
Antibody Recognizing Gametocyte/Oocyst Antigens
37
1 Excystation of Eimeria tenella Sporozoites Impaired by 1 Antibody Recognizing Gametocyte/Oocyst Antigens 2 GAM22 and GAM56* 3 4 Jürgen Krücken 1* , Ralf J. Hosse 1 , Aimdip N. Mouafo 2 , Rolf Entzeroth 2 , Stefan Bierbaum 1 , 5 Predrag Marinovski 1 , Karolina Hain 1 , Gisela Greif 3 , and Frank Wunderlich 1 6 7 8 Division of Molecular Parasitology and Biological and Medical Research Centre, Heinrich- 9 Heine-University, Düsseldorf 1 , Institute of Zoology, TU-Dresden, Dresden 2 , Animal Health 10 Business Group, Research & Development, Bayer AG Monheim 3 , Germany. 11 12 Running Title: Impaired Excystation of Eimeria tenella 13 14 *Corresponding author. Mailing address: Division of Molecular Parasitology, Heinrich- 15 Heine-University, Universitätsstr. 1, 40225 Düsseldorf, Germany. Phone: 49-211-8114733. 16 Fax: 49-211-8114734. E-Mail: [email protected] 17 18 The nucleotide sequence data reported in this paper appear in the DDBJ, EMBL and 19 Genbank® nucleotide sequence database with accession number CS000361. 20 ACCEPTED Copyright © 2007, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved. Eukaryotic Cell doi:10.1128/EC.00292-07 EC Accepts, published online ahead of print on 14 December 2007
Transcript of Antibody Recognizing Gametocyte/Oocyst Antigens

1
Excystation of Eimeria tenella Sporozoites Impaired by 1
Antibody Recognizing Gametocyte/Oocyst Antigens 2
GAM22 and GAM56* 3
4
Jürgen Krücken1*, Ralf J. Hosse1, Aimdip N. Mouafo2, Rolf Entzeroth2, Stefan Bierbaum1, 5
Predrag Marinovski1, Karolina Hain1, Gisela Greif3, and Frank Wunderlich1 6
7
8
Division of Molecular Parasitology and Biological and Medical Research Centre, Heinrich-9
Heine-University, Düsseldorf1, Institute of Zoology, TU-Dresden, Dresden
2, Animal Health 10
Business Group, Research & Development, Bayer AG Monheim3, Germany. 11
12
Running Title: Impaired Excystation of Eimeria tenella 13
14
*Corresponding author. Mailing address: Division of Molecular Parasitology, Heinrich-15
Heine-University, Universitätsstr. 1, 40225 Düsseldorf, Germany. Phone: 49-211-8114733. 16
Fax: 49-211-8114734. E-Mail: [email protected] 17
18
The nucleotide sequence data reported in this paper appear in the DDBJ, EMBL and 19
Genbank® nucleotide sequence database with accession number CS000361. 20
ACCEPTED
Copyright © 2007, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Eukaryotic Cell doi:10.1128/EC.00292-07 EC Accepts, published online ahead of print on 14 December 2007

2
Abstract 1
Eimeria tenella is the causative agent of coccidiosis in poultry. Infection of chicken 2
intestine begins with ingestion of sporulated oocysts releasing sporocysts which in turn 3
release invasive sporozoites. The monoclonal antibody E2E5 recognizes wall-forming 4
bodies type II (WFBII) in gametocytes and the WFBII-derived inner wall of oocysts. 5
Here, we describe that this antibody also binds to the stieda body of sporocysts and 6
significantly impairs in vitro excystation of sporozoites. Using affinity chromatography 7
and protein sequence analysis, E2E5 is shown to recognize EtGAM56, the E. tenella 8
ortholog of the Eimeria maxima gametocyte-specific GAM56 protein. In addition, this 9
antibody was used to screen a genomic phage display library presenting E. tenella 10
antigens as fusion proteins with gene VIII product on the surface of phagemid particles 11
and identified the novel 22 kDa histidine- and proline-rich protein EtGAM22. The 12
Etgam22 mRNA is predominantly expressed at the gametocyte stage as detected by 13
Northern blotting. Southern blot analysis in combination with data from the E. tenella 14
genome project revealed that Etgam22 is an intronless multi-copy gene with 15
approximately 12-22 copies in head to tail arrangement. Conspicuously, Etgam56 is also 16
intronless and is localized adjacent to another gam56-like gene, Etgam59. Our data 17
suggest that amplification is common for genes encoding oocyst wall proteins. 18
ACCEPTED

3
INTRODUCTION 1
Coccidiosis in poultry is caused by protozoan parasites of the genus Eimeria. The 2
worldwide economic losses due to these parasites were estimated to exceed 1.2 billion US $ 3
per annum (41). The most virulent species is Eimeria tenella causing severe hemorrhagic 4
enteritis by infection of the epithelium and submucosa of the caeca and eventually death of 5
infected chickens (24). 6
Eimeria infections occur by ingestion of oocysts (24). In the intestine, oocysts release four 7
sporocysts each containing two sporozoites. After excystation, motile infective sporozoites 8
actively enter cells in the epithelium of the caecum. Then, three rounds of asexual 9
multiplication in epithelium and submucosa are followed by differentiation to sexual stages of 10
micro- and macrogametocytes (23). After fertilization of macrogametes, a complex, two-11
layered wall is secreted around the young oocyst by exocytosis of wall forming bodies type I 12
and type II (WFBI and WFBII) (35). While the 10 nm thick outer oocyst wall is built up by 13
the contents of WFBI, the 90 nm inner oocyst wall is mainly composed of glycoproteins that 14
were stored in the WFBII (31, 37). The oocyst displays a remarkable rigidity and protects the 15
parasite from several physically and chemically adverse influences such as commonly used 16
disinfectants (34). A potential use of gametocyte antigens involved in formation of the oocyst 17
wall as protective transmission-blocking vaccines has been described for Eimeria maxima (2, 18
25, 38-40, 46). 19
The formation of oocyst and sporocyst walls and sporozoite excystation are rather complex 20
processes that we are just beginning to understand. Only a few WFBII-localized glycoproteins 21
have been characterized for E. tenella (10) and for E. maxima. They have been shown to 22
undergo site-specific proteolysis before incorporation into the mature oocyst wall (1, 3, 5). 23
Moreover, there is compelling evidence for occurrence of cross-linking of these tyrosine-rich 24
proteins. This process of sclerotization involves the formation of dityrosine bonds as well as 25
the emergence of covalent bonds between proteins by peroxidase-mediated mechanisms 26
ACCEPTED

4
involving L-3,4-dihydroxyphenylalanine (DOPA). Incidentally, sclerotization of the oocyst 1
wall appears to be comparable to other hardening processes in extracellular matrices such as 2
insect and nematode cuticles, yeast cell walls, mussel byssal threads and sea urchin 3
fertilization membranes (3). 4
In E. tenella, the WFBII of macrogametocytes and the inner oocyst wall can be specifically 5
labeled by the monoclonal antibody E2E5 (26). This antibody recognizes a complex, 6
developmentally regulated pattern of protein bands in Western blot analysis (26). Here, we 7
show that E2E5 recognizes proteins encoded by at least two different genes and impairs 8
sporozoite excystation. 9
ACCEPTED

5
MATERIALS AND METHODS 1
Animals, parasites and infections. The strain E. tenella VT-2 was used throughout all 2
experiments. Male chicken of Leghorn type strain LSL (Josef Brinkschulte GmbH, Senden, 3
Germany) were infected with 15,000 oocysts. For preparation of oocysts, infected chicken 4
were killed, and the content of the caecum was flushed out with 2% potassium dichromate 5
solution. Sporulation of oocysts was completed after stirring in 2% potassium dichromate at 6
28 °C for 48 h. 7
Cell culture. The hybridoma cell lines E1D8 and E2E5 were previously described to 8
specifically recognize antigens in WFBI and WFBII, respectively (26). Hybridomas and the 9
human T cell lymphoma cell line Jurkat were cultivated in RPMI1640 supplemented with 10
10% fetal calf serum at 37 °C, 5% CO2, and 100% humidity. For most experiments, 11
supernatants were concentrated 50fold using Vivaspin concentrators (Sartorius AG, 12
Göttingen, Germany) with a 100 kDa cut off. 13
Immunofluorescence. Reactivity of E2E5 with intracellular E. tenella stages was analyzed 14
as described previously (26). Briefly, semithin sections of LR-White embedded caeca from E. 15
tenella infected chicken were probed with undiluted hybridoma culture supernatants before 16
detection with FITC-coupled anti-mouse immunoglobulin G (IgG) (for E2E5) or rhodamine-17
coupled anti-mouse immunoglobulin M (IgM) (for E1D8) antibodies (Sigma, Germany). 18
Sections were examined using a Zeiss Axioscop 2 microscope. 19
Oocysts were vortexed in the presence of glass beads until rupturing of most walls of 20
oocysts and sporocysts. After centrifugation, the pellet was resuspended in methanol (-20 °C), 21
incubated for 10 min at -20 °C, washed in PBS and subjected to immunofluorescence 22
microscopy as described recently (18). Oocysts and sporocysts were treated with 0.1% Triton 23
X 100 in PBS for 10 min, blocked for 1 h in PBS/1% bovine serum albumin (BSA), incubated 24
with a 1:40 dilution of concentrated E2E5 supernatant for 2 h, and finally visualized with a 25
ACCEPTED

