
TLR ligands of ryegrass pollen microbial contaminants enhance Th1 and Th2 responses and decrease...
11
Eur. J. Immunol. 2013. 43: 723–733 Immunomodulation DOI: 10.1002/eji.201242747 723 TLR ligands of ryegrass pollen microbial contaminants enhance Th1 and Th2 responses and decrease induction of Foxp3 hi regulatory T cells Diana Mittag 1,2,3 , Nirupama Varese 1,2 , Anja Scholzen 1 , Ashley Mansell 4 , Gillian Barker 5 , Gregory Rice 5 , Jennifer M. Rolland 1,2 and Robyn E. O’Hehir 1,2 1 Department of Immunology, Monash University, Melbourne, Australia 2 Department of Allergy, Immunology and Respiratory Medicine, Alfred Hospital and Monash University, Melbourne, Australia 3 Transfusion Research, Australian Red Cross Blood Services, Melbourne, Australia 4 Monash Institute of Medical Research, Monash University, Melbourne, Australia 5 Translational Proteomics, Baker Heart Institute, Melbourne, Australia Microbial contamination of grass pollens could affect sensitization, subsequent aller- gic response, and efficacy of allergen-specific immunotherapy. We investigated whether bacterial immunomodulatory substances can direct PBMC responses of allergic and nonatopic subjects against ryegrass pollen (RGP) toward Th1, Th2, or regulatory T (Treg) cells. Aqueous extracts of RGP with high or low LPS were fractionated into large and small molecular weight (MW) components by diafiltration. CFSE-labeled PBMCs from allergic and nonatopic subjects were stimulated with RGP extracts (RGPEs) and analyzed for cytokine secretion and T-cell responses. High LPS RGPE increased IFN-γ + Th1 and IL-4 + Th2 effector cell induction and consistently decreased CD4 + Foxp3 hi Treg-cell induc- tion. IL-10-producing T-cell frequency was unaltered, but IL-10 secretion was increased by high LPS RGPE. RGPE-stimulation of TLR-transfected cell lines revealed that high LPS pollen also contained a TLR2-ligand, and both batches a TLR9-ligand. Beta-1,3-glucans were detected in large and small MW fractions and were also T-cell stimulatory. In con- clusion, coexposure to allergen and proinflammatory microbial stimuli does not convert an established Th2- into a Th1-response. Instead, proinflammatory responses are exac- erbated and Foxp3 hi Treg-cell induction is decreased. These findings show that adjuvants for specific immunotherapy should enhance Treg cells rather than target immune devia- tion from Th2 to Th1. Keywords: Allergology Immune regulation Immunotherapy T helper cells Toll like receptors Additional supporting information may be found in the online version of this article at the publisher’s web-site Correspondence: Dr. Diana Mittag e-mail: [email protected] Introduction A combination of genetic predisposition and environmental fac- tors determines the development of immune-mediated inflamma- tory diseases including allergy and asthma [1–3]. An important environmental factor is exposure to microbial compounds such C 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of TLR ligands of ryegrass pollen microbial contaminants enhance Th1 and Th2 responses and decrease...

Eur. J. Immunol. 2013. 43: 723–733 ImmunomodulationDOI: 10.1002/eji.201242747 723
TLR ligands of ryegrass pollen microbial contaminantsenhance Th1 and Th2 responses and decreaseinduction of Foxp3hi regulatory T cells
Diana Mittag1,2,3, Nirupama Varese1,2, Anja Scholzen1, Ashley Mansell4,Gillian Barker5, Gregory Rice5, Jennifer M. Rolland1,2
and Robyn E. O’Hehir1,2
1 Department of Immunology, Monash University, Melbourne, Australia2 Department of Allergy, Immunology and Respiratory Medicine, Alfred Hospital and Monash
University, Melbourne, Australia3 Transfusion Research, Australian Red Cross Blood Services, Melbourne, Australia4 Monash Institute of Medical Research, Monash University, Melbourne, Australia5 Translational Proteomics, Baker Heart Institute, Melbourne, Australia
Microbial contamination of grass pollens could affect sensitization, subsequent aller-gic response, and efficacy of allergen-specific immunotherapy. We investigated whetherbacterial immunomodulatory substances can direct PBMC responses of allergic andnonatopic subjects against ryegrass pollen (RGP) toward Th1, Th2, or regulatory T (Treg)cells. Aqueous extracts of RGP with high or low LPS were fractionated into large andsmall molecular weight (MW) components by diafiltration. CFSE-labeled PBMCs fromallergic and nonatopic subjects were stimulated with RGP extracts (RGPEs) and analyzedfor cytokine secretion and T-cell responses. High LPS RGPE increased IFN-γ+ Th1 andIL-4+ Th2 effector cell induction and consistently decreased CD4+Foxp3hi Treg-cell induc-tion. IL-10-producing T-cell frequency was unaltered, but IL-10 secretion was increasedby high LPS RGPE. RGPE-stimulation of TLR-transfected cell lines revealed that high LPSpollen also contained a TLR2-ligand, and both batches a TLR9-ligand. Beta-1,3-glucanswere detected in large and small MW fractions and were also T-cell stimulatory. In con-clusion, coexposure to allergen and proinflammatory microbial stimuli does not convertan established Th2- into a Th1-response. Instead, proinflammatory responses are exac-erbated and Foxp3hi Treg-cell induction is decreased. These findings show that adjuvantsfor specific immunotherapy should enhance Treg cells rather than target immune devia-tion from Th2 to Th1.
Keywords: Allergology � Immune regulation � Immunotherapy � T helper cells � Toll likereceptors
� Additional supporting information may be found in the online version of this article at thepublisher’s web-site
Correspondence: Dr. Diana Mittage-mail: [email protected]
Introduction
A combination of genetic predisposition and environmental fac-tors determines the development of immune-mediated inflamma-tory diseases including allergy and asthma [1–3]. An importantenvironmental factor is exposure to microbial compounds such
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

724 Diana Mittag et al. Eur. J. Immunol. 2013. 43: 723–733
as lipopolysaccharide (LPS), which can modulate both innateand adaptive immune responses. This is mediated by patternrecognition receptors (PRRs) on antigen-presenting cells (APCs)such as dendritic cells (DCs) that specifically recognize pathogen-or endogenous danger-associated molecular patterns (PAMPs orDAMPs). In the T helper (Th) cell response to antigen, APCs notonly present antigen-derived peptides but also, via PRR detectedsignals, direct the type of T-cell response to be inflammatory (Th1,Th2, Th17) or tolerogenic (Tr1, anergy) (reviewed in [4,5]). Sev-eral allergens have been shown to carry PAMPs or directly stimu-late PRRs. For example, house dust mite faeces contain not onlyallergenic proteins but also bacterial DNA, endotoxin and chitin[6–8]. The peanut allergen Ara h 1 is a ligand for the carbohydratereceptor DC-SIGN that facilitates Ara h 1 uptake by DCs [9]. Birchpollen-derived substances, such as phytoprostanes and adenosine,can bias immune responses toward Th2 via modulation of DCs[10,11].
