
Plasma membrane vesicles isolated from epimastigote forms of trypanosoma cruzi
Upload
independentCategory
view
1download
0
Experimental Parasitology 125 (2010) 256–263
Contents lists available at ScienceDirect
Experimental Parasitology
journal homepage: www.elsevier .com/locate /yexpr
Detection of matrix metallopeptidase-9-like proteins in Trypanosoma cruzi
Ana Cristina Nogueira de Melo a, Edilma Paraguai de Souza a, Camila Guarany Ramalho Elias a,André Luis Souza dos Santos a, Marta Helena Branquinha a, Cláudia Masini d’Ávila-Levy b,Flávia Coelho Garcia dos Reis d, Tatiana Ferreira Rocha Costa d, Ana Paula Cabral de Araujo Lima d,Mirian Cláudia de Souza Pereira c, Maria Nazareth Leal Meirelles c, Alane Beatriz Vermelho a,*
a Departamento de Microbiologia Geral, Instituto de Microbiologia Prof. Paulo de Góes (IMPPG), Centro de Ciências da Saúde (CCS), Bloco I,Universidade Federal do Rio de Janeiro (UFRJ), Ilha do Fundão, 21941-902 Rio de Janeiro, RJ, Brazilb Departamento de Bioquímica e Biologia Molecular, Instituto Oswaldo Cruz, FIOCRUZ, AvenidaBrasil 4365, 21045-900 Rio de Janeiro, RJ, Brazilc Departamento de Ultraestrutura e Biologia Celular, Instituto Oswaldo Cruz, FIOCRUZ, AvenidaBrasil 4365, 21045-900 Rio de Janeiro, RJ, Brazild Instituto de Biofísica Carlos Chagas Filho, Centro de Ciências da Saúde (CCS), Bloco D- sala 10B, Universidade Federal do Rio de Janeiro (UFRJ), Ilha do Fundão,21941-590 Rio de Janeiro, RJ, Brazil
a r t i c l e i n f o
Article history:Received 9 September 2008Received in revised form 24 December 2009Accepted 27 January 2010Available online 6 February 2010
Keywords:Trypanosoma cruziPeptidaseMatrix metallopeptidase-9Doxycycline
0014-4894/$ - see front matter � 2010 Elsevier Inc. Adoi:10.1016/j.exppara.2010.01.023
* Corresponding author. Address: Laboratório de PDepartamento de Microbiologia Geral, Instituto deGóes (IMPPG), Centro de Ciências da Saúde (CCS), BloRio de Janeiro (UFRJ), Avenida Carlos Chagas Filho21941-902 Rio de Janeiro, RJ, Brazil. Fax: +55 21 2560
E-mail address: [email protected] (A.B. Ve1 Index descriptions and abbreviations: Trypanosoma cru
gelatin–SDS–PAGE; immunoprecipitation; MMP-9, maPAGE, sodium dodecyl sulfate–polyacrylamide gel elRoitman’s complex medium; MMP, matrix metallopeptitrans-epoxysuccinyl-L-leucylamido-(4-guanidino) butancell sorting; ECM, the extracellular matrix.
a b s t r a c t
In this study, the cell-associated and extracellular peptidases of Trypanosoma cruzi grown in modifiedRoitman’s complex (MRC) medium were analyzed by measuring peptidase activity in gelatin-containingzymograms. Our results showed that the cell-associated peptidases as well as peptidases extracellularlyreleased by T. cruzi displayed two distinct proteolytic classes: cysteine and metallopeptidase activities.The major cysteine peptidase, cruzipain, synthesized by T. cruzi cells was detected in cellular parasitecontent, as a 50 kDa reactive polypeptide, after probing with anti-cruzipain antibody. In addition,metallo-type peptidases belonging to the matrix metallopeptidase-9 (MMP-9) family were revealed, afterWestern blotting, as a 97 kDa protein band in cellular extract and an 85 kDa polypeptide in both cellularand secreted parasite extracts. The MMP-9-like activity present in cells and spent culture medium wasimmunoprecipitated by an anti-MMP-9 polyclonal antibody. The surface location of MMP-9-like proteinsin T. cruzi was also evidenced by means of flow cytometry analysis. Furthermore, doxycycline that hasdirect MMP-9 inhibiting properties in vitro, inhibited MMP-9-like activities in gel zymography, immuno-precipitation and flow cytometry analyses. This is the first report of the presence of MMP-9-likemolecules in T. cruzi. The presence of a matrix extracellular-degrading enzyme may play a role in theT. cruzi-host cell interaction, making this enzyme a potential target for future drug development againstthis pathogenic trypanosomatid.
� 2010 Elsevier Inc. All rights reserved.
1. Introduction ical disease that affects over 11 million people and causes an
The flagellated protozoan Trypanosoma cruzi1 is the causativeagent of Chagas’ disease, a pathology characterized by chronicinflammation associated with cardiomyopathy and/or digestive dis-orders (Engman and Leon, 2002). Chagas’ disease is a neglected trop-
ll rights reserved.
roteases de Microrganismos,Microbiologia Prof. Paulo deco I, Universidade Federal do, 373, Cidade Universitária,8344.
rmelho).zi; matrix metallopeptidase-9;
trix metallopeptidase-9; SDS–ectrophoresis; MRC, modifieddase; DTT, dithiothreitol; E-64,e; FACS, fluorescence activated
estimated 50,000 deaths annually in Latin America (WHO, 2006).Most human infections occur through contact with the infectedbloodsucking triatomine species. However, the urbanization processin Latin America and immigration trends have led to the diseasebeing diagnosed in non-endemic areas where, even in the absenceof the vector; the infection can still be transmitted congenitally, byblood transfusion and by organ transplantation (Bern et al., 2007).In addition, oral transmission has aroused the attention of research-ers, due to outbreak in some particular areas such as the BrazilianAmazon, mainly associated with the consumption of Amazonianpalm berry or açaí (Euterpe oleracea Mart.) juice (Yoshida, 2009).
Peptidases produced by the pathogenic trypanosomatids playan important role in several steps of the host infection including:adsorption, penetration, intracellular survival, replication, differen-tiation, infectivity, immune evasion and nutrition (Vermelho et al.,2007). Trypanosomatids elaborate a large array of peptidases,which are intracellular and/or extracellular, with different specific-

A.C. Nogueira de Melo et al. / Experimental Parasitology 125 (2010) 256–263 257
ities and many of them have been purified and characterized andtheir genes cloned and sequenced. Because of their unusual struc-tural features, parasite peptidases are considered to have a highpotential as targets for novel antiparasitic agents (Brack et al.,2008). The immunodominant nature of many peptidases offerspotential for serodiagnosis and vaccine development.
