
Platinum(II) metal complexes as potential anti-Trypanosoma cruzi agents
Upload
independentCategory
view
2download
0
International Journal for Parasitology 39 (2009) 963–973
Contents lists available at ScienceDirect
International Journal for Parasitology
journal homepage: www.elsevier .com/locate / i jpara
Trypanosoma cruzi maxicircle heterogeneity in Chagas disease patients from Brazil
Julio César Carranza a,1, Helder M.S. Valadares b, Daniella A. D’Ávila c, Rodrigo P. Baptista b,Margoth Moreno a, Lúcia M.C. Galvão c,d, Egler Chiari c, Nancy R. Sturm e, Eliane D. Gontijo f,Andrea M. Macedo b, Bianca Zingales a,*
a Departamento de Bioquímica, Instituto de Química, Universidade de São Paulo, Avenida Professor Lineu Prestes, 748, CEP 05508-000 São Paulo, SP, Brazilb Departamento de Bioquímica e Imunologia, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Brazilc Departamento de Parasitologia, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Brazild Programas de Pós-Graduação em Ciências da Saúde e Ciências Farmacêuticas, CCS, Universidade Federal do Rio Grande do Norte, Brazile Department of Microbiology, Immunology and Molecular Genetics, University of California at Los Angeles, Los Angeles, CA, USAf Departamento de Medicina Preventiva e Social, Faculdade de Medicina, Universidade Federal de Minas Gerais, Brazil
a r t i c l e i n f o
Article history:Received 18 November 2008Received in revised form 15 January 2009Accepted 15 January 2009
Keywords:Chagas diseaseClinical presentationsNADH dehydrogenase subunit 7Mitochondrial complex IMicrosatellites
0020-7519/$36.00 � 2009 Australian Society for Paradoi:10.1016/j.ijpara.2009.01.009
* Corresponding author. Tel.: +55 11 30913810x21E-mail address: [email protected] (B. Zingales).
1 Present address: Laboratorio de Investigaciones entad de Ciencias, Universidad del Tolima, A.A. No. 546, I
a b s t r a c t
The majority of individuals in the chronic phase of Chagas disease are asymptomatic (indeterminateform, IF). Each year, �3% of them develop lesions in the heart or gastrointestinal tract. Cardiomyopathy(CCHD) is the most severe manifestation of Chagas disease. The factors that determine the outcome of theinfection are unknown, but certainly depend on complex interactions amongst the genetic make-up ofthe parasite, the host immunogenetic background and environment. In a previous study we verified thatthe maxicircle gene NADH dehydrogenase (mitochondrial complex I) subunit 7 (ND7) from IF isolates hada 455 bp deletion compared with the wild type (WT) ND7 gene from CCHD strains. We proposed that ND7could constitute a valuable target for PCR assays in the differential diagnosis of the infective strain. In thepresent study we evaluated this hypothesis by examination of ND7 structure in parasites from 75 patientswith defined pathologies, from Southeast Brazil. We also analysed the structure of additional mitochon-drial genes (ND4/CR4, COIII and COII) since the maxicircle is used for clustering Trypanosoma cruzi strainsinto three clades/haplogroups. We conclude that maxicircle genes do not discriminate parasite popula-tions which induce IF or CCHD forms. Interestingly, the great majority of the analysed isolates belongto T. cruzi II (discrete typing unit, (DTU) IIb) genotype. This scenario is at variance with the prevalenceof hybrid (DTU IId) human isolates in Bolivia, Chile and Argentina. The distribution of WT and deletedND7 and ND4 genes in T. cruzi strains suggests that mutations in the two genes occurred in differentancestrals in the T. cruzi II cluster, allowing the identification of at least three mitochondrial sub-lineageswithin this group. The observation that T. cruzi strains accumulate mutations in several genes coding forcomplex I subunits favours the hypothesis that complex I may have a limited activity in this parasite.
� 2009 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved.
1. Introduction
Chagas disease is one of the most important parasitic afflictionsin Latin America. The illness, caused by the protozoan Trypanosomacruzi, is transmitted mainly by reduviid insect vectors, blood trans-fusion or infected mothers during pregnancy. In recent years, agreat number of acute cases have been reported, mostly from out-breaks in the Amazon region that were attributed to oral transmis-sion. Following infection, a short acute phase is characterized by aprofuse parasitemia and mild symptoms. The disease proceeds toa chronic phase, with scarce parasitemia and an unpredictable clin-
sitology Inc. Published by Elsevier
7; fax: +55 11 38155579.
Parasitología Tropical, Facul-bagué, Colombia.
ical course. Most of the chronic individuals are asymptomatic andshow no electrocardiographic or radiologic alterations in the heart,esophagus or colon. They present positive serological tests forT. cruzi infection and, for many of them, the xenodiagnosis andPCR results may be repeatedly positive for many years. Theseindividuals with the ‘‘indeterminate” form (IF) of the disease willremain asymptomatic for decades or the rest of their lives. Never-theless, each year approximately 3% will develop lesions in theheart or gastrointestinal tract (Dias, 2006). Chronic cardiomyopa-thy, or Chagas chronic heart disease (CCHD), is the most commonand severe manifestation of human Chagas disease, affectingapproximately 30% of patients. Its basic lesions are focal or exten-sive myocardial fibrosis, which result from myocardial cell destruc-tion due to direct parasite action, inflammatory response andneuron involvement. The gastrointestinal manifestations consistof progressive enlargement of the esophagus or colon, caused by
Ltd. All rights reserved.

964 J.C. Carranza et al. / International Journal for Parasitology 39 (2009) 963–973
chronic inflammation and destruction of parasympathetic neurons.The outcome of infection in a particular individual is the result of aset of complex interactions amongst the genetic make-up of the par-asite, the host immunogenetic background and environmental fac-tors, all of which can be complicated by mixed infections and re-infections (reviewed by Campbell et al., 2004; Macedo et al., 2004).
Geographical variations in the prevalence of clinical forms andmorbidity of Chagas disease have been reported. In Brazil, the IFis the most common (60–70%), followed by the cardiac and diges-tive forms (20–30% and 8–10%, respectively); there is also a mixedcardio-digestive form. In Central Brazil and Chile the digestiveform predominates, whilst it is practically absent in Venezuelaand Central America (Luquetti et al., 1986; Dias, 1992; Couraet al., 1999). The reason for this geographical variability and whydifferent patients develop different clinical forms is unknown,however major genetic differences between the human popula-tions who inhabit these areas are probably not the culprits. Thus,the clinical presentations are thought to be determined primarilyby parasite heterogeneity (reviewed by Macedo et al., 2004). Infact, T. cruzi is represented by a pool of populations, also namedas isolates or strains, which display great diversity in biologicalbehaviour, such as different degrees of virulence and tissue tro-pism in experimental animals and humans, and distinct suscepti-bilities to drugs. The explanation for such phenotypic diversityresides in the fact that T. cruzi is a diploid organism that multipliespredominantly by binary division. Accordingly, each isolateevolves independently and is genetically distinct (Tibayrenc andAyala, 1988). The clonal population structure of T. cruzi does notpreclude the occurrence of rare genetic exchange amongst parasitestrains (Tibayrenc and Ayala, 2002).
Different experimental approaches have yielded varying num-bers of biologically and genetically significant sub-groups ofT. cruzi (reviewed by Campbell et al., 2004; Macedo et al., 2004;Westenberger et al., 2005). Based on multilocus enzyme electro-phoresis (MLEE) T. cruzi populations were distributed in the follow-ing principal zymodemes: Z1, Z2, and Z3 (Miles et al., 1978, 1980).Z1 and Z3 comprised isolates from the silvatic cycle (opossums,armadillos and triatomines) and Z2, isolates from the domesticcycle (humans and domestic animals). This classification waswidely used by the scientific community since distinct parasitegroups could be associated with specific epidemiological transmis-sion patterns. However, the zymodeme typing approach requires asubstantial amount of parasites, which might eliminate particulargenotypes that are not adapted to culture conditions. In order toobtain faster and more sensitive characterization tools, we devel-oped two straightforward PCR assays directed to the D7 divergentdomain of the 24Sa rRNA gene and the intergenic region of themini-exon gene. The two genes showed dimorphism in the targetregions and allowed the division of T. cruzi strains into two groups(Souto and Zingales, 1993; Souto et al., 1996). A third group pre-senting both D7 PCR products and denoted rDNA group 1/2 wasalso detected and we put forward that it could correspond tohybrid isolates (Souto et al., 1996).
Several laboratories reported the division of T. cruzi into twomajor groups, which received distinct designations. To homogenisethe nomenclature, the groups were named T. cruzi I and T. cruzi IIby consensus (Anonymous, 1999). These groups correspond,respectively, to Miles’ zymodemes Z1 and Z2. Zymodeme Z3 andhybrid isolates were not included in either group, since therewas not complete agreement regarding its phylogenetic status(Anonymous, 1999). Epidemiological studies confirmed that T. cru-zi II strains were related to the domestic cycle in Southern Conecountries where Chagas disease is more severe (Fernandes et al.,1998; Zingales et al., 1998). At least in Brazil, T. cruzi II strains ap-pear to be exclusively responsible for tissue lesions (Freitas et al.,2005; Valadares et al., 2008).
Further studies based on randomly amplified polymorphic DNA(RAPD) and MLEE proposed that T. cruzi could be subdivided intosix sub-groups or discrete typing units (DTUs) designated I,IIa–IIe (Brisse et al., 2000, 2001). DTU I corresponds to the T. cruziI major group and DTU IIb to the T. cruzi II group (Brisse et al.,2001). DTUs IIa and IIc are equivalent to Miles’ Z3 and its respec-tive B and A subdivisions disclosed by regions of the ribosomalRNA cistron (Mendonça et al., 2002). The hybrid nature of DTUsIIa, IIc–IIe was supported by several lines of evidence (reviewedin Campbell et al., 2004). Gene genealogies of mitochondrialsequences were also used to analyse the population structure ofT. cruzi. An extensive maxicircle survey examined a 1.25 kb frag-ment, identifying three mitochondrial clades that hold a correla-tion with the DTUs (Machado and Ayala, 2001): clade Acorresponds to DTU I; clade B to hybrid strains of DTUs IIa andIIc (Z3), IId and IIe; and clade C, to DTU IIb strains exclusively. Morerecently, the terms haplogroups and haplotypes were applied inthe description of T. cruzi population structure (Freitas et al., 2006).
