
Detrimental role of prolonged sleep deprivation on adult neurogenesis

TLQP-21, a neuroendocrine VGF-derived peptide, preventscerebellar granule cells death induced by serum and potassiumdeprivation
Cinzia Severini,* Maria Teresa Ciotti,* Laura Biondini,* Stefania Quaresima,*Anna Maria Rinaldi,� Andrea Levi,* Claudio Frank� and Roberta Possenti*,�
*Institute of Neurobiology and Molecular Medicine, CNR, Rome, Italy
�Department of Neuroscience, University of Rome ‘‘Tor Vergata’’, Rome, Italy
�Istituto Superiore di Sanita, Viale Regina Elena, Rome, Italy
Vgf, originally identified as a nerve growth factor responsivegene (Levi et al. 1985; Salton 1991), is expressed in neuronswithin the central and peripheral nervous system and invarious endocrine cells. In the rat brain, VGF is abundant inthe cortex, hypothalamus, hippocampus, cerebellum andolfactory system and present in other brain areas in differentamounts (reviewed in Salton et al. 2000; Levi et al. 2004).
Vgf encodes for a 617 amino acid protein in rat and mouse,and 615 in human. Specific prohormone convertases PC1/3and PC2 cleave the precursor protein bearing a number ofpeptides that are stored in dense core granules and secretedthrough the regulated pathway (Trani et al. 2002; Alder et al.2003). More than 10 different VGF peptides have beendetected in rat brain, bovine hypophysis, and human CSF(Liu et al. 1994; Stark et al. 2001; Trani et al. 2002; Carretteet al. 2003). In the rat cerebellum, the expression pattern ofVGF mRNA levels were measured during the post-natalperiod of morphological and functional development both in
developing and adult animals (Salton 1991; Snyder andSalton 1998; Snyder et al. 1998), and the processing of VGFprotein to smaller polypeptides deriving from the C-terminal
Received June 12, 2007; revised manuscript received August 2, 2007;accepted September 14, 2007.Address correspondence and reprint requests to Dr Cinzia Severini,
CNR, Institute of Neurobiology and Molecular Medicine, CNR, Via delFosso di Fiorano, 65, 00143 Rome, Italy.E-mail: [email protected] used: [Ca2+]i, intracellular calcium concentration; Act-
D, actynomicine D; Akt, serine/threonine protein kinase; BME, basalEagle’s medium; CGCs, cerebellar granule cells; DIV, days in vitro; Erk,extracellular signal-regulated kinase; IGF-1, insulin-like growth factor-1;JNK, c-jun N-terminal kinase; MAPs, microtubule-associated proteins;MEK, MAPK ERK kinase; PAGE, polyacrylamide gel electrophoresis;PBS, phosphate-buffered saline; PKC, protein kinase C; S-K5, serumand potassium deprivation; UO126, 1-4-diamino-2-3-dicyano-1-4-bis(2-aminophynyltio) butadiene.
Abstract
Different VGF peptides derived from Vgf, originally identified
as a nerve growth factor responsive gene, have been
detected in neurons within the central and peripheral nervous
system and in various endocrine cells. In the current study, we
have evaluated the ability of TLQP-21, a VGF-derived pep-
tide, to protect, in a dose- and time-dependent manner, pri-
mary cultures of rat cerebellar granule cells (CGCs) from
serum and potassium deprivation-induced cell death. We
demonstrated that TLQP-21 increased survival of CGCs
by decreasing the degree of apoptosis as assessed by cell
viability and DNA fragmentation. Moreover, TLQP-21 signifi-
cantly activated extracellular signal-regulated kinase 1/2,
serine/threonine protein kinase, and c-jun N-terminal kinase
phosphorylation, while decreased the extent of protein kinase
C phosphorylation, as demonstrated by western blot analysis.
In addition, TLQP-21 induced significant increase in intra-
cellular calcium (as measured by fura-2AM) in about 60% of
the recorded neurons. Taken together, the present results
demonstrate that TLQP-21 promotes the survival of CGCs via
pathways involving, within few minutes, modulation of kinases
associated with CGCs survival, and by increasing intracellular
calcium which can contribute to the neuroprotective effect of
the peptide.
Keywords: cerebellar granule cells, neuronal death, TLQP-
21, Vgf.
J. Neurochem. (2008) 104, 534–544.
d JOURNAL OF NEUROCHEMISTRY | 2008 | 104 | 534–544 doi: 10.1111/j.1471-4159.2007.05068.x
534 Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 534–544� 2007 The Authors

region was previously investigated in rat cerebellar granulecells (CGCs) (Trani et al. 1995).
Recently, we identified and characterized in rat brainextracts a new VGF-derived peptide designated as TLQP-21,which spans from residue 556 to residue 576 of the precursorsequence. This peptide has been reported to be one of themajor peptide fragments present in the brain of the rat(Bartolomucci et al. 2006) and to exert biological activityboth in vivo (Bartolomucci et al. 2006; Jethwa et al. 2007)and in vitro (inducing contraction of gastric fundus strips,unpublished work).
In vitro cultures of newborn rat CGCs represent one ofthe most widely used experimental model to study neuronalcell death because of the high degree of cellular homo-geneity, allowing the study of the effect of substances thatcan modulate this process. CGCs survive and differentiatein vitro in the presence of depolarizing concentrations ofKCl (25 mmol/L), but undergo apoptosis when cultured inphysiological low KCl (5 mmol/L) conditions (Gallo et al.1987; D’Mello et al. 1993). The mechanism of action ofdepolarizing concentrations of KCl remains controversial.Generally, it is believed that depolarization involves a lowincrease in intracellular Ca2+ concentration (D’Mello et al.1993; Galli et al. 1995) and MAPK activation (Rosen et al.1994).
In view of the finding reporting the physiological presenceof small polypeptides deriving from processing of theC-terminal region of VGF protein in CGCs, we investigatedwhether TLQP-21 can regulate the viability of these neurons.
Our results demonstrate that: (i) TLQP-21 protects CGCsfrom apoptosis induced by serum and potassium withdrawalconditions, (ii) neuroprotective action of TLQP-21 is depen-dent on the modulation of extracellular signal-regulatedkinase (Erk) 1/2, serine/threonine protein kinase (Akt),protein kinase C (PKC) and c-jun N-terminal kinase (JNK)kinases, and (iii) neuroprotective action is mediated byintracellular calcium increase.
Materials and methods
ChemicalsRabbit polyclonal antibodies (anti-phospho-Akt), anti-Akt, anti-
phospho-Erk1/2, anti-Erk1/2, anti-PKC, anti-JNK, and anti-phos-
pho-JNK, were from Cell Signaling technology (Beverly, MA,
USA). Donkey anti-rabbit peroxidase-coupled secondary antibodies
were from Amersham-GE Healthcare (Arlington Heights, IL, USA).
Monoclonal antibodies against microtubule-associated proteins
(MAPs) from Chemicon (Temecula, CA, USA) and Tetramethyl
Rhodamine Iso-Thiocyanate conjugated secondary antibodies from
Sigma (St Louis, MO, USA).
TLQP-21 (TLQPPASSRRRHFHHALPPAR) and the scrambled
control peptide Scr-21 (LRPSHTRPAHQSFARPLHRPA) were
synthesized by PRIMM (Milan, Italy). The other reagents were
from Sigma Aldrich.
Primary cultures of cerebellar granule cellsThe regional animal Ethics committee approved the experiments.
Cultures enriched in granule neurons were obtained from dissociated
cerebella of 8-day-old Wistar rats (P8) as previously described by
Levi et al. (1984). Cells were plated in basal Eagle’s medium
(BME; Gibco, Milan, Italy) supplemented with 10% fetal calf serum
(Gibco), 25 mmol/L KCl, 2 mmol/L glutamine (Gibco) and 100 lg/mL gentamycin (Gibco) in 24-well clusters, (NUNC, Roskilde,
Denmark), previously coated with poly-L-Lysine. CGCs were plated
at a constant density of 0.5 · 106 cells/well in BME.