Trypanosoma cruzi has been shown to contain several proteolyticactivities (Vermelho et al., 2007). Cruzipain, a cysteine peptidase ofthe papain family, is the most prominent proteolytic activity synthe-sized by all developmental stages of T. cruzi. Cruzipain is also aproposed therapeutic target for treatment of Chagas’ disease (Cazz-ulo, 2002). Previous studies have demonstrated that T. cruzi infectioncan be cured in cell, mouse and dog models by treatment with irre-versible inhibitors of cruzipain (Engel et al., 1998; Barr et al., 2005).In addition to cruzipain, other cysteine peptidases identified in theparasite include a 30-kDa cathepsin B-like peptidase which exhib-ited an important hydrolytic activity on human type I collagen(Garcia et al., 1998) and TcCPmet secreted from metacyclic trypom-astigotes during metacyclogenesis (Duschak et al., 2006). Two serinepeptidases belonging to the prolyl oligopeptidase family have alsobeen characterized in T. cruzi: the parasite calcium-signaling activityis linked to the 120-kDa cytosolic serine peptidase, named Tc-OP,which generates an active calcium agonist from a cytosolic precursormolecule (Burleigh and Woolsey, 2002) and the 80-kDa serine prolyloligopeptidase (POP Tc80) facilitates T. cruzi dissemination through-out host tissues due to its ability to mediate the cleavage of someextracellular matrix components (Bastos et al., 2005). The expres-sion of metallopeptidase activities have been previously describedin various strains and clones of T. cruzi, some of them specificallyobserved during metacyclogenesis (Bonaldo et al., 1991; Lowndeset al., 1996). Moreover, Cuevas et al. (2003) described that T. cruzipossesses a family of gp63 genes composed of multiple groups.Two of the groups, Tcgp63-I and Tcgp63-II, are present as high-copy-number genes and antibodies against Tcgp63-I partiallyblocked the infection of Vero cells by trypomastigotes, which sug-gests a possible role for this metallopeptidase during the infectionprocess in vitro. Recently, Pinho et al. (2009) described for the firsttime the isolation of two aspartic peptidases from T. cruzi, whichhave been named CPZ-I and CPZ-II.
Extracellular matrix (ECM)-eukaryotic cell interactions areregulated by a family of zinc-dependent peptidases known asmatrix metallopeptidases (MMPs) that can be classified broadlyby substrate specificity (Van den Steen et al., 2002). Throughturnover of extracellular matrix proteins, MMPs are involvedeither in normal matrix remodeling or pathological tissuedestruction (Gomis-Rüth, 2009). Among the members of theMMP family, the gelatinases MMP-2 (gelatinase A) and MMP-9(gelatinase B) are known to be key players in many physiologicaland pathological processes. In this context, earlier studies fromour group showed matrix metallopeptidase-9 (MMP-9)-likeactivity in the cytoplasm of T. cruzi during in vitro infection ofembryonic hepatocyte cells (Nogueira de Melo et al., 2004). Pep-tidase-dependent extracellular matrix remodeling is one of theevents that is emerging as a key regulator of T. cruzi infectionand pathogenesis of Chagas’ disease. Here in this study we lookat the detection of MMP-9-like proteins in cells and spent cul-ture medium of T. cruzi Y strain and discuss the possible impli-cations of this enzyme in the pathology of Chagas’ disease.
2. Materials and methods
2.1. Chemicals
Media constituents, reagents used in electrophoresis, buffer com-ponents and the nitrocellulose membrane were obtained fromAmersham Life Science (Little Chalfont, England). The metallo
(1,10-phenanthroline) and the cysteine (trans-epoxysuccinyl-L-leucylamido-(4-guanidino) butane [E-64]) peptidase inhibitors,dithiothreitol (DTT) and secondary antibody (horseradish peroxi-dase-conjugated goat anti-rabbit Fc) were purchased from SigmaChemical Co. (St. Louis, MO, USA). Rat IgG2b j isotype and anti-MMP-9 polyclonal antibody were purchased from BD Pharmigenand Santa Cruz (Biotechnology), respectively. All other reagentswere analytical grade.
2.2. Parasite and growth conditions
T. cruzi Y strain is part of our trypanosomatid culture collectionand it was cultivated in modified Roitman’s complex (MRC) med-ium (0.5% yeast extract, 0.5% peptone, 2% KCl, 2% sucrose and2 mg% hemin) supplemented with 10% (v/v) heat-inactivated fetalbovine serum (FBS) for 6 days at 28 �C. Cellular viability wasassessed by motility, trypan blue cell dye exclusion and theabsence of lactate dehydrogenase, an intracellular enzyme, in thecell-free culture supernatants (d’Avila-Levy et al., 2003).
Bloodstream trypomastigote forms of T. cruzi (Y strain) weremaintained cyclically in Swiss mice and they were obtained atthe peak of parasitemia (7 days post-infection) as previouslydescribed (Meirelles et al., 1984). Briefly, immediately after eutha-nasia intra-cardiac function was made for blood collection. Theblood samples were centrifuged at 1000 rpm for 20 min and theplasma containing trypomastigote forms of T. cruzi was removed.The parasites were seeded by centrifugation (3500 rpm for15 min) and resuspended in Dulbecco’s modified Eagle medium(DMEM). The total number of parasites/ml was determined byquantification in a Neubauer chamber. Intracellular amastigoteswere obtained after the infection of Vero cell monolayers withtrypomastigote forms of T. cruzi, Y strain. The highly infected cellsruptured releasing amastigote forms of T. cruzi after 5–7 days ofinfection. The parasites were harvested, centrifuged and resus-pended in DMEM (Souto-Padrón et al., 1990).
2.3. Parasite extracts
Six-day-old cultured epimastigotes, at the log growth phase,were harvested by centrifugation (1500g, 15 min, 4 �C), washedthree times with phosphate-buffered saline (PBS; 150 mM NaCl,20 mM phosphate buffer, pH 7.2), then 1 � 107 cells were lysedat 0 �C by the addition of 100 lL of sodium dodecyl sulfate–poly-acrylamide gel electrophoresis (SDS–PAGE) sample buffer(125 mM Tris, pH 6.8, 4% SDS, 20% glycerol and 0.002% bromophe-nol blue). Amastigote and trypomastigote cells (1 � 107 cells) werealso washed three times with phosphate-buffered saline and lysedas described above. The spent culture medium was passed over a0.22-lm filtration unit and concentrated 30-fold in a 10,000-molecular-weight cut-off Centricon micropartition system (AMI-CON, Beverly, MA). The concentrated culture supernatant wasmixed with SDS–PAGE sample buffer in a sample:buffer ratio of8:2. Protein concentration was determined by the method of Lowryet al. (1951), using BSA as a standard.
2.4. Analysis of peptidase activity
The cellular and extracellular proteolytic activities were evalu-ated on 10% SDS–PAGE with 0.1% gelatin incorporated as substrate(Heussen and Dowdle, 1980). Briefly, the gels were loaded with10 lL of parasite extract (equivalent to 1 � 107 cells) and 40 lLof concentrated culture supernatant (equivalent to 100 lg of pro-tein) per slot and after electrophoresis at a constant voltage of170 V for 2 h at 4 �C, they were soaked for 1 h in 2.5% Triton X-100 under constant agitation. The gel strips containing the cellularand released peptidases were then incubated for 48 h at 37 �C in

258 A.C. Nogueira de Melo et al. / Experimental Parasitology 125 (2010) 256–263
the following buffers: 50 mM sodium phosphate (pH 5.5–7.4) inthe presence or absence of 2 mM DTT (a cysteine-peptidase activa-tor) and 50 mM glycine–NaOH, pH 10.0. The effect on cellular andextracellular proteolytic activities of peptidase inhibitors—10 lME-64, 10 mM 1,10-phenanthroline and 10 mM doxycycline—wastested by including them in the same buffers described previouslyfor 48 h at 37 �C. In order to show specific inhibition due to theaddition of 10 lM E-64, DTT was added to the samples at pH5.5 + E-64. The gels were stained for 1 h with 0.2% Coomassie bril-liant blue R-250 in methanol–acetic acid–water (50:10:40) anddestained in the same solvent. The gels were dried, scanned anddigitally processed. The apparent molecular masses of the cellularand extracellular T. cruzi peptidases were calculated by comparisonwith the mobility of Gibco BRL SDS–PAGE standards.
