Wood decay by basidiomycetes: Extracellular tripartite membranous structures
10
Trans. Br. mycol. Soc. 8S (2), 257-266 (1985) [ 257 ] Printed in Great Britain WOOD DECAY BY BASIDIOMYCETES: EXTRACELLULAR TRIPARTITE MEMBRANOUS STRUCTURES By R. FOISNER, K. MESSNER, H. STACHELBERGER* AND M. ROHR Institut fur Biochemische Technologie und Mikrobiologie, TU-Wien, 1060 Wien, Getreidemarkt 9, Austria and * Institut!ur Angewandte Botanik, Technische Mikroskopie und Organische Rohstoffiehre, TU-Wien, 1060 Wien, Getreidemarkt 9, Austria Decay of pine-wood by the brown-rot fungi Poria placenta, Fomitopsis pinicola, Serpula lacrimans, Paxillus panuoides and the white-rot fungi Trametes versicolor, Trametes hirsuta, Sporotrichum pulverulentum was investigated with the transmission electron microscope. Material stained with ruthenium red showed extracellular, tripartite structures associated with all fungi throughout all stages of wood decay. These structures surrounded the intensely stained fungal cell wall, were partly embedded in an extracellular slime and were contiguous with the wood cell wall. Besides brown-rot fungi with filter paper activity S. pulverulentum showed similar tripartite structures, however these were branched and extended from the fungal cell wall into the lumen of the wood cell. The origin, formation and function of these extracellular structures are discussed. Most electron microscopic examinations of wood decay have utilized specimens fixed with an aldehyde or potassium permanganate, postfixed with osmium tetroxide and stained with uranyl acetate and lead citrate (Chou & Levi, 1971; Bravery, Parameswaran & Liese, 1974; Wilcox, Parameswaran & Liese, 1974; Hale & Eaton, 1981; Ruel, Barnoud & Eriksson, 1981). Messner & Stachelberger (1984a, b) and Messner et al. (1985) succeeded in detecting various patterns of wood decay with brown-rot and white-rot fungi by means of specimens fixed with glutaraldehyde, followed by osmium tetroxide and stained with uranyl acetate. With this method osmiophilic particles could be detected during decay in and around the fungal hypha as well as in the wood cell wall. The presence of these particles within the wood cell wall was characteristic of both brown-rot and white-rot fungi. The use of other stains was necessary in order to determine the role of the fungal cell wall and extracellular slime during wood decay. Ruth- enium red (Fletcher et al., 1961) had been used as a tracer to demonstrate diffusion barriers, pores and channels, and also to stain acidic mucopolysac- charides (Reimann, 1961; Leightley & Armstrong, 1980). This paper reports the results of an ultrastructural study of decay of pine wood by brown-rot fungi with and without filter paper activity (FPA, see Messner & Stachelberger, 1984a) and white-rot fungi stained with ruthenium red. 9 MATERIALS AND METHODS Pine sapwood was used in all experiments with the following as test fungi: Poria placenta (Fr.) Cooke; FPRL 280 (brown-rot fungus without FPA). Fomitopsis pinicola (Schwartz: Fr.) Karst.; TU-Wien (brown-rot fungi without FPA). Serpula lacrimans (Wulf.:Fr.) Schroet.; BAM 9 (brown-rot fungus with FPA). Paxillus panuoides (Fr.) Fr.; Hamburg 75-03-B (brown-rot fungus with FPA). Trametes versicolor (L.: Fr.) Pil.; CTB 863A (white-rot fungus). T. hirsuta (Wulf.:Fr.) Pil.; TU-Wien (white-rot fungus). Sporotrichum pulverulentum Novobranova; MYCO 9145 (white-rot fungus). The