
RbrA, a cyanobacterial rubrerythrin, functions as a FNR-dependent peroxidase in heterocysts in...
12
RbrA, a cyanobacterial rubrerythrin, functions as a FNR-dependent peroxidase in heterocysts in protection of nitrogenase from damage by hydrogen peroxide in Anabaena sp. PCC 7120 Weixing Zhao, 1,2† Zi Ye 1† and Jindong Zhao 1 * 1 State Key Laboratory of Protein and Plant Genetic Engineering, College of Life Sciences, Peking University, Beijing, 100871, China. 2 The National Center for Plant Genome Research, Beijing 100101, China. Summary The heterocyst is a specialized cell for nitrogen fixa- tion in some filamentous cyanobacteria. Here we report that a rubrerythrin (RbrA) from Anabaena sp. PCC 7120 functions as a peroxidase in heterocysts and plays an important role in protection of nitroge- nase. The electron donor for RbrA in H2O2 reduction is NADPH and the electron transfer from NADPH to RbrA depends on ferredoxin:NADP + oxidoreductase. A rbrA mutant (r27) grew much more slowly than the wild type under diazotrophic conditions. Its nitroge- nase activity measured in air was only 8% of that measured under anoxic conditions. Staining r27 filaments with 2,7-dichlorodihydrofluorescein diac- etate indicated that heterocysts had a higher H2O2 concentration than the vegetative cells. The expres- sion of rbrA was controlled by two promoters and the promoter for the smaller transcript was regu- lated by HetR. Spatial expression of rbrA was studied and the results showed that the transcrip- tion is localized predominantly in heterocysts. In a mutant lacking nifH and rbrA, the H2O2 concentration in heterocysts was lower than that in the vegetative cells, suggesting that NifH is involved in H2O2 gen- eration. Our results demonstrate that RbrA is a criti- cal enzyme for H2O2 decomposition and provide evidence that nitrogenase autoprotection is impor- tant in heterocysts. Introduction In oxygenic photosynthesis electrons are transferred from water through a series of electron carriers to ferredoxin. The reduced ferredoxin can further transfer electrons to NADP + through ferredoxin:NADP + oxidoreductase (FNR), forming the stable reductant NADPH. The development of oxygenic photosynthesis by cyanobacteria in early evolu- tion, which began the release of oxygen as a byproduct of photosynthetic electron transport, created a severe oxida- tive stress. Nitrogenase and many other biochemical reactions that require low potential electron carriers are strongly inhibited by oxygen. Moreover, partial reduction of oxygen by the strong reductants produced during pho- tosynthetic electron transport can lead to the formation of reactive oxygen species (ROS) that damage cellular com- ponents such as DNA, proteins and lipids (Imlay and Linn, 1988; Cadenas, 1989; Fridovich, 1995). All organisms living the presence of oxygen have enzymatic mecha- nisms that remove ROS. One such enzyme is superoxide dismutase (Sod), which removes superoxide anion (O 2 - ) in the following reaction: 2O2 - + 2H + → H2O2 + O2. Because H2O2 is also toxic, it is usually decomposed rapidly by catalase or peroxidase. Cyanobacteria are a group of Gram-negative bacteria with diverse morphologies, ranging from unicellular to true branching filaments with one or more differentiated cell types. In some filamentous cyanobacteria, specialized cells called heterocysts are formed for nitrogen fixation when the filaments are deprived of any usable source of combined nitrogen (Wolk et al., 1994; Meeks and Elhai, 2002; Golden and Yoon, 2003; Zhang et al., 2006). The differentiation from a vegetative cell to a heterocyst is a complex process and many genes are involved in regu- lation of heterocyst formation. Among the genes that are critical to the initiation of heterocyst differentiation, ntcA (Frias et al., 1994; Wei et al., 1994) and hetR (Buikema and Haselkorn, 1991) are of central importance. NtcA is a global nitrogen control regulator that belongs to the CRP (cAMP receptor proteins) superfamily (Herrero et al., 2004). HetR is a serine-type protease that binds DNA (Zhou et al., 1998a; Huang et al., 2004), and it functions as master switch of heterocyst differentiation (Black et al., Accepted 9 October, 2007. *For correspondence. E-mail jzhao@ pku.edu.cn; Tel. (+86) 10 6275 6421; Fax (+86) 10 6275 1526. † These authors contribute equally to this work. Molecular Microbiology (2007) 66(5), 1219–1230 doi:10.1111/j.1365-2958.2007.05994.x © 2007 The Authors Journal compilation © 2007 Blackwell Publishing Ltd
-
Upload
northumbria -
Category
Documents
-
view
1 -
download
0
Transcript of RbrA, a cyanobacterial rubrerythrin, functions as a FNR-dependent peroxidase in heterocysts in...

RbrA, a cyanobacterial rubrerythrin, functions as aFNR-dependent peroxidase in heterocysts inprotection of nitrogenase from damage by hydrogenperoxide in Anabaena sp. PCC 7120
Weixing Zhao,1,2† Zi Ye1† and Jindong Zhao1*1State Key Laboratory of Protein and Plant GeneticEngineering, College of Life Sciences, PekingUniversity, Beijing, 100871, China.2The National Center for Plant Genome Research,Beijing 100101, China.
Summary
The heterocyst is a specialized cell for nitrogen fixa-tion in some filamentous cyanobacteria. Here wereport that a rubrerythrin (RbrA) from Anabaena sp.PCC 7120 functions as a peroxidase in heterocystsand plays an important role in protection of nitroge-nase. The electron donor for RbrA in H2O2 reductionis NADPH and the electron transfer from NADPH toRbrA depends on ferredoxin:NADP+ oxidoreductase.A rbrA mutant (r27) grew much more slowly than thewild type under diazotrophic conditions. Its nitroge-nase activity measured in air was only 8% of thatmeasured under anoxic conditions. Staining r27filaments with 2�,7�-dichlorodihydrofluorescein diac-etate indicated that heterocysts had a higher H2O2
concentration than the vegetative cells. The expres-sion of rbrA was controlled by two promoters andthe promoter for the smaller transcript was regu-lated by HetR. Spatial expression of rbrA wasstudied and the results showed that the transcrip-tion is localized predominantly in heterocysts. In amutant lacking nifH and rbrA, the H2O2 concentrationin heterocysts was lower than that in the vegetativecells, suggesting that NifH is involved in H2O2 gen-eration. Our results demonstrate that RbrA is a criti-cal enzyme for H2O2 decomposition and provideevidence that nitrogenase autoprotection is impor-tant in heterocysts.
Introduction
In oxygenic photosynthesis electrons are transferred fromwater through a series of electron carriers to ferredoxin.The reduced ferredoxin can further transfer electrons toNADP+ through ferredoxin:NADP+ oxidoreductase (FNR),forming the stable reductant NADPH. The development ofoxygenic photosynthesis by cyanobacteria in early evolu-tion, which began the release of oxygen as a byproduct ofphotosynthetic electron transport, created a severe oxida-tive stress. Nitrogenase and many other biochemicalreactions that require low potential electron carriers arestrongly inhibited by oxygen. Moreover, partial reductionof oxygen by the strong reductants produced during pho-tosynthetic electron transport can lead to the formation ofreactive oxygen species (ROS) that damage cellular com-ponents such as DNA, proteins and lipids (Imlay and Linn,1988; Cadenas, 1989; Fridovich, 1995). All organismsliving the presence of oxygen have enzymatic mecha-nisms that remove ROS. One such enzyme is superoxidedismutase (Sod), which removes superoxide anion(O2
-) in the following reaction: 2O2- + 2H+ → H2O2 + O2.
Because H2O2 is also toxic, it is usually decomposedrapidly by catalase or peroxidase.
Cyanobacteria are a group of Gram-negative bacteriawith diverse morphologies, ranging from unicellular to truebranching filaments with one or more differentiated celltypes. In some filamentous cyanobacteria, specializedcells called heterocysts are formed for nitrogen fixationwhen the filaments are deprived of any usable source ofcombined nitrogen (Wolk et al., 1994; Meeks and Elhai,2002; Golden and Yoon, 2003; Zhang et al., 2006). Thedifferentiation from a vegetative cell to a heterocyst is acomplex process and many genes are involved in regu-lation of heterocyst formation. Among the genes that arecritical to the initiation of heterocyst differentiation, ntcA(Frias et al., 1994; Wei et al., 1994) and hetR (Buikemaand Haselkorn, 1991) are of central importance. NtcA is aglobal nitrogen control regulator that belongs to the CRP(cAMP receptor proteins) superfamily (Herrero et al.,2004). HetR is a serine-type protease that binds DNA(Zhou et al., 1998a; Huang et al., 2004), and it functionsas master switch of heterocyst differentiation (Black et al.,
Accepted 9 October, 2007. *For correspondence. E-mail [email protected]; Tel. (+86) 10 6275 6421; Fax (+86) 10 6275 1526.†These authors contribute equally to this work.
Molecular Microbiology (2007) 66(5), 1219–1230 doi:10.1111/j.1365-2958.2007.05994.x
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd

1993, Buikema and Haselkorn, 2001). Recent evidencesuggests that HetR and NtcA function together in theregulation of heterocyst differentiation (Muro-Pastor et al.,2002; Shi et al., 2006).
