
Structural health monitoring (SHM) and Nondestructive testing ...

The use of nondestructive methods to assess a physiologicalstatus and conservation perspectives of Eryngiummaritimum L.
Una Andersone & Ieva Druva-Lūsīte & Baiba Ieviņa &
Andis Karlsons & Jevgenija Ņečajeva & Ineta Samsone &
Gederts Ievinsh
Received: 5 June 2010 /Revised: 24 November 2010 /Accepted: 25 November 2010 /Published online: 7 December 2010# Springer Science+Business Media B.V. 2010
Abstract Eryngium maritimum is a perennial speciesgrowing exclusively in a coastal littoral zone both on sanddunes and shingle beach and indicated as declining inNorthern Europe. The objective of the present study was toprove the use of nondestructive physiological measurementmethods to access physiological status of endangeredplants, using E. maritimum as a model species. Plants fromtwo Latvian sites were studied in comparison with otherpopulations in Northern Europe (Estonia, Lithuania,Poland, United Kingdom) to find out if local differencesin environmental factors affect vitality of E. maritimumindividuals. Noninvasive chlorophyll analysis and chloro-phyll a fluorescence measurements were used as indicatorsof plant physiological status through characterization ofvarious aspects of photochemistry of photosystem IIactivity. Dynamics and morphology of individuals wereinvestigated for evaluation of clonal growth potential of E.maritimum in natural conditions. Highly fluctuating trendof dynamics of individuals (within 40 groups at twoLatvian sites) was established for E. maritimum in naturalconditions over a five-year period. Disturbance of individ-
uals lead to formation of new shoots from nodal rootmeristems. An exponential regression between fluorescenceparameters FV/FM and Performance Index and summarymonthly precipitation was found indicating that E.maritimum plants had significant tolerance to watershortage together with susceptibility to increased precip-itation. Negative correlation between leaf chlorophyllcontent and more northern localization of individualswas found (r=0.95 and r=0.94, for generative andvegetative shoots, respectively). Northern populations ofE. maritimum are endangered by high precipitation inconditions of low air temperature, leading to decrease ofphotosynthetic productivity and overall physiologicalstatus. High developmental plasticity at the root levelleads to clonal growth and an efficient survival andrelatively long life span of E. maritimum individuals.Chlorophyll a fluorescence is a useful method to searchfor the effect of suboptimal conditions on physiologicalstatus of endangered plant species without elimination anddisturbance of individuals.
Keywords Clonal growth . Environmental factors .
Eryngium maritimum . Mycorrhiza . Photochemistry ofphotosynthesis . Physiological status
Introduction
Studies on biological basis of plant conservation arebecoming increasingly important in applied plant biologyover recent decades. Together with population trends andgenetical aspects, knowledge on particular life forms,adaptive morphology as well as ecophysiology of particularendangered plant species is proven to be critical in
U. Andersone : I. Druva-Lūsīte :B. Ieviņa : J. Ņečajeva :I. Samsone :G. Ievinsh (*)Department of Plant Physiology, Faculty of Biology,University of Latvia,4 Kronvalda Blvd.,Riga 1586, Latviae-mail: [email protected]
A. KarlsonsLaboratory of Plant Mineral Nutrition, Institute of Biology,University of Latvia,3 Miera Str.,Salaspils 2169, Latvia
J Coast Conserv (2011) 15:509–522DOI 10.1007/s11852-010-0139-7

understanding the major affecting environmental factorsand conservation perspectives in a light of human impactand climate change (Heywood and Iriondo 2003; Coatesand Dixon 2007).
The genus Eryngium L. (Apiaceae) comprises about 250species (Calviño et al. 2008). Reproduction biology andpopulation genetics of endangered alpine species Eryngiumalpinum L. threatened by human activities has been studiedrecently (Gaudeul et al. 2004; Gaudeul and Till-Bottraud2004). Eryngium maritimum L. is a perennial speciesgrowing exclusively in a coastal littoral zone both on sanddunes and shingle beach. E. maritimum has been indicatedas declining in Europe (van der Maarel and van der Maarel-Versluys 1996). Decrease of both distribution and numberof individuals has been especially noted for Northernregions.
A complete inventory of E. maritimum population inLithuania and Poland was initiated by K. Eringis and E.Cwiklinski, respectively (Cwiklinski 1972; Eringis et al. 1976;Cwiklinski 1979; Stasiak 1986; Eringis and Pancekauskiene1995). Various aspects of E. maritimum biology have beeninvestigated more recently along coasts of the Baltic andthe North sea including population trends in Lithuania(Olšauskas 1996; Olšauskas and Urbonienė 2008) andPoland (Łabuz 2007; Żółkoś et al. 2007) as well asdemography and germination biology in Norway (Curle etal. 2007). Biogeography and genetic diversity of E.maritimum was studied recently using molecular markers(Clausing et al. 2000; Kadereit and Westberg 2007; Ievinaet al. 2009, 2010).
Different reasons, associated with both human impactand environmental factors, are mentioned as responsible forthe decline of E. maritimum. Thus, an impact of sea-levelrise along a Polish Baltic coast resulting in over-wash of thedune ridge can lead to severe deterioration of E. maritimumpopulation (Urbanski 2001). Also, Łabuz (2007) describedstorms and human impact (including coastal protectionmeasures and offorestation) as a major treats to E.maritimum through vanishing of appropriate habitats inPoland. High juvenile mortality and low germinationcapacity in Norway was noted by Curle et al. (2007).Similarly, it was hypothesized that the main reason forreduced distribution of E. maritimum in Lithuania is relatedto a small number of mature seeds as well as tounfavourable environmental conditions for seed germination(Aviziene et al. 2008). However, no ecophysiological studieswith E. maritimum have been performed so far.
Previous observations from numerous sites with E.maritimum have indicated fluctuating numbers of bothvegetative (non-flowering) and reproductive (flowering)separate individual shoots (or individuals) between differentvegetative seasons (Aviziene et al. 2008; Olšauskas andUrbonienė 2008). It is usually assumed that the vegetative
shoots represent juvenile individuals, recently establishedfrom the seed (Curle et al. 2007). However, an existence ofgroups of closely located individual shoots indicatespossible clonal character of E. maritimum, as alreadypointed out in other studies (Żółkoś et al. 2007). Whilevegetative propagation of the species by rhizomes or rootfragments has been mentioned (Turmel 1947), no study sofar evaluated a potential for clonal growth of E. maritimumin natural conditions.
For ecophysiological studies of endangered plant speciesin natural conditions, the use of nondestructive analysismethods allowing repetitive measurements of intact plantsmight be an advantage. Leaf chlorophyll content, which isan indicator of a long-term photosynthetic adaptation tounfavourable conditions (Anderson et al. 1995), can bemeasured nondestructively with a chlorophyll meter(Samsone et al. 2007). Chlorophyll a fluorescenceanalysis is another nondestructive instrumental methodreadily performed in field conditions. Different parametersof fluorescence characterize functioning of various aspectsof photochemistry of photosynthesis and can be used assensitive means to reflect the effect of suboptimal changesin environmental conditions (Maxwell and Johnson 2000).Most importantly, fluorescence measurement helps toassess physiological status and vitality of individualplants. Recently we used chlorophyll a fluorescenceanalysis in natural conditions to study changes ofphotosynthetic performance of wild plants in respect toflooding with seawater (Druva-Lūsīte et al. 2008) andsand burial (Samsone et al. 2009).