cultivation of fungi on wood has been described in preceding papers (Messner & Stachel- berger, 1984a). Specimens were fixed for 4h in 2 % glutaraldehyde in 0'1 M sodium cacodylate buffer, pH 7'3, containing 0'01 % (wjv) ruthenium red at 4°. After fixation material was washed four times in cacodylate-buffer at room temperature, stained overnight and in the dark in a2 % (wjv) buffered solution of osmium tetroxide containing 0'01 % (wjv) ruthenium red at 4°. Fixed and stained material was dehydrated in a graded alcohol series and embedded in Spurr's low viscosity resin. Ultrathin sections were cut with a diamond knife MYC 85
Transcript of Wood decay by basidiomycetes: Extracellular tripartite membranous structures

Trans. Br. mycol. Soc. 8S (2), 257-266 (1985)
[ 257 ]
Printed in Great Britain
WOOD DECAY BY BASIDIOMYCETES: EXTRACELLULARTRIPARTITE MEMBRANOUS STRUCTURES
By R. FOISNER, K. MESSNER, H. STACHELBERGER* AND M. ROHRInstitut fur Biochemische Technologie und Mikrobiologie, TU-Wien, 1060 Wien, Getreidemarkt 9, Austriaand * Institut!ur Angewandte Botanik, Technische Mikroskopie und Organische Rohstoffiehre, TU-Wien,
1060 Wien, Getreidemarkt 9, Austria
Decay of pine-wood by the brown-rot fungi Poria placenta, Fomitopsis pinicola, Serpulalacrimans, Paxillus panuoides and the white-rot fungi Trametes versicolor, Trametes hirsuta,Sporotrichum pulverulentum was investigated with the transmission electron microscope.Material stained with ruthenium red showed extracellular, tripartite structures associated withall fungi throughout all stages of wood decay. These structures surrounded the intenselystained fungal cell wall, were partly embedded in an extracellular slime and were contiguouswith the wood cell wall. Besides brown-rot fungi with filter paper activity S. pulverulentumshowed similar tripartite structures, however these were branched and extended from thefungal cell wall into the lumen of the wood cell. The origin, formation and function of theseextracellular structures are discussed.
Most electron microscopic examinations of wooddecay have utilized specimens fixed with analdehyde or potassium permanganate, postfixedwith osmium tetroxide and stained with uranylacetate and lead citrate (Chou & Levi, 1971;Bravery, Parameswaran & Liese, 1974; Wilcox,Parameswaran & Liese, 1974; Hale & Eaton, 1981;Ruel, Barnoud & Eriksson, 1981). Messner &Stachelberger (1984a, b) and Messner et al. (1985)succeeded in detecting various patterns of wooddecay with brown-rot and white-rot fungi by meansof specimens fixed with glutaraldehyde, followedby osmium tetroxide and stained with uranylacetate. With this method osmiophilic particlescould be detected during decay in and around thefungal hypha as well as in the wood cell wall. Thepresence of these particles within the wood cell wallwas characteristic of both brown-rot and white-rotfungi. The use of other stains was necessary inorder to determine the role of the fungal cell walland extracellular slime during wood decay. Ruthenium red (Fletcher et al., 1961) had been used asa tracer to demonstrate diffusion barriers, pores andchannels, and also to stain acidic mucopolysaccharides (Reimann, 1961; Leightley & Armstrong,1980). This paper reports the results of anultrastructural study of decay of pine wood bybrown-rot fungi with and without filter paperactivity (FPA, see Messner & Stachelberger,1984a) and white-rot fungi stained with rutheniumred.
