Gastrointestinal peptides controlling body weight homeostasis
15
Review Gastrointestinal peptides controlling body weight homeostasis Hugo Mendieta-Zero ´n a,1 , Miguel Lo ´pez a,b,1 , Carlos Die ´guez a,b, * a Department of Physiology, School of Medicine, University of Santiago de Compostela, San Franscisco s/n, 15782 Santiago de Compostea, A Corun ˜a, Spain b CIBER of Obesity and Nutrition (ISCIII), Spain Received 8 May 2007; revised 6 November 2007; accepted 12 November 2007 Available online 21 November 2007 Abstract Obesity has become an international public health problem. Unfortunately, effective treatment options are limited. In the last 20 years, research in obesity and associated pathologies has derived in a significant increase in the knowledge of the physiological and molecular mechanism regulating body mass, such as gastrointestinal-neuroendocrine communications. Gut–brain peptides may provide attractive therapeutic targets against this disease. This review summarizes research into energy balance through gastrointestinal tract peptides. Understanding these molecular mechanisms will provide new pharmacological targets for the treatment of obesity and appetite disorders. Ó 2007 Elsevier Inc. All rights reserved. Keywords: Brainstem; Cholecystokinin; Food intake; Ghrelin; GLP-1; Gut; Hypothalamus; Incretins; Insulin; PYY; Vagus nerve 1. Introduction In both the developed and developing world, levels of obesity and its related disorders are increasing at a rate that could be considered of epidemic proportions. For this rea- son, in the last decade there has been a significant increase in the knowledge of the physiological and molecular mech- anisms regulating body mass. Animals are now known to regulate body weight by a complex homeostatic mechanism involving interactions between peripheral organs, such as white adipose tissue (WAT), gut, thyroid, muscle, gonads, and the central nervous system (CNS), through signals that inform brain centers of nutritional, as well as, metabolic status of the animal (Fru ¨ hbeck and Gomez-Ambrosi, 2003; Flier, 2004; Medina-Gomez and Vidal-Puig, 2005; Morton et al., 2006; Foster-Schubert and Cummings, 2006; Lo ´pez et al., 2007b). 2. Neural control of food intake In the last 20 years, the knowledge about the role of the hypothalamus on the regulation of feeding has substan- tially improved. However, since the initial ‘‘Dual Centre Hypothesis’’ (Hecherington and Ranson, 1942; Anand and Brobeck, 1951a; Anand and Brobeck, 1951b), the main concept has not changed, i.e., anatomically defined hypo- thalamic areas regulate food intake. These hypothalamic defined areas, called nuclei, form interconnected neuronal circuits that respond to changes in energy status by altering the expression of specific molecules, especially neuropep- tides, which results in changes in energy intake and expen- diture (Fru ¨ hbeck and Gomez-Ambrosi, 2003; Flier, 2004; Morton et al., 2006; Lo ´pez et al., 2007b). There are several hypothalamic nuclei involved in energy homeostasis but the arcuate nucleus (ARC) is unan- imously considered as the ‘‘master hypothalamic centre’’ for feeding control (Fig. 1). It is situated around the base of the third ventricle and lies immediately above the med- ian eminence, where the blood–brain–barrier (BBB) allows the entry of peptides and proteins from the circulation, such as leptin, ghrelin, and insulin. Two distinct neuronal populations in the ARC integrate peripheral nutritional/ 0016-6480/$ - see front matter Ó 2007 Elsevier Inc. All rights reserved. doi:10.1016/j.ygcen.2007.11.009 * Corresponding author. Fax: +34 981 574145. E-mail address: [email protected] (C. Die ´guez). 1 These authors equally contributed to this work. www.elsevier.com/locate/ygcen Available online at www.sciencedirect.com General and Comparative Endocrinology 155 (2008) 481–495
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Gastrointestinal peptides controlling body weight homeostasis

Available online at www.sciencedirect.com
www.elsevier.com/locate/ygcen
General and Comparative Endocrinology 155 (2008) 481–495
Review
Gastrointestinal peptides controlling body weight homeostasis
Hugo Mendieta-Zeron a,1, Miguel Lopez a,b,1, Carlos Dieguez a,b,*
a Department of Physiology, School of Medicine, University of Santiago de Compostela, San Franscisco s/n, 15782 Santiago de Compostea, A Coruna, Spainb CIBER of Obesity and Nutrition (ISCIII), Spain
Received 8 May 2007; revised 6 November 2007; accepted 12 November 2007Available online 21 November 2007
Abstract
Obesity has become an international public health problem. Unfortunately, effective treatment options are limited. In the last 20years, research in obesity and associated pathologies has derived in a significant increase in the knowledge of the physiological andmolecular mechanism regulating body mass, such as gastrointestinal-neuroendocrine communications. Gut–brain peptides may provideattractive therapeutic targets against this disease. This review summarizes research into energy balance through gastrointestinal tractpeptides. Understanding these molecular mechanisms will provide new pharmacological targets for the treatment of obesity and appetitedisorders.� 2007 Elsevier Inc. All rights reserved.
Keywords: Brainstem; Cholecystokinin; Food intake; Ghrelin; GLP-1; Gut; Hypothalamus; Incretins; Insulin; PYY; Vagus nerve
1. Introduction
In both the developed and developing world, levels ofobesity and its related disorders are increasing at a rate thatcould be considered of epidemic proportions. For this rea-son, in the last decade there has been a significant increasein the knowledge of the physiological and molecular mech-anisms regulating body mass. Animals are now known toregulate body weight by a complex homeostatic mechanisminvolving interactions between peripheral organs, such aswhite adipose tissue (WAT), gut, thyroid, muscle, gonads,and the central nervous system (CNS), through signals thatinform brain centers of nutritional, as well as, metabolicstatus of the animal (Fruhbeck and Gomez-Ambrosi,2003; Flier, 2004; Medina-Gomez and Vidal-Puig, 2005;Morton et al., 2006; Foster-Schubert and Cummings,2006; Lopez et al., 2007b).
0016-6480/$ - see front matter � 2007 Elsevier Inc. All rights reserved.
doi:10.1016/j.ygcen.2007.11.009
* Corresponding author. Fax: +34 981 574145.E-mail address: [email protected] (C. Dieguez).
1 These authors equally contributed to this work.
2. Neural control of food intake
In the last 20 years, the knowledge about the role of thehypothalamus on the regulation of feeding has substan-tially improved. However, since the initial ‘‘Dual CentreHypothesis’’ (Hecherington and Ranson, 1942; Anandand Brobeck, 1951a; Anand and Brobeck, 1951b), the mainconcept has not changed, i.e., anatomically defined hypo-thalamic areas regulate food intake. These hypothalamicdefined areas, called nuclei, form interconnected neuronalcircuits that respond to changes in energy status by alteringthe expression of specific molecules, especially neuropep-tides, which results in changes in energy intake and expen-diture (Fruhbeck and Gomez-Ambrosi, 2003; Flier, 2004;Morton et al., 2006; Lopez et al., 2007b).
There are several hypothalamic nuclei involved inenergy homeostasis but the arcuate nucleus (ARC) is unan-imously considered as the ‘‘master hypothalamic centre’’for feeding control (Fig. 1). It is situated around the baseof the third ventricle and lies immediately above the med-ian eminence, where the blood–brain–barrier (BBB) allowsthe entry of peptides and proteins from the circulation,such as leptin, ghrelin, and insulin. Two distinct neuronalpopulations in the ARC integrate peripheral nutritional/

482 H. Mendieta-Zeron et al. / General and Comparative Endocrinology 155 (2008) 481–495
feeding signals. One set of neurons located in the ventrome-dial part of the ARC express the orexigenic (feeding pro-moting) neuropeptides agouti-related protein (AgRP) andneuropeptide Y (NPY). In the ventrolateral part of theARC, a second population of neurons expresses theanorexigenic (feeding inhibitors) products of proopiomela-nocortin (POMC), the precursor of a-melanocyte stimulat-ing hormone (a-MSH)2, and cocaine and amphetamineregulated transcript (CART).
Neurons in the ARC project to other ‘‘second order’’neurons located in hypothalamic nuclei, such as the dorso-medial nucleus (DMH), the paraventricular nucleus(PVH), the ventromedial nucleus (VMH), and the lateralhypothalamic area (LHA). Those nuclei integrate neuro-peptide signals from the ARC and other CNS areas, suchas the brainstem, by modulating the expression of otherspecific neuropeptides. LHA-neurons primarily expressorexigenic neuropeptides, such as orexins (OX-A andOX-B), melanin-concentrating hormone (MCH) anddynorphin (DYN); on the other hand, PVH-neuronsmainly produce anorexigenic neuropeptides, such asCART, corticotrophin-releasing hormone (CRH), and thy-rotrophin-releasing hormone (TRH) (Flier, 2004; Mortonet al., 2006; Lopez et al., 2007b). The integration of thesesignals drives the adequate homeostatic response to energyhomeostatic demands.
3. Enteric nervous system
The gastrointestinal tract (GIT) receives a dual extrinsicinnervation by the autonomic nervous system, i.e., by itsparasympathetic division, including vagal and pelvicnerves, and sympathetic division comprising splanchnic
2 Abbreviations: a-MSH, a-melanocyte-stimulating hormone; AgRP,agouti related peptide; AMPK, AMP-activated protein kinase; AP, areapostrema; ARC, arcuate nucleus of the hypothalamus; BBB, brain–blood–barrier; BBS, bombesin; BBSR-3, bombesin-3 receptor; cAMP, cyclic-adenosine monophosphate; CART, cocaine and amphetamine relatedtranscript; CCK, cholecystokinin; CNS, central nervous system; CRH,corticotrophin-releasing hormone; CRH1, receptor type 1; CRH2, receptortype 2; DMH, dorsomedial nucleus of the hypothalamus; DPP-IV,dipeptidyl peptidase IV; DVC, dorsal vagal complex; ENS, entericnervous system; EX-4, exendin-4; EX9-39, exendin9–39; FFAs, free fattyacids; GH, growth hormone; GHRH, growth hormone-releasing hor-mone; GHS-R, growth hormone secretagogue receptor; GIP, gastricinhibitory polypeptide or glucose-dependent insulinotropic peptide; GIP-R, GIP receptor; GIT, gastrointestinal tract; GLP-1 and -2, glucagon-likepeptide-1 and 2; GLP-1R, GLP-1 receptor; GRP, gastrin-releasingpeptide; GRP-R, gastrin-releasing peptide receptor; HFD, high fat diet;IR, insulin receptor; LC, locus coeruleus; LHA, lateral hypothalamic area;MCH, melanin-concentrating hormone; NMB, neuromedin B; NPY,neuropeptide Y; NTS, nucleus of the solitary tract; OX1R, orexin receptor1 (hypocretin receptor 1); OX2R, orexin receptor 2 (hypocretin receptor 2);OX-A, orexin A (hypocretin 1); OX-B, orexin B (hypocretin 2); OXM,oxyntomodulin; PCs, prohormone convertases; POMC, proopiomelano-cortin; PP, pancreatic polypeptide; PVH, paraventricular nucleus of thehypothalamus; PYY, peptide YY; RYGB, Roux-en-Y gastric bypass;TRH, thyrotrophin-releasing hormone; UAG, unacylated (desoctanoyl ordesacyl) ghrelin; VMH, ventromedial nucleus of the hypothalamus; WAT,white adipose tissue.
nerves (Konturek et al., 2004; Thompson, 2006; Naslundand Hellstrom, 2007). In addition to this autonomic inner-vation, the GIT also has its own nervous system, namelythe enteric nervous system (ENS) that directly controlsthe gastrointestinal system. The ENS has two major com-ponents, the submucous plexus (Meissner’s plexus) andthe myenteric plexus (Auerbach’s plexus). The submucousplexus is mostly involved in responses to nutrients signalingthrough the gut epithelium, while the myenteric plexus, sit-uated between the longitudinal and circular muscle of theGIT, is primarily involved in the coordination of motilitypatterns (Konturek et al., 2004; Thompson, 2006; Naslundand Hellstrom, 2007).
The ENS modulates the release of peptides from endo-crine cells, known as enteroendocrine cells (Kirchgessnerand Liu, 1999; Kirchgessner, 2002; Naslund and Hell-strom, 2007). The ENS, interconnected with the autonomicnervous system, transmits information of mechanical (dis-tension, contraction), chemical (presence of nutrients inthe gut lumen) and neurohumoral stimuli (gut hormones,neurotransmitters and neuromodulators), to the CNSthrough vagal and sympathetic nerves. In return, the vagalmotor fibers influence the enteric neural patterns, such asthe peristaltic reflex and the migrating motor complex.Vagal afferent nerves are therefore the major conduit bywhich nutrients signal to the brain and influence motilityand secretion, as well as hunger and satiety (Kontureket al., 2004; Thompson, 2006; Naslund and Hellstrom,2007).
The major route by which luminal contents signal to thebrainstem is via release of molecules (mostly peptides) fromenteroendocrine cells of the gut mucosa. These cells releasefrom their basolateral surface the signaling peptides intothe submucosa, where terminal fibers of the vagal afferentnerves are located. Receptors on the afferent fibers are thenactivated by the signaling molecules, thereby informing thebrainstem of luminal composition (Konturek et al., 2004;Badman and Flier, 2005; Thompson, 2006; Naslund andHellstrom, 2007). Some signals from the gut are transmit-ted from the nucleus of the solitary tract (NTS) to higherneural centers, such as the PVH and ARC nuclei of thehypothalamus, the bed nucleus of the stria terminalis andthe ventral thalamus. The integration of all these afferentsignals related to food presence in the gut, regulates the sizeof individual meals (Berthoud et al., 1990; Konturek et al.,2004; Badman and Flier, 2005; Naslund and Hellstrom,2007).
