
Characteristics of phenanthrene-degrading bacteria isolated from soils contaminated with polycyclic...
11
Characteristics of phenanthrene-degrading bacteria isolated from soils contaminated with polycyclic aromatic hydrocarbons Michael D. Aitken, William T. Stringfellow, Robert D. Nagel, Chikoma Kazunga, and Shu-Hwa Chen Abstract: Ten bacterial strains were isolated from seven contaminated soils by enrichment with phenanthrene as the sole carbon source. These isolates and another phenanthrene-degrading strain were examined for various characteristics related to phenanthrene degradation and their ability to metabolize 12 other polycyclic aromatic hydrocarbons (PAH), ranging in size from two to five rings, after growth in the presence of phenanthrene. Fatty acid methyl ester analysis indicated that at least five genera (Agrobacterium, Bacillus, Burkholderia, Pseudomonas, and Sphingomonas) and at least three species of Pseudomonas were represented in this collection. All of the strains oxidized phenanthrene according to Michaelis–Menten kinetics, with half-saturation coefficients well below the aqueous solubility of phenanthrene in all cases. All but one of the strains oxidized 1-hydroxy-2-naphthoate following growth on phenanthrene, and all oxidized at least one downstream intermediate from either or both of the known phenanthrene degradation pathways. All of the isolates could metabolize (oxidize, mineralize, or remove from solution) a broad range of PAH, although the exact range and extent of metabolism for a given substrate were unique to the particular isolate. Benz[a]anthracene, chrysene, and benzo[a]pyrene were each mineralized by eight of the strains, while pyrene was not mineralized by any. Pyrene was, however, removed from solution by all of the isolates, and the presence of at least one significant metabolite from pyrene was observed by radiochromatography for the five strains in which such metabolites were sought. Our results support earlier indications that the mineralization of pyrene by bacteria may require unique metabolic capabilities that do not appear to overlap with the determinants for mineralization of phenanthrene or other high molecular weight PAH. Key words: kinetics, polycyclic aromatic hydrocarbons, phenanthrene, mineralization, benzo[ a]pyrene. 752 Résumé : Nous avons isolé dix souches bactériennes à partir de sept sols contaminés par culture en présence de phénanthrène comme seule source de carbone. Nous avons étudié diverses caractéristiques de la dégradation du phénanthrène chez ces trois isolats et chez une autre souche dégradant ce composé et avons aussi évalué la capacité à métaboliser 12 autres hydrocarbures aromatiques polycycliques (PAH) dont la taille variait entre deux et cinq anneaux et ce après croissance en présence de phénanthrène. L’analyse des acides gras méthyl estérifiés a confirmé que cette collection bactérienne contenait au moins cinq genres (Agrobacterium, Bacillus, Burkholderia, Pseudomonas et Sphingomonas) et au moins trois espèces de Pseudomonas. Toutes les souches oxydaient le phénanthrène selon la cinétique de Michaelis–Menten avec dans tous les cas avec des coefficients de demi-saturation bien inférieurs à la solubilité dans l’eau du phénanthracène. Toutes les souches sauf une oxydaient le 1-hydroxy-2-naphthoate après croissance en présence de phénanthrène et toutes oxydaient au moins un composé intermédiaire dérivé d’un ou des deux sentiers métaboliques connus de dégradation du phénanthrène. Chacun de ces isolats était capable de métaboliser (oxydation, minéralisation ou élimination de la solution) un large éventail de PAH, même si pour un substrat donné le spectre précis et l’importance du métabolisme des PHA étaient uniques à un isolat particulier. Le benz[ a]anthracène, le chrysène et le benzo[a]pyrène étaient chacun métabolisés par huit des souches alors que le pyrène ne l’était pas par aucune des souches. Le pyrène était cependant éliminé de la solution par tous les isolats et la présence d’au moins un métabolite significatif du pyrène a été observée par radiochromatographie chez les cinq souches où de tels métabolites ont été recherchés. Les résultats de cette étude confirment des indications précédentes à l’effet que la minéralisation du pyrène par les bactéries pourrait nécessiter des propriétés métaboliques particulières qui ne semblent pas recouper les déterminants de la minéralisation du phénanthrène ou d’autres PAH de poids moléculaire élevé. Mots clés : cinétique, hydrocarbures aromatiques polycycliques, phénanthrène, minéralisation, benzo[ a]pyrène. [Traduit par la Rédaction] Aitken et al. Can. J. Microbiol. 44: 743–752 (1998) © 1998 NRC Canada 743 Received January 26, 1998. Revision received May 20, 1998. Accepted June 4, 1998. M.D. Aitken, 1 W.T. Stringfellow, 2 R.D. Nagel, C. Kazunga, and S.-H. Chen. Department of Environmental Sciences and Engineering, School of Public Health, CB No. 7400, University of North Carolina, Chapel Hill, NC 27599-7400, U.S.A. 1 Author to whom all correspondence should be addressed (e-mail: [email protected]). 2 Present address: Center for Environmental Biotechnology, Lawrence Berkeley Laboratory, Berkeley, CA 94720, U.S.A. Can. J. Microbiol. Downloaded from www.nrcresearchpress.com by Calif Dig Lib - Lawrence Berkeley National Laboratory (LBL) on 03/04/13 For personal use only.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Characteristics of phenanthrene-degrading bacteria isolated from soils contaminated with polycyclic...

Characteristics of phenanthrene-degradingbacteria isolated from soils contaminated withpolycyclic aromatic hydrocarbons
Michael D. Aitken, William T. Stringfellow, Robert D. Nagel, Chikoma Kazunga,and Shu-Hwa Chen
Abstract: Ten bacterial strains were isolated from seven contaminated soils by enrichment with phenanthrene as thesole carbon source. These isolates and another phenanthrene-degrading strain were examined for various characteristicsrelated to phenanthrene degradation and their ability to metabolize 12 other polycyclic aromatic hydrocarbons (PAH),ranging in size from two to five rings, after growth in the presence of phenanthrene. Fatty acid methyl ester analysisindicated that at least five genera (Agrobacterium, Bacillus, Burkholderia, Pseudomonas, andSphingomonas) and atleast three species ofPseudomonaswere represented in this collection. All of the strains oxidized phenanthreneaccording to Michaelis–Menten kinetics, with half-saturation coefficients well below the aqueous solubility ofphenanthrene in all cases. All but one of the strains oxidized 1-hydroxy-2-naphthoate following growth onphenanthrene, and all oxidized at least one downstream intermediate from either or both of the known phenanthrenedegradation pathways. All of the isolates could metabolize (oxidize, mineralize, or remove from solution) a broadrange of PAH, although the exact range and extent of metabolism for a given substrate were unique to the particularisolate. Benz[a]anthracene, chrysene, and benzo[a]pyrene were each mineralized by eight of the strains, while pyrenewas not mineralized by any. Pyrene was, however, removed from solution by all of the isolates, and the presence of atleast one significant metabolite from pyrene was observed by radiochromatography for the five strains in which suchmetabolites were sought. Our results support earlier indications that the mineralization of pyrene by bacteria mayrequire unique metabolic capabilities that do not appear to overlap with the determinants for mineralization ofphenanthrene or other high molecular weight PAH.
Key words: kinetics, polycyclic aromatic hydrocarbons, phenanthrene, mineralization, benzo[a]pyrene. 752
Résumé: Nous avons isolé dix souches bactériennes à partir de sept sols contaminés par culture en présence dephénanthrène comme seule source de carbone. Nous avons étudié diverses caractéristiques de la dégradation duphénanthrène chez ces trois isolats et chez une autre souche dégradant ce composé et avons aussi évalué la capacité àmétaboliser 12 autres hydrocarbures aromatiques polycycliques (PAH) dont la taille variait entre deux et cinq anneauxet ce après croissance en présence de phénanthrène. L’analyse des acides gras méthyl estérifiés a confirmé que cettecollection bactérienne contenait au moins cinq genres (Agrobacterium, Bacillus, Burkholderia, PseudomonasetSphingomonas) et au moins trois espèces dePseudomonas. Toutes les souches oxydaient le phénanthrène selon lacinétique de Michaelis–Menten avec dans tous les cas avec des coefficients de demi-saturation bien inférieurs à lasolubilité dans l’eau du phénanthracène. Toutes les souches sauf une oxydaient le 1-hydroxy-2-naphthoate aprèscroissance en présence de phénanthrène et toutes oxydaient au moins un composé intermédiaire dérivé d’un ou desdeux sentiers métaboliques connus de dégradation du phénanthrène. Chacun de ces isolats était capable de métaboliser(oxydation, minéralisation ou élimination de la solution) un large éventail de PAH, même si pour un substrat donné lespectre précis et l’importance du métabolisme des PHA étaient uniques à un isolat particulier. Le benz[a]anthracène, lechrysène et le benzo[a]pyrène étaient chacun métabolisés par huit des souches alors que le pyrène ne l’était pas paraucune des souches. Le pyrène était cependant éliminé de la solution par tous les isolats et la présence d’au moins unmétabolite significatif du pyrène a été observée par radiochromatographie chez les cinq souches où de tels métabolitesont été recherchés. Les résultats de cette étude confirment des indications précédentes à l’effet que la minéralisation dupyrène par les bactéries pourrait nécessiter des propriétés métaboliques particulières qui ne semblent pas recouper lesdéterminants de la minéralisation du phénanthrène ou d’autres PAH de poids moléculaire élevé.
Mots clés: cinétique, hydrocarbures aromatiques polycycliques, phénanthrène, minéralisation, benzo[a]pyrène.
[Traduit par la Rédaction] Aitken et al.
Can. J. Microbiol.44: 743–752 (1998) © 1998 NRC Canada
743
Received January 26, 1998. Revision received May 20, 1998. Accepted June 4, 1998.
M.D. Aitken, 1 W.T. Stringfellow,2 R.D. Nagel, C. Kazunga, and S.-H. Chen.Department of Environmental Sciences andEngineering, School of Public Health, CB No. 7400, University of North Carolina, Chapel Hill, NC 27599-7400, U.S.A.
1Author to whom all correspondence should be addressed (e-mail: [email protected]).2Present address: Center for Environmental Biotechnology, Lawrence Berkeley Laboratory, Berkeley, CA 94720, U.S.A.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Cal
if D
ig L
ib -
Law
renc
e B
erke
ley
Nat
iona
l Lab
orat
ory
(LB
L)
on 0
3/04
/13
For
pers
onal
use
onl
y.