9
MATERIALS AND METHODS
Pine sapwood was used in all experiments with thefollowing as test fungi:Poria placenta (Fr.) Cooke; FPRL 280
(brown-rot fungus without FPA).Fomitopsis pinicola (Schwartz: Fr.) Karst.;
TU-Wien (brown-rot fungi without FPA).Serpula lacrimans (Wulf.:Fr.) Schroet.; BAM 9
(brown-rot fungus with FPA).Paxillus panuoides (Fr.) Fr.; Hamburg 75-03-B
(brown-rot fungus with FPA).Trametes versicolor (L.: Fr.) Pil.; CTB 863A
(white-rot fungus).T. hirsuta (Wulf.:Fr.) Pil.; TU-Wien (white-rot
fungus).Sporotrichum pulverulentum Novobranova; MYCO
9145 (white-rot fungus).The cultivation of fungi on wood has been
described in preceding papers (Messner & Stachelberger, 1984a). Specimens were fixed for 4 h in 2 %glutaraldehyde in 0'1 M sodium cacodylate buffer,pH 7'3, containing 0'01 % (wjv) ruthenium red at4°. After fixation material was washed four timesin cacodylate-buffer at room temperature, stainedovernight and in the dark in a 2 % (wjv) bufferedsolution of osmium tetroxide containing 0'01 %(wjv) ruthenium red at 4°. Fixed and stainedmaterial was dehydrated in a graded alcohol seriesand embedded in Spurr's low viscosity resin.Ultrathin sections were cut with a diamond knife
MYC 85

4100 omL..--.J
Wood decay by basidiomycetes
200 oml.---J
Figs 1-5. For captions see opposite
5

R. Foisner and others 259
and examined in a lEOL lOOC transmissionelectron microscope at 100 kV. Some specimenswere fixed with glutaraldehyde, osmium tetroxideand stained with uranyl acetate according to themethod described previously (Messner & Stachelberger, 1984a).
RESULTS
Brown-rot fungi without FPA: Poria placenta,Fomitopsis pinicola
The fungal cell wall becomes clearly visible ifstained with ruthenium red (Figs 1-3). The cellwall of Poria placenta is up to 300 nm thick with aninner electron dense, compact layer and an outerlayer which appears loose and slimy. After 4 daysof cultivation a membranous structure had formedon the surface of the fungal cell wall, adjacent to theloose, slimy layer (Fig. 1). These structures weretripartite and clearly visible on the surface of thefungal cell wall facing the lumen, in the comerformed between the wood cell wall and the fungalcell and spread over the surface ofthe wood cell wall(Fig. 1). These structures were present at all stagesof wood decay. Figure 2 illustrates that themembranous structures can be found over theentire surface of the fungal cell wall except wherethere is direct contact between the fungal and woodcell walls. It also shows that the membranousstructures extended into the comer between thehypha and the wood cell wall, which was frequentlyfilled with a slime, and later across the woodsurface. These structures often appeared fragmented on the surface of the fungal wall (Fig. 2).This, however, seems to be an artefact ofpreparation, as on the lumen surface thesemembranous structures often extended as continuous layers over a large region of the lumensurface (Fig. 3).
Fomitopsis pinicola showed similar structures,although these were observed less frequently.Similar structures were not found in wood that hadnot been attacked by fungi.
Brown-rot fungi with FPA: Serpula lacrimans,Paxillus panuoides
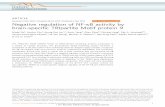
It was found that brown-rot fungi with FPAshowed the same membranous structures asbrown-rot fungi without FPA. They were observedon the fungal surface, in the comer between thehypha and the wood cell wall which was also witha slime and on the lumen surface (Figs 4, 5).Figures 4 and 8 show the similarity between thefungal plasmalemma and the extracellular membranous structure with regard to their dimensionsand tripartite organization. In brown-rot fungiwith FPA these membranous structures occurredmost frequently between 10 and 30 days after cultivation and often extended from the fungal cell walleither into the lumen or across the wood surface(Figs 6-8). The time of occurrence of the membranous structures corresponded to a weight loss of10 to 30 % of dry weight. After 15 days of cultivation of Serpula lacrimans several parallel membranous structures, surrounded by the outer slimelayer of the fungal cell wall, were found to extendfrom the hypha and over the wood surface (Fig. 6).With Paxi//us panuoides the membranous structures could be observed after 10 days of cultivation(Figs 7, 8). Some of these extended as continuouslamellae and some as shorter lengths into the lumenembedded in an extracellular slime (Figs 7, 8). Inbrown-rot fungi the membranous structures werealso present in material fixed with osmiumtetroxide and stained in uranyl acetate (Figs 9, 10).They were, however, hard to discern (Fig. 9).