4. Gastrointestinal peptides regulating food intake
The gastrointestinal tract is the body’s largest endocrineorgan and produces around 30 different peptidic hormones,which act on several tissues, such as exocrine glands, gas-trointestinal tract and CNS (Table 1 summarizes somegut peptides regulating food intake). Almost all of thosehormones are sensitive to gut nutrient content and mostof them play an important role in the control of energy
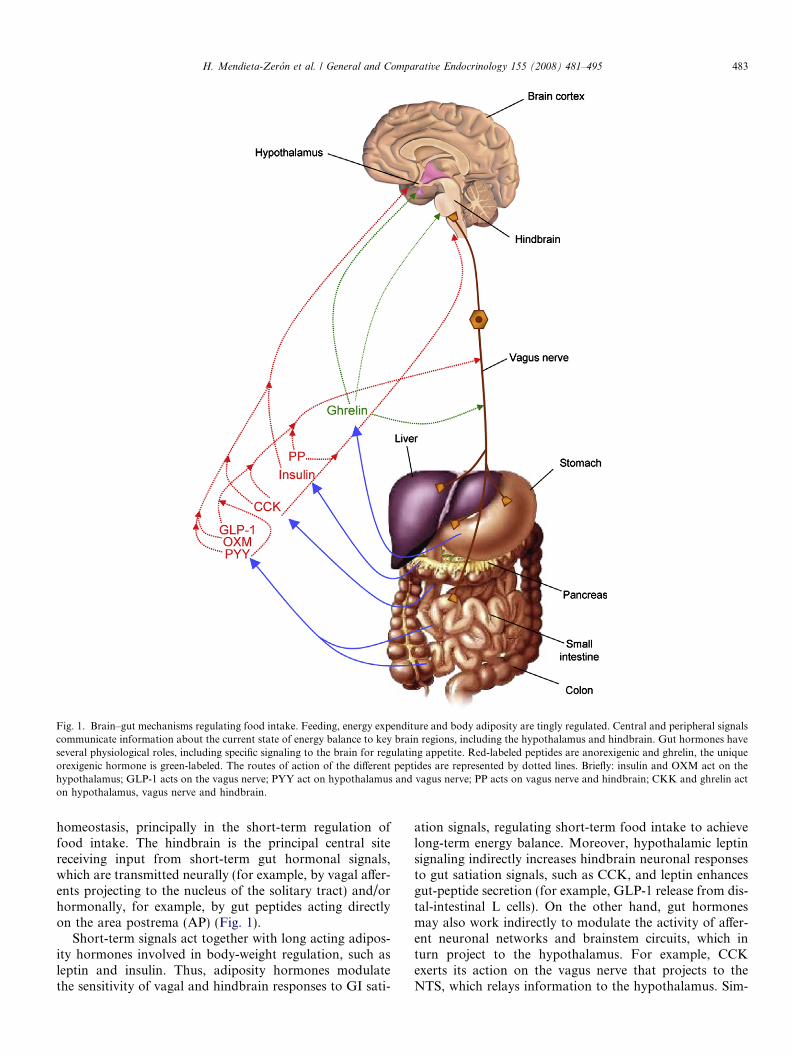
Fig. 1. Brain–gut mechanisms regulating food intake. Feeding, energy expenditure and body adiposity are tingly regulated. Central and peripheral signalscommunicate information about the current state of energy balance to key brain regions, including the hypothalamus and hindbrain. Gut hormones haveseveral physiological roles, including specific signaling to the brain for regulating appetite. Red-labeled peptides are anorexigenic and ghrelin, the uniqueorexigenic hormone is green-labeled. The routes of action of the different peptides are represented by dotted lines. Briefly: insulin and OXM act on thehypothalamus; GLP-1 acts on the vagus nerve; PYY act on hypothalamus and vagus nerve; PP acts on vagus nerve and hindbrain; CKK and ghrelin acton hypothalamus, vagus nerve and hindbrain.
H. Mendieta-Zeron et al. / General and Comparative Endocrinology 155 (2008) 481–495 483
homeostasis, principally in the short-term regulation offood intake. The hindbrain is the principal central sitereceiving input from short-term gut hormonal signals,which are transmitted neurally (for example, by vagal affer-ents projecting to the nucleus of the solitary tract) and/orhormonally, for example, by gut peptides acting directlyon the area postrema (AP) (Fig. 1).
Short-term signals act together with long acting adipos-ity hormones involved in body-weight regulation, such asleptin and insulin. Thus, adiposity hormones modulatethe sensitivity of vagal and hindbrain responses to GI sati-
ation signals, regulating short-term food intake to achievelong-term energy balance. Moreover, hypothalamic leptinsignaling indirectly increases hindbrain neuronal responsesto gut satiation signals, such as CCK, and leptin enhancesgut-peptide secretion (for example, GLP-1 release from dis-tal-intestinal L cells). On the other hand, gut hormonesmay also work indirectly to modulate the activity of affer-ent neuronal networks and brainstem circuits, which inturn project to the hypothalamus. For example, CCKexerts its action on the vagus nerve that projects to theNTS, which relays information to the hypothalamus. Sim-

Table 1Selected GI and pancreatic peptides that regulate food intake
Orexigenic: feedingstimulators
Anorexigenic: feeding inhibitors
Ghrelin Tschop et al.(2000)
Bombesin Gibbs et al. (1979)
Orexins (A and B)(Sakurai et al., 1998)
Cholecystokinin Gibbs et al. (1973)
Enterostatin Okada et al. (1992)Gastric leptin Zhang et al. (1994)Gastrin-releasing peptide Gibbs et al. (1979);Stein and Woods (1982)Glucagon-like peptide-1 Turton et al. (1996)Insulin Schwartz et al. (1992)Neuromedin B Ladenheim et al. (1994)Oxyntomodulin Dakin et al. (2004)Pancreatic polypeptide Malaisse-Lagae et al.(1977)Peptide YY3�36 Batterham et al. (2002)
484 H. Mendieta-Zeron et al. / General and Comparative Endocrinology 155 (2008) 481–495
ilarly, the GLP-1R-expressing neurons of the NTS projectto hypothalamic regions involved in appetite control suchas the ARC, DMH, and PVH (Badman and Flier, 2005;Perez-Tilve et al., 2006; Murphy and Bloom, 2006; Cum-mings and Overduin, 2007).
Next, we will review the knowledge about some gut pep-tides involved in food intake control.
4.1. Cholecystokinin (CCK)
CCK, secreted by proximal intestinal I-cells in the duo-denum and the jejunum into the circulation in response tonutrient ingestion (protein and fatty acids), was one of thefirst gastrointestinal hormones identified in regulatingenergy homeostasis (Gibbs et al., 1973a; Gibbs et al.,1973b; Larsson and Rehfeld, 1978). The major forms ofCCK in plasma are CCK-8, CCK-33, CCK-39, andCCK-58 (Reeve et al., 1994; Reeve et al., 2004; Stanleyet al., 2005; Murphy and Bloom, 2006; Cummings andOverduin, 2007). Once secreted, CCK reduces meal sizeand duration in both humans and rodents (Gibbs et al.,1973a; Muurahainen et al., 1988). and infusion of a CCKantagonist increases caloric intake in humans (Beglingeret al., 2001). However, despite its anorectic actions,repeated peripheral administration of CCK does not influ-ence body weight, as although meal frequency is increasedthere is no overall change in feeding (West et al., 1984).Thus, CCK is mostly involved in the short-term controlof food intake (Konturek et al., 2004; Badman and Flier,2005; Murphy and Bloom, 2006; Cummings and Overduin,2007). CCK is a pleiotropic hormone; besides its roles reg-ulating feeding, it is also involved in the regulation of pan-creatic and biliary secretion and gastrointestinal motility.The motor effects of CCK include postprandial inhibitionof gastric emptying and also inhibition of colonic transit(Konturek et al., 2004; Murphy and Bloom, 2006; Cum-mings and Overduin, 2007).
CCK signals via two distinct G protein-coupled recep-tors termed CCK1 and CCK2 (Wank et al., 1992a). Both
receptors are widely expressed in the CNS and in theperiphery (Moran et al., 1986; Wank et al., 1992b). Theeffect of CCK on food intake is mediated via CCK1 (Asinet al., 1992). CCK crosses the BBB (Reidelberger et al.,2004) and acts on NPY neurons in the DMH, as well asthe NTS (Moran et al., 1997; Bi et al., 2001). CCK effectson feeding are also mediated through paracrine and neuro-endocrine activation of vagal fibres (Moran et al., 1997).Consequently, vagotomy eradicates the satiating effect ofthis peptide in rats (Mercer and Lawrence, 1992). A rela-tionship between decreased plasma levels of CCK,increased hunger and decreased fullness has also beenreported in humans (French et al., 1993), indicating thatCCK acts as physiological mediator of satiety in man.
4.2. Insulin
Insulin secretion by the pancreas increases rapidly aftera meal, exerting an anorectic effect via the CNS (Schwartzet al., 2000). Insulin enters in the CNS via saturable, recep-tor-mediated transport across the BBB (Woods et al.,2003). Central administration of insulin or insulin mimeticreduces feeding and body weight in rodents and primates(Schwartz et al., 2000; Air et al., 2002). Administration ofantisense RNA against the insulin receptor (IR) induceshyperphagia and increases fat mass (Obici et al., 2002)and neuron-specific deletion of the IR results in obesity,hyperinsulinemia, and dyslipidemia in mice (Bruninget al., 2000). IRs are widespread in the brain and occurin hypothalamic nuclei involved in food intake (ARC,DMH and PVH) (Marks et al., 1990). In the hypothala-mus, the action of insulin on food intake and body weightis mediated by NPY (Schwartz et al., 2000) and the mela-nocortin system (Benoit et al., 2002).
4.3. Incretins: glucagon-like peptides (GLPs) and gastric
inhibitory polypeptide (GIP)
Preproglucagon gene is widely expressed in gut, pan-creas and NTS in the brainstem. Tissue-specific processingof preproglucagon by prohormone convertases (PCs) 1 and2 produces different products: glucagon is the main productin the pancreas and glucagon-like peptide-1 (GLP-1), glu-cagon-like peptide-2 (GLP-2) and oxyntomodulin (OXM)are the major products in CNS and gut (Tang-Christensenet al., 2001; Stanley et al., 2005). GLP-1 and OXM arereleased from distal intestinal L cells in the mucosa of theileum and colon in response to free fatty acids (FFAs)and carbohydrates (Hirasawa et al., 2005). Both peptidesinhibit feeding when they are centrally or peripherallyadministrated (Dakin et al., 2004; Murphy and Bloom,2006). Furthermore, chronic administration of GLP-1and OXM decreases weight gain and adiposity in rodents(Meeran et al., 1999; Dakin et al., 2004) and humans (Flintet al., 1998; Cohen et al., 2003; Wynne et al., 2005). On theother hand, continuous administration of GLP-1 antago-nist (namely, exendin9-39, see below) increases not only

H. Mendieta-Zeron et al. / General and Comparative Endocrinology 155 (2008) 481–495 485
feeding, but also body weight (Turton et al., 1996). In addi-tion to their effects on feeding, GLP-1 also delays gastricemptying by reducing gastric motility (Flint et al., 2001),inhibits gastric secretion (Wynne et al., 2004), inhibits glu-cagon (Drucker, 2003), and may improve insulin sensitivity(anti-diabetogenic effects) (D’Alessio et al., 1994). OXMalso reduce gastric motility (Wynne et al., 2004).
GLP-1 exists in a number of forms in the circulation.The action of GLP-17�36amide, (the most common circulat-ing form) that has attracted most attention, both from aphysiological and a therapeutic viewpoint, is its potentincretin effect: central or peripheral administration stronglystimulates insulin release (Murphy and Bloom, 2006; Cum-mings and Overduin, 2007). Consistent with its role as anincretin, GLP-17�36amide is released into the circulation inresponse to a meal in proportion to the calories ingested(Murphy and Bloom, 2006; Cummings and Overduin,2007). Although the majority of L-cells are located in thedistal gut, the presence of nutrients in the proximal smallintestine stimulates GLP-17�36amide release independentlyof the presence of nutrients within the ileum and colon(Roberge and Brubaker, 1991). This effect is abolished byvagotomy, implying the influence of neural inputs onGLP-17�36amide release (Rocca and Brubaker, 1999).
The actions of both GLP-1 and OXM on feeding may bemediated via the GLP-1 receptor (GLP-1R), expressed inthe hypothalamus, brainstem and periphery (Bullocket al., 1996). However, both peptides have different CNStargets: OXM activates neurons in the hypothalamus(Dakin et al., 2004), while GLP-1 acts in the hindbrainand other autonomic control areas, via vagus nerve, inrodents and humans (Nauck et al., 1997; Wettergrenet al., 1997; Imeryuz et al., 1997; Wettergren et al., 1998;Yamamoto et al., 2002). Vagal deafferentation or section-ing eliminates anorectic effects of GLP-1 (Abbott et al.,2005; Talsania et al., 2005). Curiously, food intake andbody weight of GLP-1-receptor-knockout mice are normal(Murphy and Bloom, 2004). Despite this evidence, it hasbeen reported that some of the anorectic effects of GLP-1may be related to taste aversion and visceral illness (Thieleet al., 1997; Lachey et al., 2005).
GLP-2 is mainly expressed in the brain and inhibits foodintake when administered centrally possibly through acti-vations of GLP-1 receptors (Badman and Flier, 2005).Peripheral circulating GLP-2, is primary involved in stim-ulating gut motility, absorption and growth; however, itdoes not appear to affect energy intake either in rodentsor humans (Schmidt et al., 2003; Murphy and Bloom,2006; Cummings and Overduin, 2007).
Gastric inhibitory polypeptide or glucose-dependentinsulinotropic peptide (GIP) is synthesized and releasedby the K-cells of the duodenum and proximal jejunum inresponse to glucose and fat ingestion (Wynne et al., 2004;Badman and Flier, 2005; Baggio and Drucker, 2007). Itis the rate of nutrient absorption rather than the mere pres-ence of nutrients in the intestine that stimulates GIPrelease. Thus, GIP secretion is reduced in individuals with
intestinal malabsorption or after the administration ofpharmacologic agents that reduce nutrient absorption(Besterman et al., 1979; Fushiki et al., 1992; Baggio andDrucker, 2007). As an incretin, GIP improve the responseof the endocrine pancreas to absorbed nutrients (Badmanand Flier, 2005; Baggio and Drucker, 2007). Remarkably,although feeding is not affected by centrally administrationof GIP (Badman and Flier, 2005), this peptide has multipleeffects on adipocytes by increasing lipogenesis, on bone for-mation and on CNS, regulating neuronal proliferation andinfluencing several aspects related to learning/memory andsensorimotor coordination (Badman and Flier, 2005; Bag-gio and Drucker, 2007).
