Activation of nuclear factor-κB by podocytes in the autologous phase of passive Heymann nephritis
9
Kidney International, Vol. 59 (2001), pp. 923–931 Activation of nuclear factor-kB by podocytes in the autologous phase of passive Heymann nephritis STUART J. MUDGE,KATHY PAIZIS,RUSSELL B. AUWARDT,RACHEL J. THOMAS, and DAVID A. POWER Department of Nephrology, St. Vincent’s Hospital, and St. Vincent’s Institute of Medical Research, Melbourne; and Department of Nephrology, Austin and Repatriation Medical Center, Heidelberg, Victoria, Australia the podocyte have not yet been determined, but may include Activation of nuclear factor-kB by podocytes in the autologous MMP-9. phase of passive Heymann nephritis. Background. The present study examined whether activa- tion of nuclear factor-kB (NF-kB) occurs within podocytes in passive Heymann nephritis (PHN) and contributes to the Human membranous glomerulonephritis (MGN) is a pathogenesis of proteinuria. common cause of nephrotic syndrome. Many insights Methods. Electrophoretic mobility shift assays (EMSAs) into the pathogenesis of MGN have been obtained from were used to detect NF-kB activation, and supershift assays were used to determine the subunits involved. Localization of the passive Heymann nephritis (PHN) model in the rat, the activated NF-kB subunit p50 was performed by immunohis- which is induced with an antiserum raised against a rat tochemistry. Expression of the NF-kB–dependent genes in- kidney proximal tubular extract [1]. Within minutes of its terleukin-1b (IL-1b) and matrix metalloproteinase-9 (MMP-9) administration, the heterologous antibody is deposited were determined by reverse transcription-polymerase chain in the glomeruli. After three to five days, subepithelial reaction and, for IL-1b, immunohistochemistry. To inhibit acti- vation of NF-kB in vivo, pyrrolidone dithiocarbamate (PDTC) electron-dense deposits are seen, comparable to those was administered for 10 days following induction of PHN. observed in early human MGN. Complement compo- Results. Glomerular nuclear extracts from rats with PHN nents are found in the immune deposits, and there is showed increased NF-kB binding activity in comparison to activation of terminal complement components (C5b-9) normal rats. The major Rel/NF-kB proteins in these activated [reviewed in 2]. The mechanism of the resultant protein- complexes were p65 and p50. Immunohistochemistry showed that nuclear translocation of p50 occurred predominantly uria is not fully understood. Proteinuria in both the heter- within podocytes. IL-1b mRNA was increased in the PHN rats, ologous and autologous phases is complement depen- and increased IL-1b protein was localized predominantly to dent and leukocyte independent. Experimental evidence podocytes by immunohistochemistry. To investigate whether suggests that fixation of terminal complement activates activation of NF-kB is involved in the pathogenesis of protein- podocytes. The activated podocytes then produce a uria, PDTC was administered to rats with PHN. Electropho- range of reactive molecules, including hydroxyl radicals, retic mobility shift assays of glomerular nuclear extracts showed a significant reduction in NF-kB binding activity in the PDTC- cytokines, prostaglandins, and proteases [3–9]. The re- treated rats with a striking reduction in MMP-9 mRNA. Com- lease of these molecules is thought to result in damage pared with control rats, there was a significant reduction in to the glomerular filtration barrier through direct injury albuminuria at days 15 (P , 0.001) and 20 (P , 0.001) when to the glomerular basement membrane or to the podo- PHN was induced with a suboptimal dose of anti-Fx1A antise- cytes themselves [10–12]. Resolution of the glomerular rum. There was no detectable difference in the systemic im- mune response to sheep Ig in the treated rats. lesion may then be hampered by the inability of fully Conclusions. These data show that NF-kB is activated within differentiated podocytes to undergo mitogenesis [13]. podocytes in PHN and suggest that it contributes to autologous A number of strategies have been employed to inter- phase proteinuria. The critical genes regulated by NF-kB in fere with the development of PHN. Reducing podocyte injury by inhibiting formation of C5b-9(m) has proven quite effective. Depletion of complement activity, how- Key words: proteinuria, MM-9, human membranous glomerulonephri- tis, glomerular lesion, inflammation, transcription factor. ever, may not be an ideal solution in patients [5, 7, 12]. Several studies have focused on the role of mediators Received for publication December 7, 1999 such as reactive oxygen species (ROS). The administra- and in revised form September 5, 2000 Accepted for publication September 14, 2000 tion of ROS scavengers and inhibition of lipid peroxida- tion have proven quite effective in reducing proteinuria 2001 by the International Society of Nephrology 923
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Activation of nuclear factor-κB by podocytes in the autologous phase of passive Heymann nephritis

Kidney International, Vol. 59 (2001), pp. 923–931
Activation of nuclear factor-kB by podocytes in theautologous phase of passive Heymann nephritis
STUART J. MUDGE, KATHY PAIZIS, RUSSELL B. AUWARDT, RACHEL J. THOMAS, andDAVID A. POWER
Department of Nephrology, St. Vincent’s Hospital, and St. Vincent’s Institute of Medical Research, Melbourne; andDepartment of Nephrology, Austin and Repatriation Medical Center, Heidelberg, Victoria, Australia
the podocyte have not yet been determined, but may includeActivation of nuclear factor-kB by podocytes in the autologousMMP-9.phase of passive Heymann nephritis.
Background. The present study examined whether activa-tion of nuclear factor-kB (NF-kB) occurs within podocytesin passive Heymann nephritis (PHN) and contributes to the Human membranous glomerulonephritis (MGN) is apathogenesis of proteinuria.
common cause of nephrotic syndrome. Many insightsMethods. Electrophoretic mobility shift assays (EMSAs)into the pathogenesis of MGN have been obtained fromwere used to detect NF-kB activation, and supershift assays
were used to determine the subunits involved. Localization of the passive Heymann nephritis (PHN) model in the rat,the activated NF-kB subunit p50 was performed by immunohis- which is induced with an antiserum raised against a rattochemistry. Expression of the NF-kB–dependent genes in- kidney proximal tubular extract [1]. Within minutes of itsterleukin-1b (IL-1b) and matrix metalloproteinase-9 (MMP-9)
administration, the heterologous antibody is depositedwere determined by reverse transcription-polymerase chainin the glomeruli. After three to five days, subepithelialreaction and, for IL-1b, immunohistochemistry. To inhibit acti-
vation of NF-kB in vivo, pyrrolidone dithiocarbamate (PDTC) electron-dense deposits are seen, comparable to thosewas administered for 10 days following induction of PHN. observed in early human MGN. Complement compo-
Results. Glomerular nuclear extracts from rats with PHN nents are found in the immune deposits, and there isshowed increased NF-kB binding activity in comparison to
activation of terminal complement components (C5b-9)normal rats. The major Rel/NF-kB proteins in these activated[reviewed in 2]. The mechanism of the resultant protein-complexes were p65 and p50. Immunohistochemistry showed
that nuclear translocation of p50 occurred predominantly uria is not fully understood. Proteinuria in both the heter-within podocytes. IL-1b mRNA was increased in the PHN rats, ologous and autologous phases is complement depen-and increased IL-1b protein was localized predominantly to dent and leukocyte independent. Experimental evidencepodocytes by immunohistochemistry. To investigate whether suggests that fixation of terminal complement activatesactivation of NF-kB is involved in the pathogenesis of protein-
podocytes. The activated podocytes then produce auria, PDTC was administered to rats with PHN. Electropho-range of reactive molecules, including hydroxyl radicals,retic mobility shift assays of glomerular nuclear extracts showed
a significant reduction in NF-kB binding activity in the PDTC- cytokines, prostaglandins, and proteases [3–9]. The re-treated rats with a striking reduction in MMP-9 mRNA. Com- lease of these molecules is thought to result in damagepared with control rats, there was a significant reduction in to the glomerular filtration barrier through direct injuryalbuminuria at days 15 (P , 0.001) and 20 (P , 0.001) when
to the glomerular basement membrane or to the podo-PHN was induced with a suboptimal dose of anti-Fx1A antise-cytes themselves [10–12]. Resolution of the glomerularrum. There was no detectable difference in the systemic im-
mune response to sheep Ig in the treated rats. lesion may then be hampered by the inability of fullyConclusions. These data show that NF-kB is activated within differentiated podocytes to undergo mitogenesis [13].