The role of exposure to microbes in allergic sensitization hasbeen studied extensively in the context of the hygiene hypothesis,which states that high exposure to particular microbes protectsagainst development of Th2-based disease. In murine studies, lowlevels of LPS enhanced Th2-type responses to inhaled antigensand high LPS doses favored a Th1 response [12]. The incidenceof allergic sensitization is inversely correlated with level of expo-sure to LPS in children with a particular single nucleotide poly-morphism in the CD14 promoter region [13]. Conversely, LPSfound on pollen grains due to Gram-negative bacterial contami-nation has been suggested to facilitate sensitization to pollen aller-gens as well as aggravate allergic responses in already sensitizedindividuals [14].
Microbial contaminants in allergen extracts are also likely toaffect efficacy of allergen-specific immunotherapy (SIT), poten-tially in different ways. SIT is the only curative treatment forallergy and involves subcutaneous injection of increasing dosesof allergen extract (SCIT) or, more recently, sublingual adminis-tration of allergen extracts (SLIT). SIT is most successful in thetreatment of bee venom and pollen allergies, but is also usefulin the treatment of house dust mite and domestic pet allergies.Since allergy is characterized by a Th2-biased immune response,therapeutic approaches include the use of adjuvants to promoteimmune deviation to a Th1-polarized response [15]. The mecha-nisms of SIT, however, are still not fully understood. Trivedi et al.[16] showed that pollen extracts used in SIT have highly variableLPS content and suggested that the influence of LPS contami-nation of allergen extracts on SIT efficacy and adverse reactionsneeds to be examined. Environmental LPS exposure of individualswith allergic airway diseases, including asthma, is known to exac-erbate the disease [17]. Therefore, such “accidental” coadminis-tration of LPS with pollen allergen extracts during SIT may notbe beneficial. Clarifying the role of LPS and other microbial con-taminants in SIT efficacy is hampered by the high batch-to-batchvariation in allergen content as well as contaminating substances[18].
Previous studies on the role of microbial stimuli in SIT havefocused largely on shifting the Th2 bias to Th1. A deficiency in
immune regulation, however, is now recognized as a major under-lying mechanism of allergic responses and therapeutic approachesfor allergy are now targeting induction of tolerance, in particularvia antigen-specific regulatory T (Treg) cells [19]. We recentlydemonstrated that allergen-mediated induction of Treg cells wasimpaired in allergic compared with nonatopic subjects [20], andin a clinical study of SLIT in patients with house dust mite allergy,we showed that in the active group, improved symptoms wereassociated with increased numbers of Treg cells [21].
In the present study, we performed a detailed investigationof the immunomodulatory effects of microbial contamination ofallergen extracts on human PBMC responses to ryegrass pollen(RGP) allergens, separating small and large molecular weight(MW) components of the pollen extracts. The presence of beta-1,3-glucans and particular TLR ligands in the pollen fractions wasdetermined and effector and Treg-cell responses of allergic andnonatopic individuals to aqueous extracts from RGP with high orlow LPS content were compared.
Results
RGP extracts have highly variable LPS content, whichis not removed by diafiltration
Three different RGP batches were extracted (1:20 w/v) andextracts tested for endotoxin content by the manufacturer GreerLaboratories with results ranging from 50 to 5000 EU/mL (Sup-porting Information Fig. 1A). We chose the high and low LPSbatches (A and C) for our further studies and prepared aque-ous extracts. We then fractionated these extracts by diafiltrationusing a 10 kDa cut-off that separated proteins and LPS from smallmolecule components (Supporting Information Fig. 1B) similarto dialysis. Diafiltration is faster than dialysis and thus decreasesthe risk of allergen protein degradation. Furthermore, small MWcomponents remained concentrated in the filtrate, which allowedtheir further analysis in comparison with the large MW fractions.
Protein and allergen content of RGPE fractions
Protein content of RGPEs was analyzed by two different proteinassays, Bradford and BCA. While these assays gave similar resultsfor the diafiltrated large MW fractions, there was a large dis-crepancy between the two methods when testing the small MWfractions and unfractionated total RGPE (Supporting InformationFig. 1C). The BCA protein assay detected a relatively high sig-nal in the small MW fraction despite minimal amounts of proteindetected by silver staining after SDS-PAGE (Supporting Informa-tion Fig. 2A) and a very low result in the Bradford assay. TheBCA assay also gave a higher signal in the unfractionated extractthan the Bradford assay. This indicates that nonprotein small MWcomponents cause a signal in the BCA, but not the Bradford assay.Therefore, the Bradford assay was used to quantify proteins in thetotal unfractionated extract and the large MW fraction, while the
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Eur. J. Immunol. 2013. 43: 723–733 Immunomodulation 725
BCA assay “protein concentration” readout was used to standard-ize the small MW fractions.
Allergens were identified using monoclonal antibodies againstthe major RGP allergens Lol p 1 and Lol p 5 as well as serumfrom RGP allergic subjects. Similar protein and allergen profileswere observed for high and low LPS RGPEs (Supporting Informa-tion Fig. 2). The majority of proteins were retained in the largeMW fractions (Supporting Information Fig. 2A). Nonatopic donorsera had negligible IgE binding to small MW components andthe allergen-specific monoclonal antibodies detected no Lol p 1and Lol p 5 (Supporting Information Fig. 2C). Sera from RGP-allergic donors had IgE-reactivity to small MW fractions of bothlow (Supporting Information Fig. 2C) and high LPS RGPE (datanot shown).
LPS content affects T-cell proliferative activity, but notmitogenicity of RGPEs
The protein concentration of RGPE for suboptimal stimulation ofT-cell proliferation was determined by 3H-thymidine incorpora-tion assay (Supporting Information Fig. 3A). Extracts from highand low LPS RGP differed in their capacity to stimulate prolifer-ation of PBMCs from allergic donors. At high protein concentra-tions, both the total extract and the large MW fraction of the highLPS RGPE had a decreased capacity to stimulate T-cell prolifer-ation, compared with the low LPS extracts at higher concentra-tions, suggesting a component that is toxic or reduces prolifera-tion. Despite the very low actual protein concentration, the smallMW extract fractions were able to induce PBMC proliferation,albeit at low levels.
Extracts were further characterized for toxic or inhibitory prop-erties and mitogenicity. Proliferation of a latex allergen Hev b5-specific T-cell line in response to IL-2 was measured in the pres-ence or absence of RGPE. Decreased proliferation was observedin the presence of the high LPS RGPE, and to a lesser extent withthe low LPS extracts (Supporting Information Fig. 3B). The effectwas greater for both small MW fractions compared with the largeMW fractions, confirming that dialysis or diafiltration is an impor-tant step for removing potentially toxic or immune modulatorycomponents in preparation of pollen extracts for SIT (SupportingInformation Fig. 3B). None of the extracts induced nonspecificT-cell proliferation in a latex allergen Hev b 5-specific T-cell line(Supporting Information Fig. 3C) showing that the RGPEs do notcontain mitogens.
