
Tubulin assembly probed with antibodies to synthetic peptides
16
J. Mol. Biol. (1990) 214, 105-120 Tubulin Assembly Probed with Antibodies to Synthetic Peptides Maha Angeles Arbvalo’, Juan Miguel Nieto’, David Andreu’ and Josh Manuel Andreu’ ‘Centro de Investigaciones Bioldgicas, C.S.I.C. Veldzquez 144, 28006 Madrid, Spain “Department0 de Q&mica Orgdnica, Universidad de Barcelona Marti i Franque’s 1, 08028 Barcelona, Spain (Received 30 October 1989; accepted 5 March 1990) Antibodies to synthetic peptides from the u and #?-tubulin sequences were employed to study zones of this protein active in microtubule assembly. In purified calf brain tubulin, six short sequences, selected according to their hydrophilicity and conservation, were found to be accessible to their affinity-purified immunoglobulin G (IgG) antibodies, in a competition radioimmunoassay performed under non-assembly native conditions. This indicated that the six sequences arc exposed on the surface of the tubulin UP heterodimer. I@;G antibodies to the a(430-443) and &412-431) sequences perturbed substoichiometrically the assembly of purified tubulin, inducing microtubule bundling and the formation of opened up structures. These positions, which are close to the C termini, were accessible to the anti-peptide antibodies in taxol-induced microtubules, Zn2+ -induced tubulin sheets, Mg2+ -induced tubulin rings and in PtK2 cell microtubules. This, together with the comparison of the sizes and gross shapes of the antibody probes and microtubules, suggested that these sequences might be located at the protruding parts of the protofilaments. Antibodies to positions a( 155-168) did not react with microtubules, while the equivalent zone 8(153-165) was accessible. The ~(214-226) and /?(241-256) sequences were antigenically occluded in the taxol microtubules, Zn2+-induced sheets and Mg’+-induced ring arrays, as well as in native microtubules from PtK2 cells, though they became reactive by fixation. This result strongly suggested that these two zones are close to tubulin-tubulin contact sites. A working model is proposed in which the positions a(214-226) and fi(241-256) are close to the axial contacts between heterodimers, which lead to protofilament formation, while the positions a(241-256) and &214-226) are suggested to be related to the a-p binding interface within the heterodimer. 1. Introduction The structure of microtubules is known, to low resolution, from electron microscopy and image reconstruction (Amos, 1979; Amos & Eagles, 1987; Mandelkow & Mandelkow, 1985) and X-ray fibre diffraction studies (Mandelkow et al., 1977; Beese et al., 1987; Wais-Steider et al., 1987). However, tubulin crystals suitable for high-resolution X-ray diffraction structural determination have not been reported. Therefore the intermolecular interactions responsible for the assembly of tubulin into micro- tubules are essentially unknown. Hydrophobic interactions are involved in tubulin self-assembly (Lee & Timasheff, 1977; Hinz et al., 1979; Andreu, 1982; Andreu et al., 1986). Ionic interactions are clearly involved in microtubule assembly, since the proper divalent cations are required (Weisenberg, 1972), the polymerization is very sensitive to pH and ionic strength (Lee & Timasheff, 1977), and the acidic carboxy-terminal regions of a and /I-tubulin play an important role in the regulation assembly (Serrano et al., 1984). It is accepted that the extreme carboxy-terminal regions of both subunits are exposed to the solvent in microtubules (Blose et aZ., 1984; Breitling & Little, 1986; Wehland et al., 1983; Sackett & Wolf, 1986). Limited proteolysis and subunit-specific anti- bodies were employed to study the domain struc- ture of tubulin (Mandelkow et al., 1985). It was proposed from proteolysis and crosslinking results that the large N-terminal fragment of a-tubulin 0022-2S36/90/130105-16 $03.00/O 105 0 1990 Academic Press Limited
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Tubulin assembly probed with antibodies to synthetic peptides

J. Mol. Biol. (1990) 214, 105-120
Tubulin Assembly Probed with Antibodies to Synthetic Peptides
Maha Angeles Arbvalo’, Juan Miguel Nieto’, David Andreu’ and Josh Manuel Andreu’
‘Centro de Investigaciones Bioldgicas, C.S.I.C. Veldzquez 144, 28006 Madrid, Spain
“Department0 de Q&mica Orgdnica, Universidad de Barcelona Marti i Franque’s 1, 08028 Barcelona, Spain
(Received 30 October 1989; accepted 5 March 1990)
Antibodies to synthetic peptides from the u and #?-tubulin sequences were employed to study zones of this protein active in microtubule assembly. In purified calf brain tubulin, six short sequences, selected according to their hydrophilicity and conservation, were found to be accessible to their affinity-purified immunoglobulin G (IgG) antibodies, in a competition radioimmunoassay performed under non-assembly native conditions. This indicated that the six sequences arc exposed on the surface of the tubulin UP heterodimer.
I@;G antibodies to the a(430-443) and &412-431) sequences perturbed substoichiometrically the assembly of purified tubulin, inducing microtubule bundling and the formation of opened up structures. These positions, which are close to the C termini, were accessible to the anti-peptide antibodies in taxol-induced microtubules, Zn2+ -induced tubulin sheets, Mg2+ -induced tubulin rings and in PtK2 cell microtubules. This, together with the comparison of the sizes and gross shapes of the antibody probes and microtubules, suggested that these sequences might be located at the protruding parts of the protofilaments.
Antibodies to positions a( 155-168) did not react with microtubules, while the equivalent zone 8(153-165) was accessible. The ~(214-226) and /?(241-256) sequences were antigenically occluded in the taxol microtubules, Zn2+-induced sheets and Mg’+-induced ring arrays, as well as in native microtubules from PtK2 cells, though they became reactive by fixation. This result strongly suggested that these two zones are close to tubulin-tubulin contact sites. A working model is proposed in which the positions a(214-226) and fi(241-256) are close to the axial contacts between heterodimers, which lead to protofilament formation, while the positions a(241-256) and &214-226) are suggested to be related to the a-p binding interface within the heterodimer.
1. Introduction
The structure of microtubules is known, to low resolution, from electron microscopy and image reconstruction (Amos, 1979; Amos & Eagles, 1987; Mandelkow & Mandelkow, 1985) and X-ray fibre diffraction studies (Mandelkow et al., 1977; Beese et al., 1987; Wais-Steider et al., 1987). However, tubulin crystals suitable for high-resolution X-ray diffraction structural determination have not been reported. Therefore the intermolecular interactions responsible for the assembly of tubulin into micro- tubules are essentially unknown.
Hydrophobic interactions are involved in tubulin self-assembly (Lee & Timasheff, 1977; Hinz et al., 1979; Andreu, 1982; Andreu et al., 1986). Ionic
interactions are clearly involved in microtubule assembly, since the proper divalent cations are required (Weisenberg, 1972), the polymerization is very sensitive to pH and ionic strength (Lee & Timasheff, 1977), and the acidic carboxy-terminal regions of a and /I-tubulin play an important role in the regulation assembly (Serrano et al., 1984).
It is accepted that the extreme carboxy-terminal regions of both subunits are exposed to the solvent in microtubules (Blose et aZ., 1984; Breitling & Little, 1986; Wehland et al., 1983; Sackett & Wolf, 1986). Limited proteolysis and subunit-specific anti- bodies were employed to study the domain struc- ture of tubulin (Mandelkow et al., 1985). It was proposed from proteolysis and crosslinking results that the large N-terminal fragment of a-tubulin
0022-2S36/90/130105-16 $03.00/O 105
0 1990 Academic Press Limited