The main objective of the present study was to prove theuse of nondestructive physiological measurement methodsto access physiological status of endangered plants, usingE. maritimum as a model species. Plants from two Latviansites were studied in comparison with other populations inNorthern Europe to find out if local differences inenvironmental factors affect vitality of E. maritimumindividuals. It was specifically asked if noninvasivechlorophyll analysis and chlorophyll a fluorescence meas-urements could be used in plant conservation biologystudies as indicators of plant physiological status. Inaddition, individual dynamics and morphology was inves-tigated for evaluation of clonal growth potential of E.maritimum in natural conditions.
Materials and methods
Study sites
In Estonia, E. maritimum can be found on several islands inthe Baltic Sea. In Latvia, E. maritimum is located only intwo sites at the western part on the coast of the Baltic Sea
510 U. Andersone et al.

between Liepaja and Ventspils. In the territory of Lithuania,E. maritimum presently is located only in a 30-km zone ondunes of Curonian Spit (Aviziene et al. 2008). In Poland, E.maritimum can be mainly found on coastal dunes of spitsand sandbars between the Baltic Sea and coastal lakes(Łabuz 2007; Żółkoś et al. 2007). In contrast, relativelyabundant distribution of E. maritimum is characteristic forcoastal areas of the UK. However, it is becoming extinct inthe northeastern part of the British Isles (Clausing et al.2000). A location map of study sites is given in Fig. 1 witha summary presented in Table 1. As E. maritimum inEstonia, Latvia, Lithuania and Poland is protected bylegislation, only nondestructive methods of analysis andsampling were used in this study.
Climate data were obtained from the Latvian Environ-ment, Geology and Meteorology Centre at http://www.meteo.lv.
Mapping and counting of individuals
Mapping of individuals were performed within fiveconsecutive years (2005–2009) in the two Latvian sitesZiemupe and Uzava (locations no. 3 and 4; Table 1, Fig. 1).A local map with microrelief objects and positions ofindividual plants (shoots) were prepared in the firstvegetation season. According to the map, number and anexact location of individual shoots (flowering and vegeta-tive) at the particular microsite were determined every yearat the end of August. Counting of individuals wasperformed only in separate, clearly distinguishable groups,occupying no more than 1 m2 of ground. The number ofgroups continuously counted was 25 and 15, for Ziemupeand Uzava, respectively.
Root sampling and analysis of arbuscular mycorrhiza
Root sampling was performed in two Latvian sites Ziemupeand Uzava during the vegetation seasons of 2007 and 2008.Root samples were obtained from a rhizosphere of intactgrowing plants by digging a slanted hole from a side of a
plant. Three individual plants per site were sampled once amonth from May to September. Root samples with soilwere placed in sealed polyethylene bags, transported to thelaboratory and kept at 4°C until analysis.
Root samples were prepared according to a modifiedmethod of Hayman (1970) by boiling washed root frag-ments in 10% KOH for 1 h, washing and staining for 5 minin 0.05% trypan blue (Druva-Lūsīte et al. 2008). Afterwashing, samples were stored in lactoglycerol. Mycorrhizalcolonization was assessed under microscope (Eclipse E200,Nicon, Tokyo, Japan) at 400 × magnification using 10 rootsegments from each individual plant (Trouvelot et al. 1986).Both intensity (M%) and frequency (F%) of mycorrhizalcolonization were calculated with the computer programMycocalc as described previously (Druva-Lūsīte et al.2008).
Chlorophyll content and chlorophyll a fluorescencemeasurements
Physiological analysis of E. maritimum at the two Latviansites Ziemupe and Uzava (locations no. 3 and 4; Table 1,Fig. 1) were repeated within two consecutive vegetationseasons (2007, 2008) with similar results. Plant analysis inPoland (locations no. 6, 7 and 8) and UK (locations no. 9and 10) was performed in August of 2007. Only the datafrom 2007 are shown in the present paper representing theseason with measurements from the largest number of sites.At the end of the vegetation season of 2009, comparativephysiological analysis was performed in five North Europeansites (locations no. 1 to 5, Table 1, Fig. 1).
Leaf chlorophyll content was measured in naturalconditions by SPAD-502 chlorophyll meter (Konica-Minolta, Osaka, Japan). Measurement was performed on30 intact leaves from at least five individual plants foreach site. Five consecutive readings were made acrossthe surface of each leaf. The mean value was calculatedusing the internal function of the chlorophyll meter.
Chlorophyll a fluorescence was measured in fieldconditions using two different methods, e.g., pulse amplitude
Fig. 1 The map of study sitesof E. maritimum in NorthernEurope. Location numbers as inTable 1
The use of nondestructive methods in plant conservation biology 511

modulated and continuous measurement. For a pulse modu-lated measurement portable fluorometer (PAM2100, H.Walz,Effeltrich, Germany) and leaf clip holder (2030-B, H. Walz,Effeltrich, Germany) with integrated micro quantum-temperature sensor were used. A laptop computer equippedwith an appropriate software (DA-2000, H. Walz, Effeltrich,Germany) was used to drive the measurements. Leaves weredark adapted for 30 min prior the measurements usingappropriate leaf clips provided by the supplier. Five indepen-dent measurements on individual leaves were performed forevery site. Fluorescence measurements were performedaccording to manufacturer’s instructions as described previ-ously (Samsone et al. 2009). First, potential maximumquantum yield (FV/FM) of photosystem II was measured.The minimal fluorescence level (F0) was measured by lowmodulated light and the maximal fluorescence level (FM)was determined by a saturating pulse on dark-adapted leaves.The ratio FV/FM was calculated, where FV is the differencebetween the maximal fluorescence and the minimal fluores-cence level. After that, fluorescence induction curve withquenching analysis at 10 ms p−1 was recorded within 6 minusing a built-in standard procedure of DA-2000. After lightadaptation of 6 min, the steady-state fluorescence FS wasrecorded. Then the maximal fluorescence level in the lightadapted state (F′M) and the minimal fluorescence level (F′0)was measured by means of saturating pulse and far-red lightpulse, respectively. Relative photochemical quantum yield ofphotosystem II [Y(II)] was calculated as Y(II)=ΔF/FM′;where ΔF=FM′ – FS. Light-dependent non-photochemicalquenching (NPQ) was calculated according to the equationNPQ=(FM – F′M)/F′M.