LPS enhances proinflammatory cytokine productionas well as IL-10 secretion
Cytokine secretion was determined by a 12-cytokine multiplexassay of cell culture supernatants after 48 h based on a time-courseexperiment (Fig. 1A). High LPS RGPE increased proinflammatoryand effector Th1 and Th2 cytokine levels in PBMCs from allergic
(IFN-γ (p = 0.0039), TNF-α (p = 0.0039), IL-5 (p = 0.0156),IL-13 (p = 0.0156)) and nonatopic (IFN-γ (p = 0.0156), TNF-α(0.0039), IL-13 (p = 0.0313)) donors as well as IL-10 (allergic:p = 0.0039, nonallergic: p = 0.0156) when compared with lowLPS RGPE (Fig. 1C). IL-12p70 was only above the detection limitin five subjects if they were stimulated with high LPS RGPE orlow LPS RGPE with added LPS, but not in the absence of LPS.Only very low levels of IL-4 were observed and no RGPE frac-tion induced IL-17. LPS alone induced IFN-γ and TNF-α as well asIL-10 (Fig. 1B and D). Addition of LPS to low LPS RGPE frac-tions increased secretion of these cytokines in allergic (IFN-γ,p = 0.0039, TNF-α, p = 0.0039, IL-10, p = 0.0078) and nonatopic(IFN-γ, p = 0.0223, TNF-α, p = 0.0156, IL-10, p = 0.0313) donors,indicating they were induced by LPS itself. Consistently, block-ing LPS with an antagonist decreased secretion of IFN-γ, TNF-α,and IL-10 (allergic: IFN-γ, p = 0.0039, TNF-α, p = 0.0039, IL-10,p = 0.0039, and nonatopic: IFN-γ, p = 0.0156, TNF-α, p =0.0156, IL-10, p = 0.0156) in response to high LPS RGPE(Fig. 1D).
Both high and low LPS RGPEs induced higher Th2 cytokinesecretion in allergic subjects compared with that of nonatopic con-trols (IL-5, p = 0.0120, IL-13, p = 0.0166 for combined high andlow LPS RGPEs) (Fig. 1C). High LPS RGPE induced more IL-13secretion than low LPS RGPE. Addition of LPS increased IL-13secretion above the detection limit in PBMCs of two nonatopicsubjects that did not secrete IL-13 in response to low LPS RGPElarge MW fraction alone (Fig. 1D). Blocking LPS in the high LPSRGPE large MW fraction had no effect on IL-13 secretion indicat-ing that it is induced by components other than LPS.
The small MW fractions of both low and high LPS RGPEsinduced similar cytokine levels. Interestingly, the low LPS RGPEsmall MW fraction induced higher IL-10 secretion than the largeMW fraction (p = 0.0010) (Fig. 1C). This indicates that dialysismay remove small MW components that could have a tolerogeniceffect, in particular when contamination with Gram-negative bac-teria is low.
LPS increases induction of both IFN-γ and IL-4producing effector T cells
Consistent with the increased cytokine secretion, induction of Th1and Th2 effector cells (dividing CD4+IFN-γ+ and IL-4+ T cells)was increased in response to high LPS RGPE (Fig. 2A–C). Therewas a trend for increased IL-4+CD4+ Th2 effector cell inductionin allergic subjects with both RGPEs compared with nonatopicsubjects (Fig. 2B). This indicates that the Th2-biased responsein allergic subjects could not be overcome by natural adjuvantspresent in the high LPS RGPE. Interestingly, the induction ofIFN-γ+ CD4+ T cells in response to high LPS RGPE was signifi-cantly higher in nonatopic than in allergic subjects (Fig. 2B). Theratio of IL-4+/IFN-γ+ dividing CD4+ T cells was not significantlydifferent between high and low LPS extracts (Fig. 2C).
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
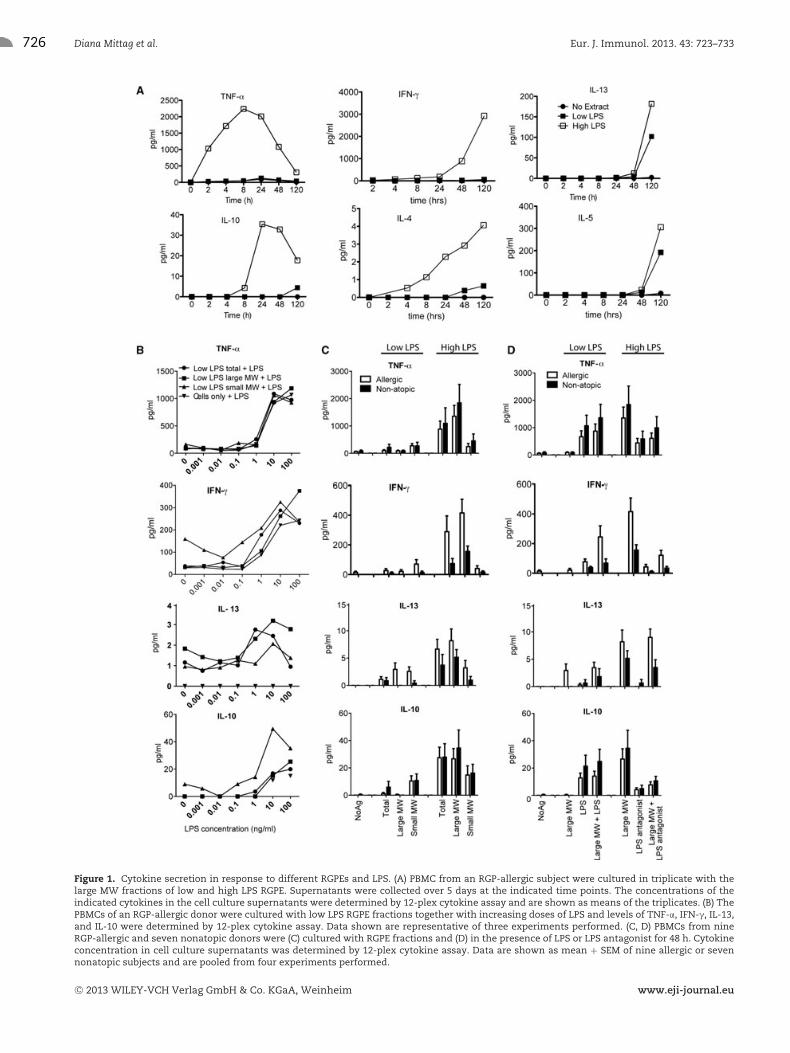
726 Diana Mittag et al. Eur. J. Immunol. 2013. 43: 723–733
Figure 1. Cytokine secretion in response to different RGPEs and LPS. (A) PBMC from an RGP-allergic subject were cultured in triplicate with thelarge MW fractions of low and high LPS RGPE. Supernatants were collected over 5 days at the indicated time points. The concentrations of theindicated cytokines in the cell culture supernatants were determined by 12-plex cytokine assay and are shown as means of the triplicates. (B) ThePBMCs of an RGP-allergic donor were cultured with low LPS RGPE fractions together with increasing doses of LPS and levels of TNF-α, IFN-γ, IL-13,and IL-10 were determined by 12-plex cytokine assay. Data shown are representative of three experiments performed. (C, D) PBMCs from nineRGP-allergic and seven nonatopic donors were (C) cultured with RGPE fractions and (D) in the presence of LPS or LPS antagonist for 48 h. Cytokineconcentration in cell culture supernatants was determined by 12-plex cytokine assay. Data are shown as mean + SEM of nine allergic or sevennonatopic subjects and are pooled from four experiments performed.