1b-Arabinofuranosylcytosine (10 lmol/L) was added to the
culture medium 18–20 h after plating to prevent proliferation of
non-neuronal cells. After 6 days ‘in vitro’ (DIV), extracellular KClwas lowered from 25 to 5 mmol/L for neuronal cell death.
Cultured neurons were washed (2X) with serum-free BME
containing 5 mmol/L KCl incubated in the same medium up to
48 h (D’Mello et al. 1993). Neuronal viability was assessed by
counting the number of intact nuclei according to the method
described by Volonte et al. (1994). The culture medium was
removed and replaced with 0.5 mL of a detergent containing
lysing solution (0.5% ethylhexadecyldimethylammonium bromide,
0.28% acetic acid, 0.5% Triton X-100, 3 mmol/L NaCl, 2 mmol/L
MgCl2, in phosphate-buffered saline (PBS), pH 7.4 diluted 1/10).
After 2 min, the cells were collected and the solution consisted of
a uniform suspension of single intact and nuclei that were then
quantified by counting in a hemocytometer (Fortuna, Wertheim,
Germany). Broken or damaged nuclei were not included in the
counts. Alternatively, CGCs were fixed in 4% paraformaldehyde
and permeabilized with 0.2% Triton X-100 in Tris–HCl 0.1 mol/L,
pH 7.4 for 5¢ and then incubated with Hoechst 33258
(0.25 lg/mL) for 5 min at 20�C. After washing with PBS, the
percentage of shrunken and condensed nuclei was assessed.
To verify the effect of TLQP-21 on neuronal cell death, at 7 or 6
DIV CGCs were incubated for 24 or 48 h, respectively, with the
peptide tested and simultaneously exposed to KCl 5 mmol/L.
Counting intact nuclei as previously described assessed cell
viability.
ImmunocytochemistryCerebellar granule cells were fixed and permeabilized with 3.5%
formaldehyde, 70% ethanol and 5% acetic acid for 15 min at 20�Cand fixed with 4% (w/v in PBS) paraformaldehyde for 15 min at
20�C. Fixed cells were washed in PBS, pH 7.4, and permeabilized
with 0.2% Triton X-100-Tris–HCl, pH 7.4, for 5 min. The
coverslips were treated with a monoclonal antibody against MAPs
1 : 200 overnight at 4�C in a moist chamber, rinsed in PBS, and
stained with Tetramethyl Rhodamine Iso-Thiocyanate conjugated
secondary antibodies for 30 min at 20�C.To visualize nuclear morphology, cells were incubated for 5 min
with 1 lg/mL of the DNA dye Hoechst 33258 and then visualized
by a Laica fluorescent photomicroscope (Wetzlar, Germany).
Cell death ELISA assayTo detect the enrichment of histone-associated DNA fragments
(mononucleosomes and oligonucleosomes) in cytoplasmic fractions
of CGCs, the samples were processed and analyzed with the Cell
Death Detection ELISAPLUS kit (Roche, Mannheim, Germany).
After plating at the density of 0.5 · 106 cells/well in 24-well clusters,
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 534–544
TLQP-21 neuroprotection of cerebellar granule cells | 535
CGCs were treated with TLQP-21 (1 lmol/L) or insulin-like
growth factor-1 (IGF-1) (25 ng/mL) and simultaneously exposed
to KCl 5 mmol/L. After 12 h, cells were analyzed into 96-well plates
at 1 · 103 cells/well density, according to the kit instructions. The
absorbance was measured with a Victor 3V 1420 multilabel counter
(Perkin Elmer, Turku, Finland). The bound anti-DNA-peroxidase
was quantified by using the substrate 2,2¢-azino-di-(3-ethyl) benz-isothiazoline sulfonate measured at an absorbance of 405 nm.
Background absorbance values, determined by measuring samples
of cell lysates and reaction buffer, were subtracted from the
sample absorbance values. The enrichment of histone-associated
DNA fragments was calculated considering the values found in
K25-treated cells as one.
Western blottingFor whole-cell lysates, the culture medium was discarded, the
neurons washed twice with ice-cold PBS, and lysed in lysis buffer
[50 mmol/L Tris–HCl (pH 7.5), 150 mmol/L NaCl, 10 mmol/L
EDTA, 10 mmol/L EGTA, 50 lmol/L NaF, 2.5 lmol/L sodium
pyrophosphate, 1 lmol/L beta-glycerophosphate, 1 lmol/L
Na3VO4, 1X protease inhibitor mixture, Sigma]. Equivalent
amounts of cell extracts (5 · 105 cells) were mixed with sodium
dodecyl sulfate-reducing sample buffer according to Invitrogen Nu
polyacrylamide gel electrophoresis (PAGE) (Invitrogen, Carlsbad,
CA, USA). Following heating at 70�C for 10 min, proteins were
subjected to sodium dodecyl sulfate–PAGE (Nu PAGE) and
transferred electrophoretically to polyvinylidene difluoride mem-
brane (Amersham). After staining with Ponceau S to verify
uniformity of protein loads/transfer, the membranes were analyzed
for immunoreactivity. Incubation with primary antibodies was
performed overnight at 4�C (rabbit anti-Erk1/2/p-Erk1/2, anti-Akt/
p-Akt, anti-JNK/p-JNK, p-a-PKC, 1 : 1000) and then membranes
were incubated with donkey anti-rabbit peroxidase-coupled second-
ary antibodies (1 : 5000) for 1 h at 20�C. Immunoreactivity was
developed by enhanced chemiluminescence system (Amersham). To
normalize loaded sample, the blots were stripped (Restore western
blot Stripping; Pierce, Rockford, IL, USA) and reprobed with an
antibody against a-tubulin (Sigma, 1/10.000, overnight at 4�C). Foranalysis of the western blotting data, densitometric analysis was
performed using Scan Analysis software for Mc Intosh (Biosoft,
Ferguson, MO, USA).
Fura-2AM experimentsOptical fluorimetric recordings with fura-2AM were used to
evaluate the intracellular calcium concentration ([Ca2+]i). Fura-
2AM stock solutions were obtained by adding 50 lg of fura-2AM to
50 lL of 75% dimethylsulfoxide plus 25% pluronic acid. Cells were
bathed for 60 min at 20�C in 1 mL of extracellular solution
(in mmol/L: 125 NaCl, one KCl, five CaCl2, one MgCl2, eight
glucose, and 20 HEPES, pH 7.35) with a final fura concentration of
5 lmol/L. This solution was then removed and replaced with
extracellular solution, and the dishes were quickly placed on the
microscope stage. To measure fluorescence changes, an Hamamatsu
(Shizouka, Japan) Argus 50 computerized analysis system was
used, recording every 6 s the ratio between the values of light
intensity at 340 and 380 nm stimulation (Frank et al. 2004). Thisprogram allowed analyzing all the selected cells as digital data,
subsequently used for statistical analysis.
Data analysisAnalyses of data from treated cultures and the corresponding control
condition were performed by one-way ANOVA for repeated measures
followed byDunnett’s test formultiple comparisons. Data are reported
as mean ± SEM, n being the number of independent experiments.
The significance level was set at p < 0.05 (*) and p < 0.01 (**).
