
Using enzymes to remove biofilms of bacterial isolates sampled in the food-industry
12
PLEASE SCROLL DOWN FOR ARTICLE This article was downloaded by: [lequette, yannick] On: 1 March 2010 Access details: Access Details: [subscription number 919582326] Publisher Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37- 41 Mortimer Street, London W1T 3JH, UK Biofouling Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t713454511 Using enzymes to remove biofilms of bacterial isolates sampled in the food-industry Yannick Lequette a ; Gauthier Boels b ; Martine Clarisse a ; Christine Faille b a INRA, Villeneuve d'Ascq, France b REALCO S.A., Louvain-la-Neuve, Belgium First published on: 01 March 2010 To cite this Article Lequette, Yannick, Boels, Gauthier, Clarisse, Martine and Faille, Christine(2010) 'Using enzymes to remove biofilms of bacterial isolates sampled in the food-industry', Biofouling, 26: 4, 421 — 431, First published on: 01 March 2010 (iFirst) To link to this Article: DOI: 10.1080/08927011003699535 URL: http://dx.doi.org/10.1080/08927011003699535 Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article may be used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
Transcript of Using enzymes to remove biofilms of bacterial isolates sampled in the food-industry

PLEASE SCROLL DOWN FOR ARTICLE
This article was downloaded by: [lequette, yannick]On: 1 March 2010Access details: Access Details: [subscription number 919582326]Publisher Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
BiofoulingPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713454511
Using enzymes to remove biofilms of bacterial isolates sampled in thefood-industryYannick Lequette a; Gauthier Boels b; Martine Clarisse a; Christine Faille b
a INRA, Villeneuve d'Ascq, France b REALCO S.A., Louvain-la-Neuve, Belgium
First published on: 01 March 2010
To cite this Article Lequette, Yannick, Boels, Gauthier, Clarisse, Martine and Faille, Christine(2010) 'Using enzymes toremove biofilms of bacterial isolates sampled in the food-industry', Biofouling, 26: 4, 421 — 431, First published on: 01March 2010 (iFirst)To link to this Article: DOI: 10.1080/08927011003699535URL: http://dx.doi.org/10.1080/08927011003699535
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

Using enzymes to remove biofilms of bacterial isolates sampled in the food-industry
Yannick Lequettea*, Gauthier Boelsb, Martine Clarissea and Christine Failleb
aINRA, PIHM, UR638, Villeneuve d’Ascq, France; bREALCO S.A., Louvain-la-Neuve, Belgium
(Received 23 October 2009; final version received 9 February 2010)
The aim of this study was to analyze the cleaning efficiency of polysaccharidases and proteolytic enzymes againstbiofilms of bacterial species found in food industry processing lines and to study enzyme effects on the compositionof extracellular polymeric substances (EPS) and biofilm removal in a Clean-in-Place (CIP) procedure. The screeningof 7 proteases and polysaccharidases for removal of biofilms of 16 bacterial species was first evaluated using amicrotiter plate assay. The alkaline pH buffer removed more biofilm biomass as well as affecting a larger range ofbacterial species. The two serine proteases and a-amylase were the most efficient enzymes. Proteolytic enzymespromoted biofilm removal of a larger range of bacterial species than polysaccharidases. Using three isolates derivedfrom two bacterial species widely found in food processing lines (Pseudomonas fluorescens and the Bacillus cereusgroup), biofilms were developed on stainless steel slides and enzymatic solutions were used to remove the biofilmsusing CIP procedure. Serine proteases were more efficient in removing cells of Bacillus biofilms thanpolysaccharidases. However, polysaccharidases were more efficient in removing P. fluorescens biofilms than serineproteases. Solubilization of enzymes with a buffer containing surfactants, and dispersing and chelating agentsenhanced the efficiency of polysaccharidases and proteases respectively in removing biofilms of Bacillus andP. fluorescens. A combination of enzymes targeting several components of EPS, surfactants, dispersing and chelatingagents would be an efficient alternative to chemical cleaning agents.
Keywords: biofilm removal; enzyme cleaners; Clean-in-Place (CIP); stainless steel; food; extracellular polymericsubstances (EPS)
Introduction
Bacterial spoilage is a major concern in the foodindustry entailing both economic and public healthconsequences (Carpentier and Cerf 1993; Kumar andAnand 1998). Equipment contamination has beenreported to account for around 40% of food-bornediseases caused by bacteria in France (Haeghebaertet al. 2001). Biofilms consist of communities ofmicroorganisms attached to a surface and embeddedin self-made extracellular polymeric substances (EPS)that protect the cells. Thus, biofilms are a main factorin surface-to-food cross contamination (Sutherland2001; Perez-Rodrıguez et al. 2008). Many species ofbacteria can be isolated from these biofilms. EPScomposition in biofilms varies greatly from onebacterial species to another and is strongly affectedby growth conditions (Costerton et al. 1995; Kolterand Greenberg 2006). Studies showed EPS influencesbiofilm shape and size (Sutherland 2001), resistance toshear forces (Stoodley et al. 2002; Sharma et al. 2005;Garny et al. 2008), and resistance against antimicro-bial compounds (Stewart and Costerton 2001).Extracellular polysaccharides (e-polysaccharides) areusually the predominant component of the biofilm
matrix (Sutherland 2001). Other polymers such asproteins and DNA have also been identified in thebiofilm matrix, where they could be involved ininteractions with surfaces and cells or modify thestructural features of biofilms (Whitchurch et al.2002; Branda et al. 2005, 2006; Allesen-Holm et al.2006; Barnhart and Chapman 2006; Spoering andGilmore 2006).
Biofilms can become resistant to the chemical andphysical treatments applied during cleaning andsanitizing procedures in the food industry (Chmielews-ki and Frank 2003). Therefore, the use of enzymetreatments to break down EPS in biofilms is a possiblealternative when standard cleaning agents do not givesatisfactory results in removing biofilms. Proteases incommercially available detergents are already used toclean for example ultrafiltration units, contact lenses,medical apparatus, and laundry. Amylases are thesecond type of enzyme widely used in the formulationof enzyme detergents, mainly to remove food residuesof starch-based foods. Because of the wide hetero-geneity of EPS between bacterial species (Augustinet al. 2004), a search for new enzymes and forunderstanding the role of EPS in biofilm cohesiveness
*Corresponding author. Email: [email protected]
Biofouling
Vol. 26, No. 4, May 2010, 421–431
ISSN 0892-7014 print/ISSN 1029-2454 online
� 2010 Taylor & Francis
DOI: 10.1080/08927011003699535
http://www.informaworld.com
Downloaded By: [lequette, yannick] At: 12:13 1 March 2010

is required to improve biofilm removal during Clean-in-Place (CIP) procedures in industry.
The aim of the present study was to analyze theefficiency of polysaccharidases and proteolytic enzymesin removing biofilms developed by different bacterialspecies commonly found in food industry processinglines. Twenty five bacterial isolates sampled from foodindustry environments were used to screen sevenenzymes for their ability to remove biofilms fromfood-relevant media using a microtiter plate assay.From this screening, four enzymes were selected toanalyze the effects of enzymes in removing biofilms(cells and EPS) developed on stainless steel (SS) duringa CIP procedure.
