
The Orbitofacial Glands of Bats: An Investigation of the Potential Correlation of Gland Structure...
16
THE ANATOMICAL RECORD 293:1433–1448 (2010) The Orbitofacial Glands of Bats: An Investigation of the Potential Correlation of Gland Structure with Social Organization SUSAN J. REHOREK, 1 * TIMOTHY D. SMITH, 2,3 AND KUNWAR P. BHATNAGAR 3,4 1 Department of Biology, Slippery Rock University, Slippery Rock, Pennsylvania 2 School of Physical Therapy, Slippery Rock University, Slippery Rock, Pennsylvania 3 Carnegie Museum of Natural History, Section of Mammals, Pittsburgh, Pennsylvania 4 Department of Anatomical Sciences and Neurobiology, School of Medicine, University of Louisville, Louisville, Kentucky ABSTRACT The facial glands of bats are modified skin glands, whereas there are up to three different orbital glands: Harderian, lacrimal, and Meibomian glands. Scattered studies have described the lacrimal and Meibomian glands in a handful of bat species, but there is as yet no description of a Harderian gland in bats. In this study we examined serial sections of orbitofacial glands in eight families of bats. Much variation amongst spe- cies was observed, with few phylogenetic patterns emerging. Enlarged fa- cial glands, either sudoriparous (five genera) or sebaceous (vespertilionids only) were observed. Meibomian and lacrimal glands were present in most species examined (except Antrozous), though the relative level of development varied. Two types of anterior orbital glands were distin- guished: the Harderian gland (tubulo-acinar: observed in Rousettus, Atri- beus, Desmodus and Miniopterus) and caruncular (sebaceous: observed in Eptesicus and Dieamus). The relative development of the nasolacrimal duct and the vomeronasal organ did not appear to be correlated with the development of any of the exocrine glands examined. There does, how- ever, appear to be a correlation between the presence of at least one well developed exocrine gland and the level of communality and known olfac- tory acuity, best documented in Artibeus, Desmodus, and Miniopterus. Anat Rec, 293:1433–1448, 2010. V V C 2010 Wiley-Liss, Inc. Key words: bats; caruncular sebaceous gland; Harderian gland; lacrimal gland; Meibomian gland; orbitofacial glands; social organization Very little is known about the cranial exocrine glands of the head in bats. Two main groups of such glands are those in the facial and orbital regions (Fig. 1). Some in- formation is available about the relative development of facial glands in bats (Quay, 1970; Kulzer et al., 1985; Haffner, 1995, 2000), though most of this literature focuses on the glands in the mandibular region (chin, gular, intermandibular: see Caspers et al., 2009), with relatively little attention paid to the glands in the maxil- lary region. Generally speaking, the facial glands are modified sweat (sudoriparous) or sebaceous glands. There are three exocrine skin gland types, classified by mechanism of secretion and opening of duct. Eccrine (merocrine secretion) sweat glands release their contents onto the surface of the skin. Apocrine sweat and seba- ceous (holocrine) glands release their secretions into the *Correspondence to: Susan J. Rehorek, Department of Biol- ogy, Slippery Rock University, Slippery Rock, PA 16057-1326. Fax: þ724 738 4782. E-mail: [email protected] Received 12 August 2009; Accepted 27 August 2009 DOI 10.1002/ar.21046 Published online 19 July 2010 in Wiley InterScience (www. interscience.wiley.com). V V C 2010 WILEY-LISS, INC.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The Orbitofacial Glands of Bats: An Investigation of the Potential Correlation of Gland Structure...

THE ANATOMICAL RECORD 293:1433–1448 (2010)
The Orbitofacial Glands of Bats: AnInvestigation of the Potential
Correlation of Gland Structure withSocial Organization
SUSAN J. REHOREK,1* TIMOTHY D. SMITH,2,3 AND KUNWAR P. BHATNAGAR3,4
1Department of Biology, Slippery Rock University, Slippery Rock, Pennsylvania2School of Physical Therapy, Slippery Rock University, Slippery Rock, Pennsylvania
3Carnegie Museum of Natural History, Section of Mammals, Pittsburgh, Pennsylvania4Department of Anatomical Sciences and Neurobiology, School of Medicine,
University of Louisville, Louisville, Kentucky
ABSTRACTThe facial glands of bats are modified skin glands, whereas there are
up to three different orbital glands: Harderian, lacrimal, and Meibomianglands. Scattered studies have described the lacrimal and Meibomianglands in a handful of bat species, but there is as yet no description of aHarderian gland in bats. In this study we examined serial sections oforbitofacial glands in eight families of bats. Much variation amongst spe-cies was observed, with few phylogenetic patterns emerging. Enlarged fa-cial glands, either sudoriparous (five genera) or sebaceous (vespertilionidsonly) were observed. Meibomian and lacrimal glands were present inmost species examined (except Antrozous), though the relative level ofdevelopment varied. Two types of anterior orbital glands were distin-guished: the Harderian gland (tubulo-acinar: observed in Rousettus, Atri-beus, Desmodus and Miniopterus) and caruncular (sebaceous: observed inEptesicus and Dieamus). The relative development of the nasolacrimalduct and the vomeronasal organ did not appear to be correlated with thedevelopment of any of the exocrine glands examined. There does, how-ever, appear to be a correlation between the presence of at least one welldeveloped exocrine gland and the level of communality and known olfac-tory acuity, best documented in Artibeus, Desmodus, and Miniopterus.Anat Rec, 293:1433–1448, 2010. VVC 2010 Wiley-Liss, Inc.