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Eur. J. Immunol. 2013. 43: 723–733 Immunomodulation 727
Figure 2. High LPS RGPE increases both Th1- and Th2-cytokine producing CD4+ T cells as well as IL-10+Foxp3int CD4+ T cells. (A) CFSE-labeledPBMCs were cultured with RGPE for 7 days, restimulated for 6 h with anti-CD3 and anti-CD28 in the presence of Brefeldin A, stained for surfacemarkers and intracellular cytokines and analyzed by flow cytometry as shown. (B) Analysis of RGP-allergic (n = 14) and nonatopic subjects (n = 12)for the frequency of dividing CD4+ IL-4+ or IFN-γ+ cells after stimulation with the large MW RGPE fraction is shown. (C) The ratio of IL-4+/IFN-γ+
T cells within the CD4+ dividing T cells is shown. (D) CFSE-labeled PBMCs were cultured with RGPE for 6 days, restimulated overnight with anti-CD3and anti-CD28, stained for secreted surface-captured IL-10 and analyzed by flow cytometry as shown. (E, F) Analysis of RGP-allergic (n = 14) andnonatopic subjects (n = 12) for frequency of IL-10+ cells in (E) dividing CD4+ Foxp3hi or (F) Foxp3int cells is shown. Data shown are pooled from fourexperiments performed. Statistical significance was assessed by either two-tailed paired Students t-test for paired values (comparing low withhigh LPS in the same subject) or Mann–Whitney test (nonparametric) for independent values (comparing allergic with nonatopic subjects). *p <
0.05, **p < 0.01, ***p < 0.001; ns = not significant.
Differential induction of IL-10 in Foxp3hi and Foxp3int
dividing CD4+ T cells by LPS
A subset of Treg cells characterized by IL-10 secretion (Tr1)has been described [22]. Tr1 have been shown to regulatethe adverse Th2 response in allergic disease [23]. Inductionof dividing IL-10+CD4+ T cells was not significantly different
between high and low LPS RGPE (Fig. 2D and E). When CD4+
T cells, however, were subdivided based on Foxp3 expression,opposite effects of LPS on Foxp3hi and Foxp3int CD4+ T cellsbecame apparent. There was a trend for fewer Foxp3hi but moreFoxp3int cells to secrete IL-10 in response to high comparedwith low LPS RGPE (Fig. 2F) in both groups. These differenceswere statistically significant (Foxp3hi cells: p = 0.006; Foxp3int
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

728 Diana Mittag et al. Eur. J. Immunol. 2013. 43: 723–733
Figure 3. Treg-cell induction is reduced by high LPS RGPE. (A) CFSE-labeled PBMCs were cultured for 7 days and then stained for surface markersand Foxp3 for flow cytometry analysis. The strategy for gating Foxp3hi Treg cells in dividing CD4+ T cells is shown. (B) CD4+ T-cell prolifer-ation and (C) CFSE-labeled PBMCs from RGP-allergic (n = 20) and nonatopic (n = 12) subjects were cultured for 7 days with RGPE fractions.The frequency of dividing Foxp3hi CD4+ T cells was analyzed by flow cytometry. (D) CFSE-labeled PBMCs from allergic (n = 6) and nonatopic(n = 5) subjects were cultured for 7 days with RGPE fractions and LPS or LPS antagonist. Data shown are pooled from five experiments per-formed. Statistical significance was assessed by either two-tailed paired Students t-test for paired values (comparing low with high LPS in thesame subject) or Mann–Whitney test (nonparametric) for independent values (comparing allergic with nonatopic subjects). *p < 0.05, **p < 0.01,***p < 0.001.
cells: p = 0.05) when all subjects (allergic and nonatopic) werecompared.
Induction of CD4+Foxp3hi Treg cells
Recently, we described a subset of CD4+ Treg cells induced inresponse to RGPE stimulation. These cells are proliferating, but incontrast to T-helper cells they express very low levels of inflam-matory cytokine and suppressed CD4+ T-cell proliferation andIFN-γ production. Instead, these cells are characterized by higherand more stable levels of Foxp3 expression than observed onactivated Th cells [20]. In the present study, the frequency ofthese induced dividing CD4+Foxp3hi Treg cells was decreased forallergic compared with nonatopic subjects (Fig. 3B) while CD4+
T-cell proliferation in response to RGPE was generally higher forallergic than nonatopic subjects (Fig. 3B). This is consistent withour previous finding that RGPE (low LPS) stimulation inducesboth effector T cells and Treg cells, with a reduced Treg-cell fre-
quency in allergic compared to nonatopic subjects [20]. FewerTreg cells were induced with high LPS RGPE compared withlow LPS in both allergic and nonatopic subjects (Fig. 3C). It isimportant to note, that the difference in Treg-cell induction withlarge MW RGPE fractions between the two subject groups (aller-gic and nonatopic) was only significant with low LPS RGPE, butlost when using the high LPS RGPE. Blocking LPS with its antag-onist increased Treg-cell induction in some donors while addingLPS to the low LPS RGPE decreased Treg-cell induction in mostdonors (Fig. 3D). LPS blocking or addition had limited effects inmimicking low and high LPS RGPEs indicating that RGPE com-ponents other than LPS contribute to the differences in Treg-cellinduction.
Effects of non-LPS pollen components
Overall, high LPS RGPE increased proinflammatory and effectorTh1 and Th2 responses as well as IL-10 and reduced induction of
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Eur. J. Immunol. 2013. 43: 723–733 Immunomodulation 729
Figure 4. Contaminants other than LPS are present in high and low LPS RGPE. (A) Beta-(1,3)-D-glucan levels in RGPE fractions were determinedusing the Glucatell kit (Associates of Cape Cod) based on a modified Limulus Amebocyte Lysate reagent. (B) RGPE fractions were screened forthe ability to stimulate NF-κB in mouse RAW264.7 macrophages (n = 3). (C) Total and large MW fractions of the high LPS RGPE were testedfor their ability to stimulate TLR4-transfected cells (n = 3). (D, E) The effects of (D) high (n = 3) and (E) low (n = 2) LPS RGPE on TLR2-, TLR3-,and TLR9-transfected cells were also tested. All data are shown as mean + SD of the indicated number of experiments each performed intriplicate.