6
1:100 dilution of a secondary goat anti-mouse antibody coupled to Alexa Fluor 488 1
(Molecular Probes, Karlsruhe, Germany). 2
Excystation assay. Freshly sporulated oocysts were ruptured in a glass teflon potter. Free 3
sporocysts were incubated overnight at 4 °C in PBS containing 10 µg/ml Baytril (BAYER, 4
Leverkusen, Germany). Then, sporocyts were collected by centrifugation and resuspended in 5
20 ml PBS containing 2.5 µg/ml trypsin before adding 1 ml of chicken bile. Aliquots of 250 6
µl were incubated in the presence of either 12.5 µl E2E5 50-fold concentrated or control 7
supernatants at 41.5 °C and 5% CO2 for 5 h. Numbers of sporocyts before and of free 8
sporozoites after excystation were counted in a Neubauer chamber. Statistical comparisons 9
between groups were done using a paired Student’s t-test. 10
Affinity chromatography and Edman degradation. E. tenella gametocytes were purified 11
as described recently (26). Proteins were solubilized with 0.5% Triton X-100/PBS containing 12
1 mM phenylmethanesulfonyl fluoride. Columns containing 4 ml protein A sepharose CL-4B 13
covalently crosslinked to E2E5 (Amersham Biosciences, Freiburg, Germany) were loaded 14
with detergent solubilized gametocytes at a flow rate of 0.5 ml/min at 4 °C. Unbound material 15
was washed off with 100 ml PBS supplemented with 1 M NaCl, 0.5% Triton X-100 and 1 16
mM EDTA. Elution of antigen was performed with 0.1 M diethylamine (pH 11.5), 0.1% 17
Triton X-100. Fractions of 1 ml were immediately neutralized with 200 µl TrisCl (pH 7.5), 18
then dialyzed against 0.1 mM TrisCl (pH 6.8) before lyophilizing. Purified 51 kDa protein 19
was separated by sodiumdodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE). 20
Gels were either silver stained or transferred to PVDF membranes (Millipore, Schwalbach, 21
Germany). The NH2-terminus of the protein was determined using Edman degradation at the 22
University of Gent (Belgium). 23
Construction of the phage display library. Genomic DNA was isolated from oocysts 24
according to Blin and Stafford (7). Sheared DNA (150 bp – 800 bp) was ligated into SnaBI 25
digested pG8SAET (45) and transformed into electrocompetent Escherichia coli TG1 cells 26
ACCEPTED

7
(Stratagene, Heidelberg, Germany). Clones from several transformations were pooled to 1
obtain the final library with 4.7 × 106 independent clones (≈ 95% recombinant clones). 2
Phagemids were prepared using the helper phage R408 (Promega, Heidelberg, Germany). 3
Screening of the phage display library. In order to identify phage clones expressing 4
fusion proteins reacting with E2E5, the latter was immobilized on magnetic DYNABEADS® 5
Pan Mouse IgG (Invitrogen) according to the manufacturer’s instructions. All incubations 6
containing DYNABEADS® were carried out at 4 °C under rotation. After blocking with 7
PBS/0.1% BSA, beads were collected using a Dynal MPC®-S magnet (Deutsche Dynal 8
GmbH). For every screening round, two parallel binding reactions containing 2 × 107 control 9
or E2E5-coupled beads and 200 µl phagemids in a final volume of 400 µl PBS/0.1% BSA 10
were set up. After overnight binding, beads were washed 10 times for 5 min with 2 ml 11
PBS/0.1% BSA and once for 15 min with 150 mM NaCl, 50 mM sodium citrate (pH 4.5) 12
before phagemids were eluted with 400 µl 150 mM NaCl, 50 mM sodium citrate (pH 1.8). 13
After neutralization with 40 µl 2 M TrisCl (pH 8.6), the phagemids were used to reinfect E. 14
coli TG1 cells, titered, and amplified before use in consecutive screening rounds. 15
RNA and protein isolation. For reverse transcription and polymerase chain reaction (RT-16
PCR), DNA-free total RNA from ruptured oocysts and chicken caecum was isolated using the 17
Invisorb® RNA Kit II (Invitek, Berlin, Germany). For Northern blot analysis, RNA was 18
isolated from caeca and oocysts using Trizol (Invitrogen). Subsequently, proteins were 19
extracted from the same samples for Western blotting. 20
Western blotting. Protein extracts from E. coli TG1 overnight cultures were separated by 21
SDS-PAGE according to Laemmli (20) and transferred to membranes. A 1:200 dilution of 22
concentrated E2E5 hybridoma supernatant in 1 × RotiBlock (Roth, Karlsruhe) was used as 23
primary antibody. For detection, a 1:4000 dilution of goat anti-mouse IgG coupled to 24
horseradish peroxidase (Jackson Immuno Research Laboratories, Cambridge, UK) was 25
combined with the ECL detection system (Amersham Biosciences, Freiburg). For analysis of 26
ACCEPTED

8
protein expression in E. tenella, proteins were isolated from infected caeca or oocysts using 1
Trizol (Invitrogen). Then, 60 µg samples from caeca or 20 µg samples from oocysts were 2
separated by 9-15% gradient SDS-PAGE, transferred to nitrocellulose membrane and 3
developed as described above. 4
Sequence analyses. DNA sequencing was performed as described previously (19) using a 5
LI-COR 4000 automatic DNA Sequencer. The nucleotide sequence data reported in this paper 6
appear in the DDBJ, EMBL and Genbank® nucleotide sequence database with accession 7
number CS000361. All nucleic acid and deduced protein sequences were compared to 8
GenBank and the E. tenella genome project databases using BLAST algorithms. Protein 9
sequences were analyzed using the InterProScan server (44). NH2-terminal signal peptides 10
were identified using SignalP 3.0 (6) and glycosylation site prediction was performed with 11
NetOGlyc 3.1 (13). The genomic sequence data for Eimeria tenella were produced by the 12
Eimeria Sequencing Group at the Sanger Institute and can be obtained from 13
ftp://ftp.sanger.ac.uk/pub/pathogens/Eimeria/tenella/. For Toxoplasma gondii, preliminary 14
cDNA sequence data were accessed via http://ToxoDB.org. Genomic data were provided by 15
The Institute for Genomic Research (supported by the NIH grant #AI05093), and by the 16
Sanger Center (Wellcome Trust). EST sequences were generated by Washington University 17
(NIH grant #1R01AI045806-01A1). 18
Cloning of the full length Etgam22 cDNA. Rapid amplification of cDNA ends (RACE) 19
was carried out with RNA from sporulated oocysts as template using a 5'/3' RACE kit (Roche, 20
Mannheim, Germany). For 3'-RACE, cDNA was synthesized from 2 µg total RNA using the 21
oligo dT anchor primer. PCR reactions contained 1 µl of cDNA, 10 µl Q solution (Qiagen, 22
Hilden, Germany), 0.4 mM dNTPs, 0.25 µM of the PCR anchor primer, the primer 5'-23
TGAGGACTATCCTAGCCACCCTAGTCGGTTTC-3' and 5 U Expand High Fidelity DNA 24
polymerase (Roche, Mannheim, Germany) in 50 µl High Fidelity buffer. The enzyme was 25
added during the first annealing phase to achieve a hot start. 26
ACCEPTED