Keywords: bats; caruncular sebaceous gland; Harderian gland;lacrimal gland; Meibomian gland; orbitofacial glands;social organization
Very little is known about the cranial exocrine glandsof the head in bats. Two main groups of such glands arethose in the facial and orbital regions (Fig. 1). Some in-formation is available about the relative development offacial glands in bats (Quay, 1970; Kulzer et al., 1985;Haffner, 1995, 2000), though most of this literaturefocuses on the glands in the mandibular region (chin,gular, intermandibular: see Caspers et al., 2009), withrelatively little attention paid to the glands in the maxil-lary region. Generally speaking, the facial glands aremodified sweat (sudoriparous) or sebaceous glands.There are three exocrine skin gland types, classified by
mechanism of secretion and opening of duct. Eccrine(merocrine secretion) sweat glands release their contentsonto the surface of the skin. Apocrine sweat and seba-ceous (holocrine) glands release their secretions into the
*Correspondence to: Susan J. Rehorek, Department of Biol-ogy, Slippery Rock University, Slippery Rock, PA 16057-1326.Fax: þ724 738 4782. E-mail: [email protected]
Received 12 August 2009; Accepted 27 August 2009
DOI 10.1002/ar.21046Published online 19 July 2010 in Wiley InterScience (www.interscience.wiley.com).
VVC 2010 WILEY-LISS, INC.

hair follicle. These glands together appear to produce acompound which plays a role in chemical communication(scent marking etc.) (see Castaneda et al., 2007 forreview).
Considerably less is known about the orbital glands ofbats. In other mammals there are typically three maintypes of large orbital glands present: lacrimal, Meibo-mian (tarsal) and Harderian. Of these the lacrimal glandis usually found in the outer canthus, though it may bepresent as a palpebral structure in some tetrapods(Rehorek et al., 2005a). This gland has been examinedonly in one species of bat, Hipposideros speoris (Koradand Joshi, 1992). The Meibomian (or tarsal) glands arecommonly found in the eyelids. The nomenclature ofthese glands appears to be interchangeable, and their
association with the tarsus of the eyelid (Gartner andHiatt, 2007) has often led to them being referred to astarsal glands. There are no known descriptions of thebat Meibominan glands. In general, the secretions ofboth the lacrimal and Meibomian glands are presumedto be a source of lubrication for the cornea, the formerproducing tears (functioning to protect and moisten thesurface of the cornea and associated areas) and the lat-ter creating an oily layer on the tear film to reduce evap-oration rates (Gartner and Hiatt, 2007).
The Harderian gland is an anterior orbital gland, oftenassociated with the nictitating membrane. It is present inmost terrestrial vertebrates, but the function of this glandremains elusive (see Payne, 1994 for review). It has beenproposed that the Harderian gland may play a role in thevomeronasal system: this is suggested by the anatomicalconnection between the Harderian gland and the vomero-nasal organ (VNO) via the nasolacrimal duct (NLD) whichcan be observed in many tetrapods (Hillenius andRehorek, 2005). Harderian gland secretions are known topass into the VNO, via the NLD in caecilians (Wake,1985), snakes (Rehorek et al., 2000) and may also do so infrogs (Hillenius et al., 2001). In mammals, however, theconnections of these three structures are less clear (e.g.,Rossie and Smith, 2007). Not all mammals possess Har-derian glands (Paule, 1957; Sakai, 1981), although insome, the Harderian gland was only recently discovered.For example, a recent re-examination of primates showedthe presence of a Harderian gland across the order(Rehorek and Smith, 2006). This raises the question ofwhether the Harderian glands of bats, like that of prima-tes, may have been overlooked in earlier reports. The soleprevious study on bats examined only a few specimenswith little or no detailed description of the methodologyused. There were only two species of bats (Desmodusrotundus and Myotis lucifugus) examined by Paule (1957)with no Harderian gland observed in either one. In prima-tes, histological examination was necessary before thepresence of the Harderian gland was discovered (Rehorekand Smith, 2006). Consequently, careful histological ex-amination is needed before the presence or absence of thisgland in bats is deduced. Considerably more is knownabout the NLD (Gobbel, 2002) and the VNO (Cooper andBhatnagar, 1976; Wible and Bhatnagar, 1996) of bats.
