
Direct characterization of target podocyte antigens and auto-antibodies in human membranous...
10
Direct characterization of target podocyte antigens and auto-antibodies in human membranous glomerulonephritis: Alfa-enolase and borderline antigens Maurizio Bruschi a, 1 , Maria Luisa Carnevali c , 1 , Corrado Murtas c , Giovanni Candiano a , Andrea Petretto b , Marco Prunotto a, 2 , Rita Gatti d , Lucia Argentiero c , Riccardo Magistroni e , Giacomo Garibotto f , Francesco Scolari g , Pietro Ravani i , Loreto Gesualdo h , Landino Allegri c , Gian Marco Ghiggeri a, ⁎ a Division of Nephrology and Laboratory on Pathophysiology of Uremia, G. Gaslini Children Hospital, Genoa, Italy b Laboratory of Experimental and Clinical Immunology-Mass Spectrometry Core Facility, G. Gaslini Children Hospital, Genoa, Italy c Department of Clinical Medicine, Nephrology and Health Sciences and University of Parma, Parma, Italy d Department of Experimental Medicine, Section of Histology, University of Parma, Parma, Italy e Chair of Nephrology, University of Modena, Italy f Chair of Nephrology, University of Genoa, Italy g Chair of Nephrology, University of Brescia and Division of Nephrology Montechiari Hospital, Italy h Chair of Nephrology, University of Foggia, Italy i Division of Nephrology, University of Calgary, Calgary, Canada ARTICLE INFO ABSTRACT Article history: Received 4 February 2011 Accepted 10 May 2011 Available online 19 May 2011 The identification of glomerular auto-antigens in idiopathic human membranous glomerulonephritis (MGN) is a crucial step towards the definition of the mechanisms of the disease. Recent ‘in vivo’ studies demonstrated a heterogeneous composition of glomerular immune-deposits in MGN biopsies only a part of which have been characterized. We studied with a proteomical approach IgGs eluted from laser capture microdissected glomeruli of 8 MGN patients and showed the existence of other three immune proteins in MGN glomeruli (α-enolase, elongation factor 2 and Glycyl Aminoacyl-tRNA Synthetase). One of these, i.e. α-enolase, fulfilled all criteria for being considered an auto-antigen. Specific IgG 1 and IgG 4 reacting with podocyte α-enolase were, in fact, eluted from microdissected glomeruli and Confocal- and Immuno Electron-Microscopy showed co- localization of α-enolase with IgG 4 and C5b-9 in immune-deposits. Serum levels of anti a- enolase IgG4 were determined in 131 MGN patients and were found elevated in 25% of cases. Overall, our data demonstrate that glomerular α-enolase is a target antigen of autoimmunity in human MGN. Circulating anti α-enolase auto-antibodies can be detected Keywords: Autoimmune glomerulonephritis Renal auto-antigen Circulating auto-antibodies Membranous glomerulonephritis α-enolase Proteomics JOURNAL OF PROTEOMICS 74 (2011) 2008 – 2017 Abbreviations: MGN, membranous glomerulonephritis; PLA2r, phospholipase A2 receptor; SOD2, superoxide dismutase 2; AR, aldose reductase; NEP, neutral endopeptidase; HN, passive Heymann nephritis. ⁎ Corresponding author at: Nephrology, Dialysis, Transplantation, Laboratory on Pathophysiology of Uremia, G. Gaslini Children Hospital, Largo G. Gaslini, 5. 16148 Genova, Italy. Tel.: + 39 010 380742; fax: + 39 010 395214. E-mail address: [email protected] (G.M. Ghiggeri). 1 Contributed equally to the work. 2 Present address: Pharma Research Development and Metabolic Disease, Hoffmann-La Roche Ltd., Basel, Switzerland. 1874-3919/$ – see front matter © 2011 Elsevier B.V. All rights reserved. doi:10.1016/j.jprot.2011.05.021 available at www.sciencedirect.com www.elsevier.com/locate/jprot
Transcript of Direct characterization of target podocyte antigens and auto-antibodies in human membranous...

J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 2 0 0 8 – 2 0 1 7
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te / j p ro t
Direct characterization of target podocyte antigens andauto-antibodies in human membranous glomerulonephritis:Alfa-enolase and borderline antigens
Maurizio Bruschia, 1, Maria Luisa Carnevalic, 1, Corrado Murtasc, Giovanni Candianoa,Andrea Petrettob, Marco Prunottoa, 2, Rita Gattid, Lucia Argentieroc, Riccardo Magistronie,Giacomo Garibotto f, Francesco Scolarig, Pietro Ravanii, Loreto Gesualdoh,Landino Allegric, Gian Marco Ghiggeria,⁎aDivision of Nephrology and Laboratory on Pathophysiology of Uremia, G. Gaslini Children Hospital, Genoa, ItalybLaboratory of Experimental and Clinical Immunology-Mass Spectrometry Core Facility, G. Gaslini Children Hospital, Genoa, ItalycDepartment of Clinical Medicine, Nephrology and Health Sciences and University of Parma, Parma, ItalydDepartment of Experimental Medicine, Section of Histology, University of Parma, Parma, ItalyeChair of Nephrology, University of Modena, ItalyfChair of Nephrology, University of Genoa, ItalygChair of Nephrology, University of Brescia and Division of Nephrology Montechiari Hospital, ItalyhChair of Nephrology, University of Foggia, ItalyiDivision of Nephrology, University of Calgary, Calgary, Canada
A R T I C L E I N F O
Abbreviations: MGN, membranous glomerreductase; NEP, neutral endopeptidase; HN, p⁎ Corresponding author at: Nephrology, Dialys
Largo G. Gaslini, 5. 16148 Genova, Italy. Tel.:E-mail address: labnefro@ospedale-gaslin
1 Contributed equally to the work.2 Present address: Pharma Research Develo
1874-3919/$ – see front matter © 2011 Elsevdoi:10.1016/j.jprot.2011.05.021
A B S T R A C T
Article history:Received 4 February 2011Accepted 10 May 2011Available online 19 May 2011
The identification of glomerular auto-antigens in idiopathic human membranousglomerulonephritis (MGN) is a crucial step towards the definition of the mechanisms ofthe disease. Recent ‘in vivo’ studies demonstrated a heterogeneous composition ofglomerular immune-deposits in MGN biopsies only a part of which have beencharacterized.We studied with a proteomical approach IgGs eluted from laser capture microdissectedglomeruli of 8 MGN patients and showed the existence of other three immune proteins inMGN glomeruli (α-enolase, elongation factor 2 and Glycyl Aminoacyl-tRNA Synthetase).One of these, i.e. α-enolase, fulfilled all criteria for being considered an auto-antigen.Specific IgG1 and IgG4 reacting with podocyte α-enolase were, in fact, eluted frommicrodissected glomeruli and Confocal- and Immuno Electron-Microscopy showed co-localization of α-enolase with IgG4 and C5b-9 in immune-deposits. Serum levels of anti a-enolase IgG4 were determined in 131 MGN patients and were found elevated in 25% ofcases.Overall, our data demonstrate that glomerular α-enolase is a target antigen ofautoimmunity in human MGN. Circulating anti α-enolase auto-antibodies can be detected
Keywords:Autoimmune glomerulonephritisRenal auto-antigenCirculating auto-antibodiesMembranous glomerulonephritisα-enolaseProteomics
ulonephritis; PLA2r, phospholipase A2 receptor; SOD2, superoxide dismutase 2; AR, aldoseassive Heymann nephritis.is, Transplantation, Laboratory on Pathophysiology of Uremia, G. Gaslini Children Hospital,+39 010 380742; fax: +39 010 395214.i.ge.it (G.M. Ghiggeri).
pment and Metabolic Disease, Hoffmann-La Roche Ltd., Basel, Switzerland.
ier B.V. All rights reserved.