For continuous measurement, a Handy PEA chlorophyllfluorometer (Hansatech, Kings Lynn, Norfolk, UK) wasused. Before analysis, intact leaves were dark adapted for30 min using appropriate leaf clips. For every site, 10 to 15independent measurements on individual leaves wereperformed. The data were analyzed by PEA Plus software(Hansatech, Kings Lynn, Norfolk, UK). Several fluores-
cence parameters obtained by the continuous fluorescencemeasurement method were analyzed in the present study.Performance Index (PI) is a combination of the threeindependent parameters, expressing cumulated response ofphotosystem II – total number of active reaction centres perabsorption, yield of primary photochemistry, and efficiencywith which a trapped exciton can move an electron into theelectron transport chain (Appenroth et al. 2001). Theparameter Area describes the pool size of electron acceptorsfrom photosystem II and is proportional to the oxidizedplastoquinone pool. The ratio between total chlorophyll andthat of antenna complexes (Chl RC/Chl antenna) charac-terize energy fluxes for reaction centres. The parameter canbe replaced by the ratio RC/ABS, where RC is the numberof active photosystem II reaction centres, and ABS is thequantity of light absorbed (Clark et al. 2000).
Soil analysis
Soil analysis was performed in two Latvian sites Ziemupeand Uzava during the vegetation season of 2007. Soilsamples were collected from the rhizosphere of E.maritimum plants at the depth of 5 to 20 cm. For everysite three to five subsamples were taken once a month fromMay to September and were thoroughly mixed to form onesample. Soil samples were dried in air and sieved. Mineralnutrients were extracted with 1 M HCl at 1:5 soil toextractant volume. Soil pH and electrical conductivity wasmeasured, and nutrient concentrations in soil samples wereanalyzed by atomic absorption spectrophotometry (Ca, Mg,Fe, Cu, Zn, Mn), colorimetry (N, P, Mo, B), turbidimetry(S), flame photometry (K, Na) and AgNO3 titration (Cl) aspreviously described (Karlsons et al. 2008).
Data analysis
Data analysis was performed by KaleidaGraph (version4.0; Synergy Software). Statistically significant differ-
Table 1 Study sites of E. maritimum
Location no. Locality Country Geographical coordinates Year of analysis
1 Island of Kihnu Estonia 58°07′N; 23°57′E 2009
2 Kihelkonna, Island of Saaremaa Estonia 58°28′N; 21°53′E 2009
3 Ziemupe Latvia 56°48′N; 21°04′E 2005–2009
4 Uzava Latvia 57°15′N; 21°25′E 2005–2009
5 Nida on Curonian spit Lithuania 55°22′N; 21°01′E 2009
6 Krynica Morska on Vistula spit Poland 54°23′N; 19°28′E 2007
7 Dabki Poland 54°22′N; 16°18′E 2007
8 Miedzyzdroje Poland 53°54′N; 14°22′E 2007
9 Formby, England UK 53°33′N; 3°06′E 2007
10 Newborough, Isle of Anglesey, Wales UK 53°08′N; 4°21′E 2007
512 U. Andersone et al.
ences between the parameters were determined atP≤0.05. Correlations between the parameters were estimatedat P≤0.05.
Results
Dynamics of individuals and morphological adaptations
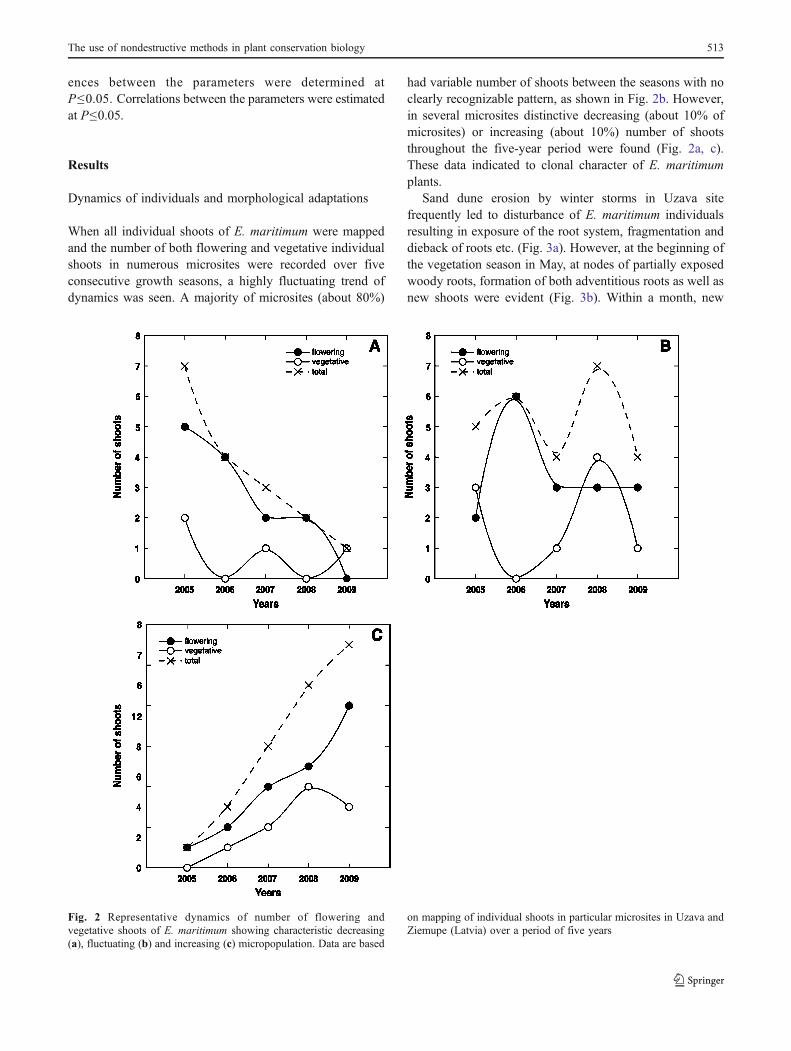
When all individual shoots of E. maritimum were mappedand the number of both flowering and vegetative individualshoots in numerous microsites were recorded over fiveconsecutive growth seasons, a highly fluctuating trend ofdynamics was seen. A majority of microsites (about 80%)
had variable number of shoots between the seasons with noclearly recognizable pattern, as shown in Fig. 2b. However,in several microsites distinctive decreasing (about 10% ofmicrosites) or increasing (about 10%) number of shootsthroughout the five-year period were found (Fig. 2a, c).These data indicated to clonal character of E. maritimumplants.
Sand dune erosion by winter storms in Uzava sitefrequently led to disturbance of E. maritimum individualsresulting in exposure of the root system, fragmentation anddieback of roots etc. (Fig. 3a). However, at the beginning ofthe vegetation season in May, at nodes of partially exposedwoody roots, formation of both adventitious roots as well asnew shoots were evident (Fig. 3b). Within a month, new
Fig. 2 Representative dynamics of number of flowering andvegetative shoots of E. maritimum showing characteristic decreasing(a), fluctuating (b) and increasing (c) micropopulation. Data are based
on mapping of individual shoots in particular microsites in Uzava andZiemupe (Latvia) over a period of five years
The use of nondestructive methods in plant conservation biology 513

individual vegetative shoots were established on the newlyformed dune ridge (Fig. 3c). By detailed inspection ofdisturbed E. maritimum plants on eroded sand dunes, it wasnoticed that many individuals with partially exposed rootsystem bore signs of previous disturbances at the root level.Most importantly, when the top of the taproot was injuredor killed by an infection, new individual shoots wereformed from the root top 10 to 30 cm below the ground(Fig. 4).
