
Initiation factors IF1 and IF2 synergistically remove peptidyl-tRNAs with short polypeptides from...
12
Initiation Factors IF1 and IF2 Synergistically Remove Peptidyl-tRNAs with Short Polypeptides from the P-site of Translating Escherichia coli ribosomes Reza Karimi 1 , Michael Yu. Pavlov 1 , Vale ´ rie Heurgue ´ -Hamard 2 Richard H. Buckingham 2 and Ma ˚ ns Ehrenberg 1 * 1 Department of Molecular Biology, BMC, Box 590 S-75124 Uppsala, Sweden 2 UPR 9073 du CNRS, Institut de Biologie Physico-Chimique 13 rue Pierre et Marie Curie Paris 75005, France A novel function of initiation factors IF1 and IF2 in Escherichia coli trans- lation has been identified. It is shown that these factors efficiently cata- lyse dissociation of peptidyl-tRNAs with polypeptides of different length from the P-site of E. coli ribosomes, and that the simultaneous presence of both factors is required for induction of drop-off. The factor-induced drop-off occurs with both sense and stop codons in the A-site and com- petes with peptide elongation or termination. The efficiency with which IF1 and IF2 catalyse drop-off decreases with increasing length of the nas- cent polypeptide, but is quite significant for hepta-peptidyl-tRNAs, the longest polypeptide chains studied. In the absence of IF1 and IF2 the rate of drop-off varies considerably for different peptidyl-tRNAs, and depends both on the length and sequence of the nascent peptide. Efficient factor-catalysed drop-off requires GTP but not GTP hydrolysis, as shown in experiments without guanine nucleotides, with GDP or with the non- cleavable analogue GMP-PNP. Simultaneous overexpression of IF1 and IF2 in vivo inhibits cell growth specifically in some peptidyl-tRNA hydrolase deficient mutants, suggesting that initiation factor-catalysed drop-off of peptidyl-tRNA can occur on a significant scale in the bacterial cell. Consequences for the bac- terial physiology of this previously unknown function of IF1 and IF2 are discussed. # 1998 Academic Press Keywords: initiation factors; peptidyl-tRNAs; drop-off; ribosomal P-site; translation *Corresponding author Introduction During protein synthesis on the ribosome, the polypeptide chain grows by one amino acid resi- due at a time as a result of successive transfers of the chain from the peptidyl-tRNA in the P-site to the amino group of aminoacyl-tRNA in the A-site (for a review, see Hershey, 1987). Peptidyl-tRNAs with polypeptides varying in length from the dipeptide to the final protein are thus necessary intermediates of protein synthesis, and must remain stably bound to the ribosome during pro- tein elongation. Nevertheless, protein synthesis is not perfectly processive, and drop-off of peptidyl- tRNAs from translating ribosomes is considered to account for the major part of failures in processiv- ity (reviewed by Kurland et al., 1996). Increased drop-off can be induced by certain drugs or by starvation (Menninger, 1985; Menninger et al., 1994; Karimi & Ehrenberg, 1994, 1996). It has been suggested that drop-off of peptidyl-tRNAs is a mechanism of error-correction (Menninger, 1976, 1977), though this has been questioned on theoreti- cal grounds (Kurland & Ehrenberg, 1985), due to the high energy costs of such a mechanism. Peptidyl-tRNA drop-off leads to accumulation of peptidyl-tRNAs in the cytoplasm of the cell, which may lead to lethal sequestering of tRNA isoaccep- tors or be poisonous for translating ribosomes E-mail address of the corresponding author: [email protected] Abbreviations used: Pth, peptidyl-tRNA hydrolase; PMSF, phenylmethylsulphonyl fluoride; PEP, phosphoenol pyruvate; DTE, dithioerythritol; PK, pyruvate kinase; MK, myokinase. Article No. mb981953 J. Mol. Biol. (1998) 281, 241–252 0022 – 2836/98/320241–12 $30.00/0 # 1998 Academic Press
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Initiation factors IF1 and IF2 synergistically remove peptidyl-tRNAs with short polypeptides from...

Article No. mb981953 J. Mol. Biol. (1998) 281, 241±252
Initiation Factors IF1 and IF2 Synergistically RemovePeptidyl-tRNAs with Short Polypeptides from theP-site of Translating Escherichia coli ribosomes
Reza Karimi1, Michael Yu. Pavlov1, ValeÂrie Heurgue -Hamard2
Richard H. Buckingham2 and MaÊns Ehrenberg1*
t
1Department of MolecularBiology, BMC, Box 590S-75124 Uppsala, Sweden2UPR 9073 du CNRS, Institude Biologie Physico-Chimique13 rue Pierre et Marie CurieParis 75005, France
0022±2836/98/320241±12 $30.00/0
A novel function of initiation factors IF1 and IF2 in Escherichia coli trans-lation has been identi®ed. It is shown that these factors ef®ciently cata-lyse dissociation of peptidyl-tRNAs with polypeptides of different lengthfrom the P-site of E. coli ribosomes, and that the simultaneous presenceof both factors is required for induction of drop-off. The factor-induceddrop-off occurs with both sense and stop codons in the A-site and com-petes with peptide elongation or termination. The ef®ciency with whichIF1 and IF2 catalyse drop-off decreases with increasing length of the nas-cent polypeptide, but is quite signi®cant for hepta-peptidyl-tRNAs, thelongest polypeptide chains studied. In the absence of IF1 and IF2 the rateof drop-off varies considerably for different peptidyl-tRNAs, anddepends both on the length and sequence of the nascent peptide. Ef®cientfactor-catalysed drop-off requires GTP but not GTP hydrolysis, as shownin experiments without guanine nucleotides, with GDP or with the non-cleavable analogue GMP-PNP.
Simultaneous overexpression of IF1 and IF2 in vivo inhibits cell growthspeci®cally in some peptidyl-tRNA hydrolase de®cient mutants,suggesting that initiation factor-catalysed drop-off of peptidyl-tRNA canoccur on a signi®cant scale in the bacterial cell. Consequences for the bac-terial physiology of this previously unknown function of IF1 and IF2 arediscussed.
# 1998 Academic Press
Keywords: initiation factors; peptidyl-tRNAs; drop-off; ribosomal P-site;translation
*Corresponding authorIntroduction
During protein synthesis on the ribosome, thepolypeptide chain grows by one amino acid resi-due at a time as a result of successive transfers ofthe chain from the peptidyl-tRNA in the P-site tothe amino group of aminoacyl-tRNA in the A-site(for a review, see Hershey, 1987). Peptidyl-tRNAswith polypeptides varying in length from thedipeptide to the ®nal protein are thus necessaryintermediates of protein synthesis, and must
E-mail address of the corresponding author:[email protected]
Abbreviations used: Pth, peptidyl-tRNA hydrolase;PMSF, phenylmethylsulphonyl ¯uoride; PEP,phosphoenol pyruvate; DTE, dithioerythritol; PK,pyruvate kinase; MK, myokinase.
remain stably bound to the ribosome during pro-tein elongation. Nevertheless, protein synthesis isnot perfectly processive, and drop-off of peptidyl-tRNAs from translating ribosomes is considered toaccount for the major part of failures in processiv-ity (reviewed by Kurland et al., 1996). Increaseddrop-off can be induced by certain drugs or bystarvation (Menninger, 1985; Menninger et al.,1994; Karimi & Ehrenberg, 1994, 1996). It has beensuggested that drop-off of peptidyl-tRNAs is amechanism of error-correction (Menninger, 1976,1977), though this has been questioned on theoreti-cal grounds (Kurland & Ehrenberg, 1985), due tothe high energy costs of such a mechanism.
Peptidyl-tRNA drop-off leads to accumulation ofpeptidyl-tRNAs in the cytoplasm of the cell, whichmay lead to lethal sequestering of tRNA isoaccep-tors or be poisonous for translating ribosomes
# 1998 Academic Press

242 IF1 and IF2 Remove Peptidyl-tRNAs from Ribosomes
(Menninger, 1976; Atherly, 1978). The experimentalevidence supports sequestering of tRNA isoaccep-tors as the principal explanation for growthinhibition due to enhanced peptidyl-tRNAaccumulation in cells de®cient in peptidyl-tRNAhydrolase activity off the ribosomes (Menninger,1976; HeurgueÂ-Hamard et al., 1996). This activity iscarried out by the essential enzyme (Atherly &Menninger, 1972) peptidyl-tRNA hydrolase (Pth;EC 3.1.1.29), which, under normal conditions, pre-vents excessive peptidyl-tRNA accumulation inthe cytoplasm (Cuzin et al., 1967; Kossel &RajBhandary, 1968; Chapeville et al., 1969).
Recent in vitro measurements in a poly(U) trans-lation system (Karimi & Ehrenberg, 1996) showthat the drop-off rate of NAc-Phe-Phe-tRNAPhe
from the P-site is too slow to account for the drop-off frequencies of about 4 � 10ÿ4 per translatedcodon measured for peptidyl-tRNA in vivo(Menninger, 1976; reviewed by Kurland et al.,1996). In the present study, the drop-off rates ofdifferent peptidyl-tRNAs were measured underconditions more relevant to translation in vivo.A newly developed in vitro translation systemassembled from puri®ed components (Pavlov &Ehrenberg, 1996) has been employed to translateshort synthetic mRNAs in a recycling mode, whereribosomes go through initiation, elongation, ter-mination and then back to initiation, generatingmany completed polypeptides per ribosome(Pavlov et al., 1997a). Surprisingly, when releasefactors were omitted from this translation system,ribosome recycling due to peptidyl-tRNA drop-offwas found to be about 15 times faster than pre-dicted from the drop-off rates measured in apoly(U) translation system (Karimi & Ehrenberg,1996). This indicated that the rate of peptidyl-tRNA drop-off was signi®cantly enhanced by somefactors present in the more complete translationsystem.