Table 1 – Patient characteristics at the onas median and interquartile range due
Patients n Age
Overall 131 61 (48–Class I 29 59 (42–Class II 57 64 (50–Class III 28 67 (58–Class IV 17 47 (33–
2009J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 2 0 0 8 – 2 0 1 7
in sera of a significant quota of MGN patients. Like other auto-antigens, α-enolase may beimplicated in the pathogenesis of human MGN.
© 2011 Elsevier B.V. All rights reserved.
1. Introduction
Primary membranous glomerulonephritis (MGN) is a commoncause of nephrotic syndrome in adults. It is the prototype of anautoimmunedisease characterized by glomerular sub-epithelialimmune-deposits (mainly composed by IgG4 and C5b-9) but itspathogenesis remains unknown.Most of our current knowledgeon mechanisms for antibody entrapping are based on experi-mental models of passive Heymann nephritis (HN) that isinduced in susceptible rat strains by injection of heterologousantisera from sheep or rabbit immunizedwith a crude extractof rat tubular antigen known as Fx1A [1]. Megalin is thepodocyte antigen involved in HN; anti-megalin IgG and C5b-9are major components of immune-deposits [2–6], while thereceptor associated protein (RAP), its antibody and clusterin[7–10] are regarded as subordinate players with a somehowunclear role in the pathogenesis of HN. Some of thesecomponents have been also detected in human MGN [11,12]but megalin and/or its homologue (LDL-receptor) [13] are notpresent in human glomeruli suggesting that different podo-cyte antigens are implicated in human MGN. Although theidentification of podocyte antigens is crucial for defining thepathogenesis of human MGN, technology problems mainlyconcerning dissection and purification of glomeruli fromhuman biopsies have limited the experimental approach foryears.
Since 2002, major advances have been achieved based ontechnology developments. Debiec et al. [14,15] first showed thatneutral endopeptidase (NEP) emerges as podocyte antigen incongenital MGN due tomaternal NEP deficiency and alloimmu-nizationduringpregnancy.This isa rare formofMGNthatarisesinnewborns fromamother carrying a genetic deficiency ofNEP:the protein is expressed by the podocytes of the foetus, andNEPdeficient mothers, during a previous miscarriage, producedanti-NEP antibodies since their immune system recognized it asa non-self protein.
More recently, two independent groups [16,17] have shownspecific IgG4 against the phospholipase A2 receptor (PLA2R),aldose reductase (AR) and superoxide dismutase 2 (SOD2) in
set of clinical signs of nto their non-normal dis
Serum cre(mg/
70) 1 (0.870) 0.9 (0.770) 1.1 (0.870) 1.2 (0.958) 1 (0.8
glomerular eluates and in sera of a substantial part of patientswith MGN. These Authors could demonstrate glomerular co-localizationof IgG4with their respectiveauto-antigens (i.e. PLA2r,ARandSOD2)bydouble stainingandbyelectronmicroscopy thatsuggests an implication of all these antigens in sub-epithelialdeposit formation. These are not, however, the unique auto-antigens that induce the autoimmune response. In fact, IgG4
eluted from glomeruli of MGN patients recognize other proteinsthat represent good candidates for new auto-antigens (Ghiggeri,personal observation). Technology developments based on tissuemicrodissection and proteomical analysis have revolutionizedthe strategies for the basic approach to autoimmune glomerulardiseases in humans allowing a direct analysis of renal tissue ‘invivo’ [18].
In this paper we investigated with an integrated proteomics-pathology approach other proteins recognized by IgG4 inglomeruli. The ‘in vivo’ model consisting of renal biopsyspecimen and serum of a large cohort of patients affected byMGN was utilized in order to gain information directly fromhuman pathology.
2. Materials and methods
2.1. Patients
Overall, 131 MGN patients were included in the study (Table 1)and were utilized for studies on circulating auto-antibodies.Eight patients were also utilized for the proteomics approachto renal biopsies and IMF (Table 2). Criteria for enrollmentwere: a) a biopsy-based diagnosis of MGN; b) negative tests forserum auto-antibodies (ANA, nDNA, ANCA), for cryoglobulins,and for viral markers (HBsAg, HIV, HCV); c) absence of anyprevious immunosuppressive treatment with exception ofanti-hypertensive drugs. For histological evaluation of kidneydisease, Dubosq-Bresil solution-fixed tissues were embeddedin paraffin, sectioned and stained with hematoxylin/eosin,Masson's trichrome, silver methenamine and periodic-acidSchiff (PAS). Specimens for classic EMwere fixed in Karnovsky
ephrotic syndrome. Data for quantitative variable are giventribution.
atininedl)
Serum cholesterol(mg/dl)
Proteinuria(g/24 h)
–1.4) 262 (214–317) 5.8 (3.3–8.8)–1) 274 (206–328) 6.1 (2.1–7.3)–1.7) 261 (228–310) 6 (4–10)–1.4) 228 (174–312) 3.9 (2.6–6.6)–1.4) 307 (219–328) 6 (4.6–9)

Table2 – Patient characteristicsat theonsetof clinical signsofnephrotic syndrome. In thoseMGNpatientswho underwenta detailed analysis of glomerular tissue (i.e. tissuedissection,proteomic characterization of eluates).
Patient ID 1 2 3 4 5 6 7 8
Sex F F M M M F F FAge at diagnosis 58 68 42 77 72 46 39 75Histological stage I–II II II II I II–III I IISerum creatinine (mg/dl)At diagnosis 0.7 1.1 1.2 0.7 2.6 0.7 0.7 1.4After 6 months 0.7 1.0 1.0 0.7 1.6 0.7 0.6 1.9After 1 year 0.6 NA 1.0 0.6 NA NA NA NA
Serum albumin (g/dl)At diagnosis 2.4 2.2 1.5 2.6 3.3 3.0 2.1 1.9After 6 months 3.0 4.0 4.0 2.6 3.4 NA 3.9 3.1After 1 year 3.9 NA NA 3.7 NA NA NA NA
Proteinuria (g/day)At diagnosis 2.8 4.7 13.0 4.0 7.6 2.7 6.5 9.7After 6 months 3.6 4.4 2.7 2.2 1.8 0.7 0.1 2.8After 1 year 0.9 NA 1.4 1.9 NA NA NA NA
Immunosuppressanttherapy
Y Y Y Y Y
ACE I/ARB Y Y Y Y Y Y Yα-enolase IMF Y Y Y Y Y Y Y YGlomerular eluates Y Y Y Y Y Y Y Y
2010 J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 2 0 0 8 – 2 0 1 7
solution and processed using standard methods. Routine IMFstudies on frozen sections were performed utilizing antihuman IgG, IgA, IgM,C1q, C3 and Fibrinogen antibody.