White-rot fungi: Trametes versicolor, Trameteshirsuta, Sporotrichum pulverulentum
The cell walls of white-rot fungi appeared slightlymore electron-dense than that of brown-rot fungi(Figs 11, 12). During wood decay white-rot fungialso possessed membranous structures, both closeto the fungal cell wall and over the lumen surface.While membranous structures of brown-rot fungi
Fig. 1. Poria placenta, 4 d ofcultivation. An extracellular membranous structure appears on the outer surfaceof the hyphal cell wall, in the cell comer and on the wood cell wall.Fig. 2. Poria pkuenta, 8 d ofcultivation. The extracellular tripartite membranous structure covers the hyphaexcept where it is in contact with the wood cell wall.Fig. 3. Poria placenra, 8 d ofcultivation. The extracellular membranous structure extends from the cell comerand over the lumen surface.Fig. 4. Serpula lacrimans, 30 d of cultivation. Extracellular structures embedded in slime occur close to theouter surface of the hyphal cell wall and in the cell comer. Note the similarity of the plasmalemma to theextracellular membranous structure.Fig. 5. Serpula lacrimans, 15 d of cultivation. The extracellular membranous structure extends from the cellcomer and over the wood cell wall.
260 Wood decay by basidiomycetes
8
10
.l~~:~:;::r.;;~;---.:..---.:.::.-.-OP
.;~/"......... •••.• EMS
. ';-'j:.~ \-A ""f It. •
o· -. .,......~• .-WCW
300 nmL...--....J
.' .9
......
\EMS
I
r ..
Figs. ~10. For captions see opposite.

R. Foisner and others 261
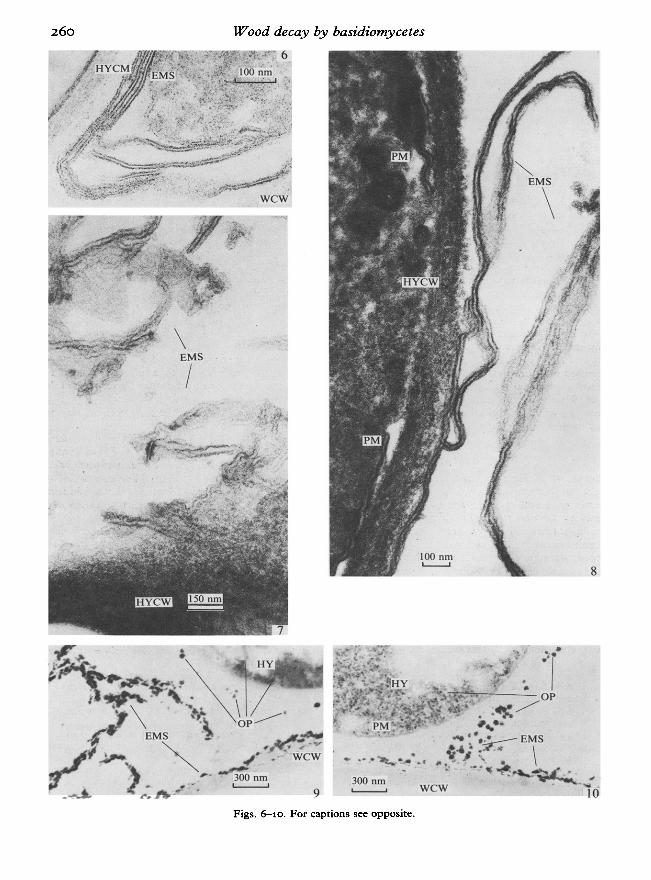
only extend across the lumen surface followingcontact between the hypha and the wood cell wall(Figs 1, 2, 4, 5), those of white-rot fungi extenddirectly from the surface ofthe fungal hypha facingthe lumen to the wood cell wall, the warty layer ofwhich is retained (Figs 11, 12). Several fungalhyphae were often surrounded by one continuousmembranous structure (Figs 11, 13). Membranousstructures were also found associated with Trametessp. (Figs 14, 15) (Messner et al., 1984b). Althoughthe membranous structures can normally beobserved 10 days after cultivation, in Trametesversicolor, which attacks wood very slowly (after120 days ofcultivation a loss in weight of4 % of dryweight) their occurrence is delayed.
