
A Glimpse into the Evolutionary Journey of Podocytes in Culture
Upload
independentCategory
view
1download
0
The American Journal of Pathology, Vol. 178, No. 5, May 2011
Copyright © 2011 American Society for Investigative Pathology.
Published by Elsevier Inc. All rights reserved.
DOI: 10.1016/j.ajpath.2011.01.017
Cell Injury, Repair, Aging, and Apoptosis
Ubiquitin C-Terminal Hydrolase-L1 Activity InducesPolyubiquitin Accumulation in Podocytes andIncreases Proteinuria in Rat Membranous
NephropathyCatherine Meyer-Schwesinger,* Tobias N. Meyer,*Henning Sievert,† Elion Hoxha,* Marlies Sachs,*Eva-Maria Klupp,* Silvia Münster,*Stefan Balabanov,† Lucie Carrier,‡§¶
Udo Helmchen,� Friedrich Thaiss,* andRolf A.K. Stahl*From the Department of Internal Medicine, Nephrology,* and the
the Department of Experimental and Clinical Pharmacology and
Toxicology,‡ Cardiovascular Research Center, Department of
Pathology,� Nierenregister, University Medical Center Hamburg-
Eppendorf, Hamburg, Germany; University Cancer Center
Hamburg-Eppendorf,† Hamburg, Germany; Inserm U974,§ Paris,
France; and the Institut de Myologie,¶ Université Pierre et Marie
Curie-Paris, UMR-S974, CNRS UMR7215, IFR14, Paris, France
Ubiquitin C-terminal hydrolase L1 (UCH-L1), a keyprotease of the ubiquitin-proteasome system (UPS),is associated with neurodegenerative diseases andcancer. Recently, de novo expression of UCH-L1 wasdescribed in podocytes in patients with membra-nous nephropathy (MN), in which UCH-L1 expres-sion correlated with increased ubiquitin content.The objective of the present study was to investigatethe role of UCH-L1 in ubiquitin homeostasis andproteasomal degradation in a rat model of MN. Afterdisease induction, UCH-L1 expression increased inpodocytes and coincided with decreased glomeru-lar monoubiquitin content. After an initial increasein proteasomal activity, the UPS was impaired. Inaddition to an increase of ubiquitin in podocytes,aggregates were observed 1 year after disease induc-tion, as in MN in human beings. Inhibition ofUCH-L1 hydrolase function in MN reduced UPS im-pairment and ameliorated proteinuria. In contrast,inhibition of proteasomal activity enhanced UPSimpairment, resulting in increased proteinuria. Sta-ble UCH-L1 overexpression in cultured podocytesresulted in accumulation of monoubiquitin and
polyubiquitin proteins. In contrast, stable knock-2044
down of UCH-L1 reduced monoubiquitin and polyu-biquitin proteins and significantly increased pro-teasomal activity, indicating that the observedeffects in rat MN also occurred in cultured podo-cytes. These data demonstrate that UCH-L1 activityresults in polyubiquitin accumulation, proteasomeinhibition, and disease aggravation in experimentalmodels of MN. (Am J Pathol 2011, 178:2044–2057; DOI:10.1016/j.ajpath.2011.01.017)
Podocytes have a major role in maintaining integrity of theglomerular filtration barrier.1 Podocyte injury is a key el-ement of proteinuric glomerular diseases.2–4 Studies per-formed using a rat model of membranous nephropathy(MN), passive Heymann nephritis (PHN), suggest thatimmune deposits damage podocytes through comple-ment-dependent processes (C5b-9), which lead to alter-ations of the podocyte cytoskeleton.5 Intracellular proteindegradation is a tightly regulated process that maintainsnormal cellular homeostasis; however, little is knownabout its role in podocyte injury. Multiple systems exist forproteolysis, the best described of which is the highlyconserved nonlysosomal proteolytic ubiquitin-protea-some system (UPS). This pathway comprises enzymesthat ubiquitinate or deubiquitinate target proteins and the26S multimeric proteasome complex that degrades ubiq-uitin-conjugated proteins. The selective degradation ofproteins via the UPS involves activation of a signaling
Supported by Werner-Otto Stiftung 2/74 and Else-Kröner-Fresenius Stif-tung P64/09 // A111/09 (C.M.-S.), and Deutsche ForschungsgemeinschaftFOR-604/2, CA 618/1-2 (L.C.).
C.M.-S. and T.N.M. contributed equally to this work
Accepted for publication January 31, 2011.
Supplemental material for this article can be found at http://ajp.amjpathol.org or at doi: 10.1016/j.ajpath.2011.01.017.
Address reprint requests to Catherine Meyer-Schwesinger, M.D.,Department of Internal Medicine, Nephrology, University Medical Cen-ter Hamburg-Eppendorf, Martinistrasse 52, 20246 Hamburg, Germany.
E-mail: [email protected].
UCH-L1 in Membranous Nephropathy 2045AJP May 2011, Vol. 178, No. 5
cascade that generates the covalent attachment of apolyubiquitin chain to protein targets at Lys48.6 Specificpolyubiquitin at Lys48 acts as a signal for degradation bythe proteasome.7 Deubiquitinating activities can promoteaccumulation of monomeric ubiquitin and can also coun-teract the effects of ubiquitin conjugation by removing thepolyubiquitin chain from conjugated proteins before theirdegradation by the proteasome.8,9 Deubiquitinating en-zymes can be subdivided into two groups, high-molecu-lar-weight ubiquitin isopeptidases and low-molecular-weight ubiquitin C-terminal hydrolases (UCHs).10 Threehuman UCH isoenzymes have been cloned, which ex-hibit marked tissue specificity.10 UCH-L1 is a member ofthe UCH family, and is widely expressed in neuronaltissues, testis, ovaries, and synovial membranes.11 Bio-chemically, UCH isoforms process precursors of freeubiquitin and recycle ubiquitin from degraded sub-strates.12 Several biological effects of UCH-L1 are basedon its ability to bind to and stabilize monoubiquitin13 andto perform ubiquitin ligase activity at Lys63.14
UCH-L1 is associated with neurodegenerative disor-ders such as Parkinson’s and Alzheimer’s disea-ses.14–16 In these diseases, protein degradation via theubiquitin pathway is deregulated, leading to polyubiquitinaccumulation.17 In the kidney, UCH-L1 is expressed intubular collecting duct epithelial cells and parietal epithe-lial cells.18,19 Studies of kidney cells or animal models ofrenal diseases have suggested a role of UCH familyenzymes in kidney development, cellular differentiation ofgrowing renal tubules, and cell cycle regulation.20,21 Werecently described an up-regulation of UCH-L1 and acorrelation between UCH-L1 and ubiquitin levels in podo-cytes in human glomerular injuries, in particular, MN. Wedemonstrated that in vitro UCH-L1 expression was thefeature of an undifferentiated podocyte and that podo-cyte differentiation into an arborized phenotype requiresUCH-L1 down-regulation.22 The present study investi-gated UCH-L1 expression and its functional role for ubiq-uitin homeostasis and proteasomal degradation in MN inthe rat and in cultured podocytes.
Materials and Methods
Passive Heymann Nephritis
PHN was induced in 200- to 250-g male Sprague-Dawleyrats via intravenous injection of 500 �L (day 1) and 750�L (day 0) of sheep anti-Fx1A antiserum (PHN rats) orcontrol sheep preimmune serum (control rats). Rats re-
Table 1. shRNA Sequences against UCH-L1 Used in the Present
Oligo forpLKO.1 shRNA
inserts Forward primer
#344 5=-CCGGTAAACTTTAGGGCTAACTTCCTGGACAGGAAGTTAGCCCTAAAGTTTATTT
#627 5=-CCGGGGGTAGATGACAAAGTGAATTTCTGGAAATTCACTTTGTCATCTACCCTTT
#817 5=-CCGGCGTGGCTCTCTGCAAAGCAGCTT
TTAAGCTGCTTTGCAGAGAGCCACGTTTTTG-3ceiving neither FX1A antiserum or preimmune serumwere termed untreated rats. Anti-Fx1A antiserum wasinduced via repeated immunization of sheep with isolatedbrush borders of proximal tubules from rat kidneys, asdescribed.23 Kidneys were harvested at 0, 2, 4, 6, 9, 14,21, and 28 days, 5 weeks, and 1 year. In one set ofexperiments, PHN was induced successively (days 4, 6,and 28), and glomeruli were isolated at the same times.For UCH-L1 inhibition, LDN57444 [40 �g/kg body weightin dimethyl sulfoxide (DMSO)], and for proteasomal inhi-bition, MG132 (60 �g/kg body weight in 15% ethanol),were introduced in Alzet osmotic minipumps (model 2ML;Alza Corp., Palo Alto, CA) and implanted subcutaneouslyon day 4 or 14 after the last serum injection in control andPHN rats.
Animal Care
Rats were housed in a controlled animal facility with freeaccess to water and standard rat chow. Animal experimentswere performed according to national and institutional ani-mal care and ethical guidelines, and were approved by thelocal ethics committees (82/06 and 107/09).
Cell Culture
Murine podocytes were cultured as described previ-ously.24 For differentiation, podocytes were cultured for14 days under nonpermissive conditions (37°C, 7.4%CO2, RPMI 1640 supplemented with 10% fetal calf se-rum, 10 mmol/L N-2-hydroxyethylpiperazine-N0-2-eth-anesulfonic acid, 1 mmol/L sodium pyruvate, 100 U/mLpenicillin, and 100 mg/mL streptomycin). For protea-somal inhibition experiments, podocytes were culturedfor 7 hours with 2 �mol/L MG132 (Calbiochem, San Di-ego, CA) in DMSO or equal amounts of DMSO for controlconditions.