Several features of heterocysts are required for theprotection of nitrogenase against inactivation by molecu-lar oxygen (Fay, 1992; Wolk et al., 1994). Heterocystslack photosystem II and do not evolve oxygen. Hetero-cysts have a thick envelop that limits oxygen penetration(Walsby, 1985; 2007). The oxygen concentration in het-erocysts is reduced by respiration. When the heterocyst-specific terminal respiratory oxidase genes (cox2 andcox3) are deleted, Anabaena sp. PCC 7120 is unable tofix nitrogen (Valladares et al., 2003; 2007). Because ROScan be generated by photosystem I (PSI) and respiratoryelectron transport in heterocysts (Krieg and Hoffman,1986; Asada, 1999), Sod and other enzymes are neededfor their removal. A recent study (Zhao et al., 2007) of asodA mutant of Anabaena sp. PCC 7120 has demon-strated that a membrane-bound MnSod (Li et al., 2002) isrequired for protection of nitrogenase against superoxideanions in heterocysts. A possible mechanism for protec-tion of nitrogenase from oxygen damage is ‘autoprotec-tion’, as proposed by Thorneley and Ashby (1989). Inautoprotection of nitrogenase, which requires that themolar ratio of nifH-encoded nitrogenase reductase tooxygen be greater than 4, nitrogenase reductase reducesoxygen to H2O2 without being inactivated by oxygen. TheH2O2 produced in this reaction must be removed promptlyby catalase or peroxidase (Thorneley and Ashby, 1989).Although autoprotection may function in heterocysts(Bergman et al., 1997), experimental evidence is lacking.A crucial factor for autoprotection is the presence of anactive peroxidase or a catalase. No such enzyme has yetbeen reported in heterocysts.
The enzymes required for the removal of H2O2 in cyano-bacteria seem to vary by species. A catalase-peroxidase(KatG) is found in some unicellular cyanobacteria(Mutsuda et al., 1996; Smulevich et al., 2006) and dele-tion of katG in Synechocystis sp. PCC 6803 (Tichy andVermaas, 1999) led to diminished H2O2 decomposition.While no katG has been found in heterocyst-formingcyanobacteria, several thiol-dependent peroxidases havebeen reported from Anabaena sp. PCC 7120 (Cha et al.,2007; Latifi et al., 2007). The main function of thethiol-peroxidases appears to be protection from lipidperoxidation. A putative Mn-catalase gene (alr0998)exists in Anabaena sp. PCC 7120, although its function isnot yet known. At present, little information is available onhow heterocysts remove H2O2.
Rubrerythrin (Rbr) facilitates electron transfer to hydro-gen peroxide in some anaerobic organisms (Jenney et al.,1999; Kurtz, 2006). Rbr proteins contain two distinctivedomains: a rubredoxin domain that contains a Fe(Cys)4
centre and a diiron domain that is similar to the diironcentre of haemerythrin (Sieker et al., 2000; Iyer et al.,2005). Rbr is critical to the protection of anaerobic bacte-ria against ROS (Coulter et al., 1999; Coulter and Kurtz,2001; Lumppio et al., 2001; Sztukowska et al., 2002;Weinberg et al., 2004; Kawasaki et al., 2007). A putativerbr gene is present in Anabaena sp. PCC 7120 (Kanekoet al., 2001). Here we report that the rbr gene from Ana-baena sp. PCC 7120 encodes an active Rbr whichfunctions as a NADPH-dependent peroxidize. We alsodemonstrate that FNR is required for electron transferbetween NADPH and Rbr. Our results show that the Rbris required for the protection of nitrogenase from ROS inAnabaena sp. PCC 7120.
Results
Rubrerythrin from Anabaena sp. PCC 7120 is aFNR-dependent peroxidase
The protein encoded by alr1174 of Anabaena sp. PCC7120 has only a moderate similarity to Rbr from anaerobicmicroorganisms such as Desulfovibrio vulgaris, but theamino acid residues required for diiron centre formationand for the rubredoxin domain are both conserved (SeeFig. S1 and Fig. S2). We named this gene rbrA. To identifyits physiological electron donor, recombinant RbrAwas produced in Escherichia coli and then purified,which appeared as a band in sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE; Fig. 1A).The monomer form of RbrA had an apparent molecularmass of 32 kDa, 5 kDa larger than the mass calculatedfrom its amino acid sequence. The air-exposed RbrA waspurple in colour. The absorption spectrum of oxidizedRbrA had peaks at 378, 492 and 580 nm (Fig. 1B), whichis characteristic of all-iron Rbr. The absorption peaks dis-appeared when a strong reductant such as dithionite wasadded to the protein solution. Neither NADH nor FADH2
was capable of directly reducing RbrA (data not shown),nor was NADPH (Fig. 1C). Recombinant FNR from Ana-baena sp. PCC 7120 was overproduced in E. coli andpurified (Fig. S3). The addition of a catalytic amount(0.5 mM) of FNR with 25 mM NADPH led to a completereduction of RbrA (Fig. 1C), demonstrating that FNR iseffective in mediating electron transfer from NADPH toRbrA.
We investigated the oxidation of reduced RbrA. While aslow oxidation of RbrA occurred in an air-saturated solu-tion, oxidation was much more rapid in the presence of0.5 mM H2O2 (Fig. 1D), indicating that RbrA has a peroxi-dase activity. The Km value for H2O2 in its reductionby RbrA was 2.25 � 0.34 mM and the Vmax was183 � 8 mmol min-1. The kinetics of FNR-dependentreduction of H2O2 by RbrA was measured as the rate of
1220 W. Zhao, Z. Ye and J. Zhao
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1219–1230
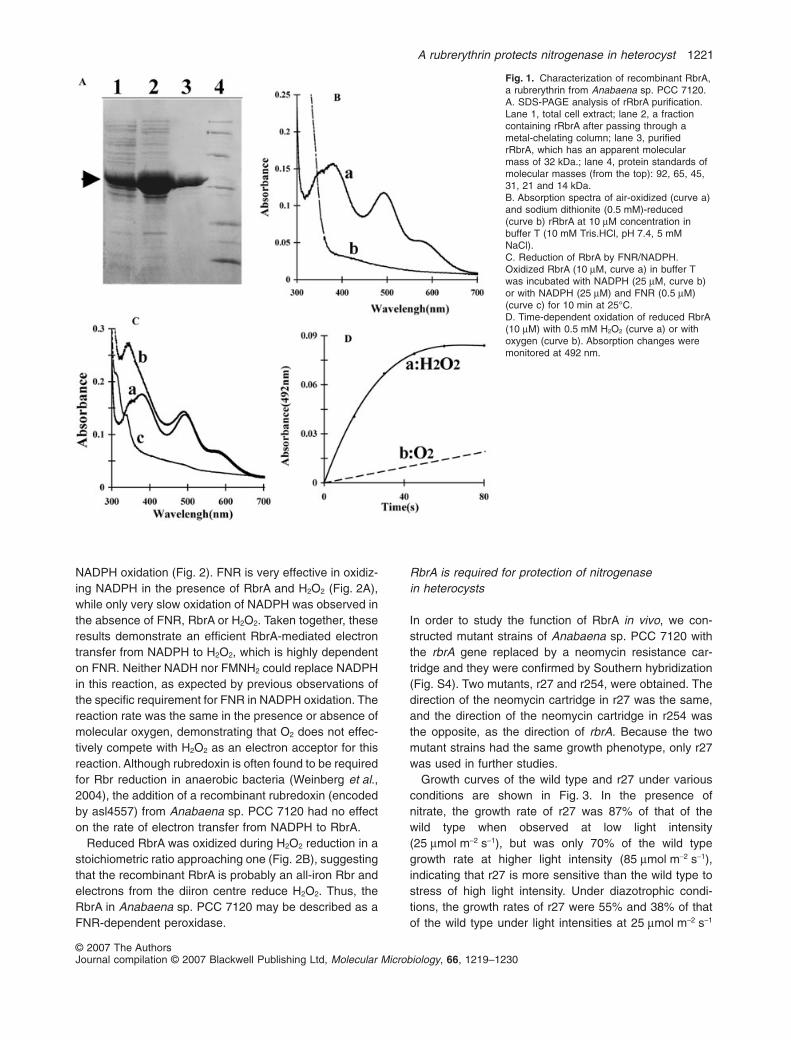
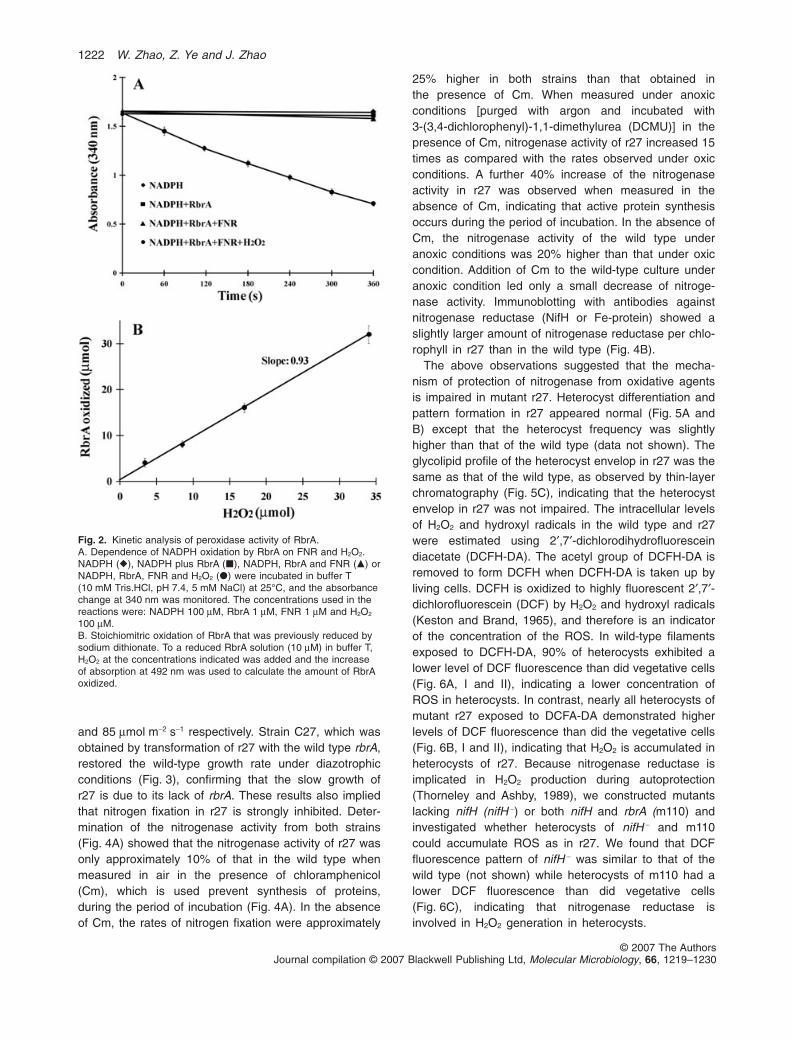
NADPH oxidation (Fig. 2). FNR is very effective in oxidiz-ing NADPH in the presence of RbrA and H2O2 (Fig. 2A),while only very slow oxidation of NADPH was observed inthe absence of FNR, RbrA or H2O2. Taken together, theseresults demonstrate an efficient RbrA-mediated electrontransfer from NADPH to H2O2, which is highly dependenton FNR. Neither NADH nor FMNH2 could replace NADPHin this reaction, as expected by previous observations ofthe specific requirement for FNR in NADPH oxidation. Thereaction rate was the same in the presence or absence ofmolecular oxygen, demonstrating that O2 does not effec-tively compete with H2O2 as an electron acceptor for thisreaction. Although rubredoxin is often found to be requiredfor Rbr reduction in anaerobic bacteria (Weinberg et al.,2004), the addition of a recombinant rubredoxin (encodedby asl4557) from Anabaena sp. PCC 7120 had no effecton the rate of electron transfer from NADPH to RbrA.
