
The use of biostimulation and bioaugmentation to remove phenanthrene from soil
21
Int. J. Oil, Gas and Coal Technology, Vol. 3, No. 1, 2010 39 Copyright © 2010 Inderscience Enterprises Ltd. The use of biostimulation and bioaugmentation to remove phenanthrene from soil M.A. Amezcua-Allieri* Dirección de Seguridad y Medio Ambiente, Instituto Mexicano del Petróleo, Eje Central L. Cárdenas No. 152, Col. San B. Atepehuacan, C.P. 07730, Mexico, D.F., Mexico Fax: +5255-9175-8000 E-mail: [email protected] *Corresponding author A. Rodríguez-Dorantes Departamento de Botánica, ENCB, Instituto Politécnico Nacional, Prol. Carpio y Plan de Ayala s/n, C.P. 11340, Mexico, D.F., Mexico E-mail: [email protected] J. Meléndez-Estrada Departamento de Ciencias Básicas, Universidad Autónoma Metropolitana, Área de Química, Av. San Pablo No. 180, Col. Reynosa Tamaulipas, C.P. 02200, Mexico, D.F., Mexico E-mail: [email protected] Abstract: Biostimulation and bioaugmentation are commonly employed as technologies to remediate numerous environmental problems. In the last few decades, both technologies have been used to treat polyaromatic hydrocarbons (PAHs). This paper presents a brief survey of some bioremediation aspects, with particular reference to phenanthrene removal from soil, since this compound is the main model substrate for studies of carcinogenic PAHs. In addition, some cases of studies related are mentioned, emphasising the importance of the pollutant, oxygen concentration and type of microorganisms. The use of plant associated microorganisms for bioaugmentation is also described, emphasising the selection of suitable bacteria or communities obtained from plant rhizosphere, in order to improve phenanthrene bioremediation in soil. [Received: September 11, 2009; Accepted: December 10, 2009] Keywords: polyaromatic hydrocarbons; PAHs; phenanthrene; biostimulation; bioaugmentation; rhizobacteria; plant growth promoting rhizobacteria; PGPR; Penicillium frequntans; Cynodon dactylon; phytohormones; polymerase chain reaction; PCR.
Transcript of The use of biostimulation and bioaugmentation to remove phenanthrene from soil

Int. J. Oil, Gas and Coal Technology, Vol. 3, No. 1, 2010 39
Copyright © 2010 Inderscience Enterprises Ltd.
The use of biostimulation and bioaugmentation to remove phenanthrene from soil
M.A. Amezcua-Allieri* Dirección de Seguridad y Medio Ambiente, Instituto Mexicano del Petróleo, Eje Central L. Cárdenas No. 152, Col. San B. Atepehuacan, C.P. 07730, Mexico, D.F., Mexico Fax: +5255-9175-8000 E-mail: [email protected] *Corresponding author
A. Rodríguez-Dorantes Departamento de Botánica, ENCB, Instituto Politécnico Nacional, Prol. Carpio y Plan de Ayala s/n, C.P. 11340, Mexico, D.F., Mexico E-mail: [email protected]
J. Meléndez-Estrada Departamento de Ciencias Básicas, Universidad Autónoma Metropolitana, Área de Química, Av. San Pablo No. 180, Col. Reynosa Tamaulipas, C.P. 02200, Mexico, D.F., Mexico E-mail: [email protected]
Abstract: Biostimulation and bioaugmentation are commonly employed as technologies to remediate numerous environmental problems. In the last few decades, both technologies have been used to treat polyaromatic hydrocarbons (PAHs). This paper presents a brief survey of some bioremediation aspects, with particular reference to phenanthrene removal from soil, since this compound is the main model substrate for studies of carcinogenic PAHs. In addition, some cases of studies related are mentioned, emphasising the importance of the pollutant, oxygen concentration and type of microorganisms. The use of plant associated microorganisms for bioaugmentation is also described, emphasising the selection of suitable bacteria or communities obtained from plant rhizosphere, in order to improve phenanthrene bioremediation in soil. [Received: September 11, 2009; Accepted: December 10, 2009]
Keywords: polyaromatic hydrocarbons; PAHs; phenanthrene; biostimulation; bioaugmentation; rhizobacteria; plant growth promoting rhizobacteria; PGPR; Penicillium frequntans; Cynodon dactylon; phytohormones; polymerase chain reaction; PCR.

40 M.A. Amezcua-Allieri et al.
Reference to this paper should be made as follows: Amezcua-Allieri, M.A., Rodríguez-Dorantes, A. and Meléndez-Estrada, J. (2010) ‘The use of biostimulation and bioaugmentation to remove phenanthrene from soil’, Int. J. Oil, Gas and Coal Technology, Vol. 3, No. 1, pp.39–59.
Biographical notes: Myriam A. Amezcua-Allieri is a Researcher at the Instituto Mexicano del Petróleo. She received her BSc in Biology (first class honours) from the Universidad Nacional Autónoma de México (UNAM), Master in Science degree (Environmental Engineering) from the Instituto Politécnico Nacional and PhD (honours) in Geography and Environmental Sciences from The University of Birmingham, UK. She is a member of the Sistema Nacional de Investigadores since 2004. Her areas of research interest include bioremediation of soil contaminated with hydrocarbons and metals, contaminant bioavailability and speciation, and impact of petroleum industry in environment. She is an International Consultant of the United Nations Development Programme and a Full Member of the Third World Organization for Women in Science, Royal Geographical Society and Institute of British Geographers.
Angélica Rodríguez-Dorantes is a full time Professor at plant physiology area at the Botany Department of the Escuela Nacional de Ciencias Biológicas from the National Polytechnic Institute (IPN). She obtained her BSc in Biology and her PhD in Biological and Chemical Sciences from IPN. Her main areas of research interest are those related with phytoremediation, especially with plant response to contaminated environments with hydrocarbons and metals.
Jorge Meléndez-Estrada is a full time Professor at the Chemistry Department of the Autonomous Metropolitan University, Mexico city. He received his PhD in Biotechnology from CINVESTAV; Master in Sciences (Environmental Engineering) from IPN and BSc in Hydrobiology from Universidad Autónoma Metropolitana. His areas of interest include ecophysiology of potential microorganisms to be used in waste water treatment, development of biotechnical and biochemical processes applied to waste water treatment and the use of bacteria and fungi with potential to remove xenobiotic compounds from soils and water. He is a Research Candidate in the Sistema Nacional de Investigadores.
1 Biostimulation and bioaugmentation
Biostimulation and bioaugmentation are common technologies used to remediate sites with environmental problems. Biostimulation or enhanced biodegradation is the stimulation of microbial degradation of organic contaminants by the addition of nutrients for optimisation of environmental factors onsite or in situ (Riser-Roberts, 1998). Biostimulation involves the modification of the environment to stimulate existing bacteria capable of bioremediation. Bioaugmentation of polluted soil is an approach whereby microorganisms are amended to a contaminated site to hasten the degradation of pollutants. It is through the introduction of excess, active, degrader microorganisms that the pollutant is mineralised, thereby providing remediation at a faster rate than would otherwise occur by means of the indigenous microorganisms (Singer et al., 2005).
The use of native microbiota for bioremediation of contaminated soil is of great interest as these microorganisms would be expected to be more adapted to the particular

The use of biostimulation and bioaugmentation to remove phenanthrene 41
soil environment than non-indigenous commercial microbial inocula and would likely out-compete introduced microorganisms. However, when the natural microbiota does not show the ability to efficiently degrade the pollutant compounds, bioaugmentation, using introduced microorganisms with the desired degradation capability, is a possible means to enhance the success of the process (Serrano et al., 2009). We direct the reader to Abdulsalam and Omale (2009), Gentry et al. (2004), Mrozik and Piotrowska-Seget (2009), Singer et al. (2005), and Thomson et al. (2005) for reviews on biostimulation and bioaugmentation techniques.
The addition of nutrients can lead to an increased development of indigenous microorganisms, which themselves either biostimulate the process or hinder the process by consuming the added nutrient or carbon source (Scullion, 2006). Therefore, it is necessary to study both the addition of nutrients and microorganisms to remove persistent pollutants.
Some of the most important pollutants are the polyaromatic hydrocarbons (PAHs). The majority of these substances are highly persistent within ecosystems in unaltered, less degradable chemical forms because of their low water solubility, their intrinsic chemical stability, and high recalcitrance to degradation. Therefore, hydrocarbon-polluted sites may represent a long-term source of pollution and pose a severe risk to environmental health (Andreoni and Gianfreda, 2007).
2 Polyaromatic hydrocarbons as recalcitrant compounds in the environment
PAHs constitute a group of priority pollutants (WHO, 2001), which are present in the soils of many industrially contaminated sites, particularly those associated with the petroleum industry (Samanta et al., 2002). PAHs are hydrophobic compounds occurring in soil, sediment, water, air, and plants as the result of the incomplete combustion of coal, oil, petrol, and wood or of petrochemical industries.
Several hundred different combinations of PAHs exist, but up to 28 compounds have recently been identified as hazardous contaminants in 2008 by the US Environmental Protection Agency (USEPA, 2008). The distinguishing feature of these toxic compounds is that they are highly hydrophobic. As such, PAHs easily adsorb onto the organic matter of solid particles, forming persistent micropollutants in the environment (Gan et al., 2009). Air, soil, water and vegetation all act as environmental sinks for PAHs, although a preliminary inventory of PAHs in the UK environment showed that soil was the major repository for PAHs (Wild and Jones, 1995). Soils contaminated with PAHs pose potential risks to human and ecological health. The assessment of the actual risk for mutagenic and carcinogenic effects requires accurate exposure information which is extremely difficult to assess in practice (White and Claxton, 2004).
The chemical properties, and hence the environmental fate, of a PAH molecule are dependent in part upon both molecular size, i.e., the number of aromatic rings, and molecule topology or the pattern of ring linkage (Kanaly and Harayama, 2000).
Most PAHs occur as hybrids encompassing various structural components, such as in the PAH benzo[a]pyrene (BaP). Generally, an increase in the size and angularity of a PAH molecule results in a concomitant increase in hydrophobicity and electrochemical stability. PAH molecule stability and hydrophobicity are two primary factors which