The purpose of this study was to examine the relativedevelopment of facial and orbital glands in a wide phylo-genetic survey of bats. Bats, which may exceed 1,100species (Locke, 2008), are highly diverse especially interms of facial morphology (Fenton, 2001). Thus anygeneralizations about gland structure are not plausibleuntil a significant number of bats is examined.
MATERIALS AND METHODS
Coronal serial sections of the heads of 20 adult and 5fetal bat specimens (21 species) were examined (Table1). This sample included representative species fromeach of the three major groupings of bats: Pteropodid(Pteropodidae, Rhinopomatidae, Megadermatidae, Rhi-nolophidae and Hipposideridae: 5 species) and the twoderived lineages of Phyllostomatidae/Mystacinaidae (9species) and Vespertilionidae (7 species; Simmons, 2005).The sections were drawn from the vast collection of seri-ally-sectioned heads of bats from the laboratory of one ofthe authors (KPB). This collection was established over
Fig. 1. Schematic overview of orbitofacial gland location in batstransposed upon a skull (outlined in white) and soft-tissues (grey) ofArtibeus jamaicensis. (A) shows the dorsal view showing the locationof all glands. Note that the Harderian gland (HG) is the most medialgland, residing in the inferoanterior region of the orbit. The lacrimalgland (LG) lies medial to the postorbital glandular region (PoG) andthe Meibomiam glands (MG). (B) shows the lateral view, and the rela-tive position of the three most cutaneous glandular regions. Abbrevia-tions: E ¼ eye; PrG ¼ preorbital (pararhinal) glandular area. Not toscale.
1434 REHOREK ET AL.

a period of 50 years with specimens donated from allover the world and specimens collected by KPB in thefield. Donated specimens were typically preserved in10% formalin or ethyl alcohol. Whereas specimens col-
lected by KPB were always perfused cardially with ei-ther Bouin’s fluid, 10% formalin, 4% glutaraldehyde, orother fixatives. The tissues needed were dissected (eitherin the field and/or in the laboratory) and decalcified
TABLE 2. Distribution of orbital and facial glands in bats
Species FG MG LG
PteropodidRousettus leschenaulti ? (fetus) ? fetus þ (fetus)Rhinolophus lepidus ESW a: U þ L UHipposideros lankadiva N a: U þ L FMegaderma lyra N b: L LFRhinopoma microphyllum LSW b: U þ L T þ FMystacinidae/PhyllostomidaeMystacina tuberculata N d: U þ L T þ FDesmodus rotundus LSW b: U þ L T þ FDiaemus youngii N c: U þ L T þ FDiphylla ecaudata ? (fetus) b: U T þ FBrachyphylla cavernarum ? (fetus) ? fetus þ (fetus)Anoura geoffroyi N d: U þ L SOMacrotus waterhousii N c: U þ L FArtibeus jamaicensis LSW d: U þ L T þ FVespertilionidaeEptesicus fuscus CSB b: U þ L T þ FAntrozous pallidus CSB b: U þ L –sMyotis lucifugus CSB b: U þ L FMiniopterus (all 3 spp.) LSW þ LSB b: U þ L SO þ T
Phylogenetic order arranged according to Simmons, 2005.Abbreviations: FG ¼ facial glands (CSB ¼ clustered sebaceous glands; ESW ¼ elongate sweatgland; LSB ¼ enlarged sebaceous glands with multiple lobules; LSW ¼ sweat glands in lobules;N ¼ same as normal skin); MG ¼ Meibomian glands (a ¼ multiple normal sized sebaceousglands; b ¼ multiple slightly enlarged sebaceous glands; c ¼ 1 clustered sebaceous secretoryunit; d ¼ 3–4 clusters of sebaceous secretory units; L ¼ lower palpebra; U ¼ upper palpebra);LG ¼ lacrimal gland (F ¼ fused palpebrae; LF ¼ lower palpebra with continuation into fusedpalpebra; SO ¼ supraorbital; SO þ T ¼ supraorbital lobe with temporal lobe; T þ F ¼ temporallobe with small opening into fused palpebrae; U ¼ upper palprebral).