Reduced RbrA was oxidized during H2O2 reduction in astoichiometric ratio approaching one (Fig. 2B), suggestingthat the recombinant RbrA is probably an all-iron Rbr andelectrons from the diiron centre reduce H2O2. Thus, theRbrA in Anabaena sp. PCC 7120 may be described as aFNR-dependent peroxidase.
RbrA is required for protection of nitrogenasein heterocysts
In order to study the function of RbrA in vivo, we con-structed mutant strains of Anabaena sp. PCC 7120 withthe rbrA gene replaced by a neomycin resistance car-tridge and they were confirmed by Southern hybridization(Fig. S4). Two mutants, r27 and r254, were obtained. Thedirection of the neomycin cartridge in r27 was the same,and the direction of the neomycin cartridge in r254 wasthe opposite, as the direction of rbrA. Because the twomutant strains had the same growth phenotype, only r27was used in further studies.
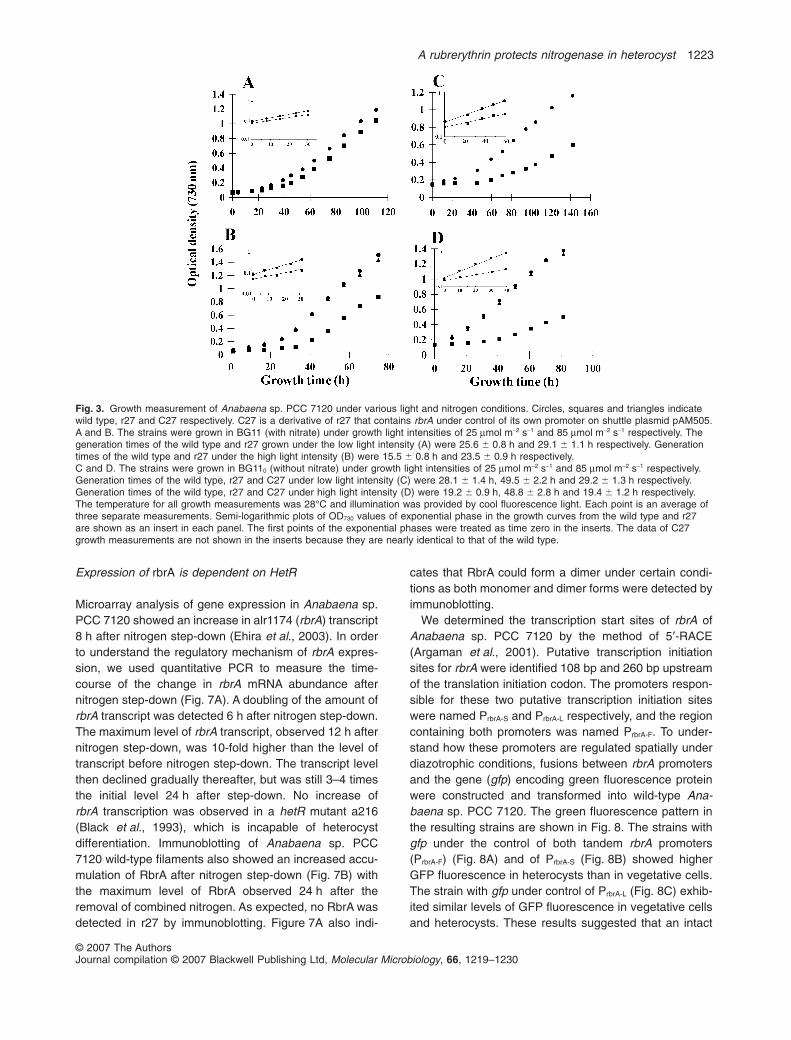
Growth curves of the wild type and r27 under variousconditions are shown in Fig. 3. In the presence ofnitrate, the growth rate of r27 was 87% of that of thewild type when observed at low light intensity(25 mmol m-2 s-1), but was only 70% of the wild typegrowth rate at higher light intensity (85 mmol m-2 s-1),indicating that r27 is more sensitive than the wild type tostress of high light intensity. Under diazotrophic condi-tions, the growth rates of r27 were 55% and 38% of thatof the wild type under light intensities at 25 mmol m-2 s-1
Fig. 1. Characterization of recombinant RbrA,a rubrerythrin from Anabaena sp. PCC 7120.A. SDS-PAGE analysis of rRbrA purification.Lane 1, total cell extract; lane 2, a fractioncontaining rRbrA after passing through ametal-chelating column; lane 3, purifiedrRbrA, which has an apparent molecularmass of 32 kDa.; lane 4, protein standards ofmolecular masses (from the top): 92, 65, 45,31, 21 and 14 kDa.B. Absorption spectra of air-oxidized (curve a)and sodium dithionite (0.5 mM)-reduced(curve b) rRbrA at 10 mM concentration inbuffer T (10 mM Tris.HCl, pH 7.4, 5 mMNaCl).C. Reduction of RbrA by FNR/NADPH.Oxidized RbrA (10 mM, curve a) in buffer Twas incubated with NADPH (25 mM, curve b)or with NADPH (25 mM) and FNR (0.5 mM)(curve c) for 10 min at 25°C.D. Time-dependent oxidation of reduced RbrA(10 mM) with 0.5 mM H2O2 (curve a) or withoxygen (curve b). Absorption changes weremonitored at 492 nm.
A rubrerythrin protects nitrogenase in heterocyst 1221
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1219–1230
and 85 mmol m-2 s-1 respectively. Strain C27, which wasobtained by transformation of r27 with the wild type rbrA,restored the wild-type growth rate under diazotrophicconditions (Fig. 3), confirming that the slow growth ofr27 is due to its lack of rbrA. These results also impliedthat nitrogen fixation in r27 is strongly inhibited. Deter-mination of the nitrogenase activity from both strains(Fig. 4A) showed that the nitrogenase activity of r27 wasonly approximately 10% of that in the wild type whenmeasured in air in the presence of chloramphenicol(Cm), which is used prevent synthesis of proteins,during the period of incubation (Fig. 4A). In the absenceof Cm, the rates of nitrogen fixation were approximately
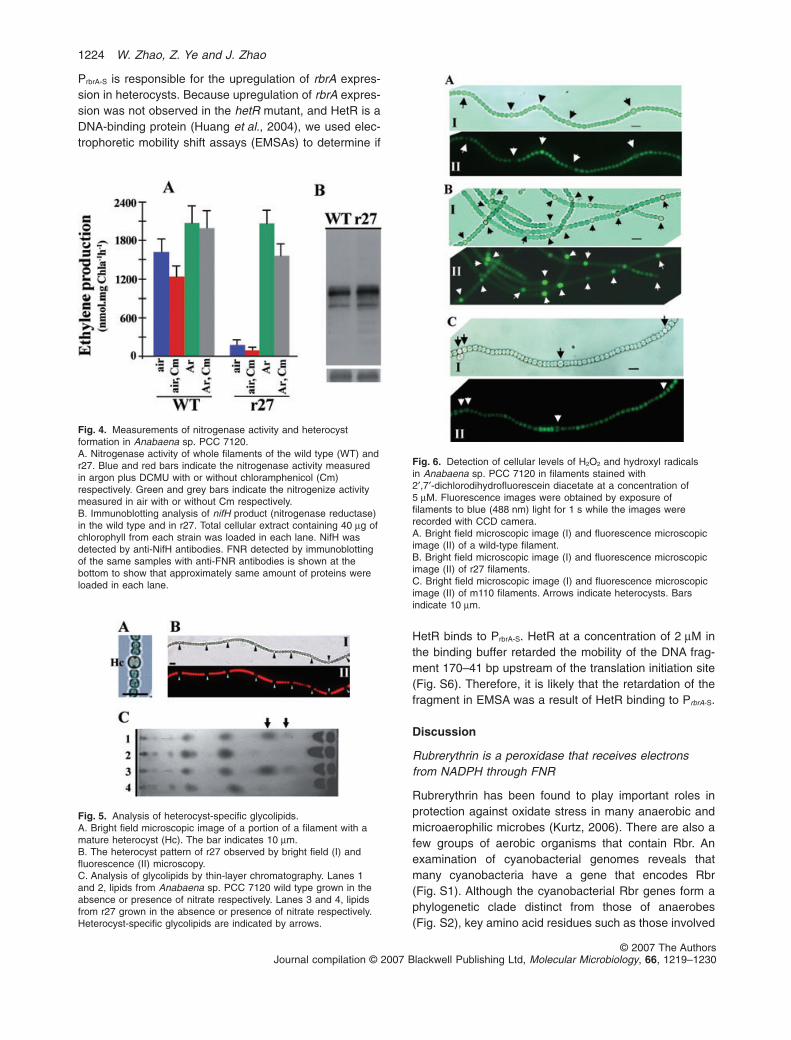
25% higher in both strains than that obtained inthe presence of Cm. When measured under anoxicconditions [purged with argon and incubated with3-(3,4-dichlorophenyl)-1,1-dimethylurea (DCMU)] in thepresence of Cm, nitrogenase activity of r27 increased 15times as compared with the rates observed under oxicconditions. A further 40% increase of the nitrogenaseactivity in r27 was observed when measured in theabsence of Cm, indicating that active protein synthesisoccurs during the period of incubation. In the absence ofCm, the nitrogenase activity of the wild type underanoxic conditions was 20% higher than that under oxiccondition. Addition of Cm to the wild-type culture underanoxic condition led only a small decrease of nitroge-nase activity. Immunoblotting with antibodies againstnitrogenase reductase (NifH or Fe-protein) showed aslightly larger amount of nitrogenase reductase per chlo-rophyll in r27 than in the wild type (Fig. 4B).