Based on the dissection of multilocus nuclear and mitochondrialgenotypes, three ancestral genotypes were proposed: T. cruzi I(DTU I), T. cruzi II (DTU IIb) and T. cruzi III (DTU IIc) (Freitas et al.,2006). Alternatively, two major hybridization events have beenpostulated, with DTU I and DTU IIb fusion resulting in the hybridDTU IIa and IIc lineages, and a more recent backcross of DTU IIband IIc producing the heterozygous hybrid DTUs IId and IIe(Westenberger et al., 2005). The genetic heterogeneity of the T. cru-zi I group was examined in Colombian isolates from different hostsand vectors (Herrera et al., 2007). Based on genealogies of themini-exon gene intergenic region, four sub-groups were disclosed,which correlated with specific transmission cycles.
Different laboratories proposed different designations for thesame T. cruzi subdivisions. Whilst illustrating the high geneticdiversity of T. cruzi, these studies failed to demonstrate a correla-tion between Chagas disease clinical presentation with particulargroups/DTUs/clades of strains. The nomenclature adopted hereand the equivalence with other designations is summarised inTable 1.
In kinetoplastids the mitochondrial genome is represented by20–50 maxicircles which, together with thousands of minicircles,constitute a dense network or kinetoplast (kDNA). Maxicirclesare the functional equivalent of the mitochondrial DNA of othereukaryotes, containing genes for mitochondrial rRNAs and mito-chondrial proteins mostly involved in the membrane-bound oxida-tive phosphorylation pathway (Simpson et al., 1987). Themaxicircle genomes of the CL Brener (DTU IIe/clade B) and Esmer-aldo (DTU IIb/clade C) strains were assembled and annotated fromdata generated by the T. cruzi Sequencing Consortium (Westenber-ger et al., 2006). The non-coding components of the maxicirclehave strain-specific repetitive regions and a variable region thatis unique for each strain. In the coding region, strain-specific inser-tions and deletions in particular genes were reported (Westenber-ger et al., 2006); whether these changes are due to prolonged timein tissue culture or characteristic of strains or larger sub-groups re-mains to be determined.
In a previous study, we showed that the NADH dehydrogenasesubunit 7 maxicircle gene (ND7) of isolates from individuals withthe IF of Chagas disease presented a 455 bp deletion in the centreof the coding region relative to wild type (WT) ND7 gene of the CLBrener strain and of isolates from patients with CCHD. The ND7gene structure correlated with disease manifestation for 20 isolatesfrom clinically characterised, chronic phase patients (Baptistaet al., 2006). Based on these observations, we proposed that ND7could constitute a valuable target for PCR assays in the differentialdiagnosis of the infective T. cruzi strain. In the present study, weevaluated this hypothesis by examining 84 recent parasite isolatesfrom 75 patients with defined pathologies. We also analysed the
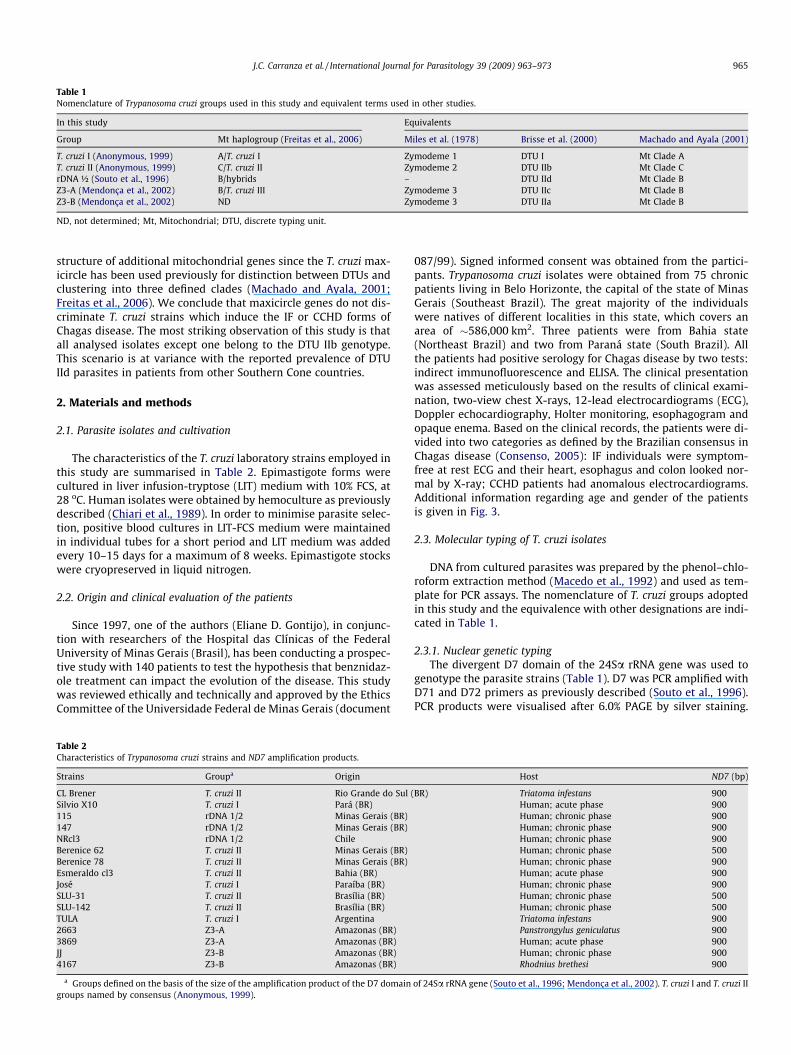
Table 1Nomenclature of Trypanosoma cruzi groups used in this study and equivalent terms used in other studies.
In this study Equivalents
Group Mt haplogroup (Freitas et al., 2006) Miles et al. (1978) Brisse et al. (2000) Machado and Ayala (2001)
T. cruzi I (Anonymous, 1999) A/T. cruzi I Zymodeme 1 DTU I Mt Clade AT. cruzi II (Anonymous, 1999) C/T. cruzi II Zymodeme 2 DTU IIb Mt Clade CrDNA ½ (Souto et al., 1996) B/hybrids – DTU IId Mt Clade BZ3-A (Mendonça et al., 2002) B/T. cruzi III Zymodeme 3 DTU IIc Mt Clade BZ3-B (Mendonça et al., 2002) ND Zymodeme 3 DTU IIa Mt Clade B
ND, not determined; Mt, Mitochondrial; DTU, discrete typing unit.
J.C. Carranza et al. / International Journal for Parasitology 39 (2009) 963–973 965
structure of additional mitochondrial genes since the T. cruzi max-icircle has been used previously for distinction between DTUs andclustering into three defined clades (Machado and Ayala, 2001;Freitas et al., 2006). We conclude that maxicircle genes do not dis-criminate T. cruzi strains which induce the IF or CCHD forms ofChagas disease. The most striking observation of this study is thatall analysed isolates except one belong to the DTU IIb genotype.This scenario is at variance with the reported prevalence of DTUIId parasites in patients from other Southern Cone countries.
2. Materials and methods
2.1. Parasite isolates and cultivation
The characteristics of the T. cruzi laboratory strains employed inthis study are summarised in Table 2. Epimastigote forms werecultured in liver infusion-tryptose (LIT) medium with 10% FCS, at28 oC. Human isolates were obtained by hemoculture as previouslydescribed (Chiari et al., 1989). In order to minimise parasite selec-tion, positive blood cultures in LIT-FCS medium were maintainedin individual tubes for a short period and LIT medium was addedevery 10–15 days for a maximum of 8 weeks. Epimastigote stockswere cryopreserved in liquid nitrogen.
2.2. Origin and clinical evaluation of the patients
Since 1997, one of the authors (Eliane D. Gontijo), in conjunc-tion with researchers of the Hospital das Clínicas of the FederalUniversity of Minas Gerais (Brasil), has been conducting a prospec-tive study with 140 patients to test the hypothesis that benznidaz-ole treatment can impact the evolution of the disease. This studywas reviewed ethically and technically and approved by the EthicsCommittee of the Universidade Federal de Minas Gerais (document
Table 2Characteristics of Trypanosoma cruzi strains and ND7 amplification products.
Strains Groupa Origin
CL Brener T. cruzi II Rio Grande do Sul (Silvio X10 T. cruzi I Pará (BR)115 rDNA 1/2 Minas Gerais (BR)147 rDNA 1/2 Minas Gerais (BR)NRcl3 rDNA 1/2 ChileBerenice 62 T. cruzi II Minas Gerais (BR)Berenice 78 T. cruzi II Minas Gerais (BR)Esmeraldo cl3 T. cruzi II Bahia (BR)José T. cruzi I Paraíba (BR)SLU-31 T. cruzi II Brasília (BR)SLU-142 T. cruzi II Brasília (BR)TULA T. cruzi I Argentina2663 Z3-A Amazonas (BR)3869 Z3-A Amazonas (BR)JJ Z3-B Amazonas (BR)4167 Z3-B Amazonas (BR)
a Groups defined on the basis of the size of the amplification product of the D7 domaingroups named by consensus (Anonymous, 1999).
