
FurA is the master regulator of iron homeostasis and modulates the expression of tetrapyrrole...
13
FurA is the master regulator of iron homeostasis and modulates the expression of tetrapyrrole biosynthesis genes in Anabaena sp. PCC 7120 Andrés González, 1,2 M. Teresa Bes, 1,2 Ana Valladares, 3 M. Luisa Peleato 1,2 and María F. Fillat 1,2 * 1 Departamento de Bioquímica y Biología Molecular y Celular, Universidad de Zaragoza, 50009 Zaragoza, Spain. 2 Instituto de Biocomputación y Física de Sistemas Complejos (BIFI), Universidad de Zaragoza, 50009 Zaragoza, Spain. 3 Instituto de Bioquímica Vegetal y Fotosíntesis, Centro de Investigaciones Científicas Isla de la Cartuja, CSIC-Universidad de Sevilla, E-41092 Seville, Spain. Summary Knowledge on the regulatory mechanisms control- ling iron homeostasis in cyanobacteria is limited. In Anabaena sp. PCC 7120, the ferric uptake regulator FurA is a constitutive and essential protein whose expression is induced under iron deprivation. Our previous analyses have shown that this protein acts as a global transcriptional regulator, controlling the expression of several genes belonging to different functional categories, including schT, a gene coding for a TonB-dependent schizokinen transporter. In the present study we analysed the impact of FurA overexpression and iron availability on the transcrip- tional modulation of a broad range of Anabaena iron uptake, transport, storage and cellular iron utilization mechanisms, including enzymes involved in sidero- phore biosynthesis, TonB-dependent siderophore outer membrane transporters, siderophore peri- plasmic binding proteins, ABC inner membrane per- meases, ferritin Dps family proteins, and enzymes involved in tetrapyrrole biosynthesis. By combining reverse transcription-PCR analyses, electrophoretic mobility shift assays and DNase I footprinting experiments, we defined a variety of novel direct iron-dependent transcriptional targets of this metal- loregulator, including genes encoding at least five enzymes involved in the tetrapyrrole biosynthesis pathway. The results unravel the role of FurA as the master regulator of iron homeostasis in Anabaena sp. PCC 7120, providing new insights into the Fur regulons in cyanobacteria. Introduction The control of iron homeostasis in oxygenic photosyn- thetic organisms requires an effective balance between the use of iron as an essential nutrient and as a catalyser of reactive oxygen species (ROS) formation. Iron plays a critical role in photosynthetic electron transfer, serving as cofactor of every membrane-bound protein complex and other mobile electron carriers on the light reactions (Ferreira and Strauss, 1994). Thus, the demand for iron far exceeds that of non-photosynthetic organisms, being this micronutrient one of the main limiting factors of ocean primary productivity (Geider and La Roche, 1994). On the other hand, photosynthesis generates substantial amounts of hydrogen peroxide and superoxide which can interact with free iron producing hydroxyl radicals through Fenton chemistry (Shcolnick and Keren, 2006). In fact, both iron excess and iron starvation lead to significant increase in ROS and induce oxidative stress in cyanobac- teria (Latifi et al., 2005; 2009); therefore, iron homeostasis must be tightly regulated. In response to iron limitation, cyanobacteria synthesize efficient siderophore-based systems to improve the scav- enging of iron from the environment (Goldman et al., 1983; Jeanjean et al., 2008). Ferric-siderophore com- plexes are recognized, bound and translocated into the periplasm by TonB-dependent transporters in the outer membrane and then passed into the cytoplasm by ABC transporters in the cytoplasmic membrane (Nicolaisen et al., 2008; 2010; Mirus et al., 2009). Inter- nalized ferric iron is then reduced and dissociated from the siderophores (Andrews et al., 2003). Once inside the cell, the metal is mostly used as cofactor (e.g. haem and Fe-S clusters) or incorporated into iron storage proteins like ferritins and bacterioferritins, in preparation for long periods of iron scarcity in habitats where its supply is often intermittent (Keren et al., 2004; Shcolnick et al., 2007; 2009). Received 12 July, 2012; revised 6 September, 2012; accepted 9 September, 2012. *For correspondence. E-mail fi[email protected]; Tel. (+34) 976 76 1282; Fax (+34) 976 76 2123. Environmental Microbiology (2012) doi:10.1111/j.1462-2920.2012.02897.x © 2012 Society for Applied Microbiology and Blackwell Publishing Ltd
Transcript of FurA is the master regulator of iron homeostasis and modulates the expression of tetrapyrrole...

FurA is the master regulator of iron homeostasis andmodulates the expression of tetrapyrrole biosynthesisgenes in Anabaena sp. PCC 7120
Andrés González,1,2 M. Teresa Bes,1,2
Ana Valladares,3 M. Luisa Peleato1,2 andMaría F. Fillat1,2*1Departamento de Bioquímica y Biología Molecular yCelular, Universidad de Zaragoza, 50009 Zaragoza,Spain.2Instituto de Biocomputación y Física de SistemasComplejos (BIFI), Universidad de Zaragoza, 50009Zaragoza, Spain.3Instituto de Bioquímica Vegetal y Fotosíntesis, Centrode Investigaciones Científicas Isla de la Cartuja,CSIC-Universidad de Sevilla, E-41092 Seville, Spain.
Summary
Knowledge on the regulatory mechanisms control-ling iron homeostasis in cyanobacteria is limited. InAnabaena sp. PCC 7120, the ferric uptake regulatorFurA is a constitutive and essential protein whoseexpression is induced under iron deprivation. Ourprevious analyses have shown that this protein actsas a global transcriptional regulator, controlling theexpression of several genes belonging to differentfunctional categories, including schT, a gene codingfor a TonB-dependent schizokinen transporter. Inthe present study we analysed the impact of FurAoverexpression and iron availability on the transcrip-tional modulation of a broad range of Anabaena ironuptake, transport, storage and cellular iron utilizationmechanisms, including enzymes involved in sidero-phore biosynthesis, TonB-dependent siderophoreouter membrane transporters, siderophore peri-plasmic binding proteins, ABC inner membrane per-meases, ferritin Dps family proteins, and enzymesinvolved in tetrapyrrole biosynthesis. By combiningreverse transcription-PCR analyses, electrophoreticmobility shift assays and DNase I footprintingexperiments, we defined a variety of novel directiron-dependent transcriptional targets of this metal-loregulator, including genes encoding at least five
enzymes involved in the tetrapyrrole biosynthesispathway. The results unravel the role of FurA as themaster regulator of iron homeostasis in Anabaenasp. PCC 7120, providing new insights into the Furregulons in cyanobacteria.
Introduction
The control of iron homeostasis in oxygenic photosyn-thetic organisms requires an effective balance betweenthe use of iron as an essential nutrient and as a catalyserof reactive oxygen species (ROS) formation. Iron plays acritical role in photosynthetic electron transfer, servingas cofactor of every membrane-bound protein complexand other mobile electron carriers on the light reactions(Ferreira and Strauss, 1994). Thus, the demand for ironfar exceeds that of non-photosynthetic organisms, beingthis micronutrient one of the main limiting factors of oceanprimary productivity (Geider and La Roche, 1994). Onthe other hand, photosynthesis generates substantialamounts of hydrogen peroxide and superoxide which caninteract with free iron producing hydroxyl radicals throughFenton chemistry (Shcolnick and Keren, 2006). In fact,both iron excess and iron starvation lead to significantincrease in ROS and induce oxidative stress in cyanobac-teria (Latifi et al., 2005; 2009); therefore, iron homeostasismust be tightly regulated.
In response to iron limitation, cyanobacteria synthesizeefficient siderophore-based systems to improve the scav-enging of iron from the environment (Goldman et al.,1983; Jeanjean et al., 2008). Ferric-siderophore com-plexes are recognized, bound and translocated intothe periplasm by TonB-dependent transporters in theouter membrane and then passed into the cytoplasmby ABC transporters in the cytoplasmic membrane(Nicolaisen et al., 2008; 2010; Mirus et al., 2009). Inter-nalized ferric iron is then reduced and dissociated fromthe siderophores (Andrews et al., 2003). Once inside thecell, the metal is mostly used as cofactor (e.g. haem andFe-S clusters) or incorporated into iron storage proteinslike ferritins and bacterioferritins, in preparation for longperiods of iron scarcity in habitats where its supply is oftenintermittent (Keren et al., 2004; Shcolnick et al., 2007;2009).