The above observations suggested that the mecha-nism of protection of nitrogenase from oxidative agentsis impaired in mutant r27. Heterocyst differentiation andpattern formation in r27 appeared normal (Fig. 5A andB) except that the heterocyst frequency was slightlyhigher than that of the wild type (data not shown). Theglycolipid profile of the heterocyst envelop in r27 was thesame as that of the wild type, as observed by thin-layerchromatography (Fig. 5C), indicating that the heterocystenvelop in r27 was not impaired. The intracellular levelsof H2O2 and hydroxyl radicals in the wild type and r27were estimated using 2′,7′-dichlorodihydrofluoresceindiacetate (DCFH-DA). The acetyl group of DCFH-DA isremoved to form DCFH when DCFH-DA is taken up byliving cells. DCFH is oxidized to highly fluorescent 2′,7′-dichlorofluorescein (DCF) by H2O2 and hydroxyl radicals(Keston and Brand, 1965), and therefore is an indicatorof the concentration of the ROS. In wild-type filamentsexposed to DCFH-DA, 90% of heterocysts exhibited alower level of DCF fluorescence than did vegetative cells(Fig. 6A, I and II), indicating a lower concentration ofROS in heterocysts. In contrast, nearly all heterocysts ofmutant r27 exposed to DCFA-DA demonstrated higherlevels of DCF fluorescence than did the vegetative cells(Fig. 6B, I and II), indicating that H2O2 is accumulated inheterocysts of r27. Because nitrogenase reductase isimplicated in H2O2 production during autoprotection(Thorneley and Ashby, 1989), we constructed mutantslacking nifH (nifH -) or both nifH and rbrA (m110) andinvestigated whether heterocysts of nifH - and m110could accumulate ROS as in r27. We found that DCFfluorescence pattern of nifH - was similar to that of thewild type (not shown) while heterocysts of m110 had alower DCF fluorescence than did vegetative cells(Fig. 6C), indicating that nitrogenase reductase isinvolved in H2O2 generation in heterocysts.
Fig. 2. Kinetic analysis of peroxidase activity of RbrA.A. Dependence of NADPH oxidation by RbrA on FNR and H2O2.NADPH (�), NADPH plus RbrA (�), NADPH, RbrA and FNR (�) orNADPH, RbrA, FNR and H2O2 (�) were incubated in buffer T(10 mM Tris.HCl, pH 7.4, 5 mM NaCl) at 25°C, and the absorbancechange at 340 nm was monitored. The concentrations used in thereactions were: NADPH 100 mM, RbrA 1 mM, FNR 1 mM and H2O2
100 mM.B. Stoichiomitric oxidation of RbrA that was previously reduced bysodium dithionate. To a reduced RbrA solution (10 mM) in buffer T,H2O2 at the concentrations indicated was added and the increaseof absorption at 492 nm was used to calculate the amount of RbrAoxidized.
1222 W. Zhao, Z. Ye and J. Zhao
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1219–1230
Expression of rbrA is dependent on HetR
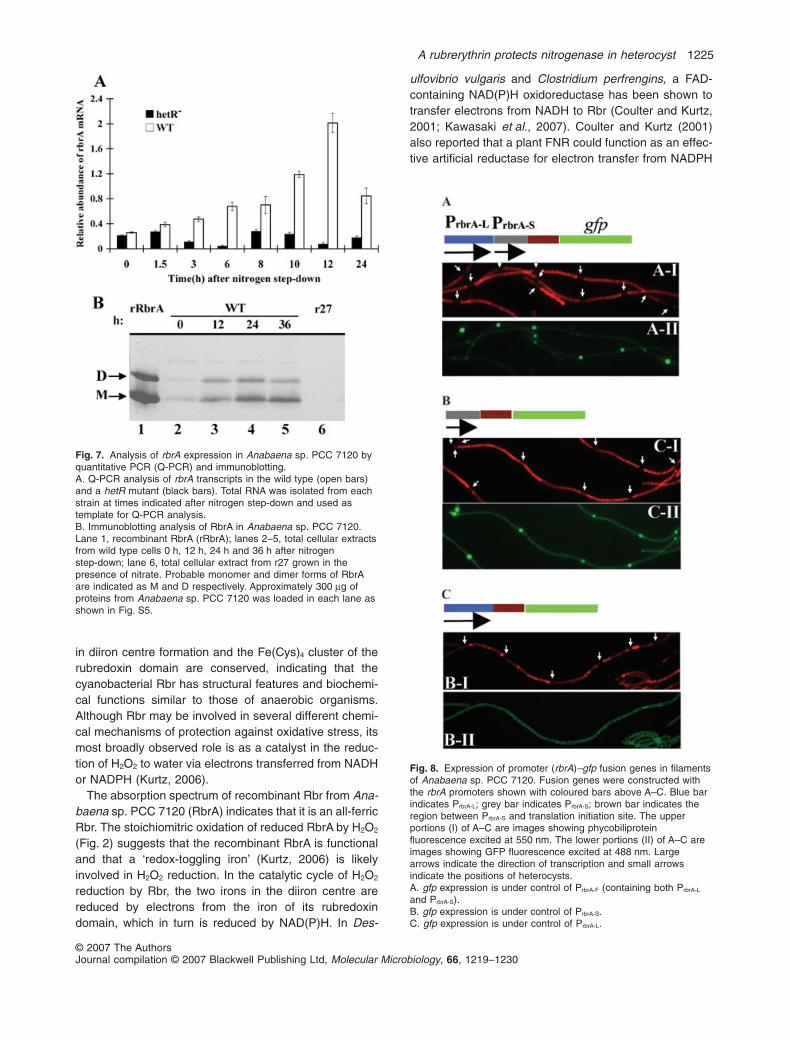
Microarray analysis of gene expression in Anabaena sp.PCC 7120 showed an increase in alr1174 (rbrA) transcript8 h after nitrogen step-down (Ehira et al., 2003). In orderto understand the regulatory mechanism of rbrA expres-sion, we used quantitative PCR to measure the time-course of the change in rbrA mRNA abundance afternitrogen step-down (Fig. 7A). A doubling of the amount ofrbrA transcript was detected 6 h after nitrogen step-down.The maximum level of rbrA transcript, observed 12 h afternitrogen step-down, was 10-fold higher than the level oftranscript before nitrogen step-down. The transcript levelthen declined gradually thereafter, but was still 3–4 timesthe initial level 24 h after step-down. No increase ofrbrA transcription was observed in a hetR mutant a216(Black et al., 1993), which is incapable of heterocystdifferentiation. Immunoblotting of Anabaena sp. PCC7120 wild-type filaments also showed an increased accu-mulation of RbrA after nitrogen step-down (Fig. 7B) withthe maximum level of RbrA observed 24 h after theremoval of combined nitrogen. As expected, no RbrA wasdetected in r27 by immunoblotting. Figure 7A also indi-
cates that RbrA could form a dimer under certain condi-tions as both monomer and dimer forms were detected byimmunoblotting.