9
Synthesis of cDNA for 5'-RACE was started with the Etgam22-specific primer 5'-1
AGGATGCGCAAAATGGTAGTCATGGTGATAAT-3'. The cDNA was purified and tailed 2
with dATP using terminal nucleotide transferase. PCR was carried out with 5 µl purified 3
cDNA as template as described for 3'-RACE using the PCR anchor primer and 5'-4
TATGTTCATGATGATGATGGTGAGGATGATGG-3' for the first round and 5'-5
GTGGGGATGATGGTCGGG-3' for a nested PCR. All PCR reactions were carried out with 6
the same cycling parameters. After 2 min denaturation at 94 °C, 35 cycles 15 s at 94 °C, 30 s 7
at 63 °C, and 4 min at 72 °C were executed with a final extension for 10 min at 72 °C. PCR 8
products were cloned into the pCR2.1 TOPO vector (Invitrogen, Karlsruhe, Germany). 9
Southern blotting. Genomic DNA was isolated from sporozoites using the Invisorb® 10
Genomic DNA Kit II. Southern blots were carried out as detailed previously (18). Briefly, 10 11
µg genomic DNA were digested with the indicated restriction enzymes, separated in 0.7% 12
agarose gels, and transferred to Hybond-N nylon membrane (Amersham). Hybridization was 13
performed using the full length Etgam22 cDNA as probe. For estimation of the Etgam22 copy 14
number, the Etgam22 cDNA in the plasmid pcDNA3.1 was linearized with XhoI, and defined 15
amounts of linearized plasmid and DraI digested genomic DNA were mixed before analyzing 16
twofold serial dilutions of the mixture by Southern blotting. Blots were exposed to Kodak 17
Biomax MS film with intensifying screen at -80 °C or to a Fuji BAS-MS imaging plate and 18
read out with a Fuji BAS-1500 phosphorimager. Densitometric analysis was performed with 19
QuantiScan 1.5 (Biosoft, Cambridge, UK). Signal intensities from the plasmid DNA were 20
plotted against the copy number, and the copy number in genomic DNA was calculated 21
assuming a haploid genome size of 60 Mb. 22
RT-PCR. RNA (3.5 µg) was reverse transcribed in 50 µl 1 × AMV buffer containing 0.4 23
mM dNTPs, 5 mM DTT, 2.5 µM random hexamer primers, 50 U AMV Reverse transcriptase 24
(all components from Roche, Mannheim, Germany) and 80 U RNasin (Promega, Heidelberg, 25
Germany). Reactions were incubated at 22 °C for 10 min, at 42 °C for 30 min, and at 55 °C 26
ACCEPTED

10
for 30 min before inactivating the enzyme by heating at 95 °C for 5 min. As negative controls, 1
parallel reactions were incubated without AMV Reverse transcriptase or without RNA. PCR 2
reactions contained 5 µl of the RT reactions, 0.4 mM dNTPs, 0.4 µM of each primer, 10 µl Q 3
solution (Qiagen, Hilden, Germany), and 5 U Expand High Fidelity DNA polymerase (Roche, 4
Mannheim, Germany) in 50 µl 1 × High Fidelity buffer. Primers for amplification of Etgam22 5
were 5'-TCCTCATCCTTATCATCCTCATCCT-3' and 5'-GTGGGGATGATGGTCGGG-3'. 6
Control PCRs amplifying an actin fragment used the primers 5'-7
CTGTGAGAAGAACCGGGTGCTCTTC-3' and 5'-8
CGTGCGAAAATGCCGGACGAAGAG-3'. The reactions were denatured for 2 min at 94 °C 9
followed by 35 cycles 15 s at 94 °C, 30 s at 63 °C and 1 min at 72 °C. A final extension for 10 10
min at 72 °C completed the reactions. 11
Amplification of full length Etgam22 cDNA was carried out with 5 µl cDNA from infected 12
caeca at 137 h p.i. Reactions were performed as described above with the primers 5'- 13
CAGGACCCCAAAATAAAATCAAAGGCTATCACA-3' and 5'- 14
TGACCGGTGGTGTGTACTTCGTAAC-3'. 15
Northern blotting. RNA (20 µg) was glyoxylated, separated in agarose gels, transferred to 16
Hybond-N membrane (Amersham) and hybridized as described recently (42). 17
In vitro translation. Coupled in vitro transcription and translation was performed using 18
the TnT Quick Coupled Transcription/Translation System (Promega). In vitro translation 19
products were separated by SDS-PAGE and subjected to Western blotting using the E2E5 20
antibody. 21
ACCEPTED

11
RESULTS 1
In vitro inhibition of excystation. The monoclonal antibody E2E5 recognizes antigens in 2
the WFBII of gametocytes and in the inner oocysts wall of E. tenella (Fig. 1A, B, and C), thus 3
confirming our previous results (26). In addition, we can show here that E2E5 reacts with the 4
apical stieda body in ruptured but not in intact sporocysts (Fig. 1D). This indicates that the 5
epitope is not accessible for the E2E5 antibody from the outside. 6
Since the apical stieda body of sporocysts is involved in the excystation process of 7
sporozoites, we also tried to inhibit in vitro excystation of E. tenella sporozoites from the 8
sporocysts with E2E5. The oocyst walls were mechanically disrupted and excystation was 9
induced by incubation of free sporocysts with trypsin and bile at 41.5 °C for 5 h. In these 10
control experiments, between 48% and 77% of all possibly available sporozoites were 11
released from sporocysts. Addition of supernatants from E2E5 significantly decreased the 12
release of sporozoites by about 50% (Fig. 1E). In contrast, neither supernatants from E1D8 13
hybridoma cells nor supernatants from Jurkat cells – with or without supplementation with an 14
irrelevant IgG2a antibody (anti-V5 tag) – had any significant effect on excystation. 15
E2E5 recognizes a complex pattern of proteins in the different developmental stages of E. 16
tenella (23). In order to identify genes encoding the corresponding proteins, we used two 17
complementary approaches: First, we characterized the largest of the proteins by affinity 18
chromatography and Edman degradation. Secondly, we used phage display to select cDNAs 19
encoding proteins recognized by E2E5. 20
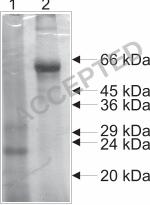
Purification and identification of EtGAM56 from gametocytes. Young gametocytes 21
express only a single protein of about 51 kDa that is recognized by E2E5. Affinity 22
chromatography was performed with E2E5 coupled to proteinA agarose. The protein fraction 23
eluted from the column was separated by SDS-PAGE and either silver-stained (Fig. 2) or 24
blotted on PVDF membrane for protein sequencing. Edman degradation of the NH2-terminus 25
resulted in the sequence VPTTVENTVHPYSEMGHYQEGRPYAAYMG. Database 26
ACCEPTED

12
screening using this sequence revealed a 78% identity of amino acids 3 to 21 of this sequence 1
to the GAM56 protein of Eimeria maxima (EmGAM56) (1, 5). Moreover, 20 out of the first 2
21 amino acids in this sequence can be deduced from the sequence of a genomic contig 3
deposited in the E. tenella database. An intronless open reading frame of 374 amino acids 4
encoding a 54 kDa protein can be deduced from the corresponding region of the genomic 5
sequence dev_EIMER_contig_00030093 - eimer-679b04.p1ka (compare supplemental Fig. 6
1). This putative protein has 63% identity and 73% similarity with EmGAM56 as revealed by 7
BLAST analysis. Since both EmGAM56 and the 54 kDa protein recognized by E2E5 are 8
selectively expressed in the macrogametocyte stage and antibodies to EmGAM56 also 9
selectively recognize the wall forming bodies II, we propose that the 54 kDa protein is the E. 10
tenella ortholog to GAM56 of E. maxima and should be designated as EtGAM56. 11
Interestingly, a second GAM56-like protein of 59 kDa is encoded on the same contig and 12
in the same orientation, about 2.3 kb upstream of Etgam56. We designate this intronless gene 13
Etgam59 though we have not shown gametocyte specific expression of Etgam59 yet. Fig. 3 14
shows a multiple sequence alignment of both deduced E. tenella proteins with the GAM56 15
sequence of E. maxima. The sequence of all three GAM56-like proteins is rich in the amino 16
acids Ala, Pro, Thr, Ser. Moreover, all these proteins contain a Tyr-rich domain. EmGAM56 17
is known to be present in the WFBII, the inner oocyst wall and the apical part of the 18
sporocyst, and this fits perfectly well to our data using immunofluorescence and 19
immunoelectron microscopy presented in Fig. 1 and in Mouafo et al. (26). Remarkably, 20
EmGAM56 has been shown to be proteolytically cleaved during oocyst formation into mature 21
polypeptides of 33 kDa and 12 kDa (3). Indeed, the protein band corresponding to EtGAM56 22
disappears in unsporulated oocysts and a strong band slightly larger than 30 kDa appears (26). 23
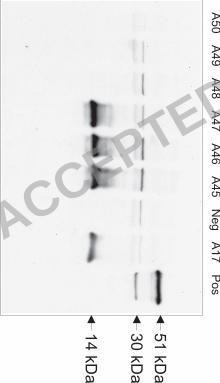
Cloning of EtGAM22 by phage display. In addition to EtGAM56 and its 33 kDa 24
proteolytic fragment, E2E5 recognizes two smaller proteins of approximately 23 kDa and 25 25
kDa in unsporulated oocysts and an about 80 kDa protein in sporulated oocysts (26). In order 26
ACCEPTED