087/99). Signed informed consent was obtained from the partici-pants. Trypanosoma cruzi isolates were obtained from 75 chronicpatients living in Belo Horizonte, the capital of the state of MinasGerais (Southeast Brazil). The great majority of the individualswere natives of different localities in this state, which covers anarea of �586,000 km2. Three patients were from Bahia state(Northeast Brazil) and two from Paraná state (South Brazil). Allthe patients had positive serology for Chagas disease by two tests:indirect immunofluorescence and ELISA. The clinical presentationwas assessed meticulously based on the results of clinical exami-nation, two-view chest X-rays, 12-lead electrocardiograms (ECG),Doppler echocardiography, Holter monitoring, esophagogram andopaque enema. Based on the clinical records, the patients were di-vided into two categories as defined by the Brazilian consensus inChagas disease (Consenso, 2005): IF individuals were symptom-free at rest ECG and their heart, esophagus and colon looked nor-mal by X-ray; CCHD patients had anomalous electrocardiograms.Additional information regarding age and gender of the patientsis given in Fig. 3.
2.3. Molecular typing of T. cruzi isolates
DNA from cultured parasites was prepared by the phenol–chlo-roform extraction method (Macedo et al., 1992) and used as tem-plate for PCR assays. The nomenclature of T. cruzi groups adoptedin this study and the equivalence with other designations are indi-cated in Table 1.
2.3.1. Nuclear genetic typingThe divergent D7 domain of the 24Sa rRNA gene was used to
genotype the parasite strains (Table 1). D7 was PCR amplified withD71 and D72 primers as previously described (Souto et al., 1996).PCR products were visualised after 6.0% PAGE by silver staining.
Host ND7 (bp)
BR) Triatoma infestans 900Human; acute phase 900Human; chronic phase 900Human; chronic phase 900Human; chronic phase 900Human; chronic phase 500Human; chronic phase 900Human; acute phase 900Human; chronic phase 900Human; chronic phase 500Human; chronic phase 500Triatoma infestans 900Panstrongylus geniculatus 900Human; acute phase 900Human; chronic phase 900Rhodnius brethesi 900
of 24Sa rRNA gene (Souto et al., 1996; Mendonça et al., 2002). T. cruzi I and T. cruzi II

966 J.C. Carranza et al. / International Journal for Parasitology 39 (2009) 963–973
Amplification products of 110 and 125 bp characterise, respec-tively, the T. cruzi I and T. cruzi II major groups (Anonymous,1999). The presence of both products identified the rDNA 1/2group (Souto et al., 1996). Z3 isolates, previously defined basedon the amplification of the non-transcribed spacer of the mini-exon gene (Fernandes et al., 2001), produced a 110 bp product(Z3-A) or a 117 bp product (Z3-B) (Mendonça et al., 2002).
2.3.2. Mitochondrial (Mt) maxicircle genetic typingThe locations of the primers employed in the PCR assays to am-
plify ND7, Cytochrome oxidase subunit III (COIII) and ND4/CR4 genesare shown in Fig. 1A. ND7 gene – PCR amplification was obtainedwith Tc.Maxi.ND7 For and Tc.Maxi.ND7 Rev primers under condi-tions described previously (Baptista et al., 2006). COIII gene– PCR amplification was performed with primers CO1 and CO2 asreported previously (Baptista et al., 2006). NADH dehydrogenasesubunit 4 (ND4) gene/intergenic region/C-rich region 4 (CR4) gene– PCR amplification was performed with primers ND4.For (50-AAA CTC TAT CTT TCG AAA ACC C-30) and ND4.Rev (50-GGG AAAAAT AGA CTT TCA AAA AGT ATC-30) designed, respectively, onCR4 (nucleotide 16,258) and ND4 genes (nucleotide 16,816) ofthe CL Brener sequence (GenBank Accession No. DQ343645). PCRconditions included initial 2 min denaturation at 94 oC, followedby 30 cycles at three temperatures: 1 min denaturation at 93 oC;1 min primer annealing at 55 oC; and 1 min elongation at 72 oC;followed by 10 min elongation at 72 oC. PCR products of ND7, COIIIand ND4/CR4 genes were separated in 0.8% agarose gels andstained with ethidium bromide (Fig. 1B). COII gene. PCR amplifica-tion was performed with primers TcMit10 (50-CCATATATTGTTGCATTATT-30) and TcMit21 (50-TTGTAATAGGAGTCATGTTT-30) (Fre-itas et al., 2006). The positions of the primer annealing sites areshown in Fig. 2A. PCR amplification consisted of an initial 5 mindenaturation at 94 �C, followed by 40 cycles of 45 s denaturationat 94 �C; 45 s annealing at 48 �C and 1 min elongation at 72 �C (Fre-itas et al., 2006). Subsequently, 10 lL of each reaction was digestedwith AluI (Promega), according to the manufacturer’s instructions.Restriction Fragment Length Polymorphism (RFLP) assays were
Fig. 1. Maxicircle typing assays. (A) Organization of maxicircle genes ND7, COIII, CR4 andand CR4-ND4 deletions are shown by hatched boxes. Oligonucleotide pairs used for PCRdiagonal lines represent discontinuities in the scale. (B) Amplification products of maxiciand stained with ethidium bromide. Sizes are given in bp. (C) ND7 and ND4/CR4 amplicona no-template control.
visualised in silver stained 6% polyacrylamide gels. The three AluIsites and the expected fragment sizes for T. cruzi I/Mt HaplogroupA; T. cruzi III + hybrids/Mt Haplogroup B and T. cruzi II/Mt Haplo-group C (Freitas et al., 2006; see Table 1) are shown in Fig. 2Aand B.
2.4. Microsatellite analysis
Amplification of microsatellites by PCR was achieved as de-scribed previously (Valadares et al., 2008). Briefly, the reactionswere performed in total volume of 15 lL containing 10 mM Tris–HCl pH 9.0, 50 mM KCl, 0.1% Triton X-100 (Buffer B, Promega,WI, USA), 2.5 mM MgCl2 (Promega), 0.5 U Taq DNA Polymerase(Promega), 250 lM of each dNTP, 0.3 lM of each primer (the se-quence of the primers is shown in Supplementary Table S1) and3 lL DNA (1 ng/lL). The amplifications were conducted in aPT100 termocycler (MJ Research) using a step-down protocol: ini-tial denaturation at 94 �C for 5 min; annealing at 58 �C for 30 s; pri-mer extension at 72 �C for 1 min and denaturation at 94 �C for 30 s.Every five cycles, the annealing temperature was decreasedsequentially to 55, 53, 51 and 48 �C. At 48 �C the number of cycleswas increased to 15, followed by a final extension at 72 �C for10 min. To determinate the allele sizes, 1–3 lL of the PCR fluores-cent products were analysed in a 6% denaturing polyacrylamide gelin an A.L.F. sequencer (GE Healthcare, WI, USA) and compared withfluorescent DNA fragments of 50–500 bp using Allelelocator soft-ware (GE Healthcare).
2.5. Phylogenetic inference
To make phylogenetic inferences, we assumed a stepwisemutation model for the microsatellites, as described previously(Oliveira et al., 1998, 1999). As a measure of genetic distance be-tween any two strains we used the minimum number of muta-tional steps necessary to transform one into the other. Themicrosatellite multilocus genotypes were transformed into binarycharacters using the FACTOR programme from the Phylogeny
ND4. CL Brener maxicircle numbering is used as a reference. The location of the ND7are shown by black (ND7), grey (COIII) or stippled (CR4-ND4) arrowheads. Double
rcle genes. After PCR for the specified target, products were run on 0.8% agarose gelsheterogeneity. The sample numbers refer to the isolate/code shown in Table 4. (-) is
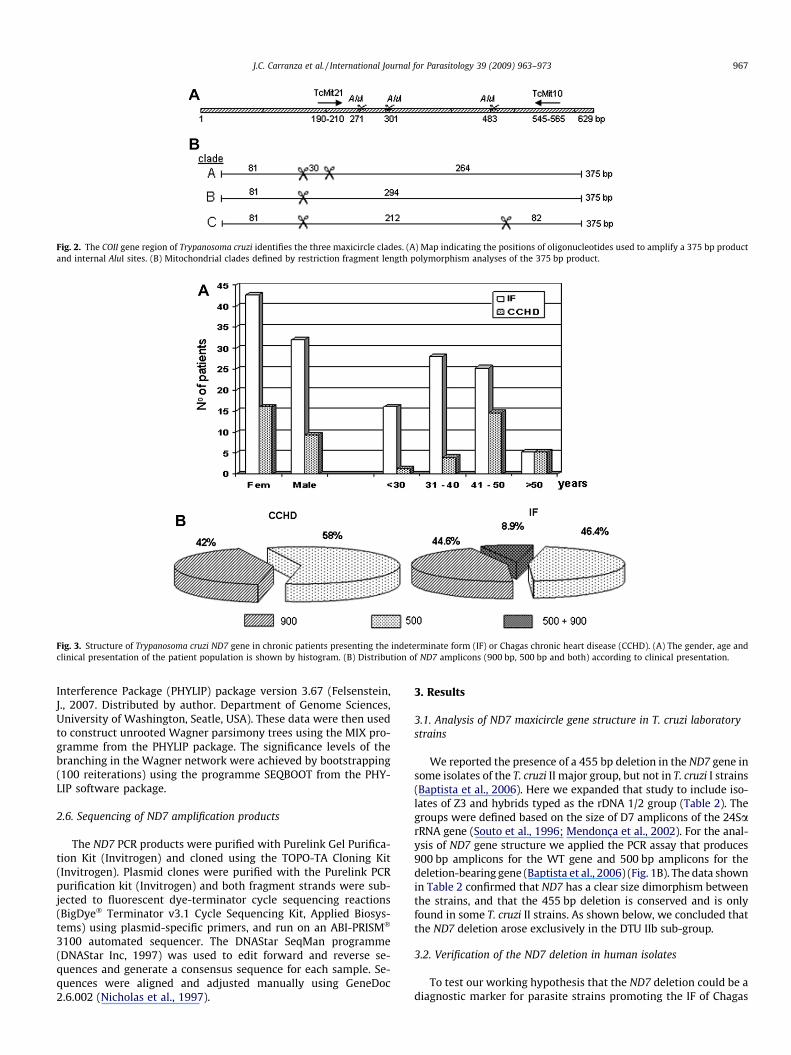
Fig. 2. The COII gene region of Trypanosoma cruzi identifies the three maxicircle clades. (A) Map indicating the positions of oligonucleotides used to amplify a 375 bp productand internal AluI sites. (B) Mitochondrial clades defined by restriction fragment length polymorphism analyses of the 375 bp product.