Plasmid Production
For overexpression experiments, the murine UCH-L1cDNA (NM_011670) coding sequence was cloned intopMSCV (murine stem cell virus) puro expression vectors(BD Bioscience, Clontech Laboratories, Inc., San Jose,CA). First, the UCH-L1 cDNA coding sequence was am-plified from total cDNA via PCR (Phusion polymerase;Finnzymes Oy, Vantaa, Finland). The forward primer usedwas 5=-CTAGAGATCTGCCACCATGCAGCTGAAGCC-GATGGA-3=, and reverse primer was 5=-CTAGCTC-
Reverse primer
GAG=
5=-AATTCAAAAATAAACTTTAGGGCTAACTTCCTGTCCTCGAGGACAGGAAGTTAGCCCTAAAGTTTA-3=
GAG=
5=-AATTCAAAAAGGGTAGATGACAAAGTGAATTTCCACTCGAGTGGAAATTCACTTTGTCATCTACCC-3=
GAG 5=-AATTCAAAAACGTGGCTCTCTGCAAAGCAGCTTA
Study
TCCTCTTG-3CACTCTTG-3AACTC
= ACTCGAGTTAAGCTGCTTTGCAGAGAGCCACG-3=

estern b
2046 Meyer-Schwesinger et alAJP May 2011, Vol. 178, No. 5
GAGTTAAGCTGCTTTGCAGAGAG-3=. PCR products andvectors were digested using BglII/XhoI (Fermentas Inc.,Glen Burnie, MD), ligated, and transformed into Escherichiacoli. For stable knock-down experiments, small hairpin RNA(shRNA) was designed based on three Stealth small inter-fering RNA sequences (Invitrogen Corp., Carlsbad, CA).The plasmids were produced as described by Moffat etal.25 In short, two oligos for each sequence (Table 1) wereannealed. pLKO.1 plasmid was digested with EcoRI andAgeI, and annealed with 2 �L oligos and transformed into E.coli. Positive clones were validated via sequencing.
Virus Production and Target Cell Infection
Phoenix ecotropic packaging cells (for packaging of pM-SCV into retroviral particles) and 293T packaging cells(for packaging of pLKO.1 plasmids into lentiviral parti-cles) were transfected using DNA-CaCl2 precipitate. Forlentiviral particles, 10 �g plasmid DNA was mixed with 10�g EcoEnv helper plasmid and 5 �g each of Rev andGag/Pol helper plasmids. The virus-containing superna-tant was transferred to a 10-cm plate containing podo-cyte target cells at approximately 50% to 60% conflu-ence, and the infection steps were repeated twice.Selection for integration of the expression plasmids wasperformed using puromycin 1.5 �g/mL or G418 500 �g/mL. For negative control experiments, the plasmids weretransduced without insert or with inserted scrambledshRNA.
Human Specimen Collection
The study was conducted according to the Declaration ofHelsinki principles, with approval from the local ethicscommittee. Tissue samples were obtained between 2007and 2008 from the Nierenregister of the University Med-ical Center Hamburg-Eppendorf.
Histologic Analysis
Kidneys were fixed in 4% paraformaldehyde, embeddedin paraffin or in Lowicryl K4M resin (Electron MicroscopySciences, Hapfield, PA), and further processed for PAS
Table 2. Antibodies Used in the Present Study
AntibodyCatalog
no./Clone
UCH-L1 13C4 Abcam PLC, Cambridge, MAUCH-L1 CL95101 Biozol Diagnostica Vertrieb GUbiquitin MAB1510 Millipore Corp. (Chemicon In�5 Xuejun Wang, PhD, UniversitFITC-C3 55500 Cappel Laboratories, Inc., CCy2-� rat 712225153 Jackson ImmunoResearch LCy2-� sheep 713225147 Jackson ImmunoResearch L�-Actin AC-15 Sigma-Aldrich, St. Louis, MOCollagen type 4 1340-01 Southern Biotech, Birmingha�-Actinin-4 0042-05 immunoGlobe Antikörpertech
IF, immunofluorescence staining; IHC, immunohistochemistry; WB, W
staining or electron microscopy as described.26
IHC (UCH-L1, Ubiquitin, Collagen Type 4,�-Actinin-4)
For immunohistochemistry (IHC), 1-�m paraffin sec-tions were deparaffinized, and antigen retrieval wasperformed via microwave boiling (10 mmol/L citratebuffer, pH 6.1) or protease digestion [protease XXIV(Sigma-Aldrich, St. Louis, MO), 5 mg/mL]. Unspecificbinding was blocked (5% horse serum for 30 minutesat room temperature). Primary antibody (Table 2) incu-bation (5% horse serum overnight at 4°C) was followedby incubation with biotinylated secondary antibodies(1:400 for 30 minutes at room temperature). Color de-velopment was performed using the ABC-AP kit (Vec-tor Laboratories, Inc., Burlingame, CA) or the TSA kit(NEN Life Science Products, Inc., Boston, MA) accord-ing to the manufacturers’ instructions; neufuchsin andnuclei were counterstained using hematoxylin. Stain-ings were evaluated using an Axioskop, and were pho-tographed using an Axiocam HRc and Axiostar soft-ware (Carl Zeiss AG, Jena, Germany) or using confocalmicroscopy with an LSM 510 Meta microscope andLSM software (Carl Zeiss AG).
Ubiquitin Immunofluorescence on Podocytes
Podocytes were fixed with 4% paraformaldehyde (EMSciences, Gibbstown, NJ) and permeabilized with0.05% Triton X-100. Unspecific binding was blockedwith 5% horse serum for 30 minutes at room tempera-ture before incubation with mouse anti-ubiquitin in 5%horse serum overnight at 4°C. Staining was visualizedusing biotinylated anti-mouse secondary antibody(Jackson ImmunoResearch Laboratories Inc., WestGrove, PA) 1:200, in 5% horse serum for 30 minutes atroom temperature followed by fluorescein isothiocya-nate (FITC)– conjugated avidin D (Vector Laboratories,Inc.), 1:100, in PBS for 30 minutes at room tempera-ture. Negative controls were performed by omitting theprimary antibody. Podocytes with ubiquitin aggregateswere counted in 40 high-power visual fields (1000�)and normalized against podocyte number per visual
Application (Dilution)
IHC (1:40)Eching, Germany WB (1:1000)
onal, Inc.), Temecula, CA WB (1:1000); IHC, IF (1:50)uth Dakota WB (1:1000)ville, PA IF (1:50)ries Inc., West Grove, PA IF (1:200)ries Inc., West Grove, PA IF (1:200)
WB (1:3000)IF (1:400)
bH, Himmelstadt, Germany IF (1:200)
lot analysis.
Source
mbH,ternatiy of Soochranaboratoaborato
m, ALnik Gm
field.
UCH-L1 in Membranous Nephropathy 2047AJP May 2011, Vol. 178, No. 5
Immunoblot Analysis
Immunoblots were performed as described previously.27
In brief, samples were lysed in tissue protein extractionreagent (T-PER; Pierce Biotechnology, Inc., Rockford, IL)containing phosphatase inhibitors, and denatured with4X lithium dodecyl sulfate. Samples were separated on a4% to 12% Bis-Tris NuPage gel (Invitrogen Corp.) inNuPage running buffer. Protein transfer was performed intransfer buffer (50 mmol/L Tris base, 0192 mmol/L glycinein double-distilled water) in a Novex Mini Cell (InvitrogenCorp.). Polyvinylidine fluoride membranes (MilliporeCorp., Bedford, MA) were blocked (3% nonfat milk) be-fore incubation with primary antibodies diluted in Super-block blocking reagent (Pierce Biotechnology, Inc.).Binding was detected via incubation with horseradishperoxidase–coupled secondary antibodies (1:10,000;3% nonfat milk). Protein expression was visualized usingECL SuperSignal (Pierce Biotechnology, Inc.) accordingto the manufacturers’ instructions, using Kodak BiomaxLight Film (Eastman Kodak Co., Rochester, NY). Immu-noblots were analyzed using ImageJ software.28
Real-Time Quantitative PCR
Total mRNA was isolated from glomeruli using the RLTPlus kit (Qiagen, Hilden, Germany) according to the man-ufacturers’ instructions, and 1 �g was reverse tran-scribed with random hexamer primer (Invitrogen Corp.)and Moloney murine leukemia virus reverse transcriptase(New England Biolabs, Inc., Ipswich, MA). mRNA expres-sion was quantified using AbiPrism NN8860, using SYBRGreen dye, as recently described.29 Exon spanning primerpairs used were as follows: UCH-L1 forward 5=-AGCTGGAATTTGAGGATGGA-3= and reverse 5=-GGCCTCGTTCTTCTCGAAA-3=; 18S forward 5=-CACGGCCGGTACAGTGAAAC-3= and reverse 5=-AGAGGAGCGAGCGACCAAA-3=. 18S was used as an internal control to cor-rect for small variations in mRNA quality and cDNA synthe-sis, essentially as described using AbiPrism. Amplicons ofrandom samples for each primer pair were determined us-ing automatic PCR sequencing to demonstrate the speci-ficity of the PCR reaction (data not shown). Relative quan-tification of gene expression was calculated using the ��CTmethod.
Assessment of Proteinuria
Urine albumin content was quantified using a commer-cially available enzyme-linked immunosorbent assay(Bethyl Laboratories, Inc., Montgomery, TX) according tothe manufacturers’ instructions and as described previ-ously.27 Urine albumin values were standardized againsturine creatinine values for the same animals.