Ten MGN patients were utilized for IMF studies on alpha-enolase glomerular expression. Glomeruli of eight patientswere microdissected with laser capture and used for eluateanalysis (see below and Table 1).
2.2. Normal kidneys, other nephropathies
Overall, eight biopsy sections from patients with other nephro-pathies (MCD n=2, MPGN n=2, IgAN n=4) and four non-carcinomatous portions of kidneys removed for renal cancerwere used as negative tissue controls. Sera obtained from 12patients with IgAN (4 F, 8 M,mean age 26 years 16–42) and from60wereutilized for theanalysisof circulating antiα-enolase IgG.
2.3. Antibodies
2.3.1. Alpha-enolase-1Rabbit anti human Non-Neuronal Enolase (NNE) (alpha-alpha)purifiedpolyclonal antibodieswerepurchased fromAbDSerotecMorphoSys Ltd, (Endeavour House, Kidlington Oxford, UK).
2.3.2. Alpha-enolase-2Anti α-enolasemonoclonal antibodies (clones 251/3 and 276/3)were a gentle gift from Prof P. Migliorini [19].
2.3.3. Aldose reductasePolyclonal goat anti human aldose reductase antibodies were agentle gift fromDr. Peter Kador. Preparation of aldose reductaseprotein and specificity testing of antibodies were performed asdescribed elsewhere [17] and is summarized in the Supplementmethods.
2.3.4. Superoxide dismutase 2Purified rabbit antiserum against human Mn-superoxide dis-mutase (SOD2) was purchased from Upstate Biotechnologies(Charlottesville, VA, U.S.A.).
2.3.5. PLA2R, Glycil-tRNA synthase, elongation factor 2Polyclonal goat anti human PLA2R, Glycil-tRNA synthase andelongation factor 2 antibodies were purchased from Abnovacorporation (Taipei, Taiwan).
2.3.6. IgG4
Mouse monoclonal antibodies against human IgG4 (Clone:HP6025, IgG1-K) were purchased from Invitrogen Corporation(Camarillo, CA).
2.3.7. C5b-9Mouse monoclonal anti human C5b-9 antibodies (clone aE11-IgG2a, kappa) were purchased fromDako (Glostrup, Denmark).
2.3.8. Secondary antibodiesAffinity-purified fluorescein isothiocyanate (FITC) F(ab′)2 donkeyanti-rabbit IgG (affinity-purified Texas Red conjugated donkeyanti-mouse IgG) were all purchased from Jackson Immunore-search (West Grove, PA, USA). For immunoelectron microscopy(IEM), donkey anti-rabbit IgG conjugated with 12 nm-colloidalgold particles (Jackson Immunoresearch) were employed.
2.4. Laser capture microdissection (LCM) and elution ofantibodies from renal biopsy tissue
Laser capture microdissection and elution of antibodieswere done as already described [17] and as reported in theSupplements.
2.5. Two-dimensional electrophoresis
Two-dimensional electrophoresis was performed in soft gelsas described [20]. A detailed description is given in theSupplement methods.
2.6. Mono-dimensional electrophoresis
Gradient polyacrylamide gel electrophoresis was done accord-ing to Laemmli [21].
2.7. Gel/membrane staining and image analysis
After separation in SDS-PAGE gels, proteins were visualized bya double staining procedure: first the methyl-trichloroacetatenegative staining [22] followed by the silver staining or Bluesilver colloidal Coomassie for preparative mass spectrometryanalysis. Images of stained gels were digitized using a GS800photometer and western blots were acquired using a VersaDOC 400. All imageswere analyzedwith the PDQuest software(Bio-Rad, Hercules, CA, USA).
2.8. Western blot
Westernblotwithglomerulareluateswasdonewithpodocytecellline whole extracts separated by either mono- or bi-dimensional

2011J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 2 0 0 8 – 2 0 1 7
electrophoresis. Equipment and technique of analysis is de-scribed in the Supplements.
2.9. Tryptic digestion and protein identification by LC–Ms
LC–ESI MS-MS/MS was utilized for characterizing single spotsderiving from two-dimensional electrophoresis. Protein spotswere excised from gel, rinse in 50% v/v acetonitrile (ACN) in5 mM ammonium bicarbonate pH 8.9 until full destaining,rinse twice in 100% v/v ACN and briefly rinse in 1 mM CaCl2and 100 mM ammonium bicarbonate pH 8.9. Enzymaticdigestions were performed using trypsin in 100 mM ammoni-um bicarbonate buffer pH 7.8 overnight at 37 °C. Afterincubation, the reaction was quenched by the addition offormic acid to pH 2. Equipment and technique for LC-ESI MS-MS/MS are described in the Supplements.
2.10. Immunofluorescence studies for α-enolase
Renal biopsy specimens were embedded in OCT (Tissue Tek,Miles Inc., Elkhart, IN, USA) and stored in liquid nitrogen.Three μm sections were cut by a cryostat (Leica CM1850, LeicaMycrosystems, Germany) and placed on poly-L-lysine coatedglass slides for indirect immunostaining. Details of stainingare given in the Supplements. Co-localization of α-enolasewith IgG4 and C5b-9 was analyzed by Confocal Microscopy(analysis) as detailed in the Supplements.
2.11. Detection of circulating antibodies against α-enolase
Detection of anti α-enolase antibodies in sera of MGN patientswas assessed by western blot of podocyte cell line membraneextracts (see below) separatedby 2-dimensional electrophoresisand by dot-blot analysis with recombinant α-enolase (AbnovaCorporation, Taiwan). Detailed technique for transblot anddetection is described in the Supplement methods.
2.12. Electron microscopy
Immunoelectron microscopy was carried out on small frag-ments of the kidney biopsies fixed in Karnovsky's solution andprocessed according to standard methods omitting post-fixation in osmium tetroxide [23]. The technique of analysisis described in the Supplements.
2.13. Detection of other circulating antibodies
Anti SOD2 and anti AR circulating antibody levels weredetermined as already described [17]. The technique for antiPLA2r is described in the Supplements.
2.14. Statistical analysis
Non parametric Mann–Whitney U-test was used for comparisonof anti α-enolase IgG4 serum titer in MGN vs. controls. Resultswere given as median. In Table 1 data are presented as medianand inter-quartile range.
For the analysis of correlation of α-enolase auto-antibodylevels with clinical variables (i.e. protenuria or creatinuria) andother antibodies (AR, SOD, PLA) we utilized nonparametric
Spearman test using GraphPad 5 software. Differences wereconsidered statistically significant at two-tailed p values<0.05.
3. Results
3.1. Characterization of IgG4 target antigens in MGNglomeruli
The experimental approach to podocyte antigens in patientswith MGN [16,17] started with the characterization of theaffinity with podocyte proteins of antibodies (IgG1–4) elutedfrom laser-captured glomeruli. MGN biopsies from 8 patientswith MGN were studied (Table 2). Validation of the relevantresults was done in a cohort of 131 patients.