podocytes in PHN and suggest that it contributes to autologous A number of strategies have been employed to inter-phase proteinuria. The critical genes regulated by NF-kB in
fere with the development of PHN. Reducing podocyteinjury by inhibiting formation of C5b-9(m) has provenquite effective. Depletion of complement activity, how-Key words: proteinuria, MM-9, human membranous glomerulonephri-
tis, glomerular lesion, inflammation, transcription factor. ever, may not be an ideal solution in patients [5, 7, 12].Several studies have focused on the role of mediators
Received for publication December 7, 1999such as reactive oxygen species (ROS). The administra-and in revised form September 5, 2000
Accepted for publication September 14, 2000 tion of ROS scavengers and inhibition of lipid peroxida-tion have proven quite effective in reducing proteinuria 2001 by the International Society of Nephrology
923

Mudge et al: NF-kB in PHN924
in PHN [3, 4]. An alternative therapeutic strategy, how- washed with cold PBS, and nuclear extracts were pre-pared from approximately one third of the glomeruliever, might be to reduce transcription factor activation
in podocytes and, subsequently, the release of these me- according to the procedure of Schreiber et al [25]. Briefly,glomeruli were suspended in 400 mL of cold buffer Adiators.
Activation of the transcription factor nuclear factor-kB [10 mmol/L HEPES, pH 7.9, 10 mmol/L KCl, 0.1 mmol/Lethylenediaminetetraacetic acid (EDTA), 0.1 mmol/L(NF-kB) is central to the inflammatory process. Activa-
tion of NF-kB in response to complement attack has egtazic acid (EGTA), 1 mmol/L dithiothreitol (DTT), and0.5 mmol/L phenylmethylsulfonyl fluoride (PMSF)] andbeen demonstrated in vitro in human umbilical vein en-
dothelial cells [14], and many of the molecules are up- homogenized with a stab Pro200 homogenizer (Pro-Science, Melbourne, Australia). Following 15 minutes onregulated in human and experimental GN [for example,
interleukin-1b (IL-1b), tumor necrosis factor-a (TNF-a), ice, 25 mL of 10% Nonidet P-40 were added, and thehomogenates were vortexed vigorously for 10 seconds.monocyte chemoattractant protein-1 (MCP-1), intercel-
lular adhesion molecule-1 (ICAM-1), E-selectin, and ma- The nuclear fraction was precipitated at 15,000 3 g forfive minutes at 48C, resuspended in 100 mL of cold Buffertrix metalloproteinase-9 (MMP-9)] are transcriptionally
regulated by NF-kB [15–22]. Furthermore, steroid ther- B (20 mmol/L HEPES, pH 7.9, 0.4 mol/L NaCl, 1 mmol/LEDTA, 1 mmol/L EGTA, 1 mmol/L DTT, and 1 mmol/Lapy, commonly used for patients with GN, is believed
to interfere with activation of NF-kB [23]. PMSF), and the mixture was left on ice for 15 minuteswith constant agitation. Nuclear debris was precipitatedIn this study, we postulated that activation of NF-kB
in podocytes might contribute to the development of by centrifugation at 15,000 3 g for five minutes at 48C,and the nuclear extract was stored at 2708C. Proteinproteinuria in PHN and addressed three questions: (1) Is
NF-kB activated in podocytes in PHN? (2) Does NF-kB concentrations were measured using the Bradford method(Bio-Rad Protein Assay kit; Bio-Rad Lab, Hercules, CA,activation in podocytes cause up-regulation of NF-kB–
dependent genes? (3) Is the development of proteinuria USA).in PHN dependent on NF-kB activation within podo-
Electrophoretic mobility shift assaycytes?Nuclear extracts were assayed for NF-kB binding ac-
tivity using [g-32P] dATP (Amersham, Little Chalfont,METHODS
Buckinghamshire, UK) end-labeled NF-kB site consen-Induction of PHN sus oligonucleotide (Promega, Madison, WI, USA).