Results
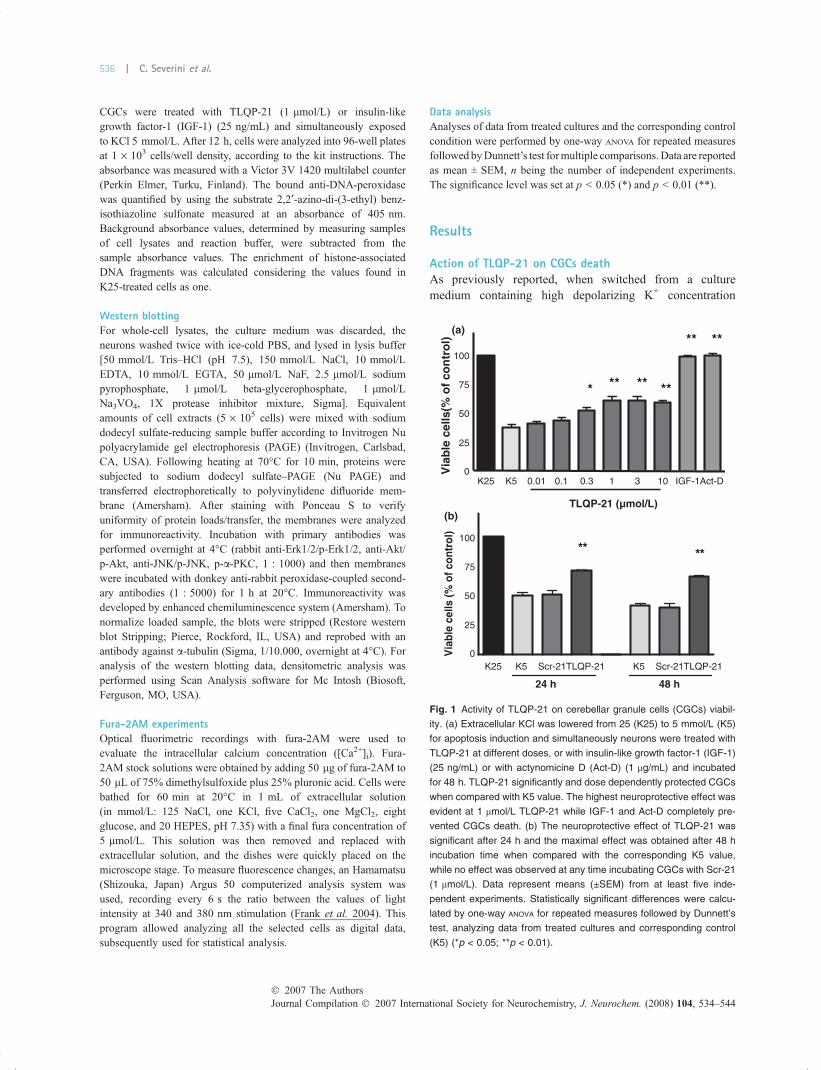
Action of TLQP-21 on CGCs deathAs previously reported, when switched from a culturemedium containing high depolarizing K+ concentration
12-rcS5K52K T 12-rcS5K12-PQL T 12-PQL
48 h24 h
** **
Via
ble
cells
(% o
f con
trol
)
(a)
(b)
0.01 0.1 0.3 1 3 10 IGF-1Act-DK5K250
100
75
50
25
0
100
75
50
25
* ****
****
**
TLQP-21 (µmol/L)
Via
ble
cel
ls(%
of
con
tro
l)
Fig. 1 Activity of TLQP-21 on cerebellar granule cells (CGCs) viabil-
ity. (a) Extracellular KCl was lowered from 25 (K25) to 5 mmol/L (K5)
for apoptosis induction and simultaneously neurons were treated with
TLQP-21 at different doses, or with insulin-like growth factor-1 (IGF-1)
(25 ng/mL) or with actynomicine D (Act-D) (1 lg/mL) and incubated
for 48 h. TLQP-21 significantly and dose dependently protected CGCs
when compared with K5 value. The highest neuroprotective effect was
evident at 1 lmol/L TLQP-21 while IGF-1 and Act-D completely pre-
vented CGCs death. (b) The neuroprotective effect of TLQP-21 was
significant after 24 h and the maximal effect was obtained after 48 h
incubation time when compared with the corresponding K5 value,
while no effect was observed at any time incubating CGCs with Scr-21
(1 lmol/L). Data represent means (±SEM) from at least five inde-
pendent experiments. Statistically significant differences were calcu-
lated by one-way ANOVA for repeated measures followed by Dunnett’s
test, analyzing data from treated cultures and corresponding control
(K5) (*p < 0.05; **p < 0.01).
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 534–544� 2007 The Authors
536 | C. Severini et al.

(25 mmol/L) to one containing a low even though morephysiological K+ concentration (5 mmol/L), CGCs degener-ate and die, prevalently by apoptosis (D’Mello et al. 1993).A dose–response curve was carried out following 48 hincubation with TLQP-21. As shown in Fig. 1a, treatmentwith 0.01, 0.1, 0.3, 1, 3, and 10 lmol/L TLQP-21 caused anincrease in cell viability of 3.2 ± 2.3%, 6.3 ± 2.7%, 14.4 ±3.0%, 23.5 ± 2.9%, 22.8 ± 3.4% and 21.4 ± 1.6%, respec-tively. Thus, the concentration of 1 lmol/L was used in allsubsequent experiments. No effect was observed using thescrambled peptide (Scr-21) at concentrations up to 10 lmol/L (data not shown). As positive controls, we used two well-established protective factors for CGCs following K+
withdrawal, IGF-1 and actynomicine D (Act-D) (D’Melloet al. 1993; Galli et al. 1995). A complete preservation ofcell viability occurred in the presence of both IGF-1(63 ± 1.3% increase at 25 ng/mL) and Act-D (60 ± 1.8%increase at 1 lg/mL).
Moreover, time-dependent response effects, incubatingCGCs with 1 lmol/L TLQP-21 simultaneously the apoptosisinduction, showed that the neuroprotective effect wassignificant after 24 h of treatment in K5 conditions (19.6 ±3.1%), while a maximal action was obtained after 48 h(24.7 ± 2.7%). On the contrary, no effect was observed usingScr-21 at the same concentration (Fig. 1b).
HoechstsPAM
52K
5K
12-PQLT+
1-FGI+
5K
5K
Fig. 2 TLQP-21 neuroprotective activity.
Cerebellar granule cells (CGCs) were
incubated with TLQP-21 (1 lmol/L) or
insulin-like growth factor-1 (IGF-1) (25 ng/
mL) and simultaneously exposed to KCl
5 mmol/L. On the left, CGCs morphological
analysis assessed upon 48 h of apoptosis
by immunofluorescence using an antibody
against the microtubule-associated proteins
(MAPs). On the right, Hoechst 33258
staining showing intact and condensed
apoptotic nuclei (2.6X magnification). Val-
ues corresponding to apoptotic nuclei as
percentages of total nuclei were: K25: 11%
(16/142), K5: 67% (98/154), K5 + TLQP-
21: 34% (55/160), K5 + IGF-1: 18% (19/
105). Scale bar: 25 lm.
D-tcA1-FGI12-PQLT5K52K
0
1
2
3
4
5
6
7
8
9
**
*
**En
rich
men
t o
f D
NA
frag
men
tati
on
5K
Fig. 3 Effect of TLQP-21 on cerebellar granule cells (CGCs) DNA
fragmentation. CGCs were treated with TLQP-21 (1 lmol/L) or insulin-
like growth factor-1 (IGF-1) (25 ng/mL) or actynomicine D (Act-D)
(1 lg/mL) and simultaneously exposed to KCl 5 mmol/L. DNA frag-
mentation was determined 12 h after the addition of the drugs and
detected by the ELISA method. Data were expressed as relative DNA
fragmentation enrichment. Data represent means (±SEM) from six
independent experiments. Statistically significant differences were
calculated by one-way ANOVA for repeated measures followed by
Dunnett’s test, analyzing data from treated cultures and corresponding
controls (K5) (*p < 0.05; **p < 0.01).
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 534–544
TLQP-21 neuroprotection of cerebellar granule cells | 537

Morphological analysis, by immunofluorescence methodusing an antibody against the MAPs, indicated that in S-K5conditions CGCs resulted severely damaged and showedmorphological alterations associated with apoptosis, dis-playing severe neuronal soma shrinkage and axonalretraction. On the contrary, CGCs incubated for 48 h withTLQP-21 or IGF-1 showed a sustained neurites network(Fig. 2).
Reported data were also confirmed by cell viabilityquantification by Hoechst staining, indicating the samepercentage obtained counting intact nuclei (Fig. 2).