Foxp3hi Treg cells. Low LPS RGPE with added LPS did resemblesome high LPS RGPE effects such as high TNF-α and IL-10 secretionand low Foxp3hi Treg-cell induction. Addition of high dose LPS orantagonising LPS, however, had no effect on IL-13 secretion andno clear effect on CD4+ T-cell proliferation, indicating a role forother pollen extract components causing the differences associatedwith varying levels of microbial contamination. Further testingrevealed the presence of beta-1,3-glucans in the extracts (Fig. 4A)that can be of plant or fungal origin. Levels of beta-1,3-glucanswere higher in the high LPS extract and in the large rather thanthe small MW fractions.
All fractions could activate the innate immune system tosome extent as assessed by activation of the NF-κB-dependent
ELAM promoter linked to luciferase reporter in mouse RAW264.7macrophages (Fig. 4B). The low LPS RGPEs only activatedweakly at the highest doses, while the high LPS RGPE frac-tions potently induced NF-κB activation. Using TLR-transfectedcell lines, the presence of a TLR4-ligand in the high LPS RGPEwas confirmed (Fig. 4C). The high LPS RGPE fractions alsoactivated TLR2- and to a lesser extent TLR9-transfected cells,consistent with contamination with other bacterial componentssuch as bacterial lipopeptides and DNA (Fig. 4D). Low LPSRGPEs only activated TLR9 at low levels (Fig. 4E). None of theRGPE preparations were recognized by TLR3-transfected cells,indicating a lack of viral nucleic acid contamination (Fig. 4Dand E).
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

730 Diana Mittag et al. Eur. J. Immunol. 2013. 43: 723–733
Discussion
After the initial discovery of the association of the allergic diathesiswith a Th2-biased T-cell response, therapeutic approaches focusedon shifting the T-cell response from Th2- to Th1-polarized usingTh1-inducing adjuvants. Bacterial extracts were used by them-selves in the absence of the allergen in the 1970s and 1980s withlittle success [24]. Later, the less toxic LPS derivative, monophos-phoryl lipid A (MPL) was tested as an adjuvant in SCIT and SLIT incombination with chemically modified rather than native allergenextract. Treatment was observed to improve rhinoconjunctivitisand asthma symptom scores, decrease allergen-specific IgE levelsand increase allergen-specific IgG compared with untreated con-trols [15]. However, it is not clear whether this treatment has anyadvantage over traditional SIT.
Grass and tree pollens have been found to be contaminatedwith bacteria and fungi, which introduce various known andunknown adjuvants including LPS. It is, however, not clear howthis might affect the use of allergen extracts from pollen inallergen-specific immunotherapy. Although small MW compo-nents are removed from SIT reagents by dialysis, this fails toremove LPS and other large MW microbial products. In this study,we compared RGPE with low and high LPS content and found thatbacterial products such as LPS augment existing T-cell responses,both Th1 and Th2. Therefore, this bacterial contamination isunlikely to be responsible for immune deviation of Th2 responsesof allergic subjects to Th1 during immunotherapy. Moreover, theinduction of Foxp3hi Treg cells that could suppress the unwantedTh2 response is not only decreased in allergic subjects [20], butalso further suppressed by high LPS. These CD4+Foxp3hi Tregcells are distinct from activated CD4 T cells, that only transientlyupregulate Foxp3 [20].
We detected no IL-17 secretion in response to RGPE frac-tions and have previously found that the frequency of IL-17+ Thcells induced by RGP is low compared with IL-4+ and IFN-γ+
Th cells [20]. Th17 cells have been shown to drive chronicinflammatory disease states such as severe asthma, while ourexperiments are focusing on the initial antigen-specific responses[25,26].
In our study, LPS increased the frequency of dividing IL-10+
Foxp3int CD4+ T cells consistent with Tr1 Treg cells. However,a population of IL-10 secreting activated IFN-γ+ Th1 effectorcells has been described and shown to be important in defenceagainst infection with the malaria parasite Toxoplasmodium gondi[27]. Costaining of IL-10 and IFN-γ would be required to clarifythe presence of this T-cell subset. Both dividing and nondividingIL-10+ CD4 T cells were very rare in our RGPE-stimulated PBMCcultures. Therefore, secreted IL-10, as detected by multiplex assayin this study, is likely to be mostly derived from non-T cells. Nev-ertheless, IL-10+ CD4 T cells could have important effects locallyon other T cells.
LPS has been shown to increase allergen-specific T-cell prolif-eration when present in relatively small concentrations of up to500 EU/mL [25, 26]. The high LPS RGPE used in this study con-tained about 10 times more LPS and the decreased proliferation
that it induced may be due to toxic effects of such high dose LPSor the presence of other toxic components. The reduced prolifera-tion was not due to increased Treg-cell induction. To the contrary,frequency of CD4+Foxp3hi Treg cells in dividing CD4+ T cellswas reduced by high LPS RGPE fractions. Therefore, an impor-tant mechanism that normally controls both Th1- and Th2-typeinflammation is impaired. Similarly, LPS was found to increaseIL-4 as well as IFN-γ production and block Treg-cell developmentin vivo in mice [28]. We also detected TLR2 ligands in the highLPS RGPE and TLR2 stimulation has been shown to favor Th2responses by reducing IFN-γ and IL-12p70 production by den-dritic cells [29]. Interestingly, the receptor for beta-1,3-glucans(which we detected in our extracts), dectin-1, has been shownto synergize with TLR-2 and TLR-4 pathways for TNF-α produc-tion by human monocytes and macrophages [30]. Thus bacterialcontamination may contribute to the exacerbation of proinflam-matory and effector Th1 and Th2 responses consistent with ourstudy, which further emphasizes that bacterial contamination ofallergen preparations for SIT is not beneficial.
Both low and high LPS RGPEs stimulated TLR9-transfectedcells pointing to the presence of CpG motifs from bacterial DNA.In the case of the low LPS RGPE, the CpG DNA may be derivedfrom contaminating Gram-positive bacteria. Both Gram-negativeand Gram-positive bacteria, as well as fungi have been identi-fied on pollen [14]. Genetic variations of TLRs and TLR pathwaygenes associated with asthma and atopy have been identified, inparticular for TLR2, for which we detected a ligand in the high LPSRGPE, and also for TLR9 [31]. CpG immunostimulatory sequences(CpG ISS) are another adjuvant that has been utilized for promot-ing Th1 responses. When CpG ISS was packaged into virus-likeparticles and administered together with house dust mite aller-gen extract, asthma, and rhinitis symptom scores improved andallergen-specific IgG increased [15]. There was also a transientincrease in allergen-specific IgE consistent with our in vitro find-ings that LPS, also a Th1 adjuvant, exacerbated both Th1 and Th2responses in allergic subjects.