Polycyclic aromatic hydrocarbons (PAH) are found incontaminated soils and groundwater at sites associated withcreosote-based treatment of wood, petroleum processing,and the production of manufactured gas. Many microorgan-isms are able to degrade PAH (Sutherland et al. 1995), sothere is a strong interest in applying bioremediation at manyof these sites. As of 1996, PAH represented the largest classof contaminants for which bioremediation was adopted as atreatment action at Superfund sites in the United States (U.S.Environmental Protection Agency 1996).
In soils and sediments contaminated with PAH, the degra-dation of high molecular weight compounds (those with fouror more rings) by indigenous microorganisms is usuallymuch slower or less extensive than the degradation of lowmolecular weight PAH (Herbes and Schwall 1978; Bossertand Bartha 1986; Shiaris 1989; Mueller et al. 1991a, 1991b;Wild and Jones 1993). The ability of indigenous microbialcommunities to mineralize or otherwise degrade the highmolecular weight compounds also appears to be site specific(Grosser et al. 1991, 1995; Kästner et al. 1994; Carmichaeland Pfaender 1997). There are several possible explanationsfor the apparent recalcitrance of these compounds and thevariability between sites, including effects of bioavailability(Weissenfels et al. 1992), the occurrence (and number) ofdegraders of a specific compound (Kästner et al. 1994), aninability of organisms to grow on the high molecular weightcompounds, or complex interactions among substrates lead-ing to the inhibition of the degradation of one PAH in thepresence of other PAH (Bouchez et al. 1995; Kelley andCerniglia 1995; Millette et al. 1995; Stringfellow and Aitken1995).
It is difficult to elucidate the mechanisms governing thebiodegradation of PAH in complex media containing com-plex mixtures of substrates and heterogeneous microbialcommunities. One approach is to study the constituent bacte-ria contributing to the biotransformation and biodegradationprocesses in these complex systems. By determining thePAH substrate range of individual organisms, for example, itmay be possible to establish links between the metabolismof various substrates or identify those compounds for whichmore specialized metabolic capabilities are required for ex-tensive degradation. It is particularly important to evaluatepotential links between the metabolism of low molecularweight PAH and high molecular weight PAH that have notbeen documented to serve as growth substrates for microor-ganisms.
In this study, we isolated ten bacterial strains from sevendifferent contaminated soils by enrichment on phenanthreneas the sole carbon source. Various characteristics of thesebacteria related to phenanthrene degradation were examined,including the ability to degrade a broad range of PAH aftergrowth in the presence of phenanthrene. In particular, wewere interested in screening these bacteria for their ability todegrade high molecular weight PAH after growth onphenanthrene. Detailed studies related to low molecularweight PAH degradation by two of these strains,Pseudomo-nas saccharophilaP15 andPseudomonas stutzeriP16, havebeen published elsewhere (Stringfellow and Aitken 1994,1995; Grimberg et al. 1996).
Chemicals and mediaPAH were purchased from various suppliers and were the
highest grade available. [9-14C]Phenanthrene (8.3 mCi/mmol;1 Ci = 37 GBq), [4,5,9,10-14C]pyrene (32.3 mCi/mmol), and[7-14C]benzo[a]pyrene (26.6 mCi/mmol) were purchased fromSigma (Milwaukee, Wis.). [5,6,11,12-14C]Chrysene (47.6 mCi/mmol)and [5,6-14C]benz[a]anthracene (54.6 mCi/mmol) were obtainedfrom Chemsyn Science Laboratories (Lenexa, Kans.). Allradiolabeled chemicals had purities >98%. Tap water buffer(TWB), liquid media containing crystalline phenanthrene in TWB(TWB/PAT), peptone plus crystalline phenanthrene (PEP/PAT), andagar media were prepared as described previously (Stringfellowand Aitken 1994).
The aqueous solubilities of the PAH used in this project, inmilligrams per litre, were 110 for indane, 31 for naphthalene,28 for 1-methylnaphthalene, 26 for 2-methylnaphthalene, 3.5 foracenaphthene, 1.9 for fluorene, 1.1 for phenanthrene, 0.07 foranthracene, 0.26 for fluoranthene, 0.13 for pyrene, 0.011 forbenz-[a]anthracene, 0.004 for chrysene, and 0.004 forbenzo-[a]pyrene (Mackay et al. 1992).
Sources of bacteriaSoil samples from several contaminated sites were obtained
from remediation contractors. The sites included a facility at whichrailroad ties were treated with creosote (soil S); a refinery (soil G);a facility used to treat wood with creosote and pentachlorophenol(soil C); and a former gas-manufacturing plant (soil R). A soilsample from the Reilly Tar and Chemical Superfund site inSt. Louis Park, Minn. (soil M), which had been used to treat woodwith creosote, was collected by F. Pfaender (University of NorthCarolina, Chapel Hill, N.C.). A sample from a motor oil contami-nated surface soil was obtained from Durham, N.C. Strains P15and P16 were isolated from creosote-contaminated soil from an-other wood treatment facility (Stringfellow and Aitken 1994).Strain VT1, originally isolated from oil-contaminated soil (Foghtand Westlake 1991), was obtained from J. Foght (University of Al-berta, Edmonton, Alta.).
Enrichment, isolation, and identification ofphenanthrene-degrading bacteria
Strain P21 was isolated from motor oil contaminated soil in thesame manner described for the isolation ofP. saccharophilaP15andP. stutzeriP16 (Stringfellow and Aitken 1994). The remainingstrains were isolated by first inoculating flasks containing 200 mLof TWB/PAT medium with 4–8 g of soil. The flasks were incu-bated at 25°C on a rotary shaker (150 rpm) for 5–7 days, 2 mL ofthe culture fluid was then transferred to fresh medium, and theprocedure was repeated for successive transfers. Cultures fromthe third transfer were dilution plated on R2A agar plates con-taining phenanthrene (R2A/PAT) for isolation of phenanthrene-degrading bacteria. Total bacterial counts ranged from 5 × 106 to5 × 107 cfu/mL except for the enrichment culture from soil R,which contained 109 cfu/mL.
Colonies were selected from dilution plates based on distinctcolony morphology. Most of the selected colonies were among themost frequent of the plated colony types except for strains C5 andM6, which were relatively rare (fewer than 1 of 50 colonies). Eachcolony was picked and streaked onto fresh R2A/PAT plates twiceto ascertain culture purity. Each isolate was then tested for its abil-ity to grow on phenanthrene as the sole carbon source in TWB/PATliquid medium. Isolates able to grow on phenanthrene in the liquidmedium were stored on R2A/PAT slants for further study. All iso-lates were periodically plated on different media (R2A/PAT, nutri-ent agar supplemented with phenanthrene, MacConkey’s, and
© 1998 NRC Canada
744 Can. J. Microbiol. Vol. 44, 1998
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Cal
if D
ig L
ib -
Law
renc
e B
erke
ley
Nat
iona
l Lab
orat
ory
(LB
L)
on 0
3/04
/13
For
pers
onal
use
onl
y.

sheep’s blood) to ascertain culture purity and to verify colonyidentity with that of the original isolate.
The cultures were sent to MIDI Inc. (Newark, Del.) for identifi-cation by fatty acid methyl ester (FAME) analysis. Each isolatewas also stab-inoculated into motility agar in a 16 × 125 mm testtube, and motility was indicated by spreading of the culture after7 days.
Kinetics of phenanthrene oxidationInitial rates of phenanthrene oxidation were determined as a
function of dissolved phenanthrene concentration by respirometryfor cultures grown on PEP/PAT liquid medium, as described previ-ously (Stringfellow and Aitken 1994). Maximum specific oxygenuptake rates (SOURmax) and half-saturation coefficients (KS) wereestimated by fitting the Michaelis–Menten equation to the data bynonlinear regression.
Inducibility of phenanthrene metabolismThe isolates were tested for their ability to oxidize phenanthrene
after growth in peptone medium or in PEP/PAT medium. Initialrates of phenanthrene oxidation were measured by respirometry ata phenanthrene concentration of 0.8 mg/L. Each measurement wasmade twice for each isolate.
Oxidation of phenanthrene degradation intermediatesPhenanthrene is metabolized by various bacteria to the interme-
diate 1-hydroxy-2-naphthoate (1H2N), at which point two diver-gent pathways have been reported (Cerniglia and Heitkamp 1989).In the first pathway, 1H2N is metabolized to salicylate and then tocatechol; in the second pathway, 1H2N is metabolized to phthalateand then to protocatechuate. Each isolate was grown on PEP/PATmedium and tested for its ability to oxidize 1H2N, salicylate,catechol, phthalate, and in some cases, protocatechuate, each at aconcentration of 5.3 mg/L. Initial rates of oxidation of each com-pound were determined by respirometry.
Screening for biosurfactant productionWashed cells grown in PEP/PAT medium were inoculated into
TWB/PAT medium or TWB containing naphthalene in 250-mLBoston round screw-capped bottles. The bottles were incubated atroom temperature in the dark on a rotary shaker (120 rpm). Theoptical absorbance was monitored every 2 or 3 days until lategrowth or early stationary phase. Then the surface tension of theculture fluid (centrifuged and filtered) was measured using a ringtensiometer (Stringfellow and Aitken 1994). The surface tension ofthe controls (uninoculated culture media) was measured at thesame time.
PAH substrate rangeVarious assays have been used to screen for PAH metabolism by
microorganisms. We chose respirometry as a rapid measure of theextensive oxidation of low molecular weight PAH with solubilitiesgreater than that of phenanthrene (indane, naphthalene,methylnaphthalenes, fluorene, and acenaphthene). Mineralizationwas selected as a measure of extensive metabolism for most of thecompounds with solubilities less than that of phenanthrene. SincePAH can be partially metabolized without mineralization, the re-moval of these substrates from solution was also tested. For thehigh molecular weight compounds that were not mineralized, evi-dence of metabolite formation was obtained in several cases as afollow-up to substrate removal assays by incubating the cultureswith radiolabeled substrates and analyzing the culture fluids byradiochromatography. For two low-solubility compounds that werenot available in our laboratory in radiolabeled form, anthraceneand fluoranthene, removal from solution was the only measure ofmetabolism evaluated.
Oxidation of low molecular weight PAHWashed cells grown in PEP/PAT medium were tested for their
ability to oxidize several two- and three-ring PAH by respirometry.The compounds were prepared in methanol and, except for indane,were injected into the respirometer vessel to a final concentrationof approximately 80% of the compound’s aqueous solubility; ow-ing to apparent inhibitory effects at concentrations near its solubil-ity limit, indane was tested at a concentration of approximately30% of its aqueous solubility. The oxidation of phenanthrene wasused as a positive control and methanol alone was used as a nega-tive control.