42 M.A. Amezcua-Allieri et al.
contribute to the persistence of high-molecular-weight (HMW) PAHs in the environment (Kanaly and Harayama, 2000).
About chemical structure, phenanthrene is the smallest aromatic hydrocarbon to have a ‘bay-region’ and a ‘K-region’ (Riser-Roberts, 1998), therefore it is used in this study as a model substrate for studies on metabolism of carcinogenic PAHs because analogues of its structure are found in carcinogenic aromatic hydrocarbons (Samanta et al., 2002). In addition, the degradation of higher-ring PAHs may also proceed through a phenanthrene intermediate and phenanthrene increases the rate of degradation of highly mutagenic benzo(a)pyrene (Cerniglia, 1992).
2.1 Biodegradation of phenanthrene
The biodegradation of PAHs with less than three rings, which are often referred as low-molecular-weight (LMW) PAHs, has been well documented for bacteria and fungi (Zhang et al., 2006), specially for phenanthrene since it is a model substrate for studies on PAHs metabolism.
Phenanthrene is degraded by some soil bacteria through one of two different routes. In one route, 1-hydroxy-2-naphthoic acid is oxidised to 1,2-dihydroxynaphthalene, which is further degraded via the naphthalene pathway to salicylate, which can be further metabolised. In the other pathway, the ring of 1-hydroxy-2-naphthoic acid is cleaved and further metabolised via the phthalate pathway. It has been demonstrated that naphthalene and phenanthrene share a common upper metabolic pathway (Kiyohara et al., 1994). In addition, the metabolism of phenanthrene by Streptomyces flavovirens and the marine cyanobacteria Agmenellum quadruplicatum PR-6 is similar to that reported in mammalian and fungal enzyme systems than those catalysed by bacteria (Cerniglia, 1992). Both oxidise phenanthrene to phenanthrene trans-9,10-dihydrodiol via a monooxygenase-epoxide hydrolase-catalysed reaction rather than by a dioxygenase. The metabolic formation of 1-methoxyphenanthrene from phenanthrene was first reported in Synechococcus sp. PR-6. This organism may detoxify other 1-phenanthrols. The metabolic fate of 1-methoxyphenanthrene in Synechococcus sp. PR-6 remains to be elucidated (Ouyang, 2002).
Fungi play an important role in the hydrocarbon-oxidising activities of the soil, and they seem to be at least as versatile as bacteria in metabolising aromatics. Most of the fungal transformation products formed are less mutagenic than the parent compound (Riser-Roberts, 1998). Fungi (such as Penicillium and Cunninghamella) exhibit greater hydrocarbon biodegradation than bacteria. Fungi appear to be predominantly involved in degradation of those xenobiotics of lower water solubility and greater adsorptivity. Soil fungi are generally believed to play a more important role in the formation, metabolism, and interactions of soil-organic matter complexes than bacteria (Cerniglia, 1992). This is because fungi have various bonding mechanisms involved with adsorbed materials (Sutherland et al., 1995). In fact, most of the fungal transformation products formed by these fungi are less mutagenic than the parent compound (Riser-Roberts, 1998). A wide range of fungi have the enzymatic capacity to oxidise PAHs when grown on an alternative (to soil organic matter) carbon source (Cerniglia, 1992). For instance, Aspergillus and Penicillium are rich in hydrocarbon-assimilating strains (Sutherland et al., 1995).
Fungi can also metabolise phenanthrene non-ligninolytic cultures of Phanerochaete chrysosporium fit the typical eukaryotic pattern. They metabolise phenanthrene to

The use of biostimulation and bioaugmentation to remove phenanthrene 43
trans-dihydrodiols and phenanthrene conjugates. Phenanthrene trans-9,10- and trans-3,4-dihydrodiols are formed by the successive activities of monooxygenases and epoxide hydrolases. The regiospecificity of P. chrysosporium differs from that of Cunninghamella elegans, which principally produces the trans-1,2-dihydrodiol with smaller amounts of the trans-3,4-dihydrodiol. There was no metabolism at 9,10-positions (‘K-region’) of phenanthrene as evidenced by the failure to detect trans-9,10-dihydrodiols (Müncnerova and Augustin, 1994). The 3-phenanthrols were most likely produced either by dehydration of the trans-dihydrodiols or by rearrangement of the postulated arene oxides. Similarly, in Cunninghamella elegans, 1-phenanthrol was produced via dehydration of the corresponding trans-1,2-dihydrodiols. The novel 9-phenanthryl-beta-D-glucopyranoside produced by P. chrysosporium differs from the 1-phenanthryl-beta-D-glucopyranoside produced by C. elegans. Those conjugates may be considered to be detoxification products of phenanthrene (Ouyang, 2002).
Unlike non-ligninolytic Phanerochaete chrysosporium, ligninolytic P. chrysosporium does not accumulate trans-dihydrodiols and phenanthrols (Müncnerova and Augustin, 1994). It gives 2,2’-diphenic acid (DPA) as a major fate of phenanthrene by 9,10-oxidation and ring cleavage. These two reactions could involve multiple steps. However, no intermediates were reported so far. The oxidation of phenanthrene-9,10-quinone (PQ) to DPA involves both fungal and abiotic mechanisms, and is unaffected by the level of nitrogen added. Phenanthrene degradation by ligninolytic P. chrysosporium involves both ligninolytic and non-ligninolytic enzymes and is not initiated by a classical microsomal cytochrome P-450. In reaction, 2,2’-biphenyldimethanol occurred as a minor phenanthrene metabolite which was probably a reduction product of DPA (Ouyang, 2002). Both phenanthrene and PQ could be mineralised to similar extents by this kind of fungus.
3 The use of bioaugmentation and biostimulation for removing PAHs
Some studies have demonstrated that bioaugmentation does not result in increased PAHs remediation (e.g. Bouchez et al., 2000). Studies observed that the number of exogenous microorganisms decreases shortly after addition to a site due to both abiotic and biotic stresses. In this last it can be found that the added microorganisms almost always face competition from indigenous organisms for limited nutrients (Gentry et al., 2004). It can also be difficult to deliver the inoculant to the desired location. Fungi, which are larger than bacteria, are usually restricted to surface applications while bacteria are more adaptable to surface or subsurface applications (Andreoni and Gianfreda, 2007). However, most of the cases cited that bioaugmentation and biostimulation have been shown to increase degradation of numerous PAHs (Gentry et al., 2004).
Low-molecular-weight (LMW-) PAHs may affect the degradation of high-molecular-weight (HMW-) PAHs, and vice versa, via co-metabolism, induction of enzyme activities by related degradation intermediates or via competitive inhibition or toxicity (Dean-Ross et al., 2002). The metabolic cooperation by several microorganisms may result in enhanced PAH utilisation, since metabolic intermediates produced by some organisms may serve as substrates for the growth of others. Fungi and bacteria are capable of degrading partially or completely HMW-PAHs by co-metabolism when LMW-PAHs or the products of their degradation are present, as shown with individual

44 M.A. Amezcua-Allieri et al.
microbial isolates or by microbial consortia (Boonchan et al., 2000). Bioremediation has been shown to be effective in LMW-PAHs contaminated soil. In contrast, HMW-PAHs are more recalcitrant and typically require more time and in some cases different microorganisms to perform such degradation. In addition, mineralisation of HMW-PAHs decreases with increasing residence time in soil due to adsorption by soil organic matter. An effective bioremediation strategy depends on the ability of microorganism to degrade the target compounds towards mineralisation or to a very low level with production of non-toxic metabolites (Serrano et al., 2009).
Highly condensed PAHs, with four or more rings and their metabolites, have a variety of mutagenic and carcinogenic effects on microorganisms. However, many reports utilising adapted or isolated bacteria or lower fungi to degrade various, generally low-condensed PAHs, have been presented (Cerniglia, 1992; Wilson and Bouwer, 1997). Fungi have been demonstrated that are able to degrade PAHs under certain culture conditions, for instance, phenanthrene (Chávez-Gómez et al., 2003). Often, cooperative processes involve a consortium of strains with complementary capacities (Bouchez et al., 2000). Naphthalene, phenanthrene, anthracene, and pyrene are biodegraded through similar steps. Under aerobic conditions, bacteria and fungi utilise oxygen for both ring activation and cleavage of the aromatic nucleus and as the electron acceptor for its complete degradation (Andreoni and Gianfreda, 2007).
Serrano et al. (2009) reported that naphthalene, phenanthrene and anthracene and the HMW-PAH pyrene were removed more rapidly in all microcosms and to a greater extent than the HMW-PAH’s benz(a)anthracene and benz(a)pyrene. However, naphthalene removal reached a plateau at relatively high levels, between 25 and 29 mg kg–1 soil, suggesting naphthalene became less bioavailable than the other compounds at relatively low concentrations.
The utility of bioaugmentation is supported by studies showing the incapacity of indigenous microorganisms in some cases and the apparent enhanced bioremediation rate after the addition of competent microorganisms. The use of pure cultures into multi-substrate-polluted systems, such as soil, showed variable results (Bouchez et al., 2000). However, bioaugmentation using strains enriched from sites containing the target contaminant, where the population has acquired the catabolic ability, has been success (Schwartz et al., 2000). According to Serrano et al. (2009), bioaugmentation with Aspergillus fungal isolate did significantly increase the extent of removal of benz(a)anthracene and benz(a)pyrene, two rather persistent PAH’s in soils. This suggests that this organism may have some potential as a bioaugmentation agent. However, in all other microcosms bioaugmentation did not result in improved PAH removal rate or extent as has been indicated in some other studies. Inoculation levels of microorganisms, e.g. 107–108, are considered sufficient to establish degradation activity (Jacobsen and Petersen, 1992), and approximately 106 cells g–1 soil was considered sufficient inoculum to degrade PAHs in impacted soil (Boonchan et al., 2000). In the study of Serrano et al. (2009) it used lower bacterial cell densities (105 cells g–1 soil), and it is possible that a higher inoculum would result in enhancement of the degradation process by decreasing the lag phase and also the PAH depletion time relative to the natural microbiota performance.
The efficiency of bioaugmentation is determined by many biotic and abiotic factors (see Mrozik and Piotrowska-Seget, 2009 for further discussion). The first include the selection of proper microorganisms that can not only degrade contaminants but can also successfully compete with indigenous microflora (Mrozik and Piotrowska-Seget, 2009).