Eight micrograms of nuclear extract were added to bind-Passive Heymann nephritis was induced with a sheepanti-Fx1A antiserum prepared as previously described ing buffer [2 mmol/L EDTA, 20 mmol/L Tris-HCl, pH 8,
10 mmol/L MgCl2, 2 mmol/L DTT, 2 mg/mL bovine[24]. Ten or 20 mg of anti-Fx1A were administered tooutbred male Sprague-Dawley rats (180 to 220 g body serum albumin (BSA), 1 mg poly(dI)-poly(dC) (Phar-
macia Biotech, Uppsala, Sweden); 5% glycerol] and in-weight) by intravenous injection on two consecutivedays. These doses were chosen to produce an optimal cubated at room temperature for 15 minutes; 150,000
cpm of the labeled NF-kB oligonucleotide were added,(20 mg) or suboptimal (10 mg) model of PHN. Therationale for the use of a suboptimal model was to detect and the binding reaction continued for another 15 min-
utes at room temperature. Supershift electrophoreticeffects of an inhibitor of NF-kB that might not be seenin the optimal model in which other transcriptional acti- mobility shift assay (EMSA) was performed by the addi-
tion of 1 mL antiserum against the Rel/NF-kB familyvators could have increased importance. Control animalswere given the same volume of phosphate-buffered sa- proteins (p65, p50, p52, RelB, and c-Rel), or the inhibi-
tory kB protein IkB-a (Santa Cruz Biotechnology, Santaline (PBS). Experiments were approved by the St. Vin-cent’s Hospital Animal Ethics Committee, using guide- Cruz, CA, USA) for 15 minutes prior to incubation with
the labeled oligonucleotide. Half of the binding reactionlines from the National Health and Medical ResearchCommittee. Urine was collected in metabolic cages, and was loaded on a 7% polyacrylamide gel (Tris, borate,
EDTA) and separated in 1 3 TBE buffer at 30 mA/urinary albumin excretion was determined using rat al-bumin radial immunodiffusion plates (The Binding Site, 150 V. Gels were dried, and complexes were localized
by autoradiography. To confirm the specificity of theBirmingham, UK).complexes, unlabeled NF-kB consensus oligonucleotide
Preparation of glomerular nuclear extracts (Santa Cruz) at .50-fold excess was added prior to thelabeled probe.Animals were sacrificed under general anesthesia at
days 5, 10, and 20 following induction of the model, andReverse transcription-polymerase chain reactionthe kidneys were harvested. One and one half kidneys
from each rat were pooled, and the glomeruli were iso- RNA was extracted from the remaining glomeruli(as described above) using Trizol (GIBCO BRL Lifelated by a fractional sieving technique. Glomeruli were

Mudge et al: NF-kB in PHN 925
Technologies, Grand Island, NY, USA) according to the The specificity of the p50 staining was confirmed usinga no primary antibody control, and a nonimmune rabbitmanufacturer’s instructions. cDNA synthesis and poly-
merase chain reaction (PCR) were performed as pre- Ig irrelevant control.Wilm’s tumor protein (WT-1) was detected with aviously described [26]. Custom oligonucleotides were
designed to detect and amplify rat IL-1b and MMP-9 rabbit polyclonal anti–WT-1 (C19; Santa Cruz Biotech-nology). Sections were microwaved for 25 minutes inmRNA. Primers for IL-1b were as follows: forward
primer, IL-1 b 59-CTGCAGCTGGAGAGTGTGG-39; citrate buffer (pH 6), and endogenous peroxidase wasquenched in 3% hydrogen peroxide/methanol for 10reverse primer, 59-CATCCCATACACACGGACAAC
TAG-39. Primers for MMP-9 were as follows: forward minutes. The primary antibody was then detected withthe LSAB 2 peroxidase kit for rat sections (Dako) ac-primer, 59-CGCCAACTATGACCAGGATA-39; re-cording to the manufacturer’s instructions. The specific-verse primer, 59-TCACACGCCAGAAGTATTTGT-39ity of the WT-1 staining was confirmed using a no pri-[27]. The specificity of the products was confirmed bymary antibody control and a nonimmune rabbit IgSouthern blot detection using a 32P-labeled internal oli-irrelevant control.gonucleotide probe (IL-1b,59-TGAGTCTGCACAGT
Sheep Ig was detected in glomeruli using a biotin-TCCCCAAC-39; MMP-9, 59-GTTGCCCCCAGTTACconjugated, affinity-purified anti-sheep Ig (Rockland,AGT-39). Reverse transcription-PCR (RT-PCR) andGilbertsville, PA, USA) diluted 1:100, followed by aSouthern blot of glyceraldehyde phosphate dehydroge-streptavidin-conjugated horseradish peroxidase (Dako)nase mRNA was performed as previously described us-1:200.ing primers GAPDH-1, GAPDH-4, and GAPDH-5 [26].
Administration of pyrrolidone dithiocarbamateImmunohistochemical stainingRats were divided into four groups of four to five rats.Tissue blocks were immersion fixed in 4% paraformal-
In groups 1 and 2, PHN was induced with 10 mg anti-dehyde and embedded in paraffin. Serial 4 mm paraffinFx1A, and in groups 3 and 4 with 20 mg anti-Fx1A.sections were cut on a microtome, dried overnight atGroups 2 and 4 were given oral pyrrolidone dithiocarba-378C, and dewaxed. Sections were blocked for one hourmate (PDTC; Sigma) at 200 mg/kg/day from day 1at room temperature with 10% normal swine serumthrough to day 10 [20]. Groups 1 and 3 received the(NSS). All sections were developed with DAB substratesame volume of water.(Dako Corporation, Carpinteria, CA, USA) and coun-
terstained with hematoxylin.Rat anti-sheep Ig titersInterleukin-1b was detected using a four-layer immu-
noperoxidase technique. A mouse anti-rat IL-1b anti- Titers of rat anti-sheep Ig antibodies were measuredbody (Serotech, Oxford, UK) diluted 1:100 was applied by enzyme-linked immunosorbent assay (ELISA) on se-overnight at 48C, followed by a second layer of rabbit rum collected at days 10 and 20 of the PDTC study.anti-mouse Ig (Dako) diluted 1:250 with 1% NSS and 5% Polystyrene microtiter plates (Dynex Technologies, Inc.,normal rat serum for 30 minutes at room temperature. Chantilly, VA, USA) were coated with 100 mL of 10
mg/mL normal sheep globulin in carbonate/bicarbonateAt this point, endogenous peroxidase was quenched bytaking the slides through graded ethanols (70, 90, and buffer (pH 9.6) by overnight incubation at 48C. After
washing with 0.1% Tween 20 in PBS, plates were blocked2 3 100%) to 2% hydrogen peroxide in methanol for10 minutes and then reversing through the ethanols to with 200 mL of 1% BSA for 30 minutes at room tempera-
ture. The blocking agent was then removed, and serialPBS. A third-layer swine anti-rabbit Ig 1:50 (Dako) andthen a fourth-layer rabbit PAP 1:60 (Dako) were applied, dilutions of rat serum were added for one hour at 378C.
Bound rat Ig was detected with rabbit–anti-rat Ig (1:2000;both diluted with 1% NSS and 5% normal rat serumand incubated for 30 minutes at room temperature. The Dako) for 30 minutes at room temperature, followed
by goat anti-rabbit Ig horseradish peroxidase conjugatespecificity of the IL-1b staining was confirmed using ano primary antibody control and an isotyped-matched (1:1000) for a further 30 minutes at room temperature.
Fifty microliters of substrate, comprising an o-phenylene-irrelevant mouse antibody control.The Rel/NF-kB protein p50 was detected with a rabbit diamine tablet (Sigma) dissolved in 10 mL citrate phos-
phate buffer with 50 mL 30% H2O2, was incubated foranti-p50 nuclear localization signal (Santa Cruz) anti-body. Endogenous peroxidase was reduced with 3% hy- 15 minutes at room temperature. The reaction was then
stopped with 50 mL of 4 mol/L H2SO4, and the absorban-drogen peroxide/methanol for 10 minutes. The rabbitanti-p50 antiserum was diluted 1:20 and applied over- ces at 492 and 630 nm were read on a microplate reader
(Behring, Marburg, Germany). Titers were compared bynight at 48C, followed by a second layer swine anti-rabbitIg (Dako) 1:50 and a third-layer rabbit PAP (Dako) averaging the absorbances of each group’s sera relative
to the absorbance of normal rat serum diluted 1 in 64.1:100, both applied for 30 minutes at room temperature.
Mudge et al: NF-kB in PHN926
Statistical methods
The INSTAT Version 2.01 (GraphPad Software) pro-gram was used for all statistical calculations. Albumin-uria in PHN and PDTC-treated PHN rats were initiallyexamined by one-way analysis of variance (ANOVA),followed by the Bonferroni multiple comparisons test.