M. A. Arkvalo et al.
(containing the non-exchangeable nucleotide- binding site) contacts the small C-terminal fragment of /?-tubulin in the a-fi-heterodimer, while the large N-terminal part of ,&tubulin (containing the exchangeable nucleotide-binding site) contacts the small C-terminal part of a-tubulin in the protofila- ments of microtubules (Kirchner & Mandelkow, 1985).
We employed five site-directed antibodies gener- ated from synthetic peptides, in combination with limited proteolysis, to study the substructure of tubulin in its a-fi-heterodimer, unassembled state. This approach revealed that each tubulin monomer is made up of three major types of limited proteoly- sis fragments (de la Viiia et aE., 1988).
Here, we report the result of probing the assembly of tubulin into microtubules with seven monospecific antibodies. Three small, chemically defined surface zones of tubulin have been identi- fied, which are occluded by assembly.
2. Materials and Methods (a) Chemicals and proteins
Chemicals, proteases and immunochemical reagents were as described (de la Vifia et al., 1988), except where indicated. Taxol was kindly provided by Dr Matthew Suffness (National Institutes of Health, Bethesda, MD). Fixed Staphylococcus aureus Cowan 1 cells were a gift from Dr Isabel Gil (Centro de Investigaciones Biologicas, Madrid). ‘Z51-labelled protein A was prepared as described (Andreu et a,Z., 1988). Circular dichroism spectra were obtained as described (Andreu et aZ., 1986). Tubulin was purified from bovine brain according to the Weisen- berg procedure (Lee et al.. 1973; Andreu & Timasheff, 1982) and assembled in 10 m&r-sodium phosphate, 16 m&r-MgCl,, 3.4 M-glycerol, 1 mlrr-EGTA, 91 mM-GTP (pH 7.0). Bovine brain microtubule protein was prepared by 2 cycles of polymerization (Karr et al., 1982) and assembled in 100 mM-Mes, 1 m&r-MgCl, 1 mM-EGTA, 1 m&r-2-mercaptoethanol, 1 mM-GTP (pH 65). Microtu- bules were adsorbed to carbon-coated grids, stained with 20/b (w/v) uranyl acetate and observed under a Philips EM 300 electron microscope.
(b) Synthetic peptides and anti-peptide antibodies
Peptides ERLSVDYGKKSKLEC and SKIREEEYP- DRIMNC were synthesized by the reported solid phase procedures (Andreu et al., 1988) in an Applied Biosystems 430A synthesizer. Peptide-resins were cleaved and depro- tected in low/high HF (Tam et al., 1983). The purified peptides were homogeneous by high-pressure liquid chromatography and gave correct amino acid analyses and mass spectra. They were coupled to keyhole limpet haemocyanin (KLH) employing m-maleimido-benzoyl-N-
t Abbreviations used: KLH, keyhole limpet haemocyanin; BSA, bovine serum albumin: PBS, phosphate-buffered saline; Ig, immunoglobulin; PG buffer, 10 m&r-sodium phosphate, 91 m&r-GTP (pH 7.0): PMEG buffer, 10 miw-sodium phosphate, 6 m&r-MgCl,. 1 mm-EGTA, 0.1 mM-GTP (pH 65); MAP, microtubule- associated protein; PEM buffer. 100 miw-piperazine-N,N’-bis(2-ethane sulphonic acid), 1 mM EGTA, 1 mM-MgCl, (pH 68).
hydroxysuccimide ester (Lin et al., 1979: (+reen uf al.. 1982). Anti-peptide antisera were obtained from rabbits injected with 0.5 mg peptide-carrier conjugate/immuniza,- tion, and were assayed by enzyme-linked immunosorbent assay and immunoblotting as described (Andreu Pt al.. 1988). The peptides DRIRKLADQCTG and VIRKE- SESSDC (corresponding to the tubulin sequence positions ~((120-131) and /?(119-129)), employed on t,hr KLH carrier or as bovine serum albumin (BSA)-glutaraldehvdr conjugates, generated anti-peptide antibodies that failed to react with native and denatured tubulin. The Tg(: fractions of antisera (Campbell, 1984) were purified by affinity chromatography on t,ubulin-Sepharose columns (eluted with 50 mM-glyCine. 05 M-NaCl (pH 10.5). neutralized immediately and dialysed against phosphan- buffered saline (PBS)). giving antibody preparations in which only their heavy chain and light chains were detected by SDS/polgacrplamide gel rlectrophoresir. Limited prot,rolysis of tubulin and immunoblotting were performed as described (de la Viria et al.. 1988).
((2) Radioimmunoassay
Tubulin (20 mg/ml in IO mM-sodium phosphat,e huller (pH 7.0)) was treated wit,h 1 o/0 [w/v) SDS at room temperature. Excess detergent was removed by Sephadex G-25 chromatography and the denatured prot,ein was loaded onto a 0.9 cm x 23 cm Sephacryl S300 column equilibrated with the same buffer. A single peak was obtained, which eluted in a position close to that of nativtl tubulin. The more concentrated protein fractions could be stored at - 20°C and gave the same Sepharryl S300 elution profile when they were rechromatographed. This SDS-treated tubulin, not significantly aggregated and in a stable state of denaturation, was iodmat,ed by the Hunt,er & Greenwood (1962) procedure. with modifications (Andreu et al., 1988). [iz51]tubulin had a typica,] specific activity of approx. 26,000 disints/min per ng (03 lz51 atoms/tubulin chain) and evenly labelled a and /?-chains. as observed by autoradiography of SDS/polyacr,ylamide gels.
Radioimmunoassay was carried out essentially as described by Van de Water $ Olmst,ed (1980) in PG buffer (see abbreviations) at 0°C. The experimental back- ground, determined by substitution of preimmune IgG for the antibodies, was less than 10% of the tot’al radioacti- vity, and was subtracted from the data. To measure the binding of [ J 125I tubulin to the anti-peptide antibodies. antibody concentrations giving approx. 509;, of maximal binding in the assay above were titrated with varying c*oncentrations of ] ‘251]tubulin. Both bhr free a,nd bound ligand ([‘25T]tubulin) concent’rations were measured in the supernatant and pellet, respectively. aft’er t,hr lg(: precipitation with 5’. aureus. Isotopic dilution of [1251]tubulin with unlabelled. SDS-treated, tubulin did not modify the net data, indicating that the radiolabelled ligand and the unlabelled one were immunochemically undistinguishable. Competition radioimmunoassay was performed by simultaneous incubation of t)he competitor (native or denatured t)ubulin) and the ligand with t.he antibody.
(d) Binding of antibodies to polymerized tuhulin
Tubulin (2 mg/ml in PMEG buffer (see abbreviations)) was centrifuged at 100,000 g (20 min at 4°C) and 20 pm-taxol was added to the supernatant, which was warmed to 37°C for 30 min. while the assembly of micro tubules was monitored by turbidimetry. Antibodies were