MineralizationMineralization of 14C-labeled benz[a]anthracene, benzo[a]-
pyrene, chrysene, phenanthrene, and pyrene was evaluated for eachisolate except strain S1, whose PAH-degrading phenotype was lostduring storage. Triplicate vials were prepared for each substrate byadding either a combination of unlabeled and14C-labeled substrate(for benz[a]anthracene, phenanthrene, or pyrene) or the14C-labeledsubstrate alone (for benzo[a]pyrene and chrysene) to provide ap-proximately 20 000 disintegrations per minute (dpm; 1 dpm =0.0167 Bq) except for vials with14C-benzo[a]pyrene, which con-tained 10 000 dpm. The final concentration of each substrate wasapproximately 80% of its aqueous solubility.
The vials were inoculated with washed cells obtained aftergrowth in TWB containing solidified phenanthrene (Grimberg etal. 1996). Triplicate killed controls were prepared by acidifyingwith H3PO4 prior to inoculation. Except for strains C2, R1, andP16, vials were closed with a cap and teflon-lined septum, fromwhich was suspended a plastic well containing a fluted strip ofWhatman No. 1 filter paper soaked with 200 µL of 2 N KOH toserve as a CO2 trap (Carmichael and Pfaender 1997). Vials wereincubated on a rotary shaker (100 rpm) at room temperature in thedark for 24 h, then H3PO4 was injected through the septa of thesample vials and the uninoculated controls. All of the vials werereturned to the shaker for another 24 h, then the filter paper stripswere removed and placed in scintillation vials containing 9 mL ofScintiSafe scintillation fluid (Fisher Scientific, Pittsburgh, Penn.).Radioactivity was counted with a Tri-Carb 1900TR liquid scintilla-tion analyzer (Packard, Meriden, Conn.). The amount of radiolabelremaining in each incubation vial was also counted and added tothe amount trapped in the headspace to determine total recovery.The recovery from all live samples for which this method was used(mean ± SD;n = 21) was 70 ± 12% for phenanthrene, 89 ± 21%for pyrene, 92 ± 6.3% for benz[a]anthracene, 88 ± 6.9% for chry-sene, and 102 ± 9.1% for benzo[a]pyrene. Recovery of radiolabelin the CO2 traps for the killed controls was 0.6 ± 0.2% forphenanthrene, 0.8 ± 0.6% for pyrene, 0.6 ± 0.3% forbenz[a]anthracene, 0.5 ± 0.3% for chrysene, and 0.6 ± 0.3% forbenzo[a]pyrene.
The system for trapping14CO2 described above was modifiedfor assays performed with strains C2, R1, and P16 to minimizelosses of phenanthrene and pyrene from control vials, which oc-curred primarily by sorption of volatilized PAH to the plastic buck-ets used to hold the CO2 traps. Such losses did not, however,elevate concentrations of radiolabel on the filter paper traps. Forthese strains, a glass tube containing a filter paper strip soakedwith 200 µL of 2 N KOH was inserted into each vial to trap14CO2.Total recovery of radiolabel in the live samples with this method(n = 9) was 65 ± 7.6% for phenanthrene, 88 ± 5.1% for pyrene, 90 ±1.8% for benz[a]anthracene, 88 ± 5.6% for chrysene, and 90 ±1.8% for benzo[a]pyrene. Recovery of radiolabel in the CO2 trapsfor the killed controls was 7.5 ± 2.4% for phenanthrene, 3.1 ±1.1% for pyrene, 0.9 ± 0.5% for benz[a]anthracene, 0.5 ± 0.2% forchrysene, and 0.8 ± 0.5% for benzo[a]pyrene. Although thismethod resulted in higher counts in the CO2 traps for phenanthreneand pyrene than the method using plastic buckets in the headspace,
© 1998 NRC Canada
Aitken et al. 745
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Cal
if D
ig L
ib -
Law
renc
e B
erke
ley
Nat
iona
l Lab
orat
ory
(LB
L)
on 0
3/04
/13
For
pers
onal
use
onl
y.
total recoveries (CO2 traps plus residual in the liquid medium) inkilled and uninoculated controls were much greater using the mod-ified method.
PAH removalThe ability of isolates to remove from solution the less soluble
PAH was determined by high pressure liquid chromatography(HPLC) analysis. Washed cells grown in PEP/PAT medium weresuspended in replicate flasks in 15 mL of TWB and then 100 µL ofa methanol solution containing the PAH of interest was injectedinto all the flasks; the concentration of each PAH after injection, inmicrograms per litre, was 25 for anthracene, 200 for fluoranthene,120 for pyrene, 2.0 for chrysene, and 2.5 for benzo[a]pyrene.Three flasks were amended with 5 mL of formalin to serve askilled controls, and three other flasks were amended with the samevolume of dechlorinated and sterilized tap water. All flasks wereincubated at 25°C on a rotary shaker for 24 (for anthracene, chry-sene and benzo[a]pyrene) or 48 h (for fluoranthene and pyrene), atwhich time 1.0 mL of culture fluid from each live flask was addedto 0.5 mL of formalin in 12 × 75 mm test tubes. The resulting sus-pensions were mixed and analyzed for the PAH of interest byHPLC. PAH removal was evaluated by performing at test on PAHconcentrations in the live samples and the killed controls; positiveremoval was considered to occur if the means were different atp <0.05.
HPLC analysis was conducted with a Waters (Milford, Mass.)600E system equipped with a model 470 scanning fluorescence de-tector and model 717 autosampler. Separation was achieved witha C18 reverse-phase column (25 cm × 4.6 mm, 5-µm-particle size;Supelco, Bellefonte, Penn.). The mobile phase was 70%acetonitrile in water at a flow rate of 1.0 mL/min. Excitation andemission wavelengths, in nanometres, were, respectively, 259 and370 for phenanthrene, 252 and 405 for anthracene, 284 and 460 forfluoranthene, 336 and 398 for pyrene, 268 and 384 for chrysene,and 378 and 406 for benzo[a]pyrene. The quantification limits, inmicrograms per litre, were 0.89 for phenanthrene, 0.025 foranthracene, 0.20 for fluoranthene, 0.12 for pyrene, 0.02 for chry-sene, and 0.025 for benzo[a]pyrene.
Detection of metabolitesSelected strains that were not able to mineralize one or more of
the high molecular weight PAH were grown on phenanthrene,washed, and inoculated into 10 mL of TWB containing 40 000 –100 000 dpm of labeled substrate. A killed control was prepared in
the same manner but was acidified with H3PO4. After a 2- to 3-dayincubation period, the live sample was acidified with H3PO4 andthe contents of each vial were extracted with two volumes of ethylacetate (10–20 mL each). The ethyl acetate solutions from a givenvial were pooled and then blown to dryness with a stream of N2,and the residue was resuspended in 1 mL of acetonitrile. Theacetonitrile solutions were analyzed by either of two HPLC tech-niques. In the first method, the solution was repeatedly injected(eight injections, 100 µL per injection) onto the HPLC column andeluent fractions were collected on a Waters fraction collector every27 s. The mobile phase was 70% acetonitrile in water acidifiedwith 0.01% trifluoroacetic acid at a flow rate of 1.0 mL/min. Thefractions from each injection were pooled in scintillation vials foreach sample, and scintillation fluid (9 mL) was added to each vialprior to scintillation counting. In the second method, aradiochromatography detector fitted with a calcium fluoride solidscintillator (Packard Radiomatic Flo-One\Beta) was used with thesame mobile phase conditions, except the water was not acidified;the injection volume was 25 µL.
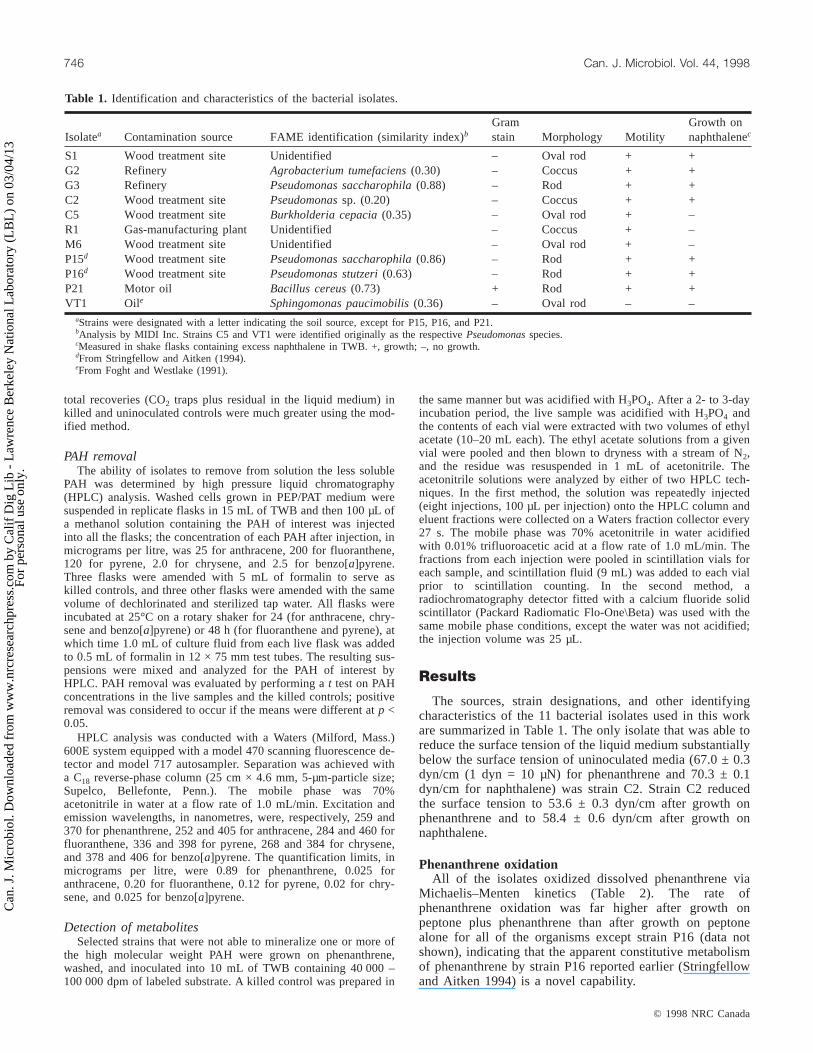
The sources, strain designations, and other identifyingcharacteristics of the 11 bacterial isolates used in this workare summarized in Table 1. The only isolate that was able toreduce the surface tension of the liquid medium substantiallybelow the surface tension of uninoculated media (67.0 ± 0.3dyn/cm (1 dyn = 10 µN) for phenanthrene and 70.3 ± 0.1dyn/cm for naphthalene) was strain C2. Strain C2 reducedthe surface tension to 53.6 ± 0.3 dyn/cm after growth onphenanthrene and to 58.4 ± 0.6 dyn/cm after growth onnaphthalene.