13
to identify additional proteins recognized by E2E5, a genomic phage display library 1
presenting recombinant E. tenella proteins as fusions with the gene VIII product of 2
filamentous phages on the surface of phagemid particles has been constructed. After four 3
rounds of library panning against E2E5 coupled to dynabeads, enrichment of a clone 4
containing an insert of 145 bp was observed (data not shown). Western blot analysis clearly 5
revealed that this phage clone A17 expresses a recombinant gene VIII product fusion-protein 6
that is recognized by E2E5 while other clones were consistently negative (Fig. 4). 7
Using 5' and 3'-RACE PCR, we could clone the complete open reading frame of the 8
corresponding mRNA. Fig. 5 shows the cDNA sequence of Etgam22 and the deduced protein 9
sequence. The EtGAM22 protein has a length of 198 amino acids, a molecular weight of 22.8 10
kDa, and a predicted pI of 6.8. At the NH2-terminus, a signal peptide for cotranslational 11
transport into the ER is predicted by the SignalP 3.0 program (6). The amino acid 12
composition of the mature EtGAM22 protein without signal peptide is very unusual since 13
only four amino acids, His (25.7%), Pro (19%), Gln (8.4%), and Ala (6.1%) account for 14
nearly 60% of all residues. In particular, an extremely His- and Pro-rich domain between 15
Pro73 and His188 can be identified. This partially resembles EmGAM56 which contains 12.8% 16
Pro and 8% Ala. In contrast to EtGAM22, however, EmGAM56 contains only 0.6% His and 17
2.9% Gln residues, whereas EtGAM22 contains no Tyr-rich domain characteristic for other 18
gametocyte-specific proteins like EmGAM56 and EmGAM82 (3). The sequence of the 19
original phage clone A17 containing the E2E5 epitope is completely localized within this 20
His/Pro-rich domain. Moreover, there are putative O-glycosylation sites at Thr74, Thr80 and 21
Thr81 as predicted by NetOGlyc 3.1. 22
Genomic organization of EtGAM22. BLAST analysis of the sequences deposited by the 23
E. tenella genome project identified several perfect matches showing that the Etgam22 gene – 24
like Emgam56, Etgam56 and Etgam59 – does not contain any intron. Conspicuously, there are 25
two contigs (dev_EIMER_contig_00030727 - eimerbac28g8Fb07.q1k (compare supplemental 26
ACCEPTED

14
Fig. 2) and dev_EIMER_contig_00016586 - eimer-437a12.q1k) with two identical head to 1
tail repeats of the Etgam22 gene. One of these contigs is schematically drawn in Fig. 6A. In 2
addition, the intergenic regions between the copies are also nearly perfectly conserved (99% 3
identity) indicating that the promoter regions are virtually identical. In order to demonstrate 4
that Etgam22 is indeed a multicopy gene, we performed genomic Southern blot analysis using 5
6 different restriction enzymes (Fig. 6B). With exception of DraI, all enzymes produced one 6
intensive band and one or two faint bands which is in accordance with several virtually 7
identical head to tail copies of Etgam22. In particular, it should be mentioned that the 8
restriction sites for all the enzymes used here are located outside of the Etgam22 open reading 9
frame indicating a high degree of conservation even in the non-coding sequences of the 10
cluster. The faint bands represent unique fragments from the end of the Etgam22 cluster. We 11
used two complementary approaches to estimate the Etgam22 copy number in the E. tenella 12
genome. The genomic DNA digested with BglI and MvaI in Fig. 6B yields two bands, i.e. a 13
faint band representing a single copy located at the end of the Etgam22 cluster and a strong 14
band due to hybridization to the other members of the cluster. After correction for different 15
overlaps of internal and flanking Etgam22 copies with the hybridization probe, it should be 16
possible to calculate the copy number from the ratio of band intensities. Our results indicate 17
that there are 14 times more hybridization targets in the strong than in the faint BglI band and 18
11 times more targets in the strong than in the faint MvaI band. Therefore, we assume that 19
there are between 12 and 15 copies of Etgam22 in the genome. 20
For an independent approach to estimate the copy number of Etgam22, we used the single 21
band produced in Southern blots after digestion with DraI. Genomic DNA from E. tenella 22
was digested with DraI and mixed with XhoI restricted Etgam22 in the plasmid vector 23
pcDNA3.1 (6,629 bp). Two-fold serial dilutions of this mixture were analyzed by Southern 24
hybridization (Fig. 6C). After densitometric evaluation of band intensities, the optical density 25
of the plasmid bands was evaluated using a phosphorimager and was plotted against the copy 26
ACCEPTED

15
number per lane. Using this standard curve, the number of Etgam22 copies in genomic DNA 1
per lane was calculated from band intensities for genomic DNA. Finally, the Etgam22 copy 2
number per genome was calculated to be 19 ± 3 assuming a genome size of 60 Mbp and using 3
only those genomic DNA bands with intensities within the range of the standard curve. 4
Presumably, this is a more accurate value since the limited linear range of X-ray film 5
compared to a phosphorimager may lead to underestimation of strong band intensities. 6
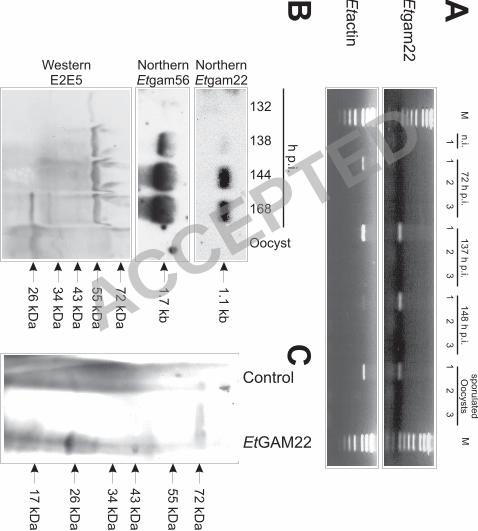
Gametocyte-specific expression of EtGAM22. RT-PCR analysis shows that the Etgam22 7
mRNA is not expressed in the early stage of infection at 72 h p.i. whereas it is readily 8
detectable at 137 h p.i., 148 h p.i. and in the sporulated oocysts (Fig. 7A). However, 9
expression levels in these stages differ widely as revealed by Northern blot analysis (Fig. 7B). 10
Thus, Etgam22 is not detectable in sporulated oocysts while a faint hybridization signal is 11
obtained from RNA of gametocytes at 138 h p.i. Then, expression of Etgam22 mRNA 12
dramatically increases at 148 h p.i. and 168 h p.i., when mature gametocytes and a mixture of 13
gametocytes and unsporulated oocysts, respectively, are present in the caeca. Expression of 14
Etgam56 mRNA is already very weakly detectable at 132 h p.i. and shows maximum 15
expression between 144 h p.i. and 168 h p.i. 16
We also extracted proteins from the same tissue samples and analyzed expression of 17
proteins using the antibody E2E5 (Fig. 7B). Weak expression of the about 60 kDa band can 18
be detected as early as 132 h p.i. Bands of about 33 kDa and 25 kDa appear not before 168 h 19
p.i. and are still detectable in sporulated oocysts. In this context, it is noteworthy that, in E. 20
maxima, the ortholog EmGAM56 is cleaved into a 33 kDa polypeptide during oocyst 21
formation (3). The 33 kDa protein in Fig. 7B is therefore assumed to represent the proteolytic 22
fragment of EtGAM56. In accordance with our previous results, a 25 kDa band is also 23
observed at 168 h p.i. However, we could not reproduce the weak 23 kDa band detected 24
previously in purified unsporulated oocysts (26). Since in vitro translation of Etgam22 cDNA 25
also produces a polypeptide of about 25 kDa (Fig. 7C), the 25 kDa band at 168 h p.i. most 26
ACCEPTED