2.5. Western blotting analysis
Total cellular protein extracts (equivalent to 1 � 107 cells) andthe cell-free concentrated culture supernatants (equivalent to100 lg of protein) were separated in 10% SDS–PAGE under reducingconditions and the polypeptides were electrophoretically trans-ferred at 4 �C at 100 V/300 mA for 2 h to nitrocellulose membranes.Each membrane was blocked in 10% low-fat dried milk in PBScontaining 0.05% Tween 20 (PBS/Tween) for 1 h at roomtemperature. Then, membranes were washed three times (10 mineach) with the blocking solution and incubated for 1 h at room tem-perature with the following primary antibodies at 1:500 dilution:rabbit anti-MMP-9 polyclonal antibody, anti-cruzipain monoclonalantibody JO1 raised against purified cruzipain from epimastigotesof T. cruzi Dm28c and anti-Dm-calpain raised against Drosophilamelanogaster calpain. In addition, the cellular and spent culturemedium extracts of T. cruzi was assayed with a polyclonal antibodyraised against gp63 Leishmania amazonensis in order to investigatewhether the metallopeptidases are related to the leishmanial gp63metallopeptidase or related to the MMP-9. The secondary antibodyused was horseradish peroxidase-conjugated goat anti-rabbit IgGat 1:2500, followed by chemiluminescence immunodetection (ECLreagent) (Nogueira de Melo et al., 2004; Elias et al., 2006).
2.6. Immunoprecipitation
Fifty microliters of protein-A agarose beads (Sigma) werewashed three times with PBS and incubated overnight in the samebuffer containing rabbit anti-MMP-9 polyclonal antibody or ratIgG2b j isotype control. The beads were washed three times with1 mL of PBS and incubated for 2 h with 600 lL of cell or spent cul-ture medium extracts, obtained as described above. Protein-A aga-rose was washed three times with PBS and subsequently eluted in0.1 M glycine HCl, pH 3.0. All the fractions were collected andanalyzed on 10% SDS–PAGE with 0.1% gelatin incorporated as sub-strate as described previously (Aparício et al., 2004).
2.7. Flow cytometry for cell surface MMP-9 detection
Epimastigotes (1 � 107 cells) used for these experiments werefixed at 4 �C in 0.4% paraformaldehyde diluted in PBS for 30 min,followed by extensive washing in the same buffer. The fixed cellsmaintained their morphological integrity, as verified by opticalmicroscopic observation. Cells were incubated for 1 h at room tem-perature with a 1:500 dilution of anti-MMP-9 polyclonal antibodyand then incubated for an additional hour with a 1:250 dilution offluorescein isothiocyanate (FITC)-labeled goat anti-rabbit IgG.Control cells were subjected to the same experimental conditions,except for the absence of anti-MMP-9 polyclonal antibody.Alternatively, epimastigotes (1 � 107 cells) were incubated with10 mM doxycycline for 24 h at 28 �C. Thereafter, cells were fixed
and processed for flow cytometry analysis as described above,employing a 1:500 dilution of anti-MMP-9 polyclonal antibody.The cells were washed three times in PBS and were analyzed byflow cytometry in FACSCalibur (BD Bioscience, USA) equipped witha 15 mW argon laser emitting at 488 nm. The mapped population(n = 10,000) was then analyzed for log green fluorescence by usinga single-parameter histogram (Elias et al., 2006).
2.8. Statistical analysis
All experiments were performed in triplicate, in 3 independentexperimental sets. The data were analyzed statistically by means ofthe ANOVA test using EPI-INFO 6.04 (Database and Statistics Pro-gram for Public Health) computer software. P values of 0.05 wereconsidered statistically significant.
3. Results
3.1. Zymographic analysis of the cell-associated and extracellularpeptidases from T. cruzi
The cell-associated and extracellular peptidases of T. cruzigrown in MRC medium were analyzed by measuring peptidaseactivity on gelatin in zymograms and they displayed two distinctproteolytic classes: cysteine- and metallo-type peptidase activities.Cysteine peptidases were detected at both pH 5.5 and 7.4 supple-mented with DTT, which is a well-known cysteine-peptidaseactivator and was totally inhibited by E-64, a powerful cysteinepeptidase inhibitor (Fig. 1A and B); meanwhile, metallopeptidaseswere active at all pHs tested and were blocked by 1,10-phenan-throline (a zinc-metallopeptidase inhibitor) (Fig. 1A and B). Theaddition of the doxycycline that has direct MMP-9 inhibiting prop-erties in vitro, inhibited metallopeptidases migrating at 110 (in thespent culture medium) and 97 kDa (in the cellular extract) (Fig. 1 Aand B). Moreover, DTT had no effect at pH 10 without metallopep-tidase inhibitor.
The cellular proteolytic profile comprises a 50 kDa cysteine pep-tidase and also a metallopeptidase with molecular mass of 97 kDa(Fig. 1A). The analyzed spent culture medium showed a morecomplex proteolytic profile displaying a cysteine peptidase of100 kDa and three metallopeptidases migrating at 110, 70 and 55(Fig. 1B). It is interesting to note that the presence of DTT inhibitedthe 55 kDa metallopeptidase, even though this proteolytic activitywas active at pH 5.5 or 7.4 supplemented with E-64. In order toinvestigate whether the 55 kDa peptidase may be inhibited byDTT, we assayed T. cruzi extract in pH 5.5 or 7.4 without DTT.Under this condition, the extracellular metallopeptidase of55 kDa was detected, suggesting the inhibitory effect of DTT(Fig. 1B). Previous work showed that metallopeptidase activitiescould be inhibited by DTT, which binds to disulfide bonds impor-tant for its activity (Xu et al., 2006). Additionally, our group dem-onstrated that some metallopeptidase activities were alsoinhibited in the presence of DTT in the supernatant of non-infectedand infected Leishmania chagasi-hepatocyte-macrophage co-cul-ture (Costa et al., 2008). The cellular viability was assessed by mon-itoring the mobility of cells and trypan blue cell dye exclusion.Also, lactate dehydrogenase, an intracellular enzyme, was notfound in the supernatant fluids, indicating that the extracellularpeptidase detected in this study was not released by autolysis(data not shown).
3.2. Immunoblotting of cellular and cell-free protein extracts
Our results also showed that T. cruzi grown on MRC displayed acellular cysteine peptidase with an apparent molecular mass of

Fig. 1. Zymogram analysis of Trypanosoma cryzi cellular and extracellular peptidases. Parasite cells and spent culture medium were analyzed through gelatin–SDS–PAGE. Gelstrips containing extracts from 1 � 107 parasites were incubated at 37 �C for 48 h in 50 mM sodium phosphate buffer, pH 5.5 or pH 7.4, supplemented or not with 2 mM DTTin the absence or in the presence of 10 lM E-64. Another set of gelatin-gel strips was incubated in 50 mM glycine–NaOH buffer, pH 10, in the absence or in the presence of2 mM DTT, 10 mM 1,10-phenanthroline (Phe) or doxycycline (Dox). The approximate molecular mass of the peptidases, expressed in kDa, are shown on the left.