Western-blot utilizing human podocyte cell lines (gentle giftfrom Dr M. A. Saleem) protein extracts separated with mono-and two-dimensional electrophoresis in reducing conditionsconfirmed the existence of 8 reacting bands in a range of MWbetween 10 and 120 kDa [17] that were concomitantly recog-nized by IgG4 eluted from glomeruli of all the 8 investigatedpatients. The major three (see Fig. 1a–b) have been alreadycharacterized as vimentin and its fragments, AR and SOD2 byLC–Ms and by specific antibodies. One more protein could bedetected at 210 kDa in non reducing conditions and wascharacterized as PLA2r by specific antibodies (Fig. 1b, d). Threeof the remaining five unknown components (see asterisks inFig. 1) were characterized by LC–Ms (Table 3) and their identitywas confirmed by western blot after one- (Fig. 1c, d) and two-dimensional electrophoresis (Fig. 2a, b). The component with95 kDawas characterized as elongation factor 2, the second oneat 85 kDawas characterized as Glycyl Aminoacyl-tRNA Synthe-tase and finally the protein at 47 kDawas shown to beα-enolase(Table 3). A fourth component (neurolysin) co-migrated withGlycyl Aminoacyl-tRNA Synthetase and actually superimposedwith spots 4 and 5 of this protein. Neurolysin could not beconfirmed with western-blot.
Among the aforementioned three podocyte immunoproteins,α-enolase was of particular interest since it was recognized byglomerular IgG4 with an intensity equivalent to SOD2 and wasalso recognizedbycirculating IgG4. For this reason,α-enolasewasstudied utilizing further techniques (see below). The other twopotential auto-antigens are also under evaluation; elongationfactor 2 is of interest since circulating antibodies against thisproteinhavebeendetectedinseraofMGNpatients (seeSection3.4on circulating antibodies).
3.2. Characterization of anti α-enolase IgG4 in MGNglomeruli
Ιn a first approach, it was defined the IgG isotype of α-enolaseantibodies. Dot-blot analysis with recombinant α-enolase asimmobilized antigen showed reactivity with IgG1 and IgG4; IgG2
was completely un-reactive and IgG3 were eluted from only aglomerular sample (Fig. 3a). Reactivity of IgG4 was abolished bypre-treatingMGNglomerular eluateswith increasing amount ofrecombinant protein (5 and 15 ng) (Fig. 3b) confirming thespecificity of the reaction.
Overall, this first set of experiments demonstrated thepresence of specific IgG1 and IgG4 against glomerular α-enolase

Fig. 1 – Characterization of podocyte antigens recognized by MGN glomerular eluates obtained with laser-capture. Podocyteextractswereseparatedbymono-dimensional electrophoresis in reducing (a,c) andnonreducing (b,d) conditionsandwerestudiedfor reactivity with eluates frommicrodissected glomeruli. Thirty micrograms of protein were utilized for the analysis; afterelectrophoresis podocyte extracts were incubated with a pool of glomerular eluates fromMN patients (Pool) and with each singleeluate deriving from renal biopsies of 8 MGN (1–8); representative normal kidney eluates were run as control; gels were thendeveloped with anti- IgG4.. Eight antigens were recognized in reducing conditions and characterized by mass spectrometry asaldose reductase (AR), superoxide dismutase 2 (SOD2) and Vimentin. One more band with a molecular weight of 210 kDa wasdetected in non-reducing conditions and was characterized as phospholipase A2 receptor (PLA2r). Five proteins remaineduncharacterized (see asterisks) and were characterized by LC–MS and western-blot. Three of themwere identified as α-enolase,elongation factor 2 and Glycyl Aminoacyl-tRNA Synthetase (c, d).
2012 J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 2 0 0 8 – 2 0 1 7
in glomeruli of all the MGN patients who were studied. Otherexperimentswere then performed to establish co-localization ofα-enolase within sub-epithelial immune-deposits with bothigG4 and C5b-9.
3.3. Alpha-enolase co-localizes with IgG4 and C5b-9 inMGN glomeruli
Immunoelectronmicroscopy (IEM) forα-enolasewasperformedin three patients affected by MGN stage II. These patients wereselected among the ones with positivity for anti α-enolasecirculating antibodies (see below). Notably, gold particles wereclearly found in glomerular basement membrane (GBM) in thecontext of electron-dense sub-epithelial immune-deposits
(Fig. 4b). Alpha-enolase was also found in the cytoplasm ofpodocytes (Fig. 4a). Nuclear staining was negative, confirmingthe specificity of the reaction (Supplement Fig. 1c). Negativecontrols were obtained by omitting specific primary antibodiesor utilizing them after pre-incubation with non-immune rabbitserum (Supplement Fig. 1b).
Confocal microscopy further confirmed IEM results indicat-ing co-localizationofα-enolasewithboth IgG4 (Fig. 5a–c) and toaminor extent with C5b-9 (Supplement Fig. 2). Co-localizationwith IgG4 was diffuse but not involving the whole glomerulus(see in Supplement Fig. 3, Z-stack confocal images showingdifferent layers of the same glomerulus). Co-localization withC5b-9 was instead less widespread, probably also because of amore limited deposition.
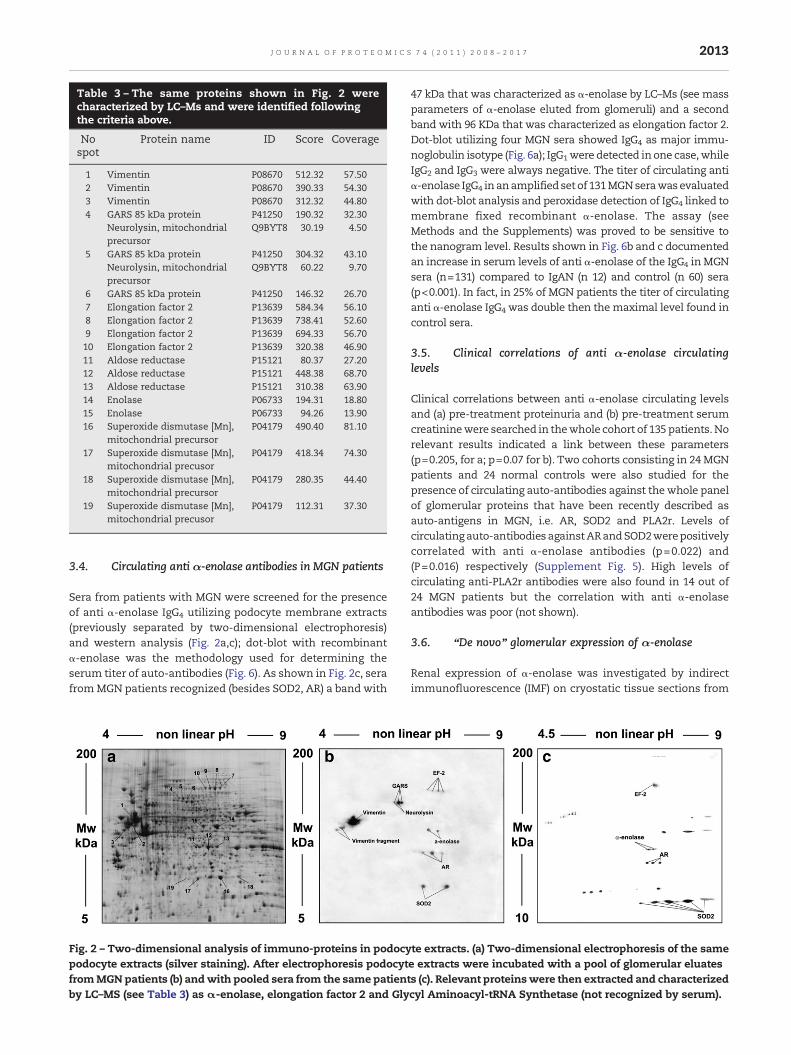
Table 3 – The same proteins shown in Fig. 2 werecharacterized by LC–Ms and were identified followingthe criteria above.