GIP binds to a G-coupled receptor, known as GIP-R(Baggio and Drucker, 2007). Mice fed with high fat diet(HFD) have increased GIP together with obesity (Wynneet al., 2004; Badman and Flier, 2005), while mice lackingthe GIP receptor (GIP-R) are protected against obesityinduced by both HFD and leptin-deficiency (ob/ob mice)(Miyawaki et al., 2002). Thus, GIP may be involved inthe development of obesity in response to high fat intake.Further work is needed to clarify the mechanism by whichGIP promotes efficient storage of ingested energy as fat.Inhibition of this system may provide a useful anti-obesitytherapeutic strategy.
4.4. PP-fold family: Pancreatic polypeptide (PP) and
peptide YY (PYY)
PP and PYY belong to the PP-fold family of peptideswhich also includes NPY (Conlon, 2002). Five receptorsfor the PP-fold family of peptides have been cloned inmammals Y1R, Y2R, Y4R, Y5R, and Y6R, which is trun-cated and non-functional in humans (Kalra and Kalra,2004). All are coupled to inhibitory G-proteins (Gi), andtherefore mediate an inhibition of intracellular cyclic-aden-osine monophosphate (cAMP) synthesis (Mullins et al.,2002).
PP is mainly produced by peripheral cells of the islets ofLangerhans, exocrine pancreas and distal GIT (Small andBloom, 2004; Stanley et al., 2005; Cummings and Overdu-in, 2007). Plasma PP concentrations increase proportion-ally to caloric intake (Small and Bloom, 2004) and theyappear to be inversely proportional to adiposity, with highlevels in anorexic subjects and reduced levels in obese sub-jects (Fujimoto et al., 1997). Both basal and postprandialPP release is subject to control by the vagus nerve (Katsu-ura et al., 2002). Peripheral PP administration reducesfeeding and body weight in obese rodents (Malaisse-Lagaeet al., 1977) and feeding in humans independently of gastricemptying (Batterham et al., 2003). Apart from its acuteeffects on appetite and food intake, PP may also modulatelong-term energy balance. Chronic administration of PP toob/ob mice does decrease body weight gain and amelioratesinsulin resistance and dyslipidemia (Asakawa et al., 2003).The acute effects of PP on appetite and its long-termactions on body weight in human obesity are unknown.

486 H. Mendieta-Zeron et al. / General and Comparative Endocrinology 155 (2008) 481–495
PP also regulates gastric motility by inhibiting gastric emp-tying when centrally administered (Murphy and Bloom,2004; Murphy and Bloom, 2006). The anorectic effect ofPP is exerted via brainstem pathways in the AP and is med-iated by Y4R. In contrast to the peripheral actions, centraladministration of PP increases food intake (Clark et al.,1984); the receptors mediating this action and the mecha-nisms involved are unclear, but it has been hypothethizedthat this effect is mediated by Y5R receptor in the hypo-thalamus (Wynne et al., 2004). However, the physiologicalsignificance of this orexigenic effect is unclear, since PPdoes not easily cross the BBB (Whitcomb et al., 1990;Banks et al., 1995).
PYY is secreted postprandially by the L cells of the gas-trointestinal tract, especially in the most distal portions,such as the ileum, colon and rectum, and its secretion iscorrelated with caloric intake (Stanley et al., 2005). Thereare two main forms of PYY in the circulation: PYY1�36
and PYY3�36 (Grandt et al., 1994; Batterham et al.,2002; Wynne et al., 2004; Stanley et al., 2005). PYY 1�36
binds Y1R, Y2R, Y4R, and Y5R receptors while PYY3�36
binds more specifically Y2R receptors (Beckoff et al.,2001). Peripheral administration of PYY has severalactions including: delay gastric emptying and gastric secre-tion, modulate bowel motility, increase ileum absorptionand act on the dorsal vagal complex (DVC) to modulategastrointestinal motility through Y1R and Y2R (Buchanet al., 1978; Al Saffar et al., 1985; Savage et al., 1987). Ithas been also reported that peripheral administration ofPYY3�36 inhibits food intake and reduces weight gain inrodents, primates and humans (Batterham et al., 2002;Challis et al., 2003; Moran et al., 2005). Chronic PYY3�36
administration reduced food intake and body weight gain(Batterham et al., 2002; Wynne et al., 2004).
PYY crosses the BBB and probably exerts its actions viathe presynaptic Y2R receptor of AgRP/NPY neurons inthe ARC, releasing inhibition of POMC neurons and con-sequently inhibiting feeding (Broberger et al., 1997; Batter-ham et al., 2002; Challis et al., 2003; Talsania et al., 2005).Besides this hypothalamic-mediated mechanisms, some evi-dence also support a vagal mediation for PYY: Y2R isexpressed by vagal-afferent terminals (Koda et al., 2005),and the anorectic effects and arcuate neuronal activationinduced by peripheral PYY3�36 were suppressed by vagot-omy or destruction of hindbrain-hypothalamic pathways(Koda et al., 2005; Abbott et al., 2005). Consistent withthese findings, PYY-knockout mice have disrupted energyhomeostasis, suggesting that the PYY system has a physi-ological role in its regulation (Boey et al., 2006; Batterhamet al., 2006).
Finally, it is interesting to note that despite this evi-dence, for some authors the anorectic effect of PYY3�36
is controversial, not easily duplicated and very vulnerableof external variables, such as animal handling, stress, routeand timing of administration (Coll et al., 2004; Tschopet al., 2004). Indeed, in contrast to peripheral injection,central administration of both PYY1�36 and PYY3�36 stim-
ulates feeding in rodents (Stanley et al., 1985; Clark et al.,1987; Hagan et al., 1998; Corpa et al., 2001), probablythrough direct activation of Y1R and Y5R in the PVH(Cummings and Overduin, 2007). It has also been sug-gested that the anorexic effect of PYY may be partiallymediated by an aversive response (Halatchev and Cone,2005).
4.5. Ghrelin
Ghrelin, initially identified as the endogenous ligand ofthe growth hormone secretagogue receptor (GHS-R) (Koj-ima et al., 1999), is a 28-amino acid acylated hormonemainly synthesized and secreted by the gut, in the gastricoxyntic cells (A/X cells) at the fundus of the stomach, aswell as, the duodenum, ileum, cecum and colon (Kojimaet al., 1999; Gualillo et al., 2003). Ghrelin exerts pleiotropicactions, such as regulating food intake (Kojima et al.,1999), GH secretion (Arvat et al., 2000; Peino et al.,2000; Seoane et al., 2000; Gualillo et al., 2003) and gastricsecretion and motility (Masuda et al., 2000).
Structural analysis of ghrelin molecule identified that thehydroxyl group of the third N-terminal amino acid serineresidue is esterified by an n-octanoic acid (Kojima et al.,1999). No other naturally occurring peptide has been pre-viously shown to have this acyl group as a post-transla-tional modification. A spliced variant of ghrelin with 27amino acids, missing the 14th amino acid (glutamine),was also identified. Later, biologically active analogues ofghrelin were described in much smaller amounts with acylchains of 10 or 11 C atoms or carbon unsaturations (Hos-oda et al., 2000b). The n-octanoyl group at the Ser3 of theghrelin molecule seems to be essential for some of the hor-mone’s bioactivity, including GH release and orexigeniceffect. Unacylated ghrelin (UAG, also called desoctanoylor desacyl) circulates in far greater concentrations thanthe acylated form, does not displace ghrelin from its hypo-thalamic and pituitary binding sites (Hosoda et al., 2000a)and is unable to stimulate GH release in rats and humans(Hosoda et al., 2000a; Broglio et al., 2003). Although inthe beginning it was widely accepted that UAG was devoidof any biological activity, increasing numbers of studiesreported different biological effects, and it is now consid-ered as a peptide with specific and wide ranging actionsin different tissues, such as regulation of cell proliferation(Cassoni et al., 2001; Nanzer et al., 2004), cardiac effects(Bedendi et al., 2003), reproduction (Martini et al., 2006;Garcıa et al., 2007), and adipogenesis (Thompson et al.,2004).
In addition to its unique post-translational modification,ghrelin displays another exclusive characteristic; so far, it isthe unique known orexigenic hormone, which confers it apredominant role on feeding control. In fact, since plasmaghrelin levels rise shortly before meals, it has been sug-gested that ghrelin might be an important signal to preparefor meal initiation in humans and rodents (Cummingset al., 2001; Drazen et al., 2006). Chronic administration

H. Mendieta-Zeron et al. / General and Comparative Endocrinology 155 (2008) 481–495 487
of ghrelin induces obesity in mice by increasing food intakeand reducing fat utilization in mice and rats. Morerecently, it has been shown that central administration ofghrelin exerts peripheral effects by promoting lipid storagein white adipose tissue, an effect that is associated withincreased expression of lipogenic enzymes and decreasedlipid oxidation (Theander-Carrillo et al., 2006). On thisbasis, ghrelin has been widely proposed as a potential ther-apeutic target against obesity and it has been recentlyshown that vaccination against the endogenous ghrelin,by using a combination of ghrelin immunoconjugates,slows weight gain in rats by decreasing feed efficiency (Hor-lick et al., 2000). Thus, identification of the molecularmechanisms mediating ghrelin effects on feeding may pro-vide new therapeutic targets for obesity.
The effects of ghrelin on feeding and GH secretion aremediated via type 1a GHS-R (Kojima et al., 1999; Gualilloet al., 2003; Nogueiras et al., 2004). However, the orexi-genic effects of ghrelin are independent of GH-releasingproperties (Tschop et al., 2000; Shintani et al., 2001). Inthe CNS the action of ghrelin on feeding is mainly exertedvia the ARC. GHS-R mRNA is expressed in neurons in theARC co-expressing NPY and AgRP (Zigman et al., 2006)and the central administration of ghrelin increases themRNA content of NPY and AgRP in the ARC in fedand fasting conditions (Nakazato et al., 2001; Seoaneet al., 2003). Recent data also indicate that ghrelin acts inthe hypothalamus by altering fatty acid metabolism andAMP-activated protein kinase (AMPK). It has been dem-onstrated that ghrelin increases hypothalamic AMPKphosphorylation levels, activating it. This action may beassociated to specific changes in hypothalamic neuropep-tides, although the exact molecular mechanisms and ana-tomical details of this interaction have not been fullyidentified (Andersson et al., 2004; Kola et al., 2005; Lopezet al., 2007a). Some evidence has also demonstrated thatorexin neurons in the lateral hypothalamic area (LHA)(Toshinai et al., 2003) and neurons in the NTS and AP,in the brainstem, may mediate the orexigenic actions ofghrelin (Nakazato et al., 2001) but this action is controver-sial (Seoane et al., 2003).
Despite ghrelin having a potent action in regulatingfood intake, both ghrelin knockout mice and mice lackingGHS-R type 1a have normal feeding patterns and bodycomposition on a standard diet (Sun et al., 2003). However,on HFD, the absence of ghrelin (Wortley et al., 2005) orthe ghrelin receptor (Zigman et al., 2005) protects againstearly-onset obesity; in both cases this reduced weight gainis associated with decreased adiposity and increased energyexpenditure and locomotor activity. These data suggestthat ghrelin, like leptin, may play an important role inthe development of hypothalamic systems regulatingenergy balance (Grove and Cowley, 2005). Very interest-ingly, ablation of ghrelin improves the diabetic but notobese phenotype of ob/ob mice (Sun et al., 2006).
Ghrelin also regulated energy homeostasis in humans.Intravenous administration of ghrelin to healthy volunteers
increases food intake (Wren et al., 2001). Moreover, therise in preprandial ghrelin correlates with hunger scoresin humans eating spontaneously (Cummings et al., 2004).Interestingly, the levels of ghrelin are correlated with adi-posity in humans, with an inverse relationship betweenplasma ghrelin levels and BMI (Tschop et al., 2001). Obesehumans show reduced levels of plasma ghrelin which rise tonormal after diet-induced weight loss (Cummings et al.,2002). Moreover, in obese individuals the postprandial reg-ulation of ghrelin seems to be altered, which may be relatedto continuous food intake and/or obesity (English et al.,2002). The severe hyperphagia seen in patients with Prad-er-Willi syndrome is associated with elevated ghrelin levels,in contrast to other forms of obesity where low ghrelin lev-els are seen (Cummings et al., 2002).
Finally, it was recently reported that obestatin a newpeptide derived from the ghrelin gene inhibits food intake,decreases gastric emptying and weight gain, by actingthrough the orphan receptor GPR39 (Zhang et al., 2005;Bresciani et al., 2006; Green et al., 2007; Lagaud et al.,2007). In spite of this evidence, there are some discrepan-cies about the anorectic effect of obestatin (Seoane et al.,2006; Sibilia et al., 2006; Nogueiras et al., 2007; Chartrelet al., 2007; Zizzari et al., 2007) as well as its binding toGPR39 (Holst et al., 2007; Tremblay et al., 2007; Zhanget al., 2007), suggesting that obestatin might not have a rolein the regulation of food intake (Gourcerol et al., 2007a;Gourcerol and Tache, 2007b).
Obestatin results from proteolysis of proghrelin at twocleavage sites, between arginine 75 and phenylalanine 76,and between lysine 100 and phenylalanine 101; it is sug-gested that arginine 75 and lysine 100 predict the monoba-sic recognition sites for an unknown protease besides theknown prohormone convertases (Zhang et al., 2005). Inter-estingly, in vitro digestion of proghrelin using several con-vertases failed to produce obestatin (Garg, 2007). Onaddition, the fact that monkeys and baboons have gluta-mine instead of arginine at position 75 suggests that thesespecies do not produce obestatin. Moreover, the relativeefficiencies for processing of proghrelin into ghrelin orobestatin in the stomach or other tissues under a varietyof conditions are also still not clear. Thus, even consideringthis broader definition of a hormone, controversies aboutobestatin and its action indicate that the main evidencedoes not support its consideration as a hormone until moreconvincing data appear.