16
likely corresponds to EtGAM22. This view is also consistent with the fact that processing of 1
EmGAM56 does not give rise to polypeptide fragments in the size range of 20-25 kDa (3). 2
ACCEPTED

17
DISCUSSION 1
The WFBII in gametocytes provide essential components for the rigid and impermeable 2
wall of oocysts, the formation of which is a critical step in the life cycle of Eimeria parasites 3
which exclusively rely on the fecal/oral route for infection of new hosts. Using the 4
monoclonal antibody E2E5, we describe the E. tenella GAM56 ortholog and confirm its 5
proteolytic processing during oocyst wall formation. Moreover, we identify a novel type of 6
small gametocyte-specific protein, EtGAM22, which also appears to be a structural 7
component of the oocyst wall. 8
The genes encoding EtGAM22 and EtGAM56 identified here in E. tenella and the 9
previously characterized genes for the WFBII-localized proteins EmGAM56 and EmGAM82 10
in E. maxima have several characteristics in common. All of them are specifically expressed 11
in gametocytes. Moreover, there is a remarkable absence of introns in these genes. Although 12
Emgam56 was described to be a single copy gene (5), our data presented here indicate the 13
presence of two closely related genes, Etgam56 and Etgam59, in the genome of E. tenella. 14
Etgam22 is the first multi-copy gene described for Eimeria species and its extraordinary high 15
copy number and the extremely conserved sequence between the copies suggest that Etgam22 16
has a particularly important role for oocyst wall formation. The high degree of conservation 17
indicates that amplification of the Etgam22 gene was either a very recent evolutionary event 18
or that the sequences of the copies are frequently equalized by unequal cross-over or gene 19
conversion which prevent sequence diversification as previously described for multi-gene 20
families including rRNA (33) and histone genes (30, 32). Apparently, such mechanisms are 21
not acting on gam56-like genes. 22
In apicomplexa, low copy head-to-tail clusters of homologous intronless genes have been 23
described for some of the T. gondii SAG genes that encode GPI-coupled surface antigens 24
expressed by the invasive parasite stages (11, 21, 36). Sequence similarity and expression 25
level in different developmental stages vary widely between family members in the same 26
ACCEPTED

18
cluster and intergenic regions are usually not conserved – with exception of SAG5B and 1
SAG5C (36). By contrast, the intergenic regions in the Etgam22 gene cluster appear to be 2
highly conserved – as far as it can be deduced from the few sequenced repeats and the 3
conservation of restriction sites, though we do not yet know whether all genes in the cluster 4
are transcriptionally active. Expression of both Etgam56 and Etgam22 mRNAs is very high, 5
since exposure times of Northern blots as short as 2.5 h and 4 h, respectively, were sufficient 6
to produce clearly detectable bands despite the fact that the RNA was not isolated from pure 7
parasites but from infected caeca and will therefore contain large amounts of chicken RNA. It 8
is therefore tempting to speculate that amplification of Etgam22 gene copy number might be a 9
mechanism to allow production of large amounts of RNA within a very short period as 10
previously described for histone genes, which are highly expressed only during a short phase 11
of the cell cycle. Despite the high level of Etgam22 mRNA, however, the protein band 12
detected in Western blot is surprisingly quite faint. There are several possible explanations for 13
this observation. First, reactivity of the antibody E2E5 might be better for EtGAM56 than for 14
EtGAM22 or extraction of EtGAM56 from the tissue might be easier than that of EtGAM22. 15
Second, translation of Etgam22 mRNA might be inefficient and only low levels of protein are 16
produced. Third, translation of Etgam22 mRNA is developmentally regulated and does not 17
occur in macrogametocytes but in oocysts after fusion of gametes or during sporulation. This 18
view is also supported by the fact that the EtGAM22 protein is not detectable at 144 h p.i. 19
when transcript levels are maximal and although transcription starts at least 6 h earlier. 20
However, the protein becomes detectable at 168 h p.i. when the first unsporulated oocysts 21
appear in the caecum as revealed by the beginning of proteolytic processing of EtGAM56. 22
At the protein level, similarity between EtGAM22 and EtGAM56 manifests itself as cross-23
reactivity with E2E5, as presence of putative N-glycosylation sites, and as NH2-terminal 24
signal peptides. The presence of this signal peptide and the fact that, in macrogametocytes, 25
E2E5 exclusively recognizes WFBII, i.e. specialized regions within the rough ER (9), suggest 26
ACCEPTED

19
that EtGAM22 – like EmGAM56 – is transported to the WFBII and participates in formation 1
of the inner oocyst wall and/or the stieda body. In E. maxima, a GAM56-specific monoclonal 2
antibody has been reported to recognize the WFBII in macrogametocytes, the oocyst wall, the 3
outer sporocyst wall, and the stieda body (5). Since this pattern largely overlaps with the 4
pattern of E2E5 labeling in E. tenella, a monoclonal antibody against EtGAM22 not cross-5
reacting with EtGAM56 will be neccessary to evaluate the exact localization of EtGAM22. 6
Other His-rich proteins are known from parasites belonging to the phylum apicomplexa 7
with the histidine-rich protein of Plasmodium lophurae (78% His residues) (12) displaying 8
the most exceptional composition. However, the low sequence complexity of these proteins 9
and EtGAM22 does not permit a phylogenetic analysis. Indeed, those His-rich proteins for 10
which a function is known, are surely not homologous to EtGAM22. For instance, the knob 11
associated histidine-rich protein of Plasmodium falciparum is involved in the interaction of 12
parasite proteins in knobs on the erythrocyte surface with the host cell cytoskeleton (27, 29) 13
whereas the histidine-rich protein 2 has been implicated in hemozoin formation of P. 14
falciparum (28). By contrast, BLAST comparison of EtGAM22 with proteins predicted from 15
the Toxoplasma gondii genome reveals the presence of at least five proteins with NH2-16
terminal signal peptides which are relatively rich in proline and histidine and have a length in 17
the range of 117 to 269 amino acids. Though there is not yet anything known about the 18
function or developmental expression pattern of these proteins, it is tempting to speculate that 19
some of them represent extracellular structural proteins such as e.g. many proline- or 20
hydroxyproline-rich glycoproteins in plant cell walls (15, 16) or histidine-rich proteins in 21
Hydra nematocysts (17). Cross-linking of EmGAM56 and EmGAM82 derived peptides via 22
dopamine has been shown in the oocyst wall (3) and His-rich proteins such as EtGAM22 23
might also be involved in stabilizing extracellular structures via cross-links between His and 24
catechols as described for insect cuticles (8, 14, 43). 25
ACCEPTED

20
The in vitro excystation inhibition assay presented here shows that the antibody E2E5 can 1
significantly interfere with parasite development. Remarkably, vaccination with a preparation 2
of native gametocyte antigens enriched for GAM56, GAM82 and GAM230, which are all 3
involved in wall forming in E. maxima, has been shown to convey at least partial protection 4
against homologous challenge (25, 39, 46). Since EtGAM22 represents a new family of 5
oocyst wall proteins it might be a useful supplement to a protective gametocyte-specific 6
cocktail vaccine. 7
ACCEPTED