RESULTS
Podocyte activation of NF-kB in PHN
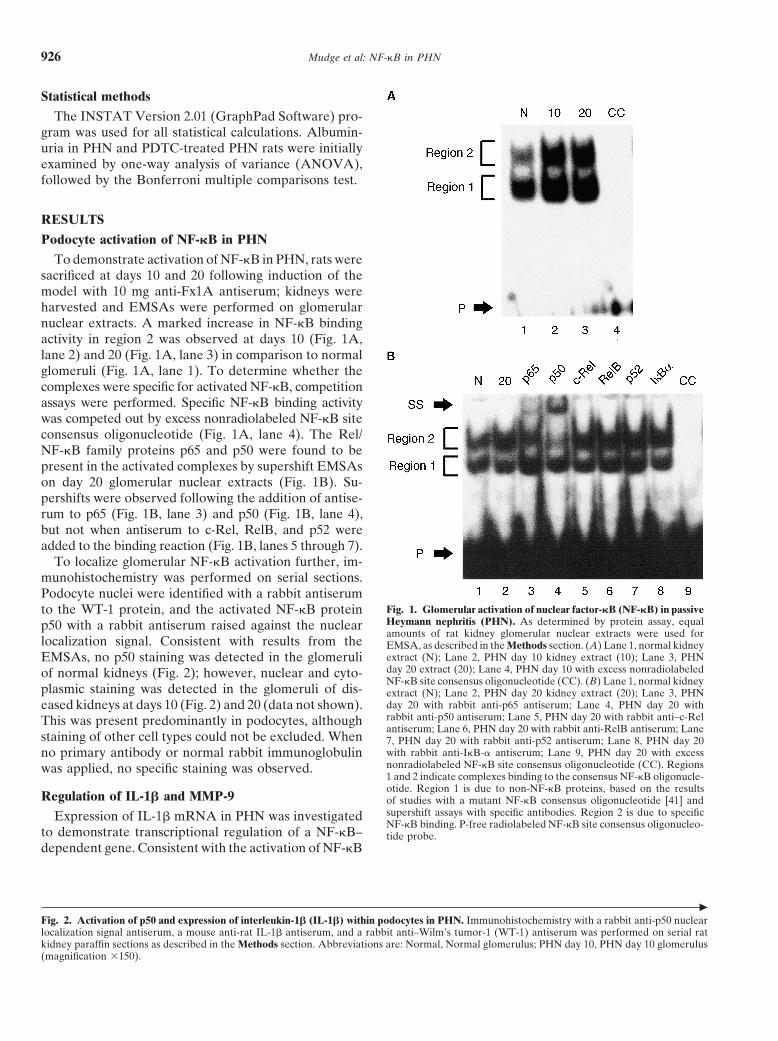
To demonstrate activation of NF-kB in PHN, rats weresacrificed at days 10 and 20 following induction of themodel with 10 mg anti-Fx1A antiserum; kidneys wereharvested and EMSAs were performed on glomerularnuclear extracts. A marked increase in NF-kB bindingactivity in region 2 was observed at days 10 (Fig. 1A,lane 2) and 20 (Fig. 1A, lane 3) in comparison to normalglomeruli (Fig. 1A, lane 1). To determine whether thecomplexes were specific for activated NF-kB, competitionassays were performed. Specific NF-kB binding activitywas competed out by excess nonradiolabeled NF-kB siteconsensus oligonucleotide (Fig. 1A, lane 4). The Rel/NF-kB family proteins p65 and p50 were found to bepresent in the activated complexes by supershift EMSAson day 20 glomerular nuclear extracts (Fig. 1B). Su-pershifts were observed following the addition of antise-rum to p65 (Fig. 1B, lane 3) and p50 (Fig. 1B, lane 4),but not when antiserum to c-Rel, RelB, and p52 wereadded to the binding reaction (Fig. 1B, lanes 5 through 7).
To localize glomerular NF-kB activation further, im-munohistochemistry was performed on serial sections.Podocyte nuclei were identified with a rabbit antiserum
Fig. 1. Glomerular activation of nuclear factor-kB (NF-kB) in passiveto the WT-1 protein, and the activated NF-kB proteinHeymann nephritis (PHN). As determined by protein assay, equalp50 with a rabbit antiserum raised against the nuclearamounts of rat kidney glomerular nuclear extracts were used for
localization signal. Consistent with results from the EMSA, as described in the Methods section. (A) Lane 1, normal kidneyextract (N); Lane 2, PHN day 10 kidney extract (10); Lane 3, PHNEMSAs, no p50 staining was detected in the glomeruliday 20 extract (20); Lane 4, PHN day 10 with excess nonradiolabeledof normal kidneys (Fig. 2); however, nuclear and cyto-NF-kB site consensus oligonucleotide (CC). (B) Lane 1, normal kidney
plasmic staining was detected in the glomeruli of dis- extract (N); Lane 2, PHN day 20 kidney extract (20); Lane 3, PHNday 20 with rabbit anti-p65 antiserum; Lane 4, PHN day 20 witheased kidneys at days 10 (Fig. 2) and 20 (data not shown).rabbit anti-p50 antiserum; Lane 5, PHN day 20 with rabbit anti–c-RelThis was present predominantly in podocytes, althoughantiserum; Lane 6, PHN day 20 with rabbit anti-RelB antiserum; Lane
staining of other cell types could not be excluded. When 7, PHN day 20 with rabbit anti-p52 antiserum; Lane 8, PHN day 20with rabbit anti-IkB-a antiserum; Lane 9, PHN day 20 with excessno primary antibody or normal rabbit immunoglobulinnonradiolabeled NF-kB site consensus oligonucleotide (CC). Regionswas applied, no specific staining was observed.1 and 2 indicate complexes binding to the consensus NF-kB oligonucle-otide. Region 1 is due to non-NF-kB proteins, based on the results
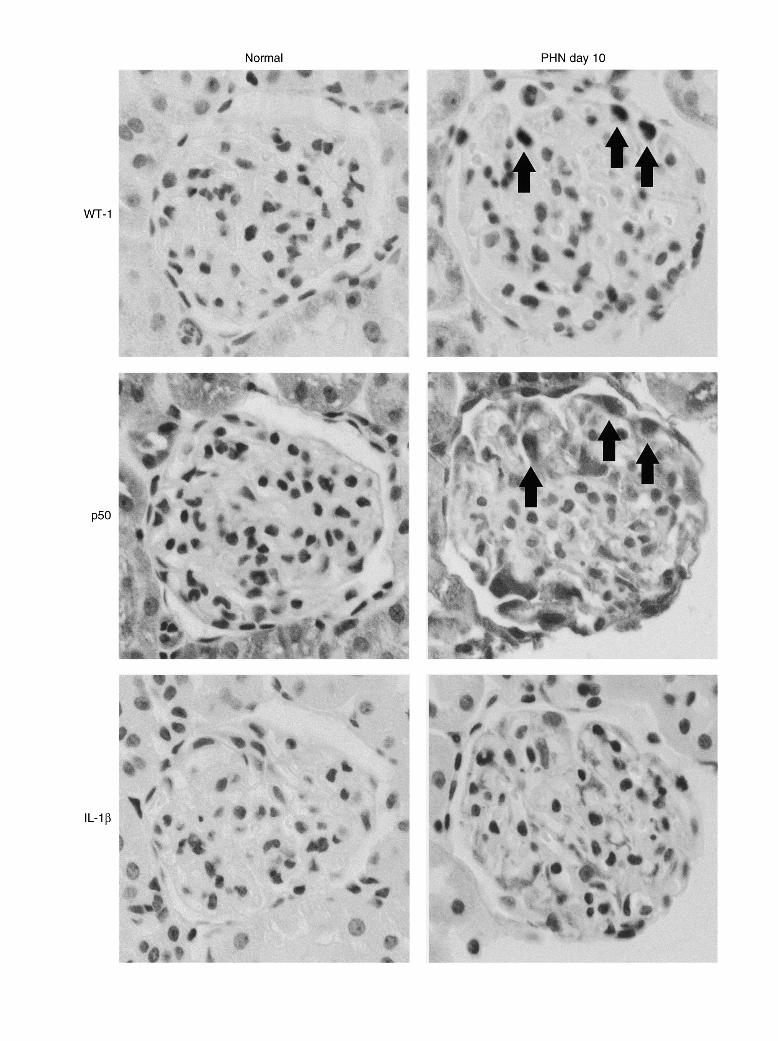
Regulation of IL-1b and MMP-9 of studies with a mutant NF-kB consensus oligonucleotide [41] andsupershift assays with specific antibodies. Region 2 is due to specificExpression of IL-1b mRNA in PHN was investigatedNF-kB binding. P-free radiolabeled NF-kB site consensus oligonucleo-
to demonstrate transcriptional regulation of a NF-kB– tide probe.dependent gene. Consistent with the activation of NF-kB
c
Fig. 2. Activation of p50 and expression of interleukin-1b (IL-1b) within podocytes in PHN. Immunohistochemistry with a rabbit anti-p50 nuclearlocalization signal antiserum, a mouse anti-rat IL-1b antiserum, and a rabbit anti–Wilm’s tumor-1 (WT-1) antiserum was performed on serial ratkidney paraffin sections as described in the Methods section. Abbreviations are: Normal, Normal glomerulus; PHN day 10, PHN day 10 glomerulus(magnification 3150).