Materials and methods
Bacterial isolates and media
The bacterial isolates are listed in the Table 1. Isolateswith a nomenclature CCL were isolated from variousindustrial origins and were from the collection of theAFSSA laboratory at Maison-Alfort, France (BrigitteCarpentier, [email protected]). Bacterial
isolates with a nomenclature D3- were from theUniversity of Cornell, Department of Food Science,Ithaca, New York14853, USA (Kathryn J. Boor,[email protected]). The D3 isolates were isolatedfrom processed milk or various surfaces from threedistinct dairy plants (Dogan and Boor 2003). Bacterialisolates with BC or PF nomenclatures were from theauthors’ collection.
Biofilms were grown in three growth media: askimmed milk medium (skimmed milk 0.11% (w/v) indistilled water), a meat extract medium (meat extractpowder (Biokar) 0.1% (w/v) in distilled water) anddiluted Trypsic Soy Broth (TSB (Biokar) 0.3% (w/v) indistilled water). Planktonic growth rates of all isolateswere monitored with a Bioscreen C microbiologicalgrowth analyzer (Thermo Labsystem).
Enzymes
The enzymes used in this study were: two serineproteases P2 and P4 (Subtilisin, Novozymes), twopapains P1 (Enzybel) and P3 (Blue Star Chemicals), 1a-amylase S1 (Novozymes). Two polysaccharidasemixes were also used in this study: mix A, whichincluded a cellulase, hemi-cellulases (Novozymes), andmix B, which included one third of mix A, one thirdof a-amylase S1 and one third of a b-glucanase(Novozymes).
Enzyme solutions were diluted 100 times in 50 mMPBS buffer (at pH 4, pH 7, and pH 10) and in Realco Bbuffer, a formulated alkaline buffer (pH 10) containinganionic surfactants with dispersing and chelatingagents (# LIQ0898, [email protected], S.A. REALCON.V, Louvain-la-Neuve, Belgium). The enzyme clean-ing solutions contained 5–10% of active proteins.
Removal of biofilms developed in microtiter plates
Biofilm formation and quantification using polystyrenemicrotiter plates were measured as previously de-scribed (O’Toole and Kolter 1998). Briefly, overnightcultures were diluted 100 times in a biofilm growthmedium and 100 ml of the suspension were loaded intomicrotiter dish wells and incubated for 24 h at 308C ina humidified atmosphere. To screen for the efficiencyof enzymes in removing biofilms, the biofilm growthmedium was gently removed, 125 ml of 458C cleaningsolution were added (458C water was used as a control)and the microtiter plates were incubated at 458C for30 min, after which the solution was gently pipettedout. Distilled water (100 ml) and 25 ml of crystal violet(1% in water) were added to each well. The microtiterplates were incubated for 15 min at room temperatureand rinsed three times with room temperature water.Crystal violet stained biofilms were solubilized in
Table 1. List of 25 bacterial isolates.
Strains Species Origin of industry
CCL488 Kocuria Cold roomCCL27a Sphingobacterium
multivorumCheese-making
premisesCCL9a Bacillus ssp. Pastry-making
premisesBC98/4 B. cereus Dairy productBCD6 B. cereus Dairy productBCD22 B. cereus Dairy productBC5832 B. cereus Dairy productCCL451a B. mycoides Cold roomCCL457 Staphylococcus xylosus Cold roomCCL15 Staphylococcus capitis Pastry-making
premisesCCL44 Staphylococcus caprae Pastry-making
premisesCCL101 Staphylococcus sciuri Processed meatCCL31 Lactococcus lactis lactis Cheese-making
premisesCCL487 Burkholderia cepacia Cold roomCCL410 Escherichia coli UnknownCT102 E. coli Dairy productCF Citrobacter freundii Dairy productCCL460 Erwinia persinius Cold roomPF22 P. fluorescens Pastry-making
premisesD3-283 P. fluorescens Dairy productD3-276 P. fluorescens Dairy productPF1a P. fluorescens CIP effluentPF22 P. fluorescens CIP effluentD3-350 P. fluorescens Dairy productCCL10 Sphingomonas spp. Dairy product
aBacterial isolates which developed a strong-adherent biofilm in twodistinct media.
422 Y. Lequette et al.
Downloaded By: [lequette, yannick] At: 12:13 1 March 2010

200 ml of 95% ethanol, and 125 ml of crystal violet-ethanol solution were used to measure absorbance at595 nm with a microplate reader (Multiskan spectrum,Thermo Labsystem). Three wells per plate wereinoculated with the same strain in each experiment.The experiments were repeated three times.
Formation and removal of biofilms developed onstainless steel slides
Biofilms were developed on AISI 316 SS slides with a2B finish (4.5 6 1.5 cm2). A 3 cm 6 1.1 cm area wasdelimited on the slides with a hydrophobic marker(Dakopen S.A.) (Sommer et al. 1999). Cell suspensions(250 ml) containing 5 6 107 cells ml71 in TSB weredeposited in the delimited area. Slides were incubatedin a humidified chamber at 308C for 2 h for celladhesion. TSB was replaced by 250 ml of a biofilmgrowth medium (TSB, skimmed milk, or meat extractmedium) and the slides were incubated for 24 h at308C. The growth medium was renewed after incuba-tion for 24 h and plates were then incubated for afurther 24 h at 308C.
To measure the efficiency of buffers and enzymesin removing biofilms in a CIP model, SS slides with 48h-old biofilms were rinsed in distilled water then slidinto square pipes connected to a CIP rig (Jullien et al.2008) and a buffer with or without enzymes wascirculated into the rig for 30 min. The temperature wasset at 458C for enzyme treatments and the controls(water and NaOH) because enzyme activities wereoptimal between 408C and 508C. A laminar flow rateof 68 l h71 (Reynolds ¼ 1500) was chosen to limit theeffect of parameters other than enzymes. The circula-tion time was set at 30 min to allow the enzymes to acton the biofilms. The number of adherent cells beforeand after treatment was determined as follows: SSslides were rinsed in sterile distilled water, inserted intoglass tubes containing 10 ml of TSB Tween 80 (0.5%)and cells were detached from the slides by applying atwo-step procedure twice: sonication for 2 min 30 (70W, 47 kHz, Bransonic) followed by 30 s of vortexing.The detached bacteria were enumerated on TrypsicSoy Agar (TSA) plates.