Received 12 July, 2012; revised 6 September, 2012; accepted 9September, 2012. *For correspondence. E-mail [email protected];Tel. (+34) 976 76 1282; Fax (+34) 976 76 2123.
bs_bs_banner
Environmental Microbiology (2012) doi:10.1111/j.1462-2920.2012.02897.x
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd

Iron availability strongly influences biosynthesis ofhaem (Chauhan et al., 1997), an abundant and ubiquitousiron-containing cyclic tetrapyrrole involved in many meta-bolic processes including respiration and oxygen metabo-lism, but especially important in cyanobacteria as theessential intermediate in the synthesis of phycobilins(Brown et al., 1990). Enzymatic steps of haem metabo-lism are well characterized (Cornah et al., 2003; Hein-emann et al., 2008; Layer et al., 2010). Haem andchlorophyll share a common biosynthetic pathway from5-aminolevulininc acid (ALA) to protoporphyrin IX. Haemis synthesized from protoporphyrin IX by insertion of Fe2+
in a ferrochelatase-mediated reaction, whereas insertionof Mg2+ by magnesium chelatase is the first step of thechlorophyll branch. Haem is cleaved to biliverdin (BV) IXaby haem oxygenase as a degradation process in animals,but this reaction constitutes one more step of the bilinsbiosynthetic pathway in photosynthetic organisms. Asoccurs with iron, the accumulation of free haem as well assome of its biosynthetic intermediates is toxic by causeoxidative damage to cellular components (Vavilin andVermaas, 2002), therefore tetrapyrrole synthesis pathwayis tightly regulated by cell, and this regulation frequentlydepends of intracellular iron levels (Chauhan et al., 1997;Taketani et al., 2000).
Iron homeostasis is predominantly controlled in bacte-ria by a global transcriptional regulator known as Fur(ferric uptake regulator), which senses intracellular freeiron and modulates transcription of iron uptake andstorage machinery in response to iron availability (Escolaret al., 1999). It is assumed that Fur works as a transcrip-tional repressor using Fe2+ as co-repressor under iron-replete conditions, and causing de-repression in theabsence of Fe2+ under iron starvation (Andrews et al.,2003). However, transcription of several genes has beenshown to be directly or indirectly upregulated by Fur,involving a variety of mechanisms (Masse and Gottes-man, 2002; Delany et al., 2004). Fur proteins havebeen described in several cyanobacterial strains like Syn-echococcus sp. PCC 7942 (Ghassemian and Straus,1996), Microcystis aeruginosa PCC 7806 (Martin-Lunaet al., 2006a) and Anabaena sp. PCC 7120 (Hernándezet al., 2004b); although fur orthologues are extensivelyfound in almost so far annotated cyanobacterial genomesaccording to CyanoBase (http://genome.kazusa.or.jp/cyanobase). As occurs with Fur from heterotrophic bacte-ria, cyanobacterial Fur proteins appears to function asglobal transcriptional regulators, controlling the expres-sion of a plethora of genes including cyanotoxins (Martin-Luna et al., 2006b), iron-stress proteins (Ghassemian andStraus, 1996), photosynthesis and nitrogen metabolism(López-Gomollón et al., 2007; González et al., 2010), zinchomeostasis (Napolitano et al., 2012), among other tran-scriptional targets; but only FurA from Anabaena sp. PCC
7120 have been previously demonstrated to directlycontrol the expression of at least one iron homeostasisgene, the TonB-dependent schizokinen transporter SchT(González et al., 2010). Furthermore, a role as haem-sensor has been proposed for FurA based on the bio-chemical characterization of the FurA-haem complex(Pellicer et al., 2012), whose ability to bind DNA isseverely impaired (Hernández et al., 2004a).
FurA is a constitutive protein whose expressionincreases under iron deprivation (Hernández et al., 2002).As occurs with other cyanobacterial Fur homologues(Ghassemian and Straus, 1996), attempts to inactivatethe gene furA have resulted in only a partial segregationof the mutated chromosomes, suggesting an essentialrole for this protein under standard culture conditions(Hernández et al., 2006a). To partially overcome this limi-tation, we have previously obtained a strain of Anabaenasp. that overproduces active FurA by using a shuttlevector with the copper-responsive petE promoter fused toa furA gene copy. This approach has been successfullyused to identify several FurA direct targets (Gonzálezet al., 2010; 2011). In the present study we have analysedthe impact of FurA overexpression and iron availability onthe transcriptional regulation of a broad range of Ana-baena iron uptake, transport, storage and cellular ironutilization mechanisms; unravelling the role of FurA as themaster regulator of iron homeostasis, as well as providingfirst evidence that FurA directly controls the tetrapyrrolebiosynthesis pathway in Anabaena sp. PCC 7120.
Results
FurA overexpression blocks the induction of ironmetabolism genes under iron-restricted conditions
Our previous studies have indicated that the Anabaenametalloregulator FurA transcriptionally modulated theexpression of the outer membrane transporter forschizokinen, SchT (González et al., 2010) and the iron-stress-induced protein IsiB (Hernández et al., 2006b). Inorder to identify novel FurA targets involved in the controlof iron homeostasis in Anabaena sp. PCC 7120, we ana-lysed the influence of furA overexpression on the tran-scriptional pattern of nine different genes involved inseveral steps of iron uptake and assimilation, includingenzymes implicated in synthesis of siderophores, TonB-dependent outer membrane ferric-siderophore transport-ers, ABC transporters of plasmatic membrane, and ironstorage proteins. Changes in mRNA levels were deter-mined by semi-quantitative reverse transcription-PCR(RT-PCR). To obtain accurate determinations, eachmeasure was performed at the early exponential phase ofthe PCR of each gene. The rnpB housekeeping gene wasincluded in all RT-PCR analyses to ensure that equivalentamounts of total RNA were being used in all reactions.
2 A. González et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

As shown in Fig. 1 and Table 1, all of the genes involvedin iron metabolism analysed in the study resulted tran-scriptionally induced to a greater or lesser extent in theAnabaena wild-type strain exposed to iron deprivation. Asexpected, the overexpression of furA under iron-repleteconditions slightly induced the transcription of iron-responsive genes, a result previously observed with schTand other FurA direct targets (González et al., 2010;2011), probably as consequence of a sudden reduction of
the free iron pool in the cytosol by a ‘buffering’ action ofthe overexpressed FurA protein. However, the inducibleeffect of iron starvation observed in the wild-type strainresulted clearly affected by the overexpression of FurA inall genes analysed (Table 1). Thus, under a FurA overex-pression background, the amount of active FurA–Fe2+
complexes would be sufficient to over-repress the tran-scription of iron-responsive FurA targets, even after exog-enous iron limitation.
WT FurA+ WT FurA+WT FurA
+ - + -
rnpB
furA
Fe2+
hemA
hemL
WT FurA
+ - + -Fe2+
all4145
all1101
alr3242
hemB
hemC
hemD
hemEall2610
alr3240
all2618
all0389
hemE
hemF
hemK
hemH1
all2648
all2646
hemH2
ho1
ho2
Fig. 1. Semi-quantitative RT-PCR analysesshowing the impact of FurA overexpressionand iron availability on the transcriptionalpattern of several genes involved in ironmetabolism and tetrapyrrole biosynthesis.Total RNA from wild-type strain PCC 7120(WT) and furA overexpressing strainAG2770FurA (FurA+) were isolated from cellsgrown in standard BG-11 medium (+Fe2+) oriron deprived medium BG-11-Fe (-Fe2+).Housekeeping gene rnpB was used ascontrol. Determinations for each gene wereperformed in the early exponential phase ofPCR. Experiments were repeated at leasttwice with independent RNA extractions, andthe relevant portions of a representative gelare shown. Relative induction ratios areshown in Tables 1 and 2.
Table 1. Relative induction ratio of selected iron metabolism genes under each experimental condition, in relation to the wild-type strain PCC 7120grown under standard conditions (BG11 medium), as result of semi-quantitative RT-PCR analyses.a
Gene IDb Protein descriptionb WT(-Fe)/WT(+Fe) FurA+(+Fe)/WT(+Fe) FurA+
(-Fe)/WT(+Fe)
all1691 Ferric uptake regulator, FurA 4.06 � 0.16 32.11 � 1.27 32.63 � 1.34all4145 Ferritin, Dps family protein 3.45 � 0.02 1.65 � 0.01 0.95 � 0.04all1101 TonB-dependent ferrichrome iron receptor 6.94 � 0.08 1.93 � 0.18 2.00 � 0.13alr3242 TonB-dependent haem receptor 2.97 � 0.07 2.27 � 0.02 2.01 � 0.04all2610 TonB-dependent ferrichrome iron receptor 1.24 � 0.02 1.22 � 0.04 1.07 � 0.01alr3240 Ferrichrome ABC transporter, permease 1.80 � 0.03 1.15 � 0.01 1.07 � 0.01all2618 Iron (III) dicitrate-binding periplasmic protein 1.87 � 0.01 1.67 � 0.02 1.75 � 0.01all0389 Iron (III) ABC transporter 2.70 � 0.04 1.75 � 0.04 1.84 � 0.01all2648 Peptide synthetase 4.49 � 0.04 1.25 � 0.06 1.75 � 0.03all2646 Polyketide synthase type I 2.45 � 0.64 2.17 � 0.01 1.96 � 0.04
a. Signal assigned to each gene corresponded to the intensity of DNA band in the agarose gel stained with ethidium bromide, normalized to thesignal observed for housekeeping gene rnpB in each condition. Values are means of two independent determination � SD.b. Gene identification and protein description according to the cyanobacterial genomes database CyanoBase (http://genome.kazusa.or.jp/cyanobase).
Iron homeostasis in Anabaena sp. 3
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology
FurA is the master transcriptional regulator of ironmetabolism in Anabaena sp.