Microtubule Mapping by Monospecific Antibodies 107
diluted in 50 ~1 of PMEG buffer containing 10 mg gelatin/ ml at 37°C. mixed with an equal vol. of microtubule suspension and incubated for 30 min at 37°C. 1251-labelled protein A (40 ~1 containing 2 pg, 1,300,OOO disints/min) were added and incubated for an additional 15 min at 37°C. The reaction mixture was centrifuged at 10.000 g for 5 min. the pellets washed with a warm solu- tion of 309, sucrose and 1 mg gelatin/ml in PMEG buffer, and counted. The experimental background was deter- mined by substitution of preimmune IgG for t,he anti- bodies and subtracted from the data. Non-tax01 controls gave insignificant pellet’s.
Microt,ubules were fixed where indicated with tk25?/, (w/v) glutaraldehyde (15 min at’ room temperature) and the rea,ction stopped with 40 miw-sodium borohydride. Taxol-induced microtubules containing microtubule-asso- ciated proteins (MAPS) were assayed as above. employing microtubule protein in 100 mM-Mes. 1 mM-MgCl,, 1 mM-EGTA. 1 mM-2-mercaptoethanol, 1 miw-GTP (pH 6.5), instead of the purified tubulin.
To prepare Zn 2+-induced tubulin sheets (Gaskin & Kress. 1977), 2 mg purified tubulin/ml in 100 mM-Mes. 1 miw-GTP (pH 65) was centrifuged at 100,000 g (20 min at 4’C) and 0.3 mM-ZnCl, was added to the supernatant, which was warmed to 37°C for 30 min. Zn’+-induced sheet,s were observed by electron microscopy and assayed as described above.
To prepare ring aggregates, 2.7 mg/ml of a preparation of substilisin-cleaved tubulin M (a differential carboxy- terminal cleavage by whirh the ~(430-443) and /(412-431) zones arr not removed; M. A. Arevalo & ,J. M. Andreu. unpublished results) in 10 mw-sodium phosphate buffer (pH 7.0) was dialysed overnight (room tempera- ture. without st’irring) against 10 mM-sodium phosphate. -50 mM-h@04, til mM-GDP (pH 6.5). Ring aggregates were observed by electron microscopy and assayed as described above.
(e) Binding of antibodies to cellular microtubules
PtK2 cells were plated onto 9 mm x 9 mm glass cover- slips (Bellco) at a density of 200,000 cells/ml and cultured for 1 day in Dulbecco’s modified Eagle’s medium (Sigma) with lo:/, foetal calf serum (Flow) at 37°C in an humidi- tied atmosphere wit)h 5O/ I0 (v/v) CO,. The coverslips were washed with PEM (see abbreviations) containing 4% (v/v) polyeth~leneglycol 8000. The cells were lysed with 05?b (v/v) Tnton X-100. 1 mM-phenylmethanesulphonyl fluoride. and 1 rnM-GTP in PEM. for 90 s at room temperature, and the coverslips were washed with PEM. polyethyleneglycol, 1 mM-GTP (Anderson et al., 1982). The coverslip-attached cytoskeletons were saturated with 1 mg BSA/ml in the same buffer for 30 min at room temperature and then they were allowed to react with anti-peptide ant’ibodies for 30 min at room temperature in the same buffer.
The binding of the anti-peptide antibody to the unfixed. stabilized, microtubules was monitored by fluor- escence microscopy as follows. The cytoskeletons were washed with PEM. polyethyleneglycol, GTP (3 times, 10 min). fixed with methanol at -20°C for 10 min and transferred to PBS. The fixed cytoskeletons were incu- bated with monoclonal antibody reacting with the other tubulin chain (DMlA or DMlB; Blose et al., 1984), processed for immunofluorescence and photographed as described (Andreu et al., 1988). The secondary antibodies employed were goat anti-rabbit Ig fluoresceinated anti- bodies (Behring, Marburg, F.R.G.) and goat anti-mouse Ig rhodaminated antibody (Pu‘ordic, Tilburg, The
Xetherlands). The absence of undesirable crossreactions or significant spillover in the double immunofluorescence was verified by the pertinent controls.
Alternatively, the cytoskeletons were fixed with 3.7% (v/v) formaldehyde or with methanol (Osborn & Weber, 1982) and processed for immunofluorescence with the anti-peptide antibodies.
3. Results
(a) Antibodies to predetermined sites of the tub&n molecule
The monospecific antibodies raised against syn- thetic peptides corresponding to tubulin positions 1x(214-226), /?(241-256), a(415-443), ~((430-443) and p(412-431), which are conserved and hydrophilic, react specifically with the homologous tubulin zones (de la Vifia et al., 1988). Following the same stra- tegy, two new peptides were synthesized, ERLSV- DYGKKSKLEC and SKIREEYPDRIMNC, which are homologous to porcine brain tubulin sequence positions a( 155-168) and /?( 153- 165), respectively (Ponstingl et al., 1981; Krauhs et al., 1981). Tubulin- crossreactive antibodies were obtained from them (see Materials and Methods).
The results of limited proteolysis of tubulin and immunoblotting with these antibodies (M. A. Arevalo & J. M. Andreu, unpublished results) complemented the previous ones and confirmed that each tubulin monomer is preferentially cleaved at three zones, giving rise to three major types of limited prot’eolysis fragment (about l/3 of the chain length each) and small C-terminal fragments (de la Viiia et al., 1988). Positions ~(155-168) and fl( 153-165) are at the first proteolysis zone. They were left within the N-terminal fragment 1, the middle fragment 2, or cleaved, depending on the protease employed. Positions a(214-226) and p(241-256) are in the middle fragment 2 of each chain. Positions a(415-443) and fi(412-431) are in fragment 3 of each chain, adjacent’ to the third preferential proteolysis zone at’ the C terminus of both chains.
(b) Interaction of anti-peptide antibodies with native and denatured tubulin in solution
The IgG anti-peptide antibodies were purified by affinity chromatography on tubulin-Sepharose and their interaction with tubulin was examined by means of liquid-phase radioimmunoassay, employing [“‘I]tubulin. Since the native structure of tubulin is labile under the usual solution condi- tions (Andreu & Timasheff, 1982; Prakash & Timasheff, 1982), we considered it necessary to use as a reference ligand a stable and defined form of the protein. This was obtained by denaturation of tubulin with SDS, followed by removal of excess detergent (see Materials and Methods). The circular dichroism spectrum of this tubulin was markedly reduced with respect to the native protein (Fig. 2(b)), indicating a partial unfolding. This pro-

108 M. A. Are’valo et al.
’ I II IO 9 8 7 6 5
-log [antIbodyI,
(0)
! :I>%0 7-m I
0 0.1 0.2 0.3 0.4 0.5 - v
(b)
Figure 1. (a) Binding of 10-l’ M-[‘251]tubulin by affinity-purified anti-peptide antibodies. Circles, anti-a(415443) (Cl02 antibody); squares, anti-a(214-226) (C97 antibody). The continuous lines are drawn solely to follow the trend of the data. (b) Isotherms of binding of [‘251]tubulin to the same antibodies as above (1.7 x lo-’ M-c102 and C97) at 0°C. The results are shown in the form of Scatchard plots (Scatchard, 1949). The continuous lines are the best fitting theoretical curves for 2 types of antibodies (high and low affinity), generated according to the expression:
where [T] is the free tubulin concentration and the n. and K values were obtained by iterative computer fitting. For antibod 1.1 x 10 B
Cl02 the values obtained were n, = @07, K, M-l, n, = 0.58, K, = 4.7 x IO’ M-l; for antibody
C97 the values were n1 = @02, K, = 1.6 x log, n2 = 052, K, =%0x lo6 M-l.
tein was labelled with “‘1 and stored frozen in this state.
Figure l(a) shows representative results of binding of increasing concentrations of anti-peptide antibodies to a constant concentration of denatured [‘2sI]tubulin. The maximum fraction of radioacti- vity bound by the different antibodies comprised between 30% and 47% (average: PO( + 5) %), indi- cating that the antibodies react with the bulk of a or /I-tubulin. That is, the epitopes probed are not particular to any minor tubulin isoform and are exposed in this partially unfolded tubulin.
Figure l(b) shows the binding of denatured [‘2sI]tubulin to a constant concentration of anti-
-log Ctubulinl, (M)
(a)
x -10
;;;
- 1
-12
5 5 -2 -2 E E E E
3 3 -4 - -4 -
E E 0 0
$ $ -6- -6-
D D - - m m -8 -8
‘0 ‘0
x -10 - ;;;
-12 - 8 8 I’ 8 8 I’
-14 - -14k ‘.t ‘.t
I I I I I I I I I I I I I 200 200 210 210 220 220 230 230 240 240 250 250
Wavelength (nm) Wavelength (nm)
(b) (b)
0 O- t
Figure 2. Conformation sensitivity of the antibodies to tubulin peptides. (a) Competition radioimmunoassay of tubulin with the anti-a(415-443) antibody C102. Curve I. partially denatured tubulin (see the text); curve 2. native tub&n. (b) Circular dichroism spectra. Curve 1, partially denatured tubulin; curve 2, native tubulin; broken line. tubulin in 1 y0 SDS, which is shown for comparison with the native and partially denatured proteins.
body. Scatchard plots for two representative anti- bodies are shown. The curvature of these plot,s (concave upwards) indicates an heterogeneous population of affinities of these monospecific poly- clonal antibodies for their determined tubulin epitopes. A simplified analysis of these results in terms of only two types of affinity fitted reasonably to the data, as shown by Figure l(b). The higher affinity class was in the minority, comprising between 1 y. and 30% of the antibody molecules in the different antibodies, with apparent binding equilibrium constants in the order of IO* to lo9 M - ’ in all cases (not shown).
The interaction of each antibody preparation with the native tubulin heterodimer was examined by competition radioimmunoassay. This was per- formed under conditions that ensured tubulin stability and attainment of equilibrium during the