Proteasome Activity Assays
For measurement of proteasomal activity, 10 �g totalprotein was diluted in incubation buffer [20 mmol/LHEPES, 0.5 mmol/L EDTA, 5 mmol/L dithiothreitol, 0.1
mg/mL ovalbumin, 6 mmol/L ATP, 50 mmol/L phospho-creatine, and 0.2 U phosphocreatine kinase in water (pH7.8)] to a final volume of 50 �L. Samples were preincu-bated in incubation buffer for 2 hours at 4°C. After pre-incubation, the substrates Suc-LLVY-AMC (chymotryp-sin-like activity), BZ-VGA-AMC (trypsin-like activity), orZ-LLE-AMC (caspase-like activity) (all from Calbiochem;EMD Chemicals, Inc, division of Merck KGaA, Darmstadt,
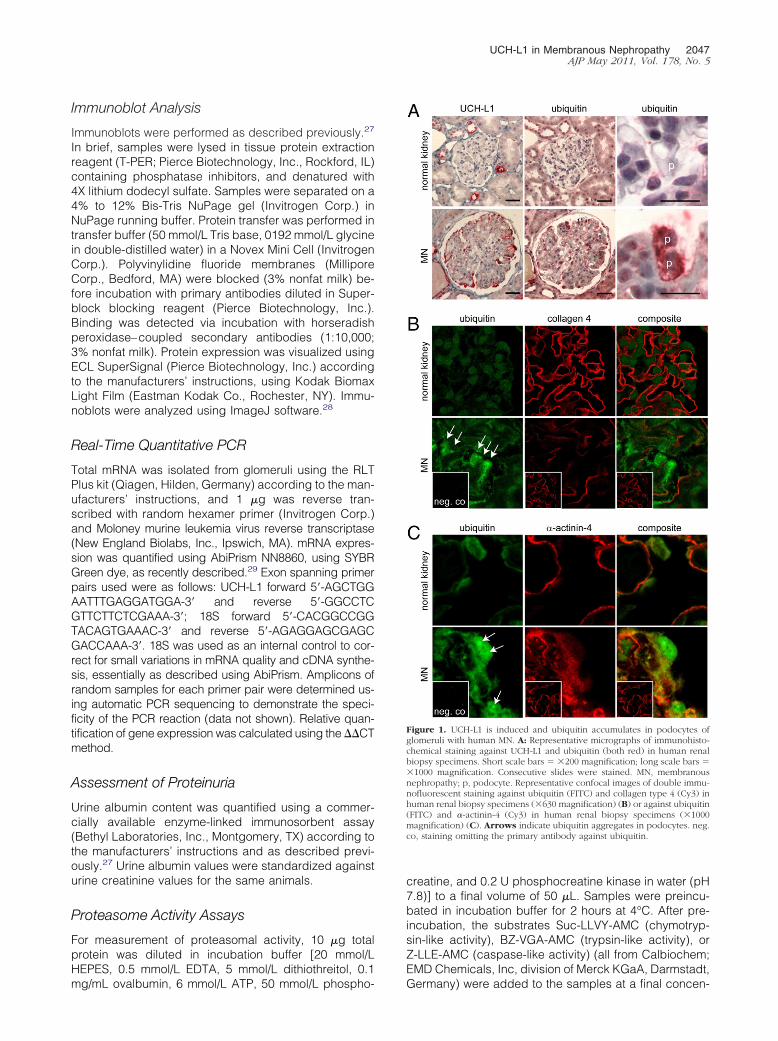
Figure 1. UCH-L1 is induced and ubiquitin accumulates in podocytes ofglomeruli with human MN. A: Representative micrographs of immunohisto-chemical staining against UCH-L1 and ubiquitin (both red) in human renalbiopsy specimens. Short scale bars � �200 magnification; long scale bars ��1000 magnification. Consecutive slides were stained. MN, membranousnephropathy; p, podocyte. Representative confocal images of double immu-nofluorescent staining against ubiquitin (FITC) and collagen type 4 (Cy3) inhuman renal biopsy specimens (�630 magnification) (B) or against ubiquitin(FITC) and �-actinin-4 (Cy3) in human renal biopsy specimens (�1000magnification) (C). Arrows indicate ubiquitin aggregates in podocytes. neg.co, staining omitting the primary antibody against ubiquitin.
Germany) were added to the samples at a final concen-

2048 Meyer-Schwesinger et alAJP May 2011, Vol. 178, No. 5
tration of 60 �mol/L (chymotrypsin-like), 20 �mol/L (tryp-sin-like), and 200 �mol/L (caspase-like). Proteasomal ac-tivity was measured at 355 and 460 nm using afluorescent spectrophotometer (Mithras LB 940; BertholdTechnologies GmbH & Co., Wildbad, Germany) after in-cubation at 37°C for 1 hour in the dark.
Statistical Analyses
Data are given as the mean (SEM). Statistical comparisonswere performed using commercially available software(PRISM; GraphPad Software, Inc., San Diego, CA). TheMann-Whitney U-test was used for nonparametric data, andthe unpaired Student’s t-test for parametric data. P � 0.05was accepted as statistically significant.
Results
In Human MN, podocytes exhibited a massive accumula-tion of UCH-L1 at immunohistochemical staining, comparedwith podocytes from normal biopsy specimens (Figure 1A),as demonstrated previously.22 Ubiquitin accumulation inpodocytes correlated with increased UCH-L1 expression inpodocytes in biopsy specimens from patients with MN.In addition, ubiquitin aggregates were observed in podo-cytes in three of eight MN cases (Figure 1, B and C).
To understand the role of UCH-L1 in podocytes andits potential function in ubiquitin accumulation in hu-man MN, experimental MN was induced in the rat.5 Toinduce MN, sheep antiserum directed against brushborders of rat proximal tubules was injected intrave-nously into rats, producing linear and granular IgGdeposits, proteinuria, and complement activation. Theprotein concentration in urine remained elevated dur-ing the entire 1-year observation, with normal serumurea nitrogen and creatinine concentrations, as de-scribed by others.30 Light microscopy demonstratedthat the podocytes had hypertrophied and the glomer-ular basement membrane had thickened. Ultrastructur-ally, foot process effacement, subepithelial immunedeposits, and disruption of the glomerular basementmembrane resembled human MN (see SupplementalFigure S1 at http://ajp.amjpathol.org).
UCH-L1 Concentration Is Increased inPodocytes in Rat MN
It was first analyzed whether podocyte injury leads to denovo expression of UCH-L1 in podocytes in rat MN (PHN)similar to the increase observed in human MN. The firstimmunohistochemical detection of UCH-L1 expression inpodocytes was observed at 6 days after disease induction(Figure 2A). Thereafter, the glomerular expression pattern ofUCH-L1 in PHN was restricted to podocytes, in addition tothe “normal” expression of UCH-L1 in parietal epithelialcells,18 and was comparable to the expression pattern ob-served in human MN. Quantification of glomerular UCH-L1levels using Western blot analysis demonstrated higherUCH-L1 levels from day 6 and thereafter in PHN rats com-
pared with control rats (Figure 2, B and C). Mean UCH-L1protein levels remained stably elevated throughout the1-year observation period despite some heterogeneityamong individual animals. RT qPCR analysis of glomerularlysates from day 0 to day 35 demonstrated increased rela-tive expression of UCH-L1 mRNA in PHN glomeruli com-pared to untreated rats, starting immediately after injectionof the antiserum (Figure 2D). Injection of unspecific preim-mune serum (control rats) also increased UCH-L1 mRNAlevels up to day 6 compared with untreated rats in a patterncomparable to that in PHN rats. However, at the proteinlevel, no UCH-L1 expression was detected in podocytes incontrol rats after injection of unspecific preimmune serum.The early increase of relative UCH-L1 mRNA expression inthe PHN and control groups compared with the untreated ratsdemonstrated that the glomerular passage of immunoglobulinwas possibly sufficient to induce changes in glomerularUCH-L1 mRNA levels, which, however, resulted in increasedUCH-L1 protein only in PHN rats. This divergence betweenmRNA and protein levels in control rats compared with PHNrats indicates another or additional pathway of UCH-L1 proteinregulation (ie, proteasomal degradation) in PHN rats beforeday 9 rather than increased mRNA levels. After day 9, therelative expression of UCH-L1 mRNA was significantly higherin PHN rats compared with preimmune serum-treated controlrats, which suggests a specific reaction of UCH-L1 to podo-cyte damage induced by specific sheep antiserum.
Ubiquitin Accumulates and Aggregates inPodocytes in Rat MN
Since ubiquitin accumulation correlated with increasedUCH-L1 expression in podocytes in human MN, we in-vestigated whether ubiquitin accumulates in rat MN. Byimmunochemistry, ubiquitin was detected at low levels incontrol podocytes and during the early stages of rat MN.At day 35, strong ubiquitin staining was observed in PHNpodocytes, confirming that the accumulation of ubiquitinoccurred predominantly in podocytes (Figure 3A). Oneyear after disease induction, immunofluorescence dem-onstrated ubiquitin aggregates in PHN podocytes, asobserved in human MN, whereas control podocytes didnot exhibit ubiquitin aggregates (Figure 3B). These datademonstrated that ubiquitin accumulation and aggregateformation occurred in human and rat MN in a comparablepattern.