21
Acknowledgements 1
We would like to thank Carsten Angenendt for valuable technical assistance. 2
ACCEPTED

22
REFERENCES 1
1. Belli, S. I., Lee, M., Thebo, P., Wallach, M. G., Schwartsburd, B., and Smith, N. C. 2
2002. Biochemical characterisation of the 56 and 82 kDa immunodominant 3
gametocyte antigens from Eimeria maxima. Int. J. Parasitol. 32: 805-816. 4
2. Belli, S. I., Mai, K., Skene, C. D., Gleeson, M. T., Witcombe, D. M., Katrib, M., 5
Finger, A., Wallach, M. G., and Smith, N. C. 2004. Characterisation of the antigenic 6
and immunogenic properties of bacterially expressed, sexual stage antigens of the 7
coccidian parasite Eimeria maxima . Vaccine 22: 4316-4325. 8
3. Belli, S. I., Wallach, M. G., Luxford, C., Davies, M. J., and Smith, N. C. 2003. Roles 9
of tyrosine-rich precursor glycoproteins and dityrosine- and 3,4-10
dihydroxyphenylalanine-mediated protein cross-linking in development of the oocyst 11
wall in the coccidian parasite Eimeria maxima. Eukaryot. Cell 2: 456-464. 12
4. Belli, S. I., Wallach, M. G., and Smith, N. C. 2003. Cloning and characterization of 13
the 82 kDa tyrosine-rich sexual stage glycoprotein, GAM82, and its role in oocyst 14
wall formation in the apicomplexan parasite Eimeria maxima. Gene 307: 201-212. 15
5. Belli, S. I., Witcombe, D., Wallach, M. G., and Smith, N. C. 2002. Functional 16
genomics of gam56: characterisation of the role of a 56 kilodalton sexual stage antigen 17
in oocyst wall formation in Eimeria maxima. Int. J. Parasitol. 32: 1727-1737. 18
6. Bendtsen, J. D., Nielsen, H., von Heijne, G., and Brunak, S. 2004. Improved 19
prediction of signal peptides: SignalP 3.0. J. Mol. Biol. 340: 783-795. 20
7. Blin, N., and Stafford, D. W. 1976. A general method for isolation of high molecular 21
weight DNA from eukaryotes. Nucleic Acids Res. 3: 2303-2308. 22
8. Christensen, A. M., Schaefer, J., Kramer, K. J., Morgan, T. D., and Hopkins, T. L. 23
1991. Detection of cross-links in insect cuticle by redor nmr-spectroscopy. J. Am. 24
Chem. Soc. 113: 6799-6802. 25
ACCEPTED

23
9. Ferguson, D. J., Belli, S. I., Smith, N. C., and Wallach, M. G. 2003. The development 1
of the macrogamete and oocyst wall in Eimeria maxima: immuno-light and electron 2
microscopy. Int. J. Parasitol. 33: 1329-1340. 3
10. Fetterer, R. H., and Barfield, R. C. 2003. Characterization of a developmentally 4
regulated oocyst protein from Eimeria tenella. J. Parasitol. 89: 553-564. 5
11. Hehl A., Krieger T., and Boothroyd J. C. 1997. Identification and characterization of 6
SRS1, a Toxoplasma gondii surface antigen upstream of and related to SAG1. 7
Mol. Biochem. Parasitol. 89: 271-282. 8
12. Irving, D. O. and Cross, G. A. 1986. Primary structure of the histidine-rich protein of 9
Plasmodium lophurae. Nature 324: 492. 10
13. Julenius, K., Molgaard, A., Gupta, R., and Brunak, S. 2005. Prediction, conservation 11
analysis, and structural characterization of mammalian mucin-type O-glycosylation 12
sites. Glycobiology 15: 153-164. 13
14. Kerwin, J. L., Turecek, F., Xu, R. D., Kramer, K. J., Hopkins, T. L., Gatlin, C. L., and 14
Yates, J. R. 1999. Mass spectrometric analysis of catechol-histidine adducts from 15
insect cuticle. Anal. Biochem. 268: 229-237. 16
15. Kieliszewski, M. J., and Lamport, D. T. 1994. Extensin: repetitive motifs, functional 17
sites, post-translational codes, and phylogeny. Plant J. 5: 157-172. 18
16. Kieliszewski, M. J., and Shpak, E. 2001. Synthetic genes for the elucidation of 19
glycosylation codes for arabinogalactan-proteins and other hydroxyproline-rich 20
glycoproteins. Cell Mol. Life Sci. 58, 1386-1398. 21
17. Koch, A. W., Holstein, T. W., Mala, C., Kurz, E., Engel, J., and David, C. N. 1998. 22
Spinalin, a new glycine- and histidine-rich protein in spines of Hydra nematocysts. J. 23
Cell Sci. 111: 1545-1554. 24
ACCEPTED

24
18. Krücken, J., Schroetel, R. M., Müller, I. U., Saïdani, N., Marinovski, P., Benten, W. 1
P., Stamm, O., and Wunderlich, F. 2004. Comparative analysis of the human gimap 2
gene cluster encoding a novel GTPase family. Gene 341: 291-304. 3
19. Krücken, J., Stamm, O., Schmitt-Wrede, H.-P., Mincheva, A., Lichter, P., and 4
Wunderlich, F. 1999. Spleen-specific expression of the malaria-inducible intronless 5
mouse gene imap38. J. Biol. Chem. 274: 24383-24391. 6
20. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head 7
of bacteriophage T4. Nature 227: 680-685. 8
21. Lekutis C., Ferguson D. J., Grigg M. E., Camps M., Boothroyd J. C. 2001. Surface 9
antigens of Toxoplasma gondii: variations on a theme. Int. J. Parasitol. 31:1285-92. 10
22. Marzluff, W. F., Gongidi, P., Woods, K. R., Jin, J., and Maltais, L. J. 2002. The 11
human and mouse replication-dependent histone genes. Genomics 80: 487-498. 12
23. McDonald, V. and Rose, M. E. (1987). Eimeria tenella and E. necatrix: a third 13
generation of schizogony is an obligatory part of the developmental cycle. J. Parasitol. 14
73, 617-622. 15
24. Mehlhorn, H. 2005. Eimeria, p. 198-199. In H. Mehlhorn (ed.), Encyclopedic 16
reference of parasitology. Biology, structure, function. Springer, Berlin, Germany. 17
25. Michael, A. The practical use of a maternal vaccine against coccidiosis. World Poultry 18
19: 24-26. 19
26. Mouafo, A. N., Weck-Heimann, A., Dubremetz, J. F., and Entzeroth, R. 2002. 20
Monoclonal antibodies specific for the two types of wall-forming bodies of Eimeria 21
tenella macrogametes (Coccidia, Apicomplexa). Parasitol. Res. 88: 217-224. 22
27. Oh, S. S., Voigt, S., Fisher, D., Yi, S. J., LeRoy, P. J., Derick, L. H., Liu, S., and 23
Chishti, A. H. 2000. Plasmodium falciparum erythrocyte membrane protein 1 is 24
anchored to the actin-spectrin junction and knob-associated histidine-rich protein in 25
the erythrocyte skeleton. Mol. Biochem. Parasitol. 108: 237-247. 26
ACCEPTED

25
28. Pandey, A. V., Babbarwal, V. K., Okoyeh, J. N., Joshi, R. M., Puri, S. K., Singh, R. 1
L., and Chauhan, V. S. 2003. Hemozoin formation in malaria: a two-step process 2
involving histidine-rich proteins and lipids. Biochem. Biophys. Res. Commun. 308: 3
736-743. 4
29. Pei, X., An, X., Guo, X., Tarnawski, M., Coppel, R., and Mohandas, N. 2005. 5
Structural and functional studies of interaction between Plasmodium falciparum knob-6
associated histidine-rich protein (KAHRP) and erythrocyte spectrin. J. Biol. Chem. 7
280: 31166-31171. 8
30. Piontkivska, H., Rooney, A. P., and Nei, M. 2002. Purifying selection and birth-and-9
death evolution in the histone H4 gene family. Mol. Biol. Evol. 19: 689-697. 10
31. Pittilo, R. M., and Ball, S. J. (1980). The ultrastructural development of the oocyst 11
wall of Eimeria maxima. Parasitology 81: 115-122. 12
32. Rooney, A. P., Piontkivska, H., and Nei, M. 2002. Molecular evolution of the 13
nontandemly repeated genes of the histone 3 multigene family. Mol. Biol. Evol. 19: 14
68-75. 15
33. Rooney, A. P., and Ward, T. J. 2005. Evolution of a large ribosomal RNA multigene 16
family in filamentous fungi: birth and death of a concerted evolution paradigm. Proc. 17
Natl. Acad. Sci. U. S. A 102: 5084-5089. 18
34. Ryley, J. F. 1973. Cytochemistry, physiology and biochemistry, p. 145-181. In D. M. 19
Hammond, and P. L. Long (ed.), The coccidia. University Park Press, London. 20
35. Scholtyseck, E., Mehlhorn, H., and Hammond, D. M. 1971. Fine structure of 21
macrogametes and oocysts of Coccidia and related organisms. Z. Parasitenkd. 37: 1-22
43. 23
36. Spano F., Ricci I., Di Cristina M., Possenti A., Tinti M., Dendouga N., Tomavo S., 24
and Crisanti A. 2002. The SAG5 locus of Toxoplasma gondii encodes three novel 25
ACCEPTED