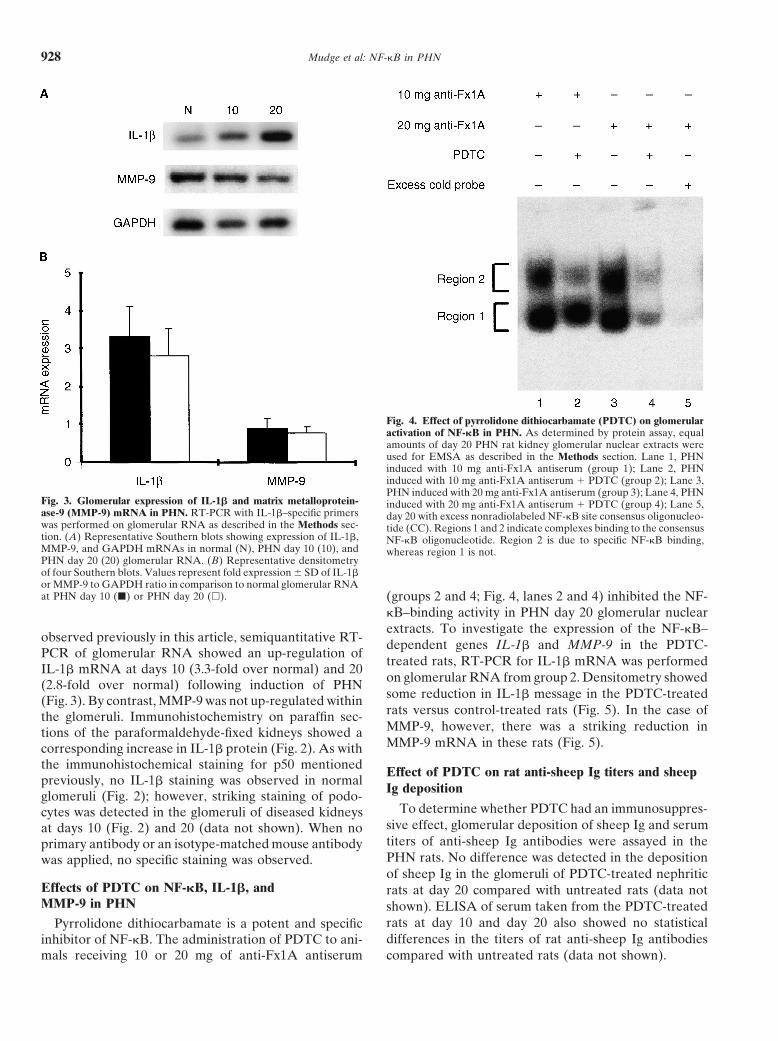
Mudge et al: NF-kB in PHN928
Fig. 4. Effect of pyrrolidone dithiocarbamate (PDTC) on glomerularactivation of NF-kB in PHN. As determined by protein assay, equalamounts of day 20 PHN rat kidney glomerular nuclear extracts wereused for EMSA as described in the Methods section. Lane 1, PHNinduced with 10 mg anti-Fx1A antiserum (group 1); Lane 2, PHNinduced with 10 mg anti-Fx1A antiserum 1 PDTC (group 2); Lane 3,PHN induced with 20 mg anti-Fx1A antiserum (group 3); Lane 4, PHN
Fig. 3. Glomerular expression of IL-1b and matrix metalloprotein- induced with 20 mg anti-Fx1A antiserum 1 PDTC (group 4); Lane 5,ase-9 (MMP-9) mRNA in PHN. RT-PCR with IL-1b–specific primers day 20 with excess nonradiolabeled NF-kB site consensus oligonucleo-was performed on glomerular RNA as described in the Methods sec- tide (CC). Regions 1 and 2 indicate complexes binding to the consensustion. (A) Representative Southern blots showing expression of IL-1b, NF-kB oligonucleotide. Region 2 is due to specific NF-kB binding,MMP-9, and GAPDH mRNAs in normal (N), PHN day 10 (10), and whereas region 1 is not.PHN day 20 (20) glomerular RNA. (B) Representative densitometryof four Southern blots. Values represent fold expression 6 SD of IL-1bor MMP-9 to GAPDH ratio in comparison to normal glomerular RNAat PHN day 10 (j) or PHN day 20 (h). (groups 2 and 4; Fig. 4, lanes 2 and 4) inhibited the NF-
kB–binding activity in PHN day 20 glomerular nuclearextracts. To investigate the expression of the NF-kB–
observed previously in this article, semiquantitative RT- dependent genes IL-1b and MMP-9 in the PDTC-PCR of glomerular RNA showed an up-regulation of
treated rats, RT-PCR for IL-1b mRNA was performedIL-1b mRNA at days 10 (3.3-fold over normal) and 20
on glomerular RNA from group 2. Densitometry showed(2.8-fold over normal) following induction of PHNsome reduction in IL-1b message in the PDTC-treated(Fig. 3). By contrast, MMP-9 was not up-regulated withinrats versus control-treated rats (Fig. 5). In the case ofthe glomeruli. Immunohistochemistry on paraffin sec-MMP-9, however, there was a striking reduction intions of the paraformaldehyde-fixed kidneys showed aMMP-9 mRNA in these rats (Fig. 5).corresponding increase in IL-1b protein (Fig. 2). As with
the immunohistochemical staining for p50 mentioned Effect of PDTC on rat anti-sheep Ig titers and sheeppreviously, no IL-1b staining was observed in normal
Ig depositionglomeruli (Fig. 2); however, striking staining of podo-
To determine whether PDTC had an immunosuppres-cytes was detected in the glomeruli of diseased kidneyssive effect, glomerular deposition of sheep Ig and serumat days 10 (Fig. 2) and 20 (data not shown). When notiters of anti-sheep Ig antibodies were assayed in theprimary antibody or an isotype-matched mouse antibodyPHN rats. No difference was detected in the depositionwas applied, no specific staining was observed.of sheep Ig in the glomeruli of PDTC-treated nephritic
Effects of PDTC on NF-kB, IL-1b, and rats at day 20 compared with untreated rats (data notMMP-9 in PHN shown). ELISA of serum taken from the PDTC-treated
rats at day 10 and day 20 also showed no statisticalPyrrolidone dithiocarbamate is a potent and specificdifferences in the titers of rat anti-sheep Ig antibodiesinhibitor of NF-kB. The administration of PDTC to ani-
mals receiving 10 or 20 mg of anti-Fx1A antiserum compared with untreated rats (data not shown).