Effect of TLQP-21 on internucleosomal DNAfragmentationNeuronal death is known to be accompanied by the charac-teristic features of apoptosis, such as DNA fragmentation(Nagata et al. 2003). To determine whether a TLQP-21neuroprotective effect is attributable to an inhibition ofapoptosis, the levels of DNA fragmentation were mea-
sured by ELISA. As shown in Fig. 3, the DNA fragmen-tation levels were drastically increased in K5 samples.At 1 lmol/L, TLQP-21 showed about 23.4% rate ofinhibition against K5-induced DNA fragmentation, whileboth IGF-1 (25 ng/mL) and Act-D (1 lg/mL) com-pletely inhibited the DNA fragmentation induced by K5conditions.
Neuroprotection by TLQP-21 was MAPK/Erk1/2 and AktdependentAs MAPK/Erk1/2 and Akt signaling pathways are two well-studied survival-promoting signaling mechanisms in neurons(Gunn-Moore et al. 1997; Bonni et al. 1999; Yuan andYankner 2000), we investigated the involvement of thesekinases in the neuroprotective effect of TLQP-21.
We used as positive control IGF-1, the most potent anti-apoptotic factor for CGCs by modulating the MAPK/Erk1/2and especially the Akt pathway (D’Mello et al. 1997; Dattaet al. 1997; Dudek et al. 1997).
12-rcS(a)′06′03′51′5C
1-FGI
2/1krE-p
12-PQLT
2/1krE
′06′03′51′5C
Protein signal intensity Protein signal intensity
6030155K5 6030155K50123456
* * *
TLQP-21 1 µmol/L IGF (25ng/mL)
P-E
rk 1
/2 le
vel
P-E
rk 1
/2 le
vel
012345
**
**
****
**
2/1krE-p
2/1krE
48 h24 h12-PQLT1FGI5K52K 12-PQLT1FGI5K52K
(b)
nilubTu
′06′03′51′5C
2/1krE-p
5K52K
h6h4h3h2
2/1krE-p
(min incubation time) (min incubation time)
Fig. 4 Action of TLQP-21 on extracellular signal-regulated kinase
(Erk) 1/2 phosphorylation during cerebellar granule cells (CGCs) death
induced by potassium and serum deprivation. CGCs were switched in
S-K5 for different times. Cells were lysed and processed for immu-
noblotting. Western blot analyses were performed using an antibody
specific for the phosphorylated (active) forms of Erk1/2 (p-Erk).
(a) CGCs were treated, in K5 conditions, with TLQP-21 (1 lmol/L) or
insulin-like growth factor-1 (IGF-1) (25 ng/mL) for different times (from
5 to 60 min). The same blots were stripped and reprobed with an
antibody against total Erk1/2 (Erk). The action of Scr-21 (1 lmol/L) on
Erk1/2 phosphorylation after different incubation times is also shown.
Quantitative analysis of Erk1/2 phosphorylation is depicted below the
respective blots and was determined by band densitometric analysis.
The densitometric values from phosphorylated Erk1/2 were normal-
ized using total Erk1/2 as internal controls. The ratio of the band
intensities was calculated considering the values found in K5 (C)
treated cells as one. Data represent means (±SEM) from four inde-
pendent experiments. Statistically significant differences were calcu-
lated by one-way ANOVA for repeated measures followed by Dunnett’s
test, analyzing data from treated cultures and corresponding controls
(K5) (*p < 0.05; **p < 0.01). (b) Representative western blots from at
least three experiments showing Erk1/2 phosphorylation of CGCs
treated with TLQP-21 (1 lmol/L) or IGF-1 (25 ng/mL) in K5 conditions
for 24 and 48 h. The same blots were stripped and reprobed with an
antibody against a-tubulin to assess the amount of proteins present in
each lane (lower panel).
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 534–544� 2007 The Authors
538 | C. Severini et al.

Cerebellar granule cells were treated for different timeswith K5 or K5 medium containing TLQP-21 (1 lmol/L)or IGF-1 (25 ng/mL). Quantitative analysis of Erk1/2 andAkt phosphorylation of different experiments was deter-mined by densitometric analysis and the densitometricvalues from phosphorylated Erk1/2 and Akt were normal-ized using total Erk1/2 and Akt as internal controls. Theratio of the band intensities was calculated considering thevalues found in K5-treated cells as one. Preliminaryexperiments were carried out to evaluate the mostfavorable time at which such signal transduction pathwayswere activated without modification in the phosphorylationbecause of K5 conditions. As a significant increase inErk1/2 phosphorylation can be detected from 3 h ofstarvation time, earlier times of incubation were used(Fig. 4a).
As shown in Fig. 4a, TLQP-21 significantly increasedlevels of phosphorylated Erk1/2 and this effect was remark-able within 15 min after the peptide administration. The
same result was obtained using IGF-1 and the highestphosphorylation was found at the same time (15 min). Inaddition, using Scr-21 at 1 lmol/L concentration, no effectwas observed at any time.
Relative high amounts of phosphorylated Erk1/2 wererevealed after 24 and 48 h but these values correlate withcorresponding high levels of total MAPKs and a-tubulin,because of an increase of viable cells (Fig. 4b).
To confirm the involvement of MAPK-Erk1/2 pathway inthe mechanism of action of TLQP-21, CGCs were treatedwith the MAPK ERK kinase (MEK) inhibitor 1-4-diamino-2-3-dicyano-1-4-bis (2-aminophynyltio) butadiene (UO126)at concentrations of 10 lmol/L. UO126 was added at 6 DIV1 h before or simultaneously the induction of apoptosis. Asthe inhibitor showed the same effect in all cases, the firstprotocol described was used for further experiments. Asshown in Fig. 5, UO126 completely prevented the increasedErk1/2 phosphorylation exerted by both TLQP-21 andIGF-1, while treatment with MEK inhibitor alone did notreduce the intensity of p-Erk immunolabeling, when com-pared with K5 culture values.
Additionally, as shown in Fig. 5, both 5 and 10 lmol/L ofUO126 completely prevented the neuroprotective activityexerted by TLQP21 1 lmol/L in K5 conditions, while aweak effect was observed on neuronal viability treatingCGCs with UO126 alone. Conversely, the same inhibitor wasunable to antagonize the neuroprotection carried out by IGF-1. To assess the effect of the MEK inhibitor on healthyneurons, equivalent experiments were carried out in K25conditions showing that the drug did not influence survival ofCGCs at different doses.
Similarly, Akt phosphorylation was found to be increasedafter 15 min of treatment with TLQP-21, while phosphory-lation of Akt increased further after that time using IGF-1(Fig. 6a). After 24 and 48 h of incubation, IGF-1 treatmentincreased Akt phosphorylation while TLQP-21 did notmodify the amount of phosphorylated Akt, although totalAkt and a-tubulin were found to be increased in both cases(Fig. 6b).
Neuroprotection by TLQP-21 was protein kinase C and JNKdependentProtein kinase C is known to play a role in various neuronalevents, including differentiation and neuroprotection (Singhet al. 1994; Zirrgiebel et al. 1995).
We performed western blots using cell lysates obtainedfrom various time points after K+ deprivation and probedthem with a phospho-specific PKC or a phospho-specific JNKantibody. Quantitative analysis of PKC phosphorylation ofdifferent experiments was determined by densitometry anal-ysis and the densitometric values from phosphorylated PKCand JNK were normalized using a-tubulin as internal control.The ratio of the band intensities was calculated consideringthe values found in K5-treated cells as one.