The use of biostimulation and bioaugmentation to remove phenanthrene 45
In turn, among abiotic factors, the chemical structure, concentration and availability of pollutants as well as physico-chemical properties of soil are also important. Moisture content, temperature, type and balance of nutrients, electron receivers and soil gas oxygen concentrations are the main abiotic factors that need to be considered (Rogers et al., 1993). However, the soil gas oxygen concentration is generally thought to be one of the limiting constituents in PAHs biodegradation, not only in its role as electron acceptor during microbial respiration but also because it serves as co-substrate of oxygenase enzymes that initiate PAHs catabolism (Hurst et al., 1996).
4 Phenanthrene removal at different O2 concentrations
Due to the importance of oxygen in the dynamics of the aerobic processes in soil, many studies have been carried out to investigate the effect of oxygen concentration in the sequestration and bioavailability of diverse aromatic hydrocarbons in soil (Chung and Alexander, 1999). Oxygen concentration varies according to soil depth and its availability depends on the oxygen diffusion and consumption rates, which in turn depend on the type of soil and the presence of bioavailable substrates. These factors can therefore affect the efficacy of aerobic remediation processes of PAHs-contaminated soils.
Porosity is related to the process of oxygen diffusion in solid medium, which enhances considerably the air diffusion, increasing the gas exchange necessary for the metabolic aerobic process. The addition of agroindustrial residues materials (bulking agents) in aerobic solid-state cultures offers several advantages for the treatment of hydrocarbon-contaminated soils (Roldán-Martín, 2006). However there is a lack of information about the effect of oxygen concentrations and the process performance in the bioremediation potential in solid-state cultures under microaerophilic conditions (Kästner and Mahro, 1996). Nevertheless, these bulking agents also contribute to the oxygen demand of the system, which is conducive to the development of microaerophilic niches. Thus, it is important to evaluate the metabolic activity and PAHs degradation capabilities under varying oxygen concentrations. Because of the reasons described in Section 2, phenanthrene was used as a model PAH to illustrate the effect of oxygen on its removal from soil.
4.1 Fungal inocula preparation in solid-state culture
The fungus Penicillium frequentans has been identified as a suitable microorganism for phenanthrene removal from soil under certain solid-state culture conditions (Amezcua-Allieri et al., 2003). Fungal inoculum was added to sterilised soil which was previously spiked with a phenanthrene solution, in order to get a final concentration of 200 mg kg–1 of phenanthrene. Subsequently the inoculum was mixed in soil until obtaining a perfectly homogeneous distribution of sugar cane bagasse pith and nutrients in the solid matrix. The oxygen concentration was adjusted by a daily supply of oxygen and nitrogen, to obtain a final concentration (%) of 21, 10, 5, 2 and nearer to 0 (Henry’s constant for oxygen = 760 atm-L mol–1). Oxygen concentrations (%) were adjusted to 2, 5, 10, and 21, inside the soil gas phase, variations among treatments were lower than
46 M.A. Amezcua-Allieri et al.
±1% (p < 0.005). The dependent variables were: CO2 evolution, O2 concentration and phenanthrene removal on a dry weight base. Average oxygen concentration was calculated, in each treatment, twice a day, before and after oxygen adjustment. Heterothrophic activity was measured by the indirect method of CO2 evolution and quantified as mg CO2 kg–1 of initial dry matter (Mitchell, 1992). O2 consumption and CO2 concentration were measured in the headspace before and after treatment daily.
Soil samples were removed from each bottle and extracted in a Soxhlet system using ketone as the solvent, for 8 h (EPA-3540). Soil extracts were analysed quantitatively by HPLC VARIAN 9012 equipment, fitted with a capillary column (Res Elut 5u C18 9090Å, 4.6 mm id, 150 mm length) equipped with a UV/VIS 9050 detector. Samples were analysed at 240 nm, methanol: water was used as the mobile phase in a 90:10 ratio and a flow velocity of 1.0. Phenanthrene removal rates at different O2 concentrations were measured to determine the effect of oxygen concentration on the ability of the fungus to remove it.
4.2 CO2 evolution and oxygen concentration
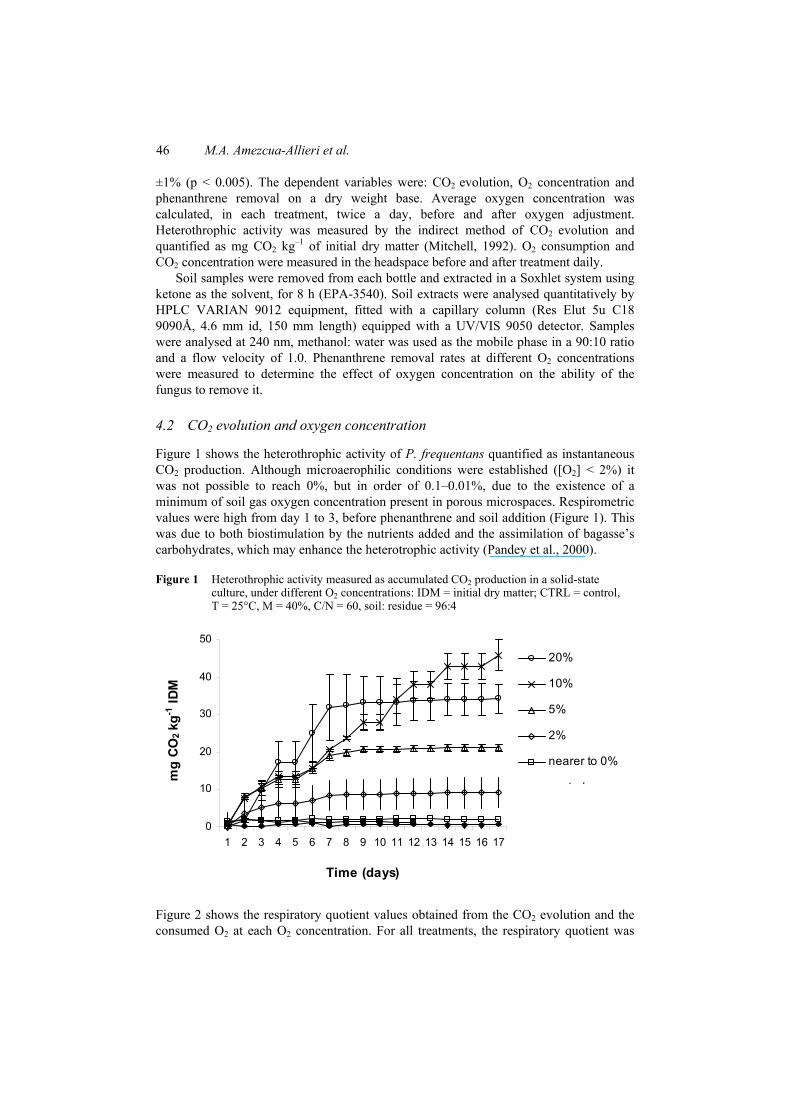
Figure 1 shows the heterothrophic activity of P. frequentans quantified as instantaneous CO2 production. Although microaerophilic conditions were established ([O2] < 2%) it was not possible to reach 0%, but in order of 0.1–0.01%, due to the existence of a minimum of soil gas oxygen concentration present in porous microspaces. Respirometric values were high from day 1 to 3, before phenanthrene and soil addition (Figure 1). This was due to both biostimulation by the nutrients added and the assimilation of bagasse’s carbohydrates, which may enhance the heterotrophic activity (Pandey et al., 2000).
Figure 1 Heterothrophic activity measured as accumulated CO2 production in a solid-state culture, under different O2 concentrations: IDM = initial dry matter; CTRL = control, T = 25°C, M = 40%, C/N = 60, soil: residue = 96:4
0
10
20
30
40
50
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
Time (days)
mg
CO
2 kg
-1 ID
M
20%
10%
5%
2%
nearer to 0%
t l
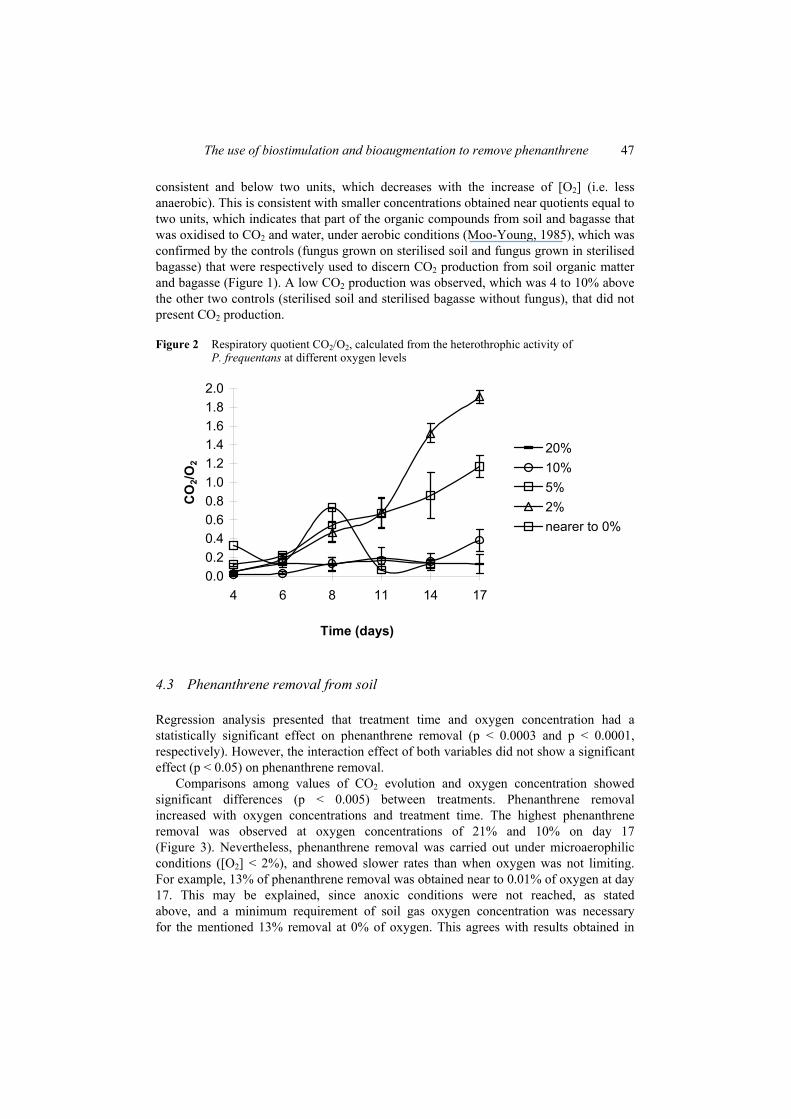
Figure 2 shows the respiratory quotient values obtained from the CO2 evolution and the consumed O2 at each O2 concentration. For all treatments, the respiratory quotient was
The use of biostimulation and bioaugmentation to remove phenanthrene 47
consistent and below two units, which decreases with the increase of [O2] (i.e. less anaerobic). This is consistent with smaller concentrations obtained near quotients equal to two units, which indicates that part of the organic compounds from soil and bagasse that was oxidised to CO2 and water, under aerobic conditions (Moo-Young, 1985), which was confirmed by the controls (fungus grown on sterilised soil and fungus grown in sterilised bagasse) that were respectively used to discern CO2 production from soil organic matter and bagasse (Figure 1). A low CO2 production was observed, which was 4 to 10% above the other two controls (sterilised soil and sterilised bagasse without fungus), that did not present CO2 production.
Figure 2 Respiratory quotient CO2/O2, calculated from the heterothrophic activity of P. frequentans at different oxygen levels
0.00.20.40.60.81.01.21.41.61.82.0
4 6 8 11 14 17
Time (days)
CO
2/O2
20%10%5%2%nearer to 0%
4.3 Phenanthrene removal from soil
Regression analysis presented that treatment time and oxygen concentration had a statistically significant effect on phenanthrene removal (p < 0.0003 and p < 0.0001, respectively). However, the interaction effect of both variables did not show a significant effect (p < 0.05) on phenanthrene removal.
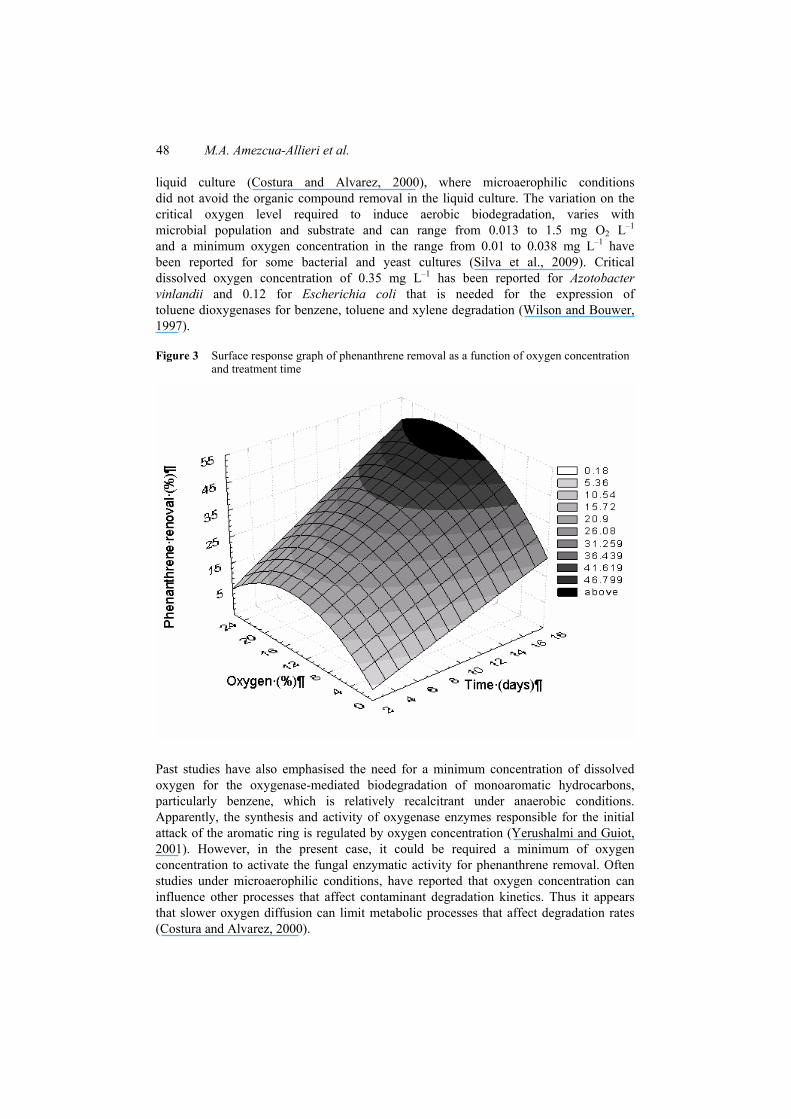
Comparisons among values of CO2 evolution and oxygen concentration showed significant differences (p < 0.005) between treatments. Phenanthrene removal increased with oxygen concentrations and treatment time. The highest phenanthrene removal was observed at oxygen concentrations of 21% and 10% on day 17 (Figure 3). Nevertheless, phenanthrene removal was carried out under microaerophilic conditions ([O2] < 2%), and showed slower rates than when oxygen was not limiting. For example, 13% of phenanthrene removal was obtained near to 0.01% of oxygen at day 17. This may be explained, since anoxic conditions were not reached, as stated above, and a minimum requirement of soil gas oxygen concentration was necessary for the mentioned 13% removal at 0% of oxygen. This agrees with results obtained in
48 M.A. Amezcua-Allieri et al.
liquid culture (Costura and Alvarez, 2000), where microaerophilic conditions did not avoid the organic compound removal in the liquid culture. The variation on the critical oxygen level required to induce aerobic biodegradation, varies with microbial population and substrate and can range from 0.013 to 1.5 mg O2 L–1 and a minimum oxygen concentration in the range from 0.01 to 0.038 mg L–1 have been reported for some bacterial and yeast cultures (Silva et al., 2009). Critical dissolved oxygen concentration of 0.35 mg L–1 has been reported for Azotobacter vinlandii and 0.12 for Escherichia coli that is needed for the expression of toluene dioxygenases for benzene, toluene and xylene degradation (Wilson and Bouwer, 1997).
Figure 3 Surface response graph of phenanthrene removal as a function of oxygen concentration and treatment time
Past studies have also emphasised the need for a minimum concentration of dissolved oxygen for the oxygenase-mediated biodegradation of monoaromatic hydrocarbons, particularly benzene, which is relatively recalcitrant under anaerobic conditions. Apparently, the synthesis and activity of oxygenase enzymes responsible for the initial attack of the aromatic ring is regulated by oxygen concentration (Yerushalmi and Guiot, 2001). However, in the present case, it could be required a minimum of oxygen concentration to activate the fungal enzymatic activity for phenanthrene removal. Often studies under microaerophilic conditions, have reported that oxygen concentration can influence other processes that affect contaminant degradation kinetics. Thus it appears that slower oxygen diffusion can limit metabolic processes that affect degradation rates (Costura and Alvarez, 2000).

