
The electrophoretic characterization of ribosomes from the blue-green alga Anabaena flos-aquae
10
Arch. Mikrobiol. 86, 39--48 (1972) by Springer-Verlag 1972 The Electrophoretic Characterization of Ribosomes from the Blue-Green Alga Anabaena flos-aquae John i~. CA~ZTON and DIANE S. H:EI~SON University of Delaware, Newark, Delaware 19711, U.S.A. l~eceived June 15, 1972 Summary. 1.1%ibosomesof the blue-green alga, Anabaena/los-aquae, were isolat- ed using a rapid lysis method and by grinding with alumina. 2. Analysis of ribosomes obtained by rapid lysis on sucrose gradients and electro- phoretie gels indicated the presence of 30S, 50S, 708, and 100S ribosomes as well as polysomes. 3. Dissociation of algal 70S and 100S ribosomes into their 30S and 50S sub- units was demonstrated using a 6000 • g preparation on a two-dimensional electro- phoretie gel as well as a 105000 • g fraction on a sucrose gradient. 4. Differences between blue-green algal and bacterial ribosomes were noted when 105000 • g fractions of both were washed in ribosomal washing buffer. A major 100S or dimer peak was observed for the bacterial preparation before and after washing. The major peak in the algal preparation shifted from 100S to 70S after washing. Most studies which have characterized proearyotic ribosomes have been conducted with Eseherichia coli ribonueleoprotein particles (Tis- si6res and Watson, i958). The techniques most commonly used in these studies were electron microscopy (Tissibres et al., i959), analytical ultracentrifugation (tIolm-Hansen, 1968), and sucrose gradient centri- fugation. Recent reports have indicated that electrophoretic characteri- zation can be done in conjunction with the above methods, tIjerten et al. (1965) used electrophoresis in an attempt to separate ribosomal particles and demonstrated E. coli 50 S and 30 S subunits. The most detailed research on the electrophoretic characterization of ribosomes was done by Dahlberg et al. with an E. coli system (1969). These workers employed slab-gel electrophoresis to demonstrate 30S, 508, 70S, and 100S ribosomal particles as well as polysomes. A turned- gel technique further enabled them to separate the 70 S and 100 S particles into their subunits. Abbreviations: O.D.26 ~ = optical density observed at 260 m~z. $6 = the super- natant fraction obtained by centrifuging cell extracts at 6000 • g for five minutes. P105 = the pellet obtained by centrifuging a ribosomal extract at 105000 • g for 90 rain. 1' ~ = indication of direction of first dimension of gel; 2' ~ = indication of direction of second dimension of gel.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of The electrophoretic characterization of ribosomes from the blue-green alga Anabaena flos-aquae

Arch. Mikrobiol. 86, 39--48 (1972) �9 by Springer-Verlag 1972
The Electrophoretic Characterization of Ribosomes from the Blue-Green Alga Anabaena flos-aquae
J o h n i~. CA~ZTON and DIANE S. H:EI~SON
University of Delaware, Newark, Delaware 19711, U.S.A.
l~eceived June 15, 1972
Summary. 1.1%ibosomes of the blue-green alga, Anabaena/los-aquae, were isolat- ed using a rapid lysis method and by grinding with alumina.
2. Analysis of ribosomes obtained by rapid lysis on sucrose gradients and electro- phoretie gels indicated the presence of 30S, 50S, 708, and 100S ribosomes as well as polysomes.
3. Dissociation of algal 70S and 100S ribosomes into their 30S and 50S sub- units was demonstrated using a 6000 • g preparation on a two-dimensional electro- phoretie gel as well as a 105000 • g fraction on a sucrose gradient.
4. Differences between blue-green algal and bacterial ribosomes were noted when 105000 • g fractions of both were washed in ribosomal washing buffer. A major 100S or dimer peak was observed for the bacterial preparation before and after washing. The major peak in the algal preparation shifted from 100S to 70S after washing.
Most studies which have characterized proearyotic ribosomes have been conducted with Eseherichia coli r ibonueleoprotein particles (Tis- si6res and Watson, i958). The techniques most commonly used in these studies were electron microscopy (Tissibres et al., i959), analyt ical u l t racent r i fugat ion ( t Io lm-Hansen, 1968), and sucrose gradient centri- fugation. Recent reports have indicated tha t electrophoretic characteri- zat ion can be done in conjunct ion with the above methods, t I j e r t en et al. (1965) used electrophoresis in an a t t empt to separate r ibosomal particles and demons t ra ted E. coli 50 S and 30 S subunits .