A.C. Nogueira de Melo et al. / Experimental Parasitology 125 (2010) 256–263 259
50 kDa. Probably, this proteolytic activity corresponds to the para-site’s cysteine peptidase named cruzipain, which migrates ingelatin–SDS–PAGE in this same molecular mass range. Supportingour idea, Western blotting analysis evidenced a faint polypeptideband with an apparent molecular mass of 50 kDa in the T. cruzicellular extracts after probing with anti-cruzipain monoclonalantibody JO1 (Fig. 2, lane a). Interestingly, in zymograms an extra-cellular cysteine peptidase with an apparent molecular mass of100 kDa was detected at pH 5.5 supplemented with DTT. The spentculture medium extract was analyzed by Western blotting usingthe same antibody described above. Our results showed no immu-no-cross-reactivity with this antibody, suggesting that thisextracellular cysteine peptidase is not a cruzipain homologue
Fig. 2. Western blotting showing the cruzipain-like (a and b), calpain-like (c and d)and MMP-9-like (e and f) polypeptides detected in the cellular extract (a, c, e) and inthe spent culture medium (b, d and f) from Trypanosoma cruzi grown in MRC for6 days. The cellular (lane g) and spent culture medium (lane h) extracts ofTrypanosoma cruzi were assayed with a polyclonal antibody raised against gp63Leishmania amazonensis in order to investigate whether the metallopeptidases arerelated to the leishmanial gp63 metallopeptidase or related to the MMP-9. Theapproximate molecular mass of the reactive polypeptides, expressed in kDa, areshown on the left.
(Fig. 2, lane b). Moreover, this enzyme is not an aggregate of cruzi-pain molecules since no cross-reactivity was observed either underSDS–PAGE conditions or the conditions used for gelatin-containinggels (not boiled and without reduction) (data not shown).
The presence of large numbers of calpain-related proteins(CALPs) in trypanosomatids has already been revealed, althoughthere are no data as yet on the specific functions of the calpain-likeproteins in these microorganisms (Ersfeld et al., 2005). Our grouphas demonstrated the presence of an 80-kDa reactive protein againstD. melanogaster calpain in the T. cruzi clone Dm28c epimastigoteform (Sangenito et al., 2009). Furthermore, different levels of CALPexpressions were also detected by flow cytometry and fluorescencemicroscopy analyses in members of distinct T. cruzi lineages: TCI (Ystrain), TCII (clone Dm28c) and Z3 zymodeme (INPA6147 strain)with higher levels (twice as many) found for the Y strain comparedto Dm28c and INPA6147 strains. Corroborating these findings, ourwork has detected a polypeptide band migrating at approximately80 kDa in cell and spent culture medium extracts after probing withD. melanogaster calpain (anti-Dm-calpain) (Fig. 2, lanes c and d).
Although Western blotting analysis using polyclonal antibodiescannot prove the protein homology with absolute certainty, thismethodology can suggest that the proteins may share some struc-tural and biochemical properties. In this sense, in order to reveal ifT. cruzi possesses proteins related to the MMP family, cellular andsecretory extracts from MRC medium were probed with the anti-MMP-9 polyclonal antibody. The anti-MMP-9 reacted with proteinbands at 97 (in the cellular extract) and 85 kDa (in both cellularand spent culture medium extracts) (Fig. 2, lanes e and f). However,no proteolytic activity band with apparent molecular mass of85 kDa was observed in the zymogram. Activated forms of MMP-9 (correspond to 85 kDa) are usually difficult to observe onzymograms, because they are highly unstable and subject to rapidturnover (Malla et al., 2008).

260 A.C. Nogueira de Melo et al. / Experimental Parasitology 125 (2010) 256–263
Cuevas et al. (2003) described that gp63 homologues in T. cruziprobably play a role in host cell-interaction. In this sense, a poly-clonal antibody raised against gp63 L. amazonensis, was assayedin order to investigate whether the metallopeptidases are relatedto the leishmanial gp63 metallopeptidase or related to theMMP-9. Our results showed that this antibody recognized onepolypeptide of 73 kDa in the cell extracts (Fig. 2, lane g). In thespent culture medium extract we were unable to detect gp63homologues (Fig. 2A, lane h). We cannot rule out the possibilitythat gp63 homologues are present in this extract, but, if present,they were so low that they were not detected under the conditionsassayed. Taken together, these data suggest that the polypeptidesof 97 and 85 kDa are not gp63, nor a related protein, but theymight correspond to MMP-9 activity.
3.3. Immunoprecipitation and flow cytometry for MMP-9-like proteindetection
Immunoprecipitation from both T. cruzi epimastigote cells andspent culture medium using the anti-MMP-9 polyclonal antibodywas carried out in order to confirm the presence of MMP-9-likeproteins in T. cruzi extracts. The immune complexes were purifiedand the proteolytic activities were examined for peptidase activityon zymogram gels with gelatin as substrate. Solely, one band ofpeptidase activity migrating approximately at 97 kDa was immu-noprecipitated by the anti-MMP-9 polyclonal antibody either cellsor spent culture medium (Fig. 3A, lanes c and g). As previouslyobserved on zymogram gels, doxycycline powerfully inhibitedthe enzymes, suggesting that these peptidases are MMP-9-likeproteins (Fig. 3A, lanes d and h). The metallopeptidase activitiespresent in both samples was not immunoprecipitated by controlIgG (Fig. 3A, lane l).
As demonstrated by flow cytometry analysis, anti-MMP-9 poly-clonal antibody was found to react with the cell surface of T. cruzi(Fig. 3B). We also aimed to assess whether the doxycycline-treatedcells of T. cruzi presented any change in levels of expression ofMMP-9-like proteins in comparison to non-treated cells. Flowcytometry analyses revealed that the expression of MMP-9-likeprotein was reduced when parasites were subjected to the pres-ence of the MMP inhibitor doxycycline (Fig. 3B and C). Althoughour results present a high standard deviation, representative dataof the analysis of 10,000 cells shown makes our results statisticallysignificant (P < 0.05) (Fig. 3C).
3.4. Detection of MMP-9-like proteins in T. cruzi amastigote andtrypomastigote forms
In order to observe the presence of MMP-9-like proteins in T. cruzievolutive forms present in the mammalian host, blood-stagetrypomastigotes and intracellular amastigotes were also investi-gated in this study. The zymogram gels showed that both evolutiveforms present a cell-associated peptidase migrating at 50 kDa(Fig. 4A). This proteolytic activity was active at pH 5.5 in thepresence of the reducing agent DTT but was completely abolishedby E-64 (Fig. 4). In addition, Western blotting analysis reacted witha single band around 50 kDa after probing with anti-cruzipainmonoclonal antibody JO1. These data are in accordance with the pre-viously described data for T. cruzi that cruzipain is found in all devel-opmental forms of T. cruzi (Fig. 4B, lanes a and c). Moreover, weobserved a metallopeptidase migrating at 97 kDa in both amastigoteand blood-stage trypomastigote forms that was only detected inalkaline conditions and was totally inhibited by 10 mM 1,10-phe-nanthroline and doxycycline (Fig. 4A). Using Western blotting withanti-MMP-9 polyclonal antibody, a protein band with a molecularmass of 97 kDa was recognized in both forms (Fig. 4, lanes b and d).