The use of biostimulation and bioaugmentation to remove phenanthrene 49
5 Bioaugmentation and plant associated microorganisms
Bioaugmentation has been utilised in agriculture for many years and the applications of it includes inoculation of plant seeds with plant growth promoting microorganisms or with plant-protecting microorganisms that are antagonistic to plant pathogens (Gentry et al., 2004).
A developing approach for bioaugmentation is to add the microbial inoculants to the soil along with a plant that supports the inoculant’s growth. The use of plants for remediation, or phytoremediation, is a relatively new technology that compared with physical and chemical remediation, has several advantages: preserves the natural properties of soil, acquires energy mainly from sunlight, high levels of microbial biomass in the rhizosphere can be achieved, it is low in cost and it has the potential to be rapid (Huang et al., 2005). Organic pollutants are transformed or degraded by plant-associated microorganisms (Karthikeyan and Kulakow, 2003). The rhizosphere, or zone of soil under the influence of plant roots, is well known as a site of elevated microbial numbers and activity and additionally, different plant roots select for different rhizosphere populations (Kowalchuk et al., 2002).
In natural environments, many of the limitations to remediation can be overcome by the dynamic synergy that exists between plant roots and soil microorganisms (El-Shatnawi and Makhadmeh, 2001). The microbial population and diversity in a rhizosphere may, however, vary with plant species, environmental factors and nutrient status of the soil. For a microbial-plant synergy to be effective, a soil microbial population must contain species that are capable of transforming soil-bound and recalcitrant organic compounds. An adaptation and selection within the micro habitats may take place and a population may change considerably with time in response to the type and degree of a contamination (Chaudhry et al., 2005).
5.1 Role of rhizospheric microorganisms in the soil remediation with petroleum contaminants
An important contribution to the degradation of pollutants is ascribed to microbes present in the rhizosphere of plants used during phytoremediation or of plants which are emerging as natural vegetation on a contaminated site. This contribution of the rhizomicrobial population is referred to as rhizoremediation (Schwab and Banks, 1994). Studies of the most suitable plant species for rhizoremediation showed that various grass varieties and leguminous plants such as alfalfa are suitable (Kuiper et al., 2004), probably due to their ability to harbour large numbers of bacteria on their highly branched root systems.
Enhanced rhizosphere biodegradation takes place in the soil immediately surrounding plant roots (Leigh et al., 2004, 2006). Natural substances released by plant roots supply nutrients to microorganisms, which enhances their biological activities and stimulate the degradation of organic chemicals by inducing enzyme systems of existing bacterial populations. Both plants and soil microorganisms have certain limitations with respect to their individual abilities to remove/breakdown organic compounds (Villacieros et al., 2005). A synergistic action of both rhizosphere microorganisms and plants can lead to increased availability of hydrophobic compounds affecting their removal and/or degradation (Mackova et al., 2007).