4.6. Bombesin (BBS)
BBS is a peptide widely distributed in the mammaliangut and plasma levels of BBS markedly increase after foodintake (Gibbs et al., 1979; Wynne et al., 2004). Peripheraland central administration of BBS is anorectic and it isnot suppressed by vagotomy (Gibbs et al., 1979; Smithet al., 1981). Chronic administration of BBS reduces bodyweight (West et al., 1982). BBS is structurally very similarto gastrin-releasing peptide (GRP) and neuromedin B

488 H. Mendieta-Zeron et al. / General and Comparative Endocrinology 155 (2008) 481–495
(NMB); and binds to their receptors (GRP-R and NMB-R,respectively). Additionally, a bombesin-3 receptor (BBSR-3) has been cloned (Ladenheim et al., 1997). Knocking outBBSR-3 induces moderate hyperphagia, obesity and meta-bolic alterations in mice (Ohki-Hamazaki et al., 1997).
4.7. Orexins or hypocretins
OX-A (hypocretin-1) and orexin-B (OX-B, hypocretin-2)are peptides derived from the same 130 amino acid long pre-cursor (prepro-orexin) that bind and activate two closelyrelated orphan G protein-coupled receptors: orexin receptor1 (OX1R) and orexin receptor 2 (OX2R) (Willie et al., 2001;Lopez et al., 2005). They are considered to be neurotrans-mitters that centrally stimulate food intake in several species(Willie et al., 2001; Lopez et al., 2005), and are also involvedin energy homeostasis, sleep–awake behavior, nociception,reward seeking, food and drug addiction, as well as repro-duction, cardiovascular and adrenal function (Willie et al.,2001; Lopez et al., 2005). Despite their effects on feedingwhen centrally administered, peripheral injections (acuteand chronic) of orexin do no elicit changes in food intakein rodents (Haynes et al., 1999). Curiously, in pigs subcuta-neous administration of OX-B stimulates feeding (Dyeret al., 1999). Orexins have been found to affect gastrointes-tinal motility and gastric, intestinal and pancreatic secre-tions (Kirchgessner and Liu, 1999; Kirchgessner, 2002).These effects were observed following central (intracerebro-ventricular) or local (intraluminal, intraarterial), but notintravenous administrations of orexins (Korczynski et al.,2006a; Korczynski et al., 2006b).
In the GIT the orexin system is highly expressed inendocrine cells. Thus, OX-A immunoreactivity is detectedin numerous endocrine enterochromaffin cells of the gastricand intestinal mucosa, as well as in the a and b cells of thepancreas (Kirchgessner and Liu, 1999; de Miguel and Bur-rell, 2002; Naslund et al., 2002; Nakabayashi et al., 2003;Ouedraogo et al., 2003; Ehrstrom et al., 2005). Since theexpression of orexins in the GIT and bloodstream isenhanced during fasting (Komaki et al., 2001), and fastingreveals many of the orexin gastrointestinal effects, it seemsprobable that on the local level, orexins keep the GIT func-tions ready during fasting and play role in brain–gut axiscontrol. Furthermore, orexin fibers innervate the DVC, akey site in the control of gastrointestinal function (Grab-auskas and Moises, 2003) where OX-A could be involvedin the responses to CCK (Burdyga et al., 2003). Despitethis evidence, there are no reported data about gut functionalteration either in orexin or orexin receptors knockoutmice. Further work with these models will help to fullyunderstand the physiological role of orexin system in gutphysiology.
4.8. Corticotropin-releasing hormone (CRH)
CRH was the first peptide isolated (Vale et al., 1981) ofa family of mammalian CRH-related peptides that now
includes urocortin 1, urocortin 2 (also known as stressc-opin-related peptide), and urocortin 3 (also known asstresscopin). CRH and the urocortins exert their biologicalactions on target cells through activation of 2 seven-trans-membrane-domain G protein-coupled receptors, known asCRH receptor type 1 (CRH1) and CRH receptor type 2(CRH2), which are encoded by 2 distinct genes (Hillhouseand Grammatopoulos, 2006).
Besides its central actions regulating feeding and stressresponses related to adrenal axis (Kalra et al., 1999; Hill-house and Grammatopoulos, 2006), CRH ligands andreceptors play a major role on gut function. Supportingthis idea, CRH ligands and receptors have been detectedin the gut myenteric nervous system, as well as in enteroen-docrine cells and lamina propia in rodents and humans(Muramatsu et al., 2000; Lewis et al., 2001; Chatzakiet al., 2004a; Chatzaki et al., 2004b; Chatzaki et al.,2006). CRH and urocortins exert inhibitory effects on gas-tric, duodenal and small intestine transit. This action isdirectly mediated by vagal nerves and it is independent ofactivation of adrenal axis (for extensive review: Tacheand Bonaz, 2007). On the other hand, CRH and urocortinsstimulate colonic transit and defecation and induce diar-rhea through increased sacral parasympathetic outflow tothe large intestine in rodents. The PVH, the locus coeruleus(LC) and the Barrington’s nucleus are the brain areaswhere CRH and stress stimulated colonic function (forextensive review: Tache and Bonaz, 2007).
In addition to their role in gut motility, both central andperipheral injection of CRH and urocortins reproducesstress-related alterations of gut motor function (gastric,small intestine, and colonic) in rodents, whereas centralinjection of CRH antagonists prevents the effects of variousstressors, suggesting a crucial role for CRH receptors in thebrain in the regulation of stress-induced alterations in gas-trointestinal motility (for extensive review: Tache andBonaz, 2007). Pharmacological studies support the notionthat gut CRH signaling occurs under stress (Williamset al., 1987; Tache et al., 2004a; Tache and Perdue,2004b). Because both central and peripheral administrationof CRH receptor antagonists is able to counteract theimpact of stress on gut motility, this supports the conceptthat stress influences the release of CRH ligands in thegut through autonomic pathways, where they can thenfunction as local effectors of altered gastrointestinal motil-ity (Tache and Bonaz, 2007). Some reports have showedthat the delayed gastric emptying induced by abdominalsurgery can be blocked by peripheral injection of CRHantagonist, such as a-helical CRH9�41, D-Phe12CRH12�41,and astressin (Martinez et al., 1999). With respect to stress-related stimulation of colonic motor function, peripheraladministration of a-helical CRH9�41, astressin, and CP-154,526, but not of astressin2-B, prevented or blunted thestimulation of distal colonic transit and defecation inducedby acute stress (Williams et al., 1987; Million et al., 2002).In patients with irritable bowel syndrome, compared withhealthy subjects, the administration of a-helical CRH9�41

H. Mendieta-Zeron et al. / General and Comparative Endocrinology 155 (2008) 481–495 489
improves colonic motility without affecting the hypotha-lamic–pituitary–adrenal axis (Sagami et al., 2004).
Finally, it is interesting to note that recent data haveimplicated both urocortin-1 and -2 in inhibiting feedingand gastric motility through CRH2 (Asakawa et al.,2001; Wang et al., 2001; Nagata et al., 2005; Czimmeret al., 2006).
5. Treatment perspectives
Gut–brain peptides may provide attractive therapeutictargets in the fight against obesity (Crowell et al., 2006;Foster-Schubert and Cummings, 2006). In this regard,one of the earliest endocrine mechanisms activated by foodintake is the insulinotropic incretin system (Drucker, 2003),since both GLP-1 and GIP highly susceptible to enzymaticdegradation in vivo by the enzyme dipeptidyl peptidase IV(DPP-IV). Therefore, modulation of incretin levels by theuse of DPP-IV inhibitors is already used for the treatmentof type 2 diabetes mellitus (Deacon, 2004).
The saliva of the Gila monster lizard (Heloderma suspec-
tum) contains a peptide called exendin-4 (EX-4), which is apotent GLP-1 agonist, with anorexigenic and fat-reducingproperties; a truncated form of this peptide, exendin9�39
(EX9-39), acts as a competitive antagonist at the samereceptor (Murphy and Bloom, 2004; Cummings and Over-duin, 2007). Clinical trials have shown that exendin-4 (alsoknown as exenatide, and marketed as Byetta) is useful forglycaemic control in people with type 2 diabetes mellitus.Interestingly, in phase III 30-week clinical trials, exenatidesignificantly reduced body weight in treated diabetics (Buseet al., 2004; DeFronzo et al., 2005; Kendall et al., 2005).Some evidence has shown that the action of EX-4 maybe related to modulation of ghrelin levels: central andperipheral administration of EX-4 reduces the levels ofghrelin by up to 74% in fasted rats (Perez-Tilve et al.,2007). These effects are dose dependent and long lasting(up to 8 h). Suppression of ghrelin was neither mimickedby GLP-1, GLP-17�36amide nor blocked by the GLP-1Rantagonist EX9-39. Moreover, it was independent of thelevels of leptin and insulin. The potent effects of EX-4 onghrelin make it tempting to speculate that EX-4 could offera therapeutic option for Prader Willi Syndrome and otherdiseases characterized by substantial amounts of circulat-ing ghrelin (Cummings et al., 2002). These results suggestthat GLP-1 system might be used to reduce body weightusing a peripherally administered drug.
Bariatric surgery can get considerable and long-termweight loss and can improve obesity associated alterations,such as diabetes, hyperlipidemia, hypertension, andobstructive sleep apnea (Badman and Flier, 2005; Le Rouxet al., 2006). The most common procedures for bariatricsurgery are jejunoileal bypass, biliopancreatic diversion,duodenal switch and Roux-en-Y gastric bypass (RYGB).In addition to the reductions stomach capacity andabsorption, as well as vagal transection, one of the primaryconsequences of gastric surgery is the alteration of the
levels of gut hormones. Increases in the levels of PYYand GLP-1 have been reported (Korner and Aronne,2004). On the other hand, decreases in plasmatic levels ofghrelin (Cummings et al., 2002; Cummings et al., 2005)and GIP are observed after RYGB surgery (Le Rouxet al., 2006), which is thought be partly responsible forthe suppression of appetite and weight loss seen after theseoperations, suggesting that ghrelin and GIP receptorantagonism may afford an alternative therapeutic optionfor treatment of obesity-diabetes (Flatt, 2007). Further-more, it is interesting to note that RYGB patients had earlyand exaggerated insulin responses, potentially mediatingimproved glycaemic control (Le Roux et al., 2006). In sum-mary, pleiotropic endocrine responses could contribute tothe improved appetite reduction, and long-term changesin body weight after bariatric surgery.
6. Conclusions
One of the main routes of communication between brainand gut in relation to energy homeostasis is via peptides.Multiple, redundant and complex peripheral-neural cir-cuits participate in the regulation of food intake and bodyweight homeostasis (Table 1 and Fig. 1). All this evidenceindicates that obesity, and associated metabolic alterations,is a complex, multifactorial and chronic pathology. Thismakes the search for and development of new weight-lossdrugs extremely complicated. In fact, the efficacy of drugsacting on a single molecular target might be limited bycompensatory feedback mechanism. In the near future,we should look for combined therapies that act on bothperipheral and central targets. Understanding these molec-ular networks regulating food intake, which include gutpeptides or gut-peptide mimetics, could lead to design bet-ter therapeutic targets for weight loss. Finally, mechanismsof weight loss following bariatric surgery require furtherstudies and a better understanding.
Acknowledgments
Dr. Hugo Mendieta-Zeron is funded by National Coun-cil of Science and Technology (CONACYT), Mexico,Scholarship. Young Talents Program, Autonomous Uni-versity of the State of Mexico (UAEMEX). Dr. Miguel Lo-pez is funded by Xunta de Galicia and Fondo deInvestigationes Sanitarias. Dr. Carlos Dieguez is fundedby European Union, Xunta de Galicia, and Fondo deInvestigationes Sanitarias.
References
Abbott, C.R., Monteiro, M., Small, C.J., Sajedi, A., Smith, K.L.,Parkinson, J.R., Ghatei, M.A., Bloom, S.R., 2005. The inhibitoryeffects of peripheral administration of peptide YY(3-36) and glucagon-like peptide-1 on food intake are attenuated by ablation of the vagal–brainstem–hypothalamic pathway. Brain Res. 1044, 127–131.
Air, E.L., Strowski, M.Z., Benoit, S.C., Conarello, S.L., Salituro, G.M.,Guan, X.M., Liu, K., Woods, S.C., Zhang, B.B., 2002. Small molecule

490 H. Mendieta-Zeron et al. / General and Comparative Endocrinology 155 (2008) 481–495
insulin mimetics reduce food intake and body weight and preventdevelopment of obesity. Nat. Med. 8, 179–183.
Al Saffar, A., Hellstrom, P.M., Nylander, G., 1985. Correlation betweenpeptide YY-induced myoelectric activity and transit of small-intestinalcontents in rats. Scand. J. Gastroenterol. 20, 577–582.
Anand, B.K., Brobeck, J.R., 1951a. Hypothalamic control of food intakein rats and cats. Yale J. Biol. Med. 24, 123–140.
Anand, B.K., Brobeck, J.R., 1951b. Localization of a ‘‘feeding center’’ inthe hypothalamus of the rat. Proc. Soc. Exp. Biol. Med. 77, 323–324.
Andersson, U., Filipsson, K., Abbott, C.R., Woods, A., Smith, K.,Bloom, S.R., Carling, D., Small, C.J., 2004. AMP-activated proteinkinase plays a role in the control of food intake. J. Biol. Chem. 279,12005–12008.
Arvat, E., Di Vito, L., Broglio, F., Papotti, M., Muccioli, G., Dieguez, C.,Casanueva, F.F., Deghenghi, R., Camanni, F., Ghigo, E., 2000.Preliminary evidence that ghrelin, the natural GH secretagogue(GHS)-receptor ligand, strongly stimulates GH secretion in humans.J. Endocrinol. Invest. 23, 493–495.
Asakawa, A., Inui, A., Ueno, N., Makino, S., Fujimiya, M., Fujino, M.A.,Kasuga, M., 2001. Urocortin reduces oxygen consumption in lean andob/ob mice. Int. J. Mol. Med. 7, 539–541.