TABLE 1. List of specimens examined
Family Species Common name Fetal Adult
PteropodidPteropodidae Rousettus leschenaulti Rousette fruit bat Fetal (N ¼ 2)Rhinolophidae Rhinolophus lepidus Horseshoe bat Adult (N ¼ 1)Hipposideridae Hipposideros lankadiva Old world leaf-nosed bat Adult (N ¼ 1)Megadermatidae Megaderma lyra Asian false vampire bat Adult (N ¼ 1)Rhinopomatidae Rhinopoma microphyllum Mouse-tailed or long-tailed bat Adult (N ¼ 1)Mystacinidae/PhyllostomidaeMystacinidae Mystacina tuberculata New Zealand short-tailed bat Adult (N ¼ 1)Phyllostomidae Desmodus rotundus Common vampire bat Adult (N ¼ 1)
Desmodontinae Diaemus youngii White-winged vampire bat Adult (N ¼ 1)Diphylla ecaudata hairy-legged vampire bat Fetal (N ¼ 1)
Brachyphyllinae Brachyphylla cavernarum Antillean fruit-eating bat Fetal (N¼ 1)Glossophaginae Anoura geoffroyi Geoffroy’s long-nosed bat Adult (N ¼ 1) and
fetus (N ¼ 1)Anoura cultrata Handley’s tailless bat Adult (N ¼ 1)
Phyllostominae Macrotus waterhousii Big-eared bat Adult (N ¼ 2)Stenodermatinae Artibeus jamaicensis Neotropical fruit bat Adult (N ¼ 1)
VespertilionidaeVespertilioninae Eptesicus fuscus House bat Adult (N ¼ 2)
Pipistrellus subflavus Pipistrelle Adult (N ¼ 1)Antrozoinae Antrozous pallidus Pallid bat Adult (N¼ 1)Myotinae Myotis lucifugus Little brown bat Adult (N ¼ 1)Miniopterinae Miniopterus australis little long-fingered bat Adult (N ¼ 1)
Miniopterus magnater Western bent-winged bat Adult (N ¼1)Miniopterus schreibersi Schreibers’s long-fingered bat Adult (N ¼ 1)
Arranged in phylogenetic order as per Simmons (2005).
BAT ORBITOFACIAL GLANDS 1435

using formic acid-sodium citrate solution (for details, seeBhatnagar and Kallen 1974a), processed for paraffinembedding, and carried through 10 lm serial sections ofthe whole head. They were then stained with theGomori trichrome procedure and other techniques.
RESULTSFacial Glands
Facial glands, in close proximity to the orbital regionwere examined. The skin in this region contained vary-ing levels of development of both apocrine sweat and se-baceous glands. Both types are associated with the hairfollicles. There was no evidence of eccrine sweat glandson the face. There were five different conditionsobserved in the relative development of these cutaneousglands (see Table 2 for summary). (1) Hipposideros lan-kadiva (Fig. 2A) represents the unmodified condition, inthat the sebaceous glands are simple branched acinarglands, and the apocrine sweat glands are coiled tubularglands, the endpieces of which pass between the seba-ceous glands. These endpieces lie perpendicular to thebases of the sebaceous glands and form a glandularlayer that is thinner than that of the sebaceous glands.This is the same condition seen in the skin on the head,and thus is referred to as unmodified. (2) Slightly elon-gate sweat glands were observed in the nasal, suboccu-lar and postoccular skin of Rhinolophus lepidus (Fig.2B). In this case, the sweat glands form identifiablelobules which are clearly separated by connective tissue.The apocrine sweat gland layer is thicker than that ofthe sebaceous gland layer. (3) Still larger apocrine sweatglands are found in the nasal, suboccular and postoccu-lar skin of both Rhinopoma microphyllum and Artibeusjamaicensis (Fig. 2C). In this case, the sweat glandsform identifiable lobules which are clearly separated byconnective tissue. In this case, the apocrine sweat glandlayer is much thicker than that observed in Rhinolophuslepidus.
The remaining two conditions are restricted to the ves-pertilionids and involved the expansion of the sebaceousglands. (4) This was observed in the superior nasalregions of Eptesicus fuscus, Antrozous pallidus and Myo-tis lucifugus (Fig. 3A) Sebaceous glands are arranged inclusters (or nests), whereby several cells open into alarger central lumen. These glandular clusters thenopened via a relatively short duct onto the surface of theskin. These sebaceous glands were not associated withhair follicles. A relatively thin layer of apocrine sweatglands, associated with hair follicles, is found deep tothese sebaceous clusters. (5) All three species of Miniop-terus, however, exhibit the expansion of both the seba-ceous and apocrine sweat glandular tissue. Enlarged,non-clustered sebaceous glands, each with multiplebranches (which open onto a hair follicle) lie superficialto the lobules of apocrine sweat glands spread through-out the skin. This occurs specifically in the nasal andperioccular regions. There is also a large suborbital cre-vasse, lined with these glands (Fig. 3B).