Polyubiquitin Proteins Accumulate in Glomeruliin Rat MN
Glomerular protein lysates were generated, and the dis-tribution and levels of ubiquitinated proteins were inves-tigated in PHN versus control animals using Western blotanalysis. The studies were performed by comparing PHNand control (preimmune serum injected) glomeruli iso-lated successively day 4, 6, 28, or 365 after diseaseinduction day 0 (Figure 4, A and B) or in glomeruli fromPHN and control rats isolated the same day after succes-sive disease induction (Figure 4, C and D). Preimmuneserum injected glomeruli at days 4, 6, and 28 demon-
strated no significant changes in monomeric and poly-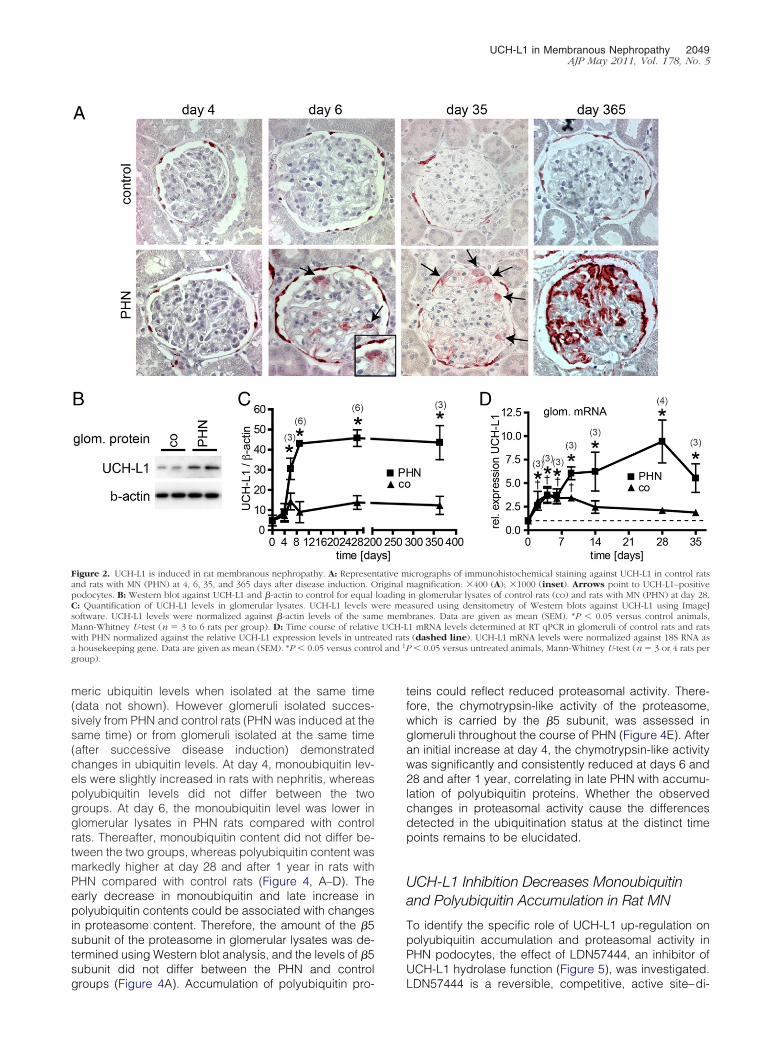
ol and P
UCH-L1 in Membranous Nephropathy 2049AJP May 2011, Vol. 178, No. 5
meric ubiquitin levels when isolated at the same time(data not shown). However glomeruli isolated succes-sively from PHN and control rats (PHN was induced at thesame time) or from glomeruli isolated at the same time(after successive disease induction) demonstratedchanges in ubiquitin levels. At day 4, monoubiquitin lev-els were slightly increased in rats with nephritis, whereaspolyubiquitin levels did not differ between the twogroups. At day 6, the monoubiquitin level was lower inglomerular lysates in PHN rats compared with controlrats. Thereafter, monoubiquitin content did not differ be-tween the two groups, whereas polyubiquitin content wasmarkedly higher at day 28 and after 1 year in rats withPHN compared with control rats (Figure 4, A–D). Theearly decrease in monoubiquitin and late increase inpolyubiquitin contents could be associated with changesin proteasome content. Therefore, the amount of the �5subunit of the proteasome in glomerular lysates was de-termined using Western blot analysis, and the levels of �5subunit did not differ between the PHN and control
Figure 2. UCH-L1 is induced in rat membranous nephropathy. A: Represenand rats with MN (PHN) at 4, 6, 35, and 365 days after disease induction. Opodocytes. B: Western blot against UCH-L1 and �-actin to control for equalC: Quantification of UCH-L1 levels in glomerular lysates. UCH-L1 levels wsoftware. UCH-L1 levels were normalized against �-actin levels of the samMann-Whitney U-test (n � 3 to 6 rats per group). D: Time course of relativwith PHN normalized against the relative UCH-L1 expression levels in untrea housekeeping gene. Data are given as mean (SEM). *P � 0.05 versus contrgroup).
groups (Figure 4A). Accumulation of polyubiquitin pro-
teins could reflect reduced proteasomal activity. There-fore, the chymotrypsin-like activity of the proteasome,which is carried by the �5 subunit, was assessed inglomeruli throughout the course of PHN (Figure 4E). Afteran initial increase at day 4, the chymotrypsin-like activitywas significantly and consistently reduced at days 6 and28 and after 1 year, correlating in late PHN with accumu-lation of polyubiquitin proteins. Whether the observedchanges in proteasomal activity cause the differencesdetected in the ubiquitination status at the distinct timepoints remains to be elucidated.
UCH-L1 Inhibition Decreases Monoubiquitinand Polyubiquitin Accumulation in Rat MN
To identify the specific role of UCH-L1 up-regulation onpolyubiquitin accumulation and proteasomal activity inPHN podocytes, the effect of LDN57444, an inhibitor ofUCH-L1 hydrolase function (Figure 5), was investigated.
icrographs of immunohistochemical staining against UCH-L1 in control ratsmagnification: �400 (A); �1000 (inset). Arrows point to UCH-L1–positivein glomerular lysates of control rats (co) and rats with MN (PHN) at day 28.asured using densitometry of Western blots against UCH-L1 using ImageJbranes. Data are given as mean (SEM). *P � 0.05 versus control animals,1 mRNA levels determined at RT qPCR in glomeruli of control rats and rats(dashed line). UCH-L1 mRNA levels were normalized against 18S RNA as� 0.05 versus untreated animals, Mann-Whitney U-test (n � 3 or 4 rats per
tative mriginalloadingere mee mem
e UCH-Lated rats
†
LDN57444 is a reversible, competitive, active site–di-

controland B)
2050 Meyer-Schwesinger et alAJP May 2011, Vol. 178, No. 5
rected isatin oxime with a median inhibition concentrationof 0.88 �mol/L for UCH-L1 and 25 �mol/L for its systemicisoform UCH-L3.31 Two weeks after disease induction,LDN57444 was applied continuously for 14 days usingsubcutaneously implanted osmotic minipumps in the
Figure 3. Ubiquitin accumulates in podocytes during rat membranous nephcontrol (upper row) and PHN rats (lower row) at day 35 after disease inducubiquitin (FITC) and collagen type 4 (Cy3) in control rats (upper panels) ain podocytes of a rat with membranous nephropathy (arrow). The negativetype 4. Short scale bars � �400 magnification; long scale bars � �1000 (A
PHN and control groups. Whereas LDN57444 reducedthe monoubiquitin level in both groups compared withuntreated animals, it reduced and increased the amountof polyubiquitin in the PHN and control groups, respec-tively (Figure 5, A and B). Immunofluorescence demon-
. A: Representative immunohistochemical staining against ubiquitin (red) inRepresentative confocal micrographs of immunofluorescent staining againstrats (lower panels) at 1 year after disease induction. Ubiquitin aggregates
was generated by omitting the primary antibody to ubiquitin and to collagen.
Figure 4. UPS impairment in late MN in the rat. A:Representative Western blots against ubiquitin, �5subunit of the proteasome, and �-actin, as ahousekeeping protein from glomerular lysates ofcontrol and PHN rats collected at days 4, 6, and 28and 1 year after disease induction. Unbrokenarrows point to reduced monoubiquitin levels,and broken arrows to increased levels of polyu-biquitin proteins in PHN rats. B: Western blotquantification of relative glomerular monoubiqui-tin and polyubiquitin levels in PHN and controlrats (co; n � 3, was set 1, dashed line) normal-ized to �-actin values of the same membrane. Dataare given as mean (SEM) *P � 0.05 versus controlrats for monoubiquitin, and **P � 0.05 versuscontrol rats for polyubiquitin, Mann-Whitney U-test (n � 3 to 6 rats per group). C: RepresentativeWestern blots from glomerular lysates collected atthe same time after successive disease induction atdays 4, 6, and 28 against ubiquitin (polyubiquitinlong exposure, monoubiquitin short exposure),and �-actin from the same membrane, as a house-keeping protein from glomerular lysates collectedat the same time after successive disease inductionat days 4, 6, and 28. Unbroken arrows point toreduced monoubiquitin levels, and broken ar-rows to increased levels of polyubiquitin proteinsin PHN rats. D: Western blot quantification ofrelative glomerular monoubiquitin and polyubiq-uitin levels in PHN and control rats (co; n � 3, wasset 1, dashed line). Monoubiquitin and polyubiq-uitin values were normalized to �-actin values ofthe same membrane. Data are given as mean(SEM) *P � 0.05 versus control rats for monoubiq-uitin, and **P � 0.05 versus control rats for polyu-biquitin, Mann-Whitney U-test (n � 3 rats pergroup). E: Measurement of glomerular chymotryp-sin-like activity of the proteasome in rats with MNat days 0, 4, 6, and 28 and 1 year after diseaseinduction. Data are given as mean (SEM) in per-centage of control rats. *P � 0.05 versus controlrats, Mann-Whitney U-test (n � 3 to 12 rats pergroup).