26
proteins belonging to the SAG1 family of surface antigens. Int. J. Parasitol. 32: 121-1
131. 2
37. Stotish, R. L., Wang, C. C., and Meyenhofer, M. 1978. Structure and composition of 3
the oocyst wall of Eimeria tenella. J. Parasitol. 64: 1074-1081. 4
38. Wallach, M. 1997. The importance of transmission-blocking immunity in the control 5
of infections by apicomplexan parasites. Int. J. Parasitol. 27: 1159-1167. 6
39. Wallach M. 2002. The development of a novel vaccine against coccidiosis. World 7
Poultry 19: 24-26. 8
40. Wallach, M. G., Mencher, D., Yarus, S., Pillemer, G., Halabi, A., and Pugatsch, T. 9
1989. Eimeria maxima: identification of gametocyte protein antigens. Exp. Parasitol. 10
68: 49-56. 11
41. Williams, R. B., Carlyle, W. W., Bond, D. R., and Brown, I. A. 1999. The efficacy 12
and economic benefits of Paracox, a live attenuated anticoccidial vaccine, in 13
commercial trials with standard broiler chickens in the United Kingdom. Int. J. 14
Parasitol. 29: 341-355. 15
42. Wunderlich, F., Dkhil, M. A., Mehnert, L. I., Braun, J. V., El Khadragy, M., Borsch, 16
E., Hermsen, D., Benten, W. P., Pfeffer, K., Mossmann, H., and Krücken, J. 2005. 17
Testosterone responsiveness of spleen and liver in female lymphotoxin beta receptor-18
deficient mice resistant to blood-stage malaria. Microbes. Infect. 7: 399-409. 19
43. Xu, R. D., Huang, X., Hopkins, T. L., and Kramer, K. J. 1997. Catecholamine and 20
histidyl protein cross-linked structures in sclerotized insect cuticle. Insect Bioch. 21
Molec. 27: 101-108. 22
44. Zdobnov, E. M., and Apweiler, R. 2001. InterProScan – an integration platform for the 23
signature-recognition methods in InterPro. Bioinformatics 17: 847-848. 24
ACCEPTED

27
45. Zhang, L., Jacobsson, K., Strom, K., Lindberg, M., and Frykberg, L. 1999. 1
Staphylococcus aureus expresses a cell surface protein that binds both IgG and beta2-2
glycoprotein I. Microbiology 145: 177-183. 3
46. Ziomko R., Karamon J., Cencek T. Gornowicz E, Skoracki A, and Ashash U. 2005. 4
Prevention of broiler chick coccidiosis using the inactivated subunit vaccine CoxAbic. 5
B. Vet. I. Pulawy 49: 299-302 6
7
ACCEPTED

28
FIGURE LEGENDS 1
2
FIG. 1. Functional characterization of antigens recognized by E2E5. (A) and (B) show 3
semithin sections of ceaca from infected chicken. Macrogamonts (A) were labeled with E2E5 4
and an anti-mouse IgG antibody coupled to FITC. The same parasites are shown by bright-5
field microscopy (B). (C) Localization of E2E5 reactive antigens at the inner oocyst wall 6
(black arrowhead) is shown by double-labeling with E2E5 and anti-mouse IgG-FITC and then 7
with E1D8 and anti-mouse IgM-rhodamine. E1D8 recognizes the outer oocyst wall (white 8
arrowhead). (D) Localization of E2E5-binding structures in sporocysts. Oocysts were 9
mechanically ruptured with glass beads before labeling with E2E5 and a goat anti-mouse 10
Alexa488-coupled secondary antibody. The arrow indicates the position of the stieda body. The 11
arrowhead marks a small rupture of the wall in the sporocyst. Separate phase contrast and 12
green fluorescent pictures were taken and merged using Corel PHOTO-PAINT 10. All bars 13
represent 10 µm. (E) Inhibition of in vitro excystation by the monoclonal antibody E2E5. In 14
vitro excystation was performed in the presence of 50-fold concentrated E2E5 or control 15
supernatants derived from the hybridoma E1D8 or from Jurkat cells. As an isotype control, 16
concentrated Jurkat cells supernatant was supplemented with 250 µg/ml anti-V5 IgG2a 17
antibody (Invitrogen). Excystation efficiencies evaluated as number of released sporozoites 18
per total number of sporozoites contained in sporocysts were calculated and normalized to the 19
mean excystation efficiency of the control. * p<0.05 vs. E2E5 supernatant. ** p<0.01 vs. 20
E2E5 supernatant. 21
22
FIG. 2. Purification of 51 kDa antigen by affinity chromatography. Protein extracts from 23
purified gametocytes (lane 1) and proteins eluted from the E2E5 affinity column (lane 2) were 24
separated by SDS-Page and silver stained. Positions of molecular weight marker bands are 25
indicated. 26
ACCEPTED

29
1
FIG. 3. Sequence alignment of GAM56-like proteins from E. tenella and E. maxima. Amino 2
acids showing identity to the sequence obtained from Edman sequencing of the NH2-terminus 3
are highlighted in grey. Invariable amino acid positions are marked by a ″*″, substitutions 4
rated as conservative or semi conservative by ClustalW using the GONNET 250 matrix are 5
marked by ″:″ and ″.″, respectively. 6
7
FIG. 4. Identification of E2E5-binding phagemid clones by Western blotting. Protein lysates 8
of bacterial cultures were separated by SDS page. As a positive control (Pos), a protein 9
extract from E. tenella infected caecum at 137 h p.i. was used. The negative control (Neg) 10
consisted of a bacterial culture containing the empty pG8SAET phagemid vector. 11
12
FIG. 5. cDNA and deduced protein sequence of EtGAM22. The NH2-terminal signal peptide 13
is shaded in light grey, the sequence corresponding to the original phage display clone in dark 14
grey. 15
16
FIG. 6. Genomic organization of Etgam22. (A) Schematic drawing showing one end of the 17
sequence contig 2257242.c007101021.contig1 containing two copies of the Etgam22 open 18
reading frame (Etgam22). Some of the predicted restriction fragments rely on the assumption 19
that at least one additional copy of Etgam22 can be found further upstream (indicated in light 20
grey on the left). (B) Genomic Southern blot using the enzymes BglI (lane 1), ClaI (lane 2), 21
KpnI (lane 3), AccI (lane 4), DraI (lane 5), and MvaI (lane 6). The blot was hybridized with 22
the Etgam22 cDNA probe indicated in (A). (C) Southern blot containing serial dilutions of 23
genomic DNA from E. tenella digested with DraI and Etgam22 cDNA in pcDNA3.1 24
linearized with XhoI. The lanes (from left to right) contained 1.7 µg (25,856,313 copies 25
assuming a genome size of 60 Mbp) 0.85 µg, 0.425 µg, 0.213 µg, 0.106 µg, 0.053 µg, and 26
ACCEPTED

30
0.027 µg of genomic DNA and 726 pg (100,000 copies), 363 pg, 181.5 pg, 90.7 pg, 45.4 pg, 1
22.7 pg, and 11.3 pg of plasmid DNA. The 6,629 bp band corresponds to the plasmid DNA 2
while the 995 bp band was shown in (B) to result from binding of the probe to DraI digested 3
genomic DNA. 4
5
Fig. 7: Expression analysis of Etgam22. (A) Reverse-Transcription PCR with primers specific 6
for Etgam22 and Etactin cDNAs. RNA was isolated from sporulated oocysts, from the 7
caecum of non-infected chicken (n.i.) or chicken infected with E. tenella for the indicated 8
times. The reactions contained reverse transcriptase and RNA (lanes 1), or were performed in 9
the absence of either the reverse transcriptase (lanes 2) or the RNA template (lanes 3). (B) 10
RNA and protein were isolated from the same samples of oocysts and infected caecae at the 11
indicated times using trizol. With these matched RNA and protein samples, Northern blot 12
analyses of Etgam22 and Etgam56 mRNA expression and Western blot analysis with E2E5 13
were performed. (C) In vitro translation of EtGAM22. Products of coupled in vitro 14
transcription and translation were separated by SDS-PAGE, transferred to nitrocellulose 15
membrane and EtGAM22 was detected using the antibody E2E5 in Western blot analysis. 16
The negative control reaction contained no plasmid DNA. 17 ACCEPTED