The most detailed research on the electrophoretic character izat ion of ribosomes was done by Dahlberg et al. with an E. coli system (1969). These workers employed slab-gel electrophoresis to demonst ra te 30S, 508, 70S, and 100S ribosomal particles as well as polysomes. A turned- gel technique fur ther enabled them to separate the 70 S and 100 S particles into their subuni ts .
Abbreviations: O.D.26 ~ = optical density observed at 260 m~z. $6 = the super- natant fraction obtained by centrifuging cell extracts at 6000 • g for five minutes. P105 = the pellet obtained by centrifuging a ribosomal extract at 105000 • g for 90 rain. 1' ~ = indication of direction of first dimension of gel; 2' ~ = indication of direction of second dimension of gel.

40 J. R. Carlton and D. S. Herson:
Blue-green algae, like the bacteria, are considered to be procaryons. Both groups of organisms possess a simple cell s t ructure and lack mem- brane enclosed organelles. Taylor and Storck (1964) demonst ra ted tha t the ribosomes fl'om Anabaena eylindrica, a f i lamentous blue-green alga, had a sedimenta t ion value of 71.6 S and ribosomes from Anacystis nidu- lans, a unicellular blue-green alga, had a sedimenta t ion value of 69.0S. Both of these values are characteristic of procaryotic (70S) ribosomes. Craig and Carr (1968) also demonst ra ted the procaryotic na ture of blue- green alga r ibonucleoprotein particles in their s tudy characterizing Anabaena variabilis ribosomes.
The present communica t ion is an a t t emp t to characterize the ribo- somes from the blue-green alga Anabaena flos-aquae using sucrose gra- dient centr ifugation and slab-gel electrophoresis.
Materials and Methods
Materials. The vertical gel electrophoresis cell was obtained from the E-C Apparatus Corporation of Philadelphia, Pennsylvania. The 4100 Pulsed Constant Power Supply was obtained from Ortec Inc. of Oak Ridge, Tennessee. Stains-all was obtained from Eastman Organic Chemicals. Brij 58 was a gift from the Atlas Chemical Company of Wilmington, Delaware.
Growth Condition~. E. coli wild-type cells used for S 6 preparations were grown in 1~ peptone broth at 37~ in a shaker water bath until a Klett reading of 150 was obtained. The ceils were then chilled within 20 sec. to 2~ and harvested by centri- fugation at 3000 • g for 15 rain. To keep polysomes intact the cells were broken immediately.
E. coli cells used for P105 preparations were obtained from Grain Processing Corporation of Muscatine, Iowa.
The blue-green alga, Anabaena/los-aquae, was grown in ]V[odified Chu :No. 10 plus trace elements (Schweiger et a[., 1969). One liter of culture was grown in a two-liter flask with 100 foot candles of light and 0.50/0 CO s plus air. Cultures were checked for contamination by streaking on Czapek and nutrient agar plates, as well as by adding aliquots to Chu and Chu plus 1~ peptone tubes. Cells were har- vested using the Sorvall KSB Continuous Flow System at 10000 rpm.
Preparation o/P105. An E. eoli P105 extract was prepared by grinding cells with an equal volume of alumina, resuspending the cell paste in ~irenberg buffer (0.1)/I Tris-HC1, pH 7.8; 0.01 M Mg Acetate; 0.06 M :NH4C1; and 0.006 M fi-mer- eaptoethanol) (:Nirenberg and Matthaei, 1961) and then centrifuging at 20000 • g for 20 rain to remove the alumina and unbroken cells. The supernatant (S 20) was then spun at 30000 • g for 30 rain and the supernatant (S 30) was spun at 105000 • g for 90 min. ]):Nase was added prior to the 30000 • g centrifugation. The 105000 • g pellet was then washed twice with ribosomal washing buffer (0.1 M tris-HC1, pH 7.8; 0.05 M KC1; 0.006 M fi-mercaptoethanol; 0.01 3/[ Mg Acetate and 0.5 M :NH4C1 ) (Stanley et al., 1966).