Phenanthrene oxidationAll of the isolates oxidized dissolved phenanthrene via
Michaelis–Menten kinetics (Table 2). The rate ofphenanthrene oxidation was far higher after growth onpeptone plus phenanthrene than after growth on peptonealone for all of the organisms except strain P16 (data notshown), indicating that the apparent constitutive metabolismof phenanthrene by strain P16 reported earlier (Stringfellowand Aitken 1994) is a novel capability.
© 1998 NRC Canada
746 Can. J. Microbiol. Vol. 44, 1998
Isolatea Contamination source FAME identification (similarity index)bGramstain Morphology Motility
Growth onnaphthalenec
S1 Wood treatment site Unidentified – Oval rod + +G2 Refinery Agrobacterium tumefaciens(0.30) – Coccus + +G3 Refinery Pseudomonas saccharophila(0.88) – Rod + +C2 Wood treatment site Pseudomonassp. (0.20) – Coccus + +C5 Wood treatment site Burkholderia cepacia(0.35) – Oval rod + –R1 Gas-manufacturing plant Unidentified – Coccus + –M6 Wood treatment site Unidentified – Oval rod + –P15d Wood treatment site Pseudomonas saccharophila(0.86) – Rod + +P16d Wood treatment site Pseudomonas stutzeri(0.63) – Rod + +P21 Motor oil Bacillus cereus(0.73) + Rod + +VT1 Oile Sphingomonas paucimobilis(0.36) – Oval rod – –
aStrains were designated with a letter indicating the soil source, except for P15, P16, and P21.bAnalysis by MIDI Inc. Strains C5 and VT1 were identified originally as the respectivePseudomonasspecies.cMeasured in shake flasks containing excess naphthalene in TWB. +, growth; –, no growth.dFrom Stringfellow and Aitken (1994).eFrom Foght and Westlake (1991).
Table 1. Identification and characteristics of the bacterial isolates.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Cal
if D
ig L
ib -
Law
renc
e B
erke
ley
Nat
iona
l Lab
orat
ory
(LB
L)
on 0
3/04
/13
For
pers
onal
use
onl
y.

Oxidation of known phenanthrene degradationintermediates
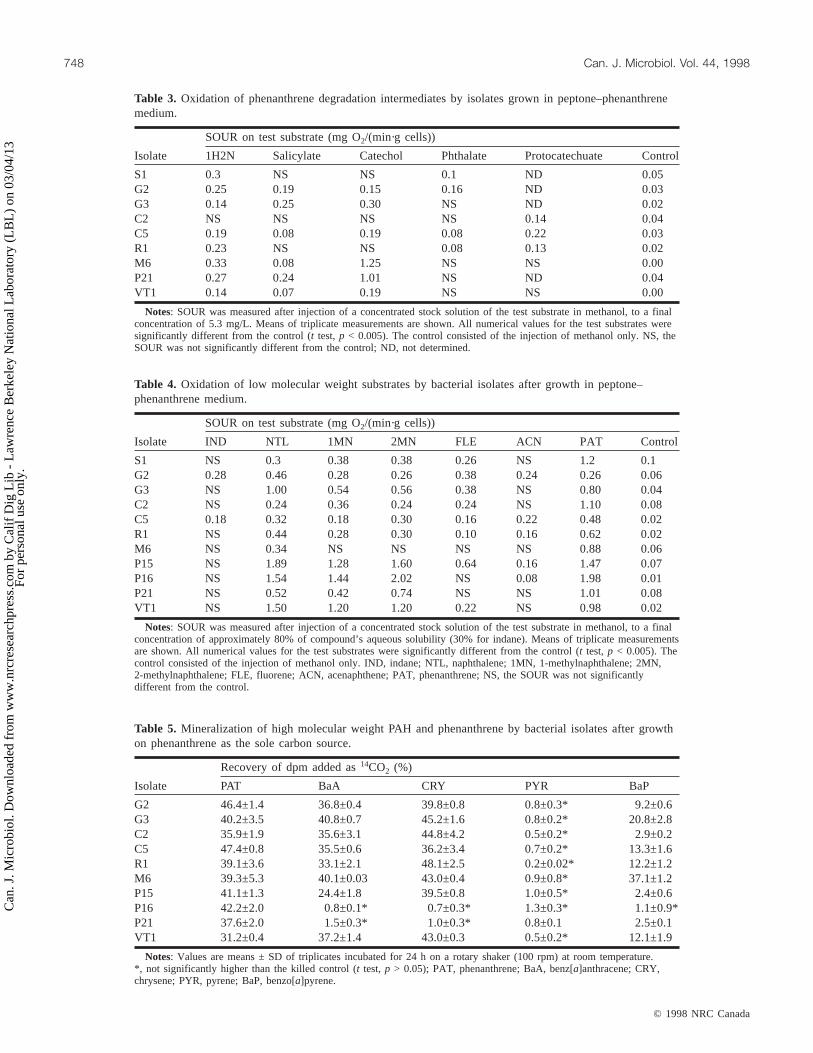
Each isolate was grown in peptone–phenanthrene mediumand tested for its ability to oxidize known intermediatesof phenanthrene metabolism (Table 3). Three strains (G3,P21, and VT1) oxidized salicylate and catechol but notphthalate, two strains (S1 and R1) oxidized phthalate but notsalicylate or catechol, and two others (G2 and C5) oxidizedintermediates of both known pathways for phenanthrenedegradation. The only intermediate oxidized by strain C2was protocatechuate.
Oxidation of low molecular weight PAHThe oxidation of several two- and three-ring PAH was
tested for each isolate after growth in peptone–phenanthrenemedium (Table 4). Strain M6 was the only isolate not able tooxidize most of the low molecular weight substrates. Of thesubstrates tested, indane and acenaphthene were oxidized bythe fewest organisms. Strains M6, P16, and P21 were theonly isolates that did not oxidize fluorene, although earlierstudies indicated that strain P16 can metabolize fluorene in amanner not detectable by respirometry (Stringfellow andAitken 1995).
Mineralization of selected high molecular weight PAHThe mineralization of benz[a]anthracene, chrysene,
pyrene, and benzo[a]pyrene after growth on phenanthrenewas evaluated for all of the isolates except strain S1. Resultsare presented in Table 5. All of the isolates were able tomineralize phenanthrene, the positive control, but none ofthe isolates was able to mineralize pyrene. In most cases the14CO2 data were unambiguous, although in general the re-covery of 14CO2 from benzo[a]pyrene was less than thatfrom the other compounds. For strains C2, P15, and P21, therecovery of14CO2 for benzo[a]pyrene was less than 3% ofthe added label but significantly higher than in the killedcontrols. Longer incubation (48 h) resulted in mineralizationof 30 ± 1.6% of the labeled benzo[a]pyrene for strain P15and 7.6 ± 0.7% for strain P21, whereas benzo[a]pyrene wasnot significantly mineralized by strain C2.
Removal of less soluble PAHMineralization of a PAH substrate requires the complete
oxidation of at least one ring (or more than one ring, de-pending on the location of the labeled carbon(s)), but it isalso possible to oxidize a ring via one or more steps withoutreleasing CO2. However, respirometry was not considered asufficiently sensitive assay for the oxidation of the PAH withaqueous solubilities lower than that of phenanthrene. There-fore, the ability of the isolates to remove these compoundsfrom aqueous solution was evaluated as an indication of atleast partial transformation. Killed controls were preparedand analyzed in the same manner to account for adsorptionof the PAH to the glassware and to cells.
All of the organisms were able to remove fluorantheneand pyrene well in excess of the amount removed by thekilled controls, and anthracene was removed by all seven ofthe strains in which it was evaluated (data not shown). Forthe organisms that did not mineralize benz[a]anthracene,chrysene, and (or) benzo[a]pyrene, in all cases the substratewas removed from solution. In several cases, we have subse-
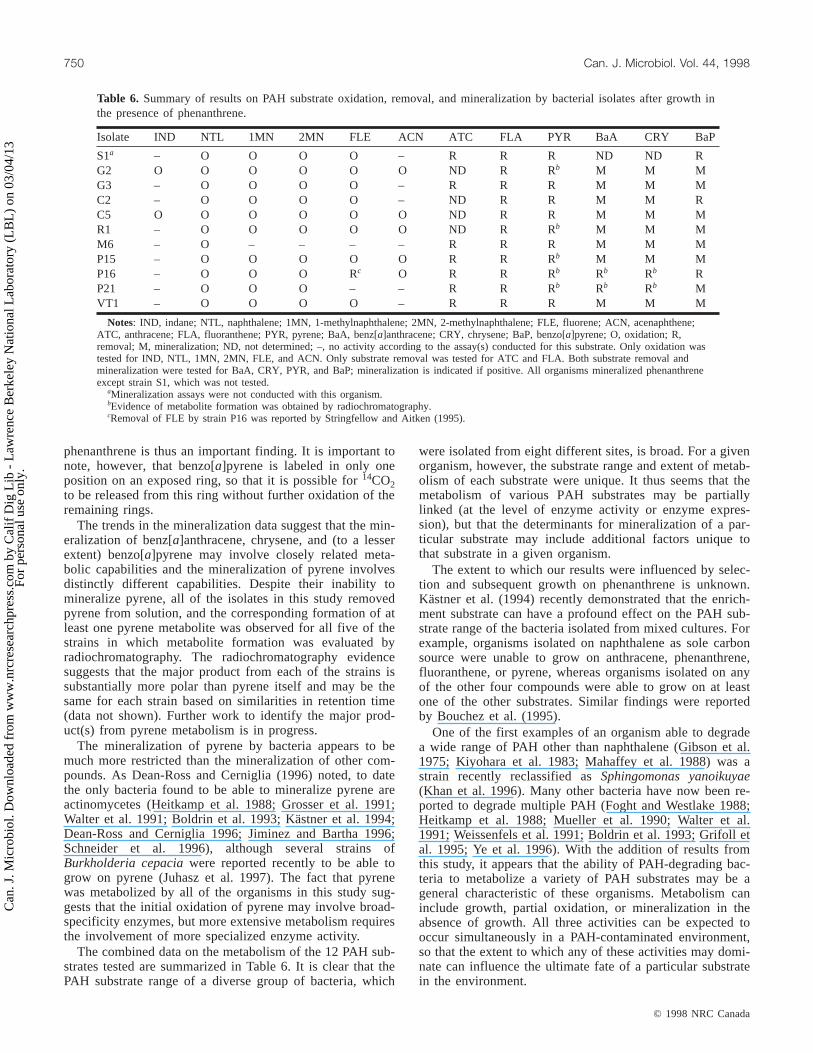
quently obtained positive evidence of metabolite formationby observing radioactive peaks in HPLC analysis indicativeof products substantially more polar than the parent com-pound. In particular, we have obtained such evidence formetabolites from pyrene for five of the isolates (G2, R1,P15, P16, and P21). The HPLC evidence for a product ofpyrene metabolism from strain P15 is shown in Fig. 1. It isunclear whether the shoulder that precedes the main peak at4 min in Fig. 1 is a second peak or simply an artifact of themethod used, which involved repeated injection of the sam-ple and subsequent fraction collection of the HPLC eluate.For all of the other isolates tested the main product peak wasin the same region as that shown in Fig. 1; a shoulder pre-ceding the main peak was also observed for the sample fromstrain R1.