Extraction, purification, and quantification of EPS ofthe biofilm matrix
EPS were purified from biofilms before and aftertreatments. After growth of biofilms for 48 h, thegrowth medium was pipetted out and replaced by250 ml of preheated buffer at 458C with or withoutenzymes. SS slides were incubated at 458C for 30 minthen rinsed in sterile distilled water. Biofilms werescraped off the SS slides using MicroAlpha swabs
(Texwipe) and resuspended in 1.5 ml of NaCl 0.9% byvortexing (30 s) (Priester et al. 2006). Biofilms fromfour slides were gathered for protein analysis or fromthree slides for polysaccharide analysis. An aliquot of200 ml was collected from biofilm suspensions for totalprotein quantification. Tubes were then centrifuged30 min at 14,000g to pellet cells and the supernatantfluids were collected. The pellet was resuspended in0.4 ml of 0.9% NaCl, sonicated for 30 s, vortexed for30 s and then tubes were centrifuged for 30 min at14,000g. Supernatant fluids were then combined withthe supernatant of the first extraction. Samples of EPSwere dried using a speed vacuum centrifuge. Distilledwater (50 ml) was added to the pellets before heatingfor 10 min at 608C and the pellets were then solubilizedby pipetting. Distinct purification steps were carriedout to extract proteins or polysaccharides from theEPS. (I) E-proteins were directly titrated from super-natant fluids using the MicroBCA protein assay kit(Pierce) and analyzed by electrophoresis. Aliquotscontaining 30 mg of e-proteins were dried out andsolubilized in electrophoresis migration buffer(TrisHCl, 125 mM; pH 6.8, SDS 4%; b-mercaptoetha-nol 10%; DTT 1mM; glycerol 10% and bromophenolblue 0.05%). Samples were heated for 2 min at 958Cjust before loading and proteins were separated bySDS-PAGE (stacking gel 4%, separation gel 12%,migration buffer: Tris HCl 25 mM, glycine 192 mM,SDS 0.1%, pH 8.6). (II) E-polysaccharides werepurified as follows: 5 ml of DNAse I (1000 U ml71,Biolabs) and 1 ml of RNAse A solution (7000 U ml71,Qiagen) were added to supernatants and incubated for1 h at 378C to degrade DNA and RNA. Then, 10 ml ofproteinase K (200 mg ml71, MP Biomedicals) wasadded and samples were incubated overnight forprotein degradation. E-polysaccharides were purifiedby hot-phenol extraction (Westphal and Jann 1965).Then, the e-polysaccharides were dialyzed againstdistilled water for 12 h using dialysis cellulose-tubingaccording to the manufacturer’s instructions (CelluSep H1, 1000 Da pores). The total carbohydrateconcentrations of the e-polysaccharides were measuredusing phenol-sulfuric assay (Dubois et al. 1951) andglucose as a standard.
Statistical analyses
Correlations were performed using the Pearson meth-ods and two-tailed p-values.
Results
Role of pH in biofilm removal
As the environmental conditions have been shown toaffect biofilm structure and resistance (Sharma et al.
Biofouling 423
Downloaded By: [lequette, yannick] At: 12:13 1 March 2010

2005; Kolter and Greenberg 2006; Garny et al. 2008),biofilms of 25 isolates (Table 1) recovered fromindustrial environments were developed in differentmedia: two food-relevant media (skimmed milk andmeat extracts) and TSB, a commonly used medium togrow biofilms in the laboratory. Twenty eight biofilms:8 in TSB, 8 in skimmed milk, and 12 in meat extractmedium were chosen for their resistance to a rinsingstep using water at 458C.
Before screening enzymes for biofilm removal, theinfluence of pH in removing biofilms was first tested.Phosphate buffer (PBS) at 458C at pH 4, pH 7, and pH10 was used to remove biofilms in microtiter plateassays (Figure 1). Water at 458C was used as control.First, acidic PBS was inefficient in removing biofilms:only two biofilms were totally removed. The growthmedia appeared to play a predominant role in biofilmremoval. Indeed, all biofilms grown in skimmed milkwere largely removed with alkaline and neutral PBSbuffers, and biofilms developed in TSB and meatextract medium were poorly removed by any of thePBS buffers: Only one biofilm developed in meatextract was totally removed with alkaline PBS buffer(Supplemental Table S1). Conversely, biofilm removalusing PBS was not related to the bacterial species.
Whether the percentage of biofilm removal wascorrelated to the amount of the initial biofilm was alsoinvestigated. When biofilms were treated with neutraland alkaline PBS, data from biofilms developed inskimmed milk were excluded because biofilms weretotally removed by this treatment. Regardless of theisolate species and the growth medium, the ease ofremoving biofilms was significantly correlated with theamount of initial biofilm whatever the PBS treatment(R ¼ 0.7641 and p 5 0.0001 at pH 10, R ¼ 0.6615and p ¼ 0.0015 at pH 7, R ¼ 0.8741 and p 5 0.0001at pH 4): the higher the initial biofilm amount, themore difficult it was to remove them.
Because alkaline PBS was the most efficienttreatment to promote biofilm removal, further experi-ments were performed using buffers at pH 10. Aformulated buffer, Realco B, containing anionicsurfactant, dispersing and chelating agents was com-pared to PBS at pH 10. Realco B promoted the totalremoval of four additional biofilms as compared to thePBS at pH 10 (Figure 1 and Supplemental Table S1).The efficacy in removing biofilms was not significantlycorrelated with the amount of biofilm removed usingRealco B (p value ¼ 0.4982, R ¼ 0.0422).
Screening for enzymes to remove biofilms developed inmicrotiter plates
Because the biofilms developed in skimmed milkwere totally removed by the buffer alone, they were
Figure 1. Quantification of biofilm removal after treatmentwith phosphate buffers at different pH using a microtiter platesassay. Eight isolates grew strongly on attached biofilms in TSB(A); 8 isolates in skimmed milk medium (B); 12 isolates in meatextract medium (C). OD at 595 nm of biofilms before and aftertreatmentwithwater, PBS buffers at pH4, 7, or 10 andRealco Bbuffers at 458Cfor 30 minwere plotted.Theblack lines representthe mean of OD at 595 nm.