Fig. 3. Structure of Trypanosoma cruzi ND7 gene in chronic patients presenting the indeterminate form (IF) or Chagas chronic heart disease (CCHD). (A) The gender, age andclinical presentation of the patient population is shown by histogram. (B) Distribution of ND7 amplicons (900 bp, 500 bp and both) according to clinical presentation.
J.C. Carranza et al. / International Journal for Parasitology 39 (2009) 963–973 967
Interference Package (PHYLIP) package version 3.67 (Felsenstein,J., 2007. Distributed by author. Department of Genome Sciences,University of Washington, Seatle, USA). These data were then usedto construct unrooted Wagner parsimony trees using the MIX pro-gramme from the PHYLIP package. The significance levels of thebranching in the Wagner network were achieved by bootstrapping(100 reiterations) using the programme SEQBOOT from the PHY-LIP software package.
2.6. Sequencing of ND7 amplification products
The ND7 PCR products were purified with Purelink Gel Purifica-tion Kit (Invitrogen) and cloned using the TOPO-TA Cloning Kit(Invitrogen). Plasmid clones were purified with the Purelink PCRpurification kit (Invitrogen) and both fragment strands were sub-jected to fluorescent dye-terminator cycle sequencing reactions(BigDye� Terminator v3.1 Cycle Sequencing Kit, Applied Biosys-tems) using plasmid-specific primers, and run on an ABI-PRISM�
3100 automated sequencer. The DNAStar SeqMan programme(DNAStar Inc, 1997) was used to edit forward and reverse se-quences and generate a consensus sequence for each sample. Se-quences were aligned and adjusted manually using GeneDoc2.6.002 (Nicholas et al., 1997).
3. Results
3.1. Analysis of ND7 maxicircle gene structure in T. cruzi laboratorystrains
We reported the presence of a 455 bp deletion in the ND7 gene insome isolates of the T. cruzi II major group, but not in T. cruzi I strains(Baptista et al., 2006). Here we expanded that study to include iso-lates of Z3 and hybrids typed as the rDNA 1/2 group (Table 2). Thegroups were defined based on the size of D7 amplicons of the 24SarRNA gene (Souto et al., 1996; Mendonça et al., 2002). For the anal-ysis of ND7 gene structure we applied the PCR assay that produces900 bp amplicons for the WT gene and 500 bp amplicons for thedeletion-bearing gene (Baptista et al., 2006) (Fig. 1B). The data shownin Table 2 confirmed that ND7 has a clear size dimorphism betweenthe strains, and that the 455 bp deletion is conserved and is onlyfound in some T. cruzi II strains. As shown below, we concluded thatthe ND7 deletion arose exclusively in the DTU IIb sub-group.
3.2. Verification of the ND7 deletion in human isolates
To test our working hypothesis that the ND7 deletion could be adiagnostic marker for parasite strains promoting the IF of Chagas
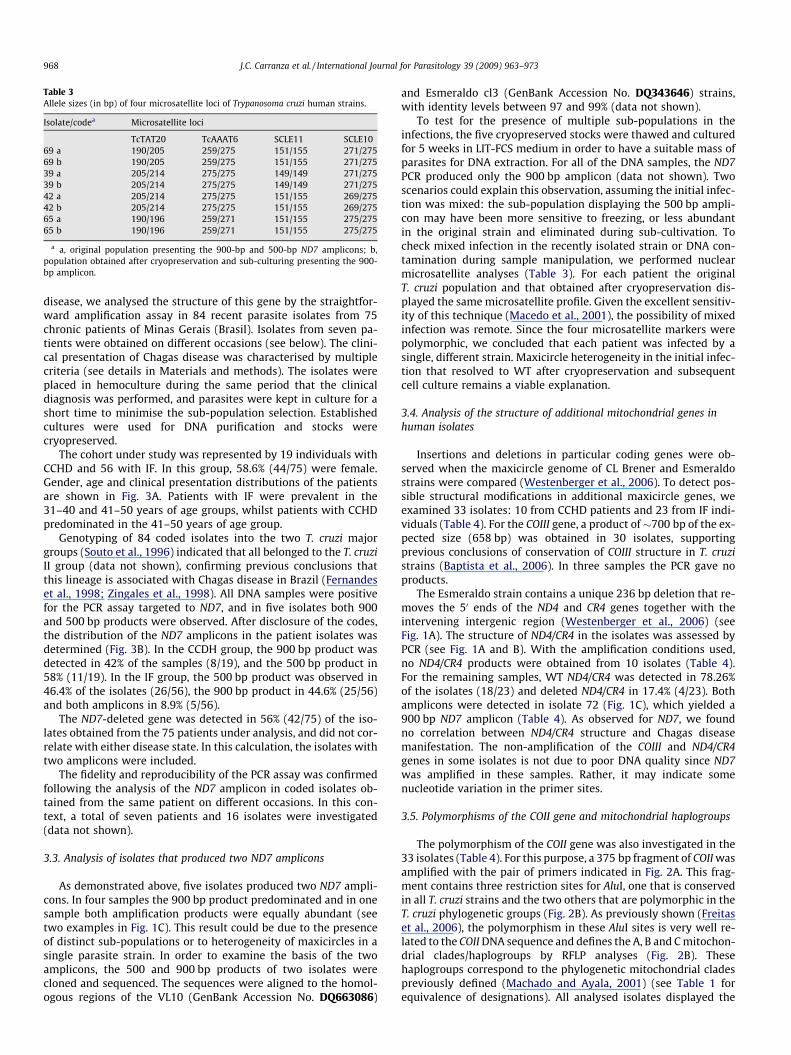
Table 3Allele sizes (in bp) of four microsatellite loci of Trypanosoma cruzi human strains.
Isolate/codea Microsatellite loci
TcTAT20 TcAAAT6 SCLE11 SCLE1069 a 190/205 259/275 151/155 271/27569 b 190/205 259/275 151/155 271/27539 a 205/214 275/275 149/149 271/27539 b 205/214 275/275 149/149 271/27542 a 205/214 275/275 151/155 269/27542 b 205/214 275/275 151/155 269/27565 a 190/196 259/271 151/155 275/27565 b 190/196 259/271 151/155 275/275
a a, original population presenting the 900-bp and 500-bp ND7 amplicons; b,population obtained after cryopreservation and sub-culturing presenting the 900-bp amplicon.
968 J.C. Carranza et al. / International Journal for Parasitology 39 (2009) 963–973
disease, we analysed the structure of this gene by the straightfor-ward amplification assay in 84 recent parasite isolates from 75chronic patients of Minas Gerais (Brasil). Isolates from seven pa-tients were obtained on different occasions (see below). The clini-cal presentation of Chagas disease was characterised by multiplecriteria (see details in Materials and methods). The isolates wereplaced in hemoculture during the same period that the clinicaldiagnosis was performed, and parasites were kept in culture for ashort time to minimise the sub-population selection. Establishedcultures were used for DNA purification and stocks werecryopreserved.
The cohort under study was represented by 19 individuals withCCHD and 56 with IF. In this group, 58.6% (44/75) were female.Gender, age and clinical presentation distributions of the patientsare shown in Fig. 3A. Patients with IF were prevalent in the31–40 and 41–50 years of age groups, whilst patients with CCHDpredominated in the 41–50 years of age group.
Genotyping of 84 coded isolates into the two T. cruzi majorgroups (Souto et al., 1996) indicated that all belonged to the T. cruziII group (data not shown), confirming previous conclusions thatthis lineage is associated with Chagas disease in Brazil (Fernandeset al., 1998; Zingales et al., 1998). All DNA samples were positivefor the PCR assay targeted to ND7, and in five isolates both 900and 500 bp products were observed. After disclosure of the codes,the distribution of the ND7 amplicons in the patient isolates wasdetermined (Fig. 3B). In the CCDH group, the 900 bp product wasdetected in 42% of the samples (8/19), and the 500 bp product in58% (11/19). In the IF group, the 500 bp product was observed in46.4% of the isolates (26/56), the 900 bp product in 44.6% (25/56)and both amplicons in 8.9% (5/56).
The ND7-deleted gene was detected in 56% (42/75) of the iso-lates obtained from the 75 patients under analysis, and did not cor-relate with either disease state. In this calculation, the isolates withtwo amplicons were included.
The fidelity and reproducibility of the PCR assay was confirmedfollowing the analysis of the ND7 amplicon in coded isolates ob-tained from the same patient on different occasions. In this con-text, a total of seven patients and 16 isolates were investigated(data not shown).
3.3. Analysis of isolates that produced two ND7 amplicons
As demonstrated above, five isolates produced two ND7 ampli-cons. In four samples the 900 bp product predominated and in onesample both amplification products were equally abundant (seetwo examples in Fig. 1C). This result could be due to the presenceof distinct sub-populations or to heterogeneity of maxicircles in asingle parasite strain. In order to examine the basis of the twoamplicons, the 500 and 900 bp products of two isolates werecloned and sequenced. The sequences were aligned to the homol-ogous regions of the VL10 (GenBank Accession No. DQ663086)
and Esmeraldo cl3 (GenBank Accession No. DQ343646) strains,with identity levels between 97 and 99% (data not shown).