Nospot
Protein name ID Score Coverage
1 Vimentin P08670 512.32 57.502 Vimentin P08670 390.33 54.303 Vimentin P08670 312.32 44.804 GARS 85 kDa protein P41250 190.32 32.30
Neurolysin, mitochondrialprecursor
Q9BYT8 30.19 4.50
5 GARS 85 kDa protein P41250 304.32 43.10Neurolysin, mitochondrialprecursor
Q9BYT8 60.22 9.70
6 GARS 85 kDa protein P41250 146.32 26.707 Elongation factor 2 P13639 584.34 56.108 Elongation factor 2 P13639 738.41 52.609 Elongation factor 2 P13639 694.33 56.7010 Elongation factor 2 P13639 320.38 46.9011 Aldose reductase P15121 80.37 27.2012 Aldose reductase P15121 448.38 68.7013 Aldose reductase P15121 310.38 63.9014 Enolase P06733 194.31 18.8015 Enolase P06733 94.26 13.9016 Superoxide dismutase [Mn],
mitochondrial precursorP04179 490.40 81.10
17 Superoxide dismutase [Mn],mitochondrial precusor
P04179 418.34 74.30
18 Superoxide dismutase [Mn],mitochondrial precursor
P04179 280.35 44.40
19 Superoxide dismutase [Mn],mitochondrial precusor
P04179 112.31 37.30
2013J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 2 0 0 8 – 2 0 1 7
3.4. Circulating anti α-enolase antibodies in MGN patients
Sera from patients with MGN were screened for the presenceof anti α-enolase IgG4 utilizing podocyte membrane extracts(previously separated by two-dimensional electrophoresis)and western analysis (Fig. 2a,c); dot-blot with recombinantα-enolase was the methodology used for determining theserum titer of auto-antibodies (Fig. 6). As shown in Fig. 2c, serafromMGN patients recognized (besides SOD2, AR) a band with
Fig. 2 – Two-dimensional analysis of immuno-proteins in podocypodocyte extracts (silver staining). After electrophoresis podocytfromMGNpatients (b) andwith pooled sera from the same patientby LC–MS (see Table 3) as α-enolase, elongation factor 2 and Gly
47 kDa that was characterized as α-enolase by LC–Ms (see massparameters of α-enolase eluted from glomeruli) and a secondband with 96 KDa that was characterized as elongation factor 2.Dot-blot utilizing four MGN sera showed IgG4 as major immu-noglobulin isotype (Fig. 6a); IgG1were detected inone case,whileIgG2 and IgG3 were always negative. The titer of circulating antiα-enolase IgG4 inanamplified set of 131MGNserawasevaluatedwith dot-blot analysis and peroxidase detection of IgG4 linked tomembrane fixed recombinant α-enolase. The assay (seeMethods and the Supplements) was proved to be sensitive tothe nanogram level. Results shown in Fig. 6b and c documentedan increase in serum levels of anti α-enolase of the IgG4 inMGNsera (n=131) compared to IgAN (n 12) and control (n 60) sera(p<0.001). In fact, in 25% of MGN patients the titer of circulatinganti α-enolase IgG4 was double then the maximal level found incontrol sera.
3.5. Clinical correlations of anti α-enolase circulatinglevels
Clinical correlations between anti α-enolase circulating levelsand (a) pre-treatment proteinuria and (b) pre-treatment serumcreatininewere searched in thewhole cohort of 135 patients.Norelevant results indicated a link between these parameters(p=0.205, for a; p=0.07 for b). Two cohorts consisting in 24MGNpatients and 24 normal controls were also studied for thepresence of circulating auto-antibodies against thewhole panelof glomerular proteins that have been recently described asauto-antigens in MGN, i.e. AR, SOD2 and PLA2r. Levels ofcirculating auto-antibodies againstARandSOD2werepositivelycorrelated with anti α-enolase antibodies (p=0.022) and(P=0.016) respectively (Supplement Fig. 5). High levels ofcirculating anti-PLA2r antibodies were also found in 14 out of24 MGN patients but the correlation with anti α-enolaseantibodies was poor (not shown).
3.6. “De novo” glomerular expression of α-enolase
Renal expression of α-enolase was investigated by indirectimmunofluorescence (IMF) on cryostatic tissue sections from
te extracts. (a) Two-dimensional electrophoresis of the samee extracts were incubated with a pool of glomerular eluatess (c). Relevant proteinswere then extracted and characterizedcyl Aminoacyl-tRNA Synthetase (not recognized by serum).

Fig. 3 – Anti α-enolase IgGs isotype in MGN glomerular eluates. (a) Dot-blot of eluates from microdissected glomeruli of threedifferent MGN patients (Gl 1–3) showed the presence of specific anti α-enolase IgG1 and IgG4. IgG3 were also present at a lowertiter; IgG2 were negative. Controls were incubated with human serum albumin (HSA) instead of glomerular eluates.(b) Competition experiment using the same glomerular eluates as in panel (a) and increasing amount ofα-enolase from5 to 15 ng.
2014 J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 2 0 0 8 – 2 0 1 7
renal biopsies of patients affected by MGN (n=10), minimalchange disease (MCD — n=2), membranoproliferative glo-merulonephtitis (MPGN — n=2) and IgA nephropathy (n=4).Non-carcinomatous portions of kidneys (n=4) removed forrenal cancer were used as negative control. Staining wererepeated on sample tissues utilizing polyclonal rabbit and twomonoclonal (clones 251/3 and 276/3) antibodies (not shown).
Polyclonal antibodies revealed the presence of glomerular α-enolase in all MGN patients, with a focal distribution (Supple-ment Fig. 4a, b). In the majority of patients α-enolase wassegmental (Supplement Fig. 4b).Glomerular stainingwasabsentin normal kidneys and in all other tested nephritides with theexception of two IgAN patients who presented scatteredglomerular deposits. Tubular positivity was present in allanalyzed kidney sections, including tubular cells in normalkidneys, indicating a good sensitivity of polyclonal antibodies(Supplement Fig. 4c, d). Blocking experiments with increasingamount of recombinant alpha-enolase abolished the staining,demonstrating specificity of employed antibodies (data notshown). Tubular cells were in all cases positive.