Orbital Glands
Eyelid glands: Meibomian glands. The Meibo-mian glands are the glands of the upper and lower eye-lids. They are all sebaceous. However, the bats examined
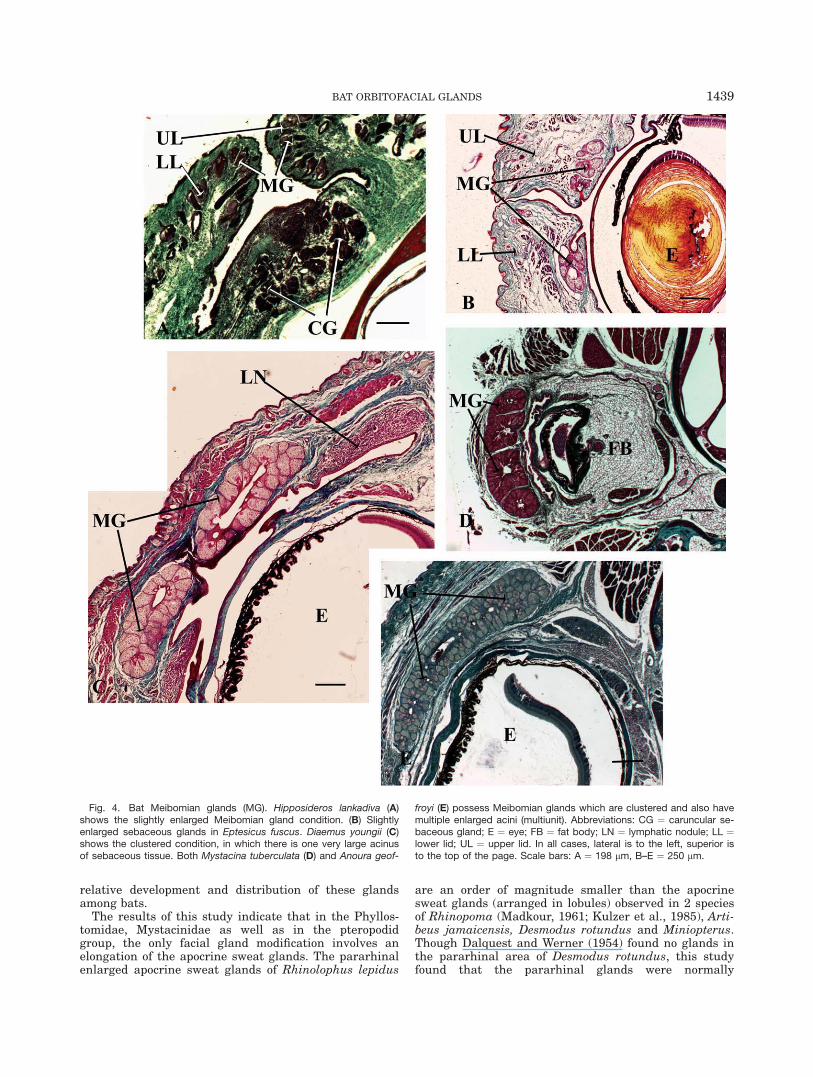
exhibit considerable variation with regards to the rela-tive development of these glands. No Meibomian glandswere observed in the eyelids of any fetal specimens. Atno stage were there more than four Meibomian glandsopening onto the either eyelid. The simplest and small-est Meibomian glands were the same sizes as, and,essentially continuous with the sebaceous glands on theouter part of the eyelid. This pattern is only found inRhinolophus lepidus and Hipposideros lankadiva (Fig.4A). Slightly larger specialized sebaceous glandsoccurred on the inner aspect of the eyelid in severalother species, including Megaderma lyra, Rhinopomamicrophyllum, Desmodus rotundus and in all vespertilio-nids (Fig. 4B). A single sebaceous cluster with a singleduct is observed in Macrotus waterhousii and Diaemusyoungii (Fig. 4C). Finally, a series of sebaceous clusters(3-4) with only 1 duct is observed in Mystacina tubercu-lata (Fig. 4D), Anoura geoffroyi and Artibeus jamaicensis(Fig. 4E).