To test for the presence of multiple sub-populations in theinfections, the five cryopreserved stocks were thawed and culturedfor 5 weeks in LIT-FCS medium in order to have a suitable mass ofparasites for DNA extraction. For all of the DNA samples, the ND7PCR produced only the 900 bp amplicon (data not shown). Twoscenarios could explain this observation, assuming the initial infec-tion was mixed: the sub-population displaying the 500 bp ampli-con may have been more sensitive to freezing, or less abundantin the original strain and eliminated during sub-cultivation. Tocheck mixed infection in the recently isolated strain or DNA con-tamination during sample manipulation, we performed nuclearmicrosatellite analyses (Table 3). For each patient the originalT. cruzi population and that obtained after cryopreservation dis-played the same microsatellite profile. Given the excellent sensitiv-ity of this technique (Macedo et al., 2001), the possibility of mixedinfection was remote. Since the four microsatellite markers werepolymorphic, we concluded that each patient was infected by asingle, different strain. Maxicircle heterogeneity in the initial infec-tion that resolved to WT after cryopreservation and subsequentcell culture remains a viable explanation.
3.4. Analysis of the structure of additional mitochondrial genes inhuman isolates
Insertions and deletions in particular coding genes were ob-served when the maxicircle genome of CL Brener and Esmeraldostrains were compared (Westenberger et al., 2006). To detect pos-sible structural modifications in additional maxicircle genes, weexamined 33 isolates: 10 from CCHD patients and 23 from IF indi-viduals (Table 4). For the COIII gene, a product of�700 bp of the ex-pected size (658 bp) was obtained in 30 isolates, supportingprevious conclusions of conservation of COIII structure in T. cruzistrains (Baptista et al., 2006). In three samples the PCR gave noproducts.
The Esmeraldo strain contains a unique 236 bp deletion that re-moves the 50 ends of the ND4 and CR4 genes together with theintervening intergenic region (Westenberger et al., 2006) (seeFig. 1A). The structure of ND4/CR4 in the isolates was assessed byPCR (see Fig. 1A and B). With the amplification conditions used,no ND4/CR4 products were obtained from 10 isolates (Table 4).For the remaining samples, WT ND4/CR4 was detected in 78.26%of the isolates (18/23) and deleted ND4/CR4 in 17.4% (4/23). Bothamplicons were detected in isolate 72 (Fig. 1C), which yielded a900 bp ND7 amplicon (Table 4). As observed for ND7, we foundno correlation between ND4/CR4 structure and Chagas diseasemanifestation. The non-amplification of the COIII and ND4/CR4genes in some isolates is not due to poor DNA quality since ND7was amplified in these samples. Rather, it may indicate somenucleotide variation in the primer sites.
3.5. Polymorphisms of the COII gene and mitochondrial haplogroups
The polymorphism of the COII gene was also investigated in the33 isolates (Table 4). For this purpose, a 375 bp fragment of COII wasamplified with the pair of primers indicated in Fig. 2A. This frag-ment contains three restriction sites for AluI, one that is conservedin all T. cruzi strains and the two others that are polymorphic in theT. cruzi phylogenetic groups (Fig. 2B). As previously shown (Freitaset al., 2006), the polymorphism in these AluI sites is very well re-lated to the COII DNA sequence and defines the A, B and C mitochon-drial clades/haplogroups by RFLP analyses (Fig. 2B). Thesehaplogroups correspond to the phylogenetic mitochondrial cladespreviously defined (Machado and Ayala, 2001) (see Table 1 forequivalence of designations). All analysed isolates displayed the

Table 4Structure of ND7, ND4/CR4 and COIII genes and the mitochondrial (Mt) haplogroupdefined by COII restriction fragment length polymorphism.
Isolate/code
Clinicalmanifestation
ND7 ND4/CR4 COIII COII Mthaplogroupa
46 IF D NA WT C59 IF D NA WT C88 CCHD D NA WT C22 IF D WT WT C23 IF D WT WT C26 CCHD D WT WT C28 IF D WT WT C38 IF D WT WT ND45 IF D WT WT C2 CCHD D WT WT C1 CCHD D WT WT C75 CCHD D WT WT C80 IF D WT WT C65 IF D/WT NA NA C69 IF D/WT NA NA C42 IF D/WT WT WT C39 IF D/WT WT NA NA40 IF D/WT WT WT C5 IF WT D WT C74 IF WT D WT C19 IF WT D WT C73 CCHD WT D WT C72 CCHD WT D/WT WT C53 IF WT NA WT C55 IF WT NA WT C36 CCHD WT NA WT C67 IF WT NA WT C21 IF WT NA WT C54 IF WT WT WT C50 IF WT WT WT C62 IF WT WT WT C24 CCHD WT WT WT C32 CCHD WT WT WT C
NA, no amplification; ND, not determined; WT, wild type gene; D, deletion-bearinggene; D/WT, both amplicons; IF, Indeterminate form; CCHD, Chagas chronic heartdisease.
a According to Freitas et al. (2006).
J.C. Carranza et al. / International Journal for Parasitology 39 (2009) 963–973 969
mitochondrial haplogroup C (Table 4), characteristic of T. cruzi II/DTU IIb. In one sample no PCR amplification was obtained. Besidesthe 33 samples, we defined the RLFP pattern of 20 additional iso-lates and all of those, except one, displayed the haplogroup C(DTU IIb) pattern. The isolate at variance was typed as mitochon-drial haplogroup B, which represents zymodeme 3 (DTU IIa/IIc) orhybrid (DTU IId/IIe) strains (Freitas et al., 2006) (see Table 1).
3.6. Genetic relationship between isolates with different mitochondrialgene structures
In order to further investigate the genetic relationship amongstthe DTU IIb isolates presenting different mitochondrial gene struc-tures, we analysed the microsatellite profile of 31 isolates: 12 WTprofiles, 10 DND7, four DND4/CR4 and five with both ND7 ampli-cons. The allele sizes of the microsatellite loci are shown in Supple-mentary Table S2. The multilocus microsatellite genotypesobtained with four microsatellite loci were used as a distance mea-sure for building a Wagner parsimony network (Fig. 4). No specificcluster correlated with the ND4 or ND7 deletions or with the twoND7 amplicons. This dataset will be discussed below.
4. Discussion
Enormous advances have been made in the understanding ofChagas disease, since Carlos Chagas first described it almost 100years ago. Still, despite the observation that CCHD affects 20–30%of asymptomatic individuals and needs 10–30 years for develop-
ment, the identification of either the genetic make-up of the infect-ing T. cruzi strains or the genetic background of the infectedindividuals for early follow-up and medical care has remainedelusive.
Recently our group has been investigating different nuclear andmitochondrial markers in an attempt to correlate the genetic var-iability of the parasites with the clinical forms of the disease.Although these studies indicated the clear predominance of theT. cruzi II major group in patients, neither RAPD analyses (D’Ávilaet al., 2006) nor kDNA minicircle signatures (Lages-Silva et al.,2006) showed any correlation with the clinical manifestations ofChagas disease.
In this study, and based on preliminary evidence (Baptista et al.,2006), we initially approached this issue by investigating the ge-netic structure of the ND7 maxicircle gene. According to our prog-nostic hypothesis, the individuals who are asymptomatic (IF) andharbour T. cruzi strains with WT ND7 would be at risk of developingchagasic cardiomyopathy in the future (Baptista et al., 2006). Thissupposition could explain the presence of 900 bp amplicons in theisolates obtained from 44.6% of IF patients (Fig. 3B). If on one sidewe verified that two patients who were asymptomatic in 1999 andwhose isolates had the WT ND7 gene developed CCHD in 2004 (onepatient) and 2006 (the other patient) (data not shown), the behav-iour of Chagas disease indicates that only 20–30% of asymptomaticindividuals develop cardiomyopathies. Additionally, the presenceof the ND7 deletion in 58% of CCHD isolates (Fig. 3B) rules outour working hypothesis. Interestingly, we noticed that 10 patientsout of 11 with CCHD harbouring isolates with the ND7 deletionwere female. This could reflect the prevalence of females (58.6%)in the cohort under study.
The examination of the genetic structure of additional maxicir-cle genes in parasites recently isolated by hemoculture indicatedthat neither ND7, nor the ND4/CR4 region showed any associationwith Chagas disease manifestations. On the other hand, the analy-sis of polymorphisms of the COII gene and mitochondrial haplo-groups lead to a remarkable observation: all analysed isolates,except one, belonged to the T. cruzi II (DTU IIb) major group.
A few reports show that the parasite population infecting spe-cific organs can be genetically distinct from the population foundin the patients’ blood (Burgos et al., 2005, 2008; Vago et al.,2000; Valadares et al., 2008). Bearing this possibility in mind, itis unlikely to occur in the majority of patients. Verification of thestrains infecting the heart tissue of living patients is not viable.
The high predominance of DTU IIb strains in Brazilian patientscontrasts with epidemiological studies related to the distributionof the strain genotypes of patients from Bolivia, Chile and Argen-tina. In 89 Bolivian patients with chronic infection, the presenceof DTU I and DTU IId was verified in 42% and 50% of the individuals,respectively (Brenière et al., 1989). A different scenario was foundin 93 young Bolivian patients where the incidence of infectioncaused by DTU IId was 85%. DTU I was identified in only one pa-tient and a mixture of the two genotypes in 18.3% (Brenièreet al., 1998). The distribution of these genotypes in the young pa-tients was significantly different from that in the vectors of thesame area (Triatoma infestans), where both groups of strains werefound with comparable frequencies. These observations suggestedsome degree of selection in the transmission of the two genotypesand a drastic control of DTU I parasitemia by the immune system ofchild patients (Brenière et al., 1998). DTU IId was also the mostprevalent genotype (frequency of 0.84) circulating in 73 chronicchagasic patients in two geographic regions of Chile (Solari et al.,2001). The possible association of certain parasite genotypes withvertical transmission was investigated in blood samples fromArgentinean and Bolivian patients from endemic areas. DTU IIdwas the prevalent genotype amongst 36/38 congenitally infectedinfants, 5/5 mothers who transmitted Chagas disease, 12/13 moth-

Fig. 4. Population structure of Trypanosoma cruzi from patients. Unrooted Wagner network of 31 T. cruzi isolates based on the amplification of four microsatellite loci (SCLE10,SCLE11, MCLF10 and MCLG10). The isolates that show deleted maxicircle amplicons are indicated. Significance levels above 75% are represented in the branching of thisnetwork and were achieved by bootstrapping (100 reiterations) using the programme SEQBOOT from the PHYLIP software package.