On habitats, different than foredunes or fixed dunes,individuals of E. maritimum can have different morphologyof the root system. Thus, in a dune plato of Abermenaidunes located at the end of an arm of the Menai Strait in theNorth Wales, UK, relatively shallow root system (up to1 m) forming an extensive clonal network of individualshoots of E. maritimum were found (Fig. 5). Similarshallow root system was evident for plants growing ongravel based low beaches of the Island of Kihnu, Estonia.
Soil properties and mycorrhizal colonization
Both intensity and frequency of mycorrhizal symbiosiswere relatively high in roots of E. maritimum throughoutthe vegetation season and were not significantly affected bylocal environmental factors (Fig. 6). However, a character-istic trend was evident for changes of intensity of symbiosisduring the season, showing continuous decrease till Augustwith subsequent significant increase in September. As thetrend seemed to be opposite to that of leaf chlorophyllcontent, an analysis of correlation between the mean monthlyvalues of the two parameters were performed, showing thatthere indeed was a linear negative correlation between leafchlorophyll content and root intensity of mycorrhizal symbi-
osis for E. maritimum plants (r=0.76 for both sites togetherand r=0.95 and r=0.81, for separate analysis of the datafrom Uzava and Ziemupe, respectively).
Soil at both Latvian sites was alkaline with low humuscontent and relatively low salinity as indicated by soilelectrical conductivity and concentrations of Na and Cl(Table 2). No significant differences were found betweenthe sites in concentration of both macronutrients andmicronutrients except relatively higher Mg and Fe contentin Ziemupe.
Photosynthesis-related parameters
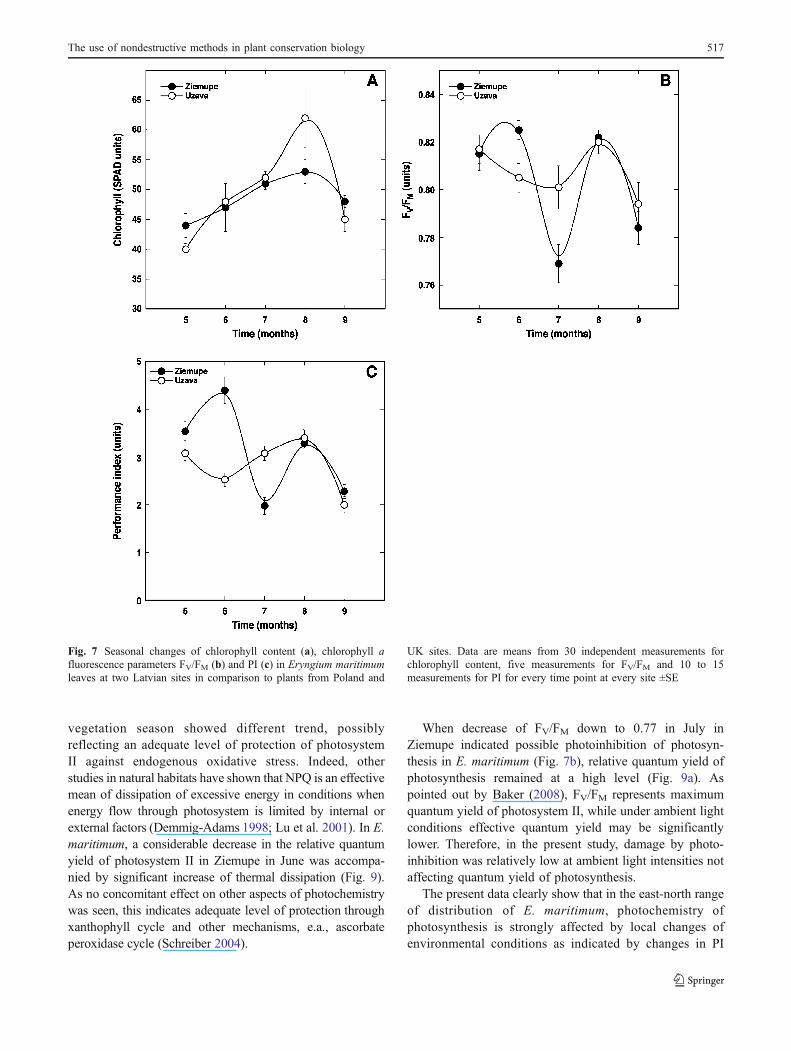
Leaf chlorophyll content showed a tendency to increasefrom May to August, with significant decrease in September(Fig. 7a). No significant differences were seen for Ziemupeand Uzava, with the exception of August, when E.maritimum plants in Uzava had higher chlorophyll level.The chlorophyll data from the three Polish sites and the twoUK sites coincided well with the trend observed for theLatvian populations.
Potential maximum quantum yield of photosystem II(FV/FM) in May was identical for plants in Ziemupe andUzava (Fig. 7b). Some further decrease of FV/FM in Juneand July were seen for plants in Uzava, while in Ziemupe,the increase in June was followed by a drastic decrease inJuly, indicating severe photoinhibition of photosynthesisand decrease in vitality. In August, FV/FM returned to asimilar level at both sites with characteristic identicaldecrease in September. In August, E. maritimum plants inPoland tended to have higher FV/FM than Latvian individ-uals, while at the UK sites FV/FM was comparably thesame. Seasonal changes in Performance Index had similar
Fig. 3 a An individual of E.maritimum on a coastal sanddune with roots exposed due todune erosion, May. b Detailedview of root segment with for-mation of new tissues. Note theformation of both shoots androots. c The same individual inJune. Uzava dunes, Latvia
514 U. Andersone et al.

character to these of FV/FM, although plants from sitesoutside Latvia had a tendency to have relatively higherlevels of Performance Index and, as a consequence, betterphysiological performance of individuals.