The present work demonstrates that initiationfactors IF1 and IF2 are responsible for the observeddrop-off acceleration, and that they synergisticallyremove peptidyl-tRNAs from the ribosomal P-sitein competition with further protein elongation ortermination. IF1 and IF2 accelerate drop-off ofdipeptidyl-tRNAs most ef®ciently but they signi®-cantly enhance the drop-off rate also of heptapepti-dyl-tRNAs carrying the longest polypeptidesstudied. These results are reminiscent of recentexperiments showing that peptidyl-tRNA drop-offis stimulated also by elongation factor EF-G actingtogether with auxiliary termination factors RF3and RRF (RF4: HeurgueÂ-Hamard et al., 1998).Taken together, these data indicate that peptidyl-tRNA drop-off in vivo is catalysed by translationfactors and not simply determined by the af®nitybetween ribosomes and peptidyl-tRNA per se,as previously thought (Menninger, 1976, 1977;Kurland et al., 1996).
Results
Ribosome recycling in the absence ofrelease factors
The in vitro translation system used in the pre-sent study employs puri®ed initiation, elongationand termination factors, and other componentsnecessary for translation of short synthetic mRNAscontaining a Shine/Dalgarno sequence, initiationcodon, a short coding sequence and a terminationsignal. In this system ribosomes perform repeatedcycles of translation, generating many completedpolypeptides per ribosome (Pavlov et al., 1997a). Inthe absence of termination factors one wouldexpect the ribosome to carry out only a singleround of translation and then pause at the stopcodon with peptidyl-tRNA in the P-site.
Figure 1(a) shows the time-course of fMet-Phe-Leu-tRNALeu
4 (fMFL-tRNA) accumulation,measured with HPLC (Materials and Methods),during translation of MFL mRNA (codingsequence AUG UUC UUG) in the absence ofrelease factors. There is an initial burst of fMFL-tRNA synthesis, measured from the intercept withthe y-axis in Figure 1(a), which corresponds to theamount of ribosomes active in translation (Pavlovet al., 1997a). After this burst, however, the amountof fMFL-tRNA continues to increase linearly withtime due to new, slow rounds of tripeptidyl-tRNAsynthesis. This additional peptidyl-tRNA synthesiscan be caused only by peptidyl-tRNA drop-off,which allows the ribosome stalled at the stopcodon of MFL mRNA to enter the next round oftranslation. Figure 1(a) shows that when the con-centrations of IF1 and IF2 are increased from0.3 mM to 1.5 mM for both factors, the ribosomalrecycling time decreases from 10.5 minutes to 1.6minutes. To test if the slow recycling at lowinitiation factor concentrations was caused by slowinitiation, recycling was also measured in the pre-sence of release factors RF1, RF3 and RRF(Figure 1(b)). Here, the recycling time is only aboutnine seconds at both the low and high concen-trations of IF1 and IF2. This shows that the rate ofre-initiation of ribosomes does not limit their recy-cling, either at high or at low concentrations of IF1and IF2. A plausible interpretation of these resultsis that the drop-off of peptidyl-tRNAs is acceler-ated when the concentrations of IF1 and IF2increase.
Table 1 summarises ribosomal recycling times,obtained in translation experiments with six differ-ent mRNAs coding for tri- and tetrapeptides. ThemRNAs were translated either in the absence ofrelease factors or of aminoacyl-tRNAs necessary tocomplete their decoding. In the former case, theribosomes are brought to pause with a stop codonand in the latter with a sense codon in the A-site.In both cases, the ribosomal recycling proceedsthrough the drop-off of peptidyl-tRNA from theribosomal P-site. In these experiments, IF1 wasalways at a low (0.1 mM) concentration, while IF2

IF1 and IF2 Remove Peptidyl-tRNAs from Ribosomes 243
was either at a low (0.1 mM) or at a high (1 mM)concentration. The experiments show that IF2 isable to accelerate the recycling of ribosomes trans-lating many different short mRNAs and thereforethe phenomenon is general. The ribosomal recy-cling times decrease between three- and sixfoldwhen the concentration of IF2 increases from its
Figure 1. (a) fMet-Phe-Leu-tRNALeu4 accumulation in
ribosome recycling experiments with 0.9 mM total(0.25 mM active) ribosomes isolated from E. coli strain017 at 0.3 mM (*) and at 1.5 mM (&) concentrations ofIF1 and IF2 in the absence of termination factors.(b) fMet-Phe-Leu tripeptide accumulation under thesame conditions as in (a) but with release factors RF1,RF3 and RRF present at 1.0 mM each. Ribosome recy-cling times were calculated by multiplying the reciprocalof the slopes of the straight lines in the Figure with theamount of active ribosomes, obtained from the inter-cepts of the lines with the y-axis. In the absence ofrelease factors, the recycling time was 10.5 minutes atthe low and 1.6 minutes at the intermediate concen-trations of IF1 and IF2. In the presence of release factors,the corresponding times were ten and nine seconds.
low to its high value. Table 1 demonstrates a cleartendency for ribosomes to recycle faster when theysynthesise di- than when they synthesise tripepti-dyl-tRNAs both at high and at low concentrationsof IF2. This most probably re¯ects faster drop-offrates for peptidyl-tRNAs with shorter polypep-tides. At the same time there is considerable vari-ation in recycling rates for a given length of thepolypeptide, presumably re¯ecting the in¯uence ofthe identity of the tRNA moiety of peptidyl-tRNAas well as the amino acid sequence of the polypep-tide on the rate of drop-off. An interesting case isthe recycling of ribosomes synthesising fMet-Phe-tRNAPhe (anticodon GAA) by translating mMFLmRNA (Phe codon UUC) in the absence of Leu-tRNALeu or mMFQI mRNA (Phe codon UUU) inthe absence of Gln-tRNAGln. At the low concen-tration of IF2, recycling is slower with a UUCcodon, which may re¯ect a stronger codon-antico-don interaction, but at high concentration ofIF2 the drop-off times are similar and close to0.5 minute.
Fast peptidyl-tRNA drop-off requirescooperative action of IF1 and IF2
The in¯uence of IF1 and IF2 on the drop-off ofpeptidyl-tRNAs was also directly measured with adifferent experimental approach. Ribosomes wereinitiated with different mRNAs, which were trans-lated up to a stop or a sense codon. Here the ribo-somes paused due to the absence of either releasefactors or ternary complex cognate for the codon inthe A-site. The ribosomes, containing a peptidyl-tRNA in the P-site and with an empty A-site, weresubsequently separated from all other componentsin the translation system by gel ®ltration (Materialsand Methods; and see Freistroffer et al., 1997). Therate of peptidyl-tRNA drop-off from these puri®edpeptidyl-tRNA:ribosome complexes was measuredas the rate of peptidyl-tRNA hydrolysis in the pre-sence of an excess amount of peptidyl-tRNAhydrolase (Pth). This can be done, since Pth doesnot hydrolyse peptidyl-tRNA on the ribosome andthe rate of hydrolysis of free peptidyl-tRNA athigh concentrations of Pth is always much fasterthan the drop-off rates. Figure 2 shows thataddition of IF1 and IF2 to ribosomes containingfMFL-tRNA strongly stimulates drop-off, asexpected from the ribosome recycling experimentsdescribed above (Figure 1(a) and Table 1). The®gure also shows that IF1 and IF2 induce drop-offin a synergistic fashion: when either IF1 or IF2 ispresent alone, there is a very small effect on drop-off, but when both factors are added, the rate ofdrop-off increases dramatically. At the same time,addition of initiation factor IF3, elongation factorsEF-Tu and EF-G, added either alone or in differentcombinations, has virtually no effect on the drop-off rate of peptidyl-tRNA (data not shown).