ropathytion. B:nd PHN

N57444gnificat
UCH-L1 in Membranous Nephropathy 2051AJP May 2011, Vol. 178, No. 5
strated no ubiquitin aggregates in the four treatmentgroups at day 28 (data not shown). In control rats,LDN57444 markedly increased UCH-L1 mRNA levels(Figure 5C). In LDN57444-treated control rats, IHC dem-onstrated higher UCH-L1 levels in parietal epithelial cells
Figure 5. UCH-L1 inhibition reduces UPS impairment in rat MN. Rats rece( � control � LDN, PHN � LDN) via continuous infusion using osmoticRepresentative Western blots against ubiquitin and �-actin, as a housekeetreatment was commenced on day 14 after disease induction. B: Western bcontrol rats (co; n � 3, was set 1, dashed line). Monoubiquitin and polyubiqas mean (SEM). *P � 0.05 versus PHN, and **P � 0.05 versus control rats, Mantranscriptomes of control and PHN rats, determined using RT qPCR. UCH-L1bar indicates the mean. Statistical significance was calculated using the MRepresentative immunohistochemical staining against UCH-L1 in control andindicates UCH-L1–positive nerve ends (positive control for UCH-L1), and sinchymotrypsin-like activity of the proteasome in control and PHN rats. Data areU-test (n � 6 rats). **P � 0.05 versus control rats, and *P � 0.001 versus contusing an enzyme-linked immunosorbent assay and normalized against urineas mean (SEM) Statistical significance was calculated using the Mann-Whitneeffacement is prevented in PHN rats with LDN57444 treatment commenced abarrier in control and PHN rats at day 18 with or without treatment with LDmembrane. Arrows indicate subepithelial immune depositions. Original ma
and additional UCH-L1 expression in podocytes, com-
pared with untreated control rats (Figure 5D). In contrastto control rats, LDN57444 did not significantly affect theUCH-L1 mRNA and protein levels in rats with PHN. To-gether, these data suggest that UCH-L1 activity in injuredpodocytes induced accumulation of monoubiquitin and
er 50% DMSO ( � control, PHN) or 40 �g/kg/d LDN57444 in 50% DMSOps during 2 weeks starting at day 4 or day 14 after disease induction. A:tein from glomerular lysates, at day 28 in control and PHN rats in whichtification of relative glomerular monoubiquitin and polyubiquitin levels in
ues were normalized to �-actin values of the same membrane. Data are giveney U-test (n � 6 rats per group). C: Relative UCH-L1 expression in glomerularvels were normalized against 18S RNA as a housekeeping gene. Horizontalhitney U-test (n � 6 rats per group). *P � 0.001 versus control rats. D:s. Original magnification: �200 (D); �1000 (insets). Double-headed arrowded arrows, positive parietal epithelial cells. E: Measurement of glomerulars mean (SEM). Statistical significance was calculated using the Mann-WhitneyF: Urine albumin concentrations (in milligrams per deciliter) were measurede concentrations (in grams per liter) in control and PHN rats. Data are given(n � 12 rats per group). *P � 0.05 versus PHN � LDN. G: Foot process (fp)Representative electron microscopic micrographs of the glomerular filtrationfor 14 days. c � capillary; e � endothelial cell; gbm, glomerular basemention: �700 �12,000 (insets).
ived eithminipumping prolot quanuitin valn-WhitnmRNA le
ann-WPHN ratgle-heagiven a
rol rats.creatininy U-testt day 4.
polyubiquitin in MN. The higher monoubiquitin content is

y U-test
2052 Meyer-Schwesinger et alAJP May 2011, Vol. 178, No. 5
consistent with the known biochemical properties ofUCH-L1, whereas the higher polyubiquitin level was sur-prising and suggests a direct inhibitory effect of UCH-L1on the proteasome.
UCH-L1 Inhibition Increases ProteasomeActivity and Decreases Proteinuria in Rat MN
The chymotrypsin-like activity of the proteasome was mark-edly increased by inhibiting UCH-L1 hydrolase function withLDN57444 in both PHN and control groups compared withuntreated animals (Figure 5E). Inhibition of UCH-L1 activitywith LDN57444 four days after disease induction also sig-nificantly reduced proteinuria (Figure 5F) without affectingdeposition of complement C3 and rat IgG in glomeruli inuntreated and treated PHN rats (data not shown). As amorphologic correlate to ameliorated proteinuria, reducedpodocyte swelling was observed at light microscopy (datanot shown). Furthermore, foot process effacement visual-ized by electron microscopy was reduced by LDN57444
Figure 6. Proteasomal inhibition induces polyubiquitin accumulation and agEtOH ( � co, PHN) or 60 �g/kg/d MG132 in 15% EtOH ( � co � MG, PHN14 days after disease induction. A: Representative Western blots against UCB: Western blot quantification of glomerular polyubiquitin levels relative to�-actin values of the same membrane. Data are given as mean (SEM). *P � 0per group). C: Measurement of chymotrypsin-like activity of the proteasomperformed by adding external MG132 to PHN � MG132 lysates. Data are gU-test (n � 6 rats per group). *P � 0.05 versus control rats, and **P � 0.05staining against ubiquitin (FITC) and collagen type 4 (Cy3) in control and Pindicate podocytes, and double-headed arrows indicate ubiquitin aggregatean enzyme-linked immunosorbent assay, and were normalized against urineas mean (SEM). Statistical significance was calculated using the Mann-WhitnePHN rats.
treatment in PHN rats (Figure 5G). Inhibition of UCH-L1
hydrolase activity at 14 days after disease induction, whenproteinuria was maximal, did not reduce proteinuria signif-icantly (data not shown).
Proteasome Inhibition Increases Monoubiquitinand Polyubiquitin Accumulation in Rat MN
To validate that reduced proteasome activity triggers accu-mulation of polyubiquitin in podocytes during MN, it wasevaluated whether further chemical inhibition of the protea-some resulted in increased polyubiquitin levels. At 14 daysafter disease induction, control and PHN rats were treatedwith MG132, a reversible inhibitor of the 26S proteasome,for 14 days using osmotic minipumps. MG132 increasedpolyubiquitin levels in both control and PHN rats comparedwith control and PHN rats receiving 15% ethanol by osmoticminipumps (untreated rats). (Figure 6, A and B). Chymot-rypsin-like activity was reduced after MG132 treatment incontrol rats, but did not decrease further in PHN rats treatedwith MG132 compared with untreated PHN rats, as would
formation and increases proteinuria in rats with MN. Rats received either 15%) via continuous infusion using osmotic minipumps during 2 weeks startingiquitin, and �-actin, as a housekeeping protein from control and PHN rats.rats (n � 3, set 1, dashed line). Polyubiquitin values were normalized to
us PHN, and **P � 0.05 versus control rats, Mann-Whitney U-test (n � 6 ratsmerular lysates of control and PHN rats. Assay control ( assay control) was
ean (SEM). Statistical significance was calculated using the Mann-WhitneyHN � MG. D: Representative confocal micrographs of immunofluorescent
. Short scale bars � �400 magnification; long scale bars � �1000. Arrowsine albumin concentrations (in milligrams per deciliter) were measured usinge concentrations (in grams per liter) in control and PHN rats. Data are given(n � 6 rats per group). *P � 0.005 versus control rats, and **P � 0.01 versus
gregate� MG
H-L1, ubcontrol.05 verse in gloiven as mversus PHN ratss. E: Urcreatinin
be expected (Figure 6C). One possible explanation is that

UCH-L1 in Membranous Nephropathy 2053AJP May 2011, Vol. 178, No. 5
MG132 is a reversible inhibitor of the chymotrypsin-like ac-tivity located in the proteasomal �5 subunit. It was specu-lated that proteasomal inhibition was reversed in PHN ratsby detachment of MG132 from the catalytic site duringglomerular isolation because addition of external MG132 tothe lysate reduced chymotrypsin-like activity (Figure 6C).MG132 induced ubiquitin aggregates in podocytes of PHNrats (Figure 6D) comparable to the aggregates observed in1-year-old PHN rats. MG132-treated control rats exhibitedincreased ubiquitin content using Western blot analysis andimmunofluorescence, but no aggregates in podocytescompared with untreated control rats. MG132 treatmentsignificantly enhanced proteinuria in MN rats (Figure 6E).Control rats treated with MG132 did not develop proteinuria.Together, these data suggest that proteasomal inhibitionwith MG132 increased monoubiquitin and polyubiquitincontent, induced ubiquitin aggregate formation in podo-cytes of PHN rats and aggravated the disease.