Microtubule Mapping by Monospecijic Antibodies 109
a
Time (min)
Figure 3. Perturbation of the assembly of purified tubulin by anti-peptide antibodies. The tracings shown are the turbidimetric assembly time-courses of 1% x 10m5 M-tubulin in assembly buffer (see Materials and Methods, section (a)). The reaction was started by a temperature increase from 4°C to 37°C. Curve a, no IgG added; curve b, 2 x 10e6 M-non-immune IgG; curves (c) to (g) 2 x 10m6 w-IgGs to positions ~(214-226) /!?(241-256), a(415443), ~~(430-443) and &412-431), respectively; curve h, 2 x 10m6 M-anti-@(415-443) IgG added to a sample identical with curve a, 20 min after the start of the reaction. Samples (a) and (b) were cooled at 20 min and samples (e), (f) and (g) at 30 min. The inset shows critical concentration plots of the assembly of tubulin with 2 x 10m6 M-anti-c((415-443) IgG (filled circles) and without it (open circles). 2 x 10e6 M-IgGs to positions ~(155-163) and &153-165) did not perturb the assembly significantly (not shown).
assay (2 h in PG buffer at 0°C). A representative comparison of native and denatured tubulin is presented in Figure 2. Native tubulin completely displaced the binding of [‘251]tubulin in all cases, indicating that the sequence zones probed are exposed in unassembled tubulin. The half-inhibitory concentrations were approximately lo-’ M-native tubulin, and were four to 15 times smaller for dena- tured tubulin depending on the antibody employed (data not shown).
(c) Perturbation of in vitro microtubule assembly by anti-peptide antibodies
The effects of antibodies on tubulin assembly were examined next. Figure 3 shows the turbidi- metric time-course of polymerization of 1.8 x 10e5 M-purified tubulin in the absence (curve a) or presence (curves b to g) of 2 x lop6 M of the different ion-exchange purified IgGs. IgGs from non-immune serum and sera to positions a(155-168), j?(153-165), ~(214-226) and &241-256) had no significant effects on the polymerization. Similar results were obtained with the affinity-puri- fied IgGs to these positions (not shown). Concentra- tions of non-immune IgG above 3 x 10e6 M
produced a non-specific increase in turbidity (not shown), which limited the assay to concentrations substoichiometric with respect to tubulin. The anti-a(415-443), anti-a(430-443) and anti-fi(412- 431) IgGs induced characteristically larger turbidi- ties (Fig. 3, curves e, f and g, respectively). Similar
results were obtained with affinity-purified IgGs. Addition of any of these three IgGs to plateau- assembled tubulin also resulted in a marked tur- bidity increase (for an example, see Fig. 3, curve h). The specific turbidity enhancement by anti-a(415-443) IgG was examined at varying total tubulin concentrations (see inset to Fig. 3). The critical protein concentration required for assembly was only slightly affected, but the slope of the plot increased from 032 to 076. Assuming that tubulin polymerization can be regarded as a nucleated condensation, these changes indicated that the apparent afinity of the growth reaction of the polymer remained essentially unchanged, but the polymer geometry was altered from that of microtu- bules to other structures scattering more light (Gaskin et al., 1974; Andreu & Timasheff, 1986). The latter was verified by electron microscopy. Microtu- bule pairs, bundles and sheet-like structures were very frequently observed, while, with non-immune IgG and the IgGs not affecting the turbidity, only microtubules and occasional microtubule pairs were seen. Figure 4 shows (a) representative micrographs of control microtubules, (b) microtubules assembled with anti-a(214-226) IgG and several degrees of abnormal microtubule polymerization (microtubule bundles, opened microtubules and sheets) exempli- fied for (c) anti-a(415-443) IgG, ((d) and (e)) anti-a(430-443) and (f) anti-/?(412-431). Therefore, the turbidity and electron microscopy results indi- cated that these antibodies to the C-terminal zones of the a and P-chains were able to perturb substoi-

110 hi’. A. A&do et al.
Figure 4. Perturbation of’ the assembly of purified tubulin by anti-prptide antibodies. Representat’ive electron micrographs of the assembly product’s of 1.8 x 1W5 M-tubulin are shown. (a) Without antibody addiCon: (h) with 2 x 10e6 M-anti-a(214-226) IgG; ( c microtubule pairs with 2 x 1W6 >I-anti-a(415-443) Ig(:; (d) microtubule bundles ) with 2 x IV6 w-anti-cr(430-443) Ig<:; (e) opened microtubules with with L? X 1OF6 M-anti-/?(412-431) Igcf. The bar represents 200 nrn.
2 x 16” M-anti-a(430-443) Ig(:: (f) sheet, formed
chiometrically the ability of purified tubulin to assemble correctly into normal microtubules.
In contrast to purified tubulin, when t,he effects of the antibodies on MAP-induced microtubule assembly were examined (2 x 1O.-6 ST of ion exchange-purified TgG and I-53 x 10m6 M of aflinity- purified IgG, employing 1 mg cycle microtubule protein/ml), non-significant effects were observed in the turbidity time-course, and only normal microtu- bules were seen under the electron microscope (see Fig. .5). This indicated that the tJubulin assembly induced by MAPS was not sensitive to the sub- stoichiometric antibody concentrations employed.
Since several of the synthetic peptides c~nxld overlap MAP-binding regions of tubulin (Serrano rf al.. 1984; Littauer et ab.. 1986), the effects of 2.5 x 10e4 M-peptide on the assembly ofmicrot~ubulr protein were examined. The three peptidrs homolo- gous to the carboxy-terminal regions were partially inhibitory (not shown). However. these peptidrs have abundant acidic residues and t,he inhibition observed could be accounted for by non-specific binding of polyanions to cationic MAP regions. In fact, 10e4 M-poly-IA-glutamic acid caused a complet,e inhibition of assembly (not shown), in support of this interpretation.

Microtubule Mapping by Monospecijic Antibodies 111
E c
H
‘;
4
1" 20 Time (min)
Figure 5. Assembly of microtubule protein in the presence of anti-peptide antibodies to tubulin. Microtubule protein (1 mg/ml) was assembled (curve a) in the absence, or (curves b to g), in the presence of 2 x 10m6 M of the same IgGs as in Fig. 3. Above is a representative electron micrograph that corresponds to Fig. 3, curve e. The bar represents 200 nm. Similar results were obtained with 1.3 x 1V6 M-affinity-purified IgGs.
(b) Binding of anti-peptide antibodies to taxol-induced microtubules, Zn ‘+ -induced sheets and
Mg *+ -induced tub&n-GDP double rings
The purpose of these experiments was to probe the exposure in the assembled protein of the small zones of the tubulin heterodimer recognized by the antibodies. Epitopes exposed in the heterodimer and occluded in microtubules are possible inter- action zones. Stable microtubules were assembled from purified tubulin with the acid of the anti- tumour drug taxol (Schiff et al., 1979). The anti- peptide antibodies were allowed to bind to them, and detected by ‘251-labelled protein A. The results
are shown in Figure 6(a). The different antibodies can be readily classified into two groups: (1) anti-a(430-443), anti-a(415443), anti-&153--165) and anti-j(412-431), which bind to microtubules; and (2) anti-a(155-168), anti-a(214-226) and anti-&241-256), which do not bind significantly to microtubules in the assay. Therefore the latter are probably occluded by the interaction between tubulin molecules in the microtubule wall. Similar experiments performed on taxol-induced MAP- containing microtubules gave qualitatively similar results, though a decrease in the binding of the anti-fl( 153-165) and anti-/?(412-431) antibodies could be observed (see Table 1).