The small MW fractions of RGPEs also induced T-cell prolif-eration and cytokine secretion, particularly in allergic subjects.This indicates that they contain T-cell activating factors, proba-bly allergen-derived peptides comprising T-cell epitopes as well asimmune modulating substances other than LPS. In addition to bac-terial and fungal contaminants, pollen-derived substances, such asphytoprostanes and adenosine, can bias immune responses towardTh2 via modulation of dendritic cells [31, 32]. While small MWsubstances are removed from pollen extracts for immunotherapyby dialysis, they may be encountered during natural exposure topollen by inhalation and could affect the type of immune responsetoward the pollen. The small MW fractions did not contain LPS, butwould be expected to contain other smaller microbial substances.Their induced T-cell proliferation was generally low, consistentwith the decreased amount of antigen and hence there was onlya small difference between the two batches of pollen (high andlow LPS) in terms of cytokine secretion and Th differentiation. Wealso observed a nonspecific reduction of T-cell proliferation whenusing high concentrations of both small MW fractions. Whether
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Eur. J. Immunol. 2013. 43: 723–733 Immunomodulation 731
this was due to inhibition of proliferation or to cell death could bedelineated in future studies.
In conclusion, coexposure to allergen and proinflammatorymicrobial stimuli does not further increase the Th1-bias innonatopic or convert an established Th2- into a Th1-response inallergic individuals. Instead, both Th1- and Th2-biased proinflam-matory responses are exacerbated in association with decreasedinduction of Foxp3hi Treg cells. These data provide importantinsight into the adjuvant properties of pollen microbial contami-nants and will aid development of suitable adjuvants for SIT thatenhance Treg cells rather than promote immune deviation fromTh2 to Th1.
Materials and methods
Donors
Twenty-six RGP-allergic donors (mean age 31.1 ± 8.9 years,17/26 male) and 12 nonatopic controls (mean age 42.1 ±15.9 years, 6/12 male) were recruited at the Alfred HospitalAllergy Clinic. A diagnosis of allergy to RGP was based on a clinicalhistory of seasonal rhinitis and evidence of specific sensitizationby positive skin prick test to RGP and/or RGP-specific IgE CAP-FEIA score (Phadia, Uppsala, Sweden) greater than 0.70 kU/L(Class 2). Blood samples were taken outside the RGP season thatin South East Australia runs from late September to early Jan-uary. No donor was suffering from symptoms of allergic rhinitisor asthma at the time of blood sampling. All donors gave writteninformed consent with study approval by The Alfred Hospital andMonash University Ethics Committees.
RGP extracts, diafiltration
Batches of RGP were purchased from Greer Laboratories Inc.(Lenoir, NC) as dry, nondefatted pollen. For RGP extraction,4 g of pollen were suspended in 40 mL PBS for 30 min at37◦C under constant rotation, followed by centrifugation at 5000× g at room temperature. Supernatants were sterile filteredthrough a 0.2-μm pore size filter (Sartorius, Goettingen, Ger-many). RGP extracts (RGPEs) were separated into large and smallMW fractions by diafiltration using Vivaspin 20 (Sartorius) with a10 kDa cut-off membrane following the manufacturer’s instruc-tions. Extracts were stored at –20◦C as allergens were stable underthese conditions as determined by IgE-immunoblotting, while IgEreactivity was decreased after storage at 4◦C for 3 weeks (data notshown).
Endotoxin and (1,3)-beta-D-glucan assay
The endotoxin content of the RGPEs was determined using thePyrochrome Chromogenic 120 Test Kit (Associates of Cape Cod, E.
Falmouth, MA) and (1,3)-beta-D-glucan was measured using theGlucatell kit (Associates of Cape Cod) as per the manufacturer’sinstructions.
Protein assays
Protein content of RGPE fractions was determined by both theBradford assay using the Coomassie Plus kit (Pierce, Rockford,IL) and the Bicinchoninic acid (BCA) assay using the BCA pro-tein assay kit (Pierce). Assays were performed as directed by themanufacturer’s instructions.
SDS-PAGE, silver staining, and immunoblotting
RGPEs were incubated with reducing sample buffer for 5 min at100◦C and proteins separated by SDS-PAGE as described previ-ously [32]. Proteins were stained in the gel using the LC6100SilverXpress Silver Staining Kit (Invitrogen, Carlsbad, CA) as perthe manufacturer’s instructions. Sera were tested for RGP-specificIgE by immunoblotting as described previously [32]. In brief, pro-teins were transferred from the gels onto nitrocellulose membraneand the membrane blocked with 0.3% Tween20/PBS followed byovernight incubation with serum (1:10 dilution). Antigen-boundIgE was detected by incubation with a rabbit-anti-human IgE mon-oclonal antibody (1:500 dilution; Dako, Glostrup, Denmark), fol-lowed by an HRP-conjugated anti-rabbit IgG antibody (1:1000dilution; Promega, Madison, WI) and 4-chloro-1-naphthol sub-strate (Sigma, St Louis, MO). Lol p 1 and Lol p 5 were detected byovernight incubation with specific monoclonal antibodies (A1 andA7, 1:10 dilution), and incubation with sheep-anti-mouse Ig-HRP(1:1000) followed by 4-chloro-1-naphthol substrate.
PBMC isolation
PBMCs were isolated from heparinized blood using Ficoll Paque(Pharmacia, Uppsala, Sweden) density gradient centrifugation.Cells from allergic and nonatopic donors used to compare induc-tion of Foxp3hi cells were cryopreserved and thawed as describedby Maecker et al. [33]. The cells were frozen in 10% DMSO infetal calf serum at 107 cells/mL and stored in liquid nitrogen forsubsequent parallel testing. The viable cell recovery on thawingwas 70 to 80%.
Cell culture
CFSE labeling of PBMCs was performed at a concentration of1 μM CFSE and 107 cells/mL in PBS. Cells were stimulated withRGPEs at 5 μg/mL for total RGPE, 20 μg/mL for large MWRGPE fractions and 100 μg/mL for the small MW RGPE frac-tions unless indicated otherwise. Cells were cultured in completemedium (RPMI supplemented with 2 mM L-glutamine, 100 IU/mL
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

732 Diana Mittag et al. Eur. J. Immunol. 2013. 43: 723–733
penicillin-streptomycin (Invitrogen, Carlsbad, CA) and 5%screened, heat-inactivated human AB serum (Sigma)) in 24-welltissue culture plates (Greiner Labortechnik, Frickenhausen, Ger-many) with 2.5 × 106 PBMCs per well in 2 mL volume at 37◦C in5% CO2 in air in a Heraeus incubator (Heraeus, S. Plainfield, NJ)for 7 days. In some experiments, LPS (Sigma) or a LPS antagonist(Invivogen, San Diego, CA) was added as indicated.
Cytokine analysis
Levels of IL-2, IL-4, IL-5, IL-6, IL-8, IL-10, IL-12p70, IL-13, IL-17,IFN-γ, TNF-α, and GM-CSF in cell culture supernatants sampled at48 h were determined simultaneously by Bioplex assay (BioRad,Hercules, CA) following the manufacturer’s instructions. Samplesfor a standard curve were measured following the manufacturer’sinstructions in parallel with the test samples. Only values withinthe detection limits as given by the manufacturer were used fordata analysis.