The same procedure was followed for the preparation of A. /los-aquae P 105 with the following exceptions: 2~ sodium desoxycholate was added to the super- natant ($20) following the 20000 • g spin and was agitated in the cold for 30 min. The green pellet obtained after the first 105000 • g spin was homogenized in a tefion

Blue-Green Alga Ribosomes 41
homogenizer with ribosomal washing buffer and spun again at 30000• The resulting $30 was spun at 105000• for 90 min. The resultant ribosomal pellet (P 105) was clear and gelatinous.
Dissociation o/ Ribosomes. The dissociation of 100S and 70S particles into 50S and 30S was accomplished by washing a ribosomal pellet twice in 0.0001 M Mg ++ Nirenberg buffer, followed by resuspension in 0.01 M phosphate buffer, pH 7.0.
Preparation o /$6. S 6 preparations were obtained from fleshly harvested E. coli or Anabaena /los-aquae cells. A modification of Godson and Sinsheimer (1967) was used as follows: Per 0.2 g of cells 0.4 ml of a 25~ sucrose solution in 0.01 M Tris- HC1, p i t 8.1 was added and well mixed. A lysozyme-EDTA solution was prepared by mixing equal volumes of EDTA (disodium salt) dissolved in distilled water at a concentration of 2.7 mg/ml and lysozyme dissolved in 0.25 M Tris-HC1, pH 8.1 at a final concentration of 0.08 mg/ml. 0.1 ml of the lysozyme-EDTA solution was added to the cell suspension and mixed by gentle swirling. After 45 to 60 sec, the suspen- sion was transferred to the lyric mixture which contained 0.1 ml of a 5~ Brij 58 solution dissolved in 0.1 M Tris-HC1, pH 7.2, 0.1 ml of 0.1 M MgSOt, 0.05 ml of a DNase stock containing 1 mg/ml in sterile distilled water and 0.2 ml of a 1~ DOC solution dissolved in 0.1 M Tris-HCl, pH 8.1. The lytic mixture was prepared just prior to use. The cell-lysozyme-EDTA-lytic mixture was well mixed and placed in an ice bath. After 2--5 rain the suspension was centrifuged at 6000 X g for 5 rain. The resulting supernate containing ribosomes, cell walls and membrane fragments is designated S 6.
Stains. The stain used in all cases was Stains-all. The staining procedure used was that of Dahlberg et al. (1969).
Gradients. A Buchler Universal Density Gradient Sedimentation System was used in the preparation of the sucrose gradients. Gradients were run in a Beckman L2-65B Ultracentrifuge with the rotor, speed and time designated in the text. Absorbance at 260 nm was monitored on a Gilford Spectrophotometer equipped with flow-ceil 203.
The following amounts of material were layered onto the sucrose gradients: E. coli P105, 1.50 0.D.26 o units; E. coli $6, 2.0 0.D.260 units; A. flos-aquae P105, 1.5 0.D.26 o units; A./los-aquae $6, 5.0 0.D.260 units.
Electrophoretic Gels. Composite agarose-acrylamide gels were prepared accord- ing to Dahlberg et al. (1969). Buffer I was composed of 0.025 M tris-HCl, 0.001 M Mg Cle, pH 8.0. Buffer I I was composed of 0.025 M tris-HC1, 0.0025 M disodium EDTA, pH 8.0.
The following amounts of material were layered onto the gels: E. coli 1)105, 0.15 0.D.260 units; E. coli $6, 0.7--0.9 0.D.26 o units; A. /los-aquae 1)105, 0.15--0.2 O.D.2s ~ units; A./los-aquae $6, 2.5--2.7 0.D.260 units.
Plates were prepared using Kodak film and a Minolta SR-T 101 camera. Densito- grams were prepared by scanning Polaroid transparency films of the gels at 540 m~.
Results
Analysis of Bacterial and Algal Ribosomes O b t a i n e d b y R a p i d Lys i s o f Cells
To o b t a i n a r i b o s o m a l prof i le a p p r o x i m a t i n g in vivo cond i t ions for
b o t h b a c t e r i a a n d algae, we used t h e r a p i d lysis m e t h o d of Godson a n d S in she imer (1967). T h e r e su l t i ng s u p e r n a t a n t f rac t ion , d e s i g n a t e d $6 ,
h a d a final m a g n e s i u m ion c o n c e n t r a t i o n o f 1 raM. A l t h o u g h the g r a d i e n t

.20
.15
0 0260
.10
.05
42 J. R. Carlton and D. S. Herson:
I E. COLI / - s T - 7os I ,oos^
! f I I I l 2 3 4
P4L.