Figure 3 shows how the drop-off time of fMFL-tRNA decreases to a minimal value of about 0.7minute, when the concentrations of IF1 and IF2 are
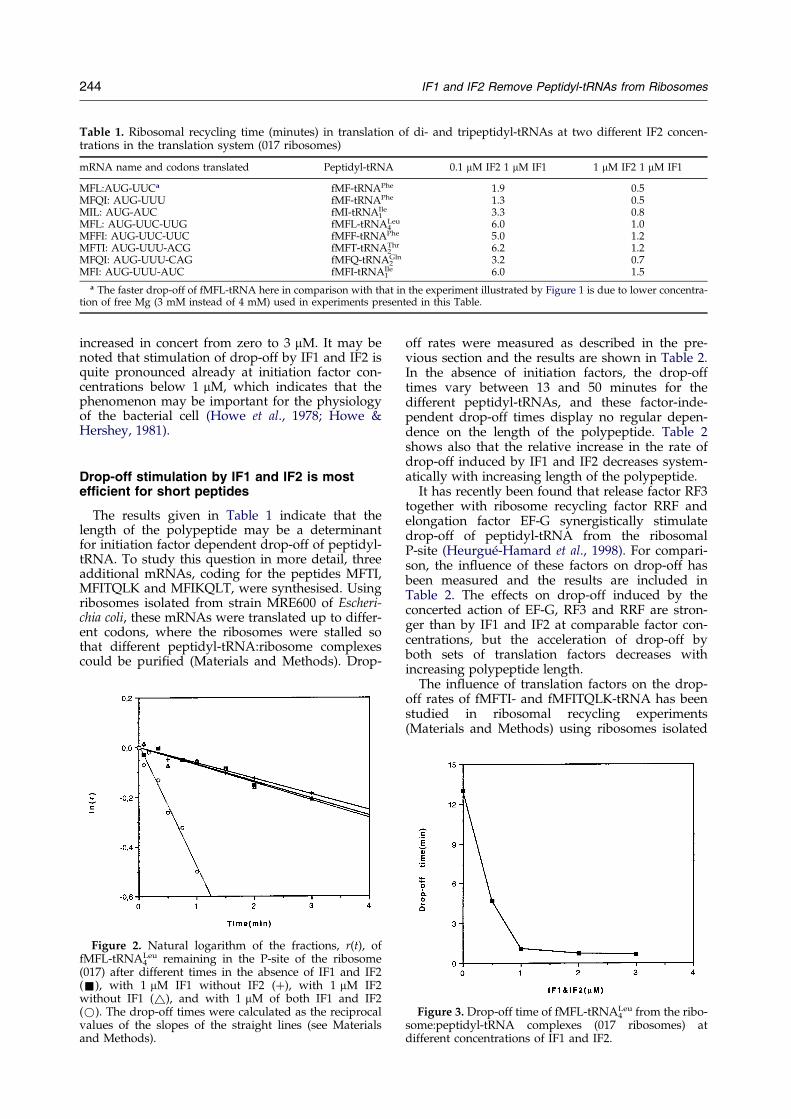
Table 1. Ribosomal recycling time (minutes) in translation of di- and tripeptidyl-tRNAs at two different IF2 concen-trations in the translation system (017 ribosomes)
mRNA name and codons translated Peptidyl-tRNA 0.1 mM IF2 1 mM IF1 1 mM IF2 1 mM IF1
MFL:AUG-UUCa fMF-tRNAPhe 1.9 0.5MFQI: AUG-UUU fMF-tRNAPhe 1.3 0.5MIL: AUG-AUC fMI-tRNA1
Ile 3.3 0.8MFL: AUG-UUC-UUG fMFL-tRNALeu
4 6.0 1.0MFFI: AUG-UUC-UUC fMFF-tRNAPhe 5.0 1.2MFTI: AUG-UUU-ACG fMFT-tRNA2
Thr 6.2 1.2MFQI: AUG-UUU-CAG fMFQ-tRNA2
Gln 3.2 0.7MFI: AUG-UUU-AUC fMFI-tRNA1
Ile 6.0 1.5
a The faster drop-off of fMFL-tRNA here in comparison with that in the experiment illustrated by Figure 1 is due to lower concentra-tion of free Mg (3 mM instead of 4 mM) used in experiments presented in this Table.
244 IF1 and IF2 Remove Peptidyl-tRNAs from Ribosomes
increased in concert from zero to 3 mM. It may benoted that stimulation of drop-off by IF1 and IF2 isquite pronounced already at initiation factor con-centrations below 1 mM, which indicates that thephenomenon may be important for the physiologyof the bacterial cell (Howe et al., 1978; Howe &Hershey, 1981).
Drop-off stimulation by IF1 and IF2 is mostefficient for short peptides
The results given in Table 1 indicate that thelength of the polypeptide may be a determinantfor initiation factor dependent drop-off of peptidyl-tRNA. To study this question in more detail, threeadditional mRNAs, coding for the peptides MFTI,MFITQLK and MFIKQLT, were synthesised. Usingribosomes isolated from strain MRE600 of Escheri-chia coli, these mRNAs were translated up to differ-ent codons, where the ribosomes were stalled sothat different peptidyl-tRNA:ribosome complexescould be puri®ed (Materials and Methods). Drop-
Figure 2. Natural logarithm of the fractions, r(t), offMFL-tRNALeu
4 remaining in the P-site of the ribosome(017) after different times in the absence of IF1 and IF2(&), with 1 mM IF1 without IF2 (�), with 1 mM IF2without IF1 (~), and with 1 mM of both IF1 and IF2(*). The drop-off times were calculated as the reciprocalvalues of the slopes of the straight lines (see Materialsand Methods).
off rates were measured as described in the pre-vious section and the results are shown in Table 2.In the absence of initiation factors, the drop-offtimes vary between 13 and 50 minutes for thedifferent peptidyl-tRNAs, and these factor-inde-pendent drop-off times display no regular depen-dence on the length of the polypeptide. Table 2shows also that the relative increase in the rate ofdrop-off induced by IF1 and IF2 decreases system-atically with increasing length of the polypeptide.
It has recently been found that release factor RF3together with ribosome recycling factor RRF andelongation factor EF-G synergistically stimulatedrop-off of peptidyl-tRNA from the ribosomalP-site (HeurgueÂ-Hamard et al., 1998). For compari-son, the in¯uence of these factors on drop-off hasbeen measured and the results are included inTable 2. The effects on drop-off induced by theconcerted action of EF-G, RF3 and RRF are stron-ger than by IF1 and IF2 at comparable factor con-centrations, but the acceleration of drop-off byboth sets of translation factors decreases withincreasing polypeptide length.
The in¯uence of translation factors on the drop-off rates of fMFTI- and fMFITQLK-tRNA has beenstudied in ribosomal recycling experiments(Materials and Methods) using ribosomes isolated
Figure 3. Drop-off time of fMFL-tRNALeu4 from the ribo-
some:peptidyl-tRNA complexes (017 ribosomes) atdifferent concentrations of IF1 and IF2.
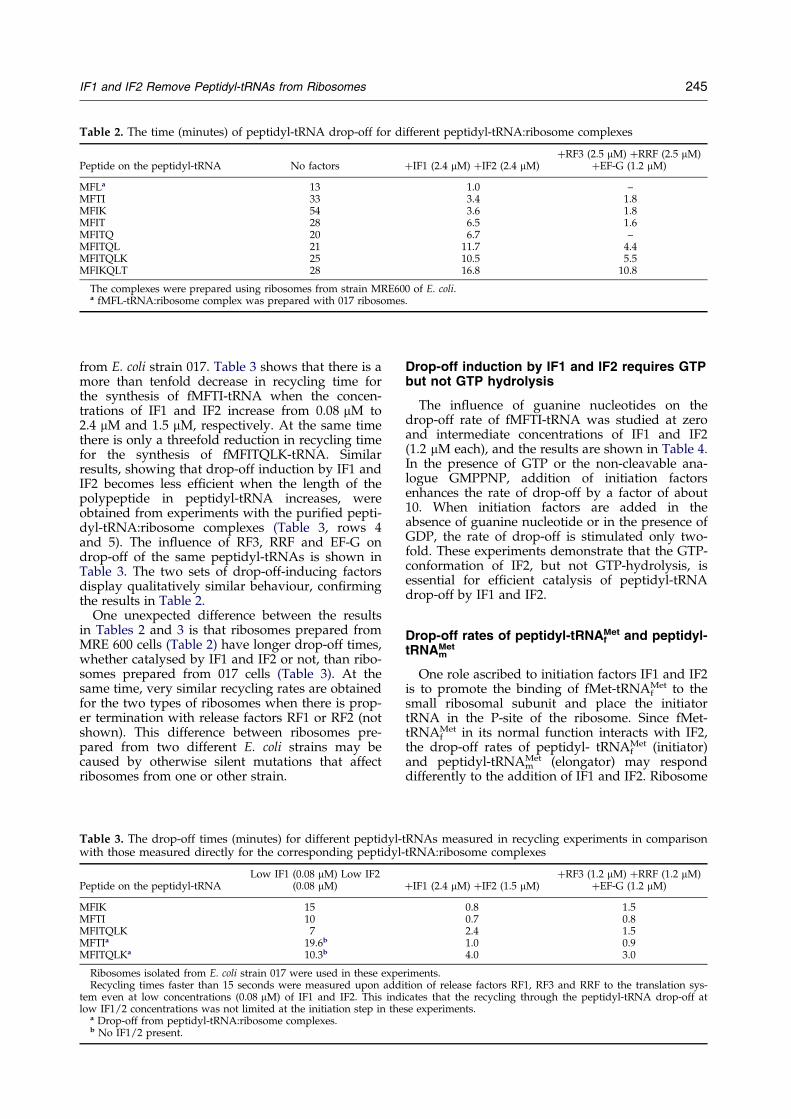
Table 2. The time (minutes) of peptidyl-tRNA drop-off for different peptidyl-tRNA:ribosome complexes
Peptide on the peptidyl-tRNA No factors �IF1 (2.4 mM) �IF2 (2.4 mM)�RF3 (2.5 mM) �RRF (2.5 mM)
�EF-G (1.2 mM)
MFLa 13 1.0 ±MFTI 33 3.4 1.8MFIK 54 3.6 1.8MFIT 28 6.5 1.6MFITQ 20 6.7 ±MFITQL 21 11.7 4.4MFITQLK 25 10.5 5.5MFIKQLT 28 16.8 10.8
The complexes were prepared using ribosomes from strain MRE600 of E. coli.a fMFL-tRNA:ribosome complex was prepared with 017 ribosomes.
IF1 and IF2 Remove Peptidyl-tRNAs from Ribosomes 245
from E. coli strain 017. Table 3 shows that there is amore than tenfold decrease in recycling time forthe synthesis of fMFTI-tRNA when the concen-trations of IF1 and IF2 increase from 0.08 mM to2.4 mM and 1.5 mM, respectively. At the same timethere is only a threefold reduction in recycling timefor the synthesis of fMFITQLK-tRNA. Similarresults, showing that drop-off induction by IF1 andIF2 becomes less ef®cient when the length of thepolypeptide in peptidyl-tRNA increases, wereobtained from experiments with the puri®ed pepti-dyl-tRNA:ribosome complexes (Table 3, rows 4and 5). The in¯uence of RF3, RRF and EF-G ondrop-off of the same peptidyl-tRNAs is shown inTable 3. The two sets of drop-off-inducing factorsdisplay qualitatively similar behaviour, con®rmingthe results in Table 2.