50 M.A. Amezcua-Allieri et al.
The hydrophobic nature of most petroleum compounds, severely limits their uptake by plants. A number of studies, however, suggest that microbial strains capable of breaking down petroleum hydrocarbons are widely present amongst soil microorganisms; the presence of some plant species seems to select and enrich these strains in the rhizosphere soils (Chaudhry et al., 2005).
Efforts to demonstrate plant or rhizosphere effects on petroleum degradation, particularly PAHs, continue, the measure of the plant effects on PAH degradation in soil and the numerous interactions among the soil-rhizosphere-microbial-plant system were done. Parts of the system interact on a continuous basis; interactions vary with conditions, such as moisture and temperature, as well as with the growth stage of both plants and microorganisms. These interactions make understanding and monitoring phytoremediation complex. Examples of the understanding of rhizosphere microbiology and its changes during phytoremediation are the works of Siciliano et al. (2003) with the observation of greater levels of hydrocarbon-related catabolic genes (ndoB, alkB and xylE) in rhizosphere soils relative to bulk soil. Muratova et al. (2003) found differences between alfalfa (Medicago sativa) and reed (Phragmites australis) rhizospheres and the alfalfa enhanced effect on the PAH degrading population in the rhizosphere. Joner et al. (2002) and Joner and Leyval (2003) found in a pot study that in PAH-contaminated soil planted with a clover–ryegrass mix PAH concentrations decreased as a function of proximity to the roots, and that mycorrhiza generally enhanced plant growth and favoured growth of clover at the expense of ryegrass. Wittig et al. (2003) and Ballach et al. (2003) found that poplar cuttings grown in a PAH amended sand-nutrient solution had similar shoot biomass, growth and leaf water content to the controls, but that transpiration, nutrient solution uptake, and root mass were reduced. Finally, the effects were also dependent on the PAH under study (Newman and Reynolds, 2004).
Microbe-assisted phytoremediation, with both naturally occurring microbes and deliberately stimulated via seed inoculation, has been investigated in the laboratory, greenhouse and field (Huang et al., 2005, Zhuang et al., 2007) for phenanthrene (Liste and Alexander, 2000). This microbial consortia can provide various benefits to plants, including the synthesis of compounds that protect the plants by decreasing plant stress hormone levels; chelators for delivering key plant nutrients; protection against plant pathogens; and degradation of contaminants before they can negatively impact the plants (Liu et al., 2007).The successful application of rhizoremediation is largely dependent on the capacity of contaminant degraders or plant growth promoting microbes to efficiently colonise growing roots (Gerhardt et al., 2009).
5.2 Plant growth promoting rhizobacteria as a tool for rhizoremediation
Although rhizoremediation occurs naturally, it can also be optimised, by using suitable plants and microbes; these can be either combinations of plants and plant growth promoting rhizobacteria (PGPR); bacteria capable of promoting plant growth by colonising the plant root, or combinations of plants and contaminant-degrading microbes (Khan, 2005).
Direct plant growth promoting mechanisms may involve production of plant growth regulators, nitrogen fixation and the mobilisation of unavailable nutrients such as phosphorus and other mineral nutrients. In addition to their beneficial effects on plant

The use of biostimulation and bioaugmentation to remove phenanthrene 51
growth, plant associated microorganisms improve the efficient degradation of organic compounds. These microbial associations increase the capacity for a stepwise transformation of organic contaminants by consortia and provide a habitat that is conducive to genetic exchange and gene rearrangements (Weyens et al., 2009).
PGPR can enhance germination and plant growth rates under stress conditions, particularly when used in conjunction with contaminant-tolerant plant species (Glick et al., 2007). Notably, PGPR promote rapid accumulation of root biomass that can provide a sink for soil contaminants (Huang et al., 2005). Many of these rhizobacteria (e.g., Pseudomonas spp.) can act both as plant growth promoters and contaminant degraders (Cherian and Oliveira, 2005). Thus, it is likely that PGPR could contribute to phytoremediation processes via multiple modes of action. PGPR-enhanced phytoremediation systems have been developed that appear to overcome the challenge of plant stress in contaminated soils, Huang et al. (2005) used grass and grain seeds (Festuca arundinacea, Lolium multiflorum, Secale cereal, Pennisetum glaucum, Hordeum vulgare, Poa pratensis, and Elymus canadensis) treated with various naturally occurring, non-pathogenic Pseudomonas strains; soils containing organic compounds have been successfully remediated.
The importance of plant–microbe partnerships in the remediation of organic contaminants was confirmed in studies at the level of the rhizosphere (Ho et al., 2007; Kidd et al., 2008) the phyllosphere (Sandhu et al., 2007), and inside the plant (Barac et al., 2009; Taghavi et al., 2005; Weyens et al., 2009).
According to Huang et al. (2004a, 2004b), the addition of PGPR increased the organic pollutant (polycyclic aromatic hydrocarbon and creosote) removal probably by enhancing plants germination and survival in soils that were heavily contaminated and by stimulating the plants to grow faster and accumulate more root biomass. The combination of PGPR and specific contaminant-degrading bacteria was found to be effective (Ajithkumar et al., 1998). Huang et al. (2005) thus developed a multi-process phytoremediation system (MPPS). They used both PGPR and specific contaminant-degrading bacteria to treat TPHs. In this system, specific contaminant-degrading bacteria can be selected according to the properties of contaminants. They can rapidly metabolise some readily available compounds while the role of PGPR is still prompting plant growth and increasing the plant tolerance to pollutants (Zhuang et al., 2007).
5.3 Evaluation of the growth response and tolerance to phenanthrene of Bermuda grass, inoculated with two phytohormone producer rhizobacteria
Recently, Guerrero-Zuñiga et al. (2008) evaluated the growth response and the tolerance to phenanthrene of Bermuda grass: Cynodon dactylon inoculated with two phytohormone producer rhizobacteria, selected and named II and III strains of the 23 rhizobacteria strains isolated from rhizospheric samples of petroleum hydrocarbons-contaminated soil, collected especially around the root debris. These strains were maintained and conserved on nutritive agar plates for the conventional bacterial analyses and they were characterised and identified as Bacillus sp. strains based on its 16S rDNA sequence homology Basic Local Alignment Search Tool (BLAST) analysis. The conventional analysis of its microscopical morphology revealed short positive Gram bacilli and sporulate bacteria.