2/1KRE-p621OU+
K25
U01
26 5
U01
26 1
0
K5
U01
26 5
U01
26 1
0
K5
U01
26 5
U01
26 1
0
K5
U01
26 5
U01
26 1
00
100
75
50
25
12-PQLT 1-FGI
***
**
Via
ble
cel
ls (
% o
f co
ntr
ol)
′06′03′51′5C
1-FGI′06′03′51′5C
12-PQLT
Fig. 5 Effect of 1-4-diamino-2-3-dicyano-1-4-bis (2-aminophynyltio)
butadiene (UO126). Representative western blots from at least three
experiments showing extracellular signal-regulated kinase (Erk) 1/2
phosphorylation of cerebellar granule cells (CGCs) in K5 conditions
treated with TLQP-21 (1 lmol/L) or insulin-like growth factor-1 (IGF-1)
(25 ng/mL) in the presence of U0126 (10 lmol/L), for different times
(from 5 to 60 min) (upper panels). CGCs in K25 and K5 conditions
were treated with UO126, a specific MAPK kinases inhibitor, at dif-
ferent doses (5 and 10 lmol/L) alone or in the presence of TLQP-21
(1 lmol/L) or IGF-1 (25 ng/mL) and incubated for 48 h. U0126 treat-
ment induced a complete inhibitory effect on the TLQP-21-promoted
neuroprotection at the concentration of 10 lmol/L while this inhibitor
did not affect IGF-1 neuroprotection (lower panel). Data represent
means (±SEM) from four independent experiments. Statistically sig-
nificant differences were calculated by one-way ANOVA for repeated
measures followed by Dunnett’s test, analyzing data from treated
cultures and corresponding control (K5) (*p < 0.05; **p < 0.01).
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 534–544
TLQP-21 neuroprotection of cerebellar granule cells | 539

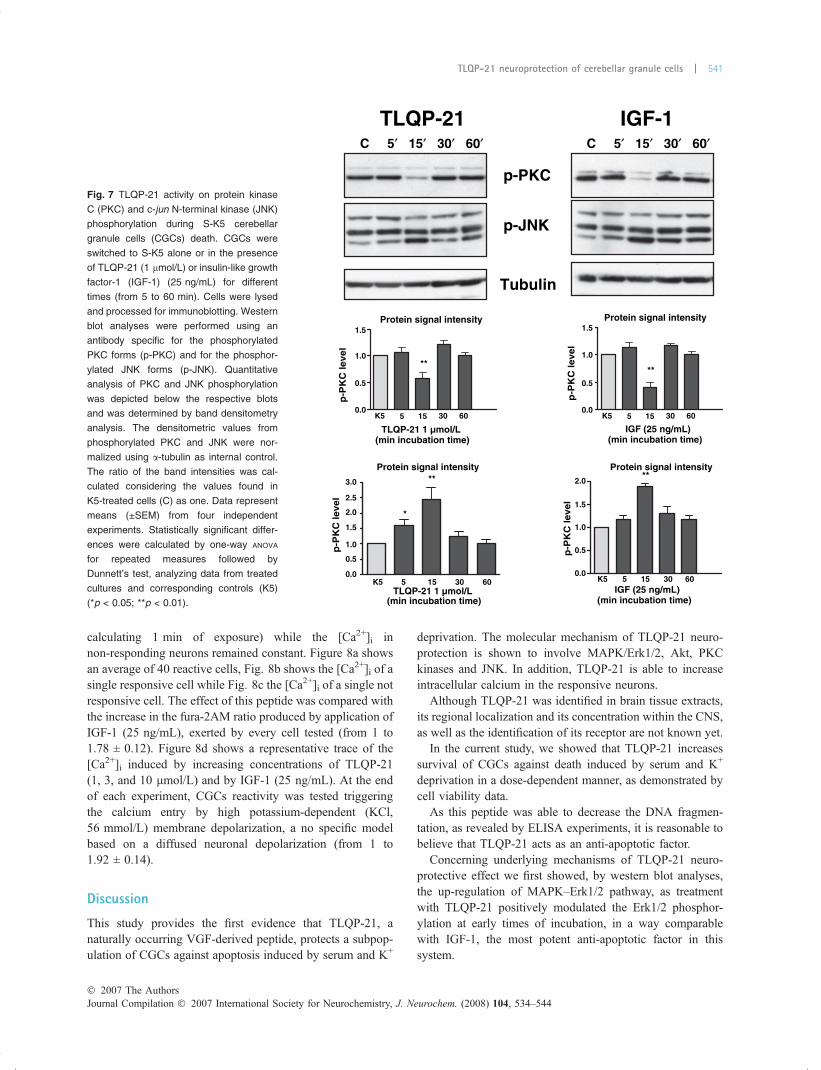
As previously demonstrated, different JNK isoformscoexist in CGCs, the phosphorylation of some of which isreportedly involved in apoptosis of these neurons (Coffeyet al. 2002).
Four major isoforms were revealed in K5 conditions bythe use of the antibody against total JNK (Cell Signaling,data not shown), and using the antibody against phospho-JNK (Cell Signaling), three phosphorylated isoforms of 54–46 and 44 kDa were showed (Fig. 7).
As shown in Fig. 7, western blot analysis indicated that intime-course experiments, a significant but transient decreasein PKC phosphorylation was obtained after 15 min, timecorresponding to the maximal activity of both Erk1/2 andAkt. Similar results were obtained using IGF-1.
Conversely, TLQP-21 treatment induced a significantincrease of one isoform of phospho-JNK (the lower bandcorresponding to 44 kDa) after 15 min, similarly to IGF-1that, however, exerted a more prolonged effect.
TLQP-21 increased intracellular calcium as measured byfura-2AMTo investigate whether neuroprotective effect of TLQP-21was associated with an increase of intracellular calcium,we performed experiments to explore whether TLQP-21application was able to modify calcium levels in CGCs inculture.
Preliminary experiments were carried out to assess theTLQP-21 concentration able to induce a statistically signif-icant increase of [Ca2+]i, resulting 10 lmol/L the mosteffective concentration.
Ten lmol/L of TLQP-21 induced a significant increase in[Ca2+]i in a subpopulation of CGCs corresponding to about60% of the analyzed cells (value obtained analyzing morethan 150 cells), while Scr-21, applied at the same concen-tration was fully inactive, as shown in Fig. 8a.
TLQP-21 induced, in responsive neurons a marked rise inthe fura-2AM ratio, from 1 to 1.42 ± 0.08 (value obtained
tkA-p
tkA
12-PQLT
′06′03′51′5C′06′03′51′5C
1-FGI
tkA-p
tkA
48 h24 h12-PQLT1-FGI5K52K 12-PQLT1-FGI5K52K
Protein signal intensity Protein signal intensity
6030155K5 6030155K50 0.0
2.5
5.0
7.5
1
2
3
TLQP-21 1 µmol/L IGF (min incubation time)
P-A
kt 1
/2 le
vel
P-A
kt 1
/2 le
vel
*
**
**
**
** **
nilubTu
(a)
(b)
(min incubation time)
Fig. 6 Action of TLQP-21 on serine/threonine protein kinase (Akt)
phosphorylation during S-K5 cerebellar granule cells (CGCs) death.
(a) CGCs were switched to S-K5 alone or in the presence of TLQP-21
(1 lmol/L) or insulin-like growth factor-1 (IGF-1) (25 ng/mL) for dif-
ferent times (from 5 to 60 min). Cells were lysed and processed for
immunoblotting. Western blot analyses were performed using an
antibody specific for the phosphorylated (active) form of Akt (p-Akt)
(upper panel). The same blot was stripped and reprobed with an
antibody against total Akt (lower panel). Quantitative analysis of Akt
phosphorylation is depicted below the respective blots and was
determined by band densitometric analysis. The densitometric values
from phosphorylated Akt were normalized using total Akt as internal
controls. The ratio of the band intensities was calculated considering
the values found in K5-treated cells (C) as one. (b) Representative
western blots from at least three experiments showing Akt phos-
phorylation of CGCs treated with TLQP-21 (1 lmol/L) or IGF-1 (25 ng/
mL) in K5 conditions for 24 and 48 h. The same blots were stripped
and reprobed with an antibody against a-tubulin to assess the amount
of proteins present in each lane (lower panel). Data represent means
(±SEM) from four independent experiments. Statistically significant
differences were calculated by one-way ANOVA for repeated measures
followed by Dunnett’s test, analyzing data from treated cultures and
corresponding controls (K5) (*p < 0.05; **p < 0.01).