To determine whether the expression of genes previ-ously analysed by RT-PCR was directly regulated byFurA, we investigated the ability of recombinant FurA tobind in vitro the promoter regions of such genes byEMSA. In the case of enzymes involved in siderophorebiosynthesis, whose genes are arranged in a clusterof non-ribosomal peptide synthetases (NRPS) andpolyketide synthases (PKS) in Anabaena sp. PCC 7120(Jeanjean et al., 2008), we analysed the ability of FurAto specifically bind to different potential promoterslocated at different points in this nine-gene cluster,denoted by P1 to P4.
As showed in Fig. 2, the EMSA experiments demon-strated that FurA specifically recognized and bound invitro the promoter regions of all iron-responsive genesanalysed. In the case of the NRPS-PKS gene cluster, gelretardation assays showed that FurA bound with higheraffinity to the promoter region denoted as P1, locatedmost upstream at the cluster, in the promoter region ofgene all2649. In vitro bindings to P2, P3 and P4 alsooccurred, but with less affinity. Taking into account apositive result in both, RT-PCR and EMSA, we con-cluded that all of iron metabolism genes analysed weredirect transcriptional targets of FurA, indicating a broadregulatory role of FurA on iron uptake and assimilationmechanisms.
As other so far recognized FurA targets, the specificbinding of the metalloregulator to these novel targets wasstrongly dependent on the presence of divalent metalions and reducing conditions. Mn2+ ions are routinely usedin Fur binding experiments instead of Fe2+ ions becauseof their greater redox stability, while the same amountsof both divalent ions have similar effect on in vitro Fur
biological activity (Delany et al., 2001). Hence, the in vitroresults suggest an iron-dependent regulation of FurA onits transcriptional targets. It is noticeable that this iron-dependent affinity results clearly affected by oxidativesignals, confirming the previous recognized connectionbetween iron homeostasis and oxidative stress in thecyanobacterial cell.
DNase I protection footprinting assay on the P1 pro-moter of the NRPS-PKS gene cluster was carried out withpurified recombinant FurA using binding conditions similarto those employed in EMSA experiments (Fig. 3). Awindow of protection due to FurA binding was observedcovering an A/T rich 28 nucleotide (nt) region, spaced 86nt upstream to the translational start point of the all2649gene.
Iron deprivation transcriptionally induces enzymesinvolved in haem metabolism
Because the synthesis of haem results in one of themost relevant iron utilization route into the cell and itconstitutes an essential intermediate to phycobilin bio-synthesis in cyanobacteria, we analysed the effect ofiron deprivation on the transcriptional profiles of haemmetabolism enzymes in Anabaena sp. PCC 7120 bymeans of RT-PCR. As expected, iron deprivationresulted in upregulation of transcription of enzymesinvolved in haem degradation and subsequent releaseof iron, the haem oxygenases I (ho1) and II (ho2). Butcuriously, limitation of iron in the wild-type strain ofAnabaena sp. additionally triggered transcriptionalinduction of all haem biosynthesis enzymes (Fig. 1).However, the increase of transcript abundance of haemmetabolism enzymes observed in the cyanobacteriumafter iron limitation was not homogenous in all genes(Table 2). Hence, while appreciable increase in transcript
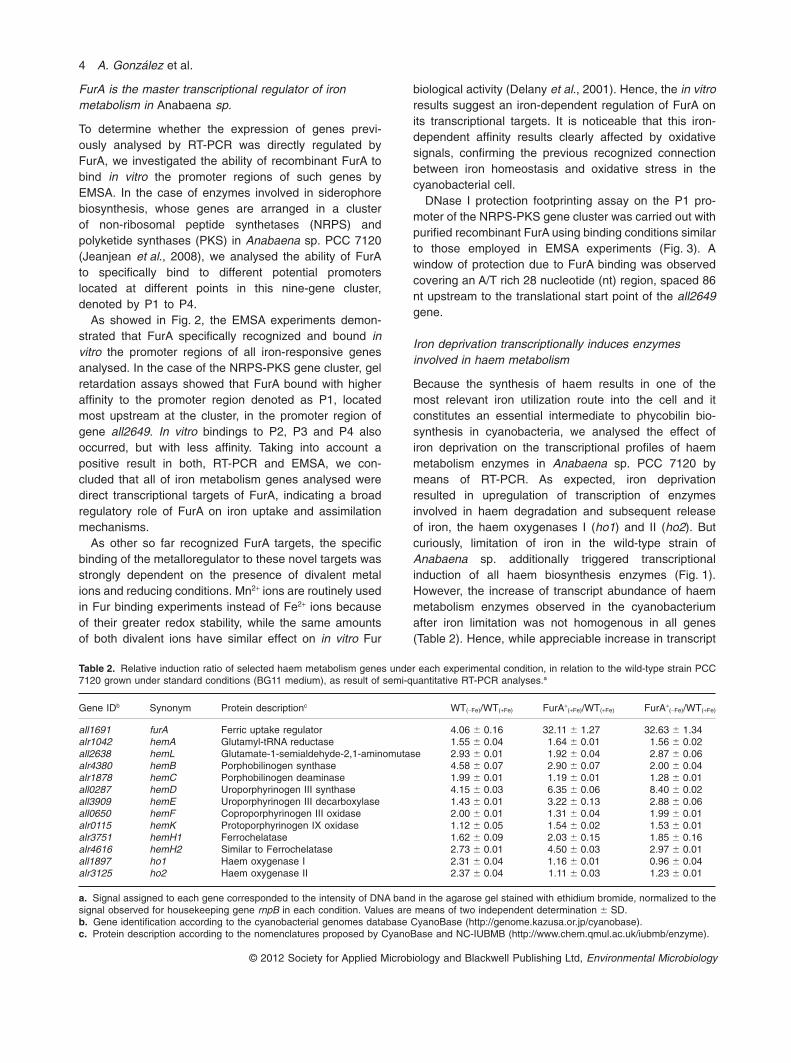
Table 2. Relative induction ratio of selected haem metabolism genes under each experimental condition, in relation to the wild-type strain PCC7120 grown under standard conditions (BG11 medium), as result of semi-quantitative RT-PCR analyses.a
Gene IDb Synonym Protein descriptionc WT(-Fe)/WT(+Fe) FurA+(+Fe)/WT(+Fe) FurA+
(-Fe)/WT(+Fe)
all1691 furA Ferric uptake regulator 4.06 � 0.16 32.11 � 1.27 32.63 � 1.34alr1042 hemA Glutamyl-tRNA reductase 1.55 � 0.04 1.64 � 0.01 1.56 � 0.02all2638 hemL Glutamate-1-semialdehyde-2,1-aminomutase 2.93 � 0.01 1.92 � 0.04 2.87 � 0.06alr4380 hemB Porphobilinogen synthase 4.58 � 0.07 2.90 � 0.07 2.00 � 0.04alr1878 hemC Porphobilinogen deaminase 1.99 � 0.01 1.19 � 0.01 1.28 � 0.01all0287 hemD Uroporphyrinogen III synthase 4.15 � 0.03 6.35 � 0.06 8.40 � 0.02all3909 hemE Uroporphyrinogen III decarboxylase 1.43 � 0.01 3.22 � 0.13 2.88 � 0.06all0650 hemF Coproporphyrinogen III oxidase 2.00 � 0.01 1.31 � 0.04 1.99 � 0.01alr0115 hemK Protoporphyrinogen IX oxidase 1.12 � 0.05 1.54 � 0.02 1.53 � 0.01alr3751 hemH1 Ferrochelatase 1.62 � 0.09 2.03 � 0.15 1.85 � 0.16alr4616 hemH2 Similar to Ferrochelatase 2.73 � 0.01 4.50 � 0.03 2.97 � 0.01all1897 ho1 Haem oxygenase I 2.31 � 0.04 1.16 � 0.01 0.96 � 0.04alr3125 ho2 Haem oxygenase II 2.37 � 0.04 1.11 � 0.03 1.23 � 0.01
a. Signal assigned to each gene corresponded to the intensity of DNA band in the agarose gel stained with ethidium bromide, normalized to thesignal observed for housekeeping gene rnpB in each condition. Values are means of two independent determination � SD.b. Gene identification according to the cyanobacterial genomes database CyanoBase (http://genome.kazusa.or.jp/cyanobase).c. Protein description according to the nomenclatures proposed by CyanoBase and NC-IUBMB (http://www.chem.qmul.ac.uk/iubmb/enzyme).