Eleven bacteria isolated from eight different contami-nated sites were examined for characteristics related tophenanthrene degradation and for their ability to metabolizea range of PAH substrates after growth in media containingphenanthrene. Except for strain VT1, these organisms wereisolated by enrichment on phenanthrene as the sole carbonsource. Eight of the isolates were identified to at least thegenus level by FAME analysis (Table 1), but the relativelylow similarity index values for strains G2, C5, and VT1 in-dicate that these are either atypical or stressed strains of therespective type culture in the MIDI library. The organismsare primarily Gram-negative, representing at least three spe-cies ofPseudomonasand four other genera (Agrobacterium,Bacillus, Burkholderia, andSphingomonas). The only Gram-positive isolate wasBacillus cereusP21. The genetic diver-sity of the cultures isolated during the present study is con-sistent with earlier findings by Foght and Westlake (1991)and by Mueller et al. (1994) on the diversity of bacterialdegradation of PAH.
© 1998 NRC Canada
Aitken et al. 747
IsolateSOURmax
(mg O2/(min·g cells))a KS (mg/L)
S1 1.83±0.10 0.15±0.07G2 0.43±0.12 0.15±0.14G3 1.04±0.10 0.12±0.05C2 1.49±0.39 0.21±0.14C5 0.66±0.06 0.075±0.028R1 0.45±0.04 0.014±0.012M6 0.76±0.11 0.17±0.08P15b 1.73±0.22 0.15±0.07P16b 3.14±0.36 0.16±0.05P21 1.31±0.22 0.38±0.15VT1 1.10±0.16 0.14±0.07
aThe estimate from nonlinear regression is shown withthe 95% confidence interval.
bFrom Stringfellow and Aitken (1994).
Table 2. Michaelis–Menten kinetic coefficients foroxidation of phenanthrene by bacterial isolatesgrown in peptone–phenanthrene medium.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Cal
if D
ig L
ib -
Law
renc
e B
erke
ley
Nat
iona
l Lab
orat
ory
(LB
L)
on 0
3/04
/13
For
pers
onal
use
onl
y.
© 1998 NRC Canada
748 Can. J. Microbiol. Vol. 44, 1998
SOUR on test substrate (mg O2/(min·g cells))
Isolate IND NTL 1MN 2MN FLE ACN PAT Control
S1 NS 0.3 0.38 0.38 0.26 NS 1.2 0.1G2 0.28 0.46 0.28 0.26 0.38 0.24 0.26 0.06G3 NS 1.00 0.54 0.56 0.38 NS 0.80 0.04C2 NS 0.24 0.36 0.24 0.24 NS 1.10 0.08C5 0.18 0.32 0.18 0.30 0.16 0.22 0.48 0.02R1 NS 0.44 0.28 0.30 0.10 0.16 0.62 0.02M6 NS 0.34 NS NS NS NS 0.88 0.06P15 NS 1.89 1.28 1.60 0.64 0.16 1.47 0.07P16 NS 1.54 1.44 2.02 NS 0.08 1.98 0.01P21 NS 0.52 0.42 0.74 NS NS 1.01 0.08VT1 NS 1.50 1.20 1.20 0.22 NS 0.98 0.02
Notes: SOUR was measured after injection of a concentrated stock solution of the test substrate in methanol, to a finalconcentration of approximately 80% of compound’s aqueous solubility (30% for indane). Means of triplicate measurementsare shown. All numerical values for the test substrates were significantly different from the control (t test,p < 0.005). Thecontrol consisted of the injection of methanol only. IND, indane; NTL, naphthalene; 1MN, 1-methylnaphthalene; 2MN,2-methylnaphthalene; FLE, fluorene; ACN, acenaphthene; PAT, phenanthrene; NS, the SOUR was not significantlydifferent from the control.
Table 4. Oxidation of low molecular weight substrates by bacterial isolates after growth in peptone–phenanthrene medium.
SOUR on test substrate (mg O2/(min·g cells))
Isolate 1H2N Salicylate Catechol Phthalate Protocatechuate Control
S1 0.3 NS NS 0.1 ND 0.05G2 0.25 0.19 0.15 0.16 ND 0.03G3 0.14 0.25 0.30 NS ND 0.02C2 NS NS NS NS 0.14 0.04C5 0.19 0.08 0.19 0.08 0.22 0.03R1 0.23 NS NS 0.08 0.13 0.02M6 0.33 0.08 1.25 NS NS 0.00P21 0.27 0.24 1.01 NS ND 0.04VT1 0.14 0.07 0.19 NS NS 0.00
Notes: SOUR was measured after injection of a concentrated stock solution of the test substrate in methanol, to a finalconcentration of 5.3 mg/L. Means of triplicate measurements are shown. All numerical values for the test substrates weresignificantly different from the control (t test,p < 0.005). The control consisted of the injection of methanol only. NS, theSOUR was not significantly different from the control; ND, not determined.
Table 3. Oxidation of phenanthrene degradation intermediates by isolates grown in peptone–phenanthrenemedium.
Recovery of dpm added as14CO2 (%)
Isolate PAT BaA CRY PYR BaP
G2 46.4±1.4 36.8±0.4 39.8±0.8 0.8±0.3* 9.2±0.6G3 40.2±3.5 40.8±0.7 45.2±1.6 0.8±0.2* 20.8±2.8C2 35.9±1.9 35.6±3.1 44.8±4.2 0.5±0.2* 2.9±0.2C5 47.4±0.8 35.5±0.6 36.2±3.4 0.7±0.2* 13.3±1.6R1 39.1±3.6 33.1±2.1 48.1±2.5 0.2±0.02* 12.2±1.2M6 39.3±5.3 40.1±0.03 43.0±0.4 0.9±0.8* 37.1±1.2P15 41.1±1.3 24.4±1.8 39.5±0.8 1.0±0.5* 2.4±0.6P16 42.2±2.0 0.8±0.1* 0.7±0.3* 1.3±0.3* 1.1±0.9*P21 37.6±2.0 1.5±0.3* 1.0±0.3* 0.8±0.1 2.5±0.1VT1 31.2±0.4 37.2±1.4 43.0±0.3 0.5±0.2* 12.1±1.9
Notes: Values are means ± SD of triplicates incubated for 24 h on a rotary shaker (100 rpm) at room temperature.*, not significantly higher than the killed control (t test,p > 0.05); PAT, phenanthrene; BaA, benz[a]anthracene; CRY,chrysene; PYR, pyrene; BaP, benzo[a]pyrene.
Table 5. Mineralization of high molecular weight PAH and phenanthrene by bacterial isolates after growthon phenanthrene as the sole carbon source.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Cal
if D
ig L
ib -
Law
renc
e B
erke
ley
Nat
iona
l Lab
orat
ory
(LB
L)
on 0
3/04
/13
For
pers
onal
use
onl
y.
Phenanthrene metabolismAll of the isolates oxidized phenanthrene at rates de-
scribed by Michaelis–Menten kinetics (Table 2) and all hadKS values well below the aqueous solubility of phenanthrene(approximately 1.1 mg/L). These results suggest that all ofthe organisms are efficient utilizers of phenanthrene. The se-lection of bacteria with similar phenanthrene utilization ki-netics is not surprising, considering that they were (exceptfor VT1) enriched in liquid media under conditions thatwould have selected against organisms that metabolizephenanthrene slowly.
The isolates differed in their ability to oxidize intermedi-ates of the two known pathways for phenanthrene metabo-lism (Table 3). For the two strains able to oxidizeintermediates of both the salicylate pathway and thephthalate pathway (G2 and C5), it is not known whetherboth pathways exist in these organisms or whether a singlepathway might contain enzymes with relaxed specificity.The pathway used by strain C2 is also ambiguous, as it wasunable to oxidize 1H2N, salicylate, or phthalate. Negativeresults from the oxidation assay on the acidic metabolitesare not conclusive, as oxidation first requires transport of theacids at least across the outer cell envelope; assays with cell-free extracts would be required to confirm these results.
Growth of the bacterial isolates on phenanthrene or naph-thalene did not appear to involve the production ofextracellular biosurfactants, with the possible exception ofstrain C2. Extracellular emulsifying agents or biosurfactantswere recently reported to be involved in PAH degradation byseveral bacteria (Burd and Ward 1996; Dagher et al. 1997).However, given the currently available evidence, it seemsthat the involvement of biosurfactants in PAH degradation isless common than their involvement in aliphatic hydrocar-bon degradation (Hommel 1990).