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 534–544� 2007 The Authors
540 | C. Severini et al.
calculating 1 min of exposure) while the [Ca2+]i innon-responding neurons remained constant. Figure 8a showsan average of 40 reactive cells, Fig. 8b shows the [Ca2+]i of asingle responsive cell while Fig. 8c the [Ca2+]i of a single notresponsive cell. The effect of this peptide was compared withthe increase in the fura-2AM ratio produced by application ofIGF-1 (25 ng/mL), exerted by every cell tested (from 1 to1.78 ± 0.12). Figure 8d shows a representative trace of the[Ca2+]i induced by increasing concentrations of TLQP-21(1, 3, and 10 lmol/L) and by IGF-1 (25 ng/mL). At the endof each experiment, CGCs reactivity was tested triggeringthe calcium entry by high potassium-dependent (KCl,56 mmol/L) membrane depolarization, a no specific modelbased on a diffused neuronal depolarization (from 1 to1.92 ± 0.14).
Discussion
This study provides the first evidence that TLQP-21, anaturally occurring VGF-derived peptide, protects a subpop-ulation of CGCs against apoptosis induced by serum and K+
deprivation. The molecular mechanism of TLQP-21 neuro-protection is shown to involve MAPK/Erk1/2, Akt, PKCkinases and JNK. In addition, TLQP-21 is able to increaseintracellular calcium in the responsive neurons.
Although TLQP-21 was identified in brain tissue extracts,its regional localization and its concentration within the CNS,as well as the identification of its receptor are not known yet.
In the current study, we showed that TLQP-21 increasessurvival of CGCs against death induced by serum and K+
deprivation in a dose-dependent manner, as demonstrated bycell viability data.
As this peptide was able to decrease the DNA fragmen-tation, as revealed by ELISA experiments, it is reasonable tobelieve that TLQP-21 acts as an anti-apoptotic factor.
Concerning underlying mechanisms of TLQP-21 neuro-protective effect we first showed, by western blot analyses,the up-regulation of MAPK–Erk1/2 pathway, as treatmentwith TLQP-21 positively modulated the Erk1/2 phosphor-ylation at early times of incubation, in a way comparablewith IGF-1, the most potent anti-apoptotic factor in thissystem.
p-P
KC
leve
l
**
**
**
**
*
nilubuT
KNJ-p
CKP-p
12-PQLT′06′03′51′5C′06′03′51′5C
1-FGI
Protein signal intensity Protein signal intensity
Protein signal intensity Protein signal intensity
6030155K5
6030155K5
6030155K5
6030155K5
0.0
0.5
1.0
1.5p
-PK
C le
vel
1.5
2.0
2.5
3.0
0.0
0.5
1.0
p-P
KC
leve
l 1.5
2.0
0.0
0.5
1.0
p-P
KC
leve
l
0.0
0.5
1.0
1.5
TLQP-21 1 µmol/L (min incubation time)
TLQP-21 1 µmol/L (min incubation time)
IGF (25 ng/mL)(min incubation time)
IGF (25 ng/mL)(min incubation time)
Fig. 7 TLQP-21 activity on protein kinase
C (PKC) and c-jun N-terminal kinase (JNK)
phosphorylation during S-K5 cerebellar
granule cells (CGCs) death. CGCs were
switched to S-K5 alone or in the presence
of TLQP-21 (1 lmol/L) or insulin-like growth
factor-1 (IGF-1) (25 ng/mL) for different
times (from 5 to 60 min). Cells were lysed
and processed for immunoblotting. Western
blot analyses were performed using an
antibody specific for the phosphorylated
PKC forms (p-PKC) and for the phosphor-
ylated JNK forms (p-JNK). Quantitative
analysis of PKC and JNK phosphorylation
was depicted below the respective blots
and was determined by band densitometry
analysis. The densitometric values from
phosphorylated PKC and JNK were nor-
malized using a-tubulin as internal control.
The ratio of the band intensities was cal-
culated considering the values found in
K5-treated cells (C) as one. Data represent
means (±SEM) from four independent
experiments. Statistically significant differ-
ences were calculated by one-way ANOVA
for repeated measures followed by
Dunnett’s test, analyzing data from treated
cultures and corresponding controls (K5)
(*p < 0.05; **p < 0.01).
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 534–544
TLQP-21 neuroprotection of cerebellar granule cells | 541

The MAPK–Erk1/2 pathway is known to be importantfor neuronal survival in different systems (Villalba et al.1997; Bonni et al. 1999; Obara et al. 2007). In particular,neurotrophin-mediated protection of neurons in vitro and invivo is mediated via the Erk pathway, resulting in phosphor-ylation of Erk1/2 and the phosphoinositide 3-kinase substrateAkt within few minutes (Han and Holtzman 2000; Willaime-Morawek et al. 2005).
It is well established that phosphorylation of Akt isassociated with CGCs survival (Gunn-Moore et al. 1997;Favata et al. 1998; Finkbeiner 2000; Zhu et al. 2004). Ithas been reported that the survival of mature CGCs afterserum and K+ withdrawal can be maintained by severaldifferent survival factors, all converging on the activation ofthe Akt (Vaillant et al. 1999; Kumari et al. 2001). Amongthese, exposure to growth factors such as IGF-1-induceactivation of Akt (D’Mello et al. 1993; Dudek et al. 1997).Using an antibody against phospho-Akt we observedin TLQP-21-treated CGCs, within 15 min, a significantincreased level of the active form of this enzyme knownto be involved in promoting the survival of differentcell types.
Protein kinase C, present in different isoforms in CGCs, isanother kinase involved in CGCs apoptosis induced by lowpotassium. In K5 conditions, levels of some PKC isoformswere markedly decreased, some not altered or even increasedduring the occurrence of death, suggesting a pivotal role for
this kinase in the maintenance of CGCs survival (Dekker andParker 1994; Lin et al. 1997).
We observed, by western blot analyses, that treatment withTLQP-21 reduced PKC phosphorylation of one isoform at15 min of incubation, time corresponding to the maximalactivity of both Erk1/2 andAkt, sharing similar qualitative andquantitative properties to the action of IGF-1. These resultsare in agreement with recent evidences showing that inhibitionof PKC promotes survival of CGCs in low potassium condi-tions through an Akt-dependent pathway (Zhu et al. 2004).
To further identify the mechanisms responsible for theneuroprotective effect of TLQP-21, we examined the effectsof modulating the activity of other death signal transductionpathways, previously shown to be involved, both in vivo andin cell culture systems in neuronal apoptosis, such as thestress-activated protein kinase JNK pathway (Miller andJohnson 1996; Watson et al. 1998; Coffey et al. 2002). Thestress-activated kinase JNK is known to regulate c-jun geneexpression and phosphorylation (Karin 1995; Ip and Davis1998; Coffey et al. 2002). As previously demonstrated,different constitutive JNK pools coexist in neurons, repre-senting different JNK isoforms (Coffey et al. 2002). In thepresent work, we demonstrated that TLQP-21 enhanced thephosphorylation of one of the three isoforms revealed by theanti-p-JNK and corresponding to 44 kDa.
The data reported in this work corroborate our findingsthat TLQP-21 acts on neuronal targets by eliciting a number
0 0 1 0 2 0 3 0 4 0 5 0 6
0
1
2
3
Time (min)
Rat
io34
0/38
0
mL / g n 5 2 - F G I mol/L m 6 5 - l C K
0 1 1 2 - r c S µ 0 1 - 1 2 - P Q L T mol/L µ mol/L
(a)
(c)
(b) 1 2 - P Q L T
F G I 6 5 K
0 10 20 30 40 50 600
1
2
3
0
1
2
3
Time (min)
0 10 20 30 40 50 60
Time (min)
0
1
2
3
0 10 20 30 40 50 60
Time (min)
(d)
1 2 - P Q L T F G I 6 5 K
1 2 - P Q L T F G I 6 5 K
0 1 0 1 3 1
Fig. 8 Effect of TLQP-21 on intracellular calcium concentration
([Ca2+]i) increase as measured by fura-2AM. (a) Trace corresponding
to the average of 40 TLQP-21 responsive cerebellar granule cells
(CGCs) from different cultures. Addition of TLQP-21 (10 lmol/L)
induced a fast [Ca2+]i increase in 6 days in vitro (DIV) CGCs, while the
scrambled peptide (Scr-21, 10 lmol/L) was completely inactive.