~ig. 1
20 -
15
o D2~ o
.I0
.05
?os
50S 30S S6
I I I I
I 2 3 4 ML.
Fig. 2
Fig. 1. E. coli $6 layered on a linear 15--30~ sucrose-Nirenberg buffer gradient and run at 48000 rpm for one hour in a Beckman SW 50.1 rotor
Fig. 2. Anabaena flos-aquae S 6 layered on a linear 15--30~ sucrose Nirenberg buffer gradient and run at 48000 rpm for one hour in a Beckman SW 50.1 rotor. For all algal figures and plate: Material designated 30S is to be interpreted as small sub-
unit, 50S as large subunit, 70S as monomer and 100S as dimer
profiles of the bacterial and algal ribosomes were similar, the rates of sedimentat ion were not identical (Figs. 1 and 2). As Tissi~res et al. (1959) have shown tha t the ribosomal particles of E. co/[+under similar buffer conditions have S values of 30S, 50S, 70S and 100S for the small sub- unit, large subunit, monomer and direct, respectively, we are designating the E. coli ribosomes by the appropriate S value. The algal ribosomal units are designated by the ]ess precise terms, small subunit, large sub- unit, monomer and dimer.The heavier material sedimenting on the gra- dients most likely indicates the presence of polysomes.
Additional information on the nature of these ribosomal particles was obtained using slab-gel electrophoresis. High molecular weight partic- les with a net negative charge such as ribosomes and polysomes can be separated on composite agarose-acrylamide gels. The smaller particles, or subunits would be expected to migrate fastest. The banding pa t te rn of the algae ribosomes on the gel was similar to t ha t of the gradient (Figs. 2 and 3).
To more conclusively demonstrate the composition of the particles in each band, we employed two-dimensionM electrophoresis, a unique feature of the slab gel-technique. Blue-green algal $6 fractions were electrophoresed in the first two slots (Fig. 4, slots A and B) to obtain a typical one-dimensional banding pat tern. Slot A was cut off and the remainder of the gel was soaked in E D T A to chelate magnesium, then
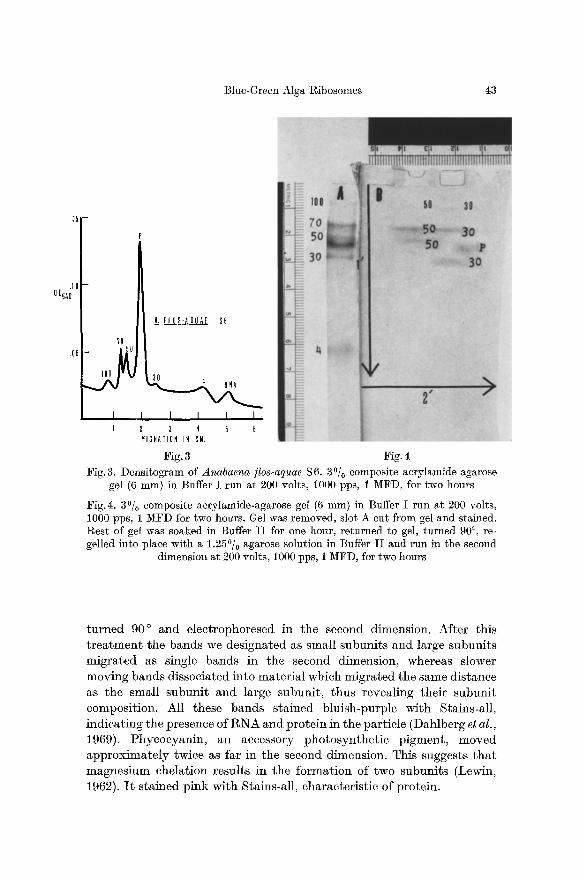
Blue-Green Alga Ribosomes 43
o D~.o I o
1
A= FLOS~AOUAE S8
I I I r I ~ 3 4 5
MIGRATION IN ;M.