We determined the transcription start sites of rbrA ofAnabaena sp. PCC 7120 by the method of 5′-RACE(Argaman et al., 2001). Putative transcription initiationsites for rbrA were identified 108 bp and 260 bp upstreamof the translation initiation codon. The promoters respon-sible for these two putative transcription initiation siteswere named PrbrA-S and PrbrA-L respectively, and the regioncontaining both promoters was named PrbrA-F. To under-stand how these promoters are regulated spatially underdiazotrophic conditions, fusions between rbrA promotersand the gene (gfp) encoding green fluorescence proteinwere constructed and transformed into wild-type Ana-baena sp. PCC 7120. The green fluorescence pattern inthe resulting strains are shown in Fig. 8. The strains withgfp under the control of both tandem rbrA promoters(PrbrA-F) (Fig. 8A) and of PrbrA-S (Fig. 8B) showed higherGFP fluorescence in heterocysts than in vegetative cells.The strain with gfp under control of PrbrA-L (Fig. 8C) exhib-ited similar levels of GFP fluorescence in vegetative cellsand heterocysts. These results suggested that an intact
Fig. 3. Growth measurement of Anabaena sp. PCC 7120 under various light and nitrogen conditions. Circles, squares and triangles indicatewild type, r27 and C27 respectively. C27 is a derivative of r27 that contains rbrA under control of its own promoter on shuttle plasmid pAM505.A and B. The strains were grown in BG11 (with nitrate) under growth light intensities of 25 mmol m-2 s-1 and 85 mmol m-2 s-1 respectively. Thegeneration times of the wild type and r27 grown under the low light intensity (A) were 25.6 � 0.8 h and 29.1 � 1.1 h respectively. Generationtimes of the wild type and r27 under the high light intensity (B) were 15.5 � 0.8 h and 23.5 � 0.9 h respectively.C and D. The strains were grown in BG110 (without nitrate) under growth light intensities of 25 mmol m-2 s-1 and 85 mmol m-2 s-1 respectively.Generation times of the wild type, r27 and C27 under low light intensity (C) were 28.1 � 1.4 h, 49.5 � 2.2 h and 29.2 � 1.3 h respectively.Generation times of the wild type, r27 and C27 under high light intensity (D) were 19.2 � 0.9 h, 48.8 � 2.8 h and 19.4 � 1.2 h respectively.The temperature for all growth measurements was 28°C and illumination was provided by cool fluorescence light. Each point is an average ofthree separate measurements. Semi-logarithmic plots of OD730 values of exponential phase in the growth curves from the wild type and r27are shown as an insert in each panel. The first points of the exponential phases were treated as time zero in the inserts. The data of C27growth measurements are not shown in the inserts because they are nearly identical to that of the wild type.
A rubrerythrin protects nitrogenase in heterocyst 1223
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1219–1230
PrbrA-S is responsible for the upregulation of rbrA expres-sion in heterocysts. Because upregulation of rbrA expres-sion was not observed in the hetR mutant, and HetR is aDNA-binding protein (Huang et al., 2004), we used elec-trophoretic mobility shift assays (EMSAs) to determine if
HetR binds to PrbrA-S. HetR at a concentration of 2 mM inthe binding buffer retarded the mobility of the DNA frag-ment 170–41 bp upstream of the translation initiation site(Fig. S6). Therefore, it is likely that the retardation of thefragment in EMSA was a result of HetR binding to PrbrA-S.
Discussion
Rubrerythrin is a peroxidase that receives electronsfrom NADPH through FNR
Rubrerythrin has been found to play important roles inprotection against oxidate stress in many anaerobic andmicroaerophilic microbes (Kurtz, 2006). There are also afew groups of aerobic organisms that contain Rbr. Anexamination of cyanobacterial genomes reveals thatmany cyanobacteria have a gene that encodes Rbr(Fig. S1). Although the cyanobacterial Rbr genes form aphylogenetic clade distinct from those of anaerobes(Fig. S2), key amino acid residues such as those involved
Fig. 4. Measurements of nitrogenase activity and heterocystformation in Anabaena sp. PCC 7120.A. Nitrogenase activity of whole filaments of the wild type (WT) andr27. Blue and red bars indicate the nitrogenase activity measuredin argon plus DCMU with or without chloramphenicol (Cm)respectively. Green and grey bars indicate the nitrogenize activitymeasured in air with or without Cm respectively.B. Immunoblotting analysis of nifH product (nitrogenase reductase)in the wild type and in r27. Total cellular extract containing 40 mg ofchlorophyll from each strain was loaded in each lane. NifH wasdetected by anti-NifH antibodies. FNR detected by immunoblottingof the same samples with anti-FNR antibodies is shown at thebottom to show that approximately same amount of proteins wereloaded in each lane.
Fig. 5. Analysis of heterocyst-specific glycolipids.A. Bright field microscopic image of a portion of a filament with amature heterocyst (Hc). The bar indicates 10 mm.B. The heterocyst pattern of r27 observed by bright field (I) andfluorescence (II) microscopy.C. Analysis of glycolipids by thin-layer chromatography. Lanes 1and 2, lipids from Anabaena sp. PCC 7120 wild type grown in theabsence or presence of nitrate respectively. Lanes 3 and 4, lipidsfrom r27 grown in the absence or presence of nitrate respectively.Heterocyst-specific glycolipids are indicated by arrows.
Fig. 6. Detection of cellular levels of H2O2 and hydroxyl radicalsin Anabaena sp. PCC 7120 in filaments stained with2′,7′-dichlorodihydrofluorescein diacetate at a concentration of5 mM. Fluorescence images were obtained by exposure offilaments to blue (488 nm) light for 1 s while the images wererecorded with CCD camera.A. Bright field microscopic image (I) and fluorescence microscopicimage (II) of a wild-type filament.B. Bright field microscopic image (I) and fluorescence microscopicimage (II) of r27 filaments.C. Bright field microscopic image (I) and fluorescence microscopicimage (II) of m110 filaments. Arrows indicate heterocysts. Barsindicate 10 mm.
1224 W. Zhao, Z. Ye and J. Zhao
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1219–1230
in diiron centre formation and the Fe(Cys)4 cluster of therubredoxin domain are conserved, indicating that thecyanobacterial Rbr has structural features and biochemi-cal functions similar to those of anaerobic organisms.Although Rbr may be involved in several different chemi-cal mechanisms of protection against oxidative stress, itsmost broadly observed role is as a catalyst in the reduc-tion of H2O2 to water via electrons transferred from NADHor NADPH (Kurtz, 2006).
The absorption spectrum of recombinant Rbr from Ana-baena sp. PCC 7120 (RbrA) indicates that it is an all-ferricRbr. The stoichiomitric oxidation of reduced RbrA by H2O2
(Fig. 2) suggests that the recombinant RbrA is functionaland that a ‘redox-toggling iron’ (Kurtz, 2006) is likelyinvolved in H2O2 reduction. In the catalytic cycle of H2O2
reduction by Rbr, the two irons in the diiron centre arereduced by electrons from the iron of its rubredoxindomain, which in turn is reduced by NAD(P)H. In Des-
ulfovibrio vulgaris and Clostridium perfrengins, a FAD-containing NAD(P)H oxidoreductase has been shown totransfer electrons from NADH to Rbr (Coulter and Kurtz,2001; Kawasaki et al., 2007). Coulter and Kurtz (2001)also reported that a plant FNR could function as an effec-tive artificial reductase for electron transfer from NADPH
Fig. 7. Analysis of rbrA expression in Anabaena sp. PCC 7120 byquantitative PCR (Q-PCR) and immunoblotting.A. Q-PCR analysis of rbrA transcripts in the wild type (open bars)and a hetR mutant (black bars). Total RNA was isolated from eachstrain at times indicated after nitrogen step-down and used astemplate for Q-PCR analysis.B. Immunoblotting analysis of RbrA in Anabaena sp. PCC 7120.Lane 1, recombinant RbrA (rRbrA); lanes 2–5, total cellular extractsfrom wild type cells 0 h, 12 h, 24 h and 36 h after nitrogenstep-down; lane 6, total cellular extract from r27 grown in thepresence of nitrate. Probable monomer and dimer forms of RbrAare indicated as M and D respectively. Approximately 300 mg ofproteins from Anabaena sp. PCC 7120 was loaded in each lane asshown in Fig. S5.
Fig. 8. Expression of promoter (rbrA)–gfp fusion genes in filamentsof Anabaena sp. PCC 7120. Fusion genes were constructed withthe rbrA promoters shown with coloured bars above A–C. Blue barindicates PrbrA-L; grey bar indicates PrbrA-S; brown bar indicates theregion between PrbrA-S and translation initiation site. The upperportions (I) of A–C are images showing phycobiliproteinfluorescence excited at 550 nm. The lower portions (II) of A–C areimages showing GFP fluorescence excited at 488 nm. Largearrows indicate the direction of transcription and small arrowsindicate the positions of heterocysts.A. gfp expression is under control of PrbrA-F (containing both PrbrA-L
and PrbrA-S).B. gfp expression is under control of PrbrA-S.C. gfp expression is under control of PrbrA-L.
A rubrerythrin protects nitrogenase in heterocyst 1225
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1219–1230

to rubredoxin/Rbr in D. vulgaris. FNR catalyses the trans-fer of electrons from reduced ferredoxin to NADP+ duringphotosynthetic electron transport. FNR can also catalysethe reverse reaction and may participate in cyclic electrontransport around PSI (Zhang et al., 2001). Based on thesestudies, we overproduced FNR of Anabaena sp. PCC7120 and found that it could use NADPH to reduce RbrA,which in turn reduces H2O2 to water. Therefore, FNR canfunction as a NADPH:RbrA oxidoreductase in addition toits roles in linear and cyclic electron transfer. Unlike inD. vulgaris and other anaerobic bacteria (Coulter andKurtz, 2001; Weinberg et al., 2004), RbrA reduction byFNR in Anabaena sp. PCC 7120 does not depend on arubredoxin, suggesting that FNR interacts with the rubre-doxin domain of RbrA and reduces it directly. Anabaenasp. PCC 7120 has two rubredoxins: RubA and a solublerubredoxin encoded by asl4557. PSI assembly and sta-bility require RubA, which is bound to thylakoid mem-branes in oxygenic photosynthetic organisms, includinghigher plants and cyanobacteria (Shen et al., 2002). Wehave constructed a mutant lacking asl4557, which has aphenotype different from that of r27 (our unpublisheddata), supporting the suggestion that a rubredoxin is notrequired for the NADPH-dependent peroxidase activity ofRbrA.