In the initial stagesofwood decay by Sporotrichumpulverulencum a dense network of extracellularmembranous structures extended from the fungalcell wall into the lumen (Figs 16, 17). Later in decaythe membranous structures spread over more thanhalf of the wood lumen either as continuous orfragmented lamellae which were sometimes surrounded by a slime. In advanced stages of wooddecay fungal hyphae were rarely found althoughramified membranous structures often with associated slime, occurred throughout the lUmen andover the wood surface. Membranous structureswere also found in regions of the wood cell wall thathad been decomposed.
DISCUSSION
The use of ruthenium red allowed the fungal cellwall, an extracellular slime, the plasmalemma,membranes and extracellular membranous structures to be observed. The fungal cell walls ofbrown-rot and white-rot fungi are very differentfrom each other. The cell walls of white-rot fungiappear strongly electron-dense and compact (Figs11, 12), whereas those of brown-rot fungi are about100 to 200 nm thicker with an outer less electrondense, amorphous layer (Figs 1-5). The membranous structures adjacent to the fungal cell walland which extended over the wood surface (Figs1-5,11-13), or into the lumen (Figs 7, 8, 16, 17),were conspicuous. Membranous structures were
detected with both white-rot and brown-rot fungiat all stages of wood decay. With brown-rot fungiwith FPA and the white-rot fungus Sporotrichumpulverulentum the membranes were occasionallyhighly branched (Figs 7, 8, 16, 17). The similarityof these membranous structures with the plasmalemma is obvious (Figs 2-5, 8). The tripartiteorganization of the plasmalemma as well as that ofthe extracellular membranous structures are clearlyvisible owing to an increase in thickness caused bythe stain. Liese (1966), Schmid & Baldermann(1967), Chou &Levi (1971), Leightley & Armstrong(1980) and Leightley (1981) described partlyfibrillar and partly granular extracellular slimelayers associated with wood decomposing fungi.They identified these layers as acidic mucopolysaccharides. Tripartite extracellular structureswere not reported. Hale & Eaton (1981) examinedsoft-rot decay of birch caused by Monodiccysputredinis (Wallr.) Hughes (Deuteromycotina) atthe transmission electron microscope level. Theyshowed a protrusion of the plasmalemma whichextended from a bore hypha across an electrontransparent region of the wood cell wall. However,bore hyphae possess very thin cell walls. Furthermore, they described in the bore hypha a •doublemembrane' which extended subapically from aregion with a mesosome-like structure across thesurface of the hypha.
From studies of shadowed surface replicas Liese& Schmid (1962) and Schmid & Liese (1964)described extracellular fibrillar elements associatedwith wood decomposing fungi. They assumed thatthe fibrillar network was identical with a thin filmwhich was found to cover hyphae in thin sectionedmaterial (Liese & Schmid, 1963).
This paper shows that the extracellular membranous structures which surround the hyphae,especially with white-rot fungi, form a continuouslayer over both the hyphae and wood surfaces (Figs11-13). This may give the impression that thefungal hypha grows under a covering membraneand may have led Meier (1955) to the assumptionthat the fungus grows under a layer of the wood cellwall.