4 A. González et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology
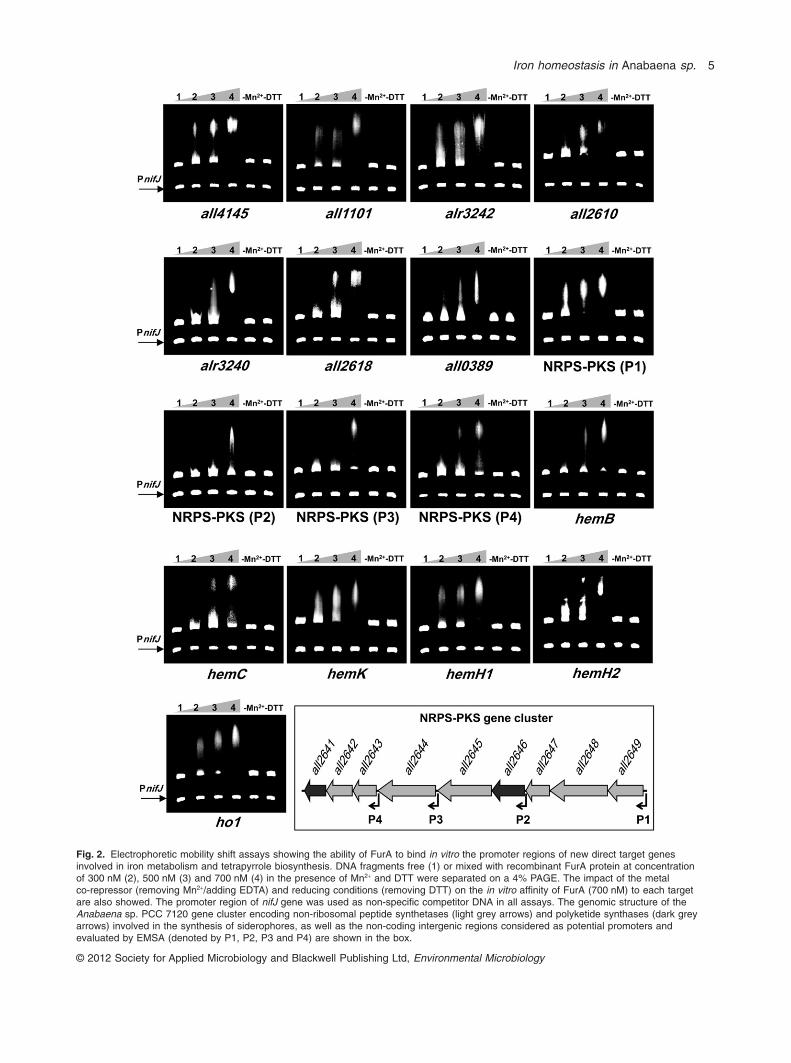
Fig. 2. Electrophoretic mobility shift assays showing the ability of FurA to bind in vitro the promoter regions of new direct target genesinvolved in iron metabolism and tetrapyrrole biosynthesis. DNA fragments free (1) or mixed with recombinant FurA protein at concentrationof 300 nM (2), 500 nM (3) and 700 nM (4) in the presence of Mn2+ and DTT were separated on a 4% PAGE. The impact of the metalco-repressor (removing Mn2+/adding EDTA) and reducing conditions (removing DTT) on the in vitro affinity of FurA (700 nM) to each targetare also showed. The promoter region of nifJ gene was used as non-specific competitor DNA in all assays. The genomic structure of theAnabaena sp. PCC 7120 gene cluster encoding non-ribosomal peptide synthetases (light grey arrows) and polyketide synthases (dark greyarrows) involved in the synthesis of siderophores, as well as the non-coding intergenic regions considered as potential promoters andevaluated by EMSA (denoted by P1, P2, P3 and P4) are shown in the box.
Iron homeostasis in Anabaena sp. 5
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

abundance was observed in some hem genes like hemBor hemD, other genes like hemK displayed a lessmarked induction after iron deprivation. These resultssuggest the existence of different ways of transcriptionalregulation and possibly the coordination of multipletranscription-control mechanisms acting on each indi-vidual gene of tetrapyrrole biosynthesis pathway underthe same environmental condition. These findings addi-tionally indicated that in contrast to non-photosyntheticorganisms where iron deprivation usually results in therepression of iron-consuming pathways including haembiosynthesis, the increased degradation of haem inAnabaena sp. PCC 7120 as iron source under iron-limited conditions transcriptionally induced the enzymesinvolved in its own biosynthesis, maybe as feedbackhomeostatic control in an attempt to maintain haem andphycobilins levels.
FurA overexpression induces changes in thetranscriptional pattern of haem metabolism enzymes
As the master transcriptional regulator of iron homeosta-sis in Anabaena sp., the influence of FurA on the tran-scriptional pattern of enzymes involved in the metabolismof one of the most important iron-containing molecules,the haem cofactor was examined. Overexpression ofFurA in Anabaena sp. PCC 7120 produced differenteffects on the transcriptional pattern of haem metabolismenzymes (Fig. 1). Several of these genes, includinghemB, hemC, ho1 and ho2 displayed a transcriptionalpattern change similar to those observed in previouslyanalysed genes coding for iron homeostasis players,where furA overexpression reduced the inducible effect ofiron starvation observed in the wild-type strain. However,the overexpression of the metalloregulator led to anappreciable induction on transcription of other hemgenes; thus, transcript abundances of hemD, hemE,hemK and hemH were significantly increased under afurA overexpression background (Table 2). In most ofthose genes, iron deprivation slightly affected the tran-scriptional induction observed with FurA overexpressionunder iron-replete conditions; while in the case of hemD,overexpression of Fur and iron starvation seemed to exerta synergistic inducible effect on transcription. On the otherhand, transcription of several hem genes appeared tooccur independently of FurA or at least, the regulatoryinfluence of this protein was only partial, indirect or coop-eratively acting with other transcriptional factors. Thus,the transcript levels of hemA, hemL or hemF under afurA overexpression background were similar to thoseobserved in the wild-type after iron-deprivation.
FurA regulates both synthesis and degradation of haem
The regulation of haem biosynthetic pathway, whichshares most enzymes with synthesis of chlorophyll andconstitutes also the initial steps of phycobilin biosynthe-sis, is quite complex and subject to several controls atdifferent levels. Therefore, all the transcriptional changesobserved in genes coding for the enzymes involved inboth biosynthesis and degradation of haem under a furAoverexpression background did not necessarily reflecta direct regulatory role of FurA on the transcription ofsuch enzymes, since some of those transcriptionalchanges could be a result of pleiotropic effects of furAoverexpression.
In order to define which enzymes involved in haemmetabolism were direct targets of FurA, we analysed theability of recombinant FurA to bind in vitro the promoterregions of all hem genes as well as those of the two haemoxygenases described in Anabaena sp. by using EMSA.As previously performed with iron metabolism genes, dif-ferent binding conditions were analysed for all genes
hemCNRPS PKS gene cl ster
-
hemC
5´-T
-
NRPS-PKS gene cluster (P1)
5´-CA
AC
TG
AC
AA
TA
TA
TT
AA
TA
GA
AA
CT
TG
C AG
AA
AT
CA
GA
AA
AT
GT
CA
AT
AA
TA
AA
TT
-3´
TT
-3´
Fig. 3. Sequences into the NRPS-PKS gene cluster promoterregion P1 and the hemC promoter protected by FurA. DNase Iprotection footprint assays were carried out with 5 fmol of32P-end-labelled 300–400 bp DNA fragment and 100, 200, 300 and500 nM of purified recombinant FurA in the presence of 100 mMMnCl2 and 1 mM DTT. Windows of protection due to FurA bindingare indicated by thick vertical lines.
6 A. González et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

tested to verify thereby all possible reaction environmentsthat could modify the in vitro affinity of FurA to potentialnew transcriptional targets.
The EMSA experiments indicated that FurA specificallybound to the promoter regions of four haem-biosynthesisenzymes encoding genes: hemB, hemC, hemK andhemH (Fig. 2), as well as to the promoter of the haem-degrading enzyme haem oxygenase I (ho1). None of theother analysed haem metabolism genes showed specificbinding to FurA according to EMSA (data not shown).Consistently with an iron-dependent regulation, the invitro affinity of FurA for these new identified targets wasmarkedly affected in the absent of divalent metal ionsand/or reducing agents, as occurs with all so far describedFurA transcriptional targets (Hernández et al., 2006b;González et al., 2010; 2011), suggesting a critical role ofmetal co-repressor and reducing conditions to the func-tion of this cyanobacterial metalloregulator in vivo.
DNase I footprinting analysis was carried out with puri-fied recombinant FurA using binding conditions similar to
those employed in EMSA experiments in order to deter-mine the FurA-binding site within the promoter of hemC.As shown in Fig. 3, a window of protection covering 28 ntinto the hemC promoter and spaced 102 nt upstream thetranslational start site was clearly observed.
Taken together, the combination of RT-PCR analysesand EMSA experiments led us to identify five novelFurA direct targets involved in tetrapyrrole biosynthesis(Fig. 4): the genes encoding haem-biosynthesis enzymesporphobilinogen synthase (hemB), porphobilinogendeaminase (hemC), protoporphyrinogen oxidase (hemK)and ferrochelatase (hemH ), but also the gene encodinghaem oxygenase I (ho1). Notably, two distinguishabletranscriptional pattern changes were observed for thesenew direct targets in relation to FurA levels and iron avail-ability. The findings suggest a dual role of FurA in thetranscriptional regulation of the tetrapryrrole biosynthesispathway, as iron-dependent transcriptional repressor ofhemB, hemC and ho1, but also as iron-dependent activa-tor of hemK and hemH, a phenomenon previously
Glutamyl-tRNA
GluTR (hemA)
Glutamate-1-semialdehyde
5-Aminolevulinic acid
Porphobilinogen
GSAM (hemL)
PBGS (hemB )
Uroporphyrinogen III
Hydroxymethylbilane
Sirohaem
Fe2+
PBGD (hemC )
UROS (hemD )
UROD (hemE )FurA
-
-
Coproporphyrinogen III
Protoporphyrinogen IX
Protoporphyrin IX Chlorophyll
CPO (hemF )
PPO (hemK )
+
+p p y
Haem
Fe2+
Mg2+
p y
FC (hemH )
-
Biliverdin IX
Fe2+
HO-1 (ho1 )
Phycobilins
Fig. 4. Model of tetrapyrrole biosynthesisregulation by FurA in Anabaena sp. PCC7120. The dual role of FurA as transcriptionalrepressor of hemB, hemC and ho1, but alsoas transcriptional activator of hemK andhemH is indicated. The full names of theenzymes are shown in Table 2.