HetR regulates rbrA expression
Although a basal expression of rbrA is observed in fila-ments grown with nitrate, the transcript level increasednearly 10-fold 12 h after nitrogen step-down. The rbrAgene of Anabaena sp. PCC 7120 has two promoters(Fig. 8), resulting in two transcripts of different lengths.Only the small transcript was greatly enhanced after nitro-gen step-down (Ehira et al., 2003). Transcriptional fusionsbetween rbrA promoters and gfp showed a higher GFPfluorescence in heterocysts when gfp was under control ofPrbrA-S, indicating that PrbrA-S is responsible for the increasein rbrA transcription after nitrogen step-down. The fusionexperiments also suggest that the increase in expressionof the small rbrA transcript occurs mostly in heterocysts.The higher GFP fluorescence in heterocysts of strainscarrying PrbrA-S–gfp or PrbrA-F–gfp could be partly due to thefact that heterocysts contain less phycobiliprotein than dovegetative cells (Zhen et al., 2002) so less GFP fluores-cence emission is absorbed in heterocysts. However,comparison of heterocysts of the strain carrying PrbrA-L–gfpwith the strain carrying PrbrA-S–gfp shows that the tran-scription of rbrA is upregulated mostly in heterocysts. AsHetR is primarily located in heterocysts (Black et al.,1993; Zhou et al., 1998b) and a hetR mutant shows noincrease of rbrA transcription (Fig. 7), we suggest that theupregulation of rbrA transcription is dependent upon HetRand occurs in heterocysts. This suggestion is supported
by our preliminary data (Fig. S6), which indicates thatHetR binds the PrbrA-S region.
Roles of rubrerythrin in nitrogen fixation
A katG gene that encodes a catalase-peroxidase ispresent in various unicellular cyanobacteria, but is absentin many filamentous cyanobacteria, including Anabaenasp. PCC 7120. Recent reports showed that Anabaena sp.PCC 7120 has several thiol-peroxidases that might beimportant when the cells are under oxidative stressessuch as illumination at high light intensities in the pres-ence of methyl viologen (Cha et al., 2007; Latifi et al.,2007). However, they could play a more important role inprotection of lipid peroxidation than in the degradation ofH2O2 as their activity toward linoleic acid hydroperoxidewas several times higher than that toward H2O2. Only onethiol-peroxidase (PrxQ-A) was found in heterocysts (Chaet al., 2007). However, the rate of H2O2 reduction cataly-sed by PrxQ-A (Cha et al., 2007) is much lower than therate catalysed by RbrA (Fig. 2). The concentration ofPrxQ-A in heterocysts was the same as that in vegetativecells and the mutant lacking PrxQ-A (Latifi et al., 2007)has no apparent phenotype when grown under diaz-otrophic conditions (C.-C. Zhang, pers. comm.). Thefinding that RbrA is a NADPH-peroxidase and isexpressed most strongly in heterocysts suggests that theRbrA functions in H2O2 decomposition within heterocystsof Anabaena sp. PCC 7120.
Mutant r27 grows more slowly than the wild type underhigh light intensity in the presence of nitrate. Strongerinhibition of the growth of r27 is observed under diaz-otrophic conditions, suggesting that the nitrogen fixation isimpaired. In the absence of the protein synthesis inhibitorCm, the nitrogenase activity on a per-chlorophyll basis inr27 is similar to that of the wild type under anoxic condi-tions, but is only 10% of that in the wild type under oxicconditions (Fig. 4). When protein synthesis is prevented,r27 had a larger decrease of nitrogenase activity than thewild type, implying a higher protein turnover rate in het-erocysts of r27. These results indicate that the peroxidaseactivity of RbrA is critical to the protection of nitrogenasefrom H2O2 inhibition.
There are two probable sources of H2O2 generation inheterocysts. One is the reaction of superoxide dismuta-tion catalysed by Sod. The requirement of MnSod in het-erocysts for protection of nitrogenase from oxidativedamage (Zhao et al., 2007) implies that a mechanism isneeded for the removal of H2O2. A second source of H2O2
may be nitrogenase reductase (Fe protein). Nitrogenasereductase is capable of reducing molecular oxygen toH2O2 under microoxic conditions when the oxygen molarconcentration is one-fourth that of nitrogenase reductase(Thorneley and Ashby, 1989). The oxygen concentration
1226 W. Zhao, Z. Ye and J. Zhao
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1219–1230

in heterocysts is estimated to be 0.6 mM (Wolk et al.,1994) and the concentration of nitrogenase reductase inheterocysts is estimated much greater than four times theoxygen concentration (Bergman et al., 1997). Therefore,nitrogenase reductase could contribute substantially tooxygen uptake in heterocysts (Smith et al., 1985). Thisnotion is supported by the results of DCFH-DA staining,which indicate that heterocysts of r27 have a higherconcentration of H2O2 and/or hydroxyl radicals than doheterocysts of the wild type, and also have a higher con-centration of H2O2 than in vegetative cells of r27. In strainm110, in which both nifH and rbrA are deleted, the con-centration of H2O2 in heterocysts is lower than the veg-etative cells as indicated by DFC fluorescence. Theseresults suggest that nitrogenase reductase contributessubstantially to the formation of H2O2 in heterocysts andthat autoprotection is a mechanism for prevention of nitro-genase in heterocysts being inactivated. In the absence ofRbrA, the H2O2 generated through both Sod and nitroge-nase reductase is accumulated in heterocysts (as shownby the higher fluorescence from DCF in heterocystsdevoid of rbrA). The accumulated H2O2 may react with theirons (Fenton reactions) to produce hydroxyl radicals. Itcould also compete for electrons from nitrogenase reduc-tase, leading to a lower nitrogenase activity. The multiplepotential substrates for FNR in heterocysts provide amechanism for allocating electrons from NADPH, so thatmaximum activity of nitrogen fixation could be achieved.
Experimental procedures
Strains and culture conditions
The wild-type and mutant strains of Anabaena sp. PCC 7120were grown in BG11 medium (Rippka et al., 1979) with orwithout nitrate at 28°C under cool white fluorescent light atlight intensities indicated in text. Liquid cultures were bubbledgently with air plus 1% CO2 and their growth was determinedby measurement of optical density at 730 nm. E. coli strainswere grown in Luria–Bertani medium supplied with appropri-ate antibiotics. E. coli strain DH5a was used for all routinecloning and strain BL21 (DE3) was used for overproduction ofrecombinant proteins.
Gene inactivation and complementation
All enzymes were purchased from Promega (Beijing) andused according to instructions. To construct rbrA (alr1174)mutants, a DNA fragment containing rbrA was amplified bythe polymerase chain reaction (PCR) with primers P1 and P2(all primers used in PCR are listed in Table S1. All PCRproducts were sequenced to confirm their correctness), usingtotal genomic DNA as template. The PCR-generated frag-ment was cloned in pGEM-T vector (Promega) to generateplasmid pTv-rbrA. The plasmid was inversely amplified byPCR with primers P3 and P4. The recovered fragment was
ligated with a blunt-ended neo R cartridge encoding resistanceto neomycin. The resulting plasmid pRbrA-neo was digestedwith SacI and XhoI and the recovered fragment was thencloned into pRL277 (Elhai and Wolk, 1988) for transformationof Anabaena sp. PCC 7120 through conjugation. Segregationof the mutant rbrA was confirmed by Southern hybridizationas described by Huang et al. (2004). Total genomic DNA ofAnabaena sp. PCC 7120 was isolated with the EZNA plantDNA miniprep kit (Omega Bio-tek, Beijing), and was thendigested with HindIII. The HindIII fragments were separatedby a 1.0% agarose gel and then transferred onto a nitrocel-lulose paper before hybridization with a DNA probe that wassynthesized by random primer extension. The template usedfor probe synthesis was the coding region of the all1173 fromAnabaena sp. PCC 7120 amplified with primers P5 and P6.Two mutants of rbrA were obtained: strain r27 had the neo R
cartridge in the same direction as rbrA and strain r254 hadthe neo R cartridge in the opposite direction. For constructionof a nifH mutant in the wild type and r27 background, afragment containing the nifH gene was amplified with PCRusing primers PnifH5 and PnifH3. The amplified fragmentwas then was cloned in pGEM-T vector (Promega) to gener-ate plasmid pTv-nifH. The plasmid was inversely amplified byPCR with primers PnifHin5 and PnifHin3. The recovered frag-ment was ligated with a blunt-ended erythromycin cartridge.Deletion of nifH was performed by the same methods thatwere used for rbrA deletion (see above). Immunoblotting fordetection of RbrA in Anabaena sp. PCC 7120 was performedas follows. The proteins from total cell extracts of Anabaenasp. PCC 7120 filaments were separated by SDS-PAGEbefore they were transferred onto a polyvinylidene difluoridemembrane. Proteins that cross-reacted with the anti-RbrAantibodies were detected using secondary antibodies conju-gated with alkaline phosphatase (Promega) as described byZhou et al. (1998b). For complementation study of r27, therbrA gene plus 480-bp upstream was amplified by PCR usingprimers P8 and P9. The amplified gene was cloned intopAM505 (Yoon and Golden, 1998) and transformed into Ana-baena sp. PCC 7120 through triparental conjugation asdescribed by Zhao et al. (2005) to generate strain C27.
Protein overproduction and enzyme assays
Overproduction of recombinant RbrA was performed asfollows. A DNA fragment containing the rbrA gene of Ana-baena sp. PCC 7120 was amplified by PCR with primers P10and P11 and cloned into pET-22a. The plasmid was trans-formed into BL21 (DE3) for overexpression. The recombinantRbrA with a His-tag fused to its N-terminus was purified witha metal-chelating column and the His-tag was removed bythrombin. The recombinant RbrA was used to raise rabbitanti-RbrA antibodies. The petH gene encoding FNR of Ana-baena sp. PCC 7120 was amplified by PCR using primersP12 and P13. The procedure used for overproduction andpurification of FNR from Anabaena sp. PCC 7120 was thesame as that used for RbrA.