Asakawa, A., Inui, A., Yuzuriha, H., Ueno, N., Katsuura, G., Fujimiya,M., Fujino, M.A., Niijima, A., Meguid, M.M., Kasuga, M., 2003.Characterization of the effects of pancreatic polypeptide in theregulation of energy balance. Gastroenterology 124, 1325–1336.
Asin, K.E., Gore Jr., P.A., Bednarz, L., Holladay, M., Nadzan, A.M.,1992. Effects of selective CCK receptor agonists on food intake aftercentral or peripheral administration in rats. Brain Res. 571, 169–174.
Badman, M.K., Flier, J.S., 2005. The gut and energy balance: visceralallies in the obesity wars. Science 307, 1909–1914.
Baggio, L.L., Drucker, D.J., 2007. Biology of incretins: GLP-1 and GIP.Gastroenterology 132, 2131–2157.
Banks, W.A., Kastin, A.J., Jaspan, J.B., 1995. Regional variation intransport of pancreatic polypeptide across the blood–brain barrier ofmice. Pharmacol. Biochem. Behav. 51, 139–147.
Batterham, R.L., Cowley, M.A., Small, C.J., Herzog, H., Cohen, M.A.,Dakin, C.L., Wren, A.M., Brynes, A.E., Low, M.J., Ghatei, M.A.,Cone, R.D., Bloom, S.R., 2002. Gut hormone PYY(3-36) physiolog-ically inhibits food intake. Nature 418, 650–654.
Batterham, R.L., Heffron, H., Kapoor, S., Chivers, J.E., Chandarana, K.,Herzog, H., Le Roux, C.W., Thomas, E.L., Bell, J.D., Withers, D.J.,2006. Critical role for peptide YY in protein-mediated satiation andbody-weight regulation. Cell Metab. 4, 223–233.
Batterham, R.L., Le Roux, C.W., Cohen, M.A., Park, A.J., Ellis, S.M.,Patterson, M., Frost, G.S., Ghatei, M.A., Bloom, S.R., 2003.Pancreatic polypeptide reduces appetite and food intake in humans.J. Clin. Endocrinol. Metab. 88, 3989–3992.
Beckoff, K., MacIntosh, C.G., Chapman, I.M., Wishart, J.M., Morris,H.A., Horowitz, M., Jones, K.L., 2001. Effects of glucose supplemen-tation on gastric emptying, blood glucose homeostasis, and appetite inthe elderly. Am. J. Physiol. Regul. Integr. Comp. Physiol. 280, R570–R576.
Bedendi, I., Alloatti, G., Marcantoni, A., Malan, D., Catapano, F., Ghe,C., Deghenghi, R., Ghigo, E., Muccioli, G., 2003. Cardiac effects ofghrelin and its endogenous derivatives des-octanoyl ghrelin and des-Gln14-ghrelin. Eur. J. Pharmacol. 476, 87–95.
Beglinger, C., Degen, L., Matzinger, D., D’Amato, M., Drewe, J., 2001.Loxiglumide, a CCK-A receptor antagonist, stimulates calorie intakeand hunger feelings in humans. Am. J. Physiol. Regul. Integr. Comp.Physiol. 280, R1149–R1154.
Benoit, S.C., Air, E.L., Coolen, L.M., Strauss, R., Jackman, A., Clegg,D.J., Seeley, R.J., Woods, S.C., 2002. The catabolic action of insulinin the brain is mediated by melanocortins. J. Neurosci. 22, 9048–9052.
Berthoud, H.R., Jedrzejewska, A., Powley, T.L., 1990. Simultaneouslabeling of vagal innervation of the gut and afferent projections fromthe visceral forebrain with dil injected into the dorsal vagal complex inthe rat. J. Comp. Neurol. 301, 65–79.
Besterman, H.S., Cook, G.C., Sarson, D.L., Christofides, N.D., Bryant,M.G., Gregor, M., Bloom, S.R., 1979. Gut hormones in tropicalmalabsorption. Br. Med. J. 2, 1252–1255.
Bi, S., Ladenheim, E.E., Schwartz, G.J., Moran, T.H., 2001. A role forNPY overexpression in the dorsomedial hypothalamus in hyperphagiaand obesity of OLETF rats. Am. J. Physiol. Regul. Integr. Comp.Physiol. 281, R254–R260.
Boey, D., Lin, S., Karl, T., Baldock, P., Lee, N., Enriquez, R., Couzens,M., Slack, K., Dallmann, R., Sainsbury, A., Herzog, H., 2006. PeptideYY ablation in mice leads to the development of hyperinsulinaemiaand obesity. Diabetologia 49, 1360–1370.
Bresciani, E., Rapetti, D., Dona, F., Bulgarelli, I., Tamiazzo, L., Locatelli,V., Torsello, A., 2006. Obestatin inhibits feeding but does notmodulate GH and corticosterone secretion in the rat. J. Endocrinol.Invest. 29, RC16–RC18.
Broberger, C., Landry, M., Wong, H., Walsh, J.N., Hokfelt, T., 1997.Subtypes Y1 and Y2 of the neuropeptide Y receptor are respectivelyexpressed in pro-opiomelanocortin- and neuropeptide-Y-containingneurons of the rat hypothalamic arcuate nucleus. Neuroendocrinology66, 393–408.
Broglio, F., Benso, A., Gottero, C., Prodam, F., Gauna, C., Filtri, L.,Arvat, E., van der Lely, A.J., Deghenghi, R., Ghigo, E., 2003. Non-acylated ghrelin does not possess the pituitaric and pancreaticendocrine activity of acylated ghrelin in humans. J. Endocrinol.Invest. 26, 192–196.
Bruning, J.C., Gautam, D., Burks, D.J., Gillette, J., Schubert, M., Orban,P.C., Klein, R., Krone, W., Muller-Wieland, D., Kahn, C.R., 2000.Role of brain insulin receptor in control of body weight andreproduction. Science 289, 2122–2125.
Buchan, A.M., Polak, J.M., Capella, C., Solcia, E., Pearse, A.G., 1978.Electron immunocytochemical evidence for the K cell localization ofgastric inhibitory polypeptide (GIP) in man. Histochemistry 56, 37–44.
Bullock, B.P., Heller, R.S., Habener, J.F., 1996. Tissue distribution ofmessenger ribonucleic acid encoding the rat glucagon-like peptide-1receptor. Endocrinology 137, 2968–2978.
Burdyga, G., Lal, S., Spiller, D., Jiang, W., Thompson, D., Attwood, S.,Saeed, S., Grundy, D., Varro, A., Dimaline, R., Dockray, G.J., 2003.Localization of orexin-1 receptors to vagal afferent neurons in the ratand humans. Gastroenterology 124, 129–139.
Buse, J.B., Henry, R.R., Han, J., Kim, D.D., Fineman, M.S., Baron,A.D., 2004. Effects of exenatide (exendin-4) on glycemic control over30 weeks in sulfonylurea-treated patients with type 2 diabetes.Diabetes Care 27, 2628–2635.
Cassoni, P., Papotti, M., Ghe, C., Catapano, F., Sapino, A., Graziani, A.,Deghenghi, R., Reissmann, T., Ghigo, E., Muccioli, G., 2001.Identification, characterization, and biological activity of specificreceptors for natural (ghrelin) and synthetic growth hormone secret-agogues and analogs in human breast carcinomas and cell lines. J.Clin. Endocrinol. Metab. 86, 1738–1745.
Challis, B.G., Pinnock, S.B., Coll, A.P., Carter, R.N., Dickson, S.L.,O’Rahilly, S., 2003. Acute effects of PYY3-36 on food intake andhypothalamic neuropeptide expression in the mouse. Biochem. Bio-phys. Res. Commun. 311, 915–919.
Chartrel, N., Alvear-Perez, R., Leprince, J., Iturrioz, X., Reaux-LeGoazigo, A., Audinot, V., Chomarat, P., Coge, F., Nosjean, O.,Rodriguez, M., Galizzi, J.P., Boutin, J.A., Vaudry, H., Llorens-Cortes,C., 2007. Comment on ‘‘obestatin, a peptide encoded by the ghrelingene, opposes ghrelin’s effects on food intake’’. Science 315, 766.
Chatzaki, E., Crowe, P.D., Wang, L., Million, M., Tache, Y., Grigoriadis,D.E., 2004a. CRF receptor type 1 and 2 expression and anatomicaldistribution in the rat colon. J. Neurochem. 90, 309–316.
Chatzaki, E., Lambropoulou, M., Constantinidis, T.C., Papadopoulos,N., Tache, Y., Minopoulos, G., Grigoriadis, D.E., 2006. Corticotro-pin-releasing factor (CRF) receptor type 2 in the human stomach:protective biological role by inhibition of apoptosis. J. Cell. Physiol.209, 905–911.
Chatzaki, E., Murphy, B.J., Wang, L., Million, M., Ohning, G.V., Crowe,P.D., Petroski, R., Tache, Y., Grigoriadis, D.E., 2004b. Differential

H. Mendieta-Zeron et al. / General and Comparative Endocrinology 155 (2008) 481–495 491
profile of CRF receptor distribution in the rat stomach and duodenumassessed by newly developed CRF receptor antibodies. J. Neurochem.88, 1–11.
Clark, J.T., Kalra, P.S., Crowley, W.R., Kalra, S.P., 1984. NeuropeptideY and human pancreatic polypeptide stimulate feeding behavior inrats. Endocrinology 115, 427–429.
Clark, J.T., Sahu, A., Kalra, P.S., Balasubramaniam, A., Kalra, S.P.,1987. Neuropeptide Y (NPY)-induced feeding behavior in female rats:comparison with human NPY ([Met17]NPY), NPY analog ([nor-Leu4]NPY) and peptide YY. Regul. Peptides 17, 31–39.
Cohen, M.A., Ellis, S.M., Le Roux, C.W., Batterham, R.L., Park, A.,Patterson, M., Frost, G.S., Ghatei, M.A., Bloom, S.R., 2003.Oxyntomodulin suppresses appetite and reduces food intake inhumans. J Clin. Endocrinol. Metab. 88, 4696–4701.
Coll, A.P., Challis, B.G., O’Rahilly, S., 2004. Peptide YY3-36 and satiety:clarity or confusion? Endocrinology 145, 2582–2584.
Conlon, J.M., 2002. The origin and evolution of peptide YY (PYY) andpancreatic polypeptide (PP). Peptides 23, 269–278.
Corpa, E.S., McQuade, J., Krasnicki, S., Conze, D.B., 2001. Feeding afterfourth ventricular administration of neuropeptide Y receptor agonistsin rats. Peptides 22, 493–499.
Crowell, M.D., Decker, G.A., Levy, R., Jeffrey, R., Talley, N.J., 2006.Gut-brain neuropeptides in the regulation of ingestive behaviors andobesity. Am. J. Gastroenterol. 101, 2848–2856.
Cummings, D.E., Frayo, R.S., Marmonier, C., Aubert, R., Chapelot, D.,2004. Plasma ghrelin levels and hunger scores in humans initiatingmeals voluntarily without time- and food-related cues. Am. J. Physiol.Endocrinol. Metab. 287, E297–E304.
Cummings, D.E., Overduin, J., 2007. Gastrointestinal regulation of foodintake. J. Clin. Invest. 117, 13–23.
Cummings, D.E., Overduin, J., Shannon, M.H., Foster-Schubert, K.E.,2005. Hormonal mechanisms of weight loss and diabetes resolutionafter bariatric surgery. Surg. Obes. Relat. Dis. 1, 358–368.
Cummings, D.E., Purnell, J.Q., Frayo, R.S., Schmidova, K., Wisse, B.E.,Weigle, D.S., 2001. A preprandial rise in plasma ghrelin levels suggestsa role in meal initiation in humans. Diabetes 50, 1714–1719.
Cummings, D.E., Weigle, D.S., Frayo, R.S., Breen, P.A., Ma, M.K.,Dellinger, E.P., Purnell, J.Q., 2002. Plasma ghrelin levels after diet-induced weight loss or gastric bypass surgery. N. Engl. J. Med. 346,1623–1630.
Czimmer, J., Million, M., Tache, Y., 2006. Urocortin 2 acts centrally todelay gastric emptying through sympathetic pathways while CRF andurocortin 1 inhibitory actions are vagal dependent in rats. Am. J.Physiol. Gastrointest. Liver Physiol. 290, G511–G518.
D’Alessio, D.A., Kahn, S.E., Leusner, C.R., Ensinck, J.W., 1994.Glucagon-like peptide 1 enhances glucose tolerance both by stimula-tion of insulin release and by increasing insulin-independent glucosedisposal. J. Clin. Invest. 93, 2263–2266.
Dakin, C.L., Small, C.J., Batterham, R.L., Neary, N.M., Cohen, M.A.,Patterson, M., Ghatei, M.A., Bloom, S.R., 2004. Peripheral oxynto-modulin reduces food intake and body weight gain in rats. Endocri-nology 145, 2687–2695.
de Miguel, M.J., Burrell, M.A., 2002. Immunocytochemical detection oforexin A in endocrine cells of the developing mouse gut. J. Histochem.Cytochem. 50, 63–70.
Deacon, C.F., 2004. Therapeutic strategies based on glucagon-like peptide1. Diabetes 53, 2181–2189.
DeFronzo, R.A., Ratner, R.E., Han, J., Kim, D.D., Fineman, M.S.,Baron, A.D., 2005. Effects of exenatide (exendin-4) on glycemiccontrol and weight over 30 weeks in metformin-treated patients withtype 2 diabetes. Diabetes Care 28, 1092–1100.
Drazen, D.L., Vahl, T.P., D’Alessio, D.A., Seeley, R.J., Woods, S.C.,2006. Effects of a fixed meal pattern on ghrelin secretion: evidence for alearned response independent of nutrient status. Endocrinology 147,23–30.
Drucker, D.J., 2003. Enhancing incretin action for the treatment of type 2diabetes. Diabetes Care 26, 2929–2940.