Iron homeostasis in Anabaena sp. 7
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

observed with other transcriptional regulators controllingthis metabolic pathway (Zappa et al., 2010).
Bioinformatic analysis of footprinted FurA binding sitesreveals a putative regulatory consensus foriron-responsive genes
The sequences of eight FurA binding sites identifiedby DNase I footprinting analysis of six different iron-responsive gene promoters were aligned in order todefine a regulatory consensus. Despite the length of theregions protected by FurA varied in different promoters, awell-conserved 26 bp A/T-rich consensus was clearlyobtained (Fig. S1). A search of recognized Fur bindingmotifs into this consensus was carried out using the pro-moter analysis tool of the Virtual Footprint software suite(Munch et al., 2005), employing all the pre-existing weightmatrices belonging to Fur proteins of the PRODORIClibrary (Munch et al., 2003). Accordingly, three Fur bindingoctamer (8-mer) motifs in a head-to-head-to-tail arraywere identified covering 23 of the 26 bp consensusinitially obtained by the alignment. Therefore, only this23 bp sequence (TGATAAAAATTCTCAATAAATAA) wasdefined as the final putative FurA regulatory consensus(Fig. 5).
Discussion
Despite the number of studies that have been undertakenon cyanobacterial response to iron deficiency (Ferreiraand Strauss, 1994; Singh and Sherman, 2000; Singhet al., 2003; Shi et al., 2007; Alexova et al., 2011), as wellas the increasing identification of iron uptake and storageplayers (Goldman et al., 1983; Keren et al., 2004;
Shcolnick et al., 2007; Nicolaisen et al., 2008; Mirus et al.,2009), knowledge on the regulating mechanisms of ironhomeostasis in cyanobacteria is limited (Nodop et al.,2008; Shcolnick et al., 2009).
In the filamentous diazotrophic cyanobacterium Ana-baena sp. PCC 7120, three Fur homologues have beendescribed (Hernández et al., 2004b), although only FurAseems to be directly involved in control of iron metabolism(González et al., 2010). In this study, we unravel the roleof FurA as the master regulator of iron metabolism ana-lysing the impact of FurA overexpression and iron avail-ability on the transcriptional modulation of a broad rangeof Anabaena iron uptake, transport, storage and cellulariron utilization mechanisms.
In cyanobacteria, siderophore-mediated iron uptake isthought to be an evolutionary advance that contributes todominate iron-limited environments. Anabaena sp. PCC7120 synthesizes schizokinen as its major siderophore,although iron uptake from aerobactin and ferrioxamine Bhas also been reported (Goldman et al., 1983). The bio-synthesis of siderophores is presumably mediated by acluster of nine open reading frames, from all2641 toall2649, encoding seven non-ribosomal peptide syn-thetases (NRPSs) and two polyketide synthases (PKSs)which are transcriptionally activated under iron limitationor oxidative stress conditions (Jeanjean et al., 2008).The comparative analysis of gene expression of two ofthese enzymes (all2646 and all2648) in the Anabaenawild-type strain PCC 7120 and the Anabaena sp. furA-overexpressing strain AG2770FurA displayed a typicaliron-responsive FurA-repressed transcriptional pattern.Both enzymes were strongly induced in the wild-typestrain under iron-restricted conditions, but the inducibleeffect of iron starvation was partially reduced under aFurA overexpression background, where the increasedamount of the transcriptional repressor hinders the com-plete downregulation of its targets. Recombinant FurAprotein specifically bound to at least four intergenicregions into this nine-gene NRPSs-PKSs cluster.
Siderophore–iron complexes are bound by outer mem-brane receptor proteins, the TonB-dependent transporters(TBDTs), which are composed of a transmembraneb-barrel domain that encloses a globular plug domain, anda periplasmic exposed TonB box (Noinaj et al., 2010).Bacteria often possess multiple TBDT receptors, eachproviding the bacterium with specificity for differentsiderophores. These outer membrane receptors are gen-erally induced by iron starvation and usually are notpresent or poorly expressed under iron-sufficient condi-tions (Andrews et al., 2003). Twenty-two TBDTs havebeen identified in the genome sequence of Anabaena sp.PCC 7120, most of them integrated into gene clustersor even putative operons containing genes coding forproteins involved in iron transport (Mirus et al., 2009). Our
A
B TGATAAAAATTCTCAATAAATAAACTATTTTTAAGAGTTATTTATT
Fig. 5. The putative FurA regulatory consensus in Anabaena sp.PCC 7120.A. The sequence logo created with the WebLogo server(http://weblogo.berkeley.edu/) was drawn from eight different FurAbinding sites identified by DNase I footprinting analysis of sixiron-responsive target promoters. The degree of sequenceconservation at each position is indicated as a total height of astack of letters, measured in arbitrary ‘bit’ units, with two bitspossible at each position.B. Putative Fur binding motifs identified by Virtual Footprint (Munchet al., 2005) are indicated by arrows.
8 A. González et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

previous results have shown that the alr0397 gene,coding for a schizokinen TBDT (Nicolaisen et al., 2008),is a direct transcriptional target of FurA (Gonzálezet al., 2010). The analysis of gene expression of theferrichrome-type TBDT coding genes all2610 and all1101,and the hutA-like haem TBDT coding the alr3242 gene inresponse to iron deprivation and/or FurA overexpressioncarried out in the present study suggests an iron-dependent FurA regulation on the transcription of thesethree TBDT; in all cases, the inducible effect of iron star-vation observed in the wild-type strain was partiallyreduced under a FurA overexpression background. Func-tional involvement of FurA was further confirmed byEMSA. A similar pattern of gene expression and EMSAresults were also observed with the alr3240, all2618 andall0389 genes, encoding periplasmic and inner mem-brane ABC transporter proteins involved in translocationof siderophore–iron complexes from periplasm to cytosol.
Due to the limited bioavailability or iron and its frequentintermittent supply in nature, bacteria have developedefficient iron storage mechanisms involving ferritinsand bacterioferritins (Andrews, 1998), ubiquitously multi-subunit proteins that can accommodate up to 4500 ironatoms into a central cavity in a form that is unlike toparticipate in ROS generation reactions (Theil, 1987). InSynechocystis sp. PCC 6803, bacterioferritins BfrA andBfrB are responsible for the storage up to 50% of intrac-ellular iron content (Keren et al., 2004), while the DPSfamily ferritin MrgA plays a critical role in the mobilizationof the stored iron within the cell (Shcolnick et al., 2007).By contrast, little is known about the mechanisms of ironstorage in Anabaena species. Only four non-haem-binding ferritin family genes have been identified in Ana-baena sp. PCC 7120 (Keren et al., 2004), includingalr3808, a DNA-binding protein homologue to DpsA fromSynechococcus sp. PCC 7942 (Peña and Bullerjahn,1995) which appears transcriptionally regulated by FurA(Hernández et al., 2007). DpsA displayed a weak catalaseactivity in vitro and is presumably involved in peroxide-consuming mechanism located on the chromosomalDNA, conferring resistance to peroxide damage duringoxidative stress conditions or long term nutrient limitation(Peña and Bullerjahn, 1995). The results presented heregive evidence that FurA additionally modulates theexpression of another DPS family protein, encoded bygene all4145, which shares more than 60% of amino acidsequence identity with MrgA from Synechocystis.
Haem biosynthesis becomes in one of the most impor-tant routes of iron utilization in both prokaryotic andeukaryotic organisms. In cyanobacteria, there is a con-tinuous requirement for haem synthesis to supply respi-ratory cytochromes and other relevant haem-proteins(Ogola et al., 2009; Sturms et al., 2011), but also as inter-mediate in the synthesis of other essential tetrapyrroles,
the light-harvesting photosynthetic accessory pigmentsphycobilins (Beale, 1994). Cyanobacteria produce fourclasses of tetrapyrroles (chlorophyll, haem, sirohaem andphycobilins) via a common branched pathway. Tetrapyr-roles biosynthesis must be tightly regulated since differentamounts of each tetrapyrrole end product are required bycells; but in addition, all the tetrapyrrole intermediates arephototoxic and generate ROS when they are excited bylight. Post-transcriptional modulation of enzyme activitiescontributes to a quite fine-tuned adaptation of tetrapyrrolebiosynthesis to environmental conditions (Czarnecki andGrimm, 2012). However, the control of enzyme abun-dance at the level of gene expression have been alsoextensively demonstrated (Qi and O’Brian, 2002; Zappaet al., 2010; Aoki et al., 2012). In the present study weprovide strong evidence that FurA modulates the expres-sion of at least five genes coding for enzymes involved intetrapyrrole biosynthesis, mediating at early stages bycontrolling porphobilinogen synthase (hemB) and porpho-bilinogen deaminase (hemC) levels, but also at the finalsteps of haem biosynthesis branch, controlling theexpression of protoporphyrinogen oxidase (hemK ) andferrochelatase (hemH ). In addition, FurA appears to regu-late the haem-degrading enzyme haem oxygenase 1(ho1), the first step of the phycobilin synthesis branch.