Peroxidase activity was measured according to Coulterand Kurtz (2001). The reduction or oxidation of RbrA wasmonitored by its absorption change at 492 nm. Chemicalreduction of RbrA was achieved by addition of dithionite andexcessive dithionite was removed by dialysis in the absence
A rubrerythrin protects nitrogenase in heterocyst 1227
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1219–1230

of oxygen. NADPH oxidation by FNR/RbrA was measured bythe absorbance change of NADPH at 340 nm. Nitrogenaseactivity was measured as ethylene production (Zhao et al.,2007) as follows. Anabaena sp. PCC 7120 strains weregrown in the absence of combined nitrogen for 48 h beforeharvesting for measurements. Filaments were suspended inBG110 medium (without nitrate) at an optical density of 0.6 at730 nm. Aliquots (8 ml) of the culture were then transferred to20 ml serum vials. The vials were purged with argon gascontaining 1% CO2 and no oxygen or else with gas containing79% argon, 1% CO2 and 20% oxygen, then were sealed. Onemillilitre of acetylene was added to each vial of the cultureand the vials were illuminated for 15 min before 100 mlvolumes of gas were withdrawn for measurements of ethyl-ene concentration by gas chromatography. When indicated,DCMU added to the medium at a final concentration of 10 mMto eliminate photosynthetic oxygen production (> 98% inhibi-tion of photosynthetic oxygen evolution). Cm was addedduring the illumination as indicated at a concentration of5 mg ml-1 to prevent protein synthesis.
The expression of rbrA and its localization
In attempt to determine rbrA transcription initiation sites inAnabaena sp. PCC 7120, 5′-RACE of rbrA transcripts wasperformed according to Argaman et al. (2001). Total RNAisolation was performed as described by Huang et al.(2004) and the reverse primer used in the 5′-Race was P7.The initiation sites of transcription were determined by DNAsequencing. The entire rbrA promoter region (containingboth promoters), the promoter that generates the large tran-script and the promoter that generates the small transcript,of rbrA were named PrbrA-F, PrbrA-L and PrbrA-S respectively.Quantitative PCR for analyses of the expression of rbrAwas performed according to Huang et al. (2004) and theprimers used for quantitative PCR of rbrA and 16 s rRNAwere Q1/Q2 and Q3/Q4. EMSAs were performed accordingto Huang et al. (2004). HetR at a concentration of 2 mM wasadded to the binding buffer and incubated for 15 min beforeelectrophoresis. The DNA fragment containing the promoterPrbrA-S was amplified with primer P15 and P17 and it waslabelled for EMSA. In a competitive assay, 10 times asmuch non-labelled PrbrA-S was also added to the bindingbuffer.
To determine which promoter is responsible for the upregu-lation of rbrA upon nitrogen step-down and the spatial patternof rbrA expression in Anabaena sp. PCC 7120, PrbrA-F andPrbrA-S were amplified with primers P14/P16 and P15/P16respectively. The amplified fragments were cloned intothe pGEM-T vector, generating pPrbrA-F and pPrbrA-Srespectively. The plasmid pPrbrA-F was inversely amplifiedby PCR to delete the region of PrbrA-S (-260 to -108) and theamplified fragment was ligated to generate plasmid pPrbrA-L.A gfp gene was amplified by PCR using primers P17 andP18 from plasmid pAM1951 (Yoon and Golden, 1998) astemplate. The fragment digested with SacI and BamHI wascloned into pAM505 to generate the plasmid pAM505–gfp.The plasmids containing rbrA promoters were digested withSalI and SacI before they were cloned into pAM505–gfp andtransformed into Anabaena sp. PCC 7120 through triparentalconjugation as described previously (Zhao et al., 2005).
Bright field and fluorescence microscopy were performedwith an Olympus BX51 microscope equipped with a CCDcamera as described by Huang et al. (2004). Observation offluorescence from photosynthetic pigments and GFP wereperformed using 466 nm excitation light according to Zhaoet al. (2005). The images of GFP fluorescence were storeddigitally and analysed with Image Pro Plus software (ApogeeAuburn, CA).
Other methods
Chlorophyll concentration was determined after extractioninto 80% acetone (MacKinney, 1941). The extraction ofheterocyst glycolipids and their analysis with thin-layerchromatography were performed according to Callahan andBuikema (2001). Staining the filaments of Anabaena sp. PCC7120 with DCFH-DA at a final concentration of 5 mM wascarried out in the dark according to He et al. (2002).DCFH-DA was purchased from Molecular Probe (Beijing)and used according to the manufacturer’s instructions. Thecyanobacterial filaments were exposed to 485 nm excitationand fluorescence images were recorded according to Zhaoet al. (2005).
Acknowledgements
We thank Dr Jerry Brand for his valuable suggestion andcomments on this work and critical reading of our manuscript.We also thank C. Dong and B. Hu for their skilful assistance.This research is supported by the National Science Founda-tion of China (30230040) and The Ministry of Science andTechnology of China (01CB108903).
References
Argaman, L., Hershberg, R., Vogel, J., Bejerano, G., Gerhart,E., Wagner, H., et al. (2001) Novel small RNA-encodinggenes in the intergenic regions of Escherichia coli. CurrBiol 11: 941–950.
Asada, K. (1999) The water–water cycle in chloroplasts:scavenging of active oxygens and dissipation of excessphotons. Annu Rev Plant Physiol Plant Mol Biol 50: 601–639.
Bergman, B., Gallon, J.R., Rai, A.N., and Stal, L.J. (1997) N2fixation by non-heterocystous cyanobacteria. FEMS Micro-biol Rev 19: 139–185.
Black, T.A., Cai, Y., and Wolk, C.P. (1993) Spatial expressionand autoregulation of hetR, a gene involved in the controlof heterocyst development in Anabaena. Mol Microbiol9: 77–84.
Buikema, W.J., and Haselkorn, R. (1991) Characterization ofa gene controlling heterocyst differentiation in the cyano-bacterium Anabaena sp. PCC 7120. Genes Dev 5: 321–330.
Buikema, W.J., and Haselkorn, R. (2001) Expression of theAnabaena hetR gene from a copper-regulated promoterleads to heterocyst differentiation under repressingconditions. Proc Natl Acad Sci USA 98: 2729–2734.
Cadenas, E. (1989) Biochemistry of oxygen toxicity. AnnuRev Biochem 58: 79–110.
1228 W. Zhao, Z. Ye and J. Zhao
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1219–1230

Callahan, S.M., and Buikema, W.J. (2001) The role of HetN inmaintenance of the heterocyst pattern in Anabaena sp.PCC 7120. Mol Microbiol 40: 941–950.
Cha, M.K., Hong, S.K., and Kim, I.H. (2007) Four thiol per-oxidases contain a conserved GCT catalytic motif and actas a versatile array of lipid peroxidases in Anabaena sp.PCC 7120. Free Radic Biol Med 42: 1736–1748.
Coulter, E.D., and Kurtz, D.M., Jr (2001) A role for rubredoxinin oxidative stress protection in Desulfovibrio vulgaris:catalytic electron transfer to rubrerythrin and two-ironsuperoxide reductase. Arch Biochem Biophys 394: 76–86.
Coulter, E.D., Shenvi, N.V., and Kurtz, D.M., Jr (1999) NADHperoxidase activity of rubrerythrin. Biochem Biophys ResCommun 255: 317–323.
Ehira, S., Ohmori, M., and Sato, N. (2003) Genome-wideexpression analysis of the responses to nitrogen depriva-tion in the heterocyst-forming cyanobacterium Anabaenasp. strain PCC 7120. DNA Res 10: 97–113.
Elhai, J., and Wolk, C.P. (1988) Conjugal transfer of DNA tocyanobacteria. Methods Enzymol 167: 747–754.
Fay, P. (1992) Oxygen relations of nitrogen fixation inCyanobacteria. Microbiol Rev 56: 340–373.
Frias, J.E., Flores, E., and Herrero, A. (1994) Requirement ofthe regulatory protein NtcA for the expression of nitrogenassimilation and heterocyst development genes in thecyanobacterium Anabaena sp. PCC 7120. Mol Microbiol14: 823–832.
Fridovich, I. (1995) Superoxide radical and superoxidedismutases. Annu Rev Biochem 64: 97–112.
Golden, J.W., and Yoon, H.S. (2003) Heterocyst develop-ment in Anabaena. Curr Opin Microbiol 6: 557–563.
He, Y., Klisch, M., and Hader, D. (2002) Adaptation of cyano-bacteria to UV-B stress correlated with oxidative stress andoxidative damage. Photochem Photobiol 76: 188–196.
Herrero, A., Muro-Pastor, A.M., Valladares, A., and Flores, E.(2004) Cellular differentiation and the NtcA transcriptionfactor in filamentous cyanobacteria. FEMS Microbiol Rev28: 469–487.
Huang, X., Dong, Y., and Zhao, J. (2004) HetR homodimer isa DNA-binding protein required for heterocyst differentia-tion, and the DNA-binding activity is inhibited by PatS. ProcNatl Acad Sci USA 101: 4848–4853.
Imlay, J.A., and Linn, S. (1988) DNA damage and oxygenradical toxicity. Science 240: 1302–1309.
Iyer, R.B., Silaghi-Dumitrescu, R., Kurtz, D.M., Jr, and Lan-zilotta, W.N. (2005) High-resolution crystal structures ofDesulfovibrio vulgaris (Hildenborough) nigerythrin: facile,redox-dependent iron movement, domain interface vari-ability, and peroxidase activity in the rubrerythrins. J BiolInorg Chem 10: 407–416.