Fig. 3 Fig. 4 Fig. 3. Densitogram of Anabaena tics-aquae $6. 30/0 composite aerylamide-agarose
gel (6 mm) in Buffer I run at 200 volts, 1000 pps, 1 MFD, for two hours
Fig.4. 3~ composite acrylamide-agarose gel (6 mm) in Buffer I run at 200 volts, 1000 pps, 1 MFD for two hours. Gel was removed, slot A cut from gel and stained. Rest of gel was soaked in Buffer I I for one hour, returned to gel, turned 90 ~ re- gelled into place with a 1.25~ agarose solution in Buffer I I and run in the second
dimension at 200 volts, 1000 pps, 1 MFD, for two hours
t u r n e d 90 ~ and e lec t rophoresed in the second dimension. Af te r this t r e a t m e n t the bands we des igna ted as small snbuni t s and large subuni t s mig ra t ed as single bands in the second dimension, whereas slower moving bands d issoc ia ted in to ma te r i a l which mig ra t ed the same d is tance as the smal l subnni t and large subuni t , thus reveal ing the i r subun i t composi t ion. All these bands s ta ined b lu ish-purple wi th Stains-al l , i nd ica t ing the presence of R N A and pro te in in the par t ic le (Dahlberg et al., 1969). Phyeocyan in , an accessory pho tosyn the t i c p igment , moved a p p r o x i m a t e l y twice as far in the second dimension. This suggests t h a t magnes ium chela t ion resul ts in the fo rma t ion of two subuni t s (Lewin, 1962). I t s t a ined p ink wi th Stains-al l , charac ter i s t ic of prote in .

.30
.25
20 00250
.15
.]0
.05
50 S .35 -
. /
.L ~OLI PI05
30S
30 50S
25 -- \ A_. FLOS-AOUAE =
20 \ P105
0 D~6 o
�9 15 - L JO -
.05 - /
I I I I I I I I 1 2 3 4 5 1 2 3 4 5
ML ML, Fig. 5 Fig. 6
Fig.5. E. coli P105 washed twice with 0.1 mM Mg ++ Nirenberg buffer, layered on a linear 15--35~ sucrose-low Mg ++ (0.1 ram) tris buffer gradient and run for 130 min
at 48 000 rpm in a SW 50.1 rotor
Fig.6. A. /los-aquae P105 washed twice with 0.1 mM Mg ++ Nirenberg buffer, layered on a linear 15--35~ sucrose-low Mg ++ (0.1 mM) tris buffer gradient and
run for 130 rain at 48000 rpm in a SW 50.1 rotor
35
100
25 L.~ LI 1QOS#~~?OS 1Of
20 PI 0 5 # v I 0 D260
15 ~ 0Ds~'0 P J I '705
i/ 'lri IOOS
I I I I I 2 3 4 5 ~ 4
4s
DH ONA
I I I B lO 12
F4L. MICRATION IN CM Fig. 7 Fig. 8
Fig.7. E. coli 1)105 layered on a linear 15 - - 30% sucrose-Nirenberg buffer gradient and run for one hour at 48000 rpm in a SW 50.1 rotor
Fig.8. Densitogram of unwashed bacterial and algal P105 fractions. 2.25~ com- posite acrylamide-agarose gel (6 mm) in Buffer I run at 200 Volts, 1000 pps, 1 MFD, for two hours. ( ) unwashed E coli t)105; ( - - - ) unwashed A. flos-aquae 1)105

J. R. Carlton and D. S. Iterson: Blue-Green Alga Ribosomes 45
Gradient Studies of Bacterial and Algal 30 S and 50 S Subunits
Subunit composition of ribosomes can be analyzed on sucrose gra- dients following washing with low magnesium buffer. Tissi~res et al. (1959) demonstrated with bacterial ribosomes that in the presence of 0.1 mM Mg +2, 70S and 100S particles would dissociate to 30S and 50S subunits.
When we washed bacterial and algal ribosomes twice with low magnesium buffer (0.1 mM Mg +2) and centrifuged them on 15--35~ sucrose gradients, typical profiles were observed (Figs. 5 and 6). Although bacterial subunits were better separated, in both bacterial and algal profiles the small subunits (30 S) and large subunits (50 S) were present in a 1:2 absorbanee ratio in agreement with Tissibres et al. (1959).