For the following reasons and based on the
Fig. 6. Serpula lacrimans, 15 d of cultivation. Extracellular membranous structures, embedded in slime,adjacent to the hyphal wall and extended over the wood cell wall.Fig. 7. Paxillus panuoitks, 10 d of cultivation. The extracellular membranous structures extend from thehyphal cell wall into the lumen and are embedded in a slime.Fig. 8. Paxillus panuoides, 10 d of cultivation. The extracellular membranous structures have separated fromthe hyphal wall and extend into the lumen. Note the similarity of the plasmalemma to the extracellularmembranous structures.Figs 9, 10. Serpula lacrimans, 4 d of cultivation. Not stained with ruthenium red. The extracellularmembranous structures in the lumen and on the wood cell wall are only visible owing to the presence ofosmiophilic particles.
Wood decay by basidiomycetes
13wcw
-.'
cc
-----------11
)t.,'
...~.~
/;:' 150 om...' ~
EMS
12wcw
wcw
200 omL...--J
200 om~
Figs. 11-13. For captions see page 264.

500 omL..--....l
wcw
R. Foisner and others
Figs. 14-17. For captions see overleaf.
wcw
17
14

Wood decay by basidiomycetesresults presented in this paper it is concluded thatthese extracellular membranous structures areproduced by the fungus: (a) They are found abovethe warty layer, which is the outermost layer of thewood cell wall (Figs 11, 12), (b) they occur onfungal hyphae outside the wood, (c) in woodnon-infected by fungi these structures were notdetected.
The formation, nature and function of theseextracellular membranous structures, remainobscure. Ruthenium red is a particularly good stainfor acidic mucopolysaccharides (Lewis & Knight,1977) but as Figs 2-5, 8, 12 and 13 illustrate,ruthenium red also stains biomembranes. Correspondingly, based on their staining, the extracellularmembranous structures may be composed of eitheracidic mucopolysaccharides or lipo- and glycoproteides. However, owing to their tripartite structureit is improbable that these structures consist ofacidic mucopolysaccharides.
The following hypothesis is presented for theformation of the extracellular membranes: lipoand glycoproteides are excreted from the wooddecay fungi to form biomembranes on the hyphalsurface. Wood decomposed by brown-rot fungi hasan average moisture content of ca 60 %, whereas inthat attacked by white-rot fungi it is nearly 100%.This means that the point of fibre saturation,30-40 %, is exceeded and the surplus moisturemust be present as a water film on the lumen surfaceand around the hyphae. Correspondingly thelipo-proteides could selfassemble on this water filmto form biomembranes. However, the compositionof the extracellular membranes seems differentfrom that of the intracellular biomembranes, sincethey are not stained as intensely as intracellularmembranes by osmium tetroxide and uranylacetate.
I t is unlikely that these extracellular membranousstructures are products of wood cell wall decom-
position as they appear at the early stages of fungalcolonization and prior to decay. In addition thetripartite structures have no similarity to woodelements and can also be found on hyphae outsidethe wood.
A further possibility is that these extracellularmembranous structures consist of regularly arranged proteinaceous macromolecules similar tothose reported for some bacteria (Glauert &Thomley, 1969). Beveridge & Murray (1974,1975,1976) and Beveridge (1979) found two additionalsuperficial layers consisting of regularly arrangedproteins with Spirillum putridiconchylium Terasaki,Spirillum metamorphum Terasaki and Sporosarcinaureae (Beijerinck) Kluyver & van Niel, whichclosely surround the outermost layer of thebacterial wall. They reported that these superficiallayers were very difficult to examine and to analyzewith the electron microscope, as during preparationthey were easily sloughed off the bacterial wall orlost their ordered structure. By the addition ofruthenium red to the staining solutions theysucceeded in resolving these ordered structures insectioned material examination with the transmission electron microscope. Previously theseworkers had only found the superficial layersin freeze-fractured and the negatively stainedmaterial. It was assumed that ruthenium red actedas a polyvalent cation to stabilize the regularlystructured layers. They further found that theseordered structures, once sloughed off the bacterialcell wall, spontaneously reassembled into regularlystructured complexes, preferentially at an air liquidinterface or again at the bacterial cell wall. It maybe that these ordered surface arrays protect themicro-organism from environmental stress factors,i.e. low pH, high temperature, high salinity, or thatthey form compartments to concentrate the substrate for the organism.