970 J.C. Carranza et al. / International Journal for Parasitology 39 (2009) 963–973
ers who delivered non-infected children, and 24/34 unrelated Cha-gas disease patients (Burgos et al., 2007).
The scenario of genotype prevalence observed in this study isalso at variance with that observed in the Brazilian Amazonia,where the most prevalent genotype causing Chagas disease is T.cruzi I, with occasional cases of Z3 infection (reviewed by Couraet al., 2002). Clinical and parasitological studies suggest that thelow morbidity in the area and the low parasitemias are correlatedwith the two genotypes, which apparently lead predominantly tothe IF of the disease (Coura et al., 2002).
The overall comprehension of the parasite and host genetic fac-tors that determine the outcome of Chagas infection will beachieved by a paired genetic analysis of the patient and of theinfecting strain. The zymodeme distribution associated with acuteand chronic Chagas disease in Central Brazil was established over20 years ago (Luquetti et al., 1986). Of 25 patients in the acutephase, Z1 was detected in 12 patients and Z2, in 13 patients. Inall 12 patients in the chronic phase only Z2 strains were found.These data suggest the potential effect of the immune system toeliminate Z1 strains. In the future, it will be of interest to classifyhuman isolates of Central Brazil in particular major lineages andsub-lineages, for comparison with the predominance of T. cruzi II/DTU IIb in patients from Southeast Brazil.
Epidemiological studies conducted in northern countries ofSouth America, Central America and in Mexico showed predomi-nance of T. cruzi I in both silvatic and domestic cycles (Herreraet al., 2007 and cited references). A survey performed in 23 acuteVenezuelan patients indicated that 74% of them were infected withT. cruzi I strains (Añez et al., 2004). Such was also the case forchronic chagasic patients of Colombia (Zafra et al., 2008). Interest-ingly, DTU IIb was detected in the blood of �10% of the individuals,indicating that in the region under study the two major T. cruzigroups induce Chagas disease (Zafra et al., 2008). In the above-
mentioned countries it will be of interest to verify whether thepathology caused by T. cruzi I strains is analogous to that observedin the Amazonian region where the same genotype circulates.
Taken together, the data in Southern Cone countries indicatethat strains of T. cruzi II (DTU IIb) and hybrids (DTU IId) promotethe indeterminate and cardiac forms of Chagas disease. Since themaxicircle genetic markers showed no correlation with the clinicalpresentation, the search for nuclear markers such as those poten-tially involved in preferential tissue tropism, virulence, resistanceto oxidative stress and escape from the immune system shouldbe pursued. Whether these markers exist or, instead, biologicalfunctions should be screened are matters for further investigation.
In addition, host genetic factors most certainly play a role in theoutcome of Chagas disease. The development of CCHD in only 20–30% of individuals infected with T. cruzi and the tendency for famil-ial aggregation (Zicker et al., 1990) suggest that host genetics maybe involved in susceptibility. The host control of parasite infectiondepends on a fine balance between the generation of host immuneresponses to keep the parasitemia restrained and the regulation ofthese responses to avoid tissue damage. The influence of the majorhuman histocompatibility system (HLA) markers in the outcome ofChagas disease was investigated in South American countries (re-viewed in Campbell et al., 2004). In the Brazilian population, noassociation between the risk of developing CCHD and two HLAclass II genes (HLA-DR and HLA-DQ) was reported (Faé et al.,2000), but the HLA class I A30 allele was over-represented in pa-tients with cardiac and digestive disorders compared with IF indi-viduals (Deghaide et al., 1998).
Patients with CCHD display high levels of circulating pro-inflammatory cytokines compared with patients with the IF.Heart-infiltrating lymphocytes from these patients also expressthe pro-inflammatory cytokines TNF-a and IFN-c (Reis et al.,1993; Abel et al., 2001; Gomes et al., 2003; Talvani et al., 2004).

Fig. 5. Events in the recent evolution of Trypanosoma cruzi mitochondrial DNA. Thethree mitochondrial (Mt) major clades and two sub-groups are shaded. Theindependent ND7 and ND4/CR4 deletions are unique to the clade C lineage andcannot be distinguished temporally. The deletions may be part of a dynamic processinvolving frequent cellular fusion to maintain maxicircle integrity or viable,functional entities. DTU, discrete typing unit.
J.C. Carranza et al. / International Journal for Parasitology 39 (2009) 963–973 971
Specific polymorphisms in BAT1 (Ramasawmy et al., 2006), whichdown-regulates TNF-a and IL6 expression, and the Lymphotoxin-a (Ramasawmy et al., 2007), a pro-inflammatory cytokine, areassociated with the risk of CCHD in Brazilian patients.
In our maxicircle analysis, a high prevalence of isolates bearingthe ND7 deletion was found (50%; 42/84). The conserved deletionsin ND7 and the ND4/CR4 region were found in only some of the DTUIIb strains. The presence of the ND4/CR4 deletion was investigatedpreviously in seven DTU IIb strains and 13 strains representing thefive other DTUs. PCR analysis indicated that only the Esmeraldostrain had this deletion, whilst neither of the DTU I representativesyielded amplification products (Westenberger et al., 2006). The un-ique appearance of this deletion in the Esmeraldo strain was inter-preted as a putative recent event tolerated in a cultured strain, butdeleterious in wild populations (Westenberger et al., 2006). Thefour human isolates that bear the identical ND4/CR4-deletion indi-cate that the loss of these genes is not terminal to survival of theparasite in the host and also that these strains may have a relation-ship with the Esmeraldo strain.
The conservation of the deletions in the ND7 or ND4/CR4 genesin some DTU IIb strains suggests that these events occurred afterthe separation of the T. cruzi II major lineage following the sche-matic shown in Fig. 5. Included are the three major T. cruzi Mtclades: clade A is shared by strains belonging to T. cruzi I/DTU I;Mt clade B is found in Z3 strains (DTU IIa, IIc/T. cruzi III) and theheterozygous hybrid strains (DTU IId, IIe); and Mt clade C is exclu-sive to DTU IIb strains. Within this latter group independent eventsdeleted ND7 and ND4/CR4, forming three distinct Mt sub-groupswithin clade C. No chronology can be determined. Further subdivi-sion of the A and B clades may be detected upon examination ofadditional strains.
Although not pursued in this study, the Esmeraldo ND5 gene,which codes for Mt complex I subunit 5, has three frameshiftmutations relative to the CL Brener sequence such that if trans-lated directly a truncated protein of 165 amino acids, comparedwith the 589 amino acid product of CL Brener, would be generated(Westenberger et al., 2006). The penetrance of these mutationscould be explored within the DTU IIb group. The abundance ofmutations in mitochondrial genes of T. cruzi II strains may be aconsequence of the decreased mismatch-repair ability conferredby specific single nucleotide polymorphisms (SNPs) in the mis-match-repair gene TcMSH2 of strains of mitochondrial clades B
and C, compared with strains assigned to clade A (Augusto-Pintoet al., 2003).
The presence and functionality of the NADH dehydrogenase, ormitochondrial complex I, in Trypanosomatidae has long been de-bated (reviewed by Opperdoes and Michels, 2008). The observationthat T. cruzi strains accumulate mutations in several genes coding forNADH dehydrogenase subunits favours the hypothesis that thiscomplex may have limited activity or altered function in thisparasite.
In this study, the presence of two ND7 amplicons in five recentparasite isolates was observed. However, after cryopreservation,thawing and sub-culturing only the WT ND7 gene was detected.Based on the consistent nuclear microsatellite profiles, we are in-clined to rule out the possibility of mixed infections in the recentisolates. An attractive possibility would be that those parasites pre-sented mitochondrial heteroplasmy, i.e., a mixture of mitochon-drial DNA genotypes within a strain, possibly as a result of recentfusions (Gaunt et al., 2003). As commonly observed, heteroplasmyis generally resolved to homoplasmy within a few generations (ref-erences in Doublet et al., 2008) and, in the present case, the deletedversions were lost. The recent isolates carrying only the deletedgenes may have resolved heteroplasmic states within the host infavour of the deletion-bearing maxicircle, or may carry a low rela-tive copy number of intact maxicircles. Alternatively, the conceptof maxicircle heterogeneity may represent another manifestationof the phenomenon of transkinetoplastidy involving bulk altera-tions of the minicircle populations in cultured T. cruzi clones (Alveset al., 1994).
A remarkable observation reported in this study was the lack ofassociation between the clusters defined by the microsatellite pro-files and the ND4/CR4 or ND7 deletions or the two ND7 amplicons.Although this could be justified by the fact that microsatellite lociare nuclear markers and the gene deletions are present in the mito-chondrial DNA, the absence of regular hybridization events withinT. cruzi taxon (Tibayrenc and Ayala, 1988; Oliveira et al., 1998; re-viewed by Macedo et al., 2004) should lead to linkage disequilib-rium between the two genomic counterparts. Our data could beexplained by the occurrence of multiclonal populations in a pro-portion below the sensitivity of the microsatellite analyses (Mac-edo et al., 2001). A provocative alternative is that hybridizationevents occur at higher frequencies than initially expected (re-viewed by Tibayrenc and Ayala, 2002). Although most of the linesin our study were characterised as DTU IIb, the microsatellite pro-files within the mitochondrial sub-clade C were distinct from oneanother. The identical mitochondrial deletion is unlikely to haveoccurred independently multiple times, thus the deletion itself isa marker of pedigree. The rapid assay of our freshly isolated strainsmay have allowed us to capture a transient state of heteroplasmythat reflects the constant state of genetic flux in the naturalpopulation.