112 M. A. Are’valo et al.
Table 1 Binding of anti-peptide antibodies to polymerized tub&in
Peptide sequence position and antibodies tested
Taxol-induced Taxol-induced microtubules Taxol-induced Zn’+ -induced Ring Epitope microtubules (fixed) MAP-microtubules sheets aggregates class?
a(1555168) (C139) 11 2 7 10 21 0 &153-165) (C141) 107 85 30 58 45 P , ~~(214-226) (C97 and C98) 17 71 5 11 13 0 8(241-256) (Cl00 and Cl 13) 0 64 18 9 9 0 a(430-443) (C85 and C103) 110 100 68 82 82 E a(415-443) (C102) 100 n.d.J 100 100 100 lx j?(412-431) (C105) 62 n.d. 29 37 37 E
Results are expressed as percentage binding, relative to the binding of anti-a(414-443) or anti-a(430-443), at 10e6 M-antibody. Where applicable, several measurements have been averaged. The standard deviation in these measurements was typically 10%. Therefore only deviations larger than 20% should be considered significant.
t 0, occluded by microtubule polymerization; E, exposed in microtubules $ n.d., not determined.
Since all of these antibodies (including anti-cr( 155168)) serve to visualize formaldehyde- fixed cellular microtubules by immunofluorescence, the above experiments were repeated on taxol- treated microtubules that had been mildly fixed by glutaraldehyde (stronger fixation conditions were not employed, to avoid non-specific crosslinking of unassembled tubulin). The results, shown in Figure 6(b), indicate that the ~(214-226) and &241-256) zones became accessible upon fixation, while ~(155-168) remained non reactive in the glutaraldehyde-fixed microtubules.
Zn2+-induced tubulin sheets are made up of protofilaments, like the microtubules. However, vicinal protofilaments in Zn2+-induced sheets have opposite orientations (Amos, 1979). The binding of the anti-peptide antibodies to Zn2+-induced tubulin sheets have opposite orientations (Amos, 1979). The binding of the anti-peptide antibodies to Zn2+-induced tubulin sheets was examined and the results, shown in Figure 7, were qualitatively similar to the binding to microtubules (Fig. 6(a)), t’hough the levels of antibody binding in this assay were smaller.
Tubulin Mg2+ -induced rings are structurally equivalent to coiled microtubule protofilaments (Mandelkow et al., 1983). Rings are the preferred form of association of tubulin in its GPD-bound, microtubule-inactive conformation (Howard & Timasheff, 1986; Melki et al., 1989). In order to have the tubulin rings easily sedimented at low speed, we prepared the partially ordered arrays of associated double rings shown in the inset to Figure 8. Binding of the different antibodies to these structures is shown (Fig. 8, which should be compared to Fig. 6(a) (microtubules)). Several differences are evident. The antibody to a(155-168), which did not bind significantly to microtubules, binds marginally to rings, in a manner similar to that of anti-&412-431). To facilitate a comparison between the polymorphic assembly products of tubulin, the binding of each antibody relative to anti-a(415-443) is displayed in Table 1, where the different epitopes
“t
6 1 -
8 7 -log [antibodyI,
(a) 1 I 1
J 8 7 6
-log hntlbodyl,
(bl
Figure 6. Binding of affinity purified anti-peptide anti- bodies to taxol-induced microtubules assembled from purified tubulin. The microtubules were observed by elec- tron microscopy (not shown). (a) Unfixed microtubules. Each antibody is indicated by the corresponding sequence position. Filled circles, ~(430-443) (average of results with C85 and Cl03 antibodies); filled squares a(415443) (C102, average of 2 experiments); filled triangles, &412-431) (Cl05 average of 2 experiments); open circles, ~~(155-168) (039, average of 2 experiments); open squares, /3( 153-165) (C141, average of 2 experiments); open trian- gles, a(214-226) (average of C97 and C98); open inverted triangles, /?(241-256) (average of Cl00 and C113). The standard error in this set of experiments was typically 300 cts/min. (b) Microtubules fixed with 0259/, glutaral- dehyde were assayed as above. The tubulin sequences a(214-226) (C97 antibody) and /?(241-256) (Cl13 anti- body) became reactive, while CI( 155-168) (C139) remained unreactive.
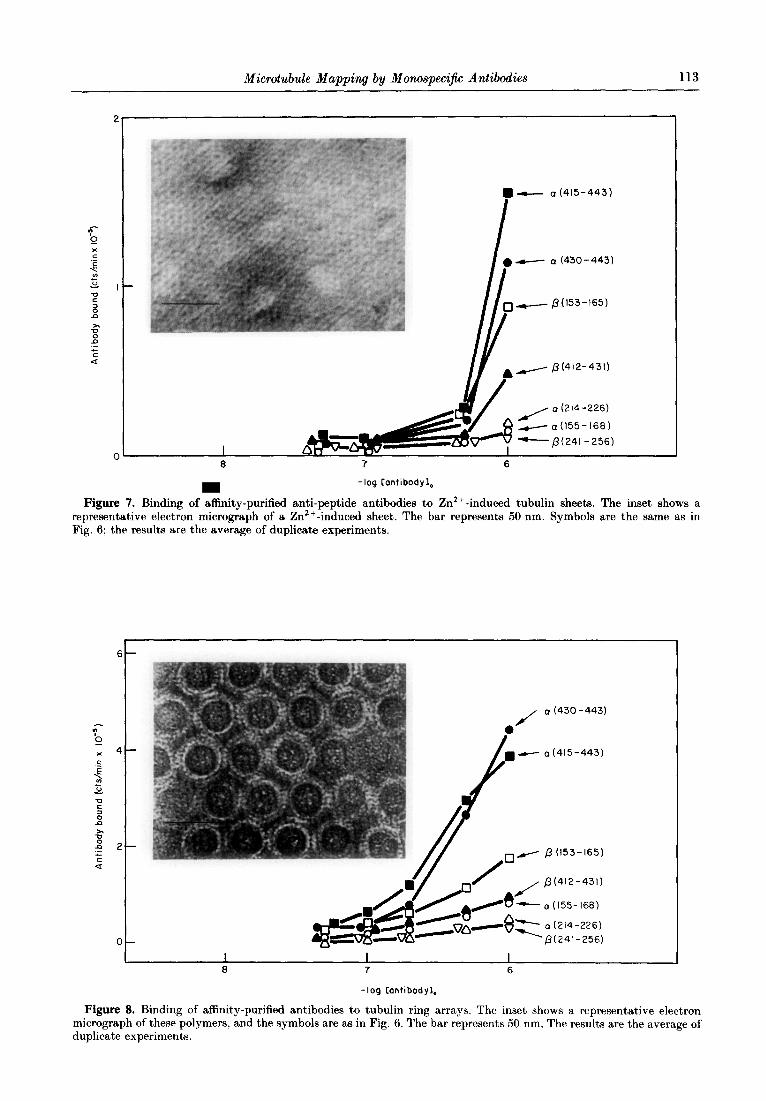
Microtubule Mapping by Monospecijic Antibodies 113
,-,- p(l53-165)
a (214-226)
- a(l55-168)
I -p(241-256)
8 7 6
-log Contibodyl,
Figure 7. Binding of affinity-purified anti-peptide antibodies to Zn2+- induced tubulin sheets. The inset shows a representative electron micrograph of a Zn2+ -induced sheet. The bar represents 50 nm. Symbols are the same as in Fig. 6; the results are the average of duplicate experiments.
6-
o-
0 (430 -443)
a(415-443)
p (153-165)
fi(412-431)
a(l55-168)
a (214-226)
‘p(241-256)
I I I 8 7 6
-log Cantibodyl,
Figure 8. Binding of affinity-purified antibodies to tubulin ring arrays. The inset shows a representative electron micrograph of these polymers, and the symbols are as in Fig. 6. The bar represents 50 nm. The results are the average of duplicate experiments.

Fi
anti.
tu
bL lli
n an
d a
seco
ndar
y rh
odam
inak
d an
tibod
y.
The
bar
repr
esen
t,s
5 pm
,