Posterior orbital glands: Lacrimal gland. Withthe exception of Antrozous pallidus, all bats examined(both adult and fetal) possessed a lacrimal gland. Theadult extent and thus position of the lacrimal gland couldnot be determined in the two taxa of which only fetalspecimens were available (Rousettus leschenaulti andBrachyphylla cavernarum). The relative position of thegland with respect to the eyeball varied among species.Though it resides superficial to the eyeball, in the poste-rior aspect of the orbital region of most bats (with theexception of Anoura, in whom the lacrimal gland is asupraorbital structure and does not seem to be associatedwith the palpebrae), its association with the palpebrashows some variation. It is found in the upper palpebra ofRhinolophus lepidus (Fig. 5A) and in the fused posteriorpalpebral region of Hipposideros lankadiva, Macrotuswaterhousii and Myotis lucifugus (Fig. 5B). Megadermalyra has an enlarged lower lid, into which the lacrimalgland extends (as it also protrudes into the fused palpe-bral region: Fig. 5C). In the remaining bats, the lacrimalgland has a small connection to the palpebral region (Fig.5D). This is connected by a few small ducts to a large tem-poral glandular mass, which consists of several lobules.Miniopterus has both a supraorbital portion and a largetemporal region. In many cases, lymphatic tissue is asso-ciated with the lacrimal gland.
Anterior orbital glands: Harderian andcaruncular sebaceous glands. Several species ofbats possess a fat body deep to the eyeball (Fig. 6A). Inseveral specimens, there is a lymphatic aggregation inthe orbit anterior to the eyeball. There was little evi-dence of extra vascularization. The Harderian glandsare tubuloacinar glands, usually located in the anteriorportion of the orbit, but which may also have lobulespenetrating deep into the orbital region. There are alsoa series of sebaceous glands, simple branched