One unexpected difference between the resultsin Tables 2 and 3 is that ribosomes prepared fromMRE 600 cells (Table 2) have longer drop-off times,whether catalysed by IF1 and IF2 or not, than ribo-somes prepared from 017 cells (Table 3). At thesame time, very similar recycling rates are obtainedfor the two types of ribosomes when there is prop-er termination with release factors RF1 or RF2 (notshown). This difference between ribosomes pre-pared from two different E. coli strains may becaused by otherwise silent mutations that affectribosomes from one or other strain.
Table 3. The drop-off times (minutes) for different peptidywith those measured directly for the corresponding peptid
Peptide on the peptidyl-tRNALow IF1 (0.08 mM) Low IF2
(0.08 mM)
MFIK 15MFTI 10MFITQLK 7MFTIa 19.6b
MFITQLKa 10.3b
Ribosomes isolated from E. coli strain 017 were used in these exRecycling times faster than 15 seconds were measured upon ad
tem even at low concentrations (0.08 mM) of IF1 and IF2. This inlow IF1/2 concentrations was not limited at the initiation step in t
a Drop-off from peptidyl-tRNA:ribosome complexes.b No IF1/2 present.
Drop-off induction by IF1 and IF2 requires GTPbut not GTP hydrolysis
The in¯uence of guanine nucleotides on thedrop-off rate of fMFTI-tRNA was studied at zeroand intermediate concentrations of IF1 and IF2(1.2 mM each), and the results are shown in Table 4.In the presence of GTP or the non-cleavable ana-logue GMPPNP, addition of initiation factorsenhances the rate of drop-off by a factor of about10. When initiation factors are added in theabsence of guanine nucleotide or in the presence ofGDP, the rate of drop-off is stimulated only two-fold. These experiments demonstrate that the GTP-conformation of IF2, but not GTP-hydrolysis, isessential for ef®cient catalysis of peptidyl-tRNAdrop-off by IF1 and IF2.
Drop-off rates of peptidyl-tRNAMetf and peptidyl-
tRNAMetm
One role ascribed to initiation factors IF1 and IF2is to promote the binding of fMet-tRNAMet
f to thesmall ribosomal subunit and place the initiatortRNA in the P-site of the ribosome. Since fMet-tRNAMet
f in its normal function interacts with IF2,the drop-off rates of peptidyl- tRNAMet
f (initiator)and peptidyl-tRNAMet
m (elongator) may responddifferently to the addition of IF1 and IF2. Ribosome
l-tRNAs measured in recycling experiments in comparisonyl-tRNA:ribosome complexes
�IF1 (2.4 mM) �IF2 (1.5 mM)�RF3 (1.2 mM) �RRF (1.2 mM)
�EF-G (1.2 mM)
0.8 1.50.7 0.82.4 1.51.0 0.94.0 3.0
periments.dition of release factors RF1, RF3 and RRF to the translation sys-dicates that the recycling through the peptidyl-tRNA drop-off at
hese experiments.
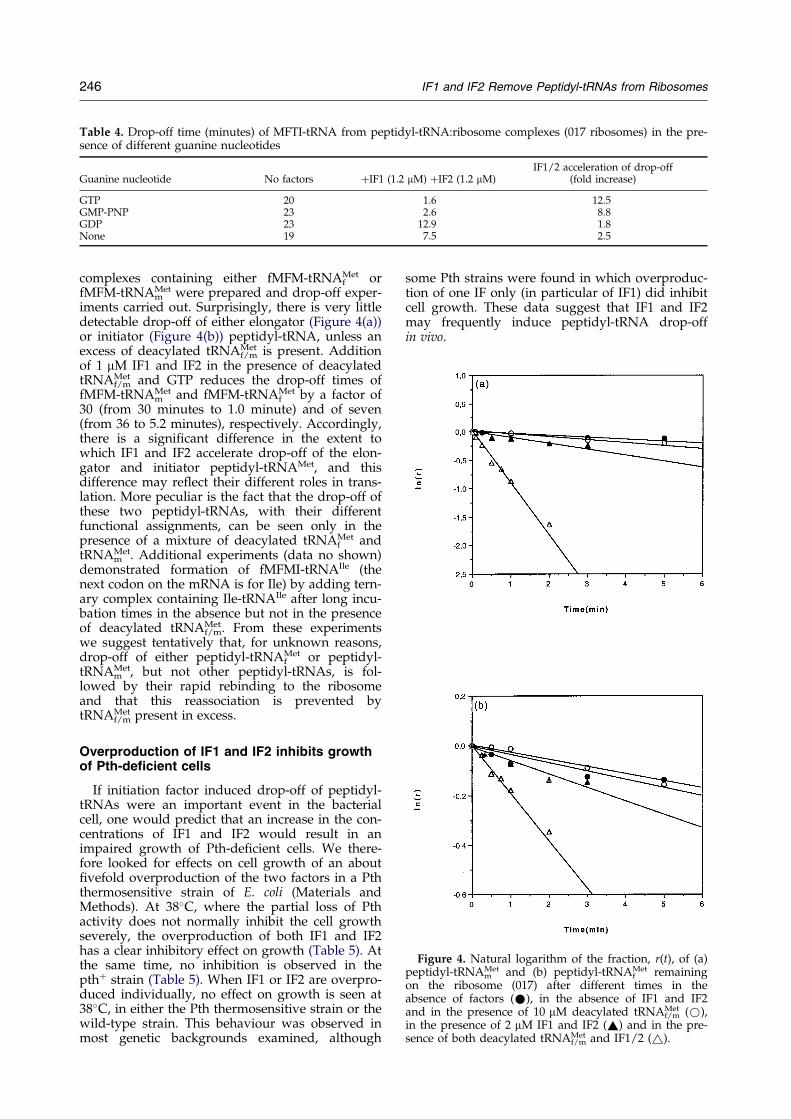
Table 4. Drop-off time (minutes) of MFTI-tRNA from peptidyl-tRNA:ribosome complexes (017 ribosomes) in the pre-sence of different guanine nucleotides
Guanine nucleotide No factors �IF1 (1.2 mM) �IF2 (1.2 mM)IF1/2 acceleration of drop-off
(fold increase)
GTP 20 1.6 12.5GMP-PNP 23 2.6 8.8GDP 23 12.9 1.8None 19 7.5 2.5
Figure 4. Natural logarithm of the fraction, r(t), of (a)peptidyl-tRNAMet
m and (b) peptidyl-tRNAMetf remaining
on the ribosome (017) after different times in theabsence of factors (*), in the absence of IF1 and IF2and in the presence of 10 mM deacylated tRNAMet
f/m (*),in the presence of 2 mM IF1 and IF2 (~) and in the pre-sence of both deacylated tRNAMet
f/m and IF1/2 (~).
246 IF1 and IF2 Remove Peptidyl-tRNAs from Ribosomes
complexes containing either fMFM-tRNAMetf or
fMFM-tRNAMetm were prepared and drop-off exper-
iments carried out. Surprisingly, there is very littledetectable drop-off of either elongator (Figure 4(a))or initiator (Figure 4(b)) peptidyl-tRNA, unless anexcess of deacylated tRNAMet
f/m is present. Additionof 1 mM IF1 and IF2 in the presence of deacylatedtRNAMet
f/m and GTP reduces the drop-off times offMFM-tRNAMet
m and fMFM-tRNAMetf by a factor of
30 (from 30 minutes to 1.0 minute) and of seven(from 36 to 5.2 minutes), respectively. Accordingly,there is a signi®cant difference in the extent towhich IF1 and IF2 accelerate drop-off of the elon-gator and initiator peptidyl-tRNAMet, and thisdifference may re¯ect their different roles in trans-lation. More peculiar is the fact that the drop-off ofthese two peptidyl-tRNAs, with their differentfunctional assignments, can be seen only in thepresence of a mixture of deacylated tRNAMet
f andtRNAMet
m . Additional experiments (data no shown)demonstrated formation of fMFMI-tRNAIle (thenext codon on the mRNA is for Ile) by adding tern-ary complex containing Ile-tRNAIle after long incu-bation times in the absence but not in the presenceof deacylated tRNAMet
f/m. From these experimentswe suggest tentatively that, for unknown reasons,drop-off of either peptidyl-tRNAMet
f or peptidyl-tRNAMet
m , but not other peptidyl-tRNAs, is fol-lowed by their rapid rebinding to the ribosomeand that this reassociation is prevented bytRNAMet
f/m present in excess.
Overproduction of IF1 and IF2 inhibits growthof Pth-deficient cells
If initiation factor induced drop-off of peptidyl-tRNAs were an important event in the bacterialcell, one would predict that an increase in the con-centrations of IF1 and IF2 would result in animpaired growth of Pth-de®cient cells. We there-fore looked for effects on cell growth of an about®vefold overproduction of the two factors in a Pththermosensitive strain of E. coli (Materials andMethods). At 38�C, where the partial loss of Pthactivity does not normally inhibit the cell growthseverely, the overproduction of both IF1 and IF2has a clear inhibitory effect on growth (Table 5). Atthe same time, no inhibition is observed in thepth� strain (Table 5). When IF1 or IF2 are overpro-duced individually, no effect on growth is seen at38�C, in either the Pth thermosensitive strain or thewild-type strain. This behaviour was observed inmost genetic backgrounds examined, although
some Pth strains were found in which overproduc-tion of one IF only (in particular of IF1) did inhibitcell growth. These data suggest that IF1 and IF2may frequently induce peptidyl-tRNA drop-offin vivo.