Fig. 4 – Immunoelectron microscopy. Alpha-enolase is present incytosol (b) of podocyte bodies (Po), while it is not present in the nprimary antibody is totally negative (see Supplement Fig. 1). Orig
4. Discussion
The knowledge onmechanisms responsible for autoimmunityin human MGN is now rapidly growing. Genetic studies aredefining the predisposing basis of the disease in humans [24]and evolutions based on proteomic approaches are producinginsights on target auto-antigens. In fact, the definition of theauto-antibody panel eluted from glomeruli of patients withMGN is an essential start up for any evolution on theunderstanding of fine mechanisms. Two independent groupshave recently addressed this point showing the existence ofauto-antibodies against a panel of podocyte antigens, three ofwhich have been characterized as PLA2r, SOD2 and AR [16,17].However, as here demonstrated, IgG4 eluted from glomeruli ofMGN patients recognized also other proteins that representgood candidates for new auto-antigens. The list of proposedimmunoproteins here presented includes α-enolase, elonga-tion factor 2 and Glycyl Aminoacyl-tRNA Synthetase. Anti-bodies versus all the antigens above been reported in patients
sub-epithelial immune-deposits (a) and appears also in theucleus (see Supplement Fig. 1) (and) Control sample lackinginal magnifications (a) ×56,000; (b): ×44,000.
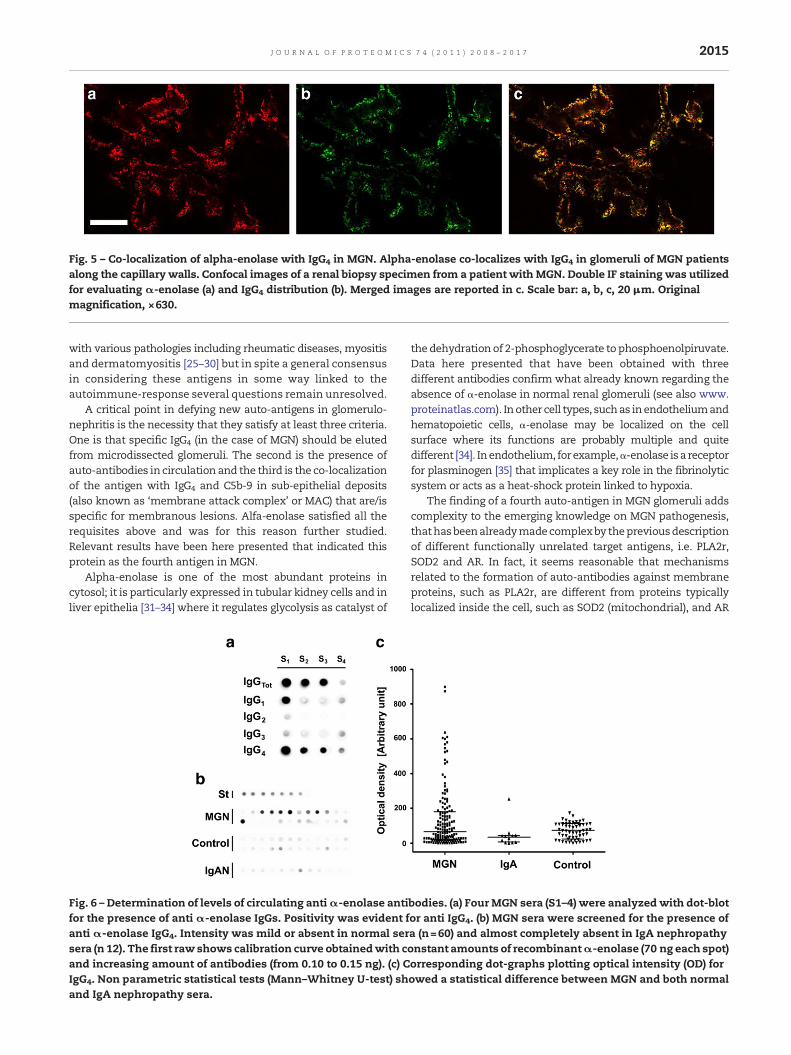
Fig. 5 – Co-localization of alpha-enolase with IgG4 in MGN. Alpha-enolase co-localizes with IgG4 in glomeruli of MGN patientsalong the capillary walls. Confocal images of a renal biopsy specimen from a patient with MGN. Double IF staining was utilizedfor evaluating α-enolase (a) and IgG4 distribution (b). Merged images are reported in c. Scale bar: a, b, c, 20 μm. Originalmagnification, ×630.
2015J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 2 0 0 8 – 2 0 1 7
with various pathologies including rheumatic diseases, myositisand dermatomyositis [25–30] but in spite a general consensusin considering these antigens in some way linked to theautoimmune-response several questions remain unresolved.
A critical point in defying new auto-antigens in glomerulo-nephritis is the necessity that they satisfy at least three criteria.One is that specific IgG4 (in the case of MGN) should be elutedfrom microdissected glomeruli. The second is the presence ofauto-antibodies in circulation and the third is the co-localizationof the antigen with IgG4 and C5b-9 in sub-epithelial deposits(also known as ‘membrane attack complex’ or MAC) that are/isspecific for membranous lesions. Alfa-enolase satisfied all therequisites above and was for this reason further studied.Relevant results have been here presented that indicated thisprotein as the fourth antigen in MGN.
Alpha-enolase is one of the most abundant proteins incytosol; it is particularly expressed in tubular kidney cells and inliver epithelia [31–34] where it regulates glycolysis as catalyst of
Fig. 6 – Determination of levels of circulating antiα-enolase antifor the presence of anti α-enolase IgGs. Positivity was evident fanti α-enolase IgG4. Intensity was mild or absent in normal sersera (n 12). The first raw shows calibration curve obtainedwith cand increasing amount of antibodies (from 0.10 to 0.15 ng). (c) CIgG4. Non parametric statistical tests (Mann–Whitney U-test) shand IgA nephropathy sera.
the dehydrationof 2-phosphoglycerate to phosphoenolpiruvate.Data here presented that have been obtained with threedifferent antibodies confirm what already known regarding theabsence of α-enolase in normal renal glomeruli (see also www.proteinatlas.com). Inother cell types, suchas inendotheliumandhematopoietic cells, α-enolase may be localized on the cellsurface where its functions are probably multiple and quitedifferent [34]. Inendothelium, for example,α-enolase isa receptorfor plasminogen [35] that implicates a key role in the fibrinolyticsystem or acts as a heat-shock protein linked to hypoxia.