Dyer, C.J., Touchette, K.J., Carroll, J.A., Allee, G.L., Matteri, R.L., 1999.Cloning of porcine prepro-orexin cDNA and effects of an intramus-cular injection of synthetic porcine orexin-B on feed intake in youngpigs. Domest. Anim. Endocrinol. 16, 145–148.
Ehrstrom, M., Gustafsson, T., Finn, A., Kirchgessner, A., Gryback, P.,Jacobsson, H., Hellstrom, P.M., Naslund, E., 2005. Inhibitory effect ofexogenous orexin a on gastric emptying, plasma leptin, and thedistribution of orexin and orexin receptors in the gut and pancreas inman. J. Clin. Endocrinol. Metab. 90, 2370–2377.
English, P.J., Ghatei, M.A., Malik, I.A., Bloom, S.R., Wilding, J.P., 2002.Food fails to suppress ghrelin levels in obese humans. J Clin.Endocrinol. Metab. 87, 2984.
Flatt, P.R., 2007. Effective surgical treatment of obesity may be mediatedby ablation of the lipogenic gut hormone gastric inhibitory polypeptide(GIP): evidence and clinical opportunity for development of newobesity-diabetes drugs? Diab. Vasc. Dis. Res. 4, 151–153.
Flier, J.S., 2004. Obesity wars: molecular progress confronts an expandingepidemic. Cell 116, 337–350.
Flint, A., Raben, A., Astrup, A., Holst, J.J., 1998. Glucagon-like peptide 1promotes satiety and suppresses energy intake in humans. J. Clin.Invest. 101, 515–520.
Flint, A., Raben, A., Ersboll, A.K., Holst, J.J., Astrup, A., 2001. Theeffect of physiological levels of glucagon-like peptide-1 on appetite,gastric emptying, energy and substrate metabolism in obesity. Int. J.Obes. Relat. Metab. Disord. 25, 781–792.
Foster-Schubert, K.E., Cummings, D.E., 2006. Emerging therapeuticstrategies for obesity. Endocr. Rev. 27, 779–793.
French, S.J., Murray, B., Rumsey, R.D., Sepple, C.P., Read, N.W., 1993.Is cholecystokinin a satiety hormone? Correlations of plasma chole-cystokinin with hunger, satiety and gastric emptying in normalvolunteers. Appetite 21, 95–104.
Fruhbeck, G., Gomez-Ambrosi, J., 2003. Control of body weight: aphysiologic and transgenic perspective. Diabetologia 46, 143–172.
Fujimoto, S., Inui, A., Kiyota, N., Seki, W., Koide, K., Takamiya, S.,Uemoto, M., Nakajima, Y., Baba, S., Kasuga, M., 1997. Increasedcholecystokinin and pancreatic polypeptide responses to a fat-richmeal in patients with restrictive but not bulimic anorexia nervosa. Biol.Psychiatry 41, 1068–1070.
Fushiki, T., Kojima, A., Imoto, T., Inoue, K., Sugimoto, E., 1992. Anextract of Gymnema sylvestre leaves and purified gymnemic acidinhibits glucose-stimulated gastric inhibitory peptide secretion in rats.J. Nutr. 122, 2367–2373.
Garcıa, M.C., Lopez, M., Alvarez, C.V., Casanueva, F., Tena-Sempere,M., Dieguez, C., 2007. Role of ghrelin in reproduction. Reproduction133, 531–540.
Garg, A., 2007. The ongoing saga of obestatin: is it a hormone? J. Clin.Endocrinol. Metab. 92, 3396–3398.
Gibbs, J., Fauser, D.J., Rowe, E.A., Rolls, B.J., Rolls, E.T., Maddison,S.P., 1979. Bombesin suppresses feeding in rats. Nature 282, 208–210.
Gibbs, J., Young, R.C., Smith, G.P., 1973a. Cholecystokinin decreasesfood intake in rats. J. Comp. Physiol. Psychol. 84, 488–495.
Gibbs, J., Young, R.C., Smith, G.P., 1973b. Cholecystokinin elicits satietyin rats with open gastric fistulas. Nature 245, 323–325.
Gourcerol, G., St Pierre, D.H., Tache, Y., 2007a. Lack of obestatin effectson food intake: should obestatin be renamed ghrelin-associatedpeptide (GAP)? Regul. Peptides 141, 1–7.
Gourcerol, G., Tache, Y., 2007b. Obestatin—a ghrelin-associated peptidethat does not hold its promise to suppress food intake and motility.Neurogastroenterol. Motil. 19, 161–165.
Grabauskas, G., Moises, H.C., 2003. Gastrointestinal-projecting neuronesin the dorsal motor nucleus of the vagus exhibit direct and viscero-topically organized sensitivity to orexin. J. Physiol. 549, 37–56.
Grandt, D., Schimiczek, M., Beglinger, C., Layer, P., Goebell, H.,Eysselein, V.E., Reeve Jr., J.R., 1994. Two molecular forms of peptideYY (PYY) are abundant in human blood: characterization of aradioimmunoassay recognizing PYY 1-36 and PYY 3-36. Regul.Peptides 51, 151–159.

492 H. Mendieta-Zeron et al. / General and Comparative Endocrinology 155 (2008) 481–495
Green, B.D., Irwin, N., Flatt, P.R., 2007. Direct and indirect effects ofobestatin peptides on food intake and the regulation of glucosehomeostasis and insulin secretion in mice. Peptides 28, 981–987.
Grove, K.L., Cowley, M.A., 2005. Is ghrelin a signal for the developmentof metabolic systems? J. Clin. Invest. 115, 3393–3397.
Gualillo, O., Lago, F., Gomez-Reino, J., Casanueva, F.F., Dieguez, C.,2003. Ghrelin, a widespread hormone: insights into molecular andcellular regulation of its expression and mechanism of action. FEBSLett. 552, 105–109.
Hagan, M.M., Castaneda, E., Sumaya, I.C., Fleming, S.M., Galloway, J.,Moss, D.E., 1998. The effect of hypothalamic peptide YY onhippocampal acetylcholine release in vivo: implications for limbicfunction in binge-eating behavior. Brain Res. 805, 20–28.
Halatchev, I.G., Cone, R.D., 2005. Peripheral administration of PYY3-36produces conditioned taste aversion in mice. Cell Metab. 1, 159–168.
Haynes, A.C., Jackson, B., Overend, P., Buckingham, R.E., Wilson, S.,Tadayyon, M., Arch, J.R., 1999. Effects of single and chronicintracerebroventricular administration of the orexins on feeding inthe rat. Peptides 20, 1099–1105.
Hecherington, A., Ranson, S., 1942. The spontaneous activity and foodintake of rats with hypothalamic lesions. Am. J. Physiol. 136, 609–617.
Hillhouse, E.W., Grammatopoulos, D.K., 2006. The molecular mecha-nisms underlying the regulation of the biological activity of cortico-tropin-releasing hormone receptors: implications for physiology andpathophysiology. Endocr. Rev. 27, 260–286.
Hirasawa, A., Tsumaya, K., Awaji, T., Katsuma, S., Adachi, T., Yamada,M., Sugimoto, Y., Miyazaki, S., Tsujimoto, G., 2005. Free fatty acidsregulate gut incretin glucagon-like peptide-1 secretion throughGPR120. Nat. Med. 11, 90–94.
Holst, B., Egerod, K.L., Schild, E., Vickers, S.P., Cheetham, S., Gerlach,L.O., Storjohann, L., Stidsen, C.E., Jones, R., Beck-Sickinger, A.G.,Schwartz, T.W., 2007. GPR39 signaling is stimulated by zinc ions butnot by obestatin. Endocrinology 148, 13–20.
Horlick, M.B., Rosenbaum, M., Nicolson, M., Levine, L.S., Fedun, B.,Wang, J., Pierson Jr., R.N., Leibel, R.L., 2000. Effect of puberty on therelationship between circulating leptin and body composition. J. Clin.Endocrinol. Metab. 85, 2509–2518.
Hosoda, H., Kojima, M., Matsuo, H., Kangawa, K., 2000a. Ghrelin anddes-acyl ghrelin: two major forms of rat ghrelin peptide in gastroin-testinal tissue. Biochem. Biophys. Res. Commun. 279, 909–913.
Hosoda, H., Kojima, M., Matsuo, H., Kangawa, K., 2000b. Purificationand characterization of rat des-Gln14-Ghrelin, a second endogenousligand for the growth hormone secretagogue receptor. J. Biol. Chem.275, 21995–22000.
Imeryuz, N., Yegen, B.C., Bozkurt, A., Coskun, T., Villanueva-Penacar-rillo, M.L., Ulusoy, N.B., 1997. Glucagon-like peptide-1 inhibitsgastric emptying via vagal afferent-mediated central mechanisms. Am.J. Physiol. 273, G920–G927.
Kalra, S.P., Dube, M.G., Pu, S., Xu, B., Horvath, T.L., Kalra, P.S., 1999.Interacting appetite-regulating pathways in the hypothalamic regula-tion of body weight. Endocr. Rev. 20, 68–100.
Kalra, S.P., Kalra, P.S., 2004. NPY and cohorts in regulating appetite,obesity and metabolic syndrome: beneficial effects of gene therapy.Neuropeptides 38, 201–211.
Katsuura, G., Asakawa, A., Inui, A., 2002. Roles of pancreatic polypep-tide in regulation of food intake. Peptides 23, 323–329.
Kendall, D.M., Riddle, M.C., Rosenstock, J., Zhuang, D., Kim, D.D.,Fineman, M.S., Baron, A.D., 2005. Effects of exenatide (exendin-4) onglycemic control over 30 weeks in patients with type 2 diabetes treatedwith metformin and a sulfonylurea. Diabetes Care 28, 1083–1091.
Kirchgessner, A.L., 2002. Orexins in the brain–gut axis. Endocr. Rev. 23,1–15.
Kirchgessner, A.L., Liu, M., 1999. Orexin synthesis and response in thegut. Neuron 24, 941–951.
Koda, S., Date, Y., Murakami, N., Shimbara, T., Hanada, T., Toshinai,K., Niiji, A., Furuya, M., Inomata, N., Osuye, K., Nakazato, M.,2005. The Role of the vagal nerve in peripheral PYY3-36-inducedfeeding reduction in rats. Endocrinology 146, 2369–2375.
Kojima, M., Hosoda, H., Date, Y., Nakazato, M., Matsuo, H., Kangawa,K., 1999. Ghrelin is a growth-hormone-releasing acylated peptide fromstomach. Nature 402, 656–660.
Kola, B., Hubina, E., Tucci, S.A., Kirkham, T.C., Garcia, E.A.,Mitchell, S.E., Williams, L.M., Hawley, S.A., Hardie, D.G.,Grossman, A.B., Korbonits, M., 2005. Cannabinoids andghrelin have both central and peripheral metabolic and cardiaceffects via AMP-activated protein kinase. J. Biol. Chem. 280,25196–25201.
Komaki, G., Matsumoto, Y., Nishikata, H., Kawai, K., Nozaki, T.,Takii, M., Sogawa, H., Kubo, C., 2001. Orexin-A and leptin changeinversely in fasting non-obese subjects. Eur. J. Endocrinol. 144, 645–651.
Konturek, S.J., Konturek, J.W., Pawlik, T., Brzozowski, T., 2004. Brain-gut axis and its role in the control of food intake. J. Physiol.Pharmacol. 55, 137–154.
Korczynski, W., Ceregrzyn, M., Kato, I., Wolinski, J., Zabielski, R.,2006a. The effect of orexins on intestinal motility in vitro in fed andfasted rats. J. Physiol. Pharmacol. 57 (Suppl 6), 43–54.
Korczynski, W., Ceregrzyn, M., Matyjek, R., Kato, I., Kuwahara, A.,Wolinski, J., Zabielski, R., 2006b. Central and local (enteric) action oforexins. J. Physiol. Pharmacol. 57 (Suppl 6), 17–42.
Korner, J., Aronne, L.J., 2004. Pharmacological approaches to weightreduction: therapeutic targets. J. Clin. Endocrinol. Metab. 89, 2616–2621.
Lachey, J.L., D’Alessio, D.A., Rinaman, L., Elmquist, J.K., Drucker,D.J., Seeley, R.J., 2005. The role of central glucagon-like peptide-1 inmediating the effects of visceral illness: differential effects in rats andmice. Endocrinology 146, 458–462.
Ladenheim, E.E., Moore, K.A., Salorio, C.F., Mantey, S.A., Taylor, J.E.,Coy, D.H., Jensen, R.T., Moran, T.H., 1997. Characterization ofbombesin binding sites in the rat stomach. Eur. J. Pharmacol. 319,245–251.
Ladenheim, E.E., Taylor, J.E., Coy, D.H., Moran, T.H., 1994. Blockadeof feeding inhibition by neuromedin B using a selective receptorantagonist. Eur. J. Pharmacol. 271, R7–R9.
Lagaud, G.J., Young, A., Acena, A., Morton, M.F., Barrett, T.D.,Shankley, N.P., 2007. Obestatin reduces food intake and suppressesbody weight gain in rodents. Biochem. Biophys. Res. Commun. 357,264–269.
Larsson, L.I., Rehfeld, J.F., 1978. Distribution of gastrin and CCK cells inthe rat gastrointestinal tract. Evidence for the occurrence of threedistinct cell types storing COOH-terminal gastrin immunoreactivity.Histochemistry 58, 23–31.
Le Roux, C.W., Aylwin, S.J., Batterham, R.L., Borg, C.M., Coyle, F.,Prasad, V., Shurey, S., Ghatei, M.A., Patel, A.G., Bloom, S.R., 2006.Gut hormone profiles following bariatric surgery favor an anorecticstate, facilitate weight loss, and improve metabolic parameters. Ann.Surg. 243, 108–114.
Lewis, K., Li, C., Perrin, M.H., Blount, A., Kunitake, K., Donaldson, C.,Vaughan, J., Reyes, T.M., Gulyas, J., Fischer, W., Bilezikjian, L.,Rivier, J., Sawchenko, P.E., Vale, W.W., 2001. Identification ofurocortin III, an additional member of the corticotropin-releasingfactor (CRF) family with high affinity for the CRF2 receptor. Proc.Natl. Acad. Sci. USA 98, 7570–7575.