acinar,that reside nearby, on the caruncle. Harderian glandswere observed in five species, possibly adding the twospecies with only fetal specimens examined (Rousettusleschenaulti and Brachyphylla cavernarum). In fetalRousettus, the Harderian gland appears to be a singletubular structure, extending into the medial orbitalregion. In fetal Brachyphylla, identification of the
1436 REHOREK ET AL.

Fig. 2. Facial glands: Hipposideros lankadiva (A) exhibits theunmodified condition, with small sebaceous (SB) and sweat (sudo-riferous: SD) glands peppered throughout the skin. Rhinolphus lep-idus (B) possess elongate sweat glands along the lateral aspectof the face (pararhinal). Sebaceous units unmodified. Artibeus
jamaicensis (C) possess lobulated sweat glands and unmodifiedsebaceous glands both pararhinal but mostly postorbital. Lateralto the left, superior to the top. Other abbrev: LG ¼ lacrimalgland; MG ¼ Meibomian gland. Scale areas: A ¼ 198 lm, B andC ¼ 250 lm.
BAT ORBITOFACIAL GLANDS 1437
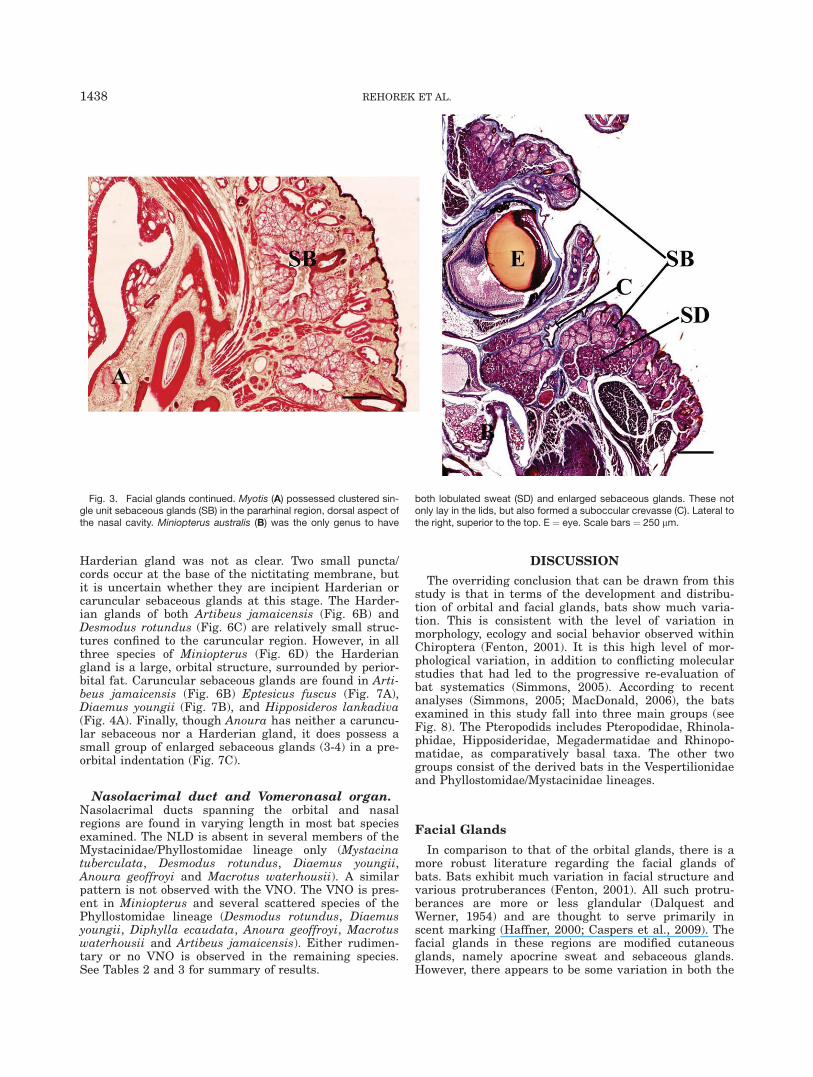
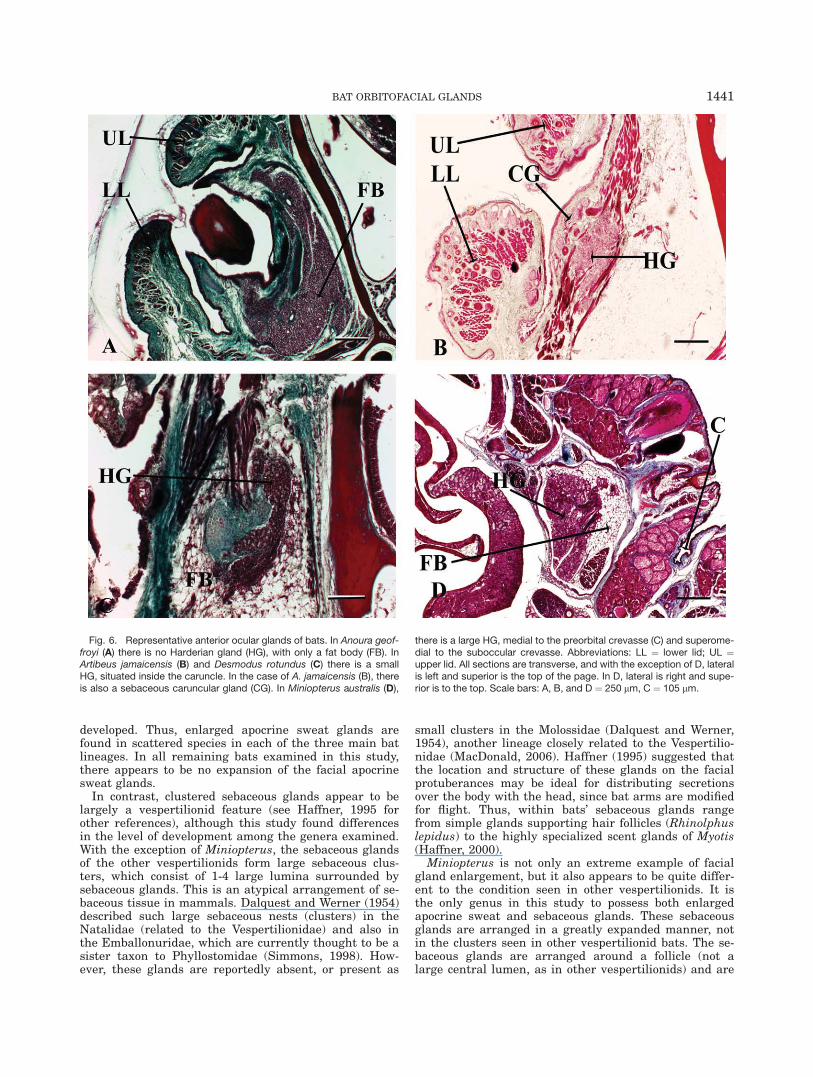
Harderian gland was not as clear. Two small puncta/cords occur at the base of the nictitating membrane, butit is uncertain whether they are incipient Harderian orcaruncular sebaceous glands at this stage. The Harder-ian glands of both Artibeus jamaicensis (Fig. 6B) andDesmodus rotundus (Fig. 6C) are relatively small struc-tures confined to the caruncular region. However, in allthree species of Miniopterus (Fig. 6D) the Harderiangland is a large, orbital structure, surrounded by perior-bital fat. Caruncular sebaceous glands are found in Arti-beus jamaicensis (Fig. 6B) Eptesicus fuscus (Fig. 7A),Diaemus youngii (Fig. 7B), and Hipposideros lankadiva(Fig. 4A). Finally, though Anoura has neither a caruncu-lar sebaceous nor a Harderian gland, it does possess asmall group of enlarged sebaceous glands (3-4) in a pre-orbital indentation (Fig. 7C).