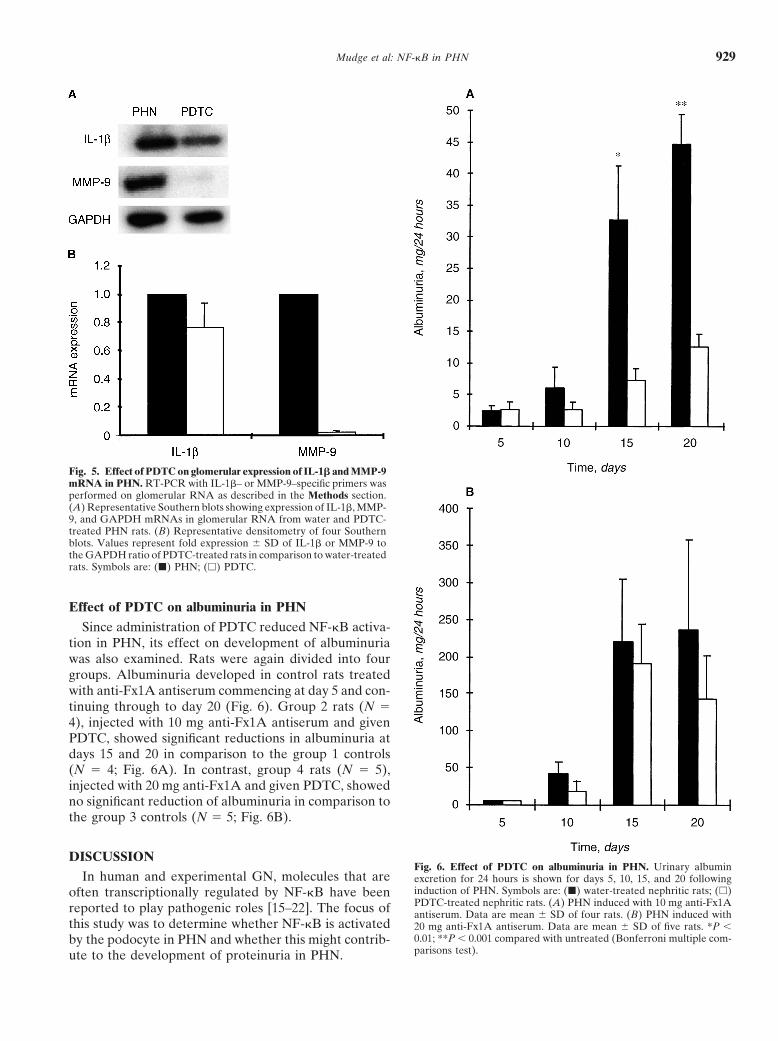
Mudge et al: NF-kB in PHN 929
Fig. 5. Effect of PDTC on glomerular expression of IL-1b and MMP-9mRNA in PHN. RT-PCR with IL-1b– or MMP-9–specific primers wasperformed on glomerular RNA as described in the Methods section.(A) Representative Southern blots showing expression of IL-1b, MMP-9, and GAPDH mRNAs in glomerular RNA from water and PDTC-treated PHN rats. (B) Representative densitometry of four Southernblots. Values represent fold expression 6 SD of IL-1b or MMP-9 tothe GAPDH ratio of PDTC-treated rats in comparison to water-treatedrats. Symbols are: (j) PHN; (h) PDTC.
Effect of PDTC on albuminuria in PHN
Since administration of PDTC reduced NF-kB activa-tion in PHN, its effect on development of albuminuriawas also examined. Rats were again divided into fourgroups. Albuminuria developed in control rats treatedwith anti-Fx1A antiserum commencing at day 5 and con-tinuing through to day 20 (Fig. 6). Group 2 rats (N 54), injected with 10 mg anti-Fx1A antiserum and givenPDTC, showed significant reductions in albuminuria atdays 15 and 20 in comparison to the group 1 controls(N 5 4; Fig. 6A). In contrast, group 4 rats (N 5 5),injected with 20 mg anti-Fx1A and given PDTC, showedno significant reduction of albuminuria in comparison tothe group 3 controls (N 5 5; Fig. 6B).
DISCUSSIONFig. 6. Effect of PDTC on albuminuria in PHN. Urinary albumin
In human and experimental GN, molecules that are excretion for 24 hours is shown for days 5, 10, 15, and 20 followinginduction of PHN. Symbols are: (j) water-treated nephritic rats; (h)often transcriptionally regulated by NF-kB have beenPDTC-treated nephritic rats. (A) PHN induced with 10 mg anti-Fx1Areported to play pathogenic roles [15–22]. The focus of antiserum. Data are mean 6 SD of four rats. (B) PHN induced with
this study was to determine whether NF-kB is activated 20 mg anti-Fx1A antiserum. Data are mean 6 SD of five rats. *P ,0.01; **P , 0.001 compared with untreated (Bonferroni multiple com-by the podocyte in PHN and whether this might contrib-parisons test).ute to the development of proteinuria in PHN.