Table 5. Effect of overproduction of IF1 and IF2 on cell growth in wild-type and Pth-de®cient strains
StrainIB188 (pth�) VH200 (pth[ts])
Factor 35�C 37�C 38�C 35�C 37�C 38�C
None �� �� �� � � �IF2 �� �� �� �� �� �IF1 �� �� �� �� � �IF1 � IF2 �� �� �� �� � ÿ
IF1, IF2 or IF1 and IF2 were overproduced from pACYC184-derived plasmids. Cells contained either the plasmid overproducing thefactor(s) or the corresponding parent plasmid without insert. Growth was judged from colony size of streaked cells on LB-chloram-phenicol plates incubated at the indicated temperature.
IF1 and IF2 Remove Peptidyl-tRNAs from Ribosomes 247
Discussion
The present study demonstrates that theinitiation factors IF1 and IF2 strongly accelerate therate of drop-off of peptidyl-tRNAs with polypep-tides containing up to seven amino acids. Theinduction of drop-off requires the synergistic actionof both factors; it becomes weaker with longerpolypeptides and requires GTP but not GTPhydrolysis. Drop-off of peptidyl-tRNAMet couldbe observed only in the presence of deacylatedtRNAMet, indicating rapid rebinding of peptidyl-tRNA to the ribosomal P-site in this special case. Itwas found also that simultaneous overexpressionof plasmid-encoded IF1 and IF2 inhibits growth ofbacterial strains that are de®cient in peptidyl-tRNAhydrolase activity, suggesting that initiation factor-dependent drop-off of peptidyl-tRNA may occurquite frequently in the bacterial cell.
The present ®ndings are surprising, in that noexperiment until now has demonstrated the inter-ference of initiation factors with protein elongationor termination. The results strikingly resemble dataobtained by HeurgueÂ-Hamard et al. (1998), whodemonstrated that elongation factor EF-G togetherwith release factors RF3 and RRF can induce drop-off of peptidyl-tRNAs both in vitro and in vivo.
A useful starting point in considering how IF1and IF2 may induce drop-off of peptidyl-tRNA,irrespective of how the phenomenon is rational-ised, is the interplay between IF1 and IF2 in theinitiation process itself. Although much detail isknown, some essential features, in particular con-cerning IF1, remain obscure (Gualerzi & Pon,1990). An active initiation complex, ready to serveas peptide donor, consists of a 70 S particle,mRNA and fMet-tRNAMet
f in the P-site. Its for-mation is strongly accelerated by the threeinitiation factors IF1, IF2 and IF3 (Gualerzi & Pon,1990; Draper, 1996 and references therein) in aGTP-dependent way (Dubnoff et al., 1972; Benneet al., 1973). All three initiation factors are essentialin vivo (Butler et al., 1986; Shiba et al., 1986;Cummings & Hershey, 1994). At the same time,fast ribosomal recycling in vitro depends strictlyonly on the presence of IF2 and IF3, while the roleof IF1 is merely stimulatory and its presence ismost important at low concentrations of fMet-tRNAMet
f (M.Y.P., unpublished results).
A direct interaction between IF2 and the 70 Sparticle, as required for its drop-off activity, hasbeen experimentally established (Hershey & Trash,1967; Fakunding & Hershey, 1973; Dubnoff et al.,1972; Benne et al., 1973). After association of the50 S subunit with a 30 S initiation complex formedin the presence of a non-cleavable GTP analogue,IF2 remains stably bound to the 70 S ribosome(Fakunding & Hershey, 1973). Removal of IF2 fromthe initiation complex seems to require GTPhydrolysis and the presence of IF1 (Benne et al.,1973 and references therein). Initiator tRNA in theP-site becomes puromycin-reactive and competentto start protein elongation only after GTP hydroly-sis on IF2 and removal of the factor from the ribo-some (Dubnoff et al., 1972). This suggests that IF2in complex with GTP is situated in the ribosomalA-site after joining of the two ribosomal subunitsin the initiation process, and that the factor sub-sequently leaves the A-site with the help of GTPhydrolysis and the action of IF1. The latter factorhas an RNA-binding motif (Sette et al., 1997) andprotects from chemical modi®cation bases in 16 SrRNA, that have been ascribed to the partial A-siteof the 30 S subunit (Moazed et al., 1995). Takentogether, these data suggest that both IF1 and IF2are present in the A-site after formation of theinitiation complex, although some controversyremains concerning IF1, which has been suggestedto leave the small subunit already when it joins50 S (Celano et al., 1988). This reÂsume of initiationsuggests that IF1 and IF2 can bind simultaneouslyto the ribosomal A-site also during the elongationcycle. Our results indicate as well that this bindingdestabilises the peptidyl-tRNA in the P-site.
Two radically different rationalisations for drop-off induction by IF1 and IF2 can be proposed. The®rst implies that the role of the factors in initiationhas the undesired but unavoidable side-effect thatthey also induce peptidyl-tRNA drop-off, therebyimpairing processivity and growth-rate. One maythus speculate that the postulated exit path for thefactors via the A-site makes it dif®cult to preventtheir rebinding to this site also during normalelongation. Accordingly, it is possible that the pre-sence of both GTP and IF1 leads to a conformationof IF2 that is an important step in its rapidremoval, but also destabilises peptidyl-tRNAsbound to the P-site.

248 IF1 and IF2 Remove Peptidyl-tRNAs from Ribosomes
The second rationalisation implies that the twoinitiation factors have dual functions. This wouldmean that they have been positively selected fortheir ability to promote initiation of translation andto catalyse drop-off of peptidyl-tRNA. One suchauxiliary function of IF1 and IF2 could be to termi-nate protein synthesis at early stages of elongationunder conditions of mild starvation. In this way,ribosomes trapped at ``hungry codons'' in thebeginning of an mRNA could be liberated and pro-duction of proteins with high missense errorscould be minimised (O'Farrel, 1978; Wagner et al.,1982). Alternatively, this IF1/IF2-induced drop-offcould be used to regulate the expression of genescontaining rare codons near their beginning (Chen& Inouye, 1990). Both these suggestions are compa-tible with the observation (R.K., unpublishedresults) that shortage of cognate ternary complexescontaining Leu-tRNALeu causes signi®cant fMet-Phe-tRNAPhe drop-off during in vitro synthesis offMet-Phe-Leu-tRNALeu. These experiments showthat, indeed, peptidyl-tRNA removal by IF1 andIF2 can effectively compete with protein elongationwhen the level of ternary complex is low, as whenthe A-site contains a rare codon or when there isamino acid starvation in the cell.
A striking observation in the present study isthat the ef®ciency with which IF1 and IF2 promotepeptidyl-tRNA drop-off decreases with increasinglength of the polypeptide (Table 2). One clue tothis length-dependence may be found in the pat-tern of cross-links between the N terminus of thenascent polypeptide and 23 S rRNA. This patternchanges dramatically when the length of the poly-peptide increases from four to six amino acid resi-dues and then remains essentially the same up to alength of 15 residues (Stade et al., 1995). Anextreme interpretation of these data is that startingwith the length of four to six residues, the growingpeptide enters a tunnel leading through the 50 Ssubunit (Yonath & Berkovitch-Yellin, 1993 andreferences therein; Frank et al., 1995; Agrawal et al.,1996). The existence of such a tunnel would effec-tively prohibit drop-off of peptidyl-tRNAs withsuf®ciently long polypeptides. Recent data (seeHeurgueÂ-Hamard et al., 1998) and previous results(Menninger, 1985; Menninger et al., 1994) indicate,however, that peptidyl-tRNAs with long polypep-tides do drop-off, and this speaks against the useof a tunnel for polypeptide export. It is likely, how-ever, that there is a length-dependent rearrange-ment of the peptidyl-tRNA in relation to the P-site,and that this structural change makes peptidyl-tRNA less sensitive to the action of IF1 and IF2.Similar polypeptide length-dependence is observedfor the rate of peptidyl-tRNA drop-off catalysed bythe concerted action of release factors RF3, RRFand elongation factor EF-G (Tables 2 and 3). Thissimilarity suggests that the drop-off activities ofthe two sets of translation factors occur via A-sitebinding, and that the initiation factors have otherfunctional features in common with EF-G, RF3 andRRF.
The estimated drop-off frequency per codonin vivo is about 4 � 10ÿ4 (Menninger, 1976). Thepresent work shows that initiation factors IF1 andIF2 accelerate drop-off rate of peptidyl-tRNA withshort peptides from around 10ÿ3 sÿ1 to 10ÿ2 sÿ1. Ifone assumes that the ``average'' peptidyl-tRNAdrops off with a rate around 10ÿ2 sÿ1 under thein¯uence of IF1/IF2 and EF-G/RRF/RF3 (seeTable 2), and that reading of a codon by ternarycomplex occurs at a rate of 30 sÿ1, then a drop-offfrequency of about 3 � 10ÿ4 (0.01/30) per codon isobtained. A possible caveat with such a rough esti-mate is that factor-dependent drop-off seems toslow with increasing length of the polypeptides(see Table 2) and may therefore be less signi®cantfor the ``average'' peptidyl-tRNA in the cell.
The factor-independent drop-off rates for allpeptidyl-tRNAs studied in the present work areslow (Tables 2 and 3) and quite similar to thosepreviously measured for poly(U) translation(Karimi & Ehrenberg, 1996). Surprisingly, the fac-tor-free drop-off rates do not decrease with peptidelength in any regular manner. In one case, a hepta-peptidyl-tRNA is even less stably bound to theP-site than a tetrapeptidyl-tRNA (Tables 2 and 3).These observations show that the rate of factor-freedrop-off is determined by features of the nascentchain other than its length.