Microtubule Mapping by Monospecific Antibodies 115
Table 2 Binding of anti-peptide antibodies to cellular microtubules
Peptide sequence position and antibodies tested
Unfixed cytoskeletonst
Formaldehyde-fixed cytoskeletonst
Methanol-fixed/ formaldehyde-fixed ratio3
Epitope class§
~((155-168) (C139) /?(153-165) (Cl40 and C141) x(214-226) (C97 and C98) &241-256) (Cl00 and C113) ~(430-443) (C85 and C103) a(415-443) (C102) 8(412-431) (C105)
nd. 0 n.d. E 0.18 0 612 0 092 E 941 E 063 E
f Binding was assayed qualitatively by indirect immunofluorescence with the appropriate controls (see ?vlaterials and 1Iethods and Figs 9 and 10).
$ Binding was assayed quantitatively, substituting 1251-labelled protein A for the secondary antibody (see Materials and Methods). The numbers given are the ratio of counts specifically bound to methanol-fixed cytoskeletons to those bound to formaldehyde-fixed cytoskeletons (see Fig. 10(d) and (c) for the equivalent fluorescence assay). This assay was not applicable to unfixed cptoskeletons due to the large non-specific background. n.d., not determined.
9 0. epitope occluded in cellular mirrotubules; E, epitope exposed.
are classified as exposed (E) or occluded (0) in the microtubules assembled in vitro.
(e) Binding of anti-peptide antibodies to cellular microtu,bules
In order to determine whether t’he tubulin zones probed in vitro were equally exposed or occluded in cells, we examined the binding of each antibody to native microtubules from PtK2 cells. Coverslip- attached cytoskeletons that had not been fixed were allowed to react with affinity-purified anti-peptide antibodies in a microtubule-stabilizing buffer (see Materials and Methods, section (e)). The microtu- bule-antibody complexes were then fixed and visua- lized with fluoresceinated secondary antibody to rabbit IgG. Microtubules were also visualized in the same specimen (after fixation) by a control parallel reaction with a non-interfering monoclonal ant’i- body and rhodaminated secondary antibody to mouse IgG. The results obtained with a reactive epitope are shown in Figure 9. It can be appreciated clearly that, the same microtubules react both with the anti-peptide antibody (Fig. 9(a)) and with the monoclonal antibody (Fig. 9(b)). The anti-peptide antibody react,ed well with formaldehyde-fixed and methanol-fixed cytoskeletons (not shown). These results were obtained with the antibodies to posi- tions p(153-165), a(430-443), ~(415-443) and p(412-431) (Table 2).
The results obtained with a non-reactive epitope are shown in Figure 10. The microtubules were not made visible a,t all by the anti-peptide antibody (Fig. 10(a)), although they were clearly detected by the control monoclonal antibody (Fig. 10(b)). The anti-peptide antibody reacted well with a parallel sample of formaldehyde-fixed microtubules (Fig. 10(c)) and did not react with methanol-fixed microtubules (Fig. 10(d)). This indicated that the tubulin epitope probed is occluded in native micro- tubules and becomes exposed depending on the structural distortion produced by the fixation.
These results were obtained with the antibodies to tubulin positions a(155-168), a(214-226) and 8(241-256) (Table 2). There was a complete agree- ment between the exposure of the different epitopes in the in vitro taxol-induced microtubules (Table 1) and in the microtubules prepared from living cells (Table 2).
4. Discussion
(a) Exposed sequences in the tu,bulin a-/I-heterodimer
Monospecific polyclonal antibodies to the conserved positions of tubulin a( 155-168), p(153-165), a(214-226), /?(241-256), a(415-443) a(430-443) and /l(412-431) consisted of populations of heterogeneous affinities (Fig. 1). All these anti- bodies were conformation-sensitive and reacted better with partially denatured tubulin than with native tubulin (Fig. 2, and see above). These anti- bodies were generated by short carrier-coupled peptide sequences and might bind better to the cognate protein sequences in the partially unfolded, mobile forms, than to their more restricted mobility forms in the native protein (Dyson et al., 1988). Nevertheless. native tubulin was able to displace [“‘T?]tubulin completely in the competition radio- immunoassay in all cases. The conditions of this radioimmunoassay, which ensured attainment of binding equilibrium, are standard to avoid any significant denaturation of tubulin (Andreu & Timasheff, 1982; Prakash & Timasheff, 1982). Therefore the results indicate that all the tubulin sequences probed are similarly accessible to their antibodies in the native protein. The simplest inter- pretation is that all these hydrophilic sequences constitute epitopes exposed at the surface of tubulin. Therefore they do not participate directly in the a-j? binding interface. This conclusion is fully consistent with the limited proteolysis results in the case of a(155-168)/fl( 1533165) and

116 M. A. Are’valo et al.
a(415443)//?(412-431) since these positions are respectively included in, and adjacent to, the first and third preferential proteolysis zones (de la Vifia et al., 1988), respectively, which are necessarily exposed. On the other hand, if some of the epitopes probed were at the a-p binding interface, but were made accessible by a process of tubulin dissociation into its subunits (Detrich et al., 1982), displacement in the radioimmunoassay at the lower tubulin con- centrations and incomplete displacement at the higher tubulin concentrations would be expected; however, this is contrary to the results.
(b) Perturbation of the assembly of tubulin by antibodies to carboxy-terminal positions
We wished to know whether any of the antibodies to the small zones of the tubulin heterodimer could interfere with the functionality of the protein. Anti- bodies to positions a(415443), ~(430-443) and /?(412-431) induced a characteristic substoichio- metric perturbation of tubulin assembly, consisting of microtubule bundling, opening and polymeriza- tion into sheets (Figs 3 and 4). The perturbation effect was not observed with Fab fragments prepared from these antibodies (J. M. Andreu, unpublished results), which indicated the require- ment of antibody bivalency or/and the presence of the Fc portion. This suggested that the ~(430-443) and /?(412-431) sequences are not contact sites, but are relatively well exposed in the microtubules made of purified tubulin; bivalent antibody binding to the carboxy termini would simply distort the polymer wall.
The fact that the antibodies to the internal chain positions did not significantly induce the substoi- chiometric perturbation suggested that these posi- tions were not within reach for bivalent antibody binding, be they exposed at the microtubule surface or occluded by polymerization. Stoichiometric assembly-blocking experiments were precluded in our case by a non-specific tubulin-IgG interaction at high antibody concentrations. Hesse et al. (1987) reported the inhibition of the assembly of microtu- bule protein by preincubation with an antibody to positions fl(154-165) in the absence of GTP. This effect might be related to the inhibition of nucleo- tide exchange at the E-site of tubulin by this anti- body (Hesse et al., 1987) and not necessarily to a, steric blocking of tubulin polymerization by the antibody.
The assembly of microtubule protein was not substoichiometrically perturbed by our antibodies. In a similar manner, Wehland et al. (1983) reported that the monoclonal antibody YL l/2, which binds to the tyrosylated C terminus of tubulin, very weakly affected the assembly of microtubule protein in vitro. In our case, the differential effect of the antibodies to positions a(415443), a(439-443) and /?(412-431) on the assembly of tubulin and tubulin plus MAPS suggests a combination of the following possibilities. (1) The MAP-induced polymerization is
stronger than the polymerization of purified tubulin. MAP-containing microtubules can not’ approach one another because of the MAP projec- tions and therefore are not bundled by the anti- bodies. (2) MAPS compete with the antibodies for binding to all the C-terminal regions probed. This second possibility is compatible with the reported locations of MAP-binding sites (Serrano et al., 1984; Littauer et aE., 1986). However, it is not sufficient, since anti-a(430-443) and anti-a(415-443) bind well to MAP-containing microtubules (Table 1).
(c) Tub&in sequences antigenically exposed or occluded in microtubules
Antibodies to positions a(415-443), ~(430-443) and &412-431) bound specifically to taxol-induced microtubules made of purified tubulin (Fig. 6), in agreement with their discussed ability to perturb assembly. When the taxol-induced microtubules containing MAPS were assayed, a significant decrease in the reactivity of anti-8(412-431) was noticed (Table 1). This would be compatible with the role proposed for the neighbouring positions p(434-440) in the binding of MAP, and tau by Littauer et al. (1986), from the results of a dot-blot assay of binding of the radioiodinated MAPS to a number of peptides. MAP-binding sites might be subjected to a detailed characterization, possibly by competition binding assays employing individually purified MAPS, antibodies, peptides and micro- tubules in solution. These antibodies bound to intact, microtubules from PtK2 cells (Fig. 9), confirming the result obtained with the microtubules assembled in vitro and indicating that the regions a(430-443) and 8(412-431) are exposed on the surface of the microtubules and not blocked by other proteins or subcellular components in the PtK2 cell models employed. This clearly is consistent’ with published results employing DMlA and DMl B monoclonal antibodies (Blose et aZ., 1984): whose epitopes overlap these positions (Breitling & Little, 1986; de la Vifia et al., 1988). It is also in agreement with the general notion that the C-terminal zones of tubulin are flexible peptides that protrude from the protein (Posting1 et al., 1979; Woody et al., 1983; Ringel Cyr Sternlicht, 1984; Serrano et al., 1984) and are exposed in microtubules (Wehland et al.. 1983: Sackett $ Wolf, 1986).
Of the two equivalent zones ~~(155~--168) and /?(153-165), both accessible in unassembled tubulin, the former is non-reactive while the latter is reactive in microtubules. The results with taxol-induced microtubules and cell models are fully consistent’. In particular, c(( 155-168) resisted glutaraldehyde fixa- tion without becoming exposed (Fig. 6(b)), but not formaldehyde fixation (as in Fig. 10(c)). One can therefore consider a( 155-168) to be a sequence anti- genitally occluded by microtubule assembly. This is consistent with the observation of a strong decrease in a subtilisin cleavage point adjacent to a( 155--16X) in taxol-induced microtubules, compared to unassembled tubulin (M. A. Arevalo & ,J. $1.