4. Discussion
In the present work, cellular and extracellular peptidases werecharacterized in T. cruzi. Our results showed the presence of cysteinepeptidases and MMP-9-like proteins in T. cruzi epimastigotes.Furthermore, we also presented evidences of MMP-9-like proteinsboth in intracellular amastigotes or blood-stage trypomastigotesforms.
Three cysteine peptidases with molecular masses range from116.1 to 101.3 have been identified in the T. cruzi genome. Interest-ingly, we have demonstrated an extracellular cysteine peptidasefrom epimastigotes with a molecular mass of 100 kDa and it wasnot recognized by anti-cruzipain monoclonal antibody JO1. Theseresults are similar to those observed by Duschak et al. (2006)who described a cysteine peptidase with molecular mass between97 and 116 kDa released into the culture medium during T. cruzimetacyclogenesis that also did not react with anti-cruzipain anti-body. The liberation of 100 kDa cysteine peptidase to the extracel-lular environment might be an important adaptive mechanismduring the life cycle of T. cruzi. The appearance of this proteolyticactivity in epimastigotes could assist the hydrolysis of a varietyof foreign proteins supplying essential amino acids and peptidesas nutrients to the flagellate cells in order to support their initialdevelopment, multiplication and differentiation within the vectormidgut. Furthermore, this enzyme might play a major role in theT. cruzi nutritional stress response that precedes metacyclogenesis.
Calpains constitute a family of calcium-dependent cytosoliccysteine peptidases that have been characterized mainly inhumans, but their role remains poorly understood, although theymay participate in a variety of cellular processes, including therearrangement of cytoskeletal proteins, different signal transduc-tion pathways and apoptosis (Goll et al., 2003).
A gene encoding the T. cruzi calpain (CALP), TcCALPx11 (GeneDBIDTc00.1047053506563.210) was identified by microarray analysis asbeing differentially expressed by T. cruzi during metacyclogenesis(Giese et al., 2008). Its expression during nutritional stress precedingT. cruzi metacyclogenesis was 2.53 times higher than that observedin epimastigotes. Antiserum against a recombinant TcCALPx11-GST-fusion protein reacted with a protein band at 80 kDa in T. cruziepimastigote protein extracts by Western blot analysis. Westernblotting with parasite extracts during metacyclogenesis showed anincrease in the abundance of TcCALPx11 in parasites undernutritional stress. Recently, a calpain inhibitor III (MDL28170) wasused to detect calpain-like molecules in the epimastigote forms ofthe T. cruzi isolate Dm28c and the inhibitor at 70 mM promoted astrong reduction in the growth rate after 48 h (Sangenito et al.,2009). In this study, we also detected a polypeptide band with anapparent molecular mass of 80 kDa both in cells and spent culturemedium from T. cruzi strain Y by Western blotting analysis. No pro-teolytic activity was observed for this 80 kDa protein in our work, inaccordance to previously demonstrated studies (Giese et al., 2008;Sangenito et al., 2009).
Members of the superfamily of metallopeptidases participate inmetabolism through both extensive and unspecific proteindegradation and deregulation of their function leads to pathology.Moreover, these enzymes may also act as virulence factors duringmicrobial infection (Gomis-Rüth, 2009). Among themetallopeptidases is the metzincin clan that is subdivided intofamilies and one of them is the matrix metallopeptidase family.Matrix metallopeptidases are able to degrade all components ofextracellular matrix. In addition, these proteolytic enzymes areinvolved in regulation of the inflammatory response under variouscircumstances, including the direct cleavage of immune systemproteins (Cockle et al., 2007). As a result, it has become clear thatthese enzymes are crucial factors in the pathogenesis of
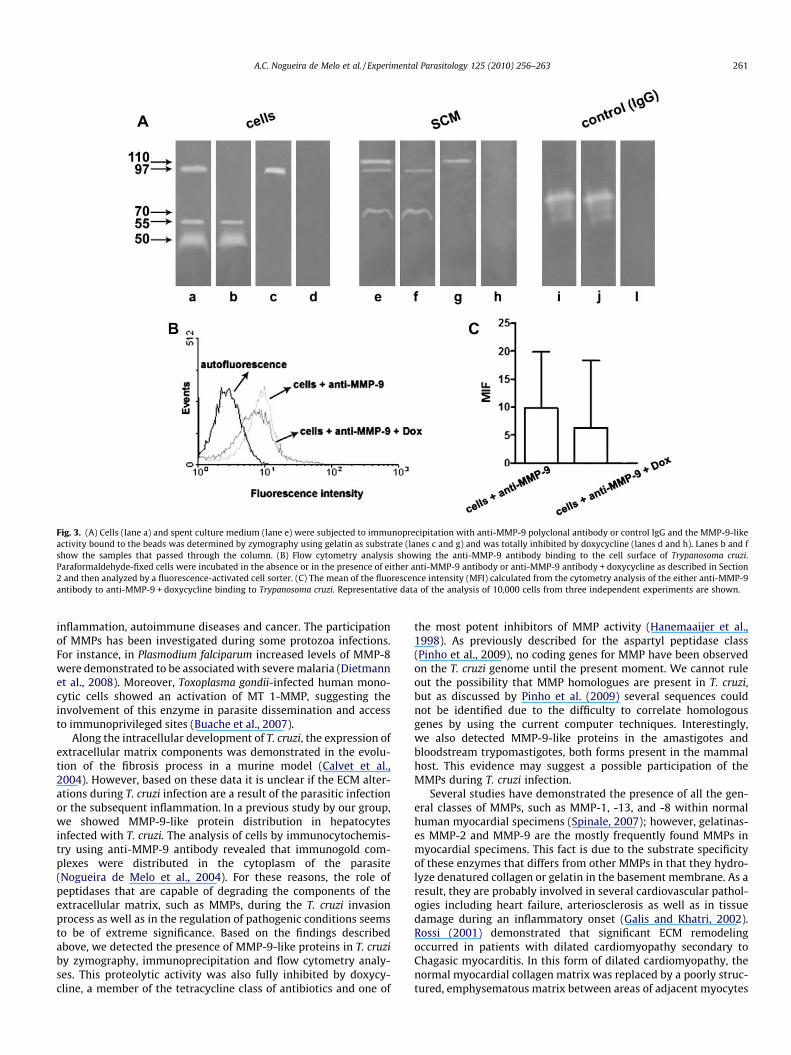
Fig. 3. (A) Cells (lane a) and spent culture medium (lane e) were subjected to immunoprecipitation with anti-MMP-9 polyclonal antibody or control IgG and the MMP-9-likeactivity bound to the beads was determined by zymography using gelatin as substrate (lanes c and g) and was totally inhibited by doxycycline (lanes d and h). Lanes b and fshow the samples that passed through the column. (B) Flow cytometry analysis showing the anti-MMP-9 antibody binding to the cell surface of Trypanosoma cruzi.Paraformaldehyde-fixed cells were incubated in the absence or in the presence of either anti-MMP-9 antibody or anti-MMP-9 antibody + doxycycline as described in Section2 and then analyzed by a fluorescence-activated cell sorter. (C) The mean of the fluorescence intensity (MFI) calculated from the cytometry analysis of the either anti-MMP-9antibody to anti-MMP-9 + doxycycline binding to Trypanosoma cruzi. Representative data of the analysis of 10,000 cells from three independent experiments are shown.