Lopez, M., Lelliott, C.J., Vidal-Puig, A., 2007a. Hypothalamic fatty acidmetabolism: a housekeeping pathway that regulates food intake.Bioessays 29, 248–261.
Lopez, M., Tena-Sempere, M., Garcıa-Caballero, T., Senarıs, R., Dieguez,C., 2005. Hypocretins in endocrine regulation. In: de Lecea, L.,Sutcliffe, J.G. (Eds.), Hypocretins. Integrators of Physiological Func-tions Springer Science, NY(USA), pp. 393–421.
Lopez, M., Tovar, S., Vazquez, M.J., Williams, L.M., Dieguez, C., 2007b.Peripheral tissue–brain interactions in the regulation of food intake.Proc. Nutr. Soc. 66, 131–155.
Malaisse-Lagae, F., Carpentier, J.L., Patel, Y.C., Malaisse, W.J., Orci, L.,1977. Pancreatic polypeptide: a possible role in the regulation of foodintake in the mouse. Hypothesis. Exp. 33, 915–917.

H. Mendieta-Zeron et al. / General and Comparative Endocrinology 155 (2008) 481–495 493
Marks, J.L., Porte Jr., D., Stahl, W.L., Baskin, D.G., 1990. Localizationof insulin receptor mRNA in rat brain by in situ hybridization.Endocrinology 127, 3234–3236.
Martinez, V., Rivier, J., Tache, Y., 1999. Peripheral injection of a newcorticotropin-releasing factor (CRF) antagonist, astressin, blocksperipheral CRF- and abdominal surgery-induced delayed gastricemptying in rats. J. Pharmacol. Exp. Ther. 290, 629–634.
Martini, A.C., Fernandez-Fernandez, R., Tovar, S., Navarro, V.M.,Vigo, E., Vazquez, M.J., Davies, J.S., Thompson, N.M., Aguilar, E.,Pinilla, L., Wells, T., Dieguez, C., Tena-Sempere, M., 2006. Com-parative analysis of the effects of ghrelin and unacylated ghrelin onluteinizing hormone secretion in male rats. Endocrinology 147, 2374–2382.
Masuda, Y., Tanaka, T., Inomata, N., Ohnuma, N., Tanaka, S., Itoh, Z.,Hosoda, H., Kojima, M., Kangawa, K., 2000. Ghrelin stimulatesgastric acid secretion and motility in rats. Biochem. Biophys. Res.Commun. 276, 905–908.
Medina-Gomez, G., Vidal-Puig, A., 2005. Gateway to the metabolicsyndrome. Nat. Med. 11, 602–603.
Meeran, K., O’Shea, D., Edwards, C.M., Turton, M.D., Heath, M.M.,Gunn, I., Abusnana, S., Rossi, M., Small, C.J., Goldstone, A.P.,Taylor, G.M., Sunter, D., Steere, J., Choi, S.J., Ghatei, M.A., Bloom,S.R., 1999. Repeated intracerebroventricular administration of gluca-gon-like peptide-1-(7-36) amide or exendin-(9-39) alters body weight inthe rat. Endocrinology 140, 244–250.
Mercer, J.G., Lawrence, C.B., 1992. Selectivity of cholecystokinin (CCK)receptor antagonists, MK-329 and L-365,260, for axonally-transportedCCK binding sites on the rat vagus nerve. Neurosci. Lett. 137, 229–231.
Million, M., Maillot, C., Saunders, P., Rivier, J., Vale, W., Tache, Y.,2002. Human urocortin II, a new CRF-related peptide, displaysselective CRF(2)-mediated action on gastric transit in rats. Am. J.Physiol. Gastrointest. Liver Physiol. 282, G34–G40.
Miyawaki, K., Yamada, Y., Ban, N., Ihara, Y., Tsukiyama, K., Zhou, H.,Fujimoto, S., Oku, A., Tsuda, K., Toyokuni, S., Hiai, H., Mizunoya,W., Fushiki, T., Holst, J.J., Makino, M., Tashita, A., Kobara, Y.,Tsubamoto, Y., Jinnouchi, T., Jomori, T., Seino, Y., 2002. Inhibitionof gastric inhibitory polypeptide signaling prevents obesity. Nat. Med.8, 738–742.
Moran, T.H., Baldessarini, A.R., Salorio, C.F., Lowery, T., Schwartz,G.J., 1997. Vagal afferent and efferent contributions to the inhibitionof food intake by cholecystokinin. Am. J. Physiol. 272, R1245–R1251.
Moran, T.H., Robinson, P.H., Goldrich, M.S., McHugh, P.R., 1986. Twobrain cholecystokinin receptors: implications for behavioral actions.Brain Res. 362, 175–179.
Moran, T.H., Smedh, U., Kinzig, K.P., Scott, K.A., Knipp, S., Laden-heim, E.E., 2005. Peptide YY(3-36) inhibits gastric emptying andproduces acute reductions in food intake in rhesus monkeys. Am. J.Physiol. Regul. Integr. Comp. Physiol. 288, R384–R388.
Morton, G.J., Cummings, D.E., Baskin, D.G., Barsh, G.S., Schwartz,M.W., 2006. Central nervous system control of food intake and bodyweight. Nature 443, 289–295.
Mullins, D.E., Zhang, X., Hawes, B.E., 2002. Activation of extracellularsignal regulated protein kinase by neuropeptide Y and pancreaticpolypeptide in CHO cells expressing the NPY Y(1), Y(2), Y(4) andY(5) receptor subtypes. Regul. Peptides 105, 65–73.
Muramatsu, Y., Fukushima, K., Iino, K., Totsune, K., Takahashi, K.,Suzuki, T., Hirasawa, G., Takeyama, J., Ito, M., Nose, M., Tashiro,A., Hongo, M., Oki, Y., Nagura, H., Sasano, H., 2000. Urocortin andcorticotropin-releasing factor receptor expression in the human colonicmucosa. Peptides 21, 1799–1809.
Murphy, K.G., Bloom, S.R., 2004. Gut hormones in the control ofappetite. Exp. Physiol. 89, 507–516.
Murphy, K.G., Bloom, S.R., 2006. Gut hormones and the regulation ofenergy homeostasis. Nature 444, 854–859.
Muurahainen, N., Kissileff, H.R., DeRogatis, A.J., Pi-Sunyer, F.X., 1988.Effects of cholecystokinin-octapeptide (CCK-8) on food intake andgastric emptying in man. Physiol. Behav. 44, 645–649.
Nagata, T., Uemoto, M., Yuzuriha, H., Asakawa, A., Inui, A., Fujimiya,M., Sakamaki, R., Kasuga, M., Shinfuku, N., 2005. Intracerebroven-tricularly administered urocortin inhibits gastric emptying in mice. Int.J. Mol. Med. 15, 1041–1043.
Nakabayashi, M., Suzuki, T., Takahashi, K., Totsune, K., Muramatsu,Y., Kaneko, C., Date, F., Takeyama, J., Darnel, A.D., Moriya, T.,Sasano, H., 2003. Orexin-A expression in human peripheral tissues.Mol. Cell. Endocrinol. 205, 43–50.
Nakazato, M., Murakami, N., Date, Y., Kojima, M., Matsuo, H.,Kangawa, K., Matsukura, S., 2001. A role for ghrelin in the centralregulation of feeding. Nature 409, 194–198.
Nanzer, A.M., Khalaf, S., Mozid, A.M., Fowkes, R.C., Patel, M.V.,Burrin, J.M., Grossman, A.B., Korbonits, M., 2004. Ghrelin exerts aproliferative effect on a rat pituitary somatotroph cell line via themitogen-activated protein kinase pathway. Eur. J. Endocrinol. 151,233–240.
Naslund, E., Ehrstrom, M., Ma, J., Hellstrom, P.M., Kirchgessner, A.L.,2002. Localization and effects of orexin on fasting motility in the ratduodenum. Am. J. Physiol. Gastrointest. Liver Physiol. 282, G470–G479.
Naslund, E., Hellstrom, P.M., 2007. Appetite signaling: from gut peptidesand enteric nerves to brain. Physiol. Behav. 92, 256–262.
Nauck, M.A., Niedereichholz, U., Ettler, R., Holst, J.J., Orskov, C.,Ritzel, R., Schmiegel, W.H., 1997. Glucagon-like peptide 1 inhibitionof gastric emptying outweighs its insulinotropic effects in healthyhumans. Am. J. Physiol. 273, E981–E988.
Nogueiras, R., Pfluger, P., Tovar, S., Myrtha, A., Mitchell, S., Morris, A.,Perez-Tilve, D., Vazquez, M.J., Wiedmer, P., Castaneda, T.R.,Dimarchi, R., Tschop, M., Schurmann, A., Joost, H.G., Williams,L.M., Langhans, W., Dieguez, C., 2007. Effects of obestatin on energybalance and growth hormone secretion in rodents. Endocrinology 148,21–26.
Nogueiras, R., Tovar, S., Mitchell, S.E., Rayner, D.V., Archer, Z.A.,Dieguez, C., Williams, L.M., 2004. Regulation of growth hormonesecretagogue receptor gene expression in the arcuate nuclei of the ratby leptin and ghrelin. Diabetes 53, 2552–2558.
Obici, S., Feng, Z., Karkanias, G., Baskin, D.G., Rossetti, L., 2002.Decreasing hypothalamic insulin receptors causes hyperphagia andinsulin resistance in rats. Nat. Neurosci. 5, 566–572.
Ohki-Hamazaki, H., Watase, K., Yamamoto, K., Ogura, H., Yamano,M., Yamada, K., Maeno, H., Imaki, J., Kikuyama, S., Wada, E.,Wada, K., 1997. Mice lacking bombesin receptor subtype-3 developmetabolic defects and obesity. Nature 390, 165–169.
Okada, S., York, D.A., Bray, G.A., Mei, J., Erlanson-Albertsson, C.,1992. Differential inhibition of fat intake in two strains of rat by thepeptide enterostatin. Am. J. Physiol. 262, R1111–R1116.
Ouedraogo, R., Naslund, E., Kirchgessner, A.L., 2003. Glucose regulatesthe release of orexin-A from the endocrine pancreas. Diabetes 52, 111–117.
Peino, R., Baldelli, R., Rodriguez-Garcia, J., Rodriguez-Segade, S.,Kojima, M., Kangawa, K., Arvat, E., Ghigo, E., Dieguez, C.,Casanueva, F.F., 2000. Ghrelin-induced growth hormone secretionin humans. Eur. J. Endocrinol. 143, R11–R14.
Perez-Tilve, D., Gonzalez-Matias, L., Alvarez-Crespo, M., Leiras, R.,Tovar, S., Dieguez, C., Mallo, F., 2007. Exendin-4 potently decreasesghrelin levels in fasting rats. Diabetes 56, 143–151.
Perez-Tilve, D., Nogueiras, R., Mallo, F., Benoit, S.C., Tschoep, M.,2006. Gut hormones ghrelin, PYY, and GLP-1 in the regulation ofenergy, balance, and metabolism. Endocrine 29, 61–72.
Reeve Jr., J.R., Eysselein, V.E., Ho, F.J., Chew, P., Vigna, S.R., Liddle,R.A., Evans, C., 1994. Natural and synthetic CCK-58. Novel reagentsfor studying cholecystokinin physiology. Ann. N. Y. Acad. Sci. 713,11–21.
Reeve Jr., J.R., Liddle, R.A., McVey, D.C., Vigna, S.R., Solomon, T.E.,Keire, D.A., Rosenquist, G., Shively, J.E., Lee, T.D., Chew, P., Green,G.M., Coskun, T., 2004. Identification of nonsulfated cholecystokinin-58 in canine intestinal extracts and its biological properties. Am. J.Physiol. Gastrointest. Liver Physiol. 287, G326–G333.

494 H. Mendieta-Zeron et al. / General and Comparative Endocrinology 155 (2008) 481–495
Reidelberger, R.D., Hernandez, J., Fritzsch, B., Hulce, M., 2004.Abdominal vagal mediation of the satiety effects of CCK in rats.Am. J. Physiol. Regul. Integr. Comp. Physiol. 286, R1005–R1012.
Roberge, J.N., Brubaker, P.L., 1991. Secretion of proglucagon-derivedpeptides in response to intestinal luminal nutrients. Endocrinology128, 3169–3174.
Rocca, A.S., Brubaker, P.L., 1999. Role of the vagus nerve in mediatingproximal nutrient-induced glucagon-like peptide-1 secretion. Endocri-nology 140, 1687–1694.
Sagami, Y., Shimada, Y., Tayama, J., Nomura, T., Satake, M., Endo, Y.,Shoji, T., Karahashi, K., Hongo, M., Fukudo, S., 2004. Effect of acorticotropin releasing hormone receptor antagonist on colonicsensory and motor function in patients with irritable bowel syndrome.Gut 53, 958–964.
Sakurai, T., Amemiya, A., Ishii, M., Matsuzaki, I., Chemelli, R.M.,Tanaka, H., Williams, S.C., Richardson, J.A., Kozlowski, G.P.,Wilson, S., Arch, J.R., Buckingham, R.E., Haynes, A.C., Carr, S.A.,Annan, R.S., McNulty, D.E., Liu, W.S., Terrett, J.A., Elshourbagy,N.A., Bergsma, D.J., Yanagisawa, M., 1998. Orexins ans orexinreceptors: a family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior. Cell 92, 573–585.
Savage, A.P., Adrian, T.E., Carolan, G., Chatterjee, V.K., Bloom, S.R.,1987. Effects of peptide YY (PYY) on mouth to caecum intestinaltransit time and on the rate of gastric emptying in healthy volunteers.Gut 28, 166–170.
Schmidt, P.T., Naslund, E., Gryback, P., Jacobsson, H., Hartmann, B.,Holst, J.J., Hellstrom, P.M., 2003. Peripheral administration of GLP-2to humans has no effect on gastric emptying or satiety. Regul. Peptides116, 21–25.
Schwartz, M.W., Figlewicz, D.P., Baskin, D.G., Woods, S.C., Porte Jr.,D., 1992. Insulin in the brain: a hormonal regulator of energy balance.Endocr. Rev. 13, 387–414.