Connection between cellular iron levels and tetrapyrrolebiosynthesis has been previously recognized. The Furfamily transcriptional regulator Irr negatively modulatesthe pathway at hemB in Bradyrhizobium japonicum underiron limitation, sensing the iron through the status of haemby a mechanism of direct interaction with ferrochelatase(Chauhan et al., 1997; Qi and O’Brian, 2002). Curiously,iron deprivation usually results in downregulation ofiron-consuming metabolic pathways in non-phototrophicorganisms, including haem biosynthesis (Schobert andJahn, 2002), a reasonable response to avoid deleteriousaccumulation of protoporphyrin IX, the immediate precur-sor of haem. By contrast, iron deprivation leads to induc-tion of porphyrin biosynthesis in plants (Duggan andGassman, 1974), a similar phenotype to that observed inAnabaena sp. under our experimental conditions. Sincehaem constitutes an intermediate in the synthesis of otheressential tetrapyrroles in plants and cyanobacteria (phy-tochromobilin and phycobilins), it is likely that increasedhaem degradation under iron limitation conditions asendogenous source of iron exerts a positive feedback onits own synthesis.
Interestingly, our results suggest a dual role of FurA onthe transcription of tetrapyrrole biosynthesis genes,repressing transcription of hemB, hemC and ho1, butactivating at the same time the expression of hemK andhemH. This dual regulatory role on the tetrapyrrole bio-synthesis pathway have been previously observed in atleast four transcriptional regulators, CrtJ, FnrL, AerR and
Iron homeostasis in Anabaena sp. 9
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

HbrL, in the facultative photosynthetic bacterium Rhodo-bacter capsulatus (Zappa et al., 2010), and seems tofacilitate adjustment between early and late steps oftetrapyrrole biosynthesis to ensure, in addition to post-translational controls, the balanced metabolic flux in thepathway by proper modulation of enzyme abundance.
FurA binds haem in the micromolar range of concentra-tion, inhibiting its in vitro DNA binding ability in aconcentration-dependent manner (Hernández et al.,2004a). The physicochemical features of the haembinding to FurA suggest a putative role of this transcrip-tional regulator as a haem sensor protein, which useshaem as a signalling molecule (Pellicer et al., 2012). Thephysiological meaning of the FurA-haem interactionremains undetermined. However, accordingly to theresults presented here, we suggest that haem levelscould directly modulate by feedback and feedforwardmechanisms the transcription of FurA-mediated genescoding for some of haem metabolism enzymes. Hence,the synthesis of haem from protoporphyrin IX by ferroche-latase (FC) or its degradation by haem oxygenase I(HO-1) would be feedback/feedforward controlled byhaem at level of enzyme synthesis via FurA. The verifica-tion of this hypothesis awaits further studies.
Sequence alignments of entire FurA target promoterregions have usually revealed minimal homology. Previ-ous attempts to define a functional FurA box sequenceinto the furA promoter have failed, since a short syntheticDNA sequence containing the putative iron boxes onlydisplayed quite feeble binding of the metalloregulator(González et al., 2011). Thus, a proper DNA conformationinto an A/T-rich operator region appeared to be essentialfor an efficient recognition and high affinity binding, ratherthan a conserved iron box sequence. In the present workwe have defined a putative FurA regulatory consensussequence, which was deduced from a well-conserved23 bp region observed in eight FurA footprinted bindingsites from six iron-responsive FurA targets. The in silicoanalysis of this putative regulatory consensus supportsthe hypothesis that cyanobacterial FurA, as occurs withFur from heterotrophic bacteria (Baichoo and Helmann,2002; Lavrrar and McIntosh, 2003), interacts with DNA asa dimeric protein recognizing arrays of conserved motifs.The presence of at least three 8-mer motifs in the con-sensus sequence, as well as the extension of the FurAprotected regions in footprinting assays are consistentwith the idea that more than one dimer of FurA binds tothe operator for transcriptional regulation.
In summary, the results presented here demonstratedthe role of FurA as the master regulator of iron homeos-tasis in Anabaena sp. PCC 7120, modulating the expres-sion of genes involved in every step of iron homeostasis.Comparative gene expression analyses under differentiron availability conditions and FurA expression levels, in
combination with in vitro binding experiments, led todefine a variety of novel direct iron-dependent transcrip-tional targets of this metalloregulator in Anabaena sp.PCC 7120, including genes encoding at least fiveenzymes involved in tetrapyrrole biosynthesis. To our bestknowledge, this is the first evidence of Fur proteins, dif-ferent of the Irr homologue from rhizobia, controlling theexpression of tetrapyrrole biosynthesis enzymes inresponse to iron availability.
Experimental procedures
Bacterial strains and culture conditions
Wild-type Anabaena sp. PCC 7120 and its derivative furA-overexpressing strain AG2770FurA (González et al., 2010)were routinely grown photoautotrophically in BG-11 medium(Rippka et al., 1979) at 30°C, supplemented with neomycin50 mg ml-1 in the case of strain AG2770FurA. Cyanobacterialstrains were cultured in 250 ml Erlenmeyer flasks containing60 ml culture medium under continuous white light illumina-tion (20 mE m-2 s-1) and shaking (120 r.p.m.). Chlorophyll a(Chl) was determined in methanol extracts (Nicolaisen et al.,2008).
For iron depletion experiments, exponentially growing cells(3–5 mg Chl ml-1) were harvested by centrifugation, washedthree times with BG-11 medium without iron (BG-11-Fe),re-suspended in the same iron-free medium and furthergrown for 24 h. Glassware used in these experiments wassoaked with 6 M HCl and extensively rinsed with Milli-Q waterto remove residual iron.
Semi-quantitative reverse transcription (RT)-PCR
Total RNA from exponentially growing Anabaena sp. strainswas isolated as described previously (Olmedo-Verd et al.,2005). Samples of 1 mg RNA were heated at 85°C for 10 minand used as templates for the first strand cDNA synthesis.Residual DNA in RNA preparations was eliminated by diges-tion with RNase-free DNase I (Roche, Basel, Switzerland).The absence of DNA was checked by PCR. Reverse tran-scription was carried out using SuperScript retrotranscriptase(Invitrogen, Carlsbad, CA, USA) in a 20 ml reaction volumecontaining 150 ng of random primers (Invitrogen, Carlsbad,CA, USA), 1 mM dNTP mix (GE Healthcare, Buckingham-shire, UK) and 10 mM DTT. The sequences of the specificprimers used for RT-PCR reactions are defined in Table S1.Housekeeping gene rnpB (Vioque, 1992) was used as acontrol to compensate for variations in the input of RNAamounts and normalize the results. Exponential phase ofRT-PCR for each gene was determined by measuring theamount of PCR product at different time intervals. For thefinal results, 20–23 cycles at the early exponential phasewere used in all genes analysed. The PCR products wereresolved by electrophoresis in 1% agarose gels, stained withethidium bromide and analysed using a Gel Doc 2000 ImageAnalyzer (Bio-Rad, Hercules, CA, USA).
Relative induction ratio for each gene was calculated as theratio between signals observed in AG2770FurA and wild-typestrains in at least two independent determinations. Signal
10 A. González et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

assigned to each gene was corresponded to the intensity ofits DNA band in the agarose gel stained with ethidiumbromide normalized to the signal observed for the house-keeping gene rnpB in each strain.
Electrophoretic mobility shift assays
The promoter regions of each gene of interest were obtainedby PCR using the primers described in Table S1. Electro-phoretic mobility shift assays (EMSA) were performed asdescribed previously (Hernández et al., 2006b). Briefly, 120–140 ng of each DNA fragment was mixed with recombinantFurA protein at concentrations of 300, 500 and 700 nM in a20 ml reaction volume containing 10 mM bis-Tris (pH 7.5),40 mM potassium chloride, 100 mg ml-1 bovine serumalbumin, 1 mM DTT, 100 mM manganese chloride, and 5%glycerol. In some experiments, EDTA was added to a finalconcentration of 200 mM. To insure the specificity of EMSA,the promoter region of Anabaena sp. nifJ (alr1911) gene(López-Gomollón et al., 2007) was included as non-specificcompetitor DNA in all assays. Mixtures were incubated atroom temperature for 20 min, and subsequently separated on4% non-denaturing polyacrylamide gels in running buffer(25 mM Tris, 190 mM glycine) at 90 V. Gels were stained withSYBR® Safe DNA gel stain (Invitrogen, Carlsbad, CA, USA)and processed with a Gel Doc 2000 Image Analyzer (Bio-Rad, Hercules, CA, USA).