Mudge et al: NF-kB in PHN930
To our knowledge, this study has demonstrated for Alternatively, down-regulation of constitutive levels ofNF-kB by PDTC may have nothing to do with NF-kB.the first time that NF-kB is activated in podocytes in
There is also uncertainty over the mechanism leadingPHN. EMSA of glomerular nuclear extracts showedto activation of NF-kB in podocytes. An intact comple-marked activation of NF-kB at days 10 and 20 followingment system is essential for this model so that comple-induction of the model. The nuclear localization signalment injury to podocytes might directly activate NF-kBof the Rel/NF-kB protein p50 was also detected in podo-within these cells, as has been reported in other cell typescytes, confirming that these cells were the major site of[14]. Alternatively, activation of NF-kB and the releaseNF-kB activation. The consequences of NF-kB activa-of IL-1b could occur indirectly. Complement injurytion in podocytes are potentially wide ranging. In manymight result in autocrine and juxtacrine stimulation ofcell types, NF-kB is an important proinflammatory tran-podocytes by inflammatory mediators such as ROS. Suchscription factor, regulating the expression of molecules,stimulation might then activate NF-kB within podocytes,including cytokines and proteases as well as enzymesamplifying the acute renal injury by increasing transcrip-involved in oxygen-free radical synthesis and prostaglan-tion of some of the many cytokines and chemokines thatdin synthesis [3–9, 28, 29].are regulated by NF-kB.In the present study, RT-PCR analysis of glomerular
Podocyte p50 nuclear translocation and IL-1b expres-RNA showed increased expression of IL-1b mRNA atsion at day 5 in PHN was slight in comparison to daysdays 10 and 20, and increased podocyte expression of10 and 20. Hence, it might be suggested that NF-kB isIL-1b protein was observed by immunohistochemistry.not activated directly by complement attack and perhapsIL-1b is one of the most important proinflammatoryplays a secondary role in the pathogenesis of heterolo-cytokines and has a wide range of actions, includinggous phase proteinuria. Podocyte NF-kB activationactivation of endothelial cells, release of chemokines andmight be of greater importance in driving the autologousother cytokines, induction of leukocyte adhesion, andphase proteinuria. This is also supported by the lack ofstimulation of inducible nitric oxide synthase [30]. Theany significant reduction in heterologous phase albumin-administration of IL-1 in experimental GN has beenuria in the PDTC-treated rats given 20 mg of anti-Fx1A.reported to exacerbate disease [31], and blocking the
In this study, PDTC was used as an inhibitor of NF-kBaction of IL-1 with anti–IL-1 antibodies or IL-1 receptoractivation. PDTC is reported to act as a scavenger ofantagonists has had some success in reducing acute renalfree radicals and a chelator of heavy metal ions [38].injury [32, 33]. The literature suggests that intrinsic glo-When inhibiting the activation of NF-kB, PDTC ismerular cells are a source of IL-1b in renal diseasethought to interfere with the signaling of intracellular[34, 35] and in vitro stimulation with IL-1b activateshydroxyl radicals, which precedes phosphorylation of
NF-kB in cultured glomerular cells [36]. It is not clear,IkBa [38]. However, the precise mechanism is not fully
however, what the role of IL-1b might be in PHN, a understood. There is no evidence in the literature of adisease that is not characterized by a marked leukocyte systemic effect of PDTC on the immune response, andinfiltrate. Perhaps it is simply a “by-product” of NF-kB this was not found in the present study.activation and has no role in this disease. Alternatively, Although PDTC was effective in inhibiting activationit could form an autocrine loop to increase synthesis of of NF-kB, it did not significantly reduce proteinuria inother molecules, such as chemokines and MMP-9 [19, rats with more severe PHN. One explanation for this28, 29, 37]. result is the redundancy of inflammatory pathways. For
Matrix metalloproteinase-9 has also been reported to example, the transcription of the human pro-IL-1b genebe induced through NF-kB activation. In this study, appears to be regulated via a variety of transcriptionMMP-9 mRNA was not increased in PHN, but levels factors, including activator protein-1 (AP-1), cAMP-re-were greatly decreased by treatment with PDTC. This sponsive element binding protein (CREB), NF-IL6, andunexpected result has no clear explanation. If the re- NF-kB [39]. Inhibition of NF-kB alone may be compen-duced levels of MMP-9 seen in rats treated with PDTC sated by signaling through other pathways. Meyer, Schreck,were simply due to inhibition of NF-kB, then it is odd and Baeuerle have demonstrated that although PDTCthat increased activation of NF-kB in the untreated dis- reduces NF-kB activation in HeLa cells, it has the reverseease did not result in increased MMP-9. The results sug- effect on AP-1 activity in the same cell line [40]. It isgest that NF-kB becomes important for maintenance of possible that additional transcription factors are acti-MMP-9 mRNA levels in disease. This could be due to vated within podocytes when the model is optimal, re-a shortened half-life of MMP-9 mRNA in disease so ducing the importance of NF-kB. A second potentialthat levels in this situation are maintained by increased explanation is that PDTC acts to reduce proteinuria intranscription mediated through NF-kB. The levels of PHN by a mechanism unrelated to NF-kB, such as scav-MMP-9 mRNA in normal glomeruli could be relatively enging of oxygen free radicals.
In summary, this study has demonstrated podocytelong lived and not indicative of active gene transcription.

Mudge et al: NF-kB in PHN 931
factor NF-kB in experimental glomerulonephritis in rats. Biochimactivation of NF-kB and increased IL-1b gene expressionBiophys Acta 1316:132–138, 1996
in PHN. The ability of PDTC to reduce albuminuria in 21. Morrissey JJ, Klahr S: Enalapril decreases nuclear factor kappaB activation in the kidney with ureteral obstruction. Kidney Intrats with PHN suggests that NF-kB activation within52:926–933, 1997podocytes might contribute to proteinuria in this model.
22. McMillan JI, Riordan JW, Couser WG, et al: Characterizationof a glomerular epithelial cell metalloproteinase as matrix metallo-
ACKNOWLEDGMENTS proteinase-9 with enhanced expression in a model of membranousThis study was supported by grants from the University of Mel- nephropathy. J Clin Invest 97:1094–1101, 1996
23. Scheinman RI, Cogswell PC, Lofquist AK, et al: Role of tran-bourne (S.M.) and the Australian Kidney Foundation.scriptional activation of IkB-a in mediation of immunosuppression
Reprint requests to David Power, M.D., Ph.D., Department of Ne- by glucocorticoids. Science 270:283–286, 1995phrology, Austin and Repatriation Medical Center, Studley Road, Hei- 24. Paizis K, Kirkland G, Polihronis M, et al: Heparin-binding epi-delberg 3084, Victoria, Australia. dermal growth factor-like growth factor in experimental modelsE-mail: [email protected] of membranous and minimal change nephropathy. Kidney Int
53:1162–1171, 199825. Schreiber E, Matthias P, Muller MM, et al: Rapid detection ofREFERENCES
octamer binding proteins with “mini-extracts,” prepared from a1. Wasserstein A: Membranous glomerulonephritis. J Am Soc small number of cells. Nucleic Acids Res 17:6419, 1989
Nephrol 8:664–674, 1997 26. Southby J, O’Keefe LM, Martin TJ, et al: Alternative promoter2. Kerjaschki D, Neale TJ: Molecular mechanisms of glomerular usage and mRNA splicing pathways for parathyroid hormone-
injury in rat experimental membranous nephropathy (Heymann related protein in normal tissues and tumours. Br J Cancer 72:702–nephritis). J Am Soc Nephrol 7:2518–2526, 1996 707, 1995
3. Neale TJ, Ojha PP, Exner M, et al: Proteinuria in passive Hey- 27. Martorana AM, Zheng G, Springall F, et al: Absolute quantita-mann nephritis is associated with lipid peroxidation and formation tion of specific mRNAs in cell and tissue samples by comparativeof adducts on type IV collagen. J Clin Invest 94:1577–1584, 1994 PCR. Biotechniques 27:136–144, 1999
4. Shah SV: Evidence suggesting a role for hydroxyl radical in passive 28. Bond M, Fabunmi RP, Baker AH, et al: Synergistic upregulation ofHeymann nephritis in rats. Am J Physiol 254(3 Pt 2):F337–F344, metalloproteinase-9 by growth factors and inflammatory cytokines:1988 An absolute requirement for transcription factor NF-kappa B.