The present data suggest that IF1 and IF2 signi®-cantly enhance the drop-off frequency in vivo(Table 5). The evidence is that when IF1 and IF2are simultaneously overexpressed in Pth-de®cient(but not in wild-type) bacteria, their growth can beseverely impaired. These ®ndings together withprevious results concerning drop-off induction bya concerted action of EF-G and release factors RF3and RRF (HeurgueÂ-Hamard et al., 1998) imply thatcurrent views of drop-off must be modi®ed. Thephenomenon is not caused by spontaneous dis-sociation of peptidyl-tRNA from translating ribo-somes as hitherto thought. The dominatingpathways for drop-off are, instead, catalysed bytwo different sets of translation factors. Thisimplies that the frequency of drop-off in the bac-terial cell can be modulated and controlled by thepool-sizes of these proteins.
Materials and Methods
Chemicals and buffers
ATP, UTP, CTP, GTP and GDP were from Pharmacia(Uppsala, Sweden). GMP-PNP (guanylyl-imidodipho-sphate) was from Boehringer (Mannheim). Phosphoenol-pyruvate (PEP), putrescine, spermidine and non-radioactive amino acids were from Sigma. L-(Methyl-3H)-methionine was from Amersham (Solna, Sweden). Allother chemicals were of analytical grade from Merck(SpaÊnga, Sweden). Polymix buffer was prepared accord-ing to Jelenc & Kurland (1979) and contained 5 mMmagnesium acetate, 0.5 mM CaCl2, 5 mM NH4Cl,95 mM KCl, 8 mM putrescine, 1 mM spermidine, 5 mMpotassium phosphate (pH 7.3) and 1 mM dithioerythritol(DTE).

IF1 and IF2 Remove Peptidyl-tRNAs from Ribosomes 249
Components of the translation system
Enzymes and other components used in proteinelongation were puri®ed according to Ehrenberg et al.(1990). Initiation factors were puri®ed from overprodu-cing strains according to Sof®entini et al. (1994) withminor modi®cations. Release factors RF1 and RF3 wereprepared according to Freistroffer et al. (1997) and RRFaccording to MacDougall et al. (1997). f[3H]Met-tRNAMet
was prepared according to Hershey & Trash (1967) withminor modi®cations.
RNAguard (porcine) was from Pharmacia (Uppsala,Sweden), pyruvate kinase (PK) from Boehringer (Man-nheim) and myokinase (MK) from Sigma. Wild-typeE. coli ribosomes (017 and MRE600) were preparedaccording to Jelenc (1980) and stored at ÿ80�C in poly-mix buffer.
Synthetic mRNAs
The following mRNAs (coding sequences in parenth-eses) were prepared according to Pavlov et al. (1997a):mMFL(AUG UUC UUG), mMFMI(AUG UUC AUGAUC), mMFFG (AUG UUU UUC GGC), mMFTI(AUGUUU ACG AUU), mMFQI(AUG UUU CAG AUC),mMFITQLK (AUG UUC AUC ACC CAG CUG AAA)and mMFIKQLT(AUG UUC AUC AAA CAG CUGACC). They code for Met-Phe-Leu, Met-Phe-Met-Ile,Met-Phe-Phe-Gly, Met-Phe-Thr-Ile, Met-Phe-Gln-Ile, Met-Phe-Ile-Thr-Glu-Leu-Lys and Met-Phe-Ile-Lys-Glu-Leu-Thr, respectively. Outside the coding region they allhave identical sequences, as illustrated by the completesequence for mMFL (gggaauucgggcccuuguuaacaauuaag-gaagguauacuAUGUUCUUGUAAcugcagaaaaaaaaaaaaa-aaaaaaaa). It contains a leader, a ribosome-binding site(underlined boldface) as for 002 mRNA (Calogero et al.,1988), a spacer, a variable coding sequence (underlinedcapital letters), the stop codon UAA (capital letters) fol-lowed by cugcag and a poly(A) tail.
Peptidyl-tRNA hydrolase (Pth) purification
Pth was puri®ed from the overproducing E. coli strainC600/pBSAE53 (Garcia Villegas et al., 1991). Inducedcells (8 g) were suspended in 50 ml of buffer A (30 mMMes-NaOH (pH 6.8), 1.0 mM DTE, 0.1 mM phenyl-methylsulphonyl ¯uoride (PMSF) and lysed in a Frenchpress. The cell lysate was clari®ed by centrifugation (Sor-vall SS-34, 17,000 rpm for 30 minutes) and then appliedto an S-Sepharose Fast Flow column (5 cm � 30 cm,Pharmacia) equilibrated with buffer A. The protein waseluted with a 500 ml � 500 ml gradient from 0 M to 1 MNaCl in buffer A. Fractions corresponding to peaks inthe A280 elution pro®le were assayed for Pth activity andfor molecular mass by SDS-(10%) PAGE. The fractionsthat contained Pth activity represented the major absor-bance-peak and were eluted at 300 mM NaCl. They werepooled, precipitated with 70% saturated ammonium sul-phate, and pelleted by centrifugation. The protein wasdissolved in buffer B (50 mM potassium phosphate(pH 7.5), 1 mM DTE, 100 mM PMSF, 0.02% NaN3) andapplied to an AcA44 (Spectracor) gel ®ltration column(5 cm � 150 cm) equilibrated with buffer B. It was elutedat a ¯ow-rate of 25 ml/hour. Pth was identi®ed by SDS-PAGE and activity. Fractions were pooled, precipitatedwith 70% ammonium sulphate and collected by centrifu-gation. Pth was dissolved in a minimal volume of poly-mix buffer, dialysed and stored at ÿ25�C in 50% (v/v)glycerol. Pth (more than 95% pure according to SDS-10%
PAGE) was quanti®ed (Bradford; 1976) with bovineserum albumin as a standard.
Peptidyl-tRNA:ribosome complexes
Peptidyl-tRNA:ribosome complexes were preparedaccording to Freistroffer et al. (1997) with minor vari-ations.
A ribosome mix (in 0.5 ml of polymix buffer) was pre-pared with 4.5 nmol of ribosomes (017 or MRE 600),2.0 nmol of IF1, 0.2 nmol of IF2, 2.0 nmol of IF3, 2.0 mMGTP, 8.6 nmol of mRNA, 40 units of RNAguard and5.0 nmol of [3H]fMet-tRNA (speci®c activity 450 cpm/pmol) and incubated for 30 minutes at 37�C for initiationof translation. A factor mix (in 0.5 ml of polymix buffer)was prepared with 20 mM PEP, 2.0 mM ATP, 300 nmolof each of Phe, Leu, Ile, Thr, Gln, Lys and Met asrequired, 300 nmol of tRNAbulk, 1.3 nmol of EF-G,4.0 nmol of EF-Ts, 12 nmol of EF-Tu, 3.0 mg of MK,50 mg of PK, 1000 units of puri®ed PheRS, LeuRS, IleRS,ThrS, GlnRS, LysRS and MetRS as required, and incu-bated for ten minutes at 37�C for ternary complex for-mation. Protein elongation was started by addingtogether the ribosome and factor mixes. After 20 secondsincubation at 37�C, the elongation mix was cooled on iceand applied to a gel-®ltration column (SephacrylS-300HR, 5 cm � 45 cm, Pharmacia). Puri®ed peptidyl-tRNA:ribosome complexes were eluted in polymix bufferat 4�C, identi®ed by measuring A280 and scintillationcounting, pooled and frozen in aliquots at ÿ80�C. Toprepare fMFIK-tRNA:ribosome complexes by translationof mMFIKQLT mRNA, ribosomes were stalled with aGln codon in the A-site by excluding Gln, Leu, Thr andtheir corresponding aminoacyl-tRNA synthetases fromthe factor mix. Ribosome complexes with fMFL-tRNA,fMFTI-tRNA, fMFIT-tRNA, fMFITQ-tRNA and fMFITQL-tRNA were prepared using analogous strategies.
Drop-off of peptidyl-tRNA from ribosomes
Pth at high concentrations very rapidly hydrolysesfree but is inert with respect to ribosome-bound pepti-dyl-tRNAs. The dissociation rates of all the different pep-tidyl-tRNA:ribosome complexes studied here wereinvariably much slower than the rate of hydrolysis of thefree peptidyl-tRNAs catalysed by Pth. Accordingly, thedrop-off rates could be followed as the disappearance ofcold trichloroacetic acid-precipitable counts from[3H]fMet, present in the polypeptide of all peptidyl-tRNAs, since the polypeptides themselves but not thepeptidyl-tRNAs are soluble in trichloroacetic acid.
Ribosome:peptidyl-tRNA complexes were prepared inpolymix buffer as described in the previous section.A Pth mix to monitor the drop-off reaction was preparedin polymix buffer containing 2 mM GTP, 2 mM ATP,20 mM PEP, 100 mg/ml PK, 17 mg/ml MK, 0.6 mM Pthand amounts of IF1 and IF2 as speci®ed. In experimentswhere the effects of different guanine nucleotides (GTP,GDP or GMP-PNP) on the rate of drop-off were com-pared, the Pth mix was changed to contain 0.6 mM gua-nine nucleotide. When GDP was used, both PK and MKwere omitted. When the drop-off rates of peptidyl-tRNMet
f and peptidyl-tRNAMetm were studied, the Pth mix
contained, in addition, 20 mM deacylated tRNAMetf/m.
