiology 96, 45–64.
K, K. N., S, J. N., H, A. R., B, A. J. &
H, C. L. (2000). Loop diuretics inhibit detubulation and
vacuolation in amphibian muscle fibres exposed to osmotic shock.
Journal of Muscle Research and Cell Motility 21, 79–90.
K, S. A., A, W. B., B, S. C., T, M. V. &
L, J. A. (1998). Accessibility of T-tubule vacuoles to
extracellular dextran and DNA: mechanism and potential
application of vacuolation. Journal of Muscle Research and CellMotility 19, 603–611.
K, S. A. & L, J. A. (2001). Reversible vacuolation of
T-tubules in skeletal muscle: mechanisms and implications for cell
biology. International Review of Cytology 202, 243–298.
L, G. D., J, P. R. & S, D. G. (1995). Raised
intracellular [Ca2+] abolishes excitation–contraction coupling in
skeletal muscle fibres of rat and toad. Journal of Physiology 489,
349–362.
L, J., B, J. D. & W, H. (2000). Vacuole
formation in fatigued skeletal muscle fibres from frog and mouse:
effects of extracellular lactate. Journal of Physiology 526, 597–611.
L, J. & W, H. (1987). The temperature
dependence of isometric contractions of single, intact fibres
dissected from a mouse foot muscle. Journal of Physiology 390,
285–293.
MB, T. A., S, B. W., G, F. A. & C, R. C.
(2000). Stretch-activated ion channels contribute to membrane
depolarization after eccentric contractions. Journal of AppliedPhysiology 88, 91–101.
MC, K. K. & F, J. A. (1985). Injury to skeletal muscle
fibers of mice following lengthening contractions. Journal ofApplied Physiology 59, 119–126.
M, D. L. (1990). New insights into the behavior of muscle
during active lengthening. Biophysical Journal 57, 209–221.
M, D. L. & A, D. G. (1999). Early events in stretch-
induced muscle damage. Journal of Applied Physiology 87,
2007–2015.
N, O. B. & C, T. (1996). The significance of active
Na+,K+ transport in the maintenance of contractility in rat skeletal
muscle. Acta Physiologica Scandinavica 157, 199–209.
P, U. & M, D. L. (2001). Muscle damage from eccentric
exercise: mechanism, mechanical signs, adaptation and clinical
applications. Journal of Physiology 537, 333–345.
T, H., F, N., N, T., O, H. &
K, N. (2001). Eccentric exercise-induced morphological
changes in the membrane systems involved in
excitation–contraction coupling in rat skeletal muscle. Journal ofPhysiology 533, 571–583.
T, J. A. & M, D. L. (1996). Quantitative analysis of
sarcomere non-uniformities in active muscle following a stretch.
Journal of Muscle Research and Cell Motility 17, 261–268.
T, J. A. & M, D. L. (1998). The effects of stretch
parameters on eccentric exercise-induced damage to toad skeletal
muscle. Journal of Muscle Research and Cell Motility 19, 237–245.
V, R. A. & H, P. (1981). Density and apparent
location of the sodium pump in frog sartorius muscle. Journal ofMembrane Biology 59, 225–232.
W, G. L., I, C. P., L, D. A. & A, R. B.
(2001). Excitation–contraction uncoupling: major role in
contraction-induced muscle injury. Exercise and Sport ScienceReview 29, 82–87.
W, G. L., L, D. A., H, D. A., F, M. A. &
A, R. B. (1995). Redistribution of cell membrane
probes following contraction-induced injury of mouse soleus
muscle. Cell Tissue Research 282, 311–320.
W, G. L., L, D. A., H, D. A., K, C. J., P,
B. M. & A, R. B. (1993). Excitation failure in eccentric
contraction-induced injury of mouse soleus muscle. Journal ofPhysiology 468, 487–499.
W, M. W., R, W. G., K, T., U, J. A.,
B, C. S., B, J. E. & B, R. J. (2001). Na,K-
ATPase in skeletal muscle: two populations of b-control
localization in the sarcolemma but not partitioning between the
sarcolemma and the transverse tubules. Journal of Cell Science 114,
751–762.
W, S. A., M, D. L. & P, U. (1993). Effects of
repeated eccentric contractions on structure and mechanical
properties of toad sartorius muscle. American Journal of Physiology265, C792–800.
Y, E. W., B, J.-P., A, D. G. & B, H. J.
(2002). The effect of eccentric contraction-induced injury on force
and intracellular pH in rat skeletal muscles. Journal of AppliedPhysiology 92, 93–99.
AcknowledgementsThe confocal microscope used in these studies was funded from aCollaborative Research Grant with Pfizer, research funds from theUniversity of Sydney and an equipment grant from the NationalHealth and Medical Research Council of Australia. The workdescribed will be submitted to the University of Hong Kong byElla W. Yeung as part of her doctoral thesis.
E. W. Yeung and others592 J. Physiol. 540.2
Copyright © 2022 FDOKUMEN




