Insulin-like growth factor (IGF) (25 ng/mL) used as positive control,
provoked a higher, fast, and persistent effect respect to TLQP-21. KCl
56 mmol/L was used to confirm the cell viability. The scale on the left
indicates the values of ratio (340/380). (b) A representative trace
showing the [Ca2+]i of a TLQP-21 responsive CGC and (c) of a not
responsive cell. (d) Representative trace of [Ca2+]i increase induced
by different doses of TLQP-21 (1, 3, and 10 lmol/L), IGF (25 ng/mL)
and KCl 56 mmol/L in one responsive cell.
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 534–544� 2007 The Authors
542 | C. Severini et al.

of signaling cascades that may share similar properties of theaction of growth factors, possibly acting as a physiologicalregulator of cell survival for CGCs.
Besides the similarities between TLQP-21 and IGF-1 inthe neuroprotective action, several differences betweenthem in the effectiveness to CGCs apoptosis can bepointed out.
While Akt seems to be the most important pathwayinvolved in the mechanism of action of IGF-1, Erk1/2 are themain kinases implicated in the signal transduction of TLQP-21, as demonstrated by the sustained phosphorylationof Erk1/2 induced by this peptide after 24 and 48 h.Corroborating the specific involvement of the MAPK–Erk1/2 pathways in the TLQP-21 neuroprotection, UO126treatment reduced TLQP-21-mediated Erk1/2 phosphoryla-tion and completely prevented the neuroprotective effectexerted by TLQP-21, as reported by neuronal viability data,confirming that this way is crucial for the TLQP-21mechanism of action.
Finally, using fura-2AM Ca2+ imaging we demonstratedthat TLQP-21 was able to induce a significant increase in[Ca2+]i in a subpopulation of CGCs.
Sustained levels of cytosolic calcium were suggested toplay a key role in the prevention of apoptosis in differentneuronal models such as rat sympathetic neurons (Koikeet al. 1989), dorsal root ganglion neurons (Eichler et al.1992), and rat CGCs (Galli et al. 1995; Marshall et al.2003). Conducting live cell imaging using fura-2AM Ca2+
imaging allowed us to visualize the [Ca2+]i rise in individualneurons, showing that TLQP-21 was able to induce a slow[Ca2+]i rise in about 60% of the total number of analyzedCGCs, while the entire population of CGCs was responsiveto IGF-1, with a more rapid and sustained action.
Once again, the molecular mechanism for TLQP-21 rescueappears to operate through a modality of action typical toneurotrophic factors such as IGF-1, although with a reducedeffectiveness.
In conclusion, the present paper has provided clues for thepossible signaling pathways that are activated by TLQP-21 toexecute its protective function in CGCs.
As previously reported, in primary cultures of rat CGCsthe processing of VGF occurs in a post-endoplasmicreticulum compartment and the resulting products arepreferentially accumulated in secretory vesicles from whichthey are released upon cell membrane depolarization.Indirect immunofluorescence in CGCs clearly demonstratesthat VGF carbossi-terminal forms are strongly enriched insecretory vesicles present in neurites. Accordingly, only theseshort C-terminal form peptides are released on cell depolar-ization (Trani et al. 1995), suggesting that TLQP-21 may beone of these secreted peptides, acting as an autocrine/paracrine anti-apoptotic factor.
As a final point, because low-molecular-weight VGFforms were previously demonstrated to be present in the rat
cerebellum, it is tempted to speculate that TLQP-21 may actalso in vivo as a physiological regulator of cell survival forCGCs.
Acknowledgements
We would like to thank Dr Lionel Marlier for critically reading this
manuscript. This work was supported by grants from FIRB
RBNE013XSXSJ-004 and FIRB RBNEO1JKLF-006 to R. Possenti
and partially by Regione Lazio funding for ‘Sviluppo della ricerca
sul cervello’.
References
Alder J., Thakker-Varia S., Bangasser D. A., Kuroiwa M., PlummerM. R., Shors T. J. and Black I. B. (2003) Derived neurotrophicfactor-induced gene expression reveals novel actions of VGF inhippocampal synaptic plasticity. J. Neurosci. 23, 10800–10808.
Bartolomucci A., La Corte G., Possenti R. et al. (2006) TLQP-21, aVGF-derived peptide, increases energy expenditure and preventsthe early phase of diet-induced obesity. Proc. Natl Acad. Sci. USA103, 14584–14589.
Bonni A., Brunet A., West A. E., Datta S. R., Takasu M. A. andGreenberg M. E. (1999) Cell survival promoted by the Ras-MAPKsignaling pathway by transcription-dependent and -independentmechanisms. Science 286, 1358–1362.
Carrette O., Demalte I., Scherl A., Yalkinoglu O., Corthals G., BurkhardP., Hochstrasser D. F. and Sanchez J. C. (2003) A panel of cere-brospinal fluid potential biomarkers for the diagnosis of Alzhei-mer’s disease. Proteomics 3, 1486–1494.
Coffey E. T., Smiciene G., Hongisto V., Cao J., Brecht S., Herdegen T.and Courtney M. J. (2002) c-Jun N-terminal protein kinase (JNK)2/3 is specifically activated by stress, mediating c-Jun activation, inthe presence of constitutive JNK1 activity in cerebellar neurons.J. Neurosci. 22, 4335–4345.
D’Mello S. R., Galli C., Ciotti T. and Calissano P. (1993) Induction ofapoptosis in cerebellar granule neurons by low potassium: inhibi-tion of death by insulin-like growth factor I and cAMP. Proc. NatlAcad. Sci. USA 90, 10989–10993.
D’Mello S. R., Borodezt K. and Soltoff S. P. (1997) Insulin-like growthfactor and potassium depolarization maintain neuronal survival bydistinct pathways: possible involvement of PI 3-kinase in IGF-1signaling. J. Neurosci. 17, 1548–1560.
Datta S. R., Dudek H., Tao X., Masters S., Fu H., Gotoh Y. and GreenbergM. E. (1997) Akt phosphorylation of BAD couples survival signalsto the cell-intrinsic death machinery. Cell 91, 231–241.
Dekker L. V. and Parker P. J. (1994) Protein kinase C – a question ofspecificity. Trends Biochem. Sci. 19, 73–77.
Dudek H., Datta S. R., Franke T. F., Birnbaum M. J., Yao R., CooperG. M., Segal R. A., Kaplan D. R. and Greenberg M. E. (1997)Regulation of neuronal survival by the serine–threonine proteinkinase Akt. Science 275, 661–665.
Eichler M. E., Dubinsky J. M. and Rich K. M. (1992) Relationship ofintracellular calcium to dependence on nerve growth factor indorsal root ganglion neurons in cell culture. J. Neurochem. 58,263–269.
Favata M. F., Horiuchi K. Y., Manos E. J. et al. (1998) Identification of anovel inhibitor of mitogen-activated protein kinase. J. Biol. Chem.273, 18623–18632.
Finkbeiner S. (2000) CREB couples neurotrophin signals to survivalmessages. Neuron 25, 11–14.
� 2007 The AuthorsJournal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 534–544
TLQP-21 neuroprotection of cerebellar granule cells | 543

Frank C., Giammarioli A. M., Pepponi R., Fiorentini C. and Rufini S.(2004) Cholesterol perturbing agents inhibit NMDA-dependentinflux in rat hippocampal primary culture. FEBS Lett. 566, 25–29.
Galli C., Meucci O., Scorziello A., Werge T. M., Calissano P. andSchettini G. (1995) Apoptosis in cerebellar granule cells is blockedby high KCl, forskolin, and IGF-1 through distinct mechanisms ofaction: the involvement of intracellular calcium and RNA synthe-sis. J. Neurosci. 15, 1172–1179.
Gallo V., Kingsbury A., Balazs R. and Jorgenses O. S. (1987) The role ofdepolarization in the survival and differentiation of cerebellargranule cells in culture. J. Neurosci. 7, 2203–2213.