UCH-L1 Overexpression Causes PolyubiquitinAccumulation in Podocytes in Culture
To evaluate whether UCH-L1 activity directly regulated
polyubiquitin accumulation and aggregate formation inpodocytes, UCH-L1 levels were manipulated in cul-tured differentiated podocytes. Stable overexpressionof UCH-L1 was associated with accumulation of polyu-biquitin proteins [Figure 7, A and B (right panel)]. Inaddition, the monoubiquitin level was higher [Figure 7, Aand B (left panel)], and diubiquitin (data not shown) wasdetectable in UCH-L1 overexpressing cells. Conversely,specific and stable knock down of UCH-L1 in three dif-ferent UCH-L1 shRNA constructs decreased the levels ofUCH-L1 and monoubiquitin and polyubiquitin proteins indifferentiated podocytes (Figure 7, A and B). The level ofproteasomal �5 subunit was not changed by UCH-L1overexpression or knock-down (data not shown). Thethree major proteolytic activities of the proteasome (chy-motrypsin-like, trypsin-like, and caspase-like activity)were significantly higher in all UCH-L1 shRNA podocytecell lines, which suggests that UCH-L1 activity inhibitedthe proteasome and caused polyubiquitin accumulation(Figure 7C). Consistent with the results of Western blotanalysis, ubiquitin accumulation was observed by immu-nofluorescence in UCH-L1 overexpressing podocytescompared with control podocytes (Figure 7D). Further-more, ubiquitin aggregates were observed in UCH-L1
Figure 7. UCH-L1 induces polyubiquitin accumu-lation and formation of ubiquitin aggregates incultured podocytes. A: Representative Westernblots against UCH-L1, ubiquitin, and �-actin,which was used to control for equal loading. Leftpanel: Differentiated podocytes stably overex-pressing UCH-L1 or a mock vector as control.Right panel: Three podocyte cell lines in whichUCH-L1 was stably knocked down using variousshRNA constructs against UCH-L1 or scrambledshRNA as a control. B: Western blot quantificationof relative monoubiquitin (left panel) and polyu-biquitin (right panel) levels to control cells(mock and scrambled were set 1, dashed line)from 3 experiments. Monoubiquitin and polyubiq-uitin values were normalized to �-actin values ofthe same membrane. Data are given as mean(SEM, unpaired Student’s t-test (n � 9 per group).*P � 0.05 versus mock, and **P � 0.01 versusscrambled. C: Measurement of chymotrypsin-like,trypsin-like, and caspase-like activity of the pro-teasome in differentiated podocytes in whichUCH-L1 was stably knocked down using shRNAtechnology or scrambled shRNA as a control. As-say control via incubation of scrambled lysateswith MG132 (final concentration 0.5 �mol/L).Data are given as mean (SEM). Statistical signifi-cance was calculated using the unpaired Student’st-test in a total of n � 12 samples of three differentshRNA cell lines or scrambled shRNA as control.*P � 0.001 versus scrambled. D: Representativemicrographs of cultured podocytes overexpress-ing UCH-L1 or mock vector as control stainedagainst ubiquitin. Arrows indicate ubiquitin ag-gregates. Original magnification: �1000. E: Rep-resentative micrographs of cultured podocytesoverexpressing UCH-L1 or mock vector as controland treated with 2 �mol/L MG132 for 7 hours.Cells were stained against ubiquitin. Arrows indi-cate ubiquitin aggregates. Original magnification:�1000.
overexpressing podocytes [mean (SD), 6.5% (4.26%)]

2054 Meyer-Schwesinger et alAJP May 2011, Vol. 178, No. 5
compared with mock-transfected podocytes [0.8%(1.12%)]. Therefore, whether aggregate formation re-quired high UCH-L1 levels and was further inducibleby decreasing proteasomal activity (Figure 7E) wasinvestigated. UCH-L1 overexpressing podocytes weretreated for 7 hours with 2 �mol/L MG132, resulting inpodocytes with high UCH-L1 levels and decreasedchymotrypsin-like activity, similar to observations inPHN rats treated with MG132 or 1-year-old PHN rats.In mock-transfected podocytes, ubiquitin accumula-tion was observed by immunofluorescence; however,no significant aggregates [2.2% (1.63%)] were formed.In contrast, UCH-L1 overexpressing podocytes exhib-ited enhanced [51.2% (2.34%)] aggregate formationafter treatment with MG132. These data indicate thatUCH-L1 activity and proteasomal inhibition together in-duced ubiquitin aggregates, whereas proteasome inhibi-tion was not sufficient to induce aggregates in podocytesin vitro.
Discussion
The role of protein degradation in protecting cells fromdefective misfolded proteins and from oxidative damagehas been the subject of increased attention. The rele-vance to human disease has become apparent in variousdiseases such as cancer and neurodegeneration, inwhich deregulation of protein turnover is associated withdisease states.32 Affected cells experience long-term ac-cumulation of proteins that engage in aberrant protein-protein interactions, form insoluble aggregates, or ac-quire toxic properties. Deregulation of protein turnover inthe course of glomerular injury could, therefore, representa new pathophysiologic concept also in the podocyte.
Studies performed in yeast demonstrated that ade-quate intracellular levels of ubiquitin, in particular at thelevel of monomeric ubiquitin, are maintained in cells andthat under stress, more deubiquitinating enzymes arerequired.33 Mutation of deubiquitinating enzymes in yeastcells resulted in insufficient amounts of monoubiquitinand increased sensitivity to an amino acid analogue,which was compensated for by expression of excessubiquitin.34–36 However, excessive ubiquitin is also notbeneficial. Overexpression of ubiquitin increased sensi-tivity to stress in yeast.37 In our previous study, de novoexpression of the deubiquitinating enzyme UCH-L1 wasdemonstrated in injured podocytes in human MN. Astrong correlation was described between ubiquitin ac-cumulation and expression of UCH-L1 in podocytes.22
Therefore, it was hypothesized that UCH-L1 is regulatedin podocyte injury during MN and that regulation of ubiq-uitin through UCH-L1 may affect protein degradation andbe involved in podocyte injury during MN. During earlydisease, podocytes reacted to antibody-mediated injuryby induction of the UPS, and lower levels of monoubiq-uitin were observed. The low monoubiquitin level possiblyreflected increased ubiquitination processes required forremoval of misfolded or damaged proteins.38 UCH-L1up-regulation and decreased monoubiquitin levels oc-
curred at the same time, which suggests a feedback loopto compensate for decreased monoubiquitin levels. Indi-rect evidence for such a feedback loop comes from gadmice, which are characterized by an intragenic deletionof UCH-L1. In these mice, the lack of UCH-L1 is accom-panied by diminished monoubiquitin levels in the brain.39
In neurons, UCH-L1 is thought to stabilize the monoubiq-uitin pool via hydrolysis of small polyubiquitin chains,monoubiquitin proteins, and ubiquitin precursors.11,40 Inaddition, UCH-L1 stabilizes the level of monoubiquitin bydirect binding to monoubiquitin in neurons.13,41 In thepresent study, UCH-L1 overexpression and knock-downin cultured podocytes resulted in higher and lowermonoubiquitin levels, respectively. In rats, inhibition ofUCH-L1 hydrolase activity lowered glomerular mono-ubiquitin in PHN and normal rats. Therefore, UCH-L1clearly regulated the pool of available monoubiquitin inpodocytes in vitro and in vivo during MN in the rat. Inhi-bition of UCH-L1 hydrolase function by LDN57444 alsoinitiated an increase in UCH-L1 mRNA and protein levelsin control rats and rats with nephritis, which supports thenotion of a feedback loop for UCH-L1 enzymatic activityin podocytes.
After disease duration of 28 days, UCH-L1 and ubiq-uitin content increased in podocytes similar to the in-crease observed in podocytes in patients with MN.Western blot analyses revealed that monoubiquitin levelsdid not differ between PHN and control rats, whereaspolyubiquitin levels were higher in PHN compared withcontrol rats. Polyubiquitin accumulation is a hallmark ofneurodegenerative disorders such as Parkinson’s andAlzheimer’s diseases.42 Modified UCH-L1 activity has animportant role in both diseases,14,43 and is thought to leadto altered protein degradation. Accumulation of polyubiq-uitinated proteins in conjunction with UCH-L1 overexpres-sion has also been observed in the testis of UCH-L1overexpressing mice44 and in UCH-L1 overexpressingCOS cells.45 Similarly, it was demonstrated that UCH-L1overexpressing podocytes contained higher polyubiqui-tin levels. Whether UCH-L1 is directly responsible for theobserved polyubiquitin accumulation is unclear. Two ex-planations are possible: First, UCH-L1 enzymatic activityand provision of monoubiquitin would induce polyubiqui-tin generation. However, ubiquitination of proteins withoutdefect would deplete valuable cellular resources and is,therefore, unlikely. Second, an increase in free ubiquitinchains (ie, generated by UCH-L1) is described to bind toand block ubiquitin binding sites of the proteasome andresult in 26S proteasome inhibition.46 This explanationwould be consistent with the observation that in UCH-L1overexpressing podocytes, not only monoubiquitin butalso ubiquitin chains or ubiquitinated proteins between14 (presumably diubiquitin) and 62 kDa were increased.Proteasomal activity in PHN glomeruli was lower than incontrol rats at all later times. It has further been proposedthat aggregated proteins themselves also directly inhibitthe 26S proteasome by “chocking” the proteases. Alter-natively, protein aggregates may indirectly interfere withUPS function by inactivating or depleting UPS compo-nents.47,48 However, the data in podocyte culture sug-gest that UCH-L1 activity had a direct inhibitory effect on
the proteasome in podocytes. UCH-L1 knock-down in
UCH-L1 in Membranous Nephropathy 2055AJP May 2011, Vol. 178, No. 5
cultured podocytes and inhibition of UCH-L1 hydrolasefunction in PHN rats both resulted in higher proteasomalactivity and lower polyubiquitin content. Direct binding ofUCH-L1 to the proteasome has been described in thesea slug Aplysia. However, in contrast to the observationsin PHN, the hydrolytic activity and the association ofAp-Uch with the proteasome was described to enhancethe proteasomal degradation of polyubiquitinated pro-teins in aplysia.12 Glomerular polyubiquitin accumulationwas also observed in mice with autophagy-deficientpodocytes when they developed severe podocyte dys-function.49 In these mice, polyubiquitin accumulation wasattributed to additional UPS impairment that developswith age in addition to the genetic impairment in au-tophagy. It will be interesting to investigate whetherpolyubiquitin accumulation in late MN is also associatedwith impairment of the autophagy or lysosomal pathwayof degradation.