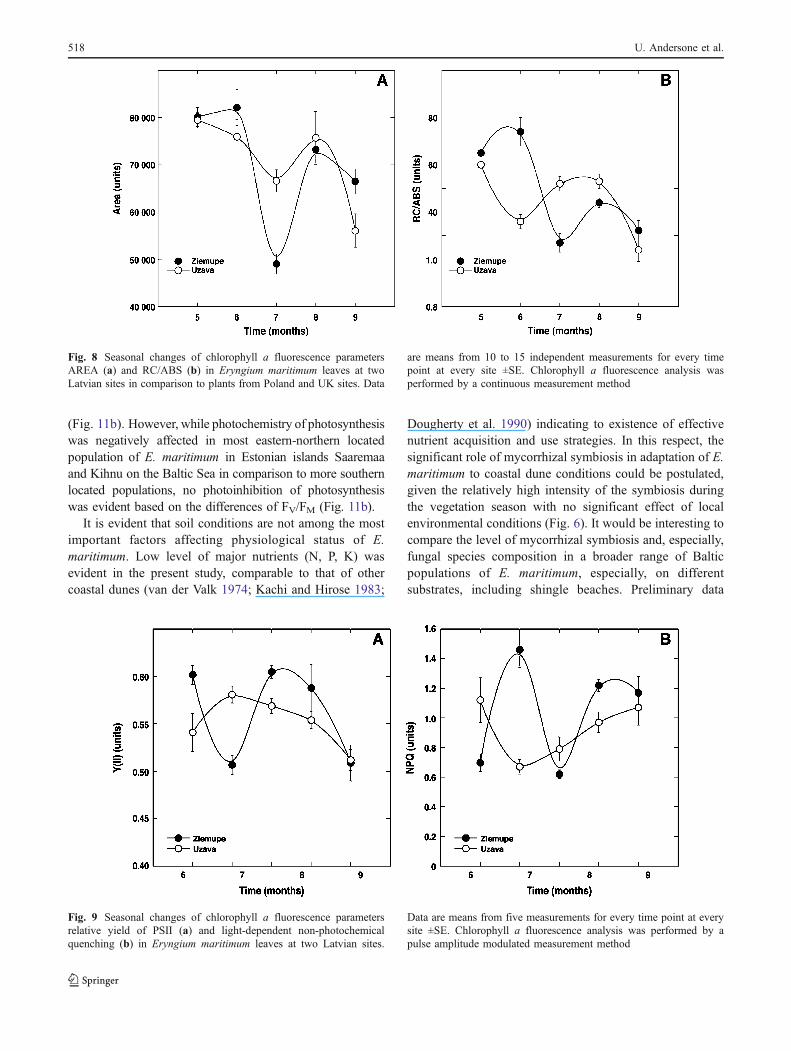
Analysis of other chlorophyll a fluorescence parametersreflecting energy flow as well as energy quenching inphotosystem II suggested that various aspects of photo-chemistry of E. maritimum were differentially affected byenvironmental conditions. While the fluorescence parame-ter Area (proportional to the oxidized plastoquinone pool)showed a trend similar to that of FV/FM (Fig. 8a), thechanges of parameter RC/ABS (reflects the proportion ofactive reaction centres of photosystem II) were similar tothese of Performance Index (Fig. 8b).
Chlorophyll fluorescence quenching analysis showedthat the relative quantum yield of photochemistry ofphotosystem II [Y(II)] significantly decreased in E. maritimumindividuals from Ziemupe site in June (Fig. 9a), whichcoincided with a significant temporary increase in non-photochemical quenching (Fig. 9b), evidently reflecting lowerphysiological performance and vitality of individuals.
In general, FV/FM positively linearly correlated withPerformance Index for all mean monthly measurements atall sites (r=0.87; data not shown). When the mean monthlyvalues of FV/FM (Fig. 10) or Performance Index at Ziemupeand Uzava were correlated with the summary precipitationfor a particular site in the respective month, both parametersdecreased with the increase of precipitation. The correlation
had a character of exponential regression (r=0.88 andr=0.70, for FV/FM and Performance Index, respectively)and was shifted to the left, indicating that E. maritimumplants had significant tolerance to water shortage togetherwith susceptibility to increased precipitation (Fig. 10).There was a strong positive linear correlation between themean number of sunny hours per day and both FV/FM(r=0.86) and Performance Index (r=0.98) for plants at thetwo Latvian sites (data not shown).
In order to find out if photosynthesis-related parametersin E. maritimum could be possibly affected by a morenorthern localization of individuals, physiological analyseswere performed in five North European populations locatedacross the Baltic coast at different latitudes vs. similarlongitudes. When leaf chlorophyll content of both genera-tive and vegetative E. maritimum individuals were plottedagainst a geographical distance in south-north direction(Fig. 11a), a distinctive negative correlation with morenorthern localization of individuals was found (r=0.95 andr=0.94, for generative and vegetative shoots, respectively).In contrast, photosynthetic performance indices FV/FM andPerformance Index (Fig. 11b) as well as other fluorescenceparameters did not show a relationship with a geographicaldistance. However, Performance Index was significantlyreduced in the two northernmost localized E. maritimumsites in Estonia, eventually showing some climate-relatedeffect on photosynthetic performance and vitality ofindividuals.
Discussion
Within the last decades, chlorophyll a fluorescence techniquehas been employed in a wide range of studies in plant
Fig. 5 An extensive clonal root network of E. maritimum plants inwind-eroded dune ridge. Abermenai dunes, Newborough, Isle ofAnglesey, Wales, UK
Fig. 4 Morphological structure and measurements of the upper part ofa root system of E. maritimum. Black line represents soil level
The use of nondestructive methods in plant conservation biology 515

biology and it was demonstrated that fluorescence parame-ters can be used for characterization of various aspects ofphotochemistry of photosystem II (Maxwell and Johnson2000). Most importantly, in the context of the present study,the use of certain fluorescence parameters for assessment ofphysiological status and vitality of plants in natural con-ditions was validated (Tsimilli-Michael and Strasser 2008).
The decline of ratio of variable to maximum fluores-cence or potential maximum quantum yield of photosystem
II (FV/FM) below 0.8 has been shown to reflect inhibition ofphotosynthesis and, therefore, is indicative of past episodesof suboptimal conditions (Öquist et al. 1992). On the otherhand, PI is a more complex parameter reflecting overallefficiency of light absorption as well as both light and darkredox reactions (Strauss et al. 2006). Therefore, PI is apotential indicator of current physiological status of a plant,reflecting both disturbance and acclimation of photosyn-thetic apparatus by changing environmental conditions. Ananalysis of PI by a continuous fluorescence meter has beenshown to be useful as a method for monitoring of treehealth (Clark et al. 2000). Recently PI was suggested to bean useful indicator of vigour and health in plant populationsin natural habitats (Thach et al. 2007).