A.C. Nogueira de Melo et al. / Experimental Parasitology 125 (2010) 256–263 261
inflammation, autoimmune diseases and cancer. The participationof MMPs has been investigated during some protozoa infections.For instance, in Plasmodium falciparum increased levels of MMP-8were demonstrated to be associated with severe malaria (Dietmannet al., 2008). Moreover, Toxoplasma gondii-infected human mono-cytic cells showed an activation of MT 1-MMP, suggesting theinvolvement of this enzyme in parasite dissemination and accessto immunoprivileged sites (Buache et al., 2007).
Along the intracellular development of T. cruzi, the expression ofextracellular matrix components was demonstrated in the evolu-tion of the fibrosis process in a murine model (Calvet et al.,2004). However, based on these data it is unclear if the ECM alter-ations during T. cruzi infection are a result of the parasitic infectionor the subsequent inflammation. In a previous study by our group,we showed MMP-9-like protein distribution in hepatocytesinfected with T. cruzi. The analysis of cells by immunocytochemis-try using anti-MMP-9 antibody revealed that immunogold com-plexes were distributed in the cytoplasm of the parasite(Nogueira de Melo et al., 2004). For these reasons, the role ofpeptidases that are capable of degrading the components of theextracellular matrix, such as MMPs, during the T. cruzi invasionprocess as well as in the regulation of pathogenic conditions seemsto be of extreme significance. Based on the findings describedabove, we detected the presence of MMP-9-like proteins in T. cruziby zymography, immunoprecipitation and flow cytometry analy-ses. This proteolytic activity was also fully inhibited by doxycy-cline, a member of the tetracycline class of antibiotics and one of
the most potent inhibitors of MMP activity (Hanemaaijer et al.,1998). As previously described for the aspartyl peptidase class(Pinho et al., 2009), no coding genes for MMP have been observedon the T. cruzi genome until the present moment. We cannot ruleout the possibility that MMP homologues are present in T. cruzi,but as discussed by Pinho et al. (2009) several sequences couldnot be identified due to the difficulty to correlate homologousgenes by using the current computer techniques. Interestingly,we also detected MMP-9-like proteins in the amastigotes andbloodstream trypomastigotes, both forms present in the mammalhost. This evidence may suggest a possible participation of theMMPs during T. cruzi infection.
Several studies have demonstrated the presence of all the gen-eral classes of MMPs, such as MMP-1, -13, and -8 within normalhuman myocardial specimens (Spinale, 2007); however, gelatinas-es MMP-2 and MMP-9 are the mostly frequently found MMPs inmyocardial specimens. This fact is due to the substrate specificityof these enzymes that differs from other MMPs in that they hydro-lyze denatured collagen or gelatin in the basement membrane. As aresult, they are probably involved in several cardiovascular pathol-ogies including heart failure, arteriosclerosis as well as in tissuedamage during an inflammatory onset (Galis and Khatri, 2002).Rossi (2001) demonstrated that significant ECM remodelingoccurred in patients with dilated cardiomyopathy secondary toChagasic myocarditis. In this form of dilated cardiomyopathy, thenormal myocardial collagen matrix was replaced by a poorly struc-tured, emphysematous matrix between areas of adjacent myocytes

Fig. 4. (A) Zymogram of Trypanosoma cruzi evolutive forms. Gel strips containing extracts from 1 � 107 parasites were incubated at 37 �C for 48 h in 50 mM sodiumphosphate buffer, pH 5.5, supplemented with 2 mM DTT in the absence or in the presence of 10 lM E-64. Another set of gelatin-gel strips was incubated in 50 mM glycine-NaOH buffer, pH 10, in the absence or in the presence of 10 mM 1,10-phenanthroline (Phe). (B) Western blotting showing the cruzipain-like polypeptides in amastigotes (lanea) and bloodstream trypomastigotes (lane c) and MMP-9-like polypeptides in amastigotes (lane b) and bloodstream trypomastigotes (lane d) from Trypanosoma cruzi. Theapproximate molecular mass of the peptidases, expressed in kDa, are shown on the left.
262 A.C. Nogueira de Melo et al. / Experimental Parasitology 125 (2010) 256–263
and myocardial fascicles. Moreover, studies performed on theexplanted myocardial specimens from patients with Chagas’ dis-ease, have identified MMPs as the main ECM proteolytic pathwaylikely to contribute to the ECM remodeling process (Li et al.,2002; Rutschow et al., 2006; Spinale, 2007). It is also likely thatreceptor pathways, such as the bradykinin receptor system, playa role in transcriptional regulation of MMPs, causing a number ofintracellular events within the coronary vasculature and myocar-dial fibroblasts (Spinale, 2007). Scharfstein et al. (2000) establishedthat the participation of several secreted cruzipain isoforms inmammalian cell invasion is linked to a mechanism involvingkinin-releasing activity. One possible explanation for the severeChagasic myocarditis during Chagas’ disease might be the induc-tion of MMP-9 via kinin-releasing, mediated by T. cruzi secretedcruzipain. This mechanism might contribute to the ECM remodel-ing process, which is considered one of the ways to arrive at asevere heart situation.
Macrophages are a primary source of MMPs. Metallopeptidaseoverexpression by macrophages can be induced by pro-inflamma-tory stimuli including cytokines, such as TNF-a (Opdenakker et al.,2001). Costa et al. (2008) observed an increase in TGF-b productionin L. chagasi-hepatocyte-macrophage co-culture supernatants dur-ing the highest leishmanicidal activity by macrophages, coincidingwith high MMP-9 activity. This fact suggests that that this cytokinecould be participating in metallopeptidase activation/conversionand probably in matrix deposition by hepatocytes in culture.Earlier studies demonstrated that the activation of MMP-9 byinflammatory cells was essential to enhance mouse heart diseaseduring sepsis (Cuenca et al., 2006). Recently, Gutierrez et al.(2008) suggested an important role for MMPs in the induction ofT. cruzi-induced acute myocarditis, since mice treated with MMPinhibitor showed a significant reduction in both NO pathwaysand in the levels of INF-c and TNF-a that are Th1 cytokines nor-mally found in the myocardium of infected mice. Thus, influx of
inflammatory cells during T. cruzi infection could increase MMP-9 expression, contributing in the tissue damage of a chagasicpatient’s heart, characteristic of Chagasic myocarditis.
The cleavage of gelatin, a denatured form of collagen, by MMP-9activity from T. cruzi would create potential binding sites toenhance T. cruzi attachment, thereby increasing colonization andpenetration of host tissues and possibly promoting disseminationof this parasite via the circulatory system. Extracellular MMP-9activity might also assist the hydrolysis of extracellular large poly-peptides into smaller peptides and amino acids, thus playing a rolein nutrition.