Substrate rangeThe substrate ranges of PAH-degrading bacteria have
been tested with a variety of assays, including growth, oxi-dation, substrate removal, the formation of clear or coloredzones around colonies on solid media, indirect measurementof metabolites (optical absorbance of filtered culture fluidsor production of phenolic compounds), and mineralization.Demonstrations of growth become increasingly ambiguousas the solubility of the substrate decreases, and growth isonly one indication of an organism’s ability to metabolize aparticular PAH. PAH can also be metabolized only partially(Gibson et al. 1975; Mueller et al. 1990; Grifoll et al. 1995;Burd and Ward 1996; Ye et al. 1996; Dagher et al. 1997) orcan be mineralized in the absence of growth (Heitkamp et al.1988; Mahaffey et al. 1988; Foght and Westlake 1991;Grosser et al. 1991; Kelley et al. 1991; Schneider et al.1996). In this study, we focused on the PAH-degrading capa-bilities of soil bacteria after growth in the presence of theenrichment substrate, phenanthrene. The metabolism of vari-ous low molecular weight substrates was tested byrespirometry, and metabolism of anthracene and varioushigh molecular weight substrates was tested by mineraliza-tion, removal from solution, or both.
Except for strain M6, growth of the isolates in the pres-ence of phenanthrene provided the capability to oxidize sev-eral other two- and three-ring substrates (Table 2). Only two
strains (G2 and C5) were able to oxidize indane. Every or-ganism oxidized naphthalene, in some cases at rates higherthan the rate of phenanthrene oxidation; such a result wasalso obtained for another PAH-degrading pseudomonad(Rogoff 1962). For most of the other substrates, the rate ofoxidation of the test substrate was not as high as the rate ofphenanthrene oxidation (Table 2).
Our results suggest that the ability to oxidize naphthalenemay be a common trait of phenanthrene-degrading bacteria.However, the observation that several strains were not ableto grow on naphthalene (Table 1) has also been observed fora variety of other bacteria able to grow on phenanthrene(Kiyohara and Nagao 1978; Foght and Westlake 1991;Kästner et al. 1994; Bouchez et al. 1995; Shuttleworth andCerniglia 1996). It therefore appears that the requirementsfor growth on naphthalene exceed the requirements for sig-nificant rates of oxidation and do not overlap entirely withthe requirements for growth on phenanthrene.
All of the bacterial isolates were able to degrade the fivehigh molecular weight PAH tested (benz[a]anthracene, chry-sene, fluoranthene, pyrene, and benzo[a]pyrene). The dataon mineralization of selected high molecular weight PAH(Table 5) revealed two trends. First, none of the organismscould mineralize pyrene, at least after growth onphenanthrene. Second, for benz[a]anthracene and chrysene,all of the isolates were either able to mineralize both or nei-ther. Strain C2 was the only isolate that mineralizedbenz[a]anthracene and chrysene but not benzo[a]pyrene,whereas strain P21 was the only organism that mineralizedbenzo[a]pyrene but not benz[a]anthracene or chrysene.
The results reported here add to a very limited data baseon the ability of pure cultures to mineralize benzo[a]pyrene(Heitkamp and Cerniglia 1989; Grosser et al. 1991; Ye et al.1996). Since no organism has yet been found that is able togrow on benzo[a]pyrene as a sole carbon and energy source,the degradation of benzo[a]pyrene and other high molecularweight PAH in the environment may involve (or require) alink to growth of the degrading populations on low molecu-lar weight compounds. The fact that 8 of the 10 strainstested mineralized benzo[a]pyrene after growth on
© 1998 NRC Canada
Aitken et al. 749
0 2 4 6 8 10 12 140
1000
2000
3000
4000
5000
6000
7000
Active Culture
Killed Control
dpm
inH
PLC
frac
tion
HPLC retention time (min)
Fig. 1. Radiochromatograms of extracts from the culture fluidsof P. saccharophilaP15 grown on phenanthrene andsubsequently incubated with14C-pyrene.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Cal
if D
ig L
ib -
Law
renc
e B
erke
ley
Nat
iona
l Lab
orat
ory
(LB
L)
on 0
3/04
/13
For
pers
onal
use
onl
y.
phenanthrene is thus an important finding. It is important tonote, however, that benzo[a]pyrene is labeled in only oneposition on an exposed ring, so that it is possible for14CO2to be released from this ring without further oxidation of theremaining rings.
The trends in the mineralization data suggest that the min-eralization of benz[a]anthracene, chrysene, and (to a lesserextent) benzo[a]pyrene may involve closely related meta-bolic capabilities and the mineralization of pyrene involvesdistinctly different capabilities. Despite their inability tomineralize pyrene, all of the isolates in this study removedpyrene from solution, and the corresponding formation of atleast one pyrene metabolite was observed for all five of thestrains in which metabolite formation was evaluated byradiochromatography. The radiochromatography evidencesuggests that the major product from each of the strains issubstantially more polar than pyrene itself and may be thesame for each strain based on similarities in retention time(data not shown). Further work to identify the major prod-uct(s) from pyrene metabolism is in progress.
The mineralization of pyrene by bacteria appears to bemuch more restricted than the mineralization of other com-pounds. As Dean-Ross and Cerniglia (1996) noted, to datethe only bacteria found to be able to mineralize pyrene areactinomycetes (Heitkamp et al. 1988; Grosser et al. 1991;Walter et al. 1991; Boldrin et al. 1993; Kästner et al. 1994;Dean-Ross and Cerniglia 1996; Jiminez and Bartha 1996;Schneider et al. 1996), although several strains ofBurkholderia cepaciawere reported recently to be able togrow on pyrene (Juhasz et al. 1997). The fact that pyrenewas metabolized by all of the organisms in this study sug-gests that the initial oxidation of pyrene may involve broad-specificity enzymes, but more extensive metabolism requiresthe involvement of more specialized enzyme activity.
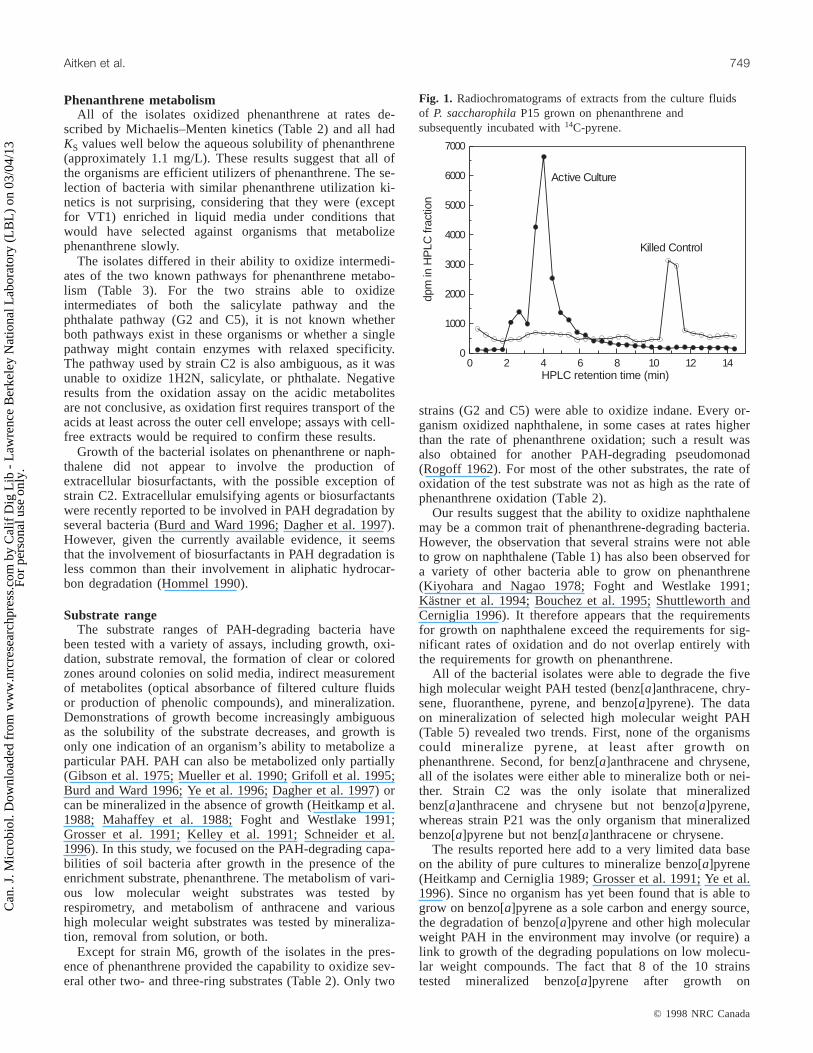
The combined data on the metabolism of the 12 PAH sub-strates tested are summarized in Table 6. It is clear that thePAH substrate range of a diverse group of bacteria, which
were isolated from eight different sites, is broad. For a givenorganism, however, the substrate range and extent of metab-olism of each substrate were unique. It thus seems that themetabolism of various PAH substrates may be partiallylinked (at the level of enzyme activity or enzyme expres-sion), but that the determinants for mineralization of a par-ticular substrate may include additional factors unique tothat substrate in a given organism.
The extent to which our results were influenced by selec-tion and subsequent growth on phenanthrene is unknown.Kästner et al. (1994) recently demonstrated that the enrich-ment substrate can have a profound effect on the PAH sub-strate range of the bacteria isolated from mixed cultures. Forexample, organisms isolated on naphthalene as sole carbonsource were unable to grow on anthracene, phenanthrene,fluoranthene, or pyrene, whereas organisms isolated on anyof the other four compounds were able to grow on at leastone of the other substrates. Similar findings were reportedby Bouchez et al. (1995).
One of the first examples of an organism able to degradea wide range of PAH other than naphthalene (Gibson et al.1975; Kiyohara et al. 1983; Mahaffey et al. 1988) was astrain recently reclassified asSphingomonas yanoikuyae(Khan et al. 1996). Many other bacteria have now been re-ported to degrade multiple PAH (Foght and Westlake 1988;Heitkamp et al. 1988; Mueller et al. 1990; Walter et al.1991; Weissenfels et al. 1991; Boldrin et al. 1993; Grifoll etal. 1995; Ye et al. 1996). With the addition of results fromthis study, it appears that the ability of PAH-degrading bac-teria to metabolize a variety of PAH substrates may be ageneral characteristic of these organisms. Metabolism caninclude growth, partial oxidation, or mineralization in theabsence of growth. All three activities can be expected tooccur simultaneously in a PAH-contaminated environment,so that the extent to which any of these activities may domi-nate can influence the ultimate fate of a particular substratein the environment.