52 M.A. Amezcua-Allieri et al.
5.4 Evaluation of the response of Cynodon dactylon inoculated with the phytohormone producing rhizobacteria Bacillus sp. strains II and III exposed to phenanthrene in hydroponic cultures
Warm-climate grasses like Bermuda grass (Cynodon dactylon L.) and other members of the Poaceae family were employed for remediation and restoration of petroleum polluted soils (EPA, 2000; Martínez et al., 2000). It has been shown that soils polluted with petroleum or its fractions, if vegetated with Poaceae plants, are more quickly remediated and the toxicity of the soil at the end of the treatment decreases (Günter et al., 1996). In addition, it has been demonstrated that the enrichment of bacteria in soil adjacent to plant roots is responsible for bulk pollutant degradation; this is known as the ‘rhizosphere effect’ (Rivera-Cruz et al., 2004). Recently, it was observed that bacterial enrichment is not only in number, but there is also a selection of microbial degradative genotypes, promoted by the grass root and the pollutant (Siciliano et al., 2003; Peña Castro et al., 2006).
Commercially obtained seeds of Cynodon dactylon were surface-sterilised and incubated for 30 minutes at room temperature either in sterile distilled water (blank control) or in a bacterial suspension with phytohormones produced by rhizobacteria Bacillus sp. strains II and III. Twenty seeds of each species treated and non treated with bacteria, were placed separately with Magenta Sigma caps with 25 mL of mineral medium and 6% bacteriological agar; supplemented in each case 100 and 200 mg L–1 of phenanthrene concentrations and the control experiments were considerate without the addition of the contaminant. All the experiments were performed by triplicate and maintained at 30°C in a growth chamber with photoperiod of 12:12, for 28 days. The plants were harvested 28 days after the inoculation treatments, the length of shoots and roots were measured at the end and the growth of plants were evaluated by the tolerance index (TI), expressed as the ratio of the shoot and root lengths of plants grown in the presence and absence of phenanthrene. All the results were analysed by ANOVA test, and Tukey-Kramer method using the statistics program Graph Pad Instat (Ver. 2.03) and the experimental designs program FAUNAL (Ver. 1.4).
5.5 Growth of Cynodon dactylon inoculated with the phytohormone producing Bacillus sp. strains II and III
The effect of phenanthrene on plants of C. dactylon showed that this species diminished its growth when the contaminant is present, without the inoculants. When the plants were inoculated with the two rhizobacteria, a particularly response has been showed, since each one was capable to promote the development of the plants.
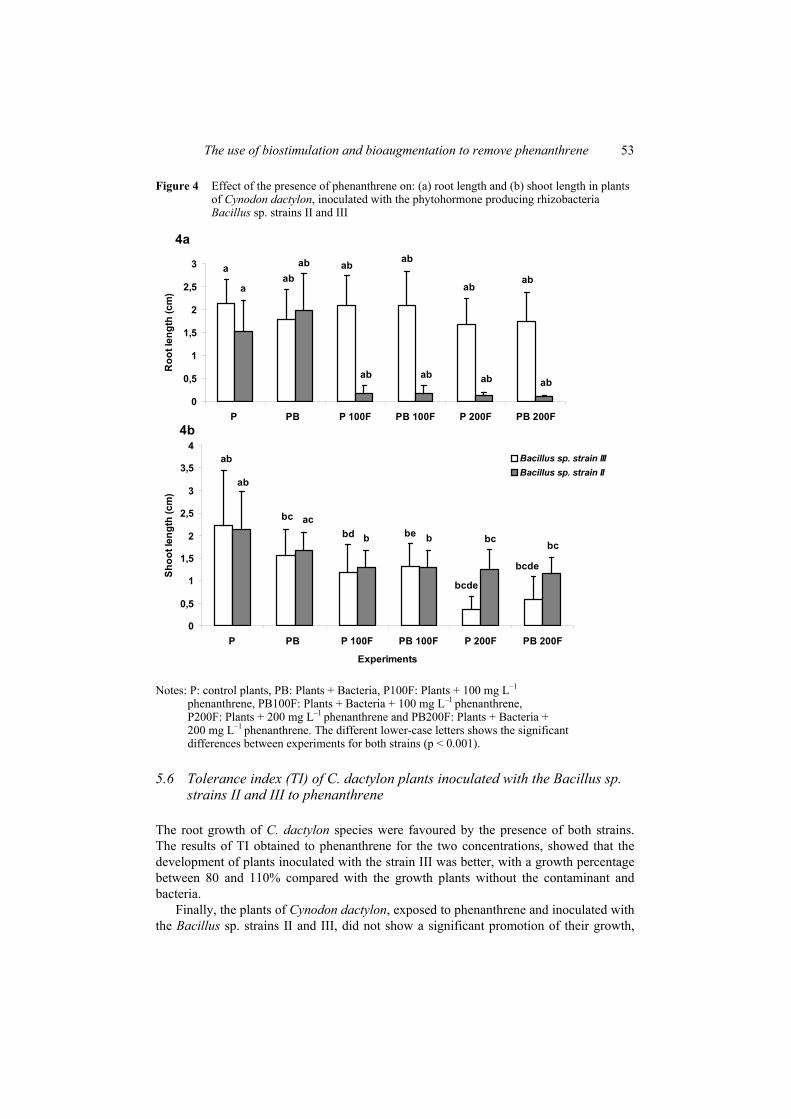
The growth of the roots of C. dactylon was favoured by the presence of the rhizobacteria III; the results obtained showed that the increase of the root length was evident in the experiments with phenanthrene with the rhizobacteria [Figure 4(a)].
For the shoots, the presence of both rhizobacteria promote the maintenance of the development of the C. dactylon plants, when the phenanthrene was present in the experiments and the effect of the protection on plants was evident for the rhizobacteria II, in the experiments with the highest concentration of phenanthrene [Figure 4(b)]. The response of C. dactylon exposed to the contaminant, showed a good tolerance to the phenanthrene concentrations probed; in this case the effect was not inhibitory at all with respect to the control plants, as the concentration of the contaminant increased.
The use of biostimulation and bioaugmentation to remove phenanthrene 53
Figure 4 Effect of the presence of phenanthrene on: (a) root length and (b) shoot length in plants of Cynodon dactylon, inoculated with the phytohormone producing rhizobacteria Bacillus sp. strains II and III
abab
ababab
a
abababab
ab
a
0
0,5
1
1,5
2
2,5
3
P PB P 100F PB 100F P 200F PB 200F
Roo
t len
gth
(cm
)
bcde
bcde
bebd
bc
ab
bcbcbb
ac
ab
0
0,5
1
1,5
2
2,5
3
3,5
4
P PB P 100F PB 100F P 200F PB 200F
Experiments
Shoo
t len
gth
(cm
)
Bacillus sp. strain IIIBacillus sp. strain II
4b
4a
Notes: P: control plants, PB: Plants + Bacteria, P100F: Plants + 100 mg L–1
phenanthrene, PB100F: Plants + Bacteria + 100 mg L–1 phenanthrene, P200F: Plants + 200 mg L–1 phenanthrene and PB200F: Plants + Bacteria + 200 mg L–1 phenanthrene. The different lower-case letters shows the significant differences between experiments for both strains (p < 0.001).
5.6 Tolerance index (TI) of C. dactylon plants inoculated with the Bacillus sp. strains II and III to phenanthrene
The root growth of C. dactylon species were favoured by the presence of both strains. The results of TI obtained to phenanthrene for the two concentrations, showed that the development of plants inoculated with the strain III was better, with a growth percentage between 80 and 110% compared with the growth plants without the contaminant and bacteria.
Finally, the plants of Cynodon dactylon, exposed to phenanthrene and inoculated with the Bacillus sp. strains II and III, did not show a significant promotion of their growth,

54 M.A. Amezcua-Allieri et al.
but the presence of these phytohormone producing rhizobacteria, helped to the protection, maintenance of development of this plant species studied, and on the tolerance to the organic contaminant. The promotion of the root and shoot lengths were evident in a separate way, rhizobacteria Bacillus sp. strain II promoted only the root growth, while rhizobacteria Bacillus sp. strain III did it with the root and shoot growth. This response could be related with the fact that some plants can release structural analogues of PAHs, such as phenols, to promote the growth of microbes (Fletcher and Hedge, 1995; Zhuang et al., 2007), and in this case favoured the growth and maintenance of the in vitro plant development.
Although this study describes data for only one pollutant and for one plant-bacterium combination; it is clear that similar microbe-plant combinations can be selected for introducing rhizosphere strains together with a suitable plant by coating bacteria on plant seed, so these well-equipped bacteria might settle on the root and could enhancing the bioremediation process for another kind of PAH pollutants.
6 Conclusions
In order to determine the potential success of bioaugmentation and biostimulation it is necessary to understand the bioavailability of the pollutant, the survival and activity of the added microorganisms, and the general environmental conditions that control soil bioremediation rates. With available genetic techniques, the comprehension of the fate of added microorganisms and of the critical parameters such as nutrients and oxygen, can be achieved. It has been illustrated that microaerophilic conditions did not preclude phenanthrene removal efficiencies, while agricultural residues can be a support and carbon source for microbial growth, can increase the porosity and surface area, which could enhance both the gas diffusion and adsorption of organic compounds.
Despite bioaugmentation is still a source of controversy within environmental microbiology, it is considered as a way of enlarging the genetic capacity present in a given site, thus corresponding to an increase in the gene pool and in the genetic diversity of that site. Therefore, further studies about the selection of suitable rhizosphere bacteria or communities, able to sustain and proliferate on the plant root system, are suitable tools to improve and develop bioremediation techniques.
Acknowledgements
Financial and material support for this study was provided by CINVESTAV-IPN. M. Amezcua-Allieri and J. Meléndez-Estrada are grateful for SNI fellowship. A. Rodríguez-Dorantes is grateful to the Secretaría de Investigación y Posgrado-IPN for the financial support to the project SIP20080061 and to the Comisión de Operación y Fomento de Actividades Académicas (COFAA) for its support throughout the development of this research work.
References Abdulsalam, S. and Omale, A.B. (2009) ‘Comparison of biostimulation and bioaugmentation
techniques for the remediation of used motor oil contaminated soil’, Brazilian Archives of Biology and Technology, Vol. 52, No. 3, pp.747–754.