Comparison of Washed and Unwashed Bacterial and Algal P 105 Fractions
The gradient profiles and banding patterns of bacterial and blue- green algal P105 fractions were examined before and after washing in ribosome washing buffer (Stanley et al., 1966). The unwashed material was resuspended in Iqirenberg buffer which has a magnesium concentration of 10 raM. In the gradient and gel profiles of unwashed bacterial P105 fractions (Figs. 7 and 8) two peaks are observed. According to the work of Tissi~res et al. (1959) the slower moving peak, or dimer, should predominate at 10 mM magnesium. No difference in the gradient profile of E. coli P105 was observed after washing, though the banding pattern on gels was slightly altered (Fig. 9). The 100S particle was still the major peak but two 70 S bands were observed. Separation on gels is based on molecular weight and charge and the eleetrophoretieal gel technique is sensitive enough to detect subtle alterations in ribosomal particles (Dahlberg et al., 1969).
The gradient profile of unwashed Anabaena ]los-aquae 1)105 is similar to tha t of E. coli with a major dimer peak (Fig. 10). The shoulder slightly heavier than the 100S peak may represent a small polysome and can also be seen on the gel (Fig. 8) as a very faint, slow-moving band. The unwashed algal 1)105 fraction was dark green, most likely due to chloro- phyll and the accessory pigment phycocyanin. Chlorophyll does not migrate into the gel. 1)hyeoeyanin enters the gel and before staining appears as a blue band. When the algal 1)105 fraction was washed the gel and gradient profiles were different from the results observed with the washed bacterial preparation (Figs. 9 and 11). In contrast to the bacterial ribosomes, the washing procedure caused dissociation of most of the algal dimer peak with concomitant increase in the monomer peak. Gel analysis shows the appearance of what may be a second monomer peak.

46 J . R. Carlton and D. S. Herson:
.,/ _
.20
.15
0 0540
.10
.05
?OS
1 IOOS i ! I I ! !
7o~,, loo j -.~..~..,
l 2 3 4 5 6 /dlGRATIOH IH CM.
Fig. 9
.4 0 0260
IOOS
A i f ,LFLOS-AOUAE
m5 I I ,.~ UN
."1" I I I I
I 2 3 4 5 ML.
Fig. 10
Fig. 9. Densitogram of washed bacterial and algal P 105 fractions. 2.25~ composite acrytamide-agarose gel (6 ram) in Buffer I run at 200 volts, 1000 pps, 1 MFD, for
two hours. ( ) washed E. coli P105; ( - - - ) washed A./los-aquae 1)105
Fig. 10. Unwashed A./los-aquae P 105 layered on a linear 15--30% sucrose-Niren- berg buffer gradient and run for one hour at 48000 rpm in a SW 50.1 rotor
.25 70S f k
.ZO l ~ A. FLOS-AQUAE I | - P]OD I ! WASHED
.15 - I ~ I v .
I 1 I I 1 1 2 3 4 5
ME.
Fig. 11. Washed A. flos-aquae F 105 layered on a linear 15--30~ sucrose-Nirenberg buffer gradient and run for one hour at 48000 rpm in a SW 50.1 rotor

Blue-Green Alga Ribosomes 47
Discussion Our results confirm and extend previous studies on blue-green algal
ribosomes. Taylor and Storck used analytical ultracentrffugation to determine the sedimentation coefficient of ribosomes isolated from molds, yeast, bacteria and blue-green algae. They found mean values of 81.3S for molds and yeast and 68.4S for bacteria and blue-green algae. The bacteria and blue-green algae had previously been considered unique because of the procaryotic nature of their cells and these workers sug- gested that possession of the "70S" type ribosome be used to further define this group (Taylor and Storck, 1964).
Craig and Cart compared the sedimentation properties of Anabaena variabilis and E. eoli ribosomes using sucrose gradients and analytical ultracentrifugation (1968). They also found that the blue-green algal ribosomes were of the 70S type but noted differences in the stability of the particles in the presence of certain buffers. They detected a protein moiety released from the ribosomes in the presence of Tris-Mg buffer which resulted in a lowered sedimentation coefficient. This was not observed when the ribosomes were isolated in the presence of phosphate buffer.
We have compared the mobility of blue-green algal and bacterial ribosomes on sucrose gradients and polyacrylamide-agarose composite gels. The latter technique can be used to analyze intact ribosomes on the basis of their net negative charge. Dahlberg et al. (1969) used gel electro- phoresis to study E. eoli polysomes. The sensitivity of the gel technique was noted when more than one type of subunit was detected. Our results indicate tha t blue-green algal and bacterial ribosomes have similar properties on sucrose gradients and polyacrylamide-agarose composite gels, though the mobility of ribosomes from the two organisms was not identical. The different mobility in the gels and gra- clients may be due to a difference in molecular weight, compactness of the particle or charge. I t was not possible, in this study, to determine ff the difference was due to a loss of protein from the Anabaena ribosomes as was noted by Craig and Cart (1968).