ACCEPTED
ACCEPTED

EmGAM56 MTRLGLAA--VALALAVGPSMAVPSTTPVENQVHPYSEMSTYQEGSAPGAP-EDTTTTTT 57
EtGAM56 MTRLSLCA--LAVALAVGQSLAVP--TTVENTVHPYSEMGTYQEGEAPGAPDESSTTTTT 56
EtGAM59 MTRLAACAALAAVALAAGQSVAMPTAIPVEVPMDEYGQAKTYQDTDIP---------TST 51
****. .* *:***.* *:*:* .** :. *.: ***: . * *:*
EmGAM56 SSPVSDGAEQWLESFVRAVQRQLQLQDQMMRQLMRDIQEYLSTAFNWAENQSTAYTRVTE 117
EtGAM56 PSPSPEAPDQWLENFVRAVQRQLQLQESMMRQLVKEIQEYLSRAFNWDENQSAAYNRVNE 116
EtGAM59 STAAPESPQQWFEGFSRAVQKQLQLQENLMRQLVRDIQEYLTEAFGWNESDSSALARVNT 111
.:. .:..:**:*.* ****:*****:.:****:::*****: **.* *.:*:* **.
EmGAM56 MMDMISNRMNAAMDSSNELMTTSDTTDPETLRRATRKYMKEVRVQDVLVDALWASLRGVQ 177
EtGAM56 MMDMITNRMTTALDGANELMATSETMDPETLRRATRKYMKEVRVQDVVVDSLWASLRGVQ 176
EtGAM59 MLDMISKRMAVTRDAANEISTT--ETEPQAVRDATRNFMKEVRVQDIVVDALWASLRGVQ 169
*:***::** .: *.:**: :* :*:::* ***::********::**:*********
EmGAM56 TAAWMNGVT-AIEKEETTPMASRAAEEFLHRMYHNLRAAGMSEEDVAKFIPRAEYN-PSE 235
EtGAM56 TSAWMSGVT-AVEKEETTPMAGRAAEEFMHRMYHNLRAAGMAEEDITRFMPKTEYTTPRE 235
EtGAM59 TSAWMSGVSSTIDERDVLAVANRAAEEFLARMYHNLRAAGVAEEDIVKYVPRTPADPSSS 229
*:***.**: ::::.:. .:*.******: **********::***:.:::*:: . .
EmGAM56 QSRNMGRKGRSFYYGG-YPSYYNSPYYSYS-SYPSYYNYSYPSYSYSS-YPS-YYRYSS- 290
EtGAM56 QTRNMGRKGR---YG--YGYSYGYPLYSYGYSYPSYS-YSYPYYSYPS-YSYPLYSYSYR 288
EtGAM59 EPRNMGKKGRSYGYGHGYGCGYSYPLYSYGCPYSSCG-YSYPFYASTWGYPSPLAWGYPS 288
:.****:*** ** * *. * ***. .*.* **** *: . *.
EmGAM56 YPYYNYSYPSY-YNYGSYPYYSYSS---YPSWYWRRLRSLATATCPDCPPLTTPSMIPTP 346
EtGAM56 YPSYSYSYPLYSYSSYSYPYYSYSYPYYSSSWYWRRLR---TASCPDCPPGVN--MPPTP 343
EtGAM59 HSSFYWRRLGAAACPDCAPAPAPEPEFIIPPTAFRGLQEEAMMGTPYANPMMGTPMMGTP 348
:. : : . * : . .. :* *: * . * * **
EmGAM56 PP------MMN--MMNTP---PPMANMMTSMMMNTPMVPPPR-------TLGT--EAMSL 386
EtGAM56 LS------PMNTPLAGTP---SPTTTPMMPPMMP-PMVPPTR-------TLGT--EPLTM 384
EtGAM59 YANPMMASPYTTPMMGTSNTNPTMATPYTNPTMGTPYTNPTMATPYTNPTMGTPYTNPTM 408
. . : .*. .. :. * * . *. *:** ::
EmGAM56 GLAPIGITGAPMTGFGVPPEFGPFGAEGIGLPTDALGSTPEM----TPFDPTTPYRTLAP 442
EtGAM56 GMGPMPGYGYPPM-MDTMPEFPPF-AEGMGYPTEPINSIPTMNNMDTPFENTTNYRNLAP 442
EtGAM59 GTPYTNSMGTPYTNSMGAPYTNSMGTPYTNPTMGAPYTNPTMATPYTNPAMGAPYTNPTM 468
* * * * .: : . . . : * * * : * . :
EmGAM56 -----MDLPPIPPPVFPETP-------MRP---------------PTPFGFGP------- 468
EtGAM56 -----IDMP----PFFPEAP-------MRPTPTPT--------PTPTPFGFG-------- 470
EtGAM59 GGPYTTPMAGQAYPAYPAAAAAGQRRSMGPTQGPMGRMGTSGSQYNSPAGYTGGYRGLSA 528
:. * :* :. * * :* *:
EmGAM56 ----------APVPPMPF- 476
EtGAM56 -----------PVPP---- 474
EtGAM59 FEAPEFFEPPMGIPSFGFE 547
:*.
ACCEPTED
ACCEPTED

1 caggacccca aaataaaatc aaaggctatc acactatttt acttcttaac cgtttactga
61 ggctacaaga acaagtttga agatgaggac tatcctagcc accctagtcg gtttcacagc
1 M R T I L A T L V G F T
121 ctgcgcagcc gttgctgcag acggagcacc tgagtatcct tctcagcttg cagttgaaat
13 A C A A V A A D G A P E Y P S Q L A V E
181 cgatccagaa gcgattattg cgatccagca agatgcaaac gccgacccac gtctcttttt
33 I D P E A I I A I Q Q D A N A D P R L F
241 cccactgagc gggcttgtct ccgccaaact tgccaaagtc tttcaaccca acatataccc
53 F P L S G L V S A K L A K V F Q P N I Y
301 aacccctcct agtccccaga caacttacca ctttcacctc catcctcatc cccattatcc
73 P T P P S P Q T T Y H F H L H P H P H Y
361 gcatcctcag ccaagttatc ctcatcctca accccatcat cctcatcctc atccttatca
93 P H P Q P S Y P H P Q P H H P H P H P Y
421 tcctcatcct catccccatc atcctcatcc tcatccccat caacatcctc atcgtcatcc
113 H P H P H P H H P H P H P H Q H P H R H
481 cgaccatcat ccccaccatc atcctcacca tcatcatcat gaacataatg ttcatgtgcc
133 P D H H P H H H P H H H H H E H N V H V 541 tcaacatcag cacgctcaac acaacggcca ccagaacaac ggtggcccag ctcattatca
153 P Q H Q H A Q H N G H Q N N G G P A H Y 601 ccatgactac cattttgcgc atcctcatca agagaaccag catcaccgcg aggaagagca
173 H H D Y H F A H P H Q E N Q H H R E E E
661 gcttaccgac atcaactaag ctattggtcg ggaattaagg tgcttagtct cagtagtcag
193 Q L T D I N -
721 tacagtacta ggctacgtct gagatcttca tggcaaagag gtaccagcca ccaagctgac
781 tcggctatgt tttattagac aaatttaaat ttaaagggtc ccagtttcag tctctgcagg
841 tctgcccctg aaagcacgag aggggcctaa agggtgattg gagctgcaaa tacagctgca
901 aatgcagctg caaagtgccg cttcaaaaaa gggacaggct tcccgccaaa atttttggat
961 catacctatc aatgcttcga gaaaacatag aaaacaaaag cactgaagaa cgttcatagt
1021 cggtagtttt aggggcatgc cgtgtgctaa aatcccatcg aaccttcagg tacacctgat
1081 cgttacgaag tacacaccac cggtcactct caacgcgcac cactagagcg agagctgctt
1141 cagggatgca gcgagatgtc gactcagagg tcctacatta aaggg(a)n
ACCEPTED

ACCEPTED
ACCEPTED




