DNase I footprinting analysis
DNase I protection assays were performed as previouslydescribed (Valladares et al., 2008). Protein–DNA complexeswere formed in a final volume of 70 ml of buffer [10 mMTris-HCl (pH 8.0), 30 mM KCl, 10 mM MgCl2, 5 mM CaCl2,2 mM DTT, 0.07 mg ml-1 bovine serum albumin, 0.07 mg ml-1
poly(dIdC)], with 10 fmol of 32P-end-labelled (with T4 polynu-cleotide kinase and [g-32P]-ATP) DNA fragment, 1 mM ATP,and the reconstituted RNAP holoenzyme. The reaction mix-tures were incubated for 30 min at 32°C. Then, 1 U DNase I(Roche, Basel, Switzerland) was added in a buffer containing50 mM sodium acetate and 6 mM MgSO4, and the reactionwas immediately interrupted with 320 ml of stop solution[0.5 mM EDTA, 0.125% SDS, 12.5 mM ammonium acetate(pH 5.2)]. The products were resolved in 6% polyacrylamide– 4.68 M urea gels next to the corresponding sequencingladder. Radioactive bands were visualized with a Cyclonestorage phosphor system and OptiQuant image analysis soft-ware (Packard Instrument, Meriden, CT, USA).
In silico analyses
Alignments of FurA binding sites were carried out usingthe AlignX software of the Vector NTI® suite (Invitrogen,Carlsbad, CA, USA). Searching of putative Fur binding motifswas performed through the promoter analysis tool of theVirtual Footprint framework program (Munch et al., 2005),using the pre-existing position weight matrices for Fur ofEscherichia coli K12, Pseudomonas aeruginosa ATCC15692/PAO1, and Helicobacter pylori ATCC 700392/26695.Sensitivity settings were varied up to 1. The relative conser-
vation of nucleotides at their respective positions were calcu-lated and graphically represented as sequence logo by usingthe WebLogo server at http://weblogo.berkeley.edu (Crookset al., 2004).
Acknowledgements
This work has been supported by grant BFU2009-07424 fromMinisterio de Ciencia e Innovación (Spain), and grant 2009/0372 from Diputación General de Aragón (Obra Social LaCaixa). Authors thank to Drs E. Flores and A. Herrero forproviding the facilities for the footprinting assays.
References
Alexova, R., Fujii, M., Birch, D., Cheng, J., Waite, T.D.,Ferrari, B.C., and Neilan, B.A. (2011) Iron uptake and toxinsynthesis in the bloom-forming Microcystis aeruginosaunder iron limitation. Environ Microbiol 13: 1064–1077.
Andrews, S.C. (1998) Iron storage in bacteria. Adv MicrobPhysiol 40: 281–351.
Andrews, S.C., Robinson, A.K., and Rodríguez-Quiñones, F.(2003) Bacterial iron homeostasis. FEMS Microbiol Rev27: 215–237.
Aoki, R., Takeda, T., Omata, T., Ihara, K., and Fujita, Y. (2012)MarR-type transcriptional regulator ChlR activates expres-sion of tetrapyrrole biosynthesis genes in response to low-oxygen conditions in cyanobacteria. J Biol Chem 287:13500–13507.
Baichoo, N., and Helmann, J.D. (2002) Recognition of DNAby Fur: a reinterpretation of the Fur box consensussequence. J Bacteriol 184: 5826–5832.
Beale, S.I. (1994) Biosynthesis of cyanobacterial tetrapyrrolepigments: hemes, chlorophylls, and phycobilins. In TheMolecular Biology of Cyanobacteria. Bryant, D.A. (ed.).Dordrecht, the Netherlands: Kluwer Academic Publishers,pp. 519–558.
Brown, S.B., Houghton, J.D., and Vernon, D.I. (1990) Biosyn-thesis of phycobilins. Formation of the chromophore ofphytochrome, phycocyanin and phycoerythrin. J Photo-chem Photobiol B 5: 3–23.
Chauhan, S., Titus, D.E., and O’Brian, M.R. (1997) Metalscontrol activity and expression of the heme biosynthesisenzyme delta-aminolevulinic acid dehydratase inBradyrhizobium japonicum. J Bacteriol 179: 5516–5520.
Cornah, J.E., Terry, M.J., and Smith, A.G. (2003) Green orred: what stops the traffic in the tetrapyrrole pathway?Trends Plant Sci 8: 224–230.
Crooks, G.E., Hon, G., Chandonia, J.M., and Brenner, S.E.(2004) WebLogo: a sequence logo generator. GenomeRes 14: 1188–1190.
Czarnecki, O., and Grimm, B. (2012) Post-translationalcontrol of tetrapyrrole biosynthesis in plants, algae, andcyanobacteria. J Exp Bot 63: 1675–1687.
Delany, I., Spohn, G., Rappuoli, R., and Scarlato, V. (2001)The Fur repressor controls transcription of iron-activatedand -repressed genes in Helicobacter pylori. Mol Microbiol42: 1297–1309.
Delany, I., Rappuoli, R., and Scarlato, V. (2004) Fur functionsas an activator and as a repressor of putative virulencegenes in Neisseria meningitidis. Mol Microbiol 52: 1081–1090.
Iron homeostasis in Anabaena sp. 11
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

Duggan, J., and Gassman, M. (1974) Induction of porphyrinsynthesis in etiolated bean leaves by chelators of iron.Plant Physiol 53: 206–215.
Escolar, L., Pérez-Martín, J., and de Lorenzo, V. (1999)Opening the iron box: transcriptional metalloregulation bythe Fur protein. J Bacteriol 181: 6223–6229.
Ferreira, F., and Strauss, N.A. (1994) Iron deprivation incyanobacteria. J Appl Phycol 6: 199–210.
Geider, R.J., and La Roche, J. (1994) The role of iron inphytoplankton photosynthesis and the potential for ironlimitation of primary productivity in the sea. Photosynth Res39: 275–301.
Ghassemian, M., and Straus, N.A. (1996) Fur regulates theexpression of iron-stress genes in the cyanobacteriumSynechococcus sp. strain PCC 7942. Microbiology 142 (Pt6): 1469–1476.
Goldman, S.J., Lammers, P.J., Berman, M.S., and Sanders-Loehr, J. (1983) Siderophore-mediated iron uptake in dif-ferent strains of Anabaena sp. J Bacteriol 156: 1144–1150.
González, A., Bes, M.T., Barja, F., Peleato, M.L., and Fillat,M.F. (2010) Overexpression of FurA in Anabaena sp. PCC7120 reveals new targets for this regulator involved inphotosynthesis, iron uptake and cellular morphology. PlantCell Physiol 51: 1900–1914.
González, A., Bes, M.T., Peleato, M.L., and Fillat, M.F. (2011)Unravelling the regulatory function of FurA in Anabaena sp.PCC 7120 through 2-D DIGE proteomic analysis. J Pro-teomics 74: 660–671.
Heinemann, I.U., Jahn, M., and Jahn, D. (2008) The bio-chemistry of heme biosynthesis. Arch Biochem Biophys474: 238–251.
Hernández, J.A., Bes, M.T., Fillat, M.F., Neira, J.L., andPeleato, M.L. (2002) Biochemical analysis of the recom-binant Fur (ferric uptake regulator) protein from AnabaenaPCC 7119: factors affecting its oligomerization state.Biochem J 366: 315–322.
Hernández, J.A., Peleato, M.L., Fillat, M.F., and Bes, M.T.(2004a) Heme binds to and inhibits the DNA-binding activ-ity of the global regulator FurA from Anabaena sp. PCC7120. FEBS Lett 577: 35–41.
Hernández, J.A., López-Gomollón, S., Bes, M.T., Fillat, M.F.,and Peleato, M.L. (2004b) Three fur homologues fromAnabaena sp. PCC 7120: exploring reciprocal protein-promoter recognition. FEMS Microbiol Lett 236: 275–282.
Hernández, J.A., Muro-Pastor, A.M., Flores, E., Bes, M.T.,Peleato, M.L., and Fillat, M.F. (2006a) Identification of afurA cis antisense RNA in the cyanobacterium Anabaenasp. PCC 7120. J Mol Biol 355: 325–334.
Hernández, J.A., López-Gomollón, S., Muro-Pastor, A., Val-ladares, A., Bes, M.T., Peleato, M.L., and Fillat, M.F.(2006b) Interaction of FurA from Anabaena sp. PCC 7120with DNA: a reducing environment and the presence ofMn(2+) are positive effectors in the binding to isiB and furApromoters. Biometals 19: 259–268.
Hernández, J.A., Pellicer, S., Huang, L., Peleato, M.L., andFillat, M.F. (2007) FurA modulates gene expression ofalr3808, a DpsA homologue in Nostoc (Anabaena) sp.PCC 7120. FEBS Lett 581: 1351–1356.
Jeanjean, R., Talla, E., Latifi, A., Havaux, M., Janicki, A., andZhang, C.C. (2008) A large gene cluster encoding peptidesynthetases and polyketide synthases is involved in pro-
duction of siderophores and oxidative stress response inthe cyanobacterium Anabaena sp. strain PCC 7120.Environ Microbiol 10: 2574–2585.
Keren, N., Aurora, R., and Pakrasi, H.B. (2004) Critical rolesof bacterioferritins in iron storage and proliferation ofcyanobacteria. Plant Physiol 135: 1666–1673.