Acknowledgements
We thank Marcelo N. Silva for technical assistance. This workwas supported by grants of Fundação de Amparo à Pesquisa doEstado de São Paulo (FAPESP), Ministério de Ciência e Tecnologia/Conselho Nacional de Desenvolvimento Científico e Tecnológico/Ministério da Saúde (MCT/CNPq/MS-SCTIE-DECIT-Edital de Doe-nças Negligenciadas) (B.Z.) and Fundação de Amparo à Pesquisado Estado de Minas Gerais (FAPEMIG) (A.M.M.). N.S. received sup-port from NIH Grant AI056034. J.C.C. was the recipient of a gradu-ation fellowship from the Universidad del Tolima (Ibagué,Colombia) and Instituto Colombiano Francisco José de Caldas(COLCIENCIAS).

972 J.C. Carranza et al. / International Journal for Parasitology 39 (2009) 963–973
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.ijpara.2009.01.009.
References
Abel, L.C., Rizzo, L.V., Ianni, B., Albuquerque, F., Bacal, F., Carrara, D., Bocchi, E.A.,Teixeira, H.C., Mady, C., Kalil, J., Cunha-Neto, E., 2001. Chronic Chagas’ diseasecardiomyopathy patients display an increased IFN-gamma response toTrypanosoma cruzi infection. J. Autoimmun. 17, 99–107.
Alves, A.M., Almeida, D.F., von Krüger, W.M., 1994. Changes in Trypanosoma cruzikinetoplast DNA minicircles induced by environmental conditions andsubcloning. J. Eukaryot. Microbiol. 41, 415–419.
Añez, N., Crisante, G., da Silva, F.M., Rojas, A., Carrasco, H., Umezawa, E.S., Stolf, A.M.,Ramírez, J.L., Teixeira, M.M., 2004. Predominance of lineage I amongTrypanosoma cruzi isolates from Venezuelan patients with different clinicalprofiles of acute Chagas’ disease. Trop. Med. Int. Health 9, 1319–1326.
Anonymous, 1999. Recommendations from a satellite meeting. Mem. Inst. OswaldoCruz 94 (Suppl. 1), 429–432.
Augusto-Pinto, L., Teixeira, S.M., Pena, S.D., Machado, C.R., 2003. Single-nucleotidepolymorphisms of the Trypanosoma cruzi MSH2 gene support the existence ofthree phylogenetic lineages presenting differences in mismatch-repairefficiency. Genetics 164, 117–126.
Baptista, C.S., Vêncio, R.Z., Abdala, S., Carranza, J.C., Westenberger, S.J., Silva, M.N.,Pereira, C.A., Galvão, L.M., Gontijo, E.D., Chiari, E., Sturm, N.R., Zingales, B., 2006.Differential transcription profiles in Trypanosoma cruzi associated with clinicalforms of Chagas disease: Maxicircle NADH dehydrogenase subunit 7 genetruncation in asymptomatic patient isolates. Mol. Biochem. Parasitol. 150, 236–248.
Brenière, S.F., Bosseno, M.F., Telleria, J., Bastrenta, B., Yacsik, N., Noireau, F., Alcazar,J.L., Barnabé, C., Wincker, P., Tibayrenc, M., 1998. Different behavior of twoTrypanosoma cruzi major clones: transmission and circulation in young Bolivianpatients. Exp. Parasitol. 89, 285–295.
Brenière, S.F., Carrasco, R., Revollo, S., Aparicio, G., Desjeux, P., Tibayrenc, M., 1989.Chagas’ disease in Bolivia: clinical and epidemiological features and zymodemevariability of Trypanosoma cruzi strains isolated from patients. Am. J. Trop. Med.Hyg. 41, 521–529.
Brisse, S., Barnabé, C., Tibayrenc, M., 2000. Identification of six Trypanosoma cruziphylogenetic lineages by random amplified polymorphic DNA and multilocusenzyme electroforesis. Int. J. Parasitol. 30, 35–44.
Brisse, S., Verhoef, J., Tibayrenc, M., 2001. Characterisation of large and smallsubunit rRNA and mini-exon genes further supports the distinction of sixTrypanosoma cruzi lineages. Int. J. Parasitol. 31, 1218–1226.
Burgos, J.M., Altcheh, J., Bisio, M., Duffy, T., Valadares, H.M., Seidenstein, M.E.,Piccinali, R., Freitas, J.M., Levin, M.J., Macchi, L., Macedo, A.M., Freilij, H.,Schijman, A.G., 2007. Direct molecular profiling of minicircle signatures andlineages of Trypanosoma cruzi bloodstream populations causing congenitalChagas disease. Int. J. Parasitol. 37, 1319–1327.
Burgos, J.M., Begher, S., Freitas, J.M., Bisio, M., Duffy, T., Altcheh, J., Teijeiro, R., LopezAlcoba, H., Deccarlini, F., Freilij, H., Levin, M.J., Levalle, J., Macedo, A.M.,Schijman, A.G., 2005. Molecular diagnosis and typing of Trypanosoma cruzipopulations and lineages in cerebral Chagas disease in a patient with AIDS. Am.J. Trop. Med. Hyg. 73, 1016–1018.
Burgos, J.M., Begher, S., Valadares Silva, H.M., Bisio, M., Duffy, T., Levin, M.J., Macedo,A.M., Schijman, A.G., 2008. Case report: molecular identification of Trypanosomacruzi I tropism for central nervous system in Chagas reactivation due to AIDS.Am. J. Trop. Med. Hyg. 78, 294–297.
Campbell, D.A., Westenberger, S.J., Sturm, N.R., 2004. The determinants of Chagasdisease: connecting parasite and host genetics. Curr. Mol. Med. 4, 549–562.
Chiari, E., Dias, J., Lana, M., Chiari, C., 1989. Hemocultures for the parasitologicaldiagnosis of human chronic Chagas’ disease. Rev. Soc. Bras. Med. Trop. 22, 19–23.
Consenso Brasileiro em Doença de Chagas. Secretaria de Vigilância em Saúde doMinistério da Saúde., 2005. Rev. Soc. Bras. Med. Trop. 38 (Supl. III).
Coura, J.R., Borges-Pereira, J., Araujo, R.M., 1999. Morbidity and regional variation ofChagas disease in Brazil. Mem. Inst. Oswaldo Cruz 94 (Suppl. II), 26–27.
Coura, J.R., Junqueira, A.C., Fernandes, O., Valente, S.A., Miles, M.A., 2002. EmergingChagas disease in Amazonian Brazil. Trends Parasitol. 18, 171–176.
D’Ávila, D.A., Gontijo, E.D., Lages-Silva, E., Meira, W.S.F., Chiari, E., Galvão, L.M.C.,2006. Random amplified polymorphic DNA profiles of Trypanosoma cruziisolates from chagasic patients with different clinical forms. Parasitol. Res. 98,455–461.
Deghaide, N.H., Dantas, R.O., Donadi, E.A., 1998. HLA class I and II profiles of patientspresenting with Chagas’ disease. Dig. Dis. Sci. 43, 246–252.
Dias, J.C.P., 1992. Epidemiology of Chagas disease. In: Wendel, S., Brener, Z.,Camargo, M., Rassi, A. (Eds.), Chagas Disease (American Trypanosomiasis): ItsImpact on Transfusion and Clinical Medicine. ISBT, Brazil’92, São Paulo, pp. 49–80.
Dias, J.C.P., 2006. The treatment of Chagas disease (South Americantrypanosomiasis). Ann. Intern. Med. 144, 772–774.
Doublet, V., Souty-Grosset, C., Bouchon, D., Cordaux, R., Marcadé, R., 2008. A thirtymillion year-old inherited heteroplasmy. PLoS ONE 3, e2938.
Faé, K.C., Drigo, S.A., Cunha-Neto, E., Ianni, B., Mady, C., Kalil, J., Goldberg, A.C., 2000.HLA and beta-myosin heavy chain do not influence susceptibility to Chagasdisease cardiomyopathy. Microbes Infect. 2, 745–751.
Fernandes, O., Santos, S.S., Cupolillo, E., Mendonça, B., Derré, R., Junqueira, A.C.,Santos, L.C., Sturm, N.R., Naiff, R.D., Barrett, T.V., Campbell, D.A., Coura, J.R.,2001. Trypanosomiasis in the Brazilian Amazon – a mini-exon multiplex PCR todistinguish the major groups of Trypanosoma cruzi and Trypanosoma rangeli.Trans. R. Soc. Trop. Med. Hyg. 95, 1–3.
Fernandes, O., Sturm, N.R., Derré, R., Campbell, D.A., 1998. The mini-exon gene: agenetic marker for zymodeme III of Trypanosoma cruzi. Mol. Biochem. Parasitol.95, 129–133.
Freitas, J., Augusto-Pinto, L., Pimenta, J., Bastos-Rodrigues, L., Gonçalves, V., Teixeira,S., Chiari, E., Junqueira, A., Fernandes, O., Macedo, A., Machado, C., Pena, S., 2006.Ancestral genomes, sex, and the population structure of Trypanosoma cruzi. PLoSPathogens 2, 226–235.
Freitas, J.M., Lages-Silva, E., Crema, E., Pena, S.D., Macedo, A.M., 2005. Real time PCRstrategy for the identification of major lineages of Trypanosoma cruzi directly inchronically infected human tissues. Int. J. Parasitol. 35, 411–417.