After separate preincubations of the Pth mix and therelease complexes for three minutes at 37�C, they wereadded together in equal volumes to form the drop-offmix, which was thoroughly vortex-mixed. Aliquots were

250 IF1 and IF2 Remove Peptidyl-tRNAs from Ribosomes
removed after different incubation times and the Pthactivity was stopped either by addition of 8 ml of ice-cold 5% (w/v) trichloroacetic acid or of an equal volumeof ice-cold 25% (v/v) formic acid.
The samples that had been quenched with trichloroa-cetic acid were collected on GF/C ®lters, dried, andcounted according to Ehrenberg et al. (1990). Plots of thenatural logarithm of the amount or fraction of peptidyl-tRNA remaining unhydrolysed after different incubationtimes were linear, demonstrating ®rst-order kinetics forall drop-off reactions studied. Drop-off rate constantswere calculated from the slopes of these lines and drop-off times were obtained as the reciprocal values of therespective rate constants. Each drop-off experiment wasrepeated at least twice, and the effects of different factorson drop-off rates were always studied in parallell exper-iments.
In those cases where the drop-off rates were particu-larly slow, the precision of the measurements wasenhanced by detection also of the amount of polypeptidecleaved off along with the amount of peptidyl-tRNAremaining after different incubation times. In those cases,the Pth activity was quenched with formic acid (seeabove in this section) rather than with trichloroaceticacid. The samples were spun down in an Eppendorf cen-trifuge and the supernatant containing peptides cleavedoff peptidyl-tRNA was applied directly to GF/C ®lters,which were dried and counted as above. The pellet, con-taining peptidyl-tRNAs, was dissolved in 0.5 M KOH,applied to GF/C ®lters, dried and counted as above.
The relative errors in drop-off times were 15% orsmaller as estimated from comparison of independentexperiments.
Ribosome recycling assays
Recycling experiments were carried out essentially asdescribed by Pavlov et al. (1997b). When drop-off stimu-lation by IF1 and IF2 was studied, release factors RF1,RF3 and RRF were excluded.
Two mixes were prepared in polymix buffer. A ribo-some mix containing (in 50 ml) 250 pmol of ribosomes(about 70 pmol active in protein elongation), 18 pmol ofIF1, 18 pmol of IF2, 250 pmol of IF3, 500 pmol ofmRNA, 1.0 mM GTP and 1 nmol of [3H]fMet-tRNA. Afactor mix containing (in 180 ml) 0.05 mM of eachrequired aminoacid, 8 nmol tRNAbulk (E. coli), 120 units(de®ned by Ehrenberg et al., 1990) of each required ami-noacyl-tRNA synthetase, 1.2 mM magnesium acetate,1.2 mM Mg(OAc)2, 12 mM PEP, 1 mM GTP, 5 mg of PK,0.3 mg of MK, 12 mM EF-Tu, 1 mM EF-Ts and 2 mM EF-G.The concentrations of release factor RF3, ribosome recy-cling factor RRF, IF1 and IF2 were as speci®ed in eachexperiment.
The ribosome and factor mixes were pre-incubated forten minutes at 37�C for initiation of translation and tern-ary complex formation, respectively. Ribosome recyclingwas started by adding 180 ml factor mix to 50 ml ribo-some mix. Aliquots (40 ml) were withdrawn after differ-ent incubation times, quenched with 20 ml of 40% formicacid and the precipitate spun down in an Eppendorf cen-trifuge. The pellet (containing peptidyl-tRNA) was dis-solved in 180 ml of 0.5 M KOH and incubated for 15minutes at 22�C for hydrolysis of peptidyl- and aminoa-cyl-tRNAs. Concentrated formic acid (10 ml) was thenadded to the samples and the solutions were clari®ed bycentrifugation. The amounts of radioactive polypeptideswere quanti®ed with HPLC according to Pavlov et al.,
(1997b). When release factors RF1 or RF2 were present inthe recycling experiments, the amounts of synthesisedpolypeptides were quanti®ed according to a proceduredescribed by Pavlov et al. (1997b).
The amount of synthesised peptidyl-tRNA plotted ver-sus time gives a straight line (as in Figure 1(a)). Theintercept at the y-axis is the amount of active ribosomesand the slope, normalised to the amount of active ribo-somes, is the ribosomal recycling rate. The recycling timeis obtained as the reciprocal value of the ribosomal recy-cling rate. In the absence of RF1 and RF2, the ribosomalrecycling time is an estimate of the drop-off time of thecorresponding peptidyl-tRNA, since drop-off is the rate-limiting step in ribosomal recycling in the absence of ter-mination by release factors.
Bacterial strains, plasmids and growth experiments
Initiation factors IF1 or IF2 alone or the two factorstogether were overexpressed from plasmids pACinfA,pACinfB (Guillon et al., 1992) and pACinfAB, respect-ively. Plasmid pACinfA (containing the infA gene) wasconstructed by cloning the HindIII-ScaI fragment con-taining infA from pTH20 (Sands et al., 1987) intopACYC184 digested with HindIII and EcoRV. PlasmidpACinfAB (containing the infA and infB genes) was con-structed by cloning the same HindIII-ScaI insert intopACinfB (Guillon et al., 1992) digested with HindIII andNruI. E. coli K12 strain IB188 has been described(Buckingham et al., 1990). The Tn 10 transposon fromVH7 (HeurgueÂ-Hamard et al., 1996) was transduced byphage P1 to strain IB188, producing VH200. Relativegrowth-rates of bacterial strains transformed with plas-mids overproducing initiation factors were compared onLB-agar medium with 15 mg/ml chloramphenicol(Miller, 1992) by colony size or patch growth after repli-ca plating (Buckingham et al., 1990).
Acknowledgements
We thank John W. B. Hershey and Jean-MichelGuillon for kind gifts of plasmids and Gabriel Guarnerosfor the Pth strain. This work was supported by theSwedish Natural Science Research Council, the SwedishResearch Council for Engineering Sciences, the CentreNational pour la Recherche Scienti®que (UPR9073),l'Association pour la Recherche sur le Cancer, the Minis-teÁre de l'Education National, de l'Enseignement SupeÂr-ieure et de la Recherche (ACCSV No. 1), the Fondationpour la Recherche Medicale, and the Human Capital andMobility Programme of the European Community.
References
Agrawal, R. K., Penczek, P., Grassucci, R. A., Li, Y.,Leith, A., Nierhaus, K. H. & Frank, J. (1996). Directvisualization of A-, P-, and E-site transfer RNAs inthe Escherichia coli ribosome. Science, 271, 1000±1002.
Atherly, A. G. (1978). Peptidyl-transfer RNA hydrolaseprevents inhibition of protein synthesis initiation.Nature, 275, 769.
Atherly, A. G. & Menninger, J. R. (1972). Mutant Escheri-chia coli strain with temperature sensitive peptidyl-transfer RNA hydrolase. Nature, 240, 245±246.

IF1 and IF2 Remove Peptidyl-tRNAs from Ribosomes 251
Benne, R., Naaktgerboren, N., Gubbens, J. & Voorma,H. O. (1973). Recycling of initiation factors IF-1, IF-2and IF-3. Eur. J. Biochem. 32, 372±380.
Bradford, M. M. (1976). A rapid and sensitive methodfor the quantitation of microgram quantities of pro-tein utilizing the principle of protein-dye binding.Anal. Biochem. 72, 248±254.
Buckingham, R. H., Murgola, E. J., Sorensen, P., Pagel,F. T., Hijazi, K. A., Mims, B. H., Figueroa, N.,Brechemier-Baey, D. & Coppin-Raynal, E. (1990).Effects of codon context on the suppression of non-sense and missense mutations in the trpA gene ofEscherichia coli. In Ribosomes: Structure, Function andEvolution (Hill, W. E., Dahlberg, A., Garrett, R. A.,Moore, P. B., Schlessinger, D. & Warner, R., eds),pp. 541±545, American Society for Microbiology,Washington, DC.
Butler, J. S., Springer, M., Dondon, J., Graffe, M. &Grunberg-Manago, M. (1986). Escherichia coli proteinsynthesis initiation factor IF3 controls its own geneexpression at the translational level in vivo. J. Mol.Biol. 192, 767±780.
Calogero, R. A., Pon, C. L., Canonaco, M. A. &Gualerzi, C. O. (1988). Selection of the mRNA trans-lation initiation region by E. coli ribosomes. Proc.Natl Acad. Sci. USA, 85, 6427±6431.
Chapeville, F., Yot, P. & Paulin, D. (1969). Enzymatichydrolysis of N-acylaminoacyl transfer RNAs. ColdSpring. Harbour. Symp. Quant. Biol. 34, 493±498.
Chen, G. T. & Inouye, M. (1990). Suppression of thenegative effect of minor arginine codons on geneexpression; preferential usage of minor codonswithin the ®rst 25 codons of the Escherichia coligenes. Nucl. Acids Res. 18, 1465±1473.
Celano, B., Pawlik, R. T. & Gualerzi, C. O. (1988). Inter-action of Escherichia coli translation-initiation factorIF-1 with ribosomes. Eur. J. Biochem. 178, 351±355.
Cummings, H. S. & Hershey, J. W. (1994). Translationinitiation factor IF1 is essential for cell viability inEscherichia coli. J. Bacteriol. 176, 198±205.
Cuzin, F., Kretchmer, N., Greenberg, R. E., Hurwitz, R.& Chapeville, F. (1967). Enzymatic hydrolysis of N-substituted aminoacyl-tRNA. Proc. Natl Acad. Sci.USA, 58, 2079±2086.