424 Y. Lequette et al.
Downloaded By: [lequette, yannick] At: 12:13 1 March 2010
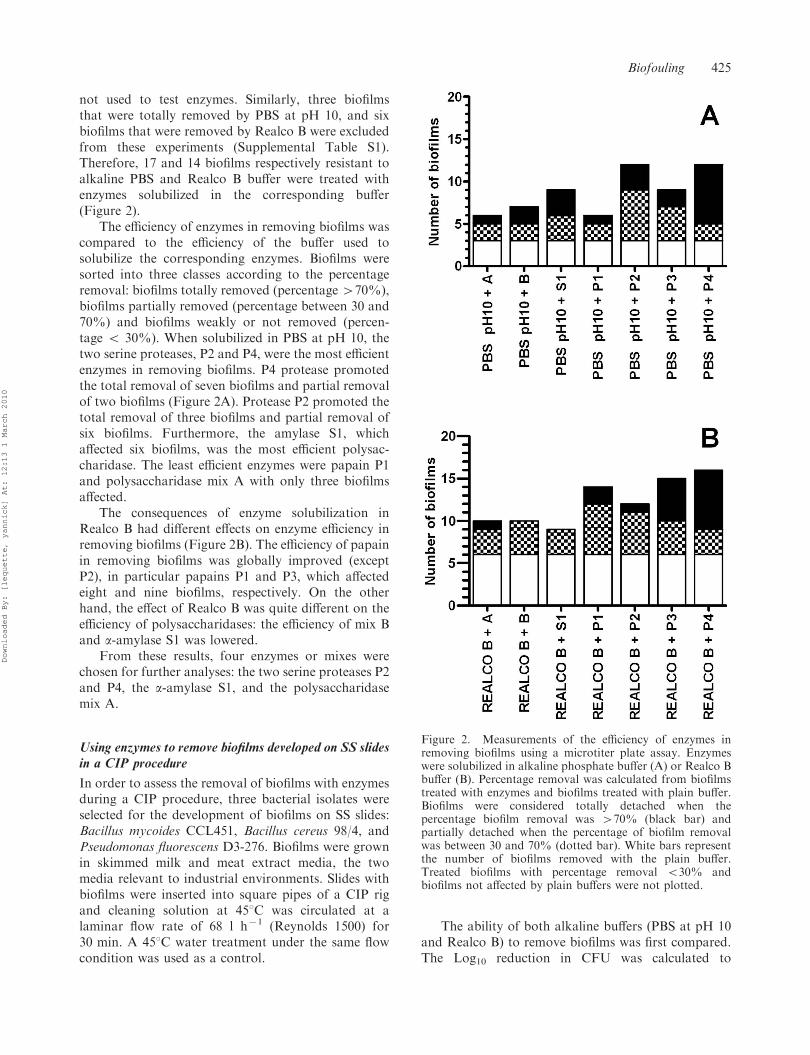
not used to test enzymes. Similarly, three biofilmsthat were totally removed by PBS at pH 10, and sixbiofilms that were removed by Realco B were excludedfrom these experiments (Supplemental Table S1).Therefore, 17 and 14 biofilms respectively resistant toalkaline PBS and Realco B buffer were treated withenzymes solubilized in the corresponding buffer(Figure 2).
The efficiency of enzymes in removing biofilms wascompared to the efficiency of the buffer used tosolubilize the corresponding enzymes. Biofilms weresorted into three classes according to the percentageremoval: biofilms totally removed (percentage 470%),biofilms partially removed (percentage between 30 and70%) and biofilms weakly or not removed (percen-tage 5 30%). When solubilized in PBS at pH 10, thetwo serine proteases, P2 and P4, were the most efficientenzymes in removing biofilms. P4 protease promotedthe total removal of seven biofilms and partial removalof two biofilms (Figure 2A). Protease P2 promoted thetotal removal of three biofilms and partial removal ofsix biofilms. Furthermore, the amylase S1, whichaffected six biofilms, was the most efficient polysac-charidase. The least efficient enzymes were papain P1and polysaccharidase mix A with only three biofilmsaffected.
The consequences of enzyme solubilization inRealco B had different effects on enzyme efficiency inremoving biofilms (Figure 2B). The efficiency of papainin removing biofilms was globally improved (exceptP2), in particular papains P1 and P3, which affectedeight and nine biofilms, respectively. On the otherhand, the effect of Realco B was quite different on theefficiency of polysaccharidases: the efficiency of mix Band a-amylase S1 was lowered.
From these results, four enzymes or mixes werechosen for further analyses: the two serine proteases P2and P4, the a-amylase S1, and the polysaccharidasemix A.
Using enzymes to remove biofilms developed on SS slidesin a CIP procedure
In order to assess the removal of biofilms with enzymesduring a CIP procedure, three bacterial isolates wereselected for the development of biofilms on SS slides:Bacillus mycoides CCL451, Bacillus cereus 98/4, andPseudomonas fluorescens D3-276. Biofilms were grownin skimmed milk and meat extract media, the twomedia relevant to industrial environments. Slides withbiofilms were inserted into square pipes of a CIP rigand cleaning solution at 458C was circulated at alaminar flow rate of 68 l h71 (Reynolds 1500) for30 min. A 458C water treatment under the same flowcondition was used as a control.
The ability of both alkaline buffers (PBS at pH 10and Realco B) to remove biofilms was first compared.The Log10 reduction in CFU was calculated to
Figure 2. Measurements of the efficiency of enzymes inremoving biofilms using a microtiter plate assay. Enzymeswere solubilized in alkaline phosphate buffer (A) or Realco Bbuffer (B). Percentage removal was calculated from biofilmstreated with enzymes and biofilms treated with plain buffer.Biofilms were considered totally detached when thepercentage biofilm removal was 470% (black bar) andpartially detached when the percentage of biofilm removalwas between 30 and 70% (dotted bar). White bars representthe number of biofilms removed with the plain buffer.Treated biofilms with percentage removal 530% andbiofilms not affected by plain buffers were not plotted.
Biofouling 425
Downloaded By: [lequette, yannick] At: 12:13 1 March 2010
measure the efficiency of the different treatments(Table 2). The use of PBS at pH 10 under dynamicconditions did not significantly improve biofilmremoval (51 log10 reduction in CFU) compared towater alone, except for P. fluorescens D3-276 biofilmsdeveloped in skimmed milk medium (3.2 log10 reduc-tion in CFU) (data are not shown). The efficiencyof Realco B was clearly depended on the growthmedium. When compared to results obtained with PBSpH 10, the removal was always more efficient forbiofilms developed in meat extract medium comparedto biofilms developed in skimmed milk medium(Table 2).
The percentages of biofilm removal promoted byenzymes solubilized in PBS at pH 10 or Realco B werethen compared to the PBS treatment. When solubilizedin PBS at pH 10, the type of enzymes efficient inremoving biofilms was affected by both the strain andthe growth medium (Table 2). A clear increase in thebiofilm removal level was observed with proteasetreatments on CCL451 biofilms grown in both meat(P4) and in skimmed milk (P2 and P4) media andBC98/4 biofilms grown in meat extract medium (P4).Conversely, proteases were poorly effective in remov-ing P. fluorescens D3-276 biofilms. Whilst the poly-saccharidases were poorly effective in removingBacillus biofilms, they were the most efficient inremoving biofilms of P. fluorescens D3-276 grown inmeat extract media (a-amylase S1 and mix A) and inskimmed milk (a-amylase S1). None of the enzymescould significantly remove BC98/4 biofilms developedin skimmed milk.
Due to the greater efficiency of Realco B alone inremoving biofilms, the biofilm reduction due toenzymes appeared less important. Solubilization of
the serine proteases in Realco B was the only treatmentthat improved the removal of P. fluorescens D3-276(P2) and B. cereus BC98/4 biofilms (P2 þ P4) devel-oped in meat extract medium while polysaccharideswere the most efficient treatment in removing P.fluorescens D3-276 (mix A) and both Bacillus biofilms(a-amylase S1) developed in skimmed milk medium(Table 2).
Enzyme treatment was then compared to treat-ments with NaOH, widely used in CIP procedures.Two NaOH concentrations were tested: 0.05% (pH9.5) and 0.5% (pH 12.5) at 458C for 30 min. The use of0.05% NaOH was less efficient than treatments usingbuffers with or without enzymes except for CCL451biofilm removal (Table 2). NaOH at 0.5% was moreefficient in removing biofilms developed in meat extractmedium, but NaOH at 0.5% showed variable efficiencycompared to the best enzyme treatment. NaOH wasless efficient on B. cereus BC98/4 biofilms, moreefficient on B. mycoides CCL451 biofilms and equallyefficient on P. fluorescens D3-276 biofilms.