While in general positive linear correlation between FV/FM and PI for North European E. maritimum individualswas evident, dynamics of changes in the individualparameters was not identical, suggesting the different effectof environmental factors on various aspects of photochem-istry of photosynthesis in E. maritimum plants (Fig. 7b, c).Monthly changes of summary precipitation explained 88%of FV/FM variation and only 70% of that for PI. On theother hand, variation in the mean monthly number of sunnyhours per day explained 98% of variation for PI and only86% for that of FV/FM. This difference can be explained, atleast in part, by different time scales of effect by changingenvironmental conditions on various aspects of the photo-chemistry, where PI shows immediate general responses ofphotosynthetic machinery while FV/FM reflects longer-lasting response. While changes in energy flow parametersof chlorophyll a fluorescence (Area and RC/ABS)exhibited close resemblance to the performance indices,energy quenching parameters Y(II) and NPQ during the
Fig. 6 Seasonal dynamics of intensity (a) and frequency (b) of mycorrhizal infection in roots of E. maritimum at two Latvian sites. Data aremeans from 3 independent measurements for every time point at every site ±SE
Table 2 Soil properties and macro- and micronutrient concentrations(mg L−1) at two Latvian sites of Eryngium maritimum. Data are meansof five independent measurements within a vegetation season ±SE
Uzava Ziemupe
Soil pH 8.52±0.03 8.40±0.07
Soil EC (dS m−1) 0.26±0.04 0.25±0.06
Soil organic matter (%) 0.4±0.1 0.7±0.2
N 17.0±1.5 18.0±2.0
P 200±32 146±22
K 42±25 53±36
Ca 20948±2126 20013±149
Mg 1261±70 2195±149
S 13±1 20±8
Fe 293±19 319±24
Mn 40±7 59±5
Zn 3.7±0.1 3.5±0.3
Cu 0.67±0.08 0.54±0.06
Mo 0.020±0.007 0.026±0.005
B 0.130±0.007 0.240±0.140
Na 32±3 26±2
Cl 18±5 12±6
516 U. Andersone et al.
vegetation season showed different trend, possiblyreflecting an adequate level of protection of photosystemII against endogenous oxidative stress. Indeed, otherstudies in natural habitats have shown that NPQ is an effectivemean of dissipation of excessive energy in conditions whenenergy flow through photosystem is limited by internal orexternal factors (Demmig-Adams 1998; Lu et al. 2001). In E.maritimum, a considerable decrease in the relative quantumyield of photosystem II in Ziemupe in June was accompa-nied by significant increase of thermal dissipation (Fig. 9).As no concomitant effect on other aspects of photochemistrywas seen, this indicates adequate level of protection throughxanthophyll cycle and other mechanisms, e.a., ascorbateperoxidase cycle (Schreiber 2004).
When decrease of FV/FM down to 0.77 in July inZiemupe indicated possible photoinhibition of photosyn-thesis in E. maritimum (Fig. 7b), relative quantum yield ofphotosynthesis remained at a high level (Fig. 9a). Aspointed out by Baker (2008), FV/FM represents maximumquantum yield of photosystem II, while under ambient lightconditions effective quantum yield may be significantlylower. Therefore, in the present study, damage by photo-inhibition was relatively low at ambient light intensities notaffecting quantum yield of photosynthesis.
The present data clearly show that in the east-north rangeof distribution of E. maritimum, photochemistry ofphotosynthesis is strongly affected by local changes ofenvironmental conditions as indicated by changes in PI
Fig. 7 Seasonal changes of chlorophyll content (a), chlorophyll afluorescence parameters FV/FM (b) and PI (c) in Eryngium maritimumleaves at two Latvian sites in comparison to plants from Poland and
UK sites. Data are means from 30 independent measurements forchlorophyll content, five measurements for FV/FM and 10 to 15measurements for PI for every time point at every site ±SE
The use of nondestructive methods in plant conservation biology 517
(Fig. 11b). However, while photochemistry of photosynthesiswas negatively affected in most eastern-northern locatedpopulation of E. maritimum in Estonian islands Saaremaaand Kihnu on the Baltic Sea in comparison to more southernlocated populations, no photoinhibition of photosynthesiswas evident based on the differences of FV/FM (Fig. 11b).
It is evident that soil conditions are not among the mostimportant factors affecting physiological status of E.maritimum. Low level of major nutrients (N, P, K) wasevident in the present study, comparable to that of othercoastal dunes (van der Valk 1974; Kachi and Hirose 1983;
Dougherty et al. 1990) indicating to existence of effectivenutrient acquisition and use strategies. In this respect, thesignificant role of mycorrhizal symbiosis in adaptation of E.maritimum to coastal dune conditions could be postulated,given the relatively high intensity of the symbiosis duringthe vegetation season with no significant effect of localenvironmental conditions (Fig. 6). It would be interesting tocompare the level of mycorrhizal symbiosis and, especially,fungal species composition in a broader range of Balticpopulations of E. maritimum, especially, on differentsubstrates, including shingle beaches. Preliminary data
Fig. 9 Seasonal changes of chlorophyll a fluorescence parametersrelative yield of PSII (a) and light-dependent non-photochemicalquenching (b) in Eryngium maritimum leaves at two Latvian sites.
Data are means from five measurements for every time point at everysite ±SE. Chlorophyll a fluorescence analysis was performed by apulse amplitude modulated measurement method
Fig. 8 Seasonal changes of chlorophyll a fluorescence parametersAREA (a) and RC/ABS (b) in Eryngium maritimum leaves at twoLatvian sites in comparison to plants from Poland and UK sites. Data
are means from 10 to 15 independent measurements for every timepoint at every site ±SE. Chlorophyll a fluorescence analysis wasperformed by a continuous measurement method
518 U. Andersone et al.

indicate that roots of E. maritimum contain a broaddiversity of mycorrhizal fungal species as revealed bymolecular analysis (at least, 16 different genotypes inconditions of Latvia; Druva-Lusite, unpublished data).
High adaptive plasticity of E. maritimum at the root levelbased on extremely high regenerative ability was revealedallowing successful growth of individuals on a wide varietyof substrates – driftline shingle, gravel beach, foredunes,semi-fixed primary dunes, fixed grey dunes, dune forest.Based on both dynamics of individuals in micropoulationsand morphological features of the root system, it could beproposed that a life span of a true individual plant of E.maritimum (a genet) lasts several decades. Demographicdata so far did not show level of individual shoots,therefore, no relation with growth form dynamics ofindividual plants were established. As an exception, Curleet al. (2007) suggested that mostly survival of fertile plantscontributed to the population growth rate, proposing alsoregrowth of individuals from root fragments. Character ofindividual dynamics on microsites in the present studyclearly indicate that, to a great extent, individual shoots ofE. maritimum are relatively independent ramets of a root-clonal genet. This conclusion was supported by the resultsfrom molecular genetic analysis of E. maritimum at the twoLatvian populations, revealing no significant genetic differ-ences between individual shoots within a local subpopula-tion (Ievina et al. 2009). Similarly, in alpine environments,individuals of some species persist even for centuriesmainly due to reproduction by clonal structures (Steingeret al. 1996). Reduced sexual reproduction, rare seedlingrecruitment and long life span of individuals are indeed
Fig. 11 Relationship between chlorophyll content in leaves ofvegetative and generative shoots (a) and leaf chlorophyll a fluores-cence parameters FV/FM and Performance Index (b) and south-northdistance of E. maritimum plants. Numbers indicate a particular siteaccording to Fig. 1 and Table 1. Data are means from 30 independent
measurements of individual leaves for chlorophyll content and 10 to15 independent measurements for fluorescence analysis for every timepoint at every site ±SE. Chlorophyll a fluorescence analysis wasperformed by a continuous measurement method
Fig. 10 Relationship between summary precipitation and FV/FM inEryngium maritimum leaves. Each point represents the data fromFig. 7b plotted against summary precipitation for the respective monthat the particular site
The use of nondestructive methods in plant conservation biology 519

characteristic features of many clonal plants (Eriksson1989).