Flow cytometry
For labeling of surface markers, cells were incubated with specificfluorochrome conjugated antibodies to CD4, CD25, and CD127(BD Biosciences, San Diego, CA) for 20 min at 4◦C. For Foxp3staining, surface marker stained cells were fixed, permeabilized,and stained for intranuclear Foxp3 using the Foxp3 staining kit(eBioscience, San Diego, CA) according to the manufacturer’sinstructions. For intracellular cytokine staining, 7-day culturedPBMCs were restimulated with phorbol myristate acetate (PMA)and ionomycin (Sigma) or with immobilized anti-CD3 monoclonalantibody (OKT-3, in-house, purified from cell culture supernatant,coated at 4 μg/mL) and 2 μg/mL soluble anti-CD28 (BD Bio-sciences) for 5 h (IL-4, IFN-γ) in the presence of 10 μg/mLBrefeldin A (Sigma) for the last 4 h. Staining with antibodies spe-cific for IL-4 and IFN-γ (BD Biosciences) was performed simultane-ously with staining for Foxp3. Staining for IL-10 secreting cells wasperformed after 16 h restimulation with anti-CD3/anti-CD28 usingthe cytokine secretion assay (CSA) kit (Miltenyi Biotec, Bergisch-Gladbach, Germany) according to the manufacturer’s instructions.Labeled cells were analyzed by flow cytometry on a FACS Calibur(BD Biosciences) or FACS Aria (BD Biosciences).
RAW264.7 mouse macrophage and TLR-transfectedcell lines and stimulation assays
RAW264.7 mouse macrophages stably expressing the κB-dependent ELAM-luciferase reporter were generated as described[34] and maintained in 10% FCS/RPMI and 5% CO2. RAW-ELAMcells were seeded at 4 × 104 cells/well in 96-well format 24 hprior to stimulation with indicated ligands for a further 6 h. Cellswere lysed with Passive Lysis Buffer (Promega) and assessed forluminescence using a FLUOstar Optima plate reader (BMG, Orten-
berg, Germany). Results are represented as fold stimulation ascompared with nonstimulated control.
HEK293 cells stably expressing individual human TLRs as indi-cated (kind gift of Eicke Latz and Douglas Golenbock, UMASS)were maintained in 10% FCS/DMEM in 5% CO2. Luciferasereporter assays were performed as previously described [35, 36]using the 5× κB-luciferase reporter construct (Stratagene, LaJolla, CA) concomitantly with Renilla luciferase-thymidine kinaseencoding plasmid (pRL-TK) (Promega) to normalize for transfec-tion efficiency. Transfected cells were lysed using Passive LysisBuffer (Promega) and assayed for luciferase and Renilla activityusing luciferase assay reagent (Promega) and coleonterazine sub-strate, respectively, and assessed using a FLUOstar Optima platereader. Luminescence readings were corrected for Renilla activityand expressed as fold increase over nonstimulated control values.
Statistical analysis
Statistical analysis was performed using GraphPad Prism softwarev5 (San Diego, CA). Statistical significance was assessed by eitherMann–Whitney test (nonparametric) for independent values, ortwo-tailed paired Students t-test for paired values. A p value <0.05was considered significant.
Acknowledgments: We thank Lorraine Baxter for collection ofblood samples, Geza Paukovics for flow cytometry advice, KathrynHjerrild and Pin Wang for technical assistance with the luciferasereporter assays, and Arnd Petersen for helpful discussion. Wegratefully acknowledge funding for this research from the NationalHealth and Medical Research Council, the Co-operative ResearchCentre for Asthma and Airways, the Victorian Government Infras-tructure Support Program and the Alfred Research Trust, Aus-tralia.
Conflict of interest: The authors declare no financial or commer-cial conflict of interest.
References
1 Ege, M. J., Mayer, M., Normand, A. C., Genuneit, J., Cookson, W. O., Braun-
Fahrlander, C., Heederik, D. et al., Exposure to environmental microor-
ganisms and childhood asthma. N. Engl. J. Med. 2011. 364: 701–709.
2 North, M. L. and Ellis, A. K., The role of epigenetics in the developmen-
tal origins of allergic disease. Ann. Allergy Asthma Immunol. 2011. 106:
355–361; quiz 362.
3 March, M. E., Sleiman, P. M. and Hakonarson, H., The genetics of asthma
and allergic disorders. Discov. Med. 2011. 11: 35–45.
4 Minnicozzi, M., Sawyer, R. T. and Fenton, M. J., Innate immunity in aller-
gic disease. Immunol. Rev. 2011. 242: 106–127.
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Eur. J. Immunol. 2013. 43: 723–733 Immunomodulation 733
5 Platts-Mills, T. A. and Woodfolk, J. A., Allergens and their role in the
allergic immune response. Immunol. Rev. 2011. 242: 51–68.
6 Valerio, C. R., Murray, P., Arlian, L. G. and Slater, J. E., Bacterial 16S
ribosomal DNA in house dust mite cultures. J. Allergy Clin. Immunol. 2005.
116: 1296–1300.
7 Douwes, J., Siebers, R., Wouters, I., Doekes, G., Fitzharris, P. and Crane,
J., Endotoxin, (1 –> 3)-beta-D-glucans and fungal extra-cellular polysac-
charides in New Zealand homes: a pilot study. Ann. Agric. Environ. Med.
2006. 13: 361–365.
8 Gregory, L. G. and Lloyd, C. M., Orchestrating house dust mite-associated
allergy in the lung. Trends Immunol. 2011. 32: 402–411.
9 Shreffler, W. G., Castro, R. R., Kucuk, Z. Y., Charlop-Powers, Z., Grishina,
G., Yoo, S., Burks, A. W. et al., The major glycoprotein allergen from
Arachis hypogaea, Ara h 1, is a ligand of dendritic cell-specific ICAM-
grabbing nonintegrin and acts as a Th2 adjuvant in vitro. J. Immunol.
2006. 177: 3677–3685.
10 Gilles, S., Fekete, A., Zhang, X., Beck, I., Blume, C., Ring, J., Schmidt-
Weber, C. et al., Pollen metabolome analysis reveals adenosine as a
major regulator of dendritic cell-primed TH cell responses. J. Allergy Clin.
Immunol. 2011. 127: 454–461.
11 Traidl-Hoffmann, C., Mariani, V., Hochrein, H., Karg, K., Wagner, H.,
Ring, J., Mueller, M. J. et al., Pollen-associated phytoprostanes inhibit
dendritic cell interleukin-12 production and augment T helper type 2 cell
polarization. J. Exp. Med. 2005. 201: 627–636.