Characterization of enzyme effects on EPS of biofilms
Biofilms were treated with buffers and with differentenzymes to further understand the effects of enzymeson EPS and in removing biofilms. Biofilms of the threeisolates B. mycoides CCL451, P. fluorescens D3-276,and B. cereus BC98/4 were grown in skimmed milk andmeat extract media, and the EPS of the biofilms werequantified before and after different enzyme treat-ments. The swabbing method was chosen as inprevious studies (Joseph et al. 2001) to limit thebreak-down of EPS, cell damage and lyses during thecollecting of biofilms (Assere et al. 2008). Visualization
Table 2. Biofilm removal estimated by the log10 reduction in CFU after treatment in a CIP procedure.
Log10 reduction of CFU after CIPa
D3-276 CCL451 BC98/4
Treatments Meat Skimmed milk Meat Skimmed milk Meat Skimmed milk
PBS pH10a
(residual CFU)(7.3 6 10þ7) +
1.3 6 10þ7(1.7 6 10þ8) +
6.5 6 10þ7(5.6 6 10þ7) +
3.5 6 10þ7(2.7 6 10þ8) +
5.9 6 10þ7(1.2 6 10þ4) +
1.7 6 10þ3(7.0 6 10þ3) +
4.0 6 10þ3
PBS pH10 þ P2 0.93 0.44 1.08 2.71 0.03 0.92PBS pH10 þ P4 0.88 0.03 2.04 3.05 1.24 0.55PBS pH10 þ A 1.69 0.00 1.14 0.01 0.05 0.08PBS pH10 þ S1 1.39 1.27 0.83 0.07 0.08 0.30Realco B 1.56 0.65 2.54 0.51 0.34 70.30Realco B þ P2 3.87 0.78 2.57 0.94 2.25 0.56Realco B þ P4 1.57 0.69 2.55 1.06 1.13 0.22Realco B þ A 1.57 1.88 2.73 0.73 0.58 0.00Realco B þ S1 1.60 0.65 2.98 1.24 0.43 1.32NaOH 0.05% 1.44 70.19 3.57 1.38 70.27 70.42NaOH 0.5% 4.06 1.60 3.70 2.11 1.42 70.12
aCell counts cm72 after PBS pH10 treatment was taken as the reference for each condition to calculate the log10 reduction in CFU.
426 Y. Lequette et al.
Downloaded By: [lequette, yannick] At: 12:13 1 March 2010
of the remaining materials after swabbing showed onlya few scattered cells remaining on the slides. E-polysaccharides were analyzed to evaluate the role ofmix A and a-amylase and e-proteins to evaluate therole of proteases P2 and P4.
For the two polysaccharidases solubilized in PBSat pH 10, the a-amylase S1 reduced more e-poly-saccharides in all biofilms than mix A (Table 3), exceptfor the D3-276 biofilms developed in skimmed milkmedium. Realco B alone was able to remove e-polysaccharides in half of the tested biofilms ascompared to PBS pH 10. When enzymes weresolubilized in Realco B buffer, mix A promoted betterremoval of e-polysaccharides in all biofilms than a-amylase S1 (Table 3). The efficiency of e-polysacchar-ide removal by polysaccharidases was better inbiofilms developed in meat extract medium exceptwith the a-amylase S1 treatment of B. mycoidesCCL451 biofilms. Then, treatments using polysacchar-idases solubilized in PBS at pH 10 or Realco B bufferswere compared with NaOH treatments in removing e-polysaccharides. NaOH (0.05%) was globally aseffective as Realco B, and consequently more effectivethan PBS pH 10. Increasing the NaOH concentrationdid not significantly increase EPS removal. As a result,
better EPS removal was obtained when polysacchar-idases were added to Realco B.
A similar analysis was performed on e-proteins.PBS at pH 10 and Realco B buffers could partiallyremove e-proteins of P. fluorescens D3-276 andB. mycoides CCL451 biofilms but not e-proteins ofB. cereus BC98/4 biofilms (Table 4). Proteases presentin the enzyme buffers biased the e-protein titration,therefore, the effects of serine proteases P2 and P4 onthe degradation of e-proteins were investigated bySDS-PAGE. When serine proteases P4 and P2 wereadded to PBS at pH 10 or Realco B, no detectable e-proteins from P. fluorescens D3-276 and B. cereusBC98/4 biofilms developed in either media wereobserved on electrophoresis gel (Supplemental FigureS1). Only bands corresponding to the serine proteaseswere observed on gels. However, a few high molecularweight proteins were still observed on gels of e-proteinsof B. mycoides CCL451 biofilms treated with proteasesP4 and P2, suggesting a partial degradation ofe-proteins (Supplemental Figure S1). Partial degrada-tion of e-proteins was observed in all biofilms when106 diluted serine proteases P2 and P4 solutions wereused, showing e-protein degradation was proteasedose-dependent. As a control, no detectable e-proteins
Table 3. Biofilm removal estimated by the ratio of reduction in residual extracellular polysaccharides after treatment.
Fold changea
D3-276 CCL451 BC98/4
Treatments Meat Skimmed milk Meat Skimmed milk Meat Skimmed milk
PBS pH10a 1.00 1.00 1.00 1.00 1.00 1.00PBS pH10 þ A 1.53 1.13 0.97 1.08 1.13 2.22PBS pH10 þ S1 2.82 0.94 1.58 1.21 1.63 2.30Realco B 1.59 0.94 1.03 0.98 3.10 1.45Realco B þ A 3.69 1.64 1.50 1.13 14.6 2.67Realco B þ S1 3.07 0.95 1.01 1.12 3.68 1.58NaOH 0.05 1.5 0.97 1.35 1.30 3.13 0.90NaOH 0.5 1.74 1.05 1.50 1.07 2.42 1.12
aPolysaccharide weight after PBS pH10 treatment was taken as the reference for each condition, and fold changes in e-polysaccharide quantitiesafter other treatments were calculated.
Table 4. Quantification of extracellular proteins present in EPS.
Quantification of e-proteins (mg e-proteins mg71 of total proteins)a
D3-276 CCL451 BC98/4
Treatments Meat Skimmed milk Meat Skimmed milk Meat Skimmed milk
Native biofilm 152 + 20 132 + 3 120 + 16 138 + 18 328 + 42 137 + 7PBS pH10 70 + 7 103 + 11 75 + 28 137 7 339 + 18 112 + 14Realco B 39 + 18 217 + 56 86 + 5 92 14 230 + 38 114 + 12NaOH 0.05% 110 + 31 169 + 8 544 + 20 253 37 280 + 44 180 + 5NaOH 0.5% 66 + 4 385 + 3 210 + 5 361 19 404 + 19 320 + 10
aExtracellular proteins are given in mg of extracellular proteins divided by mg of total proteins + SD. Total proteins in whole biofilms werequantified from aliquots containing whole biofilms (matrix and bacterial cells) (see materials and methods).