The use of biostimulation and bioaugmentation to remove phenanthrene 55
Ajithkumar, P.V., Gangadhara, K.P., Manilal, P. and Kunhi, A.A.M. (1998) ‘Soil inoculation with Pseudomonas aeruginosa 3MT eliminates the inhibitory effect of 3-chloro and 4-chlorobenzoate on tomato seed germination’, Soil Biology and Biochemistry, Vol. 30, pp.1053–1059.
Amezcua-Allieri, M.A., Lead, J.R., Meléndez-Estrada, J. and Rodríguez-Vázquez, R. (2003) ‘Phenanthrene removal in a selected Mexican soil by the fungus Penicillium frequentans: role of C:N ratio and water content’, Soil & Sediment Contamination, Vol. 12, No. 3, pp.387–399.
Andreoni, V. and Gianfreda, L. (2007) ‘Bioremediation and monitoring of aromatic-polluted habitats’, Applied Microbiology and Biotechnology, Vol. 76, pp.287–308.
Ballach, H.J., Kuhn, A. and Wittig, R. (2003) ‘Biodegradation of anthracene in the roots and growth substrate of poplar cuttings’, Environmental Science and Pollution Research International, Vol. 10, pp.308–316.
Barac, T., Weyens, N., Oeyen, L., Taghavi, S., van der Lelie, D., Dubin, D., Split, M. and Vangronsveld, J. (2009) ‘Application of poplar and its associated microorganisms for the in situ remediation of a BTEX contaminated groundwater plume’, International Journal of Phytoremediation, Vol. 11, pp.416–424.
Boonchan, S., Britz, M.L. and Stanley, G.A. (2000) ‘Degradation and mineralization of high-molecular-weight polycyclic aromatic hydrocarbons by defined fungal– bacterial cocultures’, Applied and Environmental Microbiology, Vol. 66, pp.1007–1019.
Bouchez, T., Patureau, D., Dabert, P., Juretschko, S., Dore, J., Delgenes, P., Moletta, R. and Wagner, M. (2000) ‘Ecological study of a bioaugmentation failure’, Environmental Microbiology, Vol. 2, p.179.
Cerniglia, C.E. (1992) ‘Biodegradation of polycyclic aromatic hydrocarbons’, Biodegradation, Vol. 3, pp.351–368.
Chaudhry, Q., Blom-Zandstra, M., Gupta, S. and Joner, E.J. (2005) ‘Utilizing the synergy between plants and rhizosphere microorganisms to enhance breakdown of organic pollutants in the environment’, Environmental Science and Pollution Research, Vol. 12, pp.34–48.
Chávez-Gómez, B., Quintero, R., Esparza-García, F., Mesta-Howard, A.M., Zavala Díaz de la Serna, F.J., Hernández-Rodríguez, C.H., Gillén, T., Poggi-Varaldo, H.M., Barrera-Cortés, J. and Rodríguez-Vázquez, R. (2003) ‘Removal of phenanthrene from soil by co-cultures of bacteria and fungi pregrown on sugarcane bagasse pith’, Bioresource Technology, Vol. 89, No. 2, pp.177–183.
Cherian, S. and Oliveira, M. M. (2005) ‘Transgenic plants in phytoremediation: recent advances and new possibilities’, Environmental Science & Technology, Vol. 39, pp.9377–9390.
Chung, N. and Alexander M. (1999) ‘Effect of concentration on sequestration and bioavailability of two polycyclic aromatic hydrocarbons’, Environmental Science and Technology, Vol. 33, No. 20, pp.3605–3608.
Costura, R.K. and Alvarez, P.J. (2000) ‘Expression and longevity of toluene dioxygenase in Pseudomonas putida F1 induced at different oxygen concentrations’, Water Research, Vol. 34, No. 11, pp.3014–3018.
Dean-Ross, D., Moody, J. and Cerniglia, C.E. (2002) ‘Utilization of mixtures of polycyclic aromatic hydrocarbons by bacteria isolated from contaminated sediment’, FEMS Microbiology Ecology, Vol. 41, pp.1–7.
El-Shatnawi, M.K.J. and Makhadmeh, I.M. (2001) ‘Ecophysiology of the plant-rhizosphere system’, Journal of Agronomy and Crop Science, Vol. 187, pp.1–9.
Environmental Protection Agency (EPA) (2000) ‘Introduction to Phytoremediation’, Office of Research and Development, USA, p.104.
Fletcher, J.S. and Hedge, R.S. (1995) ‘Release of phenols by perennial plant roots and their potential importance in bioremediation’, Chemosphere, Vol. 31, pp.3009–3016.
Gan, S., Lau, E.V. and Ng, H.K. (2009) ‘Remediation of soils contaminated with polycyclic aromatic hydrocarbons (PAHs)’, Journal of Hazardous Materials, Vol. 172, pp.532–549.

56 M.A. Amezcua-Allieri et al.
Gentry, T.J., Rensing, C. and Pepper, I. (2004) ‘New approaches for bioaugmentation as a remediation technology’, Critical Reviews in Environmental Science and Technology, Vol. 34, pp.447–494.
Gerhardt, K.E., Huang, X.D., Glick, B.R. and Greenberg, B.M. (2009) ‘Phytoremediation and rhizoremediation of organic contaminants: potential and challenges’, Plant Science, Vol. 176, pp.20–30.
Glick, B.R., Cheng, Z., Czarny, J. and Duan, J. (2007) ‘Promotion of plant growth by ACC deaminase-producing soil bacteria’, European Journal of Plant Pathology, Vol. 119, pp.329–339.
Guerrero-Zuñiga, A., Rojas-Contreras, A., Rodríguez-Dorantes, A. and Montes-Villafán, S. (2008) ‘Effect of two phytohormone producer rhizobacteria on the Bermuda grass growth response and tolerance to phenanthrene’, III Strategies for the Protection and Remediation of Natural Environment, Proceedings of the 3rd International Meeting on Environmental Biotechnology and Engineering, Palma de Mallorca, Spain, p.241.
Günter, T., Dornberger, U. and Fritsche, W. (1996) ‘Effects of ryegrass on biodegradation of hydrocarbons in soil’, Chemosphere, Vol. 33, pp.203–215.
Ho, C.H., Applegate, B. and Banks, M.K. (2007) ‘Impact of microbial/plant interactions on the transformation of polycyclic aromatic hydrocarbons in rhizosphere of Festuca arundinacea’, International Journal of Phytoremediation, Vol. 9, pp.107–114.
Huang, X.D., El-Alawi, Y., Penrose, D.M., Glick, B.R, and Greenberg, B.M. (2004a) ‘Responses of three grass species to creosote during phytoremediation’, Environmental Pollution, Vol. 130, pp.453–463.
Huang, X.D., El-Alawi, Y., Penrose, D.M., Glick, B.R. and Greenberg, B.M. (2004b) ‘A multiprocess phytoremediation system for removal of polycyclic aromatic hydrocarbons from contaminated soils’, Environmental Pollution, Vol. 130, pp.465–476.
Huang, X.D., El-Alawi, Y.S., Gurska, J., Glick, B.R. and Greenberg, B.M. (2005) ‘A multi-process phytoremediation system for decontamination of persistent total petroleum hydrocarbons (TPHs) from soils’, Microchemistry Journal, Vol. 81, pp.139–147.
Hurst, J.C., Sims, R., Sims, J., Sorensen, D., McLean, J. and Huling, S. (1996) ‘Polycyclic aromatic hydrocarbon biodegradation as a function of oxygen concentration in contaminated soil’, Journal of Hazardous Materials, Vol. 51, pp.193–208.
Jacobsen, C.S. and Petersen, J.C. (1992) ‘Growth and survival of Pseudomonas cepacia DBO1 (pRO101) in soil amended with 2,4-dichlorophenoxyacetic acid’, Biodegradation, Vol. 2, pp.245–252.
Joner, E.J. and Leyval, C. (2003) ‘Rhizosphere gradients of polycyclic aromatic hydrocarbon (PAH) dissipation in two industrial soils and the impact of arbuscular mycorrhiza’, Environmental Science and Technology, Vol. 37, pp.2371–2375.
Joner, E.J., Corgié, S., Amellal, N. and Leyval, C. (2002) ‘Nutritional constrains to PAH degradation in a rhizosphere model’, Soil Biology and Biochemistry, Vol. 34, pp.859–864.
Kanaly, R.A. and Harayama, S. (2000) ‘Biodegradation of high-molecular-weight polycyclic aromatic hydrocarbons by bacteria’, Journal of Bacteriology, Vol. 182, No. 8, pp.2059–2067.
Karthikeyan, R. and Kulakow, P.A. (2003) ‘Soil plant microbe interactions in phytoremediation’, Advances in Biochemistry Engineering and Biotechnology, Vol. 78, pp.51–74.
Kästner, M. and Mahro, B. (1996) ‘Microbial degradation of polycyclic aromatic hydrocarbons in soils affected by the organic matrix of compost’, Applied Microbiology & Biotechnology, Vol. 44, pp.668–675.
Khan, A.G. (2005) ‘Role of soil microbes in the rhizospheres of plants growing on trace metal contaminated soils in phytoremediation’, Journal of Trace Elements in Medicine and Biology, Vol. 18, pp.355–364.
Kidd, P.S., Prieto-Fernández, A., Monterosso, C. and Acea, M.J. (2008) ‘Rhizosphere microbial community and hexachlorocyclohexane degradative potential in contrasting plant species’, Plant and Soil, Vol. 302, pp.233–247.