Microtubule Mapping by Monospeci$c Antibodies 117
I 3gure 10. ((a) and (b)) Same assay aa in Fig. 9 made with anti-cr(214-226) (C98 antibody). ((c) and (d)) The reacti on of the same antibody with formaldehyde-fixed and methanol-fixed cytoskeletons respectively. The bar represents 10 w.
act sigi mic act Thl
dreu, unpublished results). The ~(214-226) and newly mapped zones that are antigenically occll uded :41-256) sequences, which are antigenically by microtubule assembly. The possibility ’ of :essible in the tubulin heterodimer, did not react assembly-linked conformational changes leadir 1g to nificantly in taxol-induced microtubules or in epitope occlusion cannot be excluded at pre, sent. :rotubules from PtK2 cells, though they became However, the simplest interpretation of 1 these :essible by aldehyde fixation of the microtubules. results is that the a(155-168), a(214-226) and erefore ~~(214-226) and /?(241-256) constitute /?(241-256) q se uences are part of, or close to , the

M. A. Arehalo et al.
Cm)
(a) M Figure 11. Sizes and approximate shapes of antibody
probes and microtubules, compared for the purpose of discussion. (a) Scheme of an antibody, after the model of human immunoglobulin IgG, by Pumphrey (1986) and the approximate dimensions of rabbit TgG (Roux & Metzger. 1982: Valentine & Green, 1967). The bar represents 2 nm. The position of the Fab arms in solution is flexible (Oi et al.; 1984) around mean values that, apparently depend on the IgG subclass (Gregory et al.. 1987). Each Fab is about the same mass as a tubulin monomer (M, 50,000). The distance between the 2 antigen binding sites should be considered to be variable, between approx. 6 nm minimum and 14 nm maximum for the purpose of this discussion. The shaded region on the right arm of (a) exemplifies the zone occupied by a hypothetical spherical probe of 2 nm diameter. These probes were employed in model calculations of antibody-accessible surface regions of proteins, leading to meaningful results (Novotny et al.. 1986). If a tubulin monomer was considered simplistically as a sphere of 4 nm diameter, it could be surrounded by a maximum of 26 spheres of 2 nm diameter in contact with it. On the other hand, antibody-binding sites have been described as a roughly 3 nm x 2 nm surface that contacts a complementary surface in the protein (Alzari et aZ., 1988). This would allow about 8 contacts with a 4 nm sphere. From these approximations, one might expect between 8 and 26 antigenic determinants per tubulin monomer, a part of which would be occluded by hetero- dimer formation and others by the microtubule wall. Interestingly. hydrophilicity and secondary structure anal-@s of the t,ubulin sequences (de la Viiia rt al.. 1988) predlcted 17 potent,ial epitopes for the cc-chain and 18 for the B-chain. While simultaneous binding of IgGs to neigh- bouring epitopes is expected to be sterically hindered, probing the microtubule surface with individual anti- body-binding sites should be of a resolution not much coarser than the level of resolution of present electron microscopy and fibre X-ray diffract,ion models. which is 1.8 nm at t,he best (Beese et al.. 1987); imunochemical mapping should be complementary to struct’ural deter-
regions of’ contact between tubulin heterodimers in the microtubule wall, to the resolution of the antibody probes employed.
(d) Immunochemkal mapping qf microtubulrs
Having determined several chemically defined zones of tubulin to be exposed or occluded to anti- bodies in microtubules, we wished to deduce as much as possible of the probable positions of these zones in the structure of microt,ubules. The I& molecules are not very small compared to the micro tubule wall, but the antibody binding sites art> adequately sized probes (see Fig. 11). The following discussion is based on three general assumptions regarding microtubule structure: (1) the structures of the LX and /J-monomers, apart from local differ- ences, are homologous: as are their sequences: (2) the monomers within the microtubule wall are related b-y translation operations (all have the same’ polarity); and (3) the structures of the unassembled and assembled a-/?-heterodimers are roughly similar. The following three hypotheses can be suggested.
(I) The perturbation of tubulin assembly 1, bivalent antibodies to positions ~((430-443) and /?(412-431) and in particular the formation of opened microtubules and sheets (Fig. 4(e) and (f), respectively), would be consistent) with the diskor- tion of the lateral curvature of the microtubule wall by the simultaneous binding of one antibody to two protofilaments. Comparison of the gross dimensions of an IgG molecule with the interfilament spa,cing in the microtubule wall suggests that such antibod) crosslinking could in principle be possible between alternating protofilaments and would be less likeI> between vicinal ones (see Fig. 11). The former would be equally compatible with the A or H microtubule lattices (Amos, 1987). The process would be easil! envisaged if the a(430-443) and /?(412-431) sequences were flexible peptides placed at the proto- filament ridges and not in the grooves. This t’enta- tive model is subject to experimental testing.
---
mination. (m) Microtubule cross-section, after the model of Beese et al. (1987); the contour chosen encloses 91 “b of the density and shows the protofilament ridges (arrow) and the interprotofilament grooves (broken arrow). (‘on- parison of (a) with (m) suggests the following. (I) The surface of the grooves should be partially accessibk to antibody binding, while the ridges are fully accessible. (2) Bivalent binding of the monospecific antibodies t,o rrpeti- tive epitopes in different protofilaments within t,he same microtubule wall (leading to distortion) should be possible for flexible epitopes located in alternating prot~ofilammts and only with considerable approaching of the t)wo lg(; arms for epitopes in vicinal protofilament,s. (3) Bivaltrnt binding of the antibodies to repetitive epitopes in different microtubules (leading to microt,ubule bundling) is more easily envisaged for outer-located. flexible epitopes than for more groove-located and less flexiblr epitopes. (4) The diffusion of antibody molerules into I he microtubules lumen could be sterically hindered by the inner microtubule wall.

Microtubule Mapping by Monospeeijic Antibodies 119
C-t----- l
0
2- 7 (8) 9 IO (II) 12
-
.Z
: 4’78 9 (IO) II 12
I50 200 250 300 Amino acid number
(0) (b)
Figure 12. (a) Working model of the arrangement of the 214 to 256 sequence regions within the heterodimer and t’he protofilament. The ~((214-226) and /?(241-256) sequences are occluded by polymerization, while the ~(241.-256) and /?(214-226) sequences are close to the a-b binding interface (see the text). Note that the drawing represents exclusively the microtubule wall lattice and certain interactions, and the monomer substructure has been ignored deliberately; that is, a and /l could equally stand here for a monomer domain, such as the large domain A of Beese et al. (1987). For similar reasons this model would be equally compatible with interactions between the N-terminal domain of 1 subunit (regions 1 and 2 of our proteolysis map; de la Vifia et al., 1988) and the C-terminal domain (region 3) of the next subunit, such as in the elegant model of Kirchner & Mandelkow (1985). (b) Hydrophilicity profiles of the middle regions of the a and b-tub& sequences. after de la Vifia et al. (1988).
(II) The antibody to the fl(l53-165) sequence, which is exposed in microtubules, did not induce the characteristic assembly perturbation, suggesting that these positions are less flexible or located in the microtubule grooves, or both. The fact that the a( 155-168) sequence is not antigenically accessible could be reconciled with the exposure of p(l53-165) if these two positionally equivalent regions (or their complementary positions in neighbouring monomers) had a different tertiary structure. The a( 155-168) zone is also occluded in Zn2+-induced sheets and could be partially exposed in tubulin rings (Figs 7 and 8, respectively), which suggests that it is not located in the microtubule lumen and instead might be involved in lateral interactions between protofilaments. All these clues taken jointly suggest that both c(( 155-168) and 8(153-165) might be localized at the interprotofilament grooves, the former being more buried than the latter.
(III) The fact, that the ~(214-226) and 8(241-256) sequences are antigenically occluded in the different tubulin assembly forms examined (microtubules, Zn2+-induced sheets, ring arrays) suggests that they are hindered from reacting with their antibodies by the axial contacts between tubulin heterodimers, which lead t#o protofilament formation. This is exemplified bv the scheme of Figure 12(a). In this model the axial contacts in each extreme of the heterodimer are made at positions close to
~~(214-226) and /?(241-256). An important corollary is that (on the basis of assumptions (1). (2) and (3), above) the contacts between c1 and fi-monomers within the heterodimer should be made near the ~(241-256) and fi(214-226) sequences, as depicted by the broken lines in Figure 12(a). In Figure 12(b) the hydrophilicity profiles of the central regions of the a and fi-tubulin chains are given. It is at these regions that the two sequences are in general more hydrophobic (Posting1 et al., 1983) and where conspicuous differences between the otherwise homologous profiles appear. In particular, hydro- philicity peak 10 (positions 214-226) is expressed in a but not in /? and, conversely. peak 11 (position 241-256) is expressed in /? but not in a. Equivalent sequence positions that are predicted to be exposed in one monomer and hidden in the other are possible axial interdimer and intradimer interaction zones in their respective monomers. Therefore this hydrophi- licity analysis (Fig. 12(b)) is consistent with the interaction scheme based on antigenic reactivity (Fig. 12(a)) and supports the model whereby the a(214-226) and /?(241-256) sequences are related to the axial contacts between the dimers in the proto- filament, whereas the a(241-256) sequences are related to the a-p binding interface of t’he tubulin dimer.
We thank J. Morales for technical assistance, Dr J. L. Carrascosa for helpful criticisms to the paper, Dr M.