Biofouling 427
Downloaded By: [lequette, yannick] At: 12:13 1 March 2010

were observed on electrophoresis gels when 106 or106 serine protease solutions were used on purifiede-proteins (data are not shown). NaOH at 0.05% and0.5% was less efficient in removing e-proteins thanenzyme treatments containing surfactants, dispersingand chelating agents, except for the P. fluorescens D3-276 biofilms grown in meat extract medium. Indeed,the use of 0.5% NaOH induced an increase in therelative amount of e-proteins except for the P.fluorescens D3-276 biofilms grown in meat extractmedium.
Discussion
The presence of bacterial biofilms in food processinglines is a great concern for the food industry. Chemicalsolutions such as NaOH and acids are commonly usedin CIP procedures. In some cases, these procedures arenot always sufficient for removing cells and EPS ofbiofilms (Antoniou and Frank 2005). Therefore, theidea of using enzymes to break down the EPS networkand remove biofilms became an interesting alternative(Xavier et al. 2005; de Carvalho 2007). Here, enzymeswere screened for their ability to remove biofilms ofdifferent bacterial species, and developed in differentgrowth media. Because biofilms of the selectedbacterial isolates synthesized EPS containing mostlypolysaccharides and proteins in the experimentalconditions used, different kinds of polysaccharidasesand proteases were selected.
The role of pH on the stability of the 28 biofilmsformed by 12 different bacterial species was investi-gated in microtiter plate assays. All biofilms developedin skimmed milk medium were totally removed byneutral and alkaline pH buffer. The composition of thefood matrix is known to influence the nature of EPS inbiofilms, modulating biofilm cohesiveness, its resistanceto shear stress and the cleaning efficiency (Chmielewskiand Frank 2003; Gram et al. 2007; Herrera et al. 2007).Previous studies have shown that milk reduced adhe-sion of bacteria and formation of biofilms on SS (Helkeet al. 1993). The inhibitor effect of milk on biofilmformation could explain why the present biofilmsdeveloped in milk were all also removed with neutraland alkaline pH in the microtiter plate.
The alkaline buffer removed more biomass as wellas affecting a larger range of bacterial species,including Bacillus, Pseudomonas, and Staphylococcusspecies in these experiments. Previous studies showedthat alkaline cleaners were effective in removingBacillus biofilms, but poorly effective in removingPseudomonas and Staphylococcus biofilms (Chmielews-ki and Frank 2003). An alkaline pH is known to induceswelling of polymers in protein deposits in the foodindustry and of EPS (Dogsa et al. 2005; Fryer and
Asteriadou 2009). The higher solubility of EPS and theswelling of the EPS network at alkaline pH couldreduce biofilm cohesiveness and facilitate the removalof biofilms as was observed. On the other hand, thedense network of EPS and their insolubility at acidicpH (Dogsa et al. 2005) could increase the compactnessof biofilms, making them more resistant to cleaning.Alkaline cleaners would be a preferred choice forroutine cleaning procedures and the alkaline pH bufferwas thus used to screen enzymes for biofilm removal inthis study.
In the microtiter plate model, the two serineproteases removed biofilms of a larger range ofbacterial species. The amylase S1 totally removed thebiofilms of three Pseudomonas strains. To furtherunderstand the effects of enzymes on biofilm removal,the effects of two serine proteases, amylase S1, and apolysaccharidases mix A were selected from screeningin microtiter plates to measure the removal of biofilms(EPS and cells) developed on SS slides in a CIPprocedure. The efficacy of enzymes depended on thebacterial species: proteases were more efficient thanpolysaccharidases in removing Bacillus biofilms, whilepolysaccharide-degrading enzymes were more efficientin removing P. fluorescens biofilms. In this work, theefficiency of polysaccharidase mix A (containing a- andb-polysaccharidases) and aa-amylase S1 in removingcells of P. fluorescens biofilms suggested that both a-and b-linked polysaccharides contributed to biofilmcohesiveness, as previously shown (Friedman andKolter 2004; Branda et al. 2005; Itoh et al. 2005). Thedegradation of e-proteins in all biofilms was clearlyshown when using proteases, but no clear relationshipsbetween the biofilm removal rate and e-proteindegradation were observed: serine proteases weremore efficient in removing cells of Bacillus biofilmsthan polysaccharidases, but the protein degradationwas incomplete in CCL451 biofilms. These resultssuggest that proteases targeting key proteins involvedin biofilm cohesiveness of Bacillus biofilms (Brandaet al. 2006) would be sufficient to remove biofilms.
When serine proteases were combined with anionicsurfactants, dispersing and chelating agents, theremoval of cells was enhanced in P. fluorescens biofilmsdeveloped in meat extract medium during CIP,showing proteins also contributed to the cohesivenessof P. fluorescens biofilms as previously suggested(Hinsa and O’Toole 2006). On the other hand, theaddition to enzymes of anionic surfactants withdispersing and chelating activities showed that a-amylase S1 was the most efficient enzyme treatment inremoving Bacillus biofilms, except for the B. cereusBC98/4 biofilm developed in meat extract medium(where serine protease P4 treatment was the most effi-cient treatment). The dispersal properties of surfactants
428 Y. Lequette et al.
Downloaded By: [lequette, yannick] At: 12:13 1 March 2010

are known to disrupt the hydrophobic interactionsinvolved in cross-linking the biofilm matrix. Chelatingagents also reduce the strength of alginate, the viscosityof biofilm mixture and promote cell detachment (Chenand Stewart 2000; Xavier et al. 2005), facilitating accessto and increasing the affinity of enzymes to theirsubstrates (Singh et al. 2007). As reported by Flemminget al. (2007), an increasing number of components havebeen identified in EPS, but a lot are as yet to beidentified and information about their localization andstability requires further investigation. Here, the resultssuggest that e-polysaccharides in P. fluorescens biofilmscould be directly accessible to polysaccharidases, whilee-proteins involved in biofilm cohesiveness would belocated deeper in the biofilms, and surfactants, disper-sing and chelating agents had to be combined to serineprotease for them to access their substrate. However,serine protease could directly access e-proteins inBacillus biofilm, while polysaccharidases would needthe help of surfactants, dispersing and chelating agentsto access those e-polysaccharides involved in biofilmcohesiveness.
Most biofilms developed in meat extract werebetter removed than skimmed milk biofilms whenenzymes were used in CIP procedures. Higher amountsof EPS, the substrate of enzymes, would explain abetter efficiency of enzyme cleaners in removingbiofilms developed in meat extract. This hypothesiswas verified for the P. fluorescens D3-276 and B. cereusBC98/4 biofilms, but for the B. mycoides CCL451biofilm. Biofilm growth conditions are known to applya genetic selection for biofilm formers (Kolter andGreenberg 2006). Here, it is believed that the resistanceof biofilms to enzymes, cleaners and to alkaline pHobserved above was rather due to the geneticcharacteristics of individual isolates rather than tothe general ability of a bacterial species. The specificadaptations of each isolate would be difficult to pointout by only measuring the average amount of EPScompounds and could explain why a few contradictoryresults were observed when the influences of the foodmatrix and the enzyme species-specificity in removingbiofilms were analyzed. A genomic analysis of theseisolates would allow the identification of these specificadaptations. The type of linkages in polymers andforces involved in interactions between EPS com-pounds, cells and surfaces would also be pertinentparameters in correlating the cleaning efficiency withthe biofilm cohesiveness. Atomic force microscopy, x-ray, and flow velocity to remove biofilms are methodsthat should help investigate these forces and theirs rolein cohesiveness and biofilm removal (Dogsa et al. 2005;Ahimou et al. 2007; Mohle et al. 2007).