We have demonstrated small subunits, large subunits, monomers and dimers as well as polysomes in blue-green algae. When washed with EDTA and run in a second dimension, gels of Anabaena/los-aquae mono- mers and dimers dissociated into small subunits and large subunits. Dissociation into small subunits and large subunits was also observed when an algal ribosomal pellet was twice washed in low magnesium buffer and run on a sucrose gradient. P105 fractions of bacteria and algae were isolated and analyzed on gradients and gels. Peaks of 70 S monomers and 100S dimers were present, with the 100S dimer predominating. This is expected in the presence of buffer containing 10 mM magnesium. When a bacterial P105 fraction was washed with buffer containing 0.5 M NH4§ ,

48 J. R. Carlton and D. S. Herson: Blue-Green Alga Ribosomes
the gradient profile was unaltered. On the gels two 70 S ribosome bands were observed, in addit ion to the major 100S peak. However , a difference between bacterial and algal ribosomes was noted when the Anabaena
P105 fraction was washed with r ibosomal washing buffer. On both gradients and gels the major algal r ibosomal peak shifted f rom the dimer to the monomer. I n addition, two monomer bands were observed on the gel.
As ribosome washing buffer is used to remove proteins f rom ribo- somal particles, including initiation factors, it is possible t ha t a protein or other factor which functions in stabilization of the dimer was part ial ly removed by the washing.
Acknowledgements. We thank Mrs. Linda Adams and Miss Valerie Uriah for their technical assistance.
This investigation was supported by the University of Delaware Research Foundation.
References Craig, I. W., Carr, N. G. : Ribosomes from the blue-green alga Anabaena variabilis.
Arch. Mikrobiol. 62, 167--177 (1968). Dahlberg, A. E., Dingman, C. W., Peacock, A. C. : Eleetrophoretic characterization
of bacterial polyribosomes in agarose-acrylamide composite gels. J. molec. Biol. 41, 139--147 (1969).
Godson, N., Sinsheimer, R. L. : Use of brij as a generM method to prepare poly- ribosomes from Escherichia coli. Biochim. biophys. Acta (Amst.) 149, 489--495 (1967).
Hjerten, S., Jerstedt, S., Tiselins, A.: Electrophoretie particle sieving in poly- acrylamide gels as applied to ribosomes. Analyt. Biochem. 11, 211--218 (1965).
Holm-Hansen, O. : Ecology, physiology, and biochemistry of blue-green algae. Ann. Rev. Microbiol. 22, 47--70 (1968).
Lewin, R.A.: Physiology and biochemistry of algae, 1st edition. New York: Academic Press 1962.
Nirenberg, M. W., Matthaei, J. H.: The dependence of cell-free protein synthesis in E. coli upon naturally occurring or synthetic polyribonucleotides. Proc. nat. Acad. Sci. (Wash.) 4:7, 1588--1602 (1961).
Schweiger, M., Herrlich, P., Zillig, W. : Group specificity in protein synthesis III . Hoppe-Seylers Z. physiol. Chem. 850, 775--783 (1969).
Stanley, W.M., Salas, M., Wahba, A.J., Ochoa, S.: Translation of the genetic message: Factors involved in the initiation of protein synthesis. Proc. nat. Acad. Sci. (Wash.) 56, 290--295 (1966).
Taylor, M. M., Storck, R. : Uniqueness of bacterial ribosomes. Proc. nat. Acad. Sei. (Wash.) 92, 958--965 (1964).
Tissi~res, A., Watson, J .D. : Ribonueleoprotein particles from Escherichia coti. Nature (Lond.) 182, 778--780 (1958).
Schlessinger, D., Hollingsworth, B.R.: Ribonucleoprotein particles from Escherichla coli. J. molec. Biol. 1, 221--233 (1959).
Diane S. Herson University of Delaware College of Arts and Science Dept. of Biological Sciences 117 Wolf Hall Newark, Delaware 19711, U.S.A.




