UCH-L1 is associated with formation of inclusion bodiesin Parkinson’s and Alzheimer’s diseases.50–52 In a Drosoph-ila model of polyglutamine repeat disease, inhibition of ag-gregate formation strongly correlated with suppression ofneuronal degeneration.53 Ubiquitin aggregate formationhas been observed in podocytes in human MN. In rat MN,formation of ubiquitin aggregates was observed at 1 yearafter disease induction, and was accompanied by impair-ment of the UPS. Furthermore, aggregate formation wasinducible in PHN rats and in UCH-L1 overexpressing podo-cytes via additional chemical inhibition of proteasomal ac-tivity. Aggregates were not observed in conditions of pro-teasomal inhibition without elevated UCH-L1 levels, as inMG132-treated control rats or in mock-transfected podo-cytes treated with MG132. These data support the hypoth-esis that aggregate formation in injured podocytes requireshigh levels of UCH-L1 and proteasomal inhibition. In rat MN,aggregates were evident at 1 year after disease induction.Choi et al15 proposed a direct link between oxidative dam-age of UCH-L1 and the pathogenesis of neurodegenerativedisease. In addition, oxidative modifications also affect pro-teasome subunits, resulting in loss of proteasomal activityand accumulation of ubiquitinated proteins.54 Podocyte in-jury from local accumulation of reactive oxygen species isalso considered a major effector mechanism in PHN.55 It is,therefore, possible that continuous oxidative stress leads tooxidative modification of UCH-L1 also in PHN, resulting inaggregate formation after 1 year.
The functional relevance of polyubiquitin accumulationand ubiquitin aggregate formation in human and rat MNis unclear. In the present study animals, proteinuria wassignificantly reduced by UCH-L1 inhibition when inhibi-tion was commenced at 4 days after disease induction.Early PHN is mediated by binding of the injected Fx1Aantibody to podocyte foot processes. This binding in turnmediates binding of rat anti-sheep antibodies to the in-jected Fx1A antibody. This immune deposit activates thecomplement cascade, which results in the formation of asublytical C5b-9 membrane attack complex that may in-jure the podocytes.5,56 In contrast, mediation of late PHNis not well understood. Low-dose isatin oxime(LDN57444) treatment in early PHN does not interfere
with Fx1A antibody binding, generation of rat anti-sheepantibodies, or deposition of complement C3, which indi-cates that disease initiation is not affected by LDN treat-ment commenced at day 4. It is, therefore, assumed thatthere is a direct effect of UCH-L1 inhibition on the podo-cyte. In addition, early UCH-L1 inhibition ameliorates UPSimpairment by increasing the activity of the proteasomeand by reducing polyubiquitin accumulation. It is possi-ble that this results in increased clearance of damagedproteins from the podocytes, which in turn could result inbetter “regeneration” after injury (ie, prevention of cyto-skeletal rearrangements of podocytes on injury). In linewith this argument are electron micrographs that demon-strated prevention of podocyte foot process effacementvia treatment with LDN57444 since day 4. This is mostlikely the morphologic correlate to ameliorated protein-uria. Reduction of polyubiquitin accumulation throughUCH-L1 inhibition starting on day 14 did not changeproteinuria significantly, which suggests that longer du-ration of treatment with UCH-L1 inhibitors is needed,which could result in constant polyubiquitin reductionrequired for clinical amelioration. Another explanationcould be that podocyte cytoskeletal rearrangements andother injury-related processes are too far advanced inlate PHN for UCH-L1 inhibition to be beneficial or thatmediation of late PHN is not dependent on UCH-L1. Pro-teasomal inhibition with MG132-enhanced proteinuriaand polyubiquitin accumulation and aggregate formationindicate that formation of aggregates and increase inpolyubiquitinated proteins could lead to disease progres-sion. Together, these data demonstrate that UCH-L1 ac-tivity in podocytes results in polyubiquitin accumulation,proteasome inhibition, and disease aggravation in ratMN. UPS impairment characterized by polyubiquitin ac-cumulation and aggregate formation is a feature of ratMN and possibly of human MN. Whether persistentUCH-L1 expression and ubiquitin accumulation are newpathophysiologic concepts for chronic podocyte dam-age should be further investigated.
Acknowledgments
We thank Mariola Reszka (University Medical CenterHamburg-Eppendorf, Germany) for excellent technicalassistance, Xuejun Wang (University of South Dakota,Vermillion, SD) for the �5 antibody, and Karl-Hans End-lich (Institute of Anatomy, Greifswald, Germany) for theimmortalized murine podocytes.
References
1. Kriz W, Kretzler M, Provoost AP, Shirato I: Stability and leakiness:opposing challenges to the glomerulus. Kidney Int 1996, 49:1570–1574
2. Mundel P, Shankland SJ: Podocyte biology and response to injury.J Am Soc Nephrol 2002, 13:3005–3015
3. Kriz W, Gretz N, Lemley KV: Progression of glomerular diseases: isthe podocyte the culprit? Kidney Int 1998, 54:687–697
4. Kriz W, Kretzler M, Nagata M, Provoost AP, Shirato I, Uiker S, Sakai T,Lemley KV: A frequent pathway to glomerulosclerosis: deterioration of
tuft architecture-podocyte damage-segmental sclerosis. KidneyBlood Press Res 1996, 19:245–253
2056 Meyer-Schwesinger et alAJP May 2011, Vol. 178, No. 5
5. Couser WG, Nangaku M: Cellular and molecular biology of membra-nous nephropathy. J Nephrol 2006, 19:699–705
6. Hochstrasser M: Ubiquitin, proteasomes, and the regulation of intra-cellular protein degradation. Curr Opin Cell Biol 1995, 7:215–223
7. Goldberg AL: Functions of the proteasome: the lysis at the end of thetunnel. Science 1995, 268:522–523
8. Kalderon D: Protein degradation: de-ubiquitinate to decide your fate.Curr Biol 1996, 6:662–665
9. Rolfe M, Chiu MI, Pagano M: The ubiquitin-mediated proteolytic path-way as a therapeutic area. J Mol Med 1997, 75:5–17
10. Wilkinson KD: Regulation of ubiquitin-dependent processes by deu-biquitinating enzymes. FASEB J 1997, 11:1245–1256
11. Wilkinson KD, Lee KM, Deshpande S, Duerksen-Hughes P, Boss JM,Pohl J: The neuron-specific protein PGP 9.5 is a ubiquitin carboxyl-terminal hydrolase. Science 1989, 246:670–673
12. Hegde AN, Inokuchi K, Pei W, Casadio A, Ghirardi M, Chain DG,Martin KC, Kandel ER, Schwartz JH: Ubiquitin C-terminal hydrolase isan immediate-early gene essential for long-term facilitation in Aplysia.Cell 1997, 89:115–126
13. Sakurai M, Ayukawa K, Setsuie R, Nishikawa K, Hara Y, Ohashi H,Nishimoto M, Abe T, Kudo Y, Sekiguchi M, Sato Y, Aoki S, Noda M,Wada K: Ubiquitin C-terminal hydrolase L1 regulates the morphologyof neural progenitor cells and modulates their differentiation. J CellSci 2006, 119:162–171
14. Liu Y, Fallon L, Lashuel HA, Liu Z, Lansbury PT Jr: The UCH-L1 geneencodes two opposing enzymatic activities that affect alpha-sy-nuclein degradation and Parkinson’s disease susceptibility. Cell2002, 111:209–218
15. Choi J, Levey AI, Weintraub ST, Rees HD, Gearing M, Chin LS, Li L:Oxidative modifications and down-regulation of ubiquitin carboxyl-terminal hydrolase L1 associated with idiopathic Parkinson’s andAlzheimer’s diseases. J Biol Chem 2004, 279:13256–13264
16. Gong B, Cao Z, Zheng P, Vitolo OV, Liu S, Staniszewski A, MoolmanD, Zhang H, Shelanski M, Arancio O: Ubiquitin hydrolase Uch-L1rescues beta-amyloid–induced decreases in synaptic function andcontextual memory. Cell 2006, 126:775–788
17. Chung KK, Dawson VL, Dawson TM: New insights into Parkinson’sdisease. J Neurol 2003, 250(Suppl 3):III15–III24
18. Shirato I, Asanuma K, Takeda Y, Hayashi K, Tomino Y: Protein geneproduct 9.5 is selectively localized in parietal epithelial cells of Bow-man’s capsule in the rat kidney. J Am Soc Nephrol 2000, 11:2381–2386