When the results from the microtiter plate assayswere compared with biofilms developed on SS, some
discrepancies were observed. In the microtiter plateassays, biofilms developed in skimmed milk were alleasily removed while biofilms developed on SS slides inskimmed milk could be more resistant to removal thanbiofilms developed in meat extract after a CIPprocedure. Biofilms in the microtiter plate assaydeveloped at the interface of a liquid, surfaces andair, located on the wall of the wells and at the surfaceof the liquid (O’Toole and Kolter 1998; Wijman et al.2007), while biofilms on the SS slides were totallyimmerged in the growth media. These differenceswould partially explain the discrepancies between thetwo models. Methods to measure biofilm biomasswould also influence the data. Crystal violet stainedboth cells and EPS in the microtiter plate assay, whileonly cell counts were taken into account to measurebiomass removal of biofilms developed on SS. A recentreport showed ATP quantification would be a bettermethod for correlation with viable cells in biofilms(Sule et al. 2009). Therefore, the use of ATPmeasurement in biofilms would be a pertinent choicefor determining the number of viable cells in biofilms.As EPS and cells both contribute to biofilm features,the crystal violet stain would still be a valuable choicein analyzing the role of the total biomass in biofilmformation and cohesiveness.
The efficacy of enzyme treatments was thencompared to NaOH treatments, a cleaner widelyused in current CIP procedures in food industries.NaOH is well known to promote the removal ofproteins and carbohydrates in a food matrix (Chisti1999). It also weakens the matrix of biofilms, increas-ing the sensitivity of biofilms to hydrodynamic stresses(Simoes et al. 2005). The present results show that cellswere similarly removed by NaOH and enzyme treat-ments. However, NaOH did not remove EPS asefficiently as enzyme treatments, suggesting that cellswere effectively detached by NaOH, while the EPS ofthe matrix partly remained on the surface unlike withthe enzyme treatments. A report by Li and Elimelech(2004) suggested that NaOH alone was not able toreduce the interaction between organic materials, and acombination with chelating and surface acting agentswere necessary. High temperature and a high concen-tration of NaOH are also required to remove biofilms(Antoniou and Frank 2005) and this could explain whyNaOH was poorly efficient in the conditions optimalfor enzymes.
Conclusions
The application of enzymes for the control of bacterialbiofilms in food environments would provide aninteresting alternative when the classical CIP treat-ments involving chemical agents do not give
Biofouling 429
Downloaded By: [lequette, yannick] At: 12:13 1 March 2010

satisfactory results in terms of hygiene. Resultssuggested enzymes would be more suitable for in-depth cleaning of both cells and the EPS of biofilms.Using enzymes would also help to reduce the use ofchemical agents, water consumption, and energy cost.A combination of enzymes targeting several compo-nents of EPS (such as proteases and polysaccharide-degrading enzymes) is required to degrade the varioustypes of EPS and increase the range of action of anenzymatic cleaner against multi-species biofilms. Here,the roles of nutrients, bacterial species and EPS onbiofilm removal were investigated using enzymes, aswell as several other parameters such as temperature,and flow velocity during biofilm formation. Thepresence of several types of bacterial strains inbiofilms, and the parameters of the CIP procedure(temperature, turbulence of the flow, time of cleaning)should be considered to fully understand biofilmremoval (Chmielewski and Frank 2003). The identifi-cation of key components involved in biofilm cohe-siveness and the parameters inhibiting or enhancingthe actions of enzymes in removing biofilms wouldboth help to improve enzymatic CIP procedures andthe formulation of enzyme cleaners.
Acknowledgements
This study was financed by the ‘Agence Nationale de laRecherche’ under the ‘EUREKA’ Project ‘2006 EUKA00301’. The authors are also grateful to J. Six and L.Wauquier for technical assistance in building the CIP rig andGwendoline Desailly, Marjorie Danguien, Elise Dozinel,Emma Ben Samou and Aline Delhoute for laboratoryassistance.
References
Ahimou F, Semmens MJ, Novak PJ, Haugstad G. 2007.Biofilm cohesiveness measurement using a novel atomicforce microscopy methodology. Appl Environ Microbiol73:2897–2904.
Allesen-Holm M, Barken KB, Yang L, Klausen M, Webb JS,Kjelleberg S, Molin S, Givskov M, Tolker-Nielsen T.2006. A characterization of DNA release in Pseudomonasaeruginosa cultures and biofilms. Mol Microbiol 59:1114–1128.
Antoniou K, Frank JF. 2005. Removal of Pseudomonasputida biofilm and associated extracellular polymericsubstances from stainless steel by alkali cleaning. J FoodProt 68:277–281.
Assere A, Oulahal N, Carpentier B. 2008. Comparativeevaluation of methods for counting surviving biofilmcells adhering to a polyvinyl chloride surface exposed tochlorine or drying. J Appl Microbiol 104:1692–1702.
Augustin M, Ali-Vehmas T, Atroshi F. 2004. Assessment ofenzymatic cleaning agents and disinfectants againstbacterial biofilms. J Pharm Pharmacol Sci 7:55–64.
Barnhart MM, Chapman MR. 2006. Curli biogenesis andfunction. Annu Rev Microbiol 60:131–147.
Branda SS, Vik S, Friedman L, Kolter R. 2005. Biofilms: thematrix revisited. Trends Microbiol 13:20–26.
Branda SS, Chu F, Kearns DB, Losick R, Kolter R. 2006. Amajor protein component of the Bacillus subtilis biofilmmatrix. Mol Microbiol 59:1229–1238.
Carpentier B, Cerf O. 1993. Biofilms and their consequences,with particular reference to hygiene in the food industry.J Appl Bacteriol 75:499–511.
Chen X, Stewart PS. 2000. Biofilm removal caused bychemical treatments. Water Res 34:4229–4233.
Chisti Y. 1999. Modern systems of plant cleaning. In:Robinson RK, Batt CA, Patel PD, editors. Encyclopediaof food microbiology. London: Academic Press. p. 1086–1815.
Chmielewski RAN, Frank JF. 2003. Biofilm formation andcontrol in food processing facilities. Compr Rev Food SciFood Safety 2:22–32.
Costerton JW, Lewandowski Z, Caldwell DE, Korber DR,Lappin-Scott H. 1995. Microbial biofilms. Annu RevMicrobiol 49:711–745.
de Carvalho CCRC. 2007. Biofilms: recent developments onan old battle. Recent Pat Biotechnol 1:49–57.
Dogan B, Boor KJ. 2003. Genetic diversity and spoilagepotentials among Pseudomonas spp. isolated from fluidmilk products and dairy processing plants. Appl EnvironMicrobiol 69:130–138.