Schwartz, M.W., Woods, S.C., Porte Jr., D., Seeley, R.J., Baskin, D.G.,2000. Central nervous system control of food intake. Nature 404, 661–671.
Seoane, L.M., Lopez, M., Tovar, S., Casanueva, F., Senarıs, R., Dieguez,C., 2003. Agouti-related peptide, neuropeptide Y, and somatostatin-producing neurons are targets for ghrelin actions in the rat hypothal-amus. Endocrinology 144, 544–551.
Seoane, L.M., Al Massadi, O., Pazos, Y., Pagotto, U., Casanueva, F.F.,2006. Central obestatin administration does not modify either spon-taneous or ghrelin-induced food intake in rats. J. Endocrinol. Invest.29, RC13–RC15.
Seoane, L.M., Tovar, S., Baldelli, R., Arvat, E., Ghigo, E., Casanueva,F.F., Dieguez, C., 2000. Ghrelin elicits a marked stimulatory effect onGH secretion in freely-moving rats. Eur. J. Endocrinol. 143, R7–R9.
Shintani, M., Ogawa, Y., Ebihara, K., Aizawa-Abe, M., Miyanaga, F.,Takaya, K., Hayashi, T., Inoue, G., Hosoda, K., Kojima, M.,Kangawa, K., Nakao, K., 2001. Ghrelin, an endogenous growthhormone secretagogue, is a novel orexigenic peptide that antagonizesleptin action through the activation of hypothalamic neuropeptide Y/Y1 receptor pathway. Diabetes 50, 227–232.
Sibilia, V., Bresciani, E., Lattuada, N., Rapetti, D., Locatelli, V., De, L.V.,Dona, F., Netti, C., Torsello, A., Guidobono, F., 2006. Intracerebro-ventricular acute and chronic administration of obestatin minimallyaffect food intake but not weight gain in the rat. J. Endocrinol. Invest.29, RC31–RC34.
Small, C.J., Bloom, S.R., 2004. Gut hormones and the control of appetite.Trends Endocrinol. Metab. 15, 259–263.
Smith, G.P., Jerome, C., Gibbs, J., 1981. Abdominal vagotomy does notblock the satiety effect of bombesin in the rat. Peptides 2, 409–411.
Stanley, B.G., Daniel, D.R., Chin, A.S., Leibowitz, S.F., 1985. Paraven-tricular nucleus injections of peptide YY and neuropeptide Ypreferentially enhance carbohydrate ingestion. Peptides 6, 1205–1211.
Stanley, S., Wynne, K., McGowan, B., Bloom, S., 2005. Hormonalregulation of food intake. Physiol. Rev. 85, 1131–1158.
Stein, L.J., Woods, S.C., 1982. Gastrin releasing peptide reduces meal sizein rats. Peptides 3, 833–835.
Sun, Y., Ahmed, S., Smith, R.G., 2003. Deletion of ghrelin impairs neithergrowth nor appetite. Mol. Cell. Biol. 23, 7973–7981.
Sun, Y., Asnicar, M., Saha, P.K., Chan, L., Smith, R.G., 2006. Ablationof ghrelin improves the diabetic but not obese phenotype of ob/obmice. Cell Metab. 3, 379–386.
Tache, Y., Bonaz, B., 2007. Corticotropin-releasing factor receptors andstress-related alterations of gut motor function. J. Clin. Invest. 117,33–40.
Tache, Y., Martinez, V., Wang, L., Million, M., 2004a. CRF1 receptorsignaling pathways are involved in stress-related alterations of colonicfunction and viscerosensitivity: implications for irritable bowel syn-drome. Br. J. Pharmacol. 141, 1321–1330.
Tache, Y., Perdue, M.H., 2004b. Role of peripheral CRF signallingpathways in stress-related alterations of gut motility and mucosalfunction. Neurogastroenterol. Motil. 16 (Suppl 1), 137–142.
Talsania, T., Anini, Y., Siu, S., Drucker, D.J., Brubaker, P.L., 2005.Peripheral exendin-4 and peptide yy3-36 synergistically reduce foodintake through different mechanisms in mice. Endocrinology 146,3748–3756.
Tang-Christensen, M., Vrang, N., Larsen, P.J., 2001. Glucagon-likepeptide containing pathways in the regulation of feeding behaviour.Int. J. Obes. Relat. Metab. Disord. 25 (Suppl 5), S42–S47.
Theander-Carrillo, C., Wiedmer, P., Cettour-Rose, P., Nogueiras, R.,Perez-Tilve, D., Pfluger, P., Castaneda, T.R., Muzzin, P., Schurmann,A., Szanto, I., Tschop, M.H., Rohner-Jeanrenaud, F., 2006. Ghrelinaction in the brain controls adipocyte metabolism. J. Clin. Invest. 116,1983–1993.
Thiele, T.E., van Dijk, G., Campfield, L.A., Smith, F.J., Burn, P., Woods,S.C., Bernstein, I.L., Seeley, R.J., 1997. Central infusion of GLP-1, butnot leptin, produces conditioned taste aversions in rats. Am. J. Physiol.272, R726–R730.
Thompson, D.G., 2006. Neurogastroenterology: imaging of the sensoryand motor control of the GI tract. J. Psychosom. Res. 61, 301–304.
Thompson, N.M., Gill, D.A., Davies, R., Loveridge, N., Houston, P.A.,Robinson, I.C., Wells, T., 2004. Ghrelin and des-octanoyl ghrelinpromote adipogenesis directly in vivo by a mechanism independent ofthe type 1a growth hormone secretagogue receptor. Endocrinology145, 234–242.
Toshinai, K., Date, Y., Murakami, N., Shimada, M., Mondal, M.S.,Shimbara, T., Guan, J.L., Wang, Q.P., Funahashi, H., Sakurai, T.,Shioda, S., Matsukura, S., Kangawa, K., Nakazato, M., 2003.Ghrelin-induced food intake is mediated via the orexin pathway.Endocrinology 144, 1506–1512.
Tremblay, F., Perreault, M., Klaman, L.D., Tobin, J.F., Smith, E.,Gimeno, R.E., 2007. Normal food intake and body weight in micelacking the G protein-coupled receptor GPR39. Endocrinology 148,501–506.
Tschop, M., Castaneda, T.R., Joost, H.G., Thone-Reineke, C., Ortmann,S., Klaus, S., Hagan, M.M., Chandler, P.C., Oswald, K.D., Benoit,S.C., Seeley, R.J., Kinzig, K.P., Moran, T.H., Beck-Sickinger, A.G.,Koglin, N., Rodgers, R.J., Blundell, J.E., Ishii, Y., Beattie, A.H.,Holch, P., Allison, D.B., Raun, K., Madsen, K., Wulff, B.S., Stidsen,C.E., Birringer, M., Kreuzer, O.J., Schindler, M., Arndt, K., Rudolf,K., Mark, M., Deng, X.Y., Whitcomb, D.C., Halem, H., Taylor, J.,Dong, J., Datta, R., Culler, M., Craney, S., Flora, D., Smiley, D.,Heiman, M.L., 2004. Physiology: does gut hormone PYY3-36 decreasefood intake in rodents? Nature 430, 1–2.
Tschop, M., Smiley, D.L., Heiman, M.L., 2000. Ghrelin induces adiposityin rodents. Nature 407, 908–913.
Tschop, M., Weyer, C., Tataranni, P.A., Devanarayan, V., Ravussin, E.,Heiman, M.L., 2001. Circulating ghrelin levels are decreased in humanobesity. Diabetes 50, 707–709.
Turton, M.D., O’Shea, D., Gunn, I., Beak, S.A., Edwards, C.M., Meeran,K., Choi, S.J., Taylor, G.M., Heath, M.M., Lambert, P.D., Wilding,J.P., Smith, D.M., Ghatei, M.A., Herbert, J., Bloom, S.R., 1996. Arole for glucagon-like peptide-1 in the central regulation of feeding.Nature 379, 69–72.

H. Mendieta-Zeron et al. / General and Comparative Endocrinology 155 (2008) 481–495 495
Vale, W., Spiess, J., Rivier, C., Rivier, J., 1981. Characterization of a 41-residue ovine hypothalamic peptide that stimulates secretion ofcorticotropin and beta-endorphin. Science 213, 1394–1397.
Wang, L., Martinez, V., Rivier, J.E., Tache, Y., 2001. Peripheral urocortininhibits gastric emptying and food intake in mice: differential role ofCRF receptor 2. Am. J. Physiol. Regul. Integr. Comp. Physiol. 281,R1401–R1410.
Wank, S.A., Harkins, R., Jensen, R.T., Shapira, H., de Weerth, A.,Slattery, T., 1992a. Purification, molecular cloning, and functionalexpression of the cholecystokinin receptor from rat pancreas. Proc.Natl. Acad. Sci. USA 89, 3125–3129.
Wank, S.A., Pisegna, J.R., de Weerth, A., 1992b. Brain and gastrointes-tinal cholecystokinin receptor family: structure and functional expres-sion. Proc. Natl. Acad. Sci. USA 89, 8691–8695.
West, D.B., Fey, D., Woods, S.C., 1984. Cholecystokinin persistentlysuppresses meal size but not food intake in free-feeding rats. Am. J.Physiol. 246, R776–R787.
West, D.B., Williams, R.H., Braget, D.J., Woods, S.C., 1982. Bombesinreduces food intake of normal and hypothalamically obese rats andlowers body weight when given chronically. Peptides 3, 61–67.
Wettergren, A., Wojdemann, M., Holst, J.J., 1998. Glucagon-like peptide-1 inhibits gastropancreatic function by inhibiting central parasympa-thetic outflow. Am. J. Physiol. 275, G984–G992.
Wettergren, A., Wojdemann, M., Meisner, S., Stadil, F., Holst, J.J., 1997.The inhibitory effect of glucagon-like peptide-1 (GLP-1) 7-36 amide ongastric acid secretion in humans depends on an intact vagal innerva-tion. Gut 40, 597–601.
Whitcomb, D.C., Taylor, I.L., Vigna, S.R., 1990. Characterization ofsaturable binding sites for circulating pancreatic polypeptide in ratbrain. Am. J. Physiol. 259, G687–G691.
Williams, C.L., Peterson, J.M., Villar, R.G., Burks, T.F., 1987. Cortico-tropin-releasing factor directly mediates colonic responses to stress.Am. J. Physiol. 253, G582–G586.
Willie, J.T., Chemelli, R.M., Sinton, C.M., Yanagisawa, M., 2001. To eator to sleep? Orexin in the regulation of feeding and wakefulness. Annu.Rev. Neurosci. 24, 429–458.
Woods, S.C., Seeley, R.J., Baskin, D.G., Schwartz, M.W., 2003. Insulinand the blood–brain–barrier. Curr. Pharm. Des. 9, 795–800.
Wortley, K.E., Del Rincon, J.P., Murray, J.D., Garcia, K., Iida,K., Thorner, M.O., Sleeman, M.W., 2005. Absence of ghrelin
protects against early-onset obesity. J. Clin. Invest. 115, 3573–3578.
Wren, A.M., Seal, L.J., Cohen, M.A., Brynes, A.E., Frost, G.S., Murphy,K.G., Dhillo, W.S., Ghatei, M.A., Bloom, S.R., 2001. Ghrelinenhances appetite and increases food intake in humans. J. Clin.Endocrinol. Metab. 86, 5992.
Wynne, K., Park, A.J., Small, C.J., Patterson, M., Ellis, S.M., Murphy,K.G., Wren, A.M., Frost, G.S., Meeran, K., Ghatei, M.A., Bloom,S.R., 2005. Subcutaneous oxyntomodulin reduces body weight inoverweight and obese subjects: a double-blind, randomized, controlledtrial. Diabetes 54, 2390–2395.
Wynne, K., Stanley, S., Bloom, S., 2004. The gut and regulation of bodyweight. J. Clin. Endocrinol. Metab. 89, 2576–2582.
Yamamoto, H., Lee, C.E., Marcus, J.N., Williams, T.D., Overton, J.M.,Lopez, M.E., Hollenberg, A.N., Baggio, L., Saper, C.B., Drucker,D.J., Elmquist, J.K., 2002. Glucagon-like peptide-1 receptor stimula-tion increases blood pressure and heart rate and activates autonomicregulatory neurons. J. Clin. Invest. 110, 43–52.
Zhang, J.V., Ren, P.G., Avsian-Kretchmer, O., Luo, C.W., Rauch, R.,Klein, C., Hsueh, A.J., 2005. Obestatin, a peptide encoded by theghrelin gene, opposes ghrelin’s effects on food intake. Science 310, 996–999.
Zhang, J.V., Ren, P.G., Avsian-Kretchmer, O., Luo, C.W., Rauch, R.,Klein, C., Hsueh, A.J., 2007. Response to comments on: ‘‘obestatin, apeptide encoded by the ghrelin gene, opposes ghrelin’s effects on foodintake’’. Science 315, 766.
Zhang, Y., Proenca, R., Maffei, M., Barone, M., Leopold, L., Friedman,J.M., 1994. Positional cloning of the mouse obese gene and its humanhomologue. Nature 372, 425–432.
Zigman, J.M., Jones, J.E., Lee, C.E., Saper, C.B., Elmquist, J.K., 2006.Expression of ghrelin receptor mRNA in the rat and the mouse brain.J. Comp. Neurol. 494, 528–548.
Zigman, J.M., Nakano, Y., Coppari, R., Balthasar, N., Marcus, J.N., Lee,C.E., Jones, J.E., Deysher, A.E., Waxman, A.R., White, R.D.,Williams, T.D., Lachey, J.L., Seeley, R.J., Lowell, B.B., Elmquist,J.K., 2005. Mice lacking ghrelin receptors resist the development ofdiet-induced obesity. J. Clin. Invest. 115, 3564–3572.
Zizzari, P., Longchamps, R., Epelbaum, J., Bluet-Pajot, M.T., 2007.Obestatin partially affects ghrelin stimulation of food intake andgrowth hormone secretion in rodents. Endocrinology 148, 1648–1653.