Draper, D. E. (1996). Translational initiation. In Escheri-chia coli and Salmonella: Cellular and MolecularBiology (Neidhart, F. C., et al., ed.), pp. 902±908,ASM Press, Washington, DC.
Dubnoff, J. S., Lockwood, A. H. & Maitra, U. (1972).Studies on the role of guanosine triphosphate inpolypeptide chain Initiation in Escherichia coli. J. Biol.Chem. 247, 2884±2894.
Ehrenberg, M., Bilgin, N. & Kurland, C. G. (1990).Design and use of a fast and accurate in vitro trans-lation system. In Ribosomes and Protein Synthesis(Spedding, G., ed.), pp. 101±129, IRL Press, OxfordUniversity Press, Oxford and New York.
Fakunding, J. L. & Hershey, J. W. B. (1973). The inter-action of radioactive initiation factot IF-2 with ribo-somes during initiation of protein synthesis. J. Biol.Chem. 248, 4206±4212.
Frank, J., Penczek, P., Li, Y., Srivastava, S., Verschoor,A., Radermacher, M., Grassucci, R., Lata, R. K. &Agrawal, R. K. (1995). A model of protein synthesisbased on cryo-electron microscopy of the E. coliribosome. Nature, 376, 441±444.
Freistroffer, D. V., Pavlov, M. Y., MacDougall, J.,Buckingham, R. H. & Ehrenberg, M. (1997). Releasefactor RF3 in E. coli accelerates the dissociation of
release factors RF1 and RF2 from the ribosome in aGTP dependent manner. EMBO J. 16, 4126±4133.
Garcia, Villegas M. R., De La Vega, F. M., Galindo, J. M.,Segura, M., Buckingham, R. H. & Guarneros, G.(1991). Peptidyl-tRNA hydrolase is involved inlambda inhibition of host protein synthesis. EMBOJ. 10, 3549±3555.
Gualerzi, C. O. & Pon, C. L. (1990). Initiation of mRNAtranslation in prokaryotes. Biochemistry, 29, 5881±5889.
Guillon, J. M., Meinnel, T., Mechulam, Y., Lazennec, C.,Blanquet, S. & Fayat, G. (1992). Nucleotides oftransfer RNA governing the speci®city of Escherichiacoli methionyl-transfer RNAMet
f formyltransferase.J. Mol. Biol. 224, 359±367.
Hershey, J. W. B. (1987). Protein synthesis. In Escheri-chia coli and Salmonella typhimurium: Cellular andMolecular Biology (Neidthardt, F. G., ed.), p. 613,American Society for Microbiology, Washington,DC.
Hershey, J. W. B. & Trash, R. E. (1967). Role of GTP ininitiation of peptide synthesis, I: synthesis of for-mylmethiononyl-puromycin. Proc. Natl Acad. Sci.USA, 57, 759±766.
HeurgueÂ-Hamard, V., Mora, L., Guarneros, G. &Buckingham, R. H. (1996). The growth defect inE. coli de®cient in peptidyl-tRNA hydrolase is dueto starvation for Lys tRNALys. EMBO J. 15, 2826±2833.
HeurgueÂ-Hamard, V., Karimi, R., Mora, L., MacDougall,J., Leboeuf, C'., Grentzmann, G., Ehrenberg, M. &Buckingham, R. H. (1998). Ribosome release factorRF4 and termination factor RF3 are involved in dis-sociation of peptidyl-tRNA from the ribosome.EMBO J. 17, 808±816.
Howe, J. G. & Hershey, J. W. B. (1981). A sensitiveimmunoblotting method for measuring protein syn-thesis initiation factor levels in lysates of E. coli.J. Biol. Chem. 256, 12836±12839.
Howe, J. G., Yanov, J., Meyer, L., Johnston, K. &Hershey, J. W. (1978). Determination of protein syn-thesis initiation factor levels in crude lysates ofEscherichia coli by a sensitive radioimmune assay.Arch. Biochem. Biophys. 191, 813±820.
Jelenc, P. C. (1980). Rapid puri®cation of highly activeribosomes from Escherichia coli. Anal. Biochem. 105,369±374.
Jelenc, P. C. & Kurland, C. G. (1979). Nucleotide tripho-sphate regeneration decreases the frequency oftranslation errors. Proc. Natl Acad. Sci. USA, 76,3174±3178.
Karimi, R. & Ehrenberg, M. (1994). Dissociation rate ofcognate peptidyl-tRNA from the A-site of hyperac-curate and error-prone ribosomes. Eur. J. Biochem.226, 355±360.
Karimi, R. & Ehrenberg, M. (1996). Dissociation rates ofpeptidyl-tRNA from the P-site of Escherichia coliribosomes. EMBO J. 15, 1149±1154.
Kossel, H. & RajBhandary, U. L. (1968). Studies on poly-nucleotides. LXXXVI. enzymatic hydrolysis of N-acylaminoacyl-transfer RNA. J. Mol. Biol. 35, 539±560.
Kurland, C. & Ehrenberg, M. (1985). Constraints on theaccuracy of mRNA movement. Quart. Rev. Biophys.18, 423±450.
Kurland, C. G., Hughes, D. & Ehrenberg, M. (1996).Limitations of translational accuracy. In Escherichiacoli and Salmonella: Cellular and Molecular Biology

252 IF1 and IF2 Remove Peptidyl-tRNAs from Ribosomes
(Neidhardt, F. C., ed.), pp. 979±1004, ASM Press,Washinghton, DC.
MacDougall, J., Holst-Hansen, P., Mortensen, K. K.,Freistroffer, D. V., Pavlov, M. Y., Ehrenberg, M. &Buckingham, R. H. (1997). Puri®cation of activeEscherichia coli ribosome recycling factor, RRF froman osmo-regulated expression system. Biochimie, 79,243±246.
Menninger, J. R. (1976). Peptidyl-transfer RNA dis-sociates during protein synthesis from ribosomes ofE. coli. J. Biol. Chem. 251, 3392±3398.
Menninger, J. R. (1977). Ribosome editing and the errorcatastrophe hypothesis of cellular aging. Mech. Age-ing Dev. 6, 131±142.
Menninger, J. R. (1985). Functional consequences ofbinding macrolides to ribosomes. J. Antimicrob. Che-mother. 16, 23±34.
Menninger, J. R., Coleman, R. A. & Tsai, L. N. (1994).Erythromycin, lincosamides, peptidyl-tRNA dis-sociation, and ribosome editing. Mol. Gen. Genet.243, 225±233.
Miller, J. H. (1992). A Short Course in Bacterial Genetics,Cold Spring Harbor Laboratory Press, Cold SpringHarbor, NY.
Moazed, D., Samaha, R. R., Gualerzi, C. & Noller, H. F.(1995). Speci®c protection of 16 S rRNA by transla-tional initiation factors. J. Mol. Biol. 248, 207±210.
O'Farrel, P. H. (1978). The suppression of defectivetranslation by ppGpp and its role in the stringentresponse. Cell. 14, 545±557.
Pavlov, M. & Ehrenberg, M. (1996). Rate of translationof natural mRNAs in an optimised in vitro trans-lation system. Arch. Biochem. Biophys. 328, 9±16.
Pavlov, M. Y., Freistroffer, D. V., MacDougall, J.,Buckingham, R. H. & Ehrenberg, M. (1997a). Fastrecycling of E. coli ribosomes requires both ribo-
some recycling factor (RRF) and release factor RF3.EMBO J. 16, 4134±4141.
Pavlov, M. Y., Freistroffer, D. V., HeurgueÂ-Hamard, V.,Buckingham, R. H. & Ehrenberg, M. (1997b).Release factor RF3 abolishes competition betweenrelease factor RF1 and ribosome recycling factor(RRF) for a ribosome binding site. J. Mol. Biol. 273,389±401.
Sands, J. F., Cummings, H. S., Sacerdot, C., Dondon, L.,Grunberg-Manago, M. & Hershey, J. W. (1987).Cloning and mapping of infA, the gene for proteinsynthesis initiation factor IF1. Nucl. Acids Res. 15,5157±5168.
Sette, M., van Tilborg, P., Spurio, R., Kaptein, R., Paci,M., Gualerzi, C. O. & Boelens, R. (1997). The struc-ture of the translational initiation factor IF1 fromE. coli contains an oligomer-binding motif. EMBO J.16, 1436±1443.
Shiba, K., Ito, K., Nakamura, Y., Dondon, J. &Grunberg-Manago, M. (1986). Altered translationinitiation factor 2 in the cold-sensitive ssyG mutantaffects protein export in Escherichia coli. EMBO J. 5,3001±3006.
Sof®entini, A., Lorenzetti, R., Gastado, L., Spurio, R., LaTeana, A. & Khalid, I. (1994). Puri®cation procedurefor bacterial initiation factors IF1 and IF2. Exp.Purif. 5, 118±124.
Stade, K., Junke, N. & Brimacombe, R. (1995). Mappingthe path of the nascent peptide chain through the23S RNA in the 50S ribosomal subunit. Nucl. AcidsRes. 23, 2371±2380.
Wagner, E. G. H., Ehrenberg, M. & Kurland, C. G.(1982). Kinetic suppression of translational errors by(p)ppGpp. Mol. Gen. Genet. 185, 269±274.
Yonath, A. & Berkovitch-Yellin, Z. (1993). Hollow,voids, gaps and tunnel in the ribosome. Curr. Opin.Struct. Biol. 3, 175±181.
Edited by J. Karn
(Received 16 February 1998; received in revised form 22 May 1998; accepted 27 May 1998)




