5. Hara M, Batsford SR, Mihatsch MJ, et al: Complement and FEBS Lett 435:29–34, 1998monocytes are essential for provoking glomerular injury in passive 29. Kumar A, Dhawan S, Mukhopadhyay A, et al: Human immunode-Heymann nephritis in rats: Terminal complement components are ficiency virus-1-tat induces matrix metalloproteinase-9 in monocytesnot the sole mediators of proteinuria. Lab Invest 65:168–179, 1991 through protein tyrosine phosphatase-mediated activation of nuclear
6. Salant DJ, Belok S, Madaio MP, et al: A new role for complement transcription factor NF-kappaB. FEBS Lett 462:140–144, 1999in experimental membranous nephropathy in rats. J Clin Invest 30. Dinarello CA: Interleukin-1 and interleukin-1 antagonism. Blood66:1339–1350, 1980 77:1627–1652, 1991
7. Baker P, Ochi R, Schulze M, et al: Depletion of C6 prevents 31. Tomosugi NI, Cashman SJ, Hay H, et al: Modulation of antibody-development of proteinuria in experimental membranous nephrop- mediated glomerular injury in vivo by bacterial lipopolysaccharide,athy in rats. Am J Pathol 135:185–194, 1989 tumor necrosis factor, and IL-1. J Immunol 142:3083–3090, 1989
32. Karkar AM, Tam FW, Steinkasserer A, et al: Modulation of8. Stahl RA, Adler S, Baker PJ, et al: Enhanced glomerular prosta-antibody-mediated glomerular injury in vivo by IL-1ra, solubleglandin formation in experimental membranous nephropathy. Kid-IL-1 receptor, and soluble TNF receptor. Kidney Int 48:1738–1746,ney Int 31:1126–1131, 198719959. Katoh T, Lianos EA, Fukunaga M, et al: Leukotriene D4 is a
33. Lan HY, Nikolic-Paterson DJ, Mu W, et al: Interleukin-1 recep-mediator of proteinuria and glomerular hemodynamic abnormali-tor antagonist halts the progression of established crescentic glo-ties in passive Heymann nephritis. J Clin Invest 91:1507–1515, 1993merulonephritis in the rat. Kidney Int 47:1303–1309, 199510. Panesar M, Papillon J, McTavish AJ, et al: Activation of phos-
34. Tesch GH, Yang N, Yu H, et al: Intrinsic renal cells are the majorpholipase A2 by complement C5b-9 in glomerular epithelial cells.source of interleukin-1 beta synthesis in normal and diseased ratJ Immunol 159:3584–3594, 1997kidney. Nephrol Dial Transplant 12:1109–1115, 199711. Kerjaschki D: Dysfunctions of cell biological mechanisms of vis-
35. Werber HI, Emancipator SN, Tykocinski ML, et al: The interleu-ceral epithelial cell (podocytes) in glomerular diseases. Kidney Intkin 1 gene is expressed by rat glomerular mesangial cells and is45:300–313, 1994augmented in immune complex glomerulonephritis. J Immunol12. Salant D, Quigg R, Cybulsky A: Heymann nephritis: Mechanism138:3207–3212, 1987of renal injury. Kidney Int 35:976–984, 1989
36. Richardson CA, Gordon KL, Couser WG, et al: IL-1 beta in-13. Kriz W, Gretz N, Lemley KV: Progression of glomerular diseases:creases laminin B2 chain mRNA levels and activates NF-kappa BIs the podocyte the culprit? Kidney Int 54:687–697, 1998in rat glomerular epithelial cells. Am J Physiol 268:F273–F278,14. Kilgore KS, Schmid E, Shanley TP, et al: Sublytic concentrations1995of the membrane attack complex of complement induce endothe-
37. Yokoo T, Kitamura M: Dual regulation of IL-1 beta-mediatedlial interleukin-8 and monocyte chemoattractant protein-1 through matrix metalloproteinase-9 expression in mesangial cells by NF-nuclear factor-kB activation. Am J Pathol 150:2019–2031, 1997 kappa B and AP-1. Am J Physiol 270:F123–F130, 199615. Yoshioka K, Takemura T, Murakami K, et al: In situ expression 38. Schreck R, Meier B, Mannel DN, et al: Dithiocarbamates as
of cytokines in IgA nephritis. Kidney Int 44:825–833, 1993 potent inhibitors of nuclear factor kappa B activation in intact16. Rovin BH, Rumancik M, Tan L, et al: Glomerular expression of cells. J Exp Med 175:1181–1194, 1992
monocyte chemoattractant protein-1 in experimental and human 39. Shirakawa F, Saito K, Bonagura CA, et al: The human proin-glomerulonephritis. Lab Invest 71:536–542, 1994 terleukin 1 beta gene requires DNA sequences both proximal and
17. Dal Canton A: Adhesion molecules in renal disease. Kidney Int distal to the transcription start site for tissue-specific induction.48:1687–1696, 1995 Mol Cell Biol 13:1332–1344, 1993
18. Collins T, Read MA, Neish AS, et al: Transcriptional regulation 40. Meyer M, Schreck R, Baeuerle PA: H2O2 and antioxidants haveof endothelial cells adhesion molecules: NF-kB and cytokine- opposite effects on activation of NF-kappa B and AP-1 in intactinducible enhancers. FEBS Lett 9:899–909, 1995 cells: AP-1 as secondary antioxidant-responsive factor. EMBO J
19. Rovin BH, Dickerson JA, Tan LC, et al: Activation of nuclear 12:2005–2015, 1993factor-kB correlates with MCP-1 expression by human mesangial 41. Auwardt RB, Mudge SJ, Chen CG, et al: Regulation of NF-kBcells. Kidney Int 48:1263–1271, 1995 by corticosteroids in rat mesangial cells. J Am Soc Nephrol 9:1620–
20. Sakurai H, Hisada Y, Ueno M, et al: Activation of transcription 1628, 1998