12 Eisenbarth, S. C., Piggott, D. A., Huleatt, J. W., Visintin, I., Herrick,
C. A. and Bottomly, K., Lipopolysaccharide-enhanced, toll-like receptor
4-dependent T helper cell type 2 responses to inhaled antigen. J. Exp.
Med. 2002. 196: 1645–1651.
13 Simpson, A. and Martinez, F. D., The role of lipopolysaccharide in
the development of atopy in humans. Clin. Exp. Allergy 2010. 40:
209–223.
14 Spiewak, R., Skorska, C., Prazmo, Z. and Dutkiewicz, J., Bacterial endo-
toxin associated with pollen as a potential factor aggravating pollinosis.
Ann. Agric. Environ. Med. 1996. 3: 57–69.
15 Casale, T. B. and Stokes, J. R., Future forms of immunotherapy. J. Allergy
Clin. Immunol. 2011. 127: 8–15; quiz 16–17.
16 Trivedi, B., Valerio, C. and Slater, J. E., Endotoxin content of standardized
allergen vaccines. J. Allergy Clin. Immunol. 2003. 111: 777–783.
17 Williams, L. K., Ownby, D. R., Maliarik, M. J. and Johnson, C. C., The role
of endotoxin and its receptors in allergic disease. Ann. Allergy Asthma
Immunol. 2005. 94: 323–332.
18 Suck, R. and Cromwell, O., Quality control of recombinant pollen aller-
gens. Curr. Pharm. Anal. 2006. 2: 249–257.
19 Araujo, M. I., Campos, R. A., Cardoso, L. S., Oliveira, S. C. and Carvalho,
E. M., Immunomodulation of the allergic inflammatory response: new
developments. Inflamm. Allergy Drug Targets 2010. 9: 73–82.
20 Mittag, D., Scholzen, A., Varese, N., Baxter, L., Paukovics, G., Harrison,
L. C., Rolland, J. M. et al., The effector T cell response to ryegrass pollen
is counterregulated by simultaneous induction of regulatory T cells. J.
Immunol. 2010. 184: 4708–4716.
21 O’Hehir, R. E., Gardner, L. M., de Leon, M. P., Hales, B. J., Biondo,
M., Douglass, J. A., Rolland, J. M. et al., House dust mite sublin-
gual immunotherapy: the role for transforming growth factor-beta and
functional regulatory T cells. Am. J. Respir Crit. Care Med. 2009. 180:
936–947.
22 Groux, H., O’Garra, A., Bigler, M., Rouleau, M., Antonenko, S., de
Vries, J. E. and Roncarolo, M. G., A CD4+ T-cell subset inhibits
antigen-specific T-cell responses and prevents colitis. Nature 1997. 389:
737–742.
23 Hawrylowicz, C. M. and O’Garra, A., Potential role of interleukin-10-
secreting regulatory T cells in allergy and asthma. Nat. Rev. Immunol.
2005. 5: 271–283.
24 Matricardi, P. M., Bjorksten, B., Bonini, S., Bousquet, J., Djukanovic, R.,
Dreborg, S., Gereda, J. et al., Microbial products in allergy prevention and
therapy. Allergy 2003. 58: 461–471.
25 Wang, Y. and Wills-Karp, M., The potential role of interleukin-17 in
severe asthma. Curr. Allergy Asthma Rep. 2011. 11: 388–394.
26 Maione, F., Paschalidis, N., Mascolo, N., Dufton, N., Perretti, M. and
D’Aquisto, F., Interleukin-17 sustains rather than induces inflammation.
Biochem. Pharmacol. 2008. 77: 878–887.
27 Jankovic, D., Kullberg, M. C., Feng, C. G., Goldszmid, R. S., Collazo, C. M.,
Wilson, M., Wynn, T. A. et al., Conventional T-bet(+)Foxp3(-) Th1 cells
are the major source of host-protective regulatory IL-10 during intracel-
lular protozoan infection. J. Exp. Med. 2007. 204: 273–283.
28 Duan, W., Takanori, S. and Croft, M., Antagonism of airway tolerance by
endotoxin/lipopolysaccharide through promoting OX40L and suppress-
ing antigen-specific Foxp3+ T regulatory cells. J. Immunol. 2008. 181: 8650–
8659.
29 Re, F. and Strominger, J., Toll-like receptor 2 (TLR2) and TLR4 differen-
tially activate human dendritic cells. J. Biol. Chem. 2001. 276: 37692–37699.
30 Ferwerda, G., Meyer-Wentrup, F., Kullberg, B. J., Netea, M. G. and Adema,
G. J., Dectin-1 synergises with TLR2 and TLR4 for cytokine production in
human primary monocytes and macrophages. Cell Microbiol. 2008. 10:
2058–2066.
31 Tesse, R., Pandey, R. C. and Kabesch, M., Genetic variations in toll-like
receptor pathway genes influence asthma and atopy. Allergy 2010. 66:
307–316.
32 de Leon, M. P., Glaspole, I. N., Drew, A. C., Rolland, J. M., O’Hehir, R. E.
and Suphioglu, C., Immunological analysis of allergenic cross-reactivity
between peanut and tree nuts. Clin. Exp. Allergy 2003. 33: 1273–1280.
33 Maecker, H. T., Moon, J., Bhatia, S., Ghanekar, S. A., Maino, V. C., Payne,
J. K., Kuus-Reichel, K. et al., Impact of cryopreservation on tetramer,
cytokine flow cytometry, and ELISPOT. BMC Immunol. 2005. 6: 17.
34 Hume, D. A., Underhill, D. M., Sweet, M. J., Ozinsky, A. O., Liew, F. Y. and
Aderem, A., Macrophages exposed continuously to lipopolysaccharide
and other agonists that act vial toll-like receptors exhibit a sustained
and additive activation state. BMC Immunol. 2001. 2: 11–22.
35 Mansell, A., Smith, R., Doyle, S. L., Gray, P., Fenner, J. E., Crack, P. J.,
Nicholson, S. E. et al., Suppressor of cytokine signaling 1 negatively reg-
ulates Toll-like receptor signaling by mediating Mal degradation. Nat.
Immunol. 2006. 7: 148–155.
36 Verstak, B., Nagpal, K., Bottomley, S. P., Golenbock, D. T., Hertzog, P. J.
and Mansell, A., MyD88 adapter-like (Mal)/TIRAP interaction with TRAF6
is critical for TLR2- and TLR4-mediated NF-kappaB proinflammatory
responses. J. Biol. Chem. 2009. 284: 24192–24203.
Abbreviations: MW: molecular weight · RGP: ryegrass pollen · RGPE: rye-
grass pollen extract
Full correspondence: Dr. Diana Mittag, Transfusion Research,Australian Red Cross Blood Services, 100-154 Batman St, WestMelbourne 3003, AustraliaFax: +61-3-96940-0244e-mail: [email protected]
Received: 14/6/2012Revised: 16/11/2012Accepted: 7/12/2012Accepted article online: 13/12/2012
C© 2013 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu




