This is the first report in which a metallopeptidase activity iscorrelated with MMP-9 expression in T. cruzi. The biological func-tions played by this enzyme in this parasite are still unknown andits role during infections are speculations until now. The activity ofthis metallopeptidase reported here opens up the possibility ofunderstanding its role in T. cruzi biology. Taking into account thatthere is a pressing need for the identification of novel drug targetsand virulence factors to improve the prevention and treatment ofChagas’ disease this peptidase could also be explored as a possibletarget against T. cruzi. However, further studies are necessary inorder to validate this proteolytic activity as a drug target.
Acknowledgments
We thank Dr Célia Regina Avelar Fernandez for generously pro-viding the doxycycline and Marcelo Meuser (FIOCRUZ) for his helpduring flow cytometry and statistical analyses. We also thank MsDenise da Rocha de Souza for technical assistance. This studywas supported by Coordenação de Aperfeiçoamento Pessoal deNível Superior (CAPES), Conselho Nacional de DesenvolvimentoCientífico e Tecnológico (MCT/CNPq), Conselho de Ensino paraGraduados e Pesquisas (CEPG/UFRJ), Fundação Oswaldo Cruz (FIO-CRUZ), Fundação Carlos Chagas Filho de Amparo à Pesquisa do

A.C. Nogueira de Melo et al. / Experimental Parasitology 125 (2010) 256–263 263
Estado do Rio de Janeiro (FAPERJ) and Fundação Universitária JoséBonifácio (FUJB).
References
Aparício, I.M., Scharfstein, J., Lima, A.P.C.A., 2004. A new cruzipain-mediatedpathway of human cell invasion by Trypanosoma cruzi requires trypomastigotemembranes. Infection and Immunity 77, 5892–5902.
Barr, S.C., Warner, K.L., Kornreic, B.G., Piscitelli, J., Wolfe, A., Benet, L., McKerrow,J.H., 2005. A cysteine protease inhibitor protects dogs from cardiac damageduring infection by Trypanosoma cruzi. Antimicrobial Agents and Chemotherapy49, 5160–5161.
Bastos, I.M., Grellier, P., Martins, N.F., Cadavid-Restrepo, G., de Souza-Ault, M.R.,Augustyns, K., Teixeira, A.R., Schrével, J., Maigret, B., da Silveira, J.F., Santana,J.M., 2005. Molecular, functional and structural properties of the prolyloligopeptidase of Trypanosoma cruzi (POP Tc80), which is required for parasiteentry into mammalian cells. The Biochemical Journal 15, 29–38.
Brack, K., Doyle, P.S., McKerrow, J.H., Ellman, J.A., 2008. Identification of a new classof nonpeptidic inhibitors of cruzain. The Journal of the American ChemicalSociety 130, 6404–6410.
Bern, C., Montgomery, S.P., Herwaldt, B.L., Rassi, A.Jr., Marin-Neto, J.A., Dantas, R.O.,Maguire, J.H., Acquatella, H., Morillo, C., Kirchhoff, L.V., Gilman, R.H., Reyes, P.A.,Salvatella, R., Moore, A.C., 2007. Evaluation and treatment of Chagas disease inthe United States: a systematic review. The Journal of the American MedicalAssociation 298, 2171–2181.
Bonaldo, M.C., d’Escoffier, L.N., Salles, J.M., Goldenberg, S., 1991. Characterizationand expression of proteases during Trypanosoma cruzi metacyclogenesis.Experimental Parasitology 73, 44–51.
Buache, E., Garnotel, R., Aubert, D., Gillery, P., Villena, I., 2007. Reduced secretionand expression of gelatinase profile in Toxoplasma gondii-infected humanmonocytic cells. Biochemical and Biophysical Research Communications 359,298–303.
Burleigh, B.A., Woolsey, A.M., 2002. Cell signalling and Trypanosoma cruzi invasion.Cellular Microbiology 4, 701–711.
Calvet, C.M., Meuser, M., Almeida, D., Meirelles, M.N., Pereira, M.C., 2004.Trypanosoma cruzi-cardiomyocyte interaction: role of fibronectin in therecognition process and extracellular matrix expression in vitro and in vivo.Experimental Parasitology 107, 20–30.
Cazzulo, J.J., 2002. Proteinases of Trypanosoma cruzi: potential targets for thechemotherapy of Chagas’ disease. Current Topics in Medical Chemistry 2, 1261–1271.
Cockle, J.A., Gopichandran, N., Walker, J.J., Levene, M.I., Orsi, N.M., 2007. Matrixmetalloproteinases and their tissue inhibitors in preterm perinatalcomplications. Reproductive Science 14, 629–645.
Costa, J.D., Nogueira de Melo, A.C., Vermelho, A.B., Meirelles, M.N., Porrozzi, R., 2008.In vitro evidence for metallopeptidase participation in hepatocyte damageinduced by Leishmania chagasi-infected macrophages. Acta Tropica 106, 175–183.
Cuevas, I.C., Cazzulo, J.J., Sanchez, D.O., 2003. Gp63 homologues in Trypanosomacruzi: surface antigens with metalloprotease activity and a possible role in thehost cell infection. Infection and Immunity 71, 5739–5749.
Cuenca, J., Martín-Sanz, P., Álvarez-Barrientos, A.M., Boscá, L., Goren, Nora, 2006.Infiltration of inflammatory cells plays an important role in matrixmetalloproteinase expression and activation in the heart during sepsis. TheAmerican Journal of Pathology 169, 1567–1576.
d’Avila-Levy, C.M., Souza, R.F., Gomes, R.C., Vermelho, A.B., Branquinha, M.H., 2003.A novel extracellular calcium-dependent cysteine proteinase from Crithidiadeanei. Archives of Biochemistry and Biophysics 420, 1–8.
Dietmann, A., Helbok, R., Lackner, P., Issifou, S., Lell, B., Matsiegui, P., Reindl, M.,Schmutzhard, E., Kremsner, P.G., 2008. Matrix metallopeptidases and theirtissue inhibitors (TIMPs) in Plasmodium falciparum malaria: serum levels ofTIMP-1 are associated with disease severity. The Journal of Infection Diseases197, 1614–1620.
Duschak, V.G., Barboza, M., García, G.A., Lammel, E.M., Couto, A.S., Isola, E.L.D., 2006.Novel cysteine proteinase in Trypanosoma cruzi metacyclogenesis. Parasitology132, 1–11.
Elias, C.G.R., Pereira, F.M., Silva, B.A., Alviano, C.S., Soares, R.M.A., Santos, A.L.S., 2006.Leishmanolysin (gp63 metalloproteinase)-like activity extracellularly releasedby Herpetomonas samuelpessoai. Parasitology 132, 1–11.
Engel, J.C., Doyle, P.S., Hsieh, I., McKerrow, J.H., 1998. Cysteine protease inhibitorscure na experimental Trypanosoma cruzi infection. The Journal of ExperimentalMedicine 188, 725–734.
Engman, D.M., Leon, J.S., 2002. Pathogenesis of Chagas heart disease: role ofautoimmunity. Acta Tropica 81, 123–132.
Ersfeld, K., Barraclough, H., Gull, K., 2005. Evolutionary relationships and proteindomain architecture in an expanded calpain superfamily in kinetoplastidparasites. The Journal of Molecular Evolution 61, 742–757.