At the moment it is only possible to speculate on
Fig. 11. Trametes versicolor, 120 d of cultivation. A continuous extracellular membranous structure coverstwo hyphae. The warty layer, which is the outermost layer of the wood cell wall, is clearly visible. Note therelatively large cell comer compared to that of the brown-rot fungi.Fig. 12. Trametes versicolor, 120 d of cultivation. The extracellular membranous structure extends from thehyphal surface to the wood cell wall and forms a large cell corner.Fig. 13. Trametes hirsuta, 13 d of cultivation. The extracellular membranous structure covers two hyphaeand extends to the wood cell wall to form a large cell corner.
Fig. 14. Trametes versicolor, 120 d of cultivation. The extracellular membranous structure also cover the cellwall systems typical of Trametes sp.Fig. 15. Trametes versicolor, 120 d ofcultivation. Detail ofa hyphal cell wall system covered by an extracellularmembranous structure.Fig. 16. Sporotrichum pulverulentum, 5 d of cultivation. Showing a system of extracellular membranousstructures embedded in slime.Fig. 17. Sporotrichum pulverulentum, 5 d of cultivation. Higher magnification of Fig. 16. Note the existenceof extracellular membranous structures at places of decay (arrowed).

R. Foisner and othersthe function of the extracellular membranousstructures associated with wood-decomposingBasidiomycotina. Besides protection from environmental stress and concentration of substrates,they may be associated with membrane-boundedenzymes.
The question as to whether and to what extentthere is a connexion between these extracellularmembranous structures and the appearance of theosmiophilic particles during the process of wooddecay as was described in earlier papers (Messner& Stachelberger, 1984a,b; Messner el al., 1985) isunresolved. The fact that the osmiophilic particlesas well as the membranous structures appeararound the hyphal cell wall and extend across thelumen surface of the wood cell is nevertheless moststriking.
REFERENCES
BEVERIDGE, T. J. (1979). Surface arrays on the cell wall ofSporosarcina ureae. Journal of Bacteriology 139,103!r1048.
BEVERIDGE, T. J. & MURRAY, R. G. E. (1974). Superficialmacro-molecular arrays of the cell wall of Spirillumputridiconchylium. Journal of Bacteriology 119,101!r1038.
BEVERIDGE, T. J. & MURRAY, R. G. E. (1975). Surfacearrays on the cell wall of Spirillum metamorphum.Journal of Bacteriology 1%4, 152?-1544.
BEVERIDGE, T. J. &MURRAY,R. G. E. (1976). Reassemblyin vitro of the superficial cell wall components ofSpirillum putridiconchylium. Journal of UltrastructuralResearch 55, 105-118.
BRAVERY, A. F., PARAMESWARAN, M. & LIESE, W. (1974).Electron microscopic investigation on the effect oftri-n-butyl tin oxide on the decay of beechwood byPolystietus versicolor. Material und Organismen 9,133-143·
CHOU, C. K. & LEVI, M. P. (1971). An electronmicroscopical study of the penetration and decomposition of tracheid walls of Pinus sylvestris by Poriavaillantii. Holzforschung 35, 107-112.
FLETCHER, J. M., GREENFIELD, B. F., HARDY, C. J.,SCARGILL, D. & WOODHEAD, J. L. (1961). Rutheniumred. Journal of the Chemical Society 3, 3000-2006.
GLAUERT, A. M. & THORNLEY, M. J. (1969). Thetopography of the bacterial cell wall. Annual Review ofMicrobiology 33, 15?-198.
HALE, M. D. & EATON, R. A. (1981). Soft rot ultrastructure. The International Research Group on WoodPreservation, Document No.: IRGjWPj1138.