Latifi, A., Jeanjean, R., Lemeille, S., Havaux, M., and Zhang,C.C. (2005) Iron starvation leads to oxidative stress inAnabaena sp. strain PCC 7120. J Bacteriol 187: 6596–6598.
Latifi, A., Ruiz, M., and Zhang, C.C. (2009) Oxidative stress incyanobacteria. FEMS Microbiol Rev 33: 258–278.
Lavrrar, J.L., and McIntosh, M.A. (2003) Architecture of a Furbinding site: a comparative analysis. J Bacteriol 185:2194–2202.
Layer, G., Reichelt, J., Jahn, D., and Heinz, D.W. (2010)Structure and function of enzymes in heme biosynthesis.Protein Sci 19: 1137–1161.
López-Gomollón, S., Hernández, J.A., Pellicer, S., Angarica,V.E., Peleato, M.L., and Fillat, M.F. (2007) Cross-talkbetween iron and nitrogen regulatory networks in Ana-baena (Nostoc) sp. PCC 7120: identification of overlappinggenes in FurA and NtcA regulons. J Mol Biol 374: 267–281.
Martin-Luna, B., Hernández, J.A., Bes, M.T., Fillat, M.F., andPeleato, M.L. (2006a) Identification of a ferric uptake regu-lator from Microcystis aeruginosa PCC 7806. FEMS Micro-biol Lett 254: 63–70.
Martin-Luna, B., Sevilla, E., Hernandez, J.A., Bes, M.T., Fillat,M.F., and Peleato, M.L. (2006b) Fur from Microcystis aeru-ginosa binds in vitro promoter regions of the microcystinbiosynthesis gene cluster. Phytochemistry 67: 876–881.
Masse, E., and Gottesman, S. (2002) A small RNA regulatesthe expression of genes involved in iron metabolism inEscherichia coli. Proc Natl Acad Sci USA 99: 4620–4625.
Mirus, O., Strauss, S., Nicolaisen, K., von Haeseler, A., andSchleiff, E. (2009) TonB-dependent transporters and theiroccurrence in cyanobacteria. BMC Biol 7: 68.
Munch, R., Hiller, K., Barg, H., Heldt, D., Linz, S., Wingender,E., and Jahn, D. (2003) PRODORIC: prokaryotic databaseof gene regulation. Nucleic Acids Res 31: 266–269.
Munch, R., Hiller, K., Grote, A., Scheer, M., Klein, J.,Schobert, M., and Jahn, D. (2005) Virtual Footprint andPRODORIC: an integrative framework for regulon predic-tion in prokaryotes. Bioinformatics 21: 4187–4189.
Napolitano, M., Rubio, M.A., Santamaria-Gomez, J.,Olmedo-Verd, E., Robinson, N.J., and Luque, I. (2012)Characterization of the response to zinc deficiency in thecyanobacterium Anabaena sp. strain PCC 7120. J Bacte-riol 194: 2426–2436.
Nicolaisen, K., Moslavac, S., Samborski, A., Valdebenito, M.,Hantke, K., Maldener, I., et al. (2008) Alr0397 is an outermembrane transporter for the siderophore schizokinen inAnabaena sp. strain PCC 7120. J Bacteriol 190: 7500–7507.
Nicolaisen, K., Hahn, A., Valdebenito, M., Moslavac, S., Sam-borski, A., Maldener, I., et al. (2010) The interplay betweensiderophore secretion and coupled iron and copper trans-port in the heterocyst-forming cyanobacterium Anabaenasp. PCC 7120. Biochim Biophys Acta 1798: 2131–2140.
Nodop, A., Pietsch, D., Hocker, R., Becker, A., Pistorius,E.K., Forchhammer, K., and Michel, K.P. (2008) Transcript
12 A. González et al.
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology

profiling reveals new insights into the acclimation of themesophilic fresh-water cyanobacterium Synechococcuselongatus PCC 7942 to iron starvation. Plant Physiol 147:747–763.
Noinaj, N., Guillier, M., Barnard, T.J., and Buchanan, S.K.(2010) TonB-dependent transporters: regulation, structure,and function. Annu Rev Microbiol 64: 43–60.
Ogola, H.J., Kamiike, T., Hashimoto, N., Ashida, H., Ishikawa,T., Shibata, H., and Sawa, Y. (2009) Molecular characteri-zation of a novel peroxidase from the cyanobacterium Ana-baena sp. strain PCC 7120. Appl Environ Microbiol 75:7509–7518.
Olmedo-Verd, E., Flores, E., Herrero, A., and Muro-Pastor,A.M. (2005) HetR-dependent and -independent expressionof heterocyst-related genes in an Anabaena strain overpro-ducing the NtcA transcription factor. J Bacteriol 187: 1985–1991.
Pellicer, S., González, A., Peleato, M.L., Martinez, J.I., Fillat,M.F., and Bes, M.T. (2012) Site-directed mutagenesis andspectral studies suggest a putative role of FurA from Ana-baena sp. PCC 7120 as a heme sensor protein. FEBS J279: 2231–2246.
Peña, M.M., and Bullerjahn, G.S. (1995) The DpsA protein ofSynechococcus sp. strain PCC 7942 is a DNA-bindinghemoprotein. Linkage of the Dps and bacterioferritinprotein families. J Biol Chem 270: 22478–22482.
Qi, Z., and O’Brian, M.R. (2002) Interaction between thebacterial iron response regulator and ferrochelatase medi-ates genetic control of heme biosynthesis. Mol Cell 9:155–162.
Rippka, R., Deruelles, J., Waterbury, J.B., Herdman, M., andStanier, R.Y. (1979) Generic assignments, strain historiesand properties of pure cultures of Cyanobacteria. J GenMicrobiol 111: 1–61.
Schobert, M., and Jahn, D. (2002) Regulation of heme bio-synthesis in non-phototrophic bacteria. J Mol Microbiol Bio-technol 4: 287–294.
Shcolnick, S., and Keren, N. (2006) Metal homeostasis incyanobacteria and chloroplasts. Balancing benefits andrisks to the photosynthetic apparatus. Plant Physiol 141:805–810.
Shcolnick, S., Shaked, Y., and Keren, N. (2007) A role formrgA, a DPS family protein, in the internal transport of Fein the cyanobacterium Synechocystis sp. PCC 6803.Biochim Biophys Acta 1767: 814–819.
Shcolnick, S., Summerfield, T.C., Reytman, L., Sherman,L.A., and Keren, N. (2009) The mechanism of iron home-ostasis in the unicellular cyanobacterium Synechocystissp. PCC 6803 and its relationship to oxidative stress. PlantPhysiol 150: 2045–2056.
Shi, T., Sun, Y., and Falkowski, P.G. (2007) Effects ofiron limitation on the expression of metabolic genes in
the marine cyanobacterium Trichodesmium erythraeumIMS101. Environ Microbiol 9: 2945–2956.
Singh, A.K., and Sherman, L.A. (2000) Identification ofiron-responsive, differential gene expression in the cyano-bacterium Synechocystis sp. strain PCC 6803 with acustomized amplification library. J Bacteriol 182: 3536–3543.
Singh, A.K., McIntyre, L.M., and Sherman, L.A. (2003)Microarray analysis of the genome-wide response to irondeficiency and iron reconstitution in the cyanobacteriumSynechocystis sp. PCC 6803. Plant Physiol 132: 1825–1839.
Sturms, R., DiSpirito, A.A., Fulton, D.B., and Hargrove, M.S.(2011) Hydroxylamine reduction to ammonium by plant andcyanobacterial hemoglobins. Biochemistry 50: 10829–10835.
Taketani, S., Adachi, Y., and Nakahashi, Y. (2000) Regulationof the expression of human ferrochelatase by intracellulariron levels. Eur J Biochem 267: 4685–4692.
Theil, E.C. (1987) Ferritin: structure, gene regulation, andcellular function in animals, plants, and microorganisms.Annu Rev Biochem 56: 289–315.
Valladares, A., Flores, E., and Herrero, A. (2008) Transcrip-tion activation by NtcA and 2-oxoglutarate of three genesinvolved in heterocyst differentiation in the cyanobacteriumAnabaena sp. strain PCC 7120. J Bacteriol 190: 6126–6133.
Vavilin, D.V., and Vermaas, W.F. (2002) Regulation of thetetrapyrrole biosynthetic pathway leading to heme andchlorophyll in plants and cyanobacteria. Physiol Plant 115:9–24.
Vioque, A. (1992) Analysis of the gene encoding the RNAsubunit of ribonuclease P from cyanobacteria. NucleicAcids Res 20: 6331–6337.
Zappa, S., Li, K., and Bauer, C.E. (2010) The tetrapyrrolebiosynthetic pathway and its regulation in Rhodobactercapsulatus. Adv Exp Med Biol 675: 229–250.
Supporting information
Additional Supporting Information may be found in the onlineversion of this article:
Fig. S1. Sequence alignment of FurA binding sites identifiedby DNase I footprinting analysis of six iron-responsive genepromoters.Table S1. Primers used in this study.
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting materials suppliedby the authors. Any queries (other than missing material)should be directed to the corresponding author for the article.
Iron homeostasis in Anabaena sp. 13
© 2012 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology




