Galis, Z.S., Khatri, J.J., 2002. Matrix metalloproteinases in vascular remodeling andatherogenesis: the good, the bad, and the ugly. Circulation Research 90, 251–262.
Garcia, M.P., Nóbrega, O.T., Teixeira, A.R.L., Sousa, M.V., Santana, J.M., 1998.Characterisation of a Trypanosoma cruzi acidic 30 kDa cysteine protease.Molecular and Biochemical Parasitology 91, 263–272.
Giese, V., Dallagiovanna, B., Marchini, F.K., Pavoni, D.P., Krieger, M.A., Goldenberg, S.,2008. Trypanosoma cruzi: a stage-specific calpain-like protein is induced aftervarious kinds of stress. Memórias do Instituto Oswaldo Cruz 103, 598–601.
Goll, D.E., Thompson, V.F., Li, H., Wei, W., Cong, J., 2003. The calpain system.Physiological Reviews 83, 731–801.
Gomis-Rüth, F.X., 2009. Catalytic domain architecture of metzincinmetalloproteases. The Journal of Biological Chemistry 284, 15353–15357.
Gutierrez, F.R., Lalu, M.M., Mariano, F.S., Milanezi, C.M., Cena, J., Gerlach, R.F., Santos,J.E., Torres-Dueñas, D., Cunha, F.Q., Schulz, R., Silva, J.S., 2008. Increased activitiesof cardiac matrix metalloproteinases matrix metalloproteinase (MMP)-2 andMMP-9 are associated with mortality during the acute phase of experimentalTrypanosoma cruzi infection. The Journal of Infectious Diseases 197, 1468–1476.
Hanemaaijer, R., Visser, H., Koolwijk, P., Sorsa, T., Salo, T., Golub, L.M., VanHinsbergh, V.W.M., 1998. Inhibition of MMP synthesis by doxycycline andchemically modified tetracyclines (CMTs) in human endothelial cells. Advancesin Dental Research 12, 114–118.
Heussen, C., Dowdle, E.B., 1980. Electrophoretic analysis of plasminogen activatorsin polyacrylamide gels containing sodium dodecyl sulphate and copolymerizedsubstrates. Analytical Biochemistry 102, 196–202.
Li, J., Schwimmbeck, P.L., Tschope, C., Leschka, S., Husmann, L., Rutschow, S.,Reichenbach, F., Noutsias, M., Kobalz, U., Poller, W., Spillmann, F., Zeichhardt, H.,Schultheiss, H.P., Pauschinger, M., 2002. Collagen degradation in a murinemyocarditis model: relevance of matrix metalloproteinase in association withinflammatory induction. Cardiovascular Research 56, 235–247.
Lowndes, C.M., Bonaldo, M.C., Thomaz, N., Goldenberg, S., 1996. Heterogeneity ofmetalloprotease expression in Trypanosoma cruzi. Parasitology 112, 393–399.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951. Protein measurementwith the Folin phenol reagent. The Journal of Biological Chemistry 19, 264–275.
Malla, N., Sjoli, S., Winberg, J.O., Hadler-Olsen, E., Uhlin-Hansen, L., 2008. Biologicaland pathobiological functions of gelatinase dimmers and complexes. ConnectTissue Research 49, 180–184.
Meirelles, M.N.L., Souto-Padrón, T., De Souza, W., 1984. Participation of the cellsurface anionic sites in the interaction between Trypanosoma cruzi andmacrophages. Journal of Submicroscopic Cytology and Pathology 16, 533–545.
Nogueira de Melo, A.C., Meirelles, M.N.L., Porrozzi, R., Costa, J.D., Branquinha, M.H.,Vermelho, A.B., 2004. Reduced activity of matrix metalloproteinase-9 inTrypanosoma cruzi infected mouse embryo hepatocyte cell. HepatologyResearch 28, 49–56.
Opdenakker, G., Van Den Steel, P.E., Van Damme, J., 2001. Gelatinase B: a turner andamplifier of immune functions. Trends in Immunology 22, 571–579.
Pinho, R.T., Beltramini, L.M., Alves, C.R., De-Simone, S.G., 2009. Trypanosoma cruzi:isolation and characterization of aspartyl proteases. Experimental Parasitology122, 128–133.
Rossi, M.A., 2001. Connective tissue skeleton in the normal left ventricle and inhypertensive left ventricular hypertrophy and chronic chagasic myocarditis.Medical Science Monitor 7, 820–832.
Rutschow, S., Li, J., Schultheiss, H.P., Pauschinger, M., 2006. Myocardial proteasesand matrix remodeling in inflammatory heart disease. Cardiovascular Research69, 646–656.
Sangenito, L.S., Ennes-Vidal, V., Marinho, F.A., Da Mota, F.F., Santos, A.L.S., d’Avila-Levy, C.M., Branquinha, M.H., 2009. Arrested growth of Trypanosoma cruzi by thecalpain inhibitor MDL28170 and detection of calpain homologues inepimastigote forms. Parasitology 136, 433–441.
Scharfstein, J., Schmitz, V., Morandi, V., Capella, M.M., Lima, A.P., Morrot, A., Juliano,L., Muller-Esterl, W., 2000. Host cell invasion by Trypanosoma cruzi ispotentiated by activation of bradykinin B(2) receptors. The Journal ofExperimental Medicine 192, 1289–1300.
Souto-Padrón, T., Campetella, O.E., Cazzulo, J.J., De Souza, W., 1990. Cysteineproteinase in Trypanosoma cruzi: immunocytochemical localization andinvolvement in parasite-host cell interaction. Journal of Cell Science 96, 485–490.
Spinale, F.G., 2007. Myocardial matrix remodeling and the matrixmetalloproteinases: influence on cardiac form and function. PhysiologicalReviews 87, 1285–1342.
Van den Steen, P.E., Dubois, B., Nelissen, I., Rudd, P.M., Dwek, R.A., Opdenakker, G.,2002. Biochemistry and molecular biology of gelatinase B or matrixmetalloproteinase-9 (MMP-9). Critical Reviews in Biochemistry and MolecularBiology 37, 375–536.
Vermelho, A.B., Giovanni-De-Simone, S., d’Avila-Levy, C.M., Santos, A.L.S., Nogueira deMelo, A.C., Silva Jr,, F.P., Bon, E.P., Branquinha, M.H., 2007. Trypanosomatidae:peptidases and drugs development. Current Enzyme Inhibition 3, 19–48.
Xu, L., Jiang, W., Ma, T., Qiu, P., Hou, J., Huang, Y., Chen, J., Yan, G., 2006. Expression,purification and molecular modeling of recombinant fibrinogenase [IV], ametalloproteinase from Deinakistrodon acutus venom. Toxicon 47, 241–248.
Yoshida, N., 2009. Molecular mechanisms of Trypanosoma cruzi infection by oralroute. Memórias do Instituto Oswaldo Cruz 104, 101–107.
WHO – World Health Organization, 2006. Strategic and technical meeting onintensified control of neglected tropical diseases. In: Proceedings of the Report ofan International Workshop. Berlin, April 18–20, 2005, WHO/CDS/NTD/2006.1.Available from: <http://www.who.int/neglecteddiseases/en/> (accessed inDecember 2006).
Copyright © 2022 FDOKUMEN




