© 1998 NRC Canada
750 Can. J. Microbiol. Vol. 44, 1998
Isolate IND NTL 1MN 2MN FLE ACN ATC FLA PYR BaA CRY BaP
S1a – O O O O – R R R ND ND RG2 O O O O O O ND R Rb M M MG3 – O O O O – R R R M M MC2 – O O O O – ND R R M M RC5 O O O O O O ND R R M M MR1 – O O O O O ND R Rb M M MM6 – O – – – – R R R M M MP15 – O O O O O R R Rb M M MP16 – O O O Rc O R R Rb Rb Rb RP21 – O O O – – R R Rb Rb Rb MVT1 – O O O O – R R R M M M
Notes: IND, indane; NTL, naphthalene; 1MN, 1-methylnaphthalene; 2MN, 2-methylnaphthalene; FLE, fluorene; ACN, acenaphthene;ATC, anthracene; FLA, fluoranthene; PYR, pyrene; BaA, benz[a]anthracene; CRY, chrysene; BaP, benzo[a]pyrene; O, oxidation; R,removal; M, mineralization; ND, not determined; –, no activity according to the assay(s) conducted for this substrate. Only oxidation wastested for IND, NTL, 1MN, 2MN, FLE, and ACN. Only substrate removal was tested for ATC and FLA. Both substrate removal andmineralization were tested for BaA, CRY, PYR, and BaP; mineralization is indicated if positive. All organisms mineralized phenanthreneexcept strain S1, which was not tested.
aMineralization assays were not conducted with this organism.bEvidence of metabolite formation was obtained by radiochromatography.cRemoval of FLE by strain P16 was reported by Stringfellow and Aitken (1995).
Table 6. Summary of results on PAH substrate oxidation, removal, and mineralization by bacterial isolates after growth inthe presence of phenanthrene.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Cal
if D
ig L
ib -
Law
renc
e B
erke
ley
Nat
iona
l Lab
orat
ory
(LB
L)
on 0
3/04
/13
For
pers
onal
use
onl
y.

We thank Paul Flathman (OHM Remediation ServicesInc.), Hans Stroo (Remediation Technologies Inc.), TalaatBalba (TreaTek-CRA Inc.), and Frederic Pfaender (Universityof North Carolina) for providing soil samples, and JuliaFoght (University of Alberta) for providing strain VT1. Wealso thank Randall Marx for his assistance in maintainingthe isolates. This work was supported by the NationalInstituteof Environmental Health Sciences (grant No. P42ES05948),the U.S. Geological Survey (award No. 14-08-0001-G2103)and the University of North Carolina Water Resources Re-search Institute (project No. 20162).
Boldrin, B., Tiehm, A., and Fritzsche, C. 1993. Degradation ofphenanthrene, fluorene, fluoranthene, and pyrene by aMycobac-terium sp. Appl. Environ. Microbiol.59: 1927–1930.
Bossert, I.D., and Bartha, R. 1986. Structure-biodegradability rela-tionships of polycyclic aromatic hydrocarbons in soil. Bull.Environ. Contam. Toxicol.37: 490–495.
Bouchez, M., Blanchet, D., and Vandecasteele, J.P. 1995. Degrada-tion of polycyclic aromatic hydrocarbons by pure strains and bydefined strain associations: inhibition phenomena andcometabolism. Appl. Microbiol. Biotechnol.43: 156–164.
Burd, G., and Ward, O.P. 1996. Involvement of a surface-activehigh molecular weight factor in degradation of polycyclic aro-matic hydrocarbons byPseudomonas marginalis. Can. J.Microbiol. 42: 791–797.
Carmichael, L.M., and Pfaender, F.K. 1997. Polynuclear aromatichydrocarbon metabolism in soils: relationship to soil character-istics and preexposure. Environ. Toxicol. Chem.16: 666–675.
Cerniglia, C.E., and Heitkamp, M.A. 1989. Microbial degradationof polycyclic aromatic hydrocarbons (PAH) in the aquatic envi-ronment.In Metabolism of polycyclic aromatic hydrocarbons inthe aquatic environment.Edited by U. Varanasi. CRC Press,Boca Raton, Fla. pp. 41–68.
Dagher, F., Déziel, E., Lirette, P., Paquette, G., Bisaillon, J.-G., andVillemur, R. 1997. Comparative study of five polycyclic aro-matic hydrocarbon degrading bacterial strains isolated from con-taminated soils. Can. J. Microbiol.43: 368–377.
Dean-Ross, D., and Cerniglia, C.E. 1996. Degradation of pyreneby Mycobacterium flavescens. Appl. Microbiol. Biotechnol.46:307–312.
Foght, J.M., and Westlake, D.W.S. 1988. Degradation of polycyclicaromatic hydrocarbons and aromatic heterocycles by aPseudo-monasspecies. Can. J. Microbiol.34: 1135–1141.
Foght, J.M., and Westlake, D.W.S. 1991. Cross hybridization ofplasmid and genomic DNA from aromatic and polycyclic aro-matic hydrocarbon degrading bacteria. Can. J. Microbiol.37:924–932.
Gibson, D., Mahadevan, V., Jerina, D., Yagi, H., and Yeh, H.J.C.1975. Oxidation of the carcinogens benzo[a]pyrene andbenzo[a]anthracene to dihydrodiols by a bacterium. Science(Washington, D.C.),189: 295–297.
Grifoll, M., Selifonov, S.A., Gatlin, C.V., and Chapman, P.J. 1995.Actions of a versatile fluorene-degrading bacterial isolate onpolycyclic aromatic compounds. Appl. Environ. Microbiol.61:3711–3723.
Grimberg, S.J., Stringfellow, W.T., and Aitken, M.D. 1996. Quan-tifying the biodegradation of phenanthrene byPseudomonas
stutzeri P16 in the presence of a nonionic surfactant. Appl.Environ. Microbiol. 62: 2387–2392.
Grosser, R.J., Warshawsky, D., and Vestal, J.R. 1991. Indigenousand enhanced mineralization of pyrene, benzo[a]pyrene, andcarbazole in soils. Appl. Environ. Microbiol.57: 3462–3469.
Grosser, R.J., Warshawsky, D., and Vestal, J.R. 1995. Mineraliza-tion of polycyclic andN-heterocyclic aromatic compounds inhydrocarbon-contaminated soils. Environ. Toxicol. Chem.14:375–382.
Heitkamp, M.A., and Cerniglia, C.E. 1989. Polycyclic aromatichydrocarbon degradation by aMycobacteriumsp. in micro-cosms containing sediment and water from a pristine ecosystem.Appl. Environ. Microbiol.55: 1968–1973.
Heitkamp, M.A., Freeman, J.P., Miller, D.W., and Cerniglia, C.E.1988. Pyrene degradation by aMycobacteriumsp.: identifica-tion of ring oxidation and ring fission products. Appl. Environ.Microbiol. 54: 2556–2565.
Herbes, S.E., and Schwall, L.R. 1978. Microbial transformation ofpolycyclic aromatic hydrocarbons in pristine and petroleum-contaminated sediments. Appl. Environ. Microbiol.35: 306–316.
Hommel, R.K. 1990. Formation and physiological role ofbiosurfactants produced by hydrocarbon-utilizing microorgan-isms. Biodegradation,1: 107–119.
Jiminez, I.Y., and Bartha, R. 1996. Solvent-augmented min-eralization of pyrene by aMycobacteriumsp. Appl. Environ.Microbiol. 62: 2311–2316.
Juhasz, A.L., Britz, M.L., and Stanley, G.A. 1997. Degradation offluoranthene, pyrene, benz[a]anthracene and dibenz[a,h]-anthracene byBurkholderia cepacia. J. Appl. Microbiol. 83:189–198.
Kästner, M., Breuer-Jammali, M., and Mahro, B. 1994. Enumera-tion and characterization of the soil microflora fromhydrocarbon-contaminated soil sites able to mineralizepolycyclic aromatic hydrocarbons (PAH). Appl. Microbiol.Biotechnol.41: 267–273.
Kelley, I., and Cerniglia, C.E. 1995. Degradation of a mixture ofhigh molecular-weight polycyclic aromatic hydrocarbons by aMycobacteriumstrain PYR-1. J. Soil Contam.4: 77–91.
Kelley, I., Freeman, J.P., Evans, F.E., and Cerniglia, C.E. 1991.Identification of a carboxylic acid metabolite from the catabo-lism of fluoranthene by aMycobacteriumsp. Appl. Environ.Microbiol. 57: 636–641.
Khan, A.A., Wang, R.-F., Cao, W.-W., Franklin, W., andCerniglia, C.E. 1996. Reclassification of a polycyclic aromatichydrocarbon-metabolizing bacterium,Beijerinckiasp. strain B1,asSphingomonas yanoikuyaeby fatty acid analysis, protein pat-tern analysis, DNA–DNA hybridization, and 16S ribosomalDNA sequencing. Int. J. Syst. Bacteriol.46: 466–499.
Kiyohara, H., and Nagao, K. 1978. The catabolism ofphenanthrene and naphthalene by bacteria. J. Gen. Microbiol.105: 69–75.
Kiyohara, H., Sugiyama, M., Mondello, F.J., Gibson, D.T., andYano, K. 1983. Plasmid involvement in the degradation ofpolycyclic aromatic hydrocarbons by aBeijerinckia species.Biochem. Biophys. Res. Commun.111: 939–945.
Mackay, D., Shia, Y.W., and Ma, K.C. 1992. Illustrated handbookof physical chemical properties and environmental fate for or-ganic chemicals. Vol. II. Lewis Publishers, Chelsea, Mich.
Mahaffey, W.R., Gibson, D.T., and Cerniglia, C.E. 1988. Bacterialoxidation of chemical carcinogens: formation of polycyclic aro-matic acids from benz[a]anthracene. Appl. Environ. Microbiol.54: 2415–2423.
© 1998 NRC Canada
Aitken et al. 751
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Cal
if D
ig L
ib -
Law
renc
e B
erke
ley
Nat
iona
l Lab
orat
ory
(LB
L)
on 0
3/04
/13
For
pers
onal
use
onl
y.

© 1998 NRC Canada
752 Can. J. Microbiol. Vol. 44, 1998
Millette, D., Barker, J.F., Comeau, Y., Butler, B.J., Frind, E.O.,Clement, B., and Samson, R. 1995. Substrate interaction duringaerobic biodegradation of creosote-related compounds: a facto-rial batch experiment. Environ. Sci. Technol.29: 1944–1952.
Mueller, J.G., Chapman, P.J., Blattmann, B.O., and Pritchard, P.H.1990. Isolation and characterization of a fluoranthene-utilizingstrain of Pseudomonas paucimobilis. Appl. Environ. Microbiol.56: 1079–1086.