Jenney, F.E., Jr, Verhagen, M.F., Cui, X., and Adams, M.W.(1999) Anaerobic microbes: oxygen detoxification withoutsuperoxide dismutase. Science 286: 306–309.
Kaneko, T., Nakamura, Y., Wolk, C.P., Kuritz, T., Sasamoto,S., Watanabe, A., et al. (2001) Complete genomicsequence of the filamentous nitrogen-fixing cyanobacte-rium Anabaena sp. strain PCC 7120. DNA Res 8: 205–213;227–253.
Kawasaki, S., Ono, M., Watamura, Y., Sakai, Y., Satoh, Y.,Arai, T., et al. (2007) An O2-inducible rubrerythrin-likeprotein, rubperoxin, is functional as a H2O2 reductase in an
obligatory anaerobe Clostridium acetobutylicum. FEBSLett 581: 2460–2464.
Keston, A.S., and Brand R. (1965) The fluorometric analysisof ultramicro quantities of hydrogen peroxide. AnalBiochem 11: 6–9.
Krieg, N.R., and Hoffman, P.S. (1986) Microaerophily andoxygen toxicity. Ann Rev Microbiol 40: 107–130.
Kurtz, D.M., Jr (2006) Avoiding high-valent iron intermedi-ates: superoxide reductase and rubrerythrin. J InorgBiochem 100: 679–693.
Latifi, A., Ruiz, M., Jeanjean, R., and Zhang, C.C. (2007)PrxQ-A, a member of the peroxiredoxin Q family, plays amajor role in defense against oxidative stress in the cyano-bacterium Anabaena sp. strain PCC 7120. Free Radic BiolMed 42: 424–431.
Li, T., Huang, X., Zhou, R., Liu, Y., Li, B., Nomura, C., andZhao, J. (2002) Differential expression and localization ofMn and Fe superoxide dismutases in the heterocystouscyanobacterium Anabaena sp. strain PCC 7120.J Bacteriol 184: 5096–5103.
Lumppio, H.L., Shenvi, N.V., Summers, A.O., Voordouw, G.,and Kurtz, D.M., Jr (2001) Rubrerythrin and rubredoxinoxidoreductase in Desulfovibrio vulgaris: a novel oxidativestress protection system. J Bacteriol 183: 101–108.
MacKinney, G. (1941) Absorption of light by chlorophyllsolutions. J Biol Chem 140: 315–322.
Meeks, J.C., and Elhai, J. (2002) Regulation of cellular dif-ferentiation in filamentous cyanobacteria in free-living andplant-associated symbiotic growth states. Microbiol MolBiol Rev 66: 94–121.
Muro-Pastor, A.M., Valladares, A., Flores, E., and Herrero, A.(2002) Mutual dependence of the expression of the celldifferentiation regulatory protein HetR and the globalnitrogen regulator NtcA during heterocyst development.Mol Microbiol 44: 1377–1385.
Mutsuda, M., Ishikawa, T., Takeda, T., and Shigeoka, S.(1996) The catalase-peroxidase of Synechococcus PCC7942: purification, nucleotide sequence analysis andexpression in Escherichia coli. Biochem J 316: 251–257.
Rippka, R., Deruelles, J., Waterbury, J.B., Herdman, M., andStanier, R.Y. (1979) Generic assignments, strain historiesand properties of pure cultures of cyanobacteria. J GenMicrobiol 111: 1–61.
Shen, G., Zhao, J., Reimer, S.K., Antonkine, M.L., Cai, Q.,Weiland, S.M., et al. (2002) Assembly of photosystem I. I.Inactivation of the rubA gene encoding a membrane-associated rubredoxin in the cyanobacterium Synechococ-cus sp. PCC 7002 causes a loss of photosystem I activity.J Biol Chem 277: 20343–20354.
Shi, Y., Zhao, W., Zhang, W., Ye, Z., and Zhao, J. (2006)Regulation of intracellular free calcium concentrationduring heterocyst differentiation by HetR and NtcA in Ana-baena sp. PCC 7120. Proc Natl Acad Sci USA 103: 11334–11339.
Sieker, L.C., Holmes, M., Le Trong, I., Turley, S., Liu, M.Y.,LeGall, J., and Stenkamp, R.E. (2000) The 1.9 A crystalstructure of the ‘as isolated’ rubrerythrin from Desulfovibriovulgaris: some surprising results. J Biol Inorg Chem 5:505–513.
Smith, R.L., Kumar, D., Zhang, X.K., Tabita, F.R., and VanBaalen, C. (1985) H2, N2, and O2 metabolism by isolated
A rubrerythrin protects nitrogenase in heterocyst 1229
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1219–1230

heterocysts from Anabaena sp. strain CA. J Bacteriol 162:565–570.
Smulevich, G., Jakopitsch, C., Droghetti, E., and Obinger, C.(2006) Probing the structure and bifunctionality of catalase-peroxidase (KatG). J Inorg Biochem 100: 568–585.
Sztukowska, M., Bugno, M., Potempa, J., Travis, J., andKurtz, D.M., Jr (2002) Role of rubrerythrin in the oxidativestress response of Porphyromonas gingivalis. Mol Micro-biol 44: 479–488.
Thorneley, R.N., and Ashby, G.A. (1989) Oxidation of nitro-genase iron protein by dioxygen without inactivation couldcontribute to high respiration rates of Azotobacter speciesand facilitate nitrogen fixation in other aerobicenvironments. Biochem J 261: 181–187.
Tichy, M., and Vermaas, W. (1999) In vivo role of catalase-peroxidase in Synechocystis sp. strain PCC 6803.J Bacteriol 181: 1875–1882.
Valladares, A., Herrero, A., Pils, D., Schmetterer, G., andFlores, E. (2003) Cytochrome c oxidase genes required fornitrogenase activity and diazotrophic growth in Anabaenasp. PCC 7120. Mol Microbiol 47: 1239–1249.
Valladares, A., Maldener, I., Muro-Pastor, A.M., Flores, E.,and Herrero, A. (2007) Heterocyst development anddiazotrophic metabolism in terminal respiratory oxidasemutants of the cyanobacterium Anabaena sp. strain PCC7120. J Bacteriol 189: 4425–4430.
Walsby, A.E. (1985) The permeability of heterocysts to thegases nitrogen and oxygen. Proc R Soc Lond B Biol Sci226: 345–367.
Walsby, A.E. (2007) Cyanobacterial heterocysts: terminalpores proposed as sites of gas exchange. Trends Microb15: 340–349.
Wei, T.F., Ramasubramanian, T.S., and Golden, J.W. (1994)Anabaena sp. strain PCC 7120 ntcA gene required forgrowth on nitrate and heterocyst development. J Bacteriol176: 4473–4482.
Weinberg, M.V., Jenney, F.E., Jr, Cui, X., and Adams, M.W.(2004) Rubrerythrin from the hyperthermophilic archaeonPyrococcus furiosus is a rubredoxin-dependent, iron-containing peroxidase. J Bacteriol 186: 7888–7895.
Wolk, C.P., Ernst, A., and Elhai, J. (1994) Heterocyst metabo-lism and development. In The Molecular Biology ofCyanobacteria. Bryant, D.A. (ed.). Dordrecht: Kluwer Aca-demic Press, pp. 769–823.
Yoon, H.S., and Golden, J.W. (1998) Heterocyst pattern for-
mation controlled by a diffusible peptide. Science 282:935–938.
Zhang, C.C., Laurent, S., Sakr, S., Peng, L., and Bedu, S.(2006) Heterocyst differentiation and pattern formation incyanobacteria: a chorus of signals. Mol Microbiol 59: 367–375.
Zhang, H., Whitelegge, J.P., and Cramer, W.A. (2001) Ferre-doxin: NADP+ oxidoreductase is a subunit of the chloro-plast cytochromeb6f complex. J Biol Chem 276: 38159–38165.
Zhao, W., Guo, Q., and Zhao, J. (2007) A membrane-associated Mn-superoxide dismutase protects thephotosynthetic apparatus and nitrogenase from oxidativedamage in the Cyanobacterium Anabaena sp. PCC 7120.Plant Cell Physiol 48: 563–572.
Zhao, Y., Shi, Y., Zhao, W., Huang, X., Wang, D., Brown, N.,et al. (2005) CcbP, a calcium-binding protein from Ana-baena sp. PCC 7120, provides evidence that calcium ionsregulate heterocyst differentiation. Proc Natl Acad Sci USA102: 5744–5748.
Zhen, L., Huang, X., Huang, B., Xie, J., Zhao, J., and Zhao,X. (2002) Fluorescence emission and absorption spectrumof single Anabaena sp. PCC 7120 cells. PhotochemPhotobiol 76: 310–313.
Zhou, R., Wei, X., Jiang, N., Li, H., Dong, Y., Hsi, K.L., andZhao, J. (1998a) Evidence that HetR protein is an unusualserine-type protease. Proc Natl Acad Sci USA 95: 4959–4963.
Zhou, R., Cao, Z., and Zhao, J. (1998b) Characterization ofHetR protein turnover in Anabaena sp. PCC 7120. ArchMicrobiol 169: 417–423.
Supplementary material
This material is available as part of the online article from:http://www.blackwell-synergy.com/doi/abs/10.1111/j.1365-2958.2007.05994.x(This link will take you to the article abstract).
Please note: Blackwell Publishing is not responsible for thecontent or functionality of any supplementary materials sup-plied by the authors. Any queries (other than missing mate-rial) should be directed to the corresponding author for thearticle.
1230 W. Zhao, Z. Ye and J. Zhao
© 2007 The AuthorsJournal compilation © 2007 Blackwell Publishing Ltd, Molecular Microbiology, 66, 1219–1230




