Due to the localization of meristematic tissues on themain taproots, any disturbance at the shoot or root level(dune erosion, trampling, pulling out, digging, heavyherbivory, root infections) results in root injury and canlead to induction of proliferative activity in root meriste-matic cells localized at the root nodes. As a result, both newshoots and adventitious roots are formed leading toestablishment of new ramets. Similarly, during undisturbedgrowth, new individual clones are formed every vegetationseason from vegetative buds or root meristems located 0 to30 cm above the ground level. A deducted sequence ofclonal growth phases leading to fluctuating character ofdynamics of individual shoots of E. maritimum oversequential vegetation seasons is shown in Fig. 12.
The amount of root-stored carbohydrates possibly controlsthe number and type (vegetative vs. generative) of the newlyformed shoots for a particular individual root top. Similarly,dependence of regeneration capacity on rhizome-storedreserves of carbohydrates has been shown for other clonalplant species (Klimeš et al. 1993). Therefore, highlyfluctuating character of individual shoot dynamics of E.maritimum for a particular microsite have been seen, whenseasons of a large number of flowering individual shoots arefollowed by mostly vegetative shoots, and vice versa. Owingsignificant relative contribution of vegetative shoots inproduction of photosynthates, as opposed to a high rate ofassimilate use for seed production by flowering shoots,vegetative shoots represent a producing form allowing
accumulation of root carbohydrate reserves for support ofmore flowering individual shoots in the next vegetationseason.
The results from the study of E. maritimum seed germina-tion will be published elsewhere (Necajeva and Ievinsh,unpublished). Though, in the light of the present discussion, itseems to be reasonable to mention that the generativereproduction of E. maritimum at the northeast range ofdistribution might be affected by a low number of fertileseeds produced as well as by a high mortality rate of bothseeds and seedlings. In addition, when studies on geneticaldiversity of E. maritimum in Eastern Baltics were performedusing molecular markers, it was suggested based on chloro-plast DNA sequences that Baltic populations of E. maritimumexhibit low level of polymorphism (Ievina et al. 2009).
Conclusions
Northern European populations of E. maritimum areendangered by high precipitation in conditions of low airtemperature, leading to decrease of photosynthetic produc-tivity and overall physiological status, as well as byunidentified factors negatively affecting seed ripening andgermination. However, high developmental plasticity at theroot level leads to clonal growth and, as a consequence, anefficient survival and relatively long life span of E.maritimum individuals.
Based on the present study, it can be suggested thatchlorophyll a fluorescence is useful not only for nonde-
Fig. 12 Schematic representa-tion of a deducted sequence ofclonal growth phases (A to G) ofE. maritimum on a sand dune. f,flowering individual shoot; v,vegetative individual shoot
520 U. Andersone et al.

structive measurement of stress impact on physiologicalstatus of plants, but also to search for the effect of anysuboptimal conditions through the analysis of variousaspects of photochemistry of photosynthesis. When usedin natural conditions, the method allows to study endan-gered plant species without elimination and disturbance ofindividuals. In respect to conservation perspectives of E.maritimum, decrease of photosynthetic performance innorthernmost sites due to unfavourable environmentalconditions as revealed by nondestructive methods pointsto the necessity of ex situ conservation measures of thespecies.
Acknowledgments This work was supported by the Latvian Fundfor Environment Protection (grant number 1–08/459/2006) and by theUniversity of Latvia (grant numbers 2005/ZP23, 2006/1–229708,2007/ZP20, ZP20/2008–0530). Valuable suggestions of the anony-mous referee are gratefully acknowledged.
References
Anderson JM, Chow WS, Park Y-I (1995) The grand design ofphotosynthesis: acclimation of the photosynthetic apparatus toenvironmental cues. Photosynth Res 46:129–139
Appenroth K-J, Stöckel J, Srivastava A, Strasser RJ (2001) Multipleeffects of chromate on the photosynthetic apparatus of Spirodelapolyrhiza as probed by OJIP chlorophyll a fluorescence measure-ments. Environ Pollut 115:49–64
Aviziene D, Pakalnis R, Sendzikaite J (2008) Status of red-listedspecies Eryngium maritimum L. on the Lithuanian coastal dunes.In: Cygas D, Froehner KD (eds) Environmental Engineering. The7th International Conference. Vilnius: Gediminas TechnicalUniversity, pp 22–28
Baker NR (2008) Chlorophyll fluorescence: a probe of photosynthesisin vivo. Annu Rev Plant Biol 59:89–113
Calviño CI, Martínez SG, Downie SR (2008) The evolutionary hsitoryof Eryngium (Apiaceae, Saniculoideae): Rapid radiations, longdistance dispersals, and hybridization. Mol Phylogenet Evol46:1129–1150
Clark AJ, Landolt W, Bucher JB, Strasser RJ (2000) Beech (Fagussylvatica) response to ozone exposure assessed with a chlorophylla fluorescence performance index. Environ Pollut 109:501–507
Clausing G, Vickers K, Kadereit JW (2000) Historical biogeographyin a linear system: genetic variation of Sea Rocket (Cakilemaritima) and Sea Holly (Eryngium maritimum) along Europeancoasts. Mol Ecol 9:1823–1833
Coates DJ, Dixon KW (2007) Current perspectives in plant conser-vation biology. Aust J Bot 55:187–193
Curle CM, Stabbetorp OE, Nordal I (2007) Eryngium maritimum,biology of a plant at its northernmost localities. Nord J Bot24:617–628
Cwiklinski E (1972) The distribution of the sea-holly, Eryngiummaritimum, on the Baltic sea-coast near Szczecin. ChronmyPrzyrode Ojczysta 28:21–29 (in Polish)
Cwiklinski E (1979) The population size of Eryngium maritimum L.on the Polish Baltic Sea coast. Chronmy Przyrode Ojczysta 35:5–14 (in Polish)
Demmig-Adams B (1998) Survey of thermal energy dissipation andpigment composition in sun and shade leaves. Plant Cell Physiol39:474–482
Dougherty KM, Mendelssohn IA, Monteferrante FJ (1990) Effects ofnitrogen, phosphorus and potassium additions on plant biomassand soil nutrient content of a swale barrier strand community inLouisiana. Ann Bot 66:265–271