Mueller, J.G., Lantz, S.E., Blattmann, B.O., and Chapman, P.J.1991a. Bench-scale evaluation of alternative biological treat-ment processes for the remediation of pentachlorophenol- andcreosote-contaminated materials: solid-phase bioremediation.Environ. Sci. Technol.25: 1045–1055.
Mueller, J.G., Lantz, S.E., Blattmann, B.O., and Chapman, P.J.1991b. Bench-scale evaluation of alternative biological treat-ment processes for the remediation of pentachlorophenol- andcreosote-contaminated materials: slurry-phase bioremediation.Environ. Sci. Technol.25: 1055–1061.
Mueller, J.G., Lantz, S.E.,Devereux, R., Berg,J.D., andPritchard, P.H.1994. Studies on the microbial ecology of polycyclic aromatichydrocarbon biodegradation.In Bioremediation of chlorinatedand polycyclic aromatic hydrocarbon compounds.Edited byR.E. Hinchee, A. Leeson, L. Semprini, and S.K. Ong. LewisPublishers, Boca Raton, Fla. pp. 218–230.
Rogoff, M.H. 1962. Chemistry of oxidation of polycyclic aromatichydrocarbons by soil pseudomonads. J. Bacteriol.83: 998–1004.
Schneider, J., Grosser, R., Jayasimhulu, K., Xue, W., andWarshawsky, D. 1996. Degradation of pyrene, benz[a]-anthracene, and benzo[a]pyrene by Mycobacteriumsp. strainRJGII-135, isolated from a former coal gasification site. Appl.Environ. Microbiol. 62: 13–19.
Shiaris, M.P. 1989. Seasonal biotransformation of naphthalene,phenanthrene, and benzo[a]pyrene in surficial estuarine sedi-ments. Appl. Environ. Microbiol.55: 1391–1399.
Shuttleworth, K.L., and Cerniglia, C.E. 1996. Bacterial degradationof low concentrations of phenanthrene and inhibition by naph-thalene. Microb. Ecol.31: 305–317.
Stringfellow, W.T., and Aitken, M.D. 1994. Comparative physiol-ogy of phenanthrene degradation by two dissimilar pseudo-monads isolated form a creosote-contaminated soil. Can. J.Microbiol. 40: 432–438.
Stringfellow, W.T., and Aitken, M.D. 1995. Competitive metabo-lism of naphthalene, methylnaphthalenes, and fluorene byphenanthrene-degrading pseudomonads. Appl. Environ. Micro-biol. 61: 357–362.
Sutherland, J.B., Rafii, F., Khan, A.A., and Cerniglia, C.E. 1995.Mechanisms of polycyclic aromatic hydrocarbon degradation.InMicrobial transformation and degradation of toxic organicchemicals.Edited by L.Y. Young and C.E. Cerniglia. Wiley-Liss, New York. pp. 269–306.
U.S. Environmental Protection Agency. 1996. Innovative treatmenttechnologies: annual status report. 8th ed. EPA-542-R-96-010.U.S. Environmental Protection Agency, Washington, D.C.
Walter, U., Beyer, M., Klein, J., and Rehm, H.-J. 1991. Degrada-tion of pyrene by Rhodococcussp. UW1. Appl. Microbiol.Biotechnol.34: 671–676.
Weissenfels, W.D., Beyer, M., Klein, J., and Rehm, H.J. 1991. Mi-crobial metabolism of fluoranthene: isolation and identificationof ring fission products. Appl. Microbiol. Biotechnol.34: 528–535.
Weissenfels, W.D., Klewer, H.-J., and Langhoff, J. 1992. Adsorp-tion of polycyclic aromatic hydrocarbons (PAHs) by soil parti-cles: influence on biodegradability and biotoxicity. Appl.Microbiol. Biotechnol.36: 689–696.
Wild, S.R., and Jones, K.C. 1993. Biological and abiotic losses ofpolynuclear aromatic hydrocarbons (PAHs) from soils freshlyamended with sewage sludge. Environ. Toxicol. Chem.12: 5–12.
Ye, D., Siddiqi, M.A., Maccubbin, A.E., Kumar, S., and Sikka, H.C.1996. Degradation of polynuclear aromatic hydrocarbons bySphingomonas paucimobilis. Environ. Sci. Technol.30: 136–142.
Can
. J. M
icro
biol
. Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Cal
if D
ig L
ib -
Law
renc
e B
erke
ley
Nat
iona
l Lab
orat
ory
(LB
L)
on 0
3/04
/13
For
pers
onal
use
onl
y.

This article has been cited by:
1. Flor de Ma. Guillén-Jiménez, Eliseo Cristiani-Urbina, Juan C. Cancino-Díaz, Jorge L. Flores-Moreno, Blanca E. Barragán-Huerta. 2012. Lindane biodegradation by the Fusarium verticillioides AT-100 strain, isolated from Agave tequilana leaves: Kineticstudy and identification of metabolites. International Biodeterioration & Biodegradation 74, 36-47. [CrossRef]
2. Xinying Zhang, Xiaoyan Liu, Chenglin Zhong, Zhengnan Cao, Fahui Liu, Lisha Chen, Shanshan Liu, Yan Hu. 2012. Soilmicrobial community response to pyrene at the presence of Scirpus triqueter. European Journal of Soil Biology 50, 44-50. [CrossRef]
3. Shuang Liu, Yanwei Hou, Guoxin Sun. 2012. Synergistic degradation of pyrene and volatilization of arsenic by cocultures ofbacteria and a fungus. Frontiers of Environmental Science & Engineering . [CrossRef]
4. Aruliah Rajasekar, Rajasekhar Balasubramanian, Joshua VM Kuma. 2011. Role of Hydrocarbon Degrading Bacteria Serratiamarcescens ACE2 and Bacillus cereus ACE4 on Corrosion of Carbon Steel API 5LX. Industrial & Engineering Chemistry Research50:17, 10041-10046. [CrossRef]
5. Brice Louvel, Aurélie Cébron, Corinne Leyval. 2011. Root exudates affect phenanthrene biodegradation, bacterial community andfunctional gene expression in sand microcosms. International Biodeterioration & Biodegradation . [CrossRef]
6. G.K. Parshetti, S.G. Parshetti, A.A. Telke, D.C. Kalyani, R.A. Doong, S.P. Govindwar. 2011. Biodegradation of Crystal Violetby Agrobacterium radiobacter. Journal of Environmental Sciences 23:8, 1384-1393. [CrossRef]
7. Vassilios Raikos, Sotiris S. Vamvakas, John Kapolos, Athanasia Koliadima, George Karaiskakis. 2011. Identification andcharacterization of microbial contaminants isolated from stored aviation fuels by DNA sequencing and restriction fragment lengthanalysis of a PCR-amplified region of the 16S rRNA gene. Fuel 90:2, 695-700. [CrossRef]
8. Diana V. Cortés-Espinosa, Ángel E. Absalón, Noé Sanchez, Octavio Loera, Refugio Rodríguez-Vázquez, Francisco J. Fernández.2011. Heterologous Expression of Manganese Peroxidase in <i>Aspergillus niger</i> and Its Effect on Phenanthrene Removalfrom Soil. Journal of Molecular Microbiology and Biotechnology 21:3-4, 120-129. [CrossRef]
9. Christine Lors, Annemie Ryngaert, Frédéric Périé, Ludo Diels, Denis Damidot. 2010. Evolution of bacterial community duringbioremediation of PAHs in a coal tar contaminated soil. Chemosphere 81:10, 1263. [CrossRef]
10. Evelyn Doyle, Lorraine Muckian, Anne Marie Hickey, Nicholas Clipson 65, 27. [CrossRef]11. David R. Singleton, Mark Hunt, Sabrina N. Powell, Roberto Frontera-Suau, Michael D. Aitken. 2007. Stable-isotope probing
with multiple growth substrates to determine substrate specificity of uncultivated bacteria. Journal of Microbiological Methods 69:1,180-187. [CrossRef]
12. E BRITO, R GUYONEAUD, M GONIURRIZA, A RANCHOUPEYRUSE, A VERBAERE, M CRAPEZ, J WASSERMAN,R DURAN. 2006. Characterization of hydrocarbonoclastic bacterial communities from mangrove sediments in Guanabara Bay,Brazil. Research in Microbiology 157:8, 752-762. [CrossRef]
13. M MOHAMED, M ALDOUSARY, R HAMZAH, G FUCHS. 2006. Isolation and characterization of indigenous thermophilicbacteria active in natural attenuation of bio-hazardous petrochemical pollutants. International Biodeterioration & Biodegradation58:3-4, 213-223. [CrossRef]
14. F.L. Toledo, C. Calvo, B. Rodelas, J. González-López. 2006. Selection and identification of bacteria isolated from waste crude oilwith polycyclic aromatic hydrocarbons removal capacities. Systematic and Applied Microbiology 29:3, 244-252. [CrossRef]
15. Mukesh Doble, Anil Kumar 241. [CrossRef]16. C.L. Guo, H.W. Zhou, Y.S. Wong, N.F.Y. Tam. 2005. Isolation of PAH-degrading bacteria from mangrove sediments and their
biodegradation potential. Marine Pollution Bulletin 51:8-12, 1054-1061. [CrossRef]17. L Ruberto. 2003. Effectiveness of the natural bacterial flora, biostimulation and bioaugmentation on the bioremediation of a
hydrocarbon contaminated Antarctic soil. International Biodeterioration & Biodegradation 52:2, 115-125. [CrossRef]18. William T. Stringfellow, Keun-Chan Oh. 2002. Initiation of MTBE Biotreatment in Fluidized-Bed Bioreactors. Journal of
Environmental Engineering 128:9, 852-861. [CrossRef]19. S. W. Rogers, S. K. Ong, B. H. Kjartanson, J. Golchin, G. A. Stenback. 2002. Natural Attenuation of Polycyclic Aromatic
Hydrocarbon-Contaminated Sites: Review. Practice Periodical of Hazardous, Toxic, and Radioactive Waste Management 6:3, 141.[CrossRef]
20. Albert L Juhasz, Ravendra Naidu. 2000. Bioremediation of high molecular weight polycyclic aromatic hydrocarbons: a review ofthe microbial degradation of benzo[a]pyrene. International Biodeterioration & Biodegradation 45:1-2, 57-88. [CrossRef]C
an. J
. Mic
robi
ol. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y C
alif
Dig
Lib
- L
awre
nce
Ber
kele
y N
atio
nal L
abor
ator
y (L
BL
) on
03/
04/1
3Fo
r pe
rson
al u
se o
nly.




