The use of biostimulation and bioaugmentation to remove phenanthrene 57
Kiyohara, H., Torigoe, S., Kaida, N., Asaki, T., Lida, T., Hayashi, H. and Takizawa, N. (1994) ‘Cloning and characterization of a chromosomal gene cluster, path, that encodes the upper pathway for phenanthrene and naphthalene utilization by Pseudomonas putida OUS82’, Journal of Bacteriology, Vol. 176, No. 8, pp.2439–2443.
Kowalchuk, G.A., Buma, D.S., de Boer, W., Klinkhamer, P.G.L. and van Veen, J.A. (2002) ‘Effects of above-ground plant species composition and diversity on the diversity of soil-borne microorganisms’, Antonie van Leeuwenhoek, Vol. 81, pp.509–520.
Kuiper, I., Lagendijk, E.L., Bloemberg, G.V. and Lugtenberg, B.J.J. (2004) ‘Rhizoremediation: a beneficial plant–microbe interaction’, Molecular Plant-Microbe Interactions, Vol. 17, pp.6–15.
Leigh, M.B., Fletcher, J., Nagle, D., Kucerova, P., Mackova, M. and Macek, T. (2004) ‘Rhizosphere remediation of PCBs based on field studies in the Czech Republic’, International Journal of Biodeterioration and Biodegradation, Vol. 53, p.260.
Leigh, M.B., Prouzova, P., Mackova, M., Macek, T., Nagle, D.P. and Fletcher, J.S. (2006) ‘Polychlorinated bacteria associated with the trees in a PCB-contaminated site’, Applied of Environmental Microbiology, Vol. 72, pp.2331–2342.
Liste, H.H. and Alexander, M. (2000) ‘Accumulation of phenanthrene and pyrene in rhizosphere soil’, Chemosphere, Vol. 40, pp.11–14.
Liu, L., Jiang, C.Y., Liu, X.Y., Wu, J.F., Han, J.G. and Liu, S.J. (2007) ‘Plant–microbe association for rhizoremediation of chloronitroaromatic pollutants with Comamonas sp. strain CNB-1’, Environmental Microbiology, Vol. 9, pp.465–473.
Mackova, M., Vrchotova, B., Francova, K., Sylvestre, M., Tomaniova, M., Lovecka, P., Demnerova, K. and Macek, T. (2007) ‘Biotransformation of PCBs by plants and bacteria e consequences of plant-microbe interactions’, European Journal of Soil Biology, Vol. 43, pp.233–241.
Martínez, M.G., Santos, A.G., Cruz, L.G., García, M.A.M.O., Trujillo, L.Y., Lis, J.A.Z.E. and Gutierrez-Rojas, N. (2000) ‘Diagnostic and resulting approaches to restore petroleum-contaminated soil in a Mexican tropical swamp’, Water Science and Technology, Vol. 42, pp.377–384.
Mitchell, D.A. (1992) ‘Biomass determination in solid-state cultivation’, Chapter 4, in Doelle, H., Mitchell, D.A. and Rolz, C.E.: Solid Substrate Cultivation, p.55, Elsevier, UK.
Moo-Young, M. (1985) ‘‘Comprehensive Biotechnology’: The principles, applications and regulations of biotechnology in industry, agriculture and medicine’, Chapter 10, Zabriskie, D.W.: Data analysis, p.175, Pergamon Press, UK.
Mrozik, A. and Piotrowska-Seget, Z. (2009) ‘Bioaugmentation as a strategy for cleaning up of soils contaminated with aromatic compounds’, Microbiological Research, in press, corrected proof, available online 6 September 2009.
Müncnerova, D. and Augustin, J. (1994) ‘Fungal metabolism and detoxification of polycyclic aromatic hydrocarbons: a review’, Bioresource Technology, Vol. 48, pp.97–106.
Muratova, A., Hubner, Th., Tischer, S., Turkovskaya, O., Moder, M. and Kuschk, P. (2003) ‘Plant–rhizosphere-microflora association during phytoremediation of PAH-contaminated soil’, International Journal of Phytoremediation, Vol. 5, pp.137–152.
Newman, L.A. and Reynolds, C.M. (2004) ‘Phytodegradation of organic compounds’, Current Opinion in Biotechnology, Vol. 15, pp.225–230.
Ouyang, J. (2002) ‘Phenanthrene pathway’, The University of Minnesota Biocatalysis/Biodegradation Database, University of Minnesota, available at http://umbbd.ahc.umn.edu/pha2/pha2_image_map.html.
Pandey, A., Soccol, C., Nigam, P. and Soccol, V. (2000) ‘Biotechnological potential of agroindustrial residues. I: sugarcane bagasse’, Bioresource Technology, Vol. 74, pp.69–80.

58 M.A. Amezcua-Allieri et al.
Peña-Castro, J.M., Barrera-Figueroa, B.E., Fernández-Linares, L., Ruiz-Medrano, R., Xoconostle-Cázares, B. (2006) ‘Isolation and identification of up-regulated genes in bermudagrass roots (Cynodon dactylon L.) grown under petroleum hydrocarbon stress’, Plant Science, Vol. 170, pp.724–731.
Riser-Roberts, E. (1998) Remediation of Petroleum Contaminated Soils. Biological, Physical and Chemical Processes, pp.115–198, CRC Press, Boca Raton, Florida.
Rivera-Cruz, M.C., Ferrera-Cerrato, R., Sánchez-García, P., Volke-Haller, V., Fernández-Linares, L. and Rodríguez-Vázquez, R. (2004) ‘Decontamination of soils polluted with crude petroleum using indigenous microorganisms and Aleman Grass [Echinochloa polystachya (H.B.K.) Hitchc.]’, Agrociencia, Vol. 38, pp.1–12.
Rogers, J.A., Tedaldi, D.J. and Kavanaugh, M.C. (1993) ‘A screening protocol of bioremediation of contaminated soil’, Environmental Progress, Vol. 12, pp.146–156.
Roldán-Martín, A. (2006) ‘Remoción de hidrocarburos en un sistema de biopilas con adición de texturizantes’, PhD thesis, CINVESTAV, Mexico.
Samanta, S.K., Singh, O.V. and Jain, R.K. (2002) ‘Polycyclic aromatic hydrocarbons: environmental pollution and bioremediation’, Trends Biotechnology, Vol. 20, p.243.
Sandhu, A., Halverson, L.J. and Beattie, G.A. (2007) ‘Bacterial degradation of airborne phenol in the phyllosphere’, Environmental Microbiology, Vol. 9, pp.383–392.
Schwab, A.P. and Banks, M.K. (1994) ‘Biologically mediated dissipation of polyaromatic hydrocarbons in the root zone’, in Anderson, T.A. and Coats, J.R.: Bioremediation Through Rhizosphere Technology, pp.132–141, American Chemical Society, Washington, DC.
Schwartz, E., Trinh, S.V. and Scow, K.N. (2000) ‘Measuring growth of a phenanthrene-degrading bacterial inoculum in soil with a quantitative competitive polymerase chain reaction method’, FEMS Microbiology Ecology, Vol. 34, pp.1–7.
Scullion, J. (2006) ‘Remediating polluted soils’, Naturwissenschaften, Vol. 93, pp.51–56. Serrano, S.I., da Costa, S.E., Ragagnin de Menezes, C., de Faria, A.F., Franciscon, E.,
Grossman, M. and Regina, L.D. (2009) ‘Bioremediation of a polyaromatic hydrocarbon contaminated soil by native soil microbiota and bioaugmentation with isolated microbial consortia’, Bioresource Technology, Vol. 100, No. 20, pp.4669–4675.
Siciliano, S.D., Germida, J.J., Banks, K. and Greer, C.W. (2003) ‘Changes in microbial community composition and function during a polyaromatic hydrocarbon phytoremediation field trial’, Applied of Environmental Microbiology, Vol. 69, pp.483–489.
Silva S., Grossman, M. and Durrant, L. (2009). ‘Degradation of polyciclic aromatic hydrocarbons (2-7 rings) under microaerobic and very low oxygen conditions by soil fungi’, International Biodeterioration and Biodegradation, Vol. 63, pp.224–229.
Singer, A.C., van der Gast, C.J. and Thompson, I.P. (2005) ‘Perspectives and vision for strain selection in bioaugmentation’, Trends in Biotechnology, Vol. 23, No. 2, pp.74–77.
Sutherland, J.B., Rafii, F., Khan, A.A. and Cerniglia, C.E. (1995) ‘Mechanisms of polycyclic aromatic hydrocarbon degradation’, in Young, L.Y. and Cerniglia, C.E.: Microbial transformation and degradation of toxic organic chemicals, pp.269–306, Wiley-Liss, NY.
Taghavi, S., Barac, T., Greenberg, B., Borremans, B., Vangronsveld, J. and van der Lelie, D. (2005) ‘Horizontal gene transfer to endogenous endophytic bacteria from Poplar improves phytoremediation of toluene’, Applied and Environmental Microbiology, Vol. 71, pp.8500–8505.
Thomson, I.P., van der Gast, C.J., Ciric, L. and Singer, A.C. (2005) ‘Bioaugmentation for bioremediation: the challenge of strain selection’, Environmental Microbiology, Vol. 7, pp.909–915.
USEPA (2008) ‘Polycyclic Aromatic Hydrocarbons (PAHs), 2008’, available at http://www.epa.gov/osw/hazard/wastemin/priority.htm.

The use of biostimulation and bioaugmentation to remove phenanthrene 59
Villacieros, M., Whelan, C.M., Mackova, M., Molgaard, J., Sanchez-Contreras, M., Lloret, J., Aguirre de Carcer, D., Bolanos, L., Oruezabal, R.I., Macek, T., Karlson, U., Dowling, D.N., Martin, M. and Rivilla, R. (2005) ‘PCB rhizoremediation by Pseudomonas fluorescens F113 derivatives using a Sinorhizobium meliloti nod system to drive bph gene expression’, Applied of Environmental Microbiology, Vol. 71, pp.2687–2694.
Weyens, N., van der Lelie, D., Taghavi, S. and Vangronsveld, J. (2009) ‘Phytoremediation: plant–endophyte partnerships take the challenge’, Current Opinion in Biotechnology, Vol. 20, pp.248–254.
White, P.A. and Claxton, L.D. (2004) ‘Mutagens in contaminated soil: a review’, Mutation Research, Vol. 567, pp.227–345.
WHO (2001) ‘Environmental health criteria 202: selected non-heterocyclic polycyclic aromatic hydrocarbons’, available at http://www.who.int/pcs/ehc/corrigenda/202.html.
Wild, S.R. and Jones, K.C. (1995) ‘Polynuclear aromatic hydrocarbons in the United Kingdom environment: a preliminary source inventory and budget’, Environmental Pollution, Vol. 88, pp.91–108.
Wilson, L.P. and Bouwer, E.J. (1997) ‘Biodegradation of aromatic compounds under mixed oxygen denitrifying conditions a review’, Journal of Industrial Microbiology and Biotechnology, Vol. 18, Nos. 2–3, pp.116–130.
Wittig, R., Ballach, H.J. and Kuhn, A. (2003) ‘Exposure of the roots of Populus nigra L., cv. Loenen to PAHs and its effect on growth and water balance’, Environmental Science and Pollution Research International, Vol. 10, pp.235–244.
Yerushalmi, L. and Guiot, S.R. (2001) ‘Biodegradation of benzene in a laboratory-scale bio-barrier at low dissolved oxygen concentrations’, Bioremediation Journal, Vol. 5, No. 1, pp.63–77.
Zhang, X., Cheng, S., Zhu, C. and Sun, S. (2006) ‘Microbial PAH-degradation in soil: degradation pathways and contributing factors’, Pedosphere, Vol. 16, No. 5, pp.555–565.
Zhuang, X., Chen, J., Shim, H. and Bai, Z. (2007) ‘New advances in plant growth-promoting rhizobacteria for bioremediation’, Environment International, Vol. 33, pp.406–413.




