LEIGHTLEY, L. E. (1981). Soft rot studies on CCA treatedeucalypt power transmission poles. The InternationalResearch Group on Wood Preservation, Document No.:IRGjWP/1132.
LEIGHTLEY, L. E. & ARMSTRONG, W. (1980). Ultrastructural details of the soft rot decay of Eucalyptusmaculata Hook. by Phialophora mutabilis (van Beyma)Schol Schwarz. Micron 11, 513-514.
LEWIS, P. R. & KNIGHT, D. P. (1977). Staining methodsfor sectioned material. In Practical Methods in ElectronMicroscopy V (ed. A. M. Glauen), pp. 62~6. Amsterdam-New York-Oxford: North-Holland.
LIESE, W. (1966). Mikromorphologische Veranderungenbeim Holzabbau durch Pilze. Holz und Organismen,International Symposium, Berlin-Dahlem, 1965.Material und,Organismen, Suppl. 1, 13-26.
LIESE, W. & SCHMID, R. (1962). Elektronenmikroskopische Untersuchungen iiber den Abbau des Holzesdurch Pilze. Angewandte Botanik 36, 29'1-298.
LIESE, W. & SCHMID, R. (1963). Fibrillare Strukturenan den Hyphen holzzerstorender Pilze. Naturwissenschaften 50, 103-110.
MEIER, H. (1955). Uber den Zellwandabbau durchHolzvermorschungspilze und die submikroskopischeStrukturvon Fichtentracheiden und Buchenholzfasern.Holz als Roh und Werkstojj 13, 323-338.
MESSNER, K., FOISNER, R., STACHELBERGER, H. & R6HR,M. (1985). Osmiophilic panicles as a typical aspect ofthe brown and white-rot system in transmissionelectron microscope studies. Transactions of the BritishMycological Society 84, 457-466.
MESSNER,K. & STACHELBERGER,H. (1984a). Transmissionelectron microscope observations of brown rot causedby Fomitopsis pinicola with respect to osmiophilicparticles. Transactions ofthe British Mycological Society83,113-130.
MESsNER,K. & STACHELBERGER, H.(1984b). Transmissionelectron microscope observations ofwhite rot caused byTrametes hirsuta with respect to osmiophilic particles.Transactions of the British Mycological Society 83,20?-216.
REIMANN, B. (1961). Zur Verwendbarkeit von Rutheniumrot als elektronenmikroskopisches Kontrastierungsmittel. Mikroskopie 16, 224-226.
RUEL, K., BARNotJD, F. & ERIKSSON, K. E. (1981).Micromorphological and ultrastructural aspects ofspruce wood degradation by wild type Sporotrichumpulverulentum and its cellulase-less mutant cell 44.Holzforschung 35,157-171.
SCHMID, R. & BALDERMANN, E. (1967). Elektronenoptischer Nachweis von sauren Mucopolysacharidenbei Pilzhyphen. Naturwissenschaften 19,521-5:22.
SCHMID, R. & LIESE, W. (1964). Ober die mikromorphologischen Veranderungen der Zellwandstrukturen von Buchen und Fichtenholz beim Abbau durchPolyporus versicolor (L.) Fr. Archivfur Mikrobiologie 47,26cr-276.
WILCOX, W. W., PARAMESWARAN, N. & LIESE, W. (1974).Ultrastructure of brown rot in wood treated withpentachlorophenol. Holzforschung ~8, 211-217.
(Received for publicalion J4 April J983)

266 Wood decay by basidiomycetes
LIST OF ABBREVIATIONS USED IN LEGENDSOP osmiophilic panicles.PM plasmalemma.WCW wood cell wall.WL wany layer of wood cell wallAll Figures are transmission electron micrographs.
cell comer.cell wall system.extracellular membranous structure.hypha.hyphal cell wall.
CCCWSEMSHYHYCW