The finding of a fourth auto-antigen in MGN glomeruli addscomplexity to the emerging knowledge on MGN pathogenesis,thathasbeenalreadymadecomplexby thepreviousdescriptionof different functionally unrelated target antigens, i.e. PLA2r,SOD2 and AR. In fact, it seems reasonable that mechanismsrelated to the formation of auto-antibodies against membraneproteins, such as PLA2r, are different from proteins typicallylocalized inside the cell, such as SOD2 (mitochondrial), and AR
bodies. (a) Four MGN sera (S1–4) were analyzed with dot-blotor anti IgG4. (b) MGN sera were screened for the presence ofa (n=60) and almost completely absent in IgA nephropathyonstant amounts of recombinantα-enolase (70 ng each spot)orresponding dot-graphs plotting optical intensity (OD) forowed a statistical difference between MGN and both normal

2016 J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 2 0 0 8 – 2 0 1 7
orα-enolase (cytosolic). The correlationherepresentedbetweenthe circulating titers of anti α-enolase and the other antibodiesagainst cytoplasm antigens (SOD2 and AR), but not withantibodies against the membrane antigen PLA2r, supports thisdifference. Anyway, this is not the first example of antibodypromiscuity, a relevant one being anti-endothelial antibodiesthat have been detected in a host of autoimmune diseases andin normal beings [36,37]. It is of note that anti-endothelialantibodies recognizesmost of the podocyte immunoproteins ofMGN including cytoskeleton proteins such as vimentin andenzymes of the cytoplasm such as α-enolase and AR.
Onahypothetical ground,mimicry canexplainautoimmunityfor membrane proteins whereas for molecules such as SOD2and AR or α-enolase, mechanisms of de-localization shouldhold an importance. A secondmain aspect is to define whetherthere is a hierarchy of auto-antibody deposition in which, anactivationofa first auto-antigen induces the formationofapriorclass of auto-antibodies that, in turn, stimulates the expressionof other auto-antigens. Both crucial aspects related to patho-genesis of MGN require an experimental approach based onanimal models engineered for selective hyper-expression of allthese antigens [38].
Even if α-enolase is the last protein in the list of podocyteauto-antigens, its potential implication inautoimmunediseasesis not new and poses, in some way, the question of specificity.Actually, with the methodology limitations that characterizedthe pre-proteomics era, this protein was shown as the target ofspecific circulating antibodies in patients with a variety ofautoimmune and inflammatory diseases such as in lupuserythematosus, mixed cryoglobulinemia and ANCA positivevasculitis [19,31,39–41]. Wakui and colleagues [40] showedcirculating anti α-enolase antibodies of the IgG1 and IgG3 classesin patients with primary and secondary MGN. Anti α-enolaseantibodies of the IgG1 and IgG4 class can also be detected inserum and in glomeruli of patients with lupus erythematosus(Ghiggeri et al. , personal observation) but the significance oftheir presence in LES [19,38] is still unclear and must be furtherinvestigated. In fact, a commonmotif betweenMGNand stage Vlupus nephritis is the presence of a shared panel of glomerularauto-antibodies, including anti α-enolase present in bothconditions (Ghiggeri et al., personal observation).
The presence of IgG4 in serum against a panel of podocyteantigens opens a question about validation of circulating auto-antibodies as surrogate biomarkers of their renal deposition andof the clinical outcome in patients with MGN. A strong effort inthe creation of clinical databases and bio-banks that will makeavailable a large number ofMGN seramust be undertaken. Alsothe organization of a laboratory network is necessary to avoidvariability in biochemical essays. We hope these goals will berapidly achieved among research teams involved in the finaldiscovery of mechanisms of renal autoimmunity.
In conclusion, the results of this study show the presencein glomeruli of patientswithMGN of specific antibodies of theIgG4 class against α-enolase, elongation factor 2 and GlycylAminoacyl-tRNA Synthetase. Anti α-enolase IgG4 were alsodetected by electron microscopy and immunofluorescence inglomeruli where they form electron-dense deposits and co-localizewith C5b-9. Circulating anti α-enolase IgG4were, finally,detected in serum on the same patients. Renal expression ofα-enolase was increased in MGN glomeruli implicating de-
novo synthesis and/or delocalization of this protein. Podocyteα-enolase may be considered the fourth auto-antigen ofidiopathic MGN in humans.
Supplementary materials related to this article can be foundonline at doi:10.1016/j.jprot.2011.05.021.
Acknowledgements
Weare grateful toDrM.A. Saleemfor giving ushumanpodocytecell lines and to Prof Paola Migliorini for anti α-enolasemonoclonal antibodies. This work was supported by the ItalianMinistry of Health and by the Renal Child Foundation. Authorsalso acknowledge Fondazione Mara Wilma e Bianca Querci forfinancial support to the project “Ruolo dello stress reticolare nellaprogressione del danno renale e tumurale”, Fondazione La NuovaSperanza for supporting the project ‘Progetto integrato per ladefinizione dei meccanismi implicati nella glomerulo sclerosi focale.Dalla predisposizione genetica alla regolazione della produzione difattori cellulari e circolanti’ And finally, Fondazione Compagnia diSan Paolo for the project ‘Nefropatia cronica del rene trapiantato’.
The authors gratefully acknowledge the support of theFoundation Guido Erluison for making available the lasercapture microdissection system. Data were critically discussedwith Prof. Rosanna Gusmano before her death and the paper isdedicated to her memory.
R E F E R E N C E S
[1] Heymann W, Hackel DB, Harwood S, Wilson SG, Hunter JL.Production of nephrotic syndrome in rats by Freund'sadjuvants and rat kidney suspensions. Proc Soc Exp Biol Med1959;100:660–4.
[2] Farquhar MG, Saito A, Kerjaschki D, Orlando RA. TheHeymann nephritis antigenic complex: megalin (gp330) andRAP. J Am Soc Nephrol 1995;6:35–47.
[3] Adler S, Baker PJ, Pritzl P, Couser WG. Detection of terminalcomplement components in experimental immune glomerularinjury. Kidney Int 1984;26:830–7.
[4] Kerjaschki D, Farquhar MG. The pathogenic antigen ofHeymann nephritis is a membrane glycoprotein of the renalproximal tubule brush border. Proc Natl Acad Sci USA 1982;79:5557–61.
[5] Kerjaschki D, Farquhar MG. Immunocytochemical localizationof the Heymann nephritis antigen (GP330) in glomerularepithelial cells of normal Lewis rats. J Exp Med 1983;157:667–86.
[6] Kerjaschki D, Horvat R, Binder S, Susani M, Dekan G, Ojha PP,et al. Identification of a 400-kd protein in the brush borders ofhumankidney tubules that is similar togp330, thenephritogenicantigen of rat Heymann nephritis. Am J Pathol 1987;129:183–91.
[7] CouserWG, Steinmuller DR, StilmantMM, Salant DJ, LowensteinLM. Experimental glomerulonephritis in the isolated perfusedrat kidney. J Clin Invest 1978;62:1275–87.
[8] Van Damme BJ, Fleuren GJ, Bakker WW, Vernier RL,Hoedemaeker PJ. Experimental glomerulonephritis in the ratinduced by antibodies directed against tubular antigens. V. Fixedglomerular antigens in the pathogenesis of heterologous im-mune complex glomerulonephritis. Lab Invest 1978;38:502–10.
[9] Salant DJ, Belok S, Madaio MP, Couser WG. A new role forcomplement in experimental membranous nephropathy inrats. J Clin Invest 1980;66:1339–50.

2017J O U R N A L O F P R O T E O M I C S 7 4 ( 2 0 1 1 ) 2 0 0 8 – 2 0 1 7
[10] Saito A, Pietromonaco S, Loo AK, Farquhar MG. Completecloning and sequencing of rat gp330/“megalin,” a distinctivemember of the low density lipoprotein receptor gene family.Proc Natl Acad Sci USA 1994;91:9725–9.