Gaunt, M.W., Yeo, M., Frame, I.A., Stothard, J.R., Carrasco, H.J., Taylor, M.C., Mena,S.S., Veazey, P., Miles, G.A., Acosta, N., de Arias, A.R., Miles, M.A., 2003.Mechanism of genetic exchange in American trypanosomes. Nature 421, 936–939.
Gomes, J.A., Bahia-Oliveira, L.M., Rocha, M.O., Martins-Filho, O.A., Gazzinelli, G.,Correa-Oliveira, R., 2003. Evidence that development of severe cardiomyopathyin human Chagas’ disease is due to a Th1-specific immune response. Infect.Immun. 71, 1185–1193.
Herrera, C., Bargues, M.D., Fajardo, A., Montilla, M., Triana, O., Vallejo, G.A., Guhl, F.,2007. Identifying four Trypanosoma cruzi I isolate haplotypes from differentgeographic regions in Colombia. Infect. Genet. Evol. 7, 535–539.
Lages-Silva, E., Ramírez, L.E., Pedrosa, A.L., Crema, L., Galvão, L.M.C., Pena, S.D.J.,Macedo, A.M., Chiari, E., 2006. Variability of kinetoplast DNA gene signatures ofTrypanosoma cruzi II strains from patients with different clinical forms ofChagas’ disease in Brazil. J. Clin. Microbiol. 44, 2167–2171.
Luquetti, A.O., Miles, M.A., Rassi, A., de Rezende, J.M., de Souza, A.A., Póvoa, M.M.,Rodrigues, I., 1986. Trypanosoma cruzi: zymodemes associated with acute andchronic Chagas’ disease in central Brazil. Trans. R. Soc. Trop. Med. Hyg. 80, 462–470.
Macedo, A.M., Machado, C., Oliveira, R., Pena, S., 2004. Trypanosoma cruzi: geneticstructure of populations and relevance of genetic variability to the pathogenesisof Chagas disease. Mem. Inst. Oswaldo Cruz 99, 1–12.
Macedo, A.M., Martins, M., Chiari, E., Pena, S., 1992. DNA fingerprinting ofTrypanosoma cruzi: a new tool for characterization of strains and clones. Mol.Biochem. Parasitol. 55, 147–154.
Macedo, A.M., Pimenta, J.R., Aguiar, R.S., Melo, A.I.R., Chiari, E., Zingales, B., Pena,S.D., Oliveira, R.P., 2001. Usefulness of microsatellite typing in populationgenetic studies of Trypanosoma cruzi. Mem. Inst. Oswaldo Cruz 96, 407–413.
Machado, C.A., Ayala, F.J., 2001. Nucleotide sequences provide evidence of geneticexchange among distantly related lineages of Trypanosoma cruzi. Proc. Natl.Acad. Sci. USA 98, 7396–7401.
Mendonça, M.B., Nehme, N.S., Santos, S.S., Cupolillo, E., Vargas, N., Junqueira, A.,Naiff, R.D., Barrett, T.V., Coura, J.R., Zingales, B., Fernandes, O., 2002. Two mainclusters within Trypanosoma cruzi zymodeme 3 are defined by distinct regionsof the ribosomal RNA cistron. Parasitology 124, 177–184.
Miles, M.A., Lanham, S.M., de Souza, A.A., Póvoa, M., 1980. Further enzymiccharacters of Trypanosoma cruzi and their evaluation for strain identification.Trans. R. Soc. Trop. Med. Hyg. 74, 221–242.
Miles, M.A., Souza, A., Povoa, M., Shaw, J.J., Lainson, R., Toye, P.J., 1978. Isozymicheterogeneity of Trypanosoma cruzi in the first autochthonous patients withChagas’ disease in Amazonian Brazil. Nature 272, 819–821.
Nicholas, K.B., Nicholas, H.B.J., Deerfield, D.W., 1997. GeneDoc: Analysis andVisualization of Genetic Variation. EMBNEW. NEWS 4, 14.
Oliveira, R.P., Broude, N.E., Macedo, A.M., Cantor, C.R., Smith, C.L., Pena, S.D., 1998.Probing the genetic population structure of Trypanosoma cruzi withpolymorphic microsatellites. Proc. Natl. Acad. Sci. USA 95, 3776–3780.
Oliveira, R.P., Melo, A.I.R., Macedo, A.M., Chiari, E., Pena, S.D.J., 1999. The populationstructure of Trypanosoma cruzi: expanded analysis of 54 strains using eightpolymorphic CA-repeat microsatellites. Mem. Inst. Oswaldo Cruz 94, 65–70.
Opperdoes, F., Michels, P., 2008. Complex I of Trypanosomatidae: does it exist?Trends Parasitol. 24, 310–317.
Ramasawmy, R., Cunha-Neto, E., Faé, K.C., Müller, N.G., Cavalcanti, V.L., Drigo, S.A.,Ianni, B., Mady, C., Kalil, J., Goldberg, A.C., 2006. BAT1, a putative anti-inflammatory gene, is associated with chronic Chagas cardiomyopathy. J. Infect.Dis. 193, 1394–1399.
Ramasawmy, R., Faé, K.C., Cunha-Neto, E., Müller, N.G., Cavalcanti, V.L., Ferreira, R.C.,Drigo, S.A., Ianni, B., Mady, C., Goldberg, A.C., Kalil, J., 2007. Polymorphisms inthe gene for lymphotoxin-predispose to chronic Chagas cardiomyopathy. J.Infect. Dis. 196, 1837–1843.
Reis, D.D., Jones, E.M., Tostes Jr., S., Lopes, E.R., Gazzinelli, G., Colley, D.G., McCurley,T.L., 1993. Characterization of inflammatory infiltrates in chronic chagasicmyocardial lesions: presence of tumor necrosis factor-alpha+ cells anddominance of granzyme A+, CD8+ lymphocytes. Am. J. Trop. Med. Hyg. 48,637–644.
Simpson, L., Neckelmann, N., de la Cruz, V.F., Simpson, A.M., Feagin, J.E., Jasmer, D.P.,Stuart, J.E., 1987. Comparison of the maxicircle (mitochondrial) genomes ofLeishmania tarentolae and Trypanosoma brucei at the level of nucleotidesequence. J. Biol. Chem. 262, 6182–6196.

J.C. Carranza et al. / International Journal for Parasitology 39 (2009) 963–973 973
Solari, A., Campillay, R., Ortíz, S., Wallace, A., 2001. Identification of Trypanosomacruzi genotypes circulating in Chilean chagasic patients. Exp. Parasitol. 97, 226–233.
Souto, R.P., Fernandes, O., Macedo, A.M., Campbell, D.A., Zingales, B., 1996. DNAmarkers define two major phylogenetic lineages of Trypanosoma cruzi. Mol.Biochem. Parasitol. 83, 141–152.
Souto, R.P., Zingales, B., 1993. Sensitive detection and strain classification ofTrypanosoma cruzi by amplification of a ribosomal RNA sequence. Mol. Biochem.Parasitol. 62, 45–52.
Talvani, A., Rocha, M.O., Barcelos, L.S., Gomes, Y.M., Ribeiro, A.L., Teixeira, M.M.,2004. Elevated concentrations of CCL2 and tumor necrosis factor-alpha inchagasic cardiomyopathy. Clin. Infect. Dis. 38, 943–950.
Tibayrenc, M., Ayala, F.J., 2002. The clonal theory of parasitic protozoa: 12 years on.Trends Parasitol. 18, 405–410.
Tibayrenc, M., Ayala, F.J., 1988. Isoenzyme variability in Trypanosoma cruzi, theagent of Chagas disease: genetical, taxonomic and epidemiological significance.Evolution 42, 277–292.
Vago, A.R., Andrade, L.O., Leite, A.A., d’Avila Reis, D., Macedo, A.M., Adad, S.J., TostesJr., S., Moreira, M.C., Filho, G.B., Pena, S.D.J., 2000. Genetic characterization ofTrypanosoma cruzi directly from tissues of patients with chronic Chagas disease:differential distribution of genetic types into diverse organs. Am. J. Pathol. 156,1805–1809.
Valadares, H.M., Pimenta, J.R., de Freitas, J.M., Duffy, T., Bartholomeu, D.C., Oliveira,R de P., Chiari, E., Moreira, Mda C., Filho, G.B., Schijman, A.G., Franco, G.R.,Machado, C.R., Pena, S.D., Macedo, A.M., 2008. Genetic profiling of Trypanosomacruzi directly in infected tissues using nested PCR of polymorphicmicrosatellites. Int. J. Parasitol. 38, 839–850.
Westenberger, S.J., Barnabé, C., Campbell, D.A., Sturm, N.R., 2005. Two hybridizationevents define the population structure of Trypanosoma cruzi. Genetics 171, 527–543.
Westenberger, S.J., Cerqueira, G.C., El-Sayed, N.M., Zingales, B., Campbell, D.A.,Sturm, N.R., 2006. Trypanosoma cruzi mitochondrial maxicircles display species-and strain-specific variation and a conserved element in the non-coding region.BMC Genomics 7, 60.
Zafra, G., Mantilla, J.C., Valadares, H.M., Macedo, A.M., Gonzáles, C.I., 2008. Evidenceof Trypanosoma cruzi II infection in Colombian chagasic patients. Parasitol. Res.103, 731–734.
Zicker, F., Netto, J.C., Zicker, E.M., Oliveira, R.M., Smith, P.G., 1990. Trypanosoma cruziinfection and electrocardiographic findings among active manual workers: apopulation-based study in central Brazil. Int. J. Epidemiol. 19, 182–186.
Zingales, B., Souto, R.P., Mangia, R.H., Lisboa, C.V., Campbell, D.A., Coura, J.R., Jansen,A., Fernandes, O., 1998. Molecular epidemiology of American trypanosomiasisin Brazil based on dimorphisms of rRNA and mini-exon gene sequences. Int. J.Parasitol. 28, 105–112.
Copyright © 2022 FDOKUMEN




