Druva-Lūsīte I, Karlsons A, Osvalde A, Ņečajeva J, Ievinsh G (2008)Photosytnthetic performance and mycorrhizal symbiosis of acoastal marsh plant, Glaux maritima, in conditions of fluctuatingsoil salinity. Acta Univ Latv 745:155–164
Eriksson O (1989) Seedling dynamics and life histories in clonalplants. Oikos 55:231–238
Eringis K, Pancekauskiene D (1995) Eryngium maritimum L.population dynamics in the Kuršiu Nerija National Park ofLithuania and total population inventory method. In: Gudelis V,Povilanskas R, Roepstroff A (eds) Coastal Conservation andManagement in the Baltic Region. Proceedings of the EUCC-WWF Conference. Klaipeda, Lithuania, pp 49–54
Eringis K, Preskenis J, Eringyte V (1976) Abundance and dynamicsof Eryngium maritimum on the Curonian Spit seaside dune.Phytogeographical, floristical and geobotanical characteristics ofcoastal vegetation. Lithuanian Academy of Sciences, Vilnius, pp151–153 (in Russian)
Gaudeul M, Till-Bottraud I (2004) Reproductive ecology of theendangered alpine species Eryngium alpinum L. (Apiaceae):phenology, gene dispersal and reproductive success. Ann Bot93:711–721
Gaudeul M, Till-Bottraud I, Barjon F, Manel S (2004) Geneticdiversity and differentiation in Eryngium alpinum L. (Apiaceae):comparison of AFLP and microsatellite markers. Heredity92:508–518
Hayman DS (1970) Endogone spore numbers in soil and vesicular-arbuscular mycorrhiza in wheat as influenced by season and soiltreatment. Trans Br Mycol Soc 54:53–65
Heywood VH, Iriondo JM (2003) Plant conservation: old problems,new perspectives. Biol Conserv 113:321–335
Ievina B, Rostoks N, Ievinsh G (2009) Genetic diversity in Latvianpopulations of Sea Holly Eryngium maritimum L. LatvijasVegetacija 18:13–24 (in Latvian)
Ievina B, SyedNH, Flavell NH, IevinshG, RostoksN (2010) Developmentof retrotransposon-based SSAP molecular marker system for study ofgenetic diversity in sea holly (Eryngium maritimum L.). Plant GenetRes doi:10.1017/S1479262110000316
Kachi N, Hirose T (1983) Limiting nutrients for plant growth incoastal san dune soils. J Ecol 71:937–944
Kadereit JW, Westberg E (2007) Determinants of phylogeographicstructure: a comparative study of seven coastal flowering plantspecies across their European range. Watsonia 26:229–238
Karlsons A, Osvalde A, Necajeva J, Ievinsh G (2008) Changes ofnutritional status of coastal plants Hydrocotyle vulgaris and Astertripolium at elevated soil salinity. Acta Univ Latv 745:165–177
Klimeš L, Klimešová J, Osbornová J (1993) Regeneration capacityand carbohydrate reserves in a clonal plant Rumex alpinus: effectof burial. Vegetatio 109:153–160
Łabuz TA (2007) Evaluation of past and present sea holly (Eryngiummaritimum) habitats on Polish cosatal dunes. Acta Univ Latv723:99–114
Lu C, Lu Q, Zhang J, Zhang Q, Kuang T (2001) Xanthophyll cycle,light energy dissipation and photosystem II down-regulation insenescent leaves of wheat plants grown in the field. Aust J PlantPhysiol 28:1023–1030
Maxwell K, Johnson GN (2000) Chlorophyll fluorescence—a practi-cal guide. J Exp Bot 51:659–668
Olšauskas A (1996) Influence of recreation on plant cover in WestLithuania. Klaipeda University, Klaipeda
Olšauskas AM, Urbonienė R (2008) State of Eryngium maritimum L.population on the Curonian spit coastal dunes. Env Res EngManage 2:69–74
The use of nondestructive methods in plant conservation biology 521

Öquist G, Chow WS, Anderson JM (1992) Photoinhibition ofphotosynthesis represents a mechanism for the long-term regu-lation of photosystem II. Planta 186:450–460
Samsone I, Andersone U, Vikmane M, Ieviņa B, Pakarna G, Ievinsh G(2007) Nondestructive methods in plant biology: an accuratemeasurement of chlorophyll content by a chlorophyll meter. ActaUniv Latv 723:145–154
Samsone I, Druva-Lūsīte I, Andersone U, Ņečajeva J, Karlsons A, IevinshG (2009) Plasticity of a dune plant Alyssum gmelinii in response tosand burial in natural conditions. Acta Univ Latv 753:125–136
Schreiber U (2004) Pulse-amplitude-modulation (PAM) fluorometryand saturation pulse method: an overview. In: Papageorgiou GC,Govindjee (eds) Chlorophyll a fluorescence: a signature ofphotosynthesis. Springer, Berlin, pp 279–319
Stasiak J (1986) The population size of Eryngium maritimum L. onthe Polish Baltic Sea coast. Biologia 7:65–75
Steinger T, Körner C, Schmid B (1996) Long-term persistence in achanging climate: DNA analysis suggests very old ages of clonesof alpine Carex curvula. Oecologia 105:94–99
Strauss AJ, Krüger GHJ, Strasser RJ, Van Heerden PDR (2006)Ranking of dark chilling tolerance in soybean genotypes probedby the chlorophyll a fluorescence transient O-J-I-P. Environ ExpBot 56:147–157
Thach LB, Shapcott A, Schmidt S, Critchley C (2007) The OJIP fastfluorescence rise characterizes Graptophyllum species and theirstress responses. Photosynth Res 94:423–436
Trouvelot A, Kough JL, Gianinazzi-Pearson V (1986) Mesure dutaux de mycorhization VA d’un système radiculaire. Recher-che de mèthodes d’estimation ayant une signification fonc-tionnelle. In: Gianinazzi-Pearson V, Gianinazzi S (eds)Physiological and genetical aspects of mycorrhizae. INRA,Paris, pp 217–221
Tsimilli-Michael M, Strasser RJ (2008) In vivo assessment of stressimpact on plant’s vitality: applications in detecting andevaluating the beneficial role of mycorrhization on hostplants. In: Varma A (ed) Mycorrhiza, 3rd edn. Springer,Berlin, pp 679–703
Turmel J-M (1947) Multiplication végétative des Eryngium. IIBouturage de l’Eryngium maritimum L. Bulletin de la SociétéBotanique de France 94:138–142
Urbanski JA (2001) The impact of sea-level rise along the PolishBaltic coast. J Coast Conserv 7:155–162
van der Maarel E, van der Maarel-Versluys M (1996) Distribution andconservation status of littoral vascular plant species along theEuropean coasts. J Coast Conserv 2:73–92
van der Valk AG (1974) Mineral cycling in coastal foredune plantcommunities un Cape Hatteras National Seashore. Ecology55:1349–1358
Żółkoś K, Afranowicz R, Bloch-Orlowska J, Koziel K (2007)Distribution and the resources of Sea Holly (Eryngium mar-itimum L.) on the western shore of the Gulf of Gdansk. BiodivRes Conserv 5–8:55–60
522 U. Andersone et al.
Copyright © 2022 FDOKUMEN




