Gunn-Moore F. J., Williams A. G., Toms N. J. and Tavare J. M. (1997)Activation of mitogen-activated protein kinase and p70S6 kinase isnot correlated with cerebellar granule cell survival. Biochem.J. 324, 365–369.
Han B. H. and Holtzman D. M. (2000) BDNF protects the neonatal brainfrom hypoxic-ischemic injury in vivo via the ERK pathway.J. Neurosci. 20, 5775–5781.
Ip Y. T. and Davis R. J. (1998) Signal transduction by the c-Jun N-ter-minal kinase (JNK) – from inflammation to development. Curr.Opin. Cell Biol. 10, 205–219, Review.
Jethwa P. H., Warner A., Nilaweera K. N. et al. (2007) VGF-derivedpeptide, TLQP-21, regulates food intake and body weight inSiberian hamsters. Endocrinology 148, 4044–4055.
Karin M. (1995) The regulation of AP-1 activity by mitogen-activatedprotein kinases. J. Biol. Chem. 270, 16483–16486.
Koike T., Martin D. P. and Johnson E. M. Jr (1989) Role of Ca2+channels in the ability of membrane depolarization to preventneuronal death induced by trophic-factor deprivation: evidence thatlevels of internal Ca2+ determine nerve growth factor dependenceof sympathetic ganglion cells. Proc. Natl Acad. Sci. USA 86, 6421–6425.
Kumari S., Liu X., Nguyen T., Zhang X. and D’Mello S. R. (2001)Distinct phosphorylation patterns underlie Akt activation bydifferent survival factors in neurons. Brain Res. Mol. Brain Res.96, 157–162.
Levi G., Aloisi F., Ciotti M. T. and Gallo V. (1984) Autoradiographiclocalization and depolarization-induced release of acidic aminoacids in differentiating cerebellar granule cell cultures. Brain Res.290, 77–86.
Levi A., Eldridge J. D. and Paterson B. M. (1985) Molecular cloning of agene sequence regulated by nerve growth factor. Science 229, 393–395.
Levi A., Ferri G. L., Watson E., Possenti R. and Salton S. R. (2004)Processing, distribution, and function of VGF, a neuronal andendocrine peptide precursor. Cell. Mol. Neurobiol. 24, 517–533.
Lin W. W., Wang C. W. and Chuang D. M. (1997) Effects of depolar-ization and NMDA antagonists on the role survival of cerebellargranule cells: a pivotal role for protein kinase C isoforms.J. Neurochem. 68, 2577–2586.
Liu J. W., Andrews P. C., Mershon J. L., Yan C., Allen D. L. andBen-Jonathan N. (1994) Peptide V: a VGF-derived neuropeptidepurified from bovine posterior pituitary. Endocrinology 135, 2742–2748.
Marshall J., Dolan B. M., Garcia E. P., Sathe S., Tang X., Mao Z. andBlair L. A. (2003) Calcium channel and NMDA receptor activitiesdifferentially regulate nuclear C/EBPbeta levels to control neuronalsurvival. Neuron 39, 625–639.
Miller T. M. and Johnson E. M. Jr (1996) Metabolic and genetic analysesof apoptosis in potassium/serum-deprived rat cerebellar granulecells. J. Neurosci. 16, 7487–7495.
Nagata S., Nagase H., Kawane K., Mukae N. and Fukuyama H. (2003)Degradation of chromosomal DNA during apoptosis. Cell DeathDiffer. 1, 108–116.
Obara Y., Horgan A. M. and Stork P. J. (2007) The requirement of Rasand Rap1 for the activation of ERKs by cAMP, PACAP, and KCl incerebellar granule cells. J. Neurochem. 101, 470–482.
Rosen L. B., Ginty D. D., Weber M. J. and Greenberg M. E. (1994)Membrane depolarization and calcium influx stimulate MEK andMAP kinase via activation of Ras. Neuron 12, 1207–1221.
Salton S. R. (1991) Nucleotide sequence and regulatory studies of VGF,a nervous system-specific mRNA that is rapidly and relativelyselectively induced by nerve growth factor. J. Neurochem. 57,991–996.
Salton S. R., Ferri G. L., Hahm S., Snyder S. E., Wilson A. J., PossentiR. and Levi A. (2000) VGF: a novel role for this neuronal andneuroendocrine polypeptide in the regulation of energy balance.Front. Neuroendocrinol. 21, 199–219.
Singh V. K., Cheng J. F. and Leu S. J. (1994) Effect of substance P andprotein kinase inhibitors on beta-amyloid peptide-induced prolif-eration of cultured brain cells. Brain Res. 660, 353–356.
Snyder S. E. and Salton S. R. (1998) Expression of VGF mRNA in theadult rat central nervous system. J. Comp. Neurol. 394, 91–105.
Snyder S. E., Pintar J. E. and Salton S. R. (1998) Developmentalexpression of VGF mRNA in the prenatal and postnatal rat.J. Comp. Neurol. 394, 64–90.
Stark M., Danielsson O., Griffiths W. J., Jornvall H. and Johansson J.(2001) Peptide repertoire of human cerebrospinal fluid: novelproteolytic fragments of neuroendocrine proteins. J. Chromatogr.B. Biomed. Sci. Appl. 754, 357–367.
Trani E., Ciotti M. T., Rinaldi A. M., Canu N., Ferri G. L., Levi A. andPossenti R. (1995) Tissue-specific processing of the neuroendo-crine protein VGF. J. Neurochem. 65, 2441–2449.
Trani E., Giorgi A., Canu N., Amadoro G., Rinaldi A. M., Halban P. A.,Ferri G. L., Possenti R., Schinina M. E. and Levi A. (2002) Iso-lation and characterization of VGF peptides in rat brain. Role ofPC1/3 and PC2 in the maturation of VGF precursor. J. Neurochem.81, 565–574.
Vaillant A. R., Mazzoni I., Tudan C., Boudreau M., Kaplan D. R. andMiller F. D. (1999) Depolarization and neurotrophins converge onthe phosphatidylinositol 3-kinase-Akt pathway to synergisticallyregulate neuronal survival. J. Cell Biol. 146, 955–966.
Villalba M., Bockaert J. and Journot L. (1997) Pituitary adenylatecyclase-activating polypeptide (PACAP-38) protects cerebellargranule neurons from apoptosis by activating the mitogen-activatedprotein kinase (MAP kinase) pathway. J. Neurosci. 17, 83–90.
Volonte C., Ciotti M. T. and Battistini L. (1994) Development of amethod for measuring cell number: application to CNS primaryneuronal cultures. Cytometry 17, 274–276.
Watson A., Eilers A., Lallemand D., Kyriakis J., Rubin L. L. and Ham J.(1998) Phosphorylation of c-Jun is necessary for apoptosis inducedby survival signal withdrawal in cerebellar granule neurons.J. Neurosci. 18, 751–762.
Willaime-Morawek S., Arbez N., Mariani J. and Brugg B. (2005) IGF-Iprotects cortical neurons against ceramide-induced apoptosis viaactivation of the PI-3K/Akt and ERK pathways; is this protectionindependent of CREB and Bcl-2? Brain Res. Mol. Brain Res. 42,97–106.
Yuan J. and Yankner B. A. (2000) Apoptosis in the nervous system.Nature 407, 802–809.
Zhu D., Jiang X., Wu X., Tian F., Mearow K., Lipsky R. H. and MariniA. M. (2004) Inhibition of protein kinase C promotes neuronalsurvival in low potassium through an Akt-dependent pathway.Neurotox. Res. 6, 281–289.
Zirrgiebel U., Ohga Y., Carter B., Berninger B., Inagaki N., Thoenen H.and LindholmD. (1995)Characterization of TrkB receptor-mediatedsignaling pathways in rat cerebellar granule neurons: involvement ofprotein kinaseC in neuronal survival. J. Neurochem. 65, 2241–2250.
Journal Compilation � 2007 International Society for Neurochemistry, J. Neurochem. (2008) 104, 534–544� 2007 The Authors
544 | C. Severini et al.
Copyright © 2022 FDOKUMEN




