[11] Ghiggeri GM, Bruschi M, Candiano G, Rastaldi MP, Scolari F,Passerini P, et al. Depletion of clusterin in renal diseasescausing nephrotic syndrome. Kidney Int 2002;62:2184–94.
[12] Rastaldi MP, Armelloni S, Berra S, Calvaresi N, Corbelli A,Giardino LA, et al. Glomerular podocytes contain neuron-likefunctional synaptic vesicles. FASEB J 2006;20:976–8.
[13] Bruschi M, Candiano G, Murtas C, Prunotto M, Santucci L,Carnevali ML, et al. Patients with primary membranousnephropathy lack auto-antibodies against LDL receptor, thehomologue of megalin in human glomeruli. NDT Plus 2009:sfp002.
[14] Debiec H, Guigonis V, Mougenot B, Decobert F, Haymann JP,Bensman A, et al. Antenatal membranous glomerulonephritisdue to anti-neutral endopeptidase antibodies. N Engl J Med2002;346:2053–60.
[15] Debiec H, Nauta J, Coulet F, van der Burg M, Guigonis V,Schurmans T, et al. Role of truncating mutations in MMEgene in fetomaternal alloimmunisation and antenatalglomerulopathies. Lancet 2004;364:1252–9.
[16] Beck Jr LH, Bonegio RG, Lambeau G, Beck DM, Powell DW,Cummins TD, et al. M-type phospholipase A2 receptor astarget antigen in idiopathic membranous nephropathy. NEngl J Med 2009;361:11–21.
[17] Prunotto M, Carnevali ML, Candiano G, Murtas C, Bruschi M,Corradini E, et al. Autoimmunity in membranous nephropathytargets aldose reductase and SOD2. J Am Soc Nephrol 2010;21:507–19.
[18] Murtas C, Bruschi M, Carnevali ML, Petretto A, Corradini E,Prunotto M, et al. In vivo characterization of renal auto-antigens involved in human auto-immune diseases: thecase of membranous glomerulonephritis. Proteomics ClinAppl 2011;5:90–7.
[19] Migliorini P, Pratesi F, Bongiorni F, Moscato S, Scavuzzo M,Bombardieri S. The targets of nephritogenic antibodies insystemic autoimmune disorders. Autoimmun Rev 2002;1:168–73.
[20] Bruschi M, Musante L, Candiano G, Ghiggeri GM, Herbert B,Antonucci F, et al. Soft immobilized pH gradient gels inproteome analysis: a follow-up. Proteomics 2003;3:821–5.
[21] Laemmli UK. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 1970;227:680–5.
[22] Candiano G, Bruschi M, Musante L, Santucci L, Ghiggeri GM,Carnemolla B, et al. Blue silver: a very sensitive colloidalCoomassieG-250 staining for proteomeanalysis. Electrophoresis2004;25:1327–33.
[23] D'Adda T, Bertele A, Pilato FP, Bordi C. Quantitative electronmicroscopy of endocrine cells in oxyntic mucosa of normalhuman stomach. Cell Tissue Res 1989;255:41–8.
[24] Stanescu HC, Arcos-Burgos M, Medlar A, Bockenhauer D,Kottgen A, Dragomirescu L, et al. Risk HLA-DQA1 and PLA(2)R1 alleles in idiopathic membranous nephropathy. N Engl JMed 2011;364:616–26.
[25] Alberdi F, Dadone J, Ryazanov A, Isenberg DA, Ravirajan C,Reichlin M. Cross-reaction of lupus anti-dsDNA antibodieswith protein translation factor EF-2. Clin Immunol 2001;98:293–300.
[26] Arora S, Yang JM, Craft J, Hait W. Detection of anti-elongationfactor 2 kinase (calmodulin-dependent protein kinase III)antibodies in patients with systemic lupus erythematosus.Biochem Biophys Res Commun 2002;293:1073–6.
[27] Bunn CC, Bernstein RM, Mathews MB. Autoantibodies againstalanyl-tRNAsynthetaseand tRNAAla coexist and areassociatedwith myositis. J Exp Med 1986;163:1281–91.
[28] NishikaiM,ReichlinM.Heterogeneity ofprecipitatingantibodiesin polymyositis and dermatomyositis. Characterization of theJo-1 antibody system. Arthritis Rheum 1980;23:881–8.
[29] Ge Q, Trieu EP, Targoff IN. Primary structure and functionalexpression of human Glycyl-tRNA synthetase, an autoantigenin myositis. J Biol Chem 1994;269:28790–7.
[30] Targoff IN. Autoantibodies to aminoacyl-transfer RNAsynthetases for isoleucine and glycine. Two additionalsynthetases are antigenic in myositis. J Immunol 1990;144:1737–43.
[31] Sabbatini A, Dolcher MP, Marchini B, Chimenti D, Moscato S,Pratesi F, et al. Alpha-enolase is a renal-specific antigenassociated with kidney involvement inmixed cryoglobulinemia.Clin Exp Rheumatol 1997;15:655–8.
[32] Aaronson RM, Graven KK, Tucci M, McDonald RJ, Farber HW.Non-neuronal enolase is an endothelial hypoxic stressprotein. J Biol Chem 1995;270:27752–7.
[33] Wistow GJ, Lietman T, Williams LA, Stapel SO, de Jong WW,Horwitz J, et al. Tau-crystallin/alpha-enolase: one geneencodes both an enzyme and a lens structural protein. J CellBiol 1988;107:2729–36.
[34] Miles LA, Dahlberg CM, Plescia J, Felez J, Kato K, Plow EF. Roleof cell-surface lysines in plasminogen binding to cells:identification of alpha-enolase as a candidate plasminogenreceptor. Biochemistry 1991;30:1682–91.
[35] Redlitz A, Fowler BJ, Plow EF, Miles LA. The role of anenolase-relatedmolecule in plasminogen binding to cells. Eur JBiochem 1995;227:407–15.
[36] Youinou P. New target antigens for anti-endothelial cellantibodies. Immunobiology 2005;210:789–97.
[37] ServettazA,GuilpainP,TamasN,Kaveri SV,CamoinL,MouthonL. Natural anti-endothelial cell antibodies. Autoimmun Rev2008;7:426–30.
[38] Pratesi F, Moscato S, Sabbatini A, Chimenti D, Bombardieri S,Migliorini P. Autoantibodies specific for alpha-enolase insystemic autoimmune disorders. J Rheumatol 2000;27:109–15.
[39] Moodie FD, Leaker B, Cambridge G, Totty NF, Segal AW.Alpha-enolase: a novel cytosolic autoantigen in ANCApositive vasculitis. Kidney Int 1993;43:675–81.
[40] Wakui H, Imai H, Komatsuda A, Miura AB. Circulatingantibodies against alpha-enolase in patients with primarymembranous nephropathy (MN). Clin Exp Immunol 1999;118:445–50.
[41] Terrier B, Degand N, Guilpain P, Servettaz A, Guillevin L,Mouthon L. Alpha-enolase: a target of antibodies in infectiousand autoimmune diseases. Autoimmun Rev 2007;6:176–82.




