M. A. A&valo et al.
Suffness for taxol, Dr I. Gil for the S. aureua immunopre- cipitant, Dr C. de la Terre for extended use of her photo- microscope, Servicio de Microscopia Electronica C.I.B., and A. Chao and J. F. Diaz for their help with the manuscript. This work was supported by DGICyT grant PB87-0220. Preliminary reports were presented at the EMBO Workshop on the Cytomatrix (Maria Alm, Austria, 1988) and at the 4th European Cytoskeletal Meeting (Lyon, France, 1988).
References
Alazari, P. M., Lascombe, M. B. & Poljak, R. J. (1988). Annu. Rev. Immunol. 6, 555-580.
Amos, L. A. (1979). In Microtubules (Roberts, K. & Hyams, J. S., eds), pp. l-64, Academic Press, New York.
Amos, L. A. & Eagles, P. M. A. (1987). In Fibrous Protein Structure (Squire, J. M. t Vibert, P. J., eds). pp. 215-246, Academic Press, London.
Anderson, D. C., Wible, L. J., Hughes, B. J., Smith, C. W. & Brinkley, B. R. (1982). Cell, 31, 719-729.
Andreu, J. M. (1982). EMBO J. 1, 1105-1110. Andreu, J. M. & Timasheff, S. N. (1982). Biochemistry, 21,
6465-6476. Andreu, J. M. & Timasheff, 6. N. (1986). Methods
Enzymol. 130, 47-59. Andreu, J. M., de la Torre, J. & Carrascosa, J. L. (1986).
Biochmistry, 25, 5230-5239. Andreu, D., de la Vifia, S. & Andreu, J. M. (1988). Int. J.
Peptide Protein Res. 31, 555-566. Beese, L., Stubbs, G. & Cohen, C. (1987). J. Mol. Biol.
194, 257-264. Blose, S. H., Meltzer, D. I. & Feramisco, J. R. (1984).
J. Cell Biol. 98, 847-858. Breitling, F. & Little, M. (1986). J. Mol. Biol. 189,
367-370. Campbell, A. M. (1984). Monoclonal Antibody Technology,
Elsevier, Amsterdam. de la Viiia, S., Andreu, D., Medrano, J. F., Nieto, J. M. &
Andreu, J. M. (1988). Biochemistry, 27, 5352-5356. Detrich, H. W., Williams, R. C. & Wilson, L. (1982).
Biochemistry, 21, 2392-2400. Dyson, H. J., Lerner, R. A. & Wright, P. A. (1988). Annu.
Rev. Biophys. Biophys. Chem. 17, 305-324. Gaskin, F. & Kress, Y. (1977). J. Biol. Chem. 252,
6918-6924. Gaskin, F., Cantor, C. R. & Shelanski, M. L. (1974).
J. Mol. Biol. 89, 737-758. Green, N., Alexander, H., Olson A., Alexander, S.,
Shinnick, T. M., Sutcliffe, J. G. & Lerner, R. (1982). Cell, 28, 477-487.
Gregory, L., Davis, K. G., Sheth, B., Boyd, J., Jefferis, R., Nave, C. & Burton, D. R. (1987). Mol. Immunol. 224, 821-829.
Hesse, J., Thierauf, M. & Ponstingl, H. (1987). J. Biol. Chem. 262, 15472-15475.
Hinz, H. J., Gorbunoff, M. J., Price, B. & Timasheff, S. N. (1979). Biochemistry, 18, 3084-3089.
Howard, W. D. & Timasheff, S. N. (1986). Biochemistry, 25, 8292-8300.
Hunter, W. M. & Greenwood, F. C. (1962). Nature (London), 194, 495-496.
Karr, T. L., White, H. D., Coughlin, B. A. & Purich, 1). L. (1982). Methods Cell. Biol. 24, 51-60.
Kirchner, K. & Mandelkow, E. M. (1985). EMBO J. 4, 2397-2402.
Krauhs, E., Little, M., Kempf, T., Hofer-Warbinek, R., Ade, W. & Ponstingl, H. (1981). Proc. Nat. Acad. hi.. U.S.A. 78, 4156-4160.
Lee, J. (1. & Timasheff, S. K’. (1977). Biochemistry, 16. 175991769.
Lee, J. C., Frigon, R. P. & TimashetT. S. N. (1973). ,I. Biol. Chem. 248, 7253-7262.
Lin, F. T.. Zinnecker, M., Hamaske, T. C Katz, 1). (1979). Biochemistry, 18, 690-697.
Littauer, U. Z., Giveon, D., Thierauf, M.. G&burg, I. & Postingl, H. (1986). Proc. Nat. Acad. Sci., rr.S. A. 83, 7162-7168.
Mandelkow, E. M. & Mandelkow, E. (1985). J. Mol. Biol. 181. 123-135.
Mandelkow, E., Thomas, ,J. & Cohen, (1.. (1977). Proc. Nat. Acad. Sci., U.S.A. 74, 3370-3374.
Mandelkow. E., Mandelkow, E. M. & Bordas, J, (1983). J. Mol. Biol. 181, 123-135.
Mandelkow, E. M., Hermann. M. & Riihl, L’. (1985). J. Mol. Biol. 185, 311-327.
Melki, R., Carlier. M. F., Pantaloni, D. & Timasheff. S. N. (1989). Biochemistry, 28, 9143-9152.
Novotny, J., Handschumacher, M., Haber. E., Bruceoleri. E.-E., Carlson, W., Fanning, D., Smith, J. & Rose, D. (1986). Proc. Nat. Acad. Sci., U.S.A. 83, 226-230.
Oi, V. T., Vong, T. M., Hardy, R., Reidler, tJ., Dangl, J.. Herzenberg, L. A. & Stryer, L. (1984). Nature (London), 307, 136-140.
Osborn, M. & Weber. K. (1982). Method8 Cell Biol. 24, 98-132.
Ponstingl, H.? Little, M., Krauhs, E. & Kempf, T. (1979). Nature (London), 282, 423-424.
Ponstingl, H.. Krauhs, E., Little, M. & Kempf. T. (1981). Proc. ‘Vat. Acad. Sci., 1i.S.A. 78, 2757-2761,
Ponstingl, H., Krauhs. E. & Little, M. (1983). J. Submicrosc. Cytol. 15, 359-362.
Prakash, V. 6 Timasheff, S. N. (1982). J. Mol. Biol. 160, 499-515.
Pumphrey. R. (1986). Immunol. Today, 7. 174-178. Ringel, I. & Sternlicht, H. (1984). Biochemistry, 23.
5644-5653. Roux, K. J. & Metzger. 1). W. (1982). J. Immunol. 129,
2548-2553. Sackett, D.-L. & Wolf. J. (1986). .I. Biol. Chem. 261,
9070-9076. Scatchard. G. C. (1949). dnn. N. Y. dead. Sci. 51.
600-672. Schiff, P. B., Fant, J. & Horwitz, S. H. (1979). Nature
(London), 277, 665-667. Serrano, L.. de la Torre, .J., Maccioni. R. B. & Avila, ,J.
(1984). Proc. Nat. Acad. Sci., 7J.S. A. 81, 5989-5998. Tamm. J. P., Heath, W. F. & Merrifiel, R. B. (1983).
J. Ann. Chem. Sot. 105, 6442-6455. Valentine, R. C. & Green, N. M. (1967). J. Mol. Hiol. 27.
615-617. Van de Wat,er. L. & Olmsted. ,I. B. (1980). .J. Biol. (!hem.
255, 10744-10751. Wais-Steider, C., White, N. S., Gilbert, 1). S. & Eagles.
P. A. M. (1987). J. Mol. Biol. 197, 205&218. Wehland, J., Willingham, M.-C. & Sandoval. I. V. (1983).
J. Cell. Biol. 97, 1467-1475. Weisenberg? R. C. (1972). Science, 177, 1196-l 197. Woody, R. W.. Clark, D. C., Roberts, G. C., Martin, S. &
Bayley, P. M. (1983). Biochemistry, 22. 2186-2192.
Edited by D. DeRosier




