19. Diomedi-Camassei F, Rava L, Lerut E, Callea F, Van Damme B:Protein gene product 9.5 and ubiquitin are expressed in metabolicallyactive epithelial cells of normal and pathologic human kidney. Neph-rol Dial Transplant 2005, 20:2714–2719
20. Franch HA: Pathways of proteolysis affecting renal cell growth. CurrOpin Nephrol Hypertens 2002, 11:445–450
21. Debigare R, Price SR: Proteolysis, the ubiquitin-proteasome system,and renal diseases. Am J Physiol Renal Physiol 2003, 285:F1–F8
22. Meyer-Schwesinger C, Meyer TN, Munster S, Klug P, Saleem M,Helmchen U, Stahl RA: A new role for the neuronal ubiquitin C-termi-nal hydrolase-L1 (UCH-L1) in podocyte process formation and podo-cyte injury in human glomerulopathies. J Pathol 2009, 217:452–464
23. Salant DJ, Cybulsky AV: Experimental glomerulonephritis. MethodsEnzymol 1988, 162:421–461
24. Schiwek D, Endlich N, Holzman L, Holthofer H, Kriz W, Endlich K:Stable expression of nephrin and localization to cell-cell contacts innovel murine podocyte cell lines. Kidney Int 2004, 66:91–101
25. Moffat J, Grueneberg DA, Yang X, Kim SY, Kloepfer AM, Hinkle G,Piqani B, Eisenhaure TM, Luo B, Grenier JK, Carpenter AE, Foo SY,Stewart SA, Stockwell BR, Hacohen N, Hahn WC, Lander ES, SabatiniDM, Root DE: A lentiviral RNAi library for human and mouse genesapplied to an arrayed viral high-content screen. Cell 2006, 124:1283–1298
26. Meyer-Schwesinger C, Dehde S, von Ruffer C, Gatzemeier S, Klug P,Wenzel UO, Stahl RA, Thaiss F, Meyer TN: Rho kinase inhibitionattenuates LPS-induced renal failure in mice in part by attenuation ofNF-kappaB p65 signaling. Am J Physiol Renal Physiol 2009, 296:F1088–F1099
27. Meyer TN, Schwesinger C, Wahlefeld J, Dehde S, Kerjaschki D,Becker JU, Stahl RA, Thaiss F: A new mouse model of immune-
mediated podocyte injury. Kidney Int 2007, 72:841–85228. Abramoff MMP, Ram SJ: Image processing with ImageJ. Biophoton-ics Int 2004, 11:36–42
29. Panzer U, Steinmetz OM, Reinking RR, Meyer TN, Fehr S, SchneiderA, Zahner G, Wolf G, Helmchen U, Schaerli P, Stahl RA, Thaiss F:Compartment-specific expression and function of the chemokine IP-10/CXCL10 in a model of renal endothelial microvascular injury. J AmSoc Nephrol 2006, 17:454–464
30. Bonegio RG, Fuhro R, Wang Z, Valeri CR, Andry C, Salant DJ,Lieberthal W: Rapamycin ameliorates proteinuria-associated tubulo-interstitial inflammation and fibrosis in experimental membranous ne-phropathy. J Am Soc Nephrol 2005, 16:2063–2072
31. Liu Y, Lashuel HA, Choi S, Xing X, Case A, Ni J, Yeh LA, Cuny GD,Stein RL, Lansbury PT Jr: Discovery of inhibitors that elucidate therole of UCH-L1 activity in the H1299 lung cancer cell line. Chem Biol2003, 10:837–846
32. Kundu M, Thompson CB: Autophagy: basic principles and relevanceto disease. Annu Rev Pathol 2008, 3:427–455
33. Finley D, Ozkaynak E, Varshavsky A: The yeast polyubiquitin gene isessential for resistance to high temperatures, starvation, and otherstresses. Cell 1987, 48:1035–1046
34. Chernova TA, Allen KD, Wesoloski LM, Shanks JR, Chernoff YO,Wilkinson KD: Pleiotropic effects of Ubp6 loss on drug sensitivitiesand yeast prion are due to depletion of the free ubiquitin pool. J BiolChem 2003, 278:52102–52115
35. Papa FR, Hochstrasser M: The yeast DOA4 gene encodes a deubiq-uitinating enzyme related to a product of the human tre-2 oncogene.Nature 1993, 366:313–319
36. Swaminathan S, Amerik AY, Hochstrasser M: The Doa4 deubiquiti-nating enzyme is required for ubiquitin homeostasis in yeast. Mol BiolCell 1999, 10:2583–2594
37. Chen Y, Piper PW: Consequences of the overexpression of ubiquitinin yeast: elevated tolerances of osmostress, ethanol and canavanine,yet reduced tolerances of cadmium, arsenite and paromomycin.Biochim Biophys Acta 1995, 1268:59–64
38. Wilkinson KD: Ubiquitination and deubiquitination: targeting of pro-teins for degradation by the proteasome. Semin Cell Dev Biol 2000,11:141–148
39. Saigoh K, Wang YL, Suh JG, Yamanishi T, Sakai Y, Kiyosawa H,Harada T, Ichihara N, Wakana S, Kikuchi T, Wada K: Intragenicdeletion in the gene encoding ubiquitin carboxy-terminal hydrolase ingad mice. Nat Genet 1999, 23:47–51
40. Das C, Hoang QQ, Kreinbring CA, Luchansky SJ, Meray RK, Ray SS,Lansbury PT, Ringe D, Petsko GA: Structural basis for conformationalplasticity of the Parkinson’s disease–associated ubiquitin hydrolaseUCH-L1. Proc Natl Acad Sci USA 2006, 103:4675–4680
41. Osaka H, Wang YL, Takada K, Takizawa S, Setsuie R, Li H, Sato Y,Nishikawa K, Sun YJ, Sakurai M, Harada T, Hara Y, Kimura I, Chiba S,Namikawa K, Kiyama H, Noda M, Aoki S, Wada K: Ubiquitin carboxy-terminal hydrolase L1 binds to and stabilizes monoubiquitin in neu-ron. Hum Mol Genet 2003, 12:1945–1958
42. Vigouroux S, Briand M, Briand Y: Linkage between the proteasomepathway and neurodegenerative diseases and aging. Mol Neurobiol2004, 30:201–221
43. Lansbury PT Jr: Improving synaptic function in a mouse model of AD.Cell 2006, 126:655–657
44. Wang YL, Liu W, Sun YJ, Kwon J, Setsuie R, Osaka H, Noda M, AokiS, Yoshikawa Y, Wada K: Overexpression of ubiquitin carboxyl-ter-minal hydrolase L1 arrests spermatogenesis in transgenic mice. MolReprod Dev 2006, 73:40–49
45. Ardley HC, Scott GB, Rose SA, Tan NG, Robinson PA: UCH-L1aggresome formation in response to proteasome impairment indi-cates a role in inclusion formation in Parkinson’s disease. J Neuro-chem 2004, 90:379–391
46. Piotrowski J, Beal R, Hoffman L, Wilkinson KD, Cohen RE, Pickart CM:Inhibition of the 26 S proteasome by polyubiquitin chains synthesizedto have defined lengths. J Biol Chem 1997, 272:23712–23721
47. Chen Q, Liu JB, Horak KM, Zheng H, Kumarapeli AR, Li J, Li F,Gerdes AM, Wawrousek EF, Wang X: Intrasarcoplasmic amyloidosisimpairs proteolytic function of proteasomes in cardiomyocytes bycompromising substrate uptake. Circ Res 2005, 97:1018–1026
48. Liu J, Chen Q, Huang W, Horak KM, Zheng H, Mestril R, Wang X:
Impairment of the ubiquitin-proteasome system in desminopathymouse hearts. FASEB J 2006, 20:362–364
UCH-L1 in Membranous Nephropathy 2057AJP May 2011, Vol. 178, No. 5
49. Hartleben B, Godel M, Meyer-Schwesinger C, Liu S, Ulrich T, KoblerS, Wiech T, Grahammer F, Arnold SJ, Lindenmeyer MT, Cohen CD,Pavenstadt H, Kerjaschki D, Mizushima N, Shaw AS, Walz G, HuberTB: Autophagy influences glomerular disease susceptibility andmaintains podocyte homeostasis in aging mice. J Clin Invest 2010,120:1084–1096
50. Maisonneuve P, Agodoa L, Gellert R, Stewart JH, Buccianti G, LowenfelsAB, Wolfe RA, Jones E, Disney AP, Briggs D, McCredie M, Boyle P: Distri-bution of primary renal diseases leading to end-stage renal failure in theUnited States, Europe, and Australia/New Zealand: results from an interna-tional comparative study Am J Kidney Dis 2000, 35:157–165
51. McNaught KS, Bjorklund LM, Belizaire R, Isacson O, Jenner P,Olanow CW: Proteasome inhibition causes nigral degeneration withinclusion bodies in rats. Neuroreport 2002, 13:1437–1441
52. McNaught KS, Olanow CW: Protein aggregation in the pathogenesis offamilial and sporadic Parkinson’s disease. Neurobiol Aging 2006, 27:530–
54553. Apostol BL, Kazantsev A, Raffioni S, Illes K, Pallos J, Bodai L, Slepko N,Bear JE, Gertler FB, Hersch S, Housman DE, Marsh JL, Thompson LM:A cell-based assay for aggregation inhibitors as therapeutics of polyglu-tamine-repeat disease and validation in Drosophila. Proc Natl Acad SciUSA 2003, 100:5950–5955
54. Beck LH Jr, Bonegio RG, Lambeau G, Beck DM, Powell DW, Cum-mins TD, Klein JB, Salant DJ: M-type phospholipase A2 receptor astarget antigen in idiopathic membranous nephropathy. N Engl J Med2009, 361:11–21
55. Kerjaschki D, Neale TJ: Molecular mechanisms of glomerular injury inrat experimental membranous nephropathy (Heymann nephritis).J Am Soc Nephrol 1996, 7:2518–2526
56. Kerjaschki D, Schulze M, Binder S, Kain R, Ojha PP, Susani M, HorvatR, Baker PJ, Couser WG: Transcellular transport and membraneinsertion of the C5b-9 membrane attack complex of complement byglomerular epithelial cells in experimental membranous nephropathy.
J Immunol 1989, 143:546–552Copyright © 2022 FDOKUMEN




