Dogsa I, Kriechbaum M, Stopar D, Laggner P. 2005.Structure of bacterial extracellular polymeric substancesat different pH values as determined by SAXS. Biophys J89:2711–2720.
Dubois M, Gilles K, Hamilton JK, Rebers PA, Smith F.1951. A colorimetric method for the determination ofsugars. Nature 168:167.
Flemming H-C, Neu TR, Wozniak DJ. 2007. The EPSmatrix: the ‘‘house of biofilm cells’’. J Bacteriol 189:7945–7947.
Friedman L, Kolter R. 2004. Genes involved in matrixformation in Pseudomonas aeruginosa PA14 biofilms.Mol Microbiol 51:675–690.
Fryer PJ, Asteriadou K. 2009. A prototype cleaning map: aclassification of industrial cleaning processes. TrendsFood Sci Technol 20:255–262.
Garny K, Horn H, Neu TR. 2008. Interaction betweenbiofilm development, structure and detachment in rotat-ing annular reactors. Bioproc Biosyst Eng 31:619–629.
Gram L, Bagge-Ravn D, Ng YY, Gymoese P, Vogel BF.2007. Influence of food soiling matrix on cleaning anddisinfection efficiency on surface attached Listeria mono-cytogenes. Food Control 18:1165–1171.
Haeghebaert S, Le Querrec F, Vaillant V, Astagneau ED,Bouvet P. 2001. Food poisoning incidents in France in1998. Bull Epidemiol Hebdomad 65–70.
Helke DM, Somers EB, Wong ACL. 1993. Attachment ofListeria monocytogenes and Salmonella typhimurium tostainless steel and Buna-N in the presence of milk andindividual milk components. J Food Prot 56:479–484.
Herrera JJR, Cabo ML, Gonzalez A, Pazos I, Pastoriza L.2007. Adhesion and detachment kinetics of severalstrains of Staphylococcus aureus subsp aureus underthree different experimental conditions. Food Microbiol24:585–591.
Hinsa SM, O’Toole GA. 2006. Biofilm formation byPseudomonas fluorescens WCS365: a role for LapD.Microbiology 152:1375–1383.
Itoh Y, Wang X, Hinnebusch BJ, Preston JF, III, Romeo T.2005. Depolymerization of b-1,6-N-acetyl-D-glucosaminedisrupts the integrity of diverse bacterial biofilms. JBacteriol 187:382–387.
430 Y. Lequette et al.
Downloaded By: [lequette, yannick] At: 12:13 1 March 2010

Joseph B, Otta SK, Karunasagar I, Karunasagar I. 2001.Biofilm formation by Salmonella spp. on food contactsurfaces and their sensitivity to sanitizers. Int J FoodMicrobiol 64:367–372.
Jullien C, Benezech T, Le Gentil C, Boulange-Petermann L,Dubois PE, Tissier JP, Traisnel M, Faille C. 2008.Physico-chemical and hygienic property modifications ofstainless steel surfaces induced by conditioning with foodand detergent. Biofouling 24:163–172.
Kolter R, Greenberg EP. 2006. Microbial sciences: thesuperficial life of microbes. Nature 441:300–302.
Kumar CG, Anand SK. 1998. Significance of microbialbiofilms in food industry: a review. Int J Food Microbiol42:9–27.
Li QL, Elimelech M. 2004. Organic fouling and chemicalcleaning of nanofiltration membranes: measurementsand mechanisms. Environ Sci Technol 38:4683–4693.
Mohle RB, Langemann T, Haesner M, Augustin W, SchollS, Neu TR, Hempel DC, Horn H. 2007. Structure andshear strength of microbial biofilms as determined withconfocal laser scanning microscopy and fluid dynamicgauging using a novel rotating disc biofilm reactor.Biotechnol Bioeng 98:747–755.
O’Toole GA, Kolter R. 1998. Initiation of biofilm formationin Pseudomonas fluorescens WCS365 proceeds via multi-ple, convergent signalling pathways: a genetic analysis.Mol Microbiol 28:449–461.
Perez-Rodrıguez F, Valero A, Carrasco E, Garcıa RM,Zurera G. 2008. Understanding and modelling bacterialtransfer to foods: a review. Trends Food Sci Technol19:131–144.
Priester JH, Olson SG, Webb SM, Neu MP, Hersman LE,Holden PA. 2006. Enhanced exopolymer productionand chromium stabilization in Pseudomonas putidaunsaturated biofilms. Appl Environ Microbiol 72:1988–1996.
Sharma PK, Gibcus MJ, van der Mei HC, Busscher HJ.2005. Influence of fluid shear and microbubbles onbacterial detachment from a surface. Appl EnvironMicrobiol 71:3668–3673.
Simoes M, Pereira MO, Vieira MJ. 2005. Effect of mechan-ical stress on biofilms challenged by different chemicals.Water Res 39:5142–5152.
Singh A, Van Hamme JD, Ward OP. 2007. Surfactants inmicrobiology and biotechnology. Part 2. Applicationaspects. Biotechnol Adv 25:99–121.
Sommer P, Martin-Rouas C, Mettler E. 1999. Influence ofthe adherent population level on biofilm population,structure and resistance to chlorination. Food Microbiol16:503–515.
Spoering AL, Gilmore MS. 2006. Quorum sensing and DNArelease inbacterial biofilms.CurrOpinMicrobiol 9:133–137.
Stewart PS, Costerton JW. 2001. Antibiotic resistance ofbacteria in biofilms. Lancet 358:135–138.
Stoodley P, Sauer K, Davies DG, Costerton JW. 2002. Biofilmsas complex differentiated communities. Annu Rev Micro-biol 56:187–209.
Sule P, Wadhawan T, Carr NJ, Horne SM, Wolfe AJ, PrussBM. 2009. A combination of assays reveals biomassdifferences in biofilms formed by Escherichia colimutants. Lett Appl Microbiol 49:299–304.
Sutherland IW. 2001. Biofilm exopolysaccharides: a strongand sticky framework. Microbiology 147(Part 1):3–9.
Westphal O, Jann K. 1965. Bacterial lipopolysaccharides.Extraction with phenol-water and further applications ofthe procedure. In: Whistler RL, editor. Methods incarbohydrate chemistry, Vol. 5. New York: AcademicPress. p. 83–91.
Whitchurch CB, Tolker-Nielsen T, Ragas PC, Mattick JS.2002. Extracellular DNA required for bacterial biofilmformation. Science 295:1487–1487.
Wijman JGE, de Leeuw P, Moezelaar R, Zwietering MH,Abee T. 2007. Air-liquid interface biofilms of Bacilluscereus: formation, sporulation, and dispersion. ApplEnviron Microbiol 73:1481–1488.
Xavier JB, Picioreanu C, Rani SA, van Loosdrecht MCM,Stewart PS. 2005. Biofilm-control strategies based onenzymic disruption of the extracellular polymeric sub-stance matrix – a modelling study. Microbiology151:3817–3832.
Biofouling 431
Downloaded By: [lequette, yannick] At: 12:13 1 March 2010




