Nasolacrimal duct and Vomeronasal organ.Nasolacrimal ducts spanning the orbital and nasalregions are found in varying length in most bat speciesexamined. The NLD is absent in several members of theMystacinidae/Phyllostomidae lineage only (Mystacinatuberculata, Desmodus rotundus, Diaemus youngii,Anoura geoffroyi and Macrotus waterhousii). A similarpattern is not observed with the VNO. The VNO is pres-ent in Miniopterus and several scattered species of thePhyllostomidae lineage (Desmodus rotundus, Diaemusyoungii, Diphylla ecaudata, Anoura geoffroyi, Macrotuswaterhousii and Artibeus jamaicensis). Either rudimen-tary or no VNO is observed in the remaining species.See Tables 2 and 3 for summary of results.
DISCUSSION
The overriding conclusion that can be drawn from thisstudy is that in terms of the development and distribu-tion of orbital and facial glands, bats show much varia-tion. This is consistent with the level of variation inmorphology, ecology and social behavior observed withinChiroptera (Fenton, 2001). It is this high level of mor-phological variation, in addition to conflicting molecularstudies that had led to the progressive re-evaluation ofbat systematics (Simmons, 2005). According to recentanalyses (Simmons, 2005; MacDonald, 2006), the batsexamined in this study fall into three main groups (seeFig. 8). The Pteropodids includes Pteropodidae, Rhinola-phidae, Hipposideridae, Megadermatidae and Rhinopo-matidae, as comparatively basal taxa. The other twogroups consist of the derived bats in the Vespertilionidaeand Phyllostomidae/Mystacinidae lineages.
Facial Glands
In comparison to that of the orbital glands, there is amore robust literature regarding the facial glands ofbats. Bats exhibit much variation in facial structure andvarious protruberances (Fenton, 2001). All such protru-berances are more or less glandular (Dalquest andWerner, 1954) and are thought to serve primarily inscent marking (Haffner, 2000; Caspers et al., 2009). Thefacial glands in these regions are modified cutaneousglands, namely apocrine sweat and sebaceous glands.However, there appears to be some variation in both the
Fig. 3. Facial glands continued. Myotis (A) possessed clustered sin-gle unit sebaceous glands (SB) in the pararhinal region, dorsal aspect ofthe nasal cavity. Miniopterus australis (B) was the only genus to have
both lobulated sweat (SD) and enlarged sebaceous glands. These notonly lay in the lids, but also formed a suboccular crevasse (C). Lateral tothe right, superior to the top. E ¼ eye. Scale bars ¼ 250 lm.
1438 REHOREK ET AL.
relative development and distribution of these glandsamong bats.
The results of this study indicate that in the Phyllos-tomidae, Mystacinidae as well as in the pteropodidgroup, the only facial gland modification involves anelongation of the apocrine sweat glands. The pararhinalenlarged apocrine sweat glands of Rhinolophus lepidus
are an order of magnitude smaller than the apocrinesweat glands (arranged in lobules) observed in 2 speciesof Rhinopoma (Madkour, 1961; Kulzer et al., 1985), Arti-beus jamaicensis, Desmodus rotundus and Miniopterus.Though Dalquest and Werner (1954) found no glands inthe pararhinal area of Desmodus rotundus, this studyfound that the pararhinal glands were normally
Fig. 4. Bat Meibomian glands (MG). Hipposideros lankadiva (A)shows the slightly enlarged Meibomian gland condition. (B) Slightlyenlarged sebaceous glands in Eptesicus fuscus. Diaemus youngii (C)shows the clustered condition, in which there is one very large acinusof sebaceous tissue. Both Mystacina tuberculata (D) and Anoura geof-
froyi (E) possess Meibomian glands which are clustered and also havemultiple enlarged acini (multiunit). Abbreviations: CG ¼ caruncular se-baceous gland; E ¼ eye; FB ¼ fat body; LN ¼ lymphatic nodule; LL ¼lower lid; UL ¼ upper lid. In all cases, lateral is to the left, superior isto the top of the page. Scale bars: A ¼ 198 lm, B–E ¼ 250 lm.
BAT ORBITOFACIAL GLANDS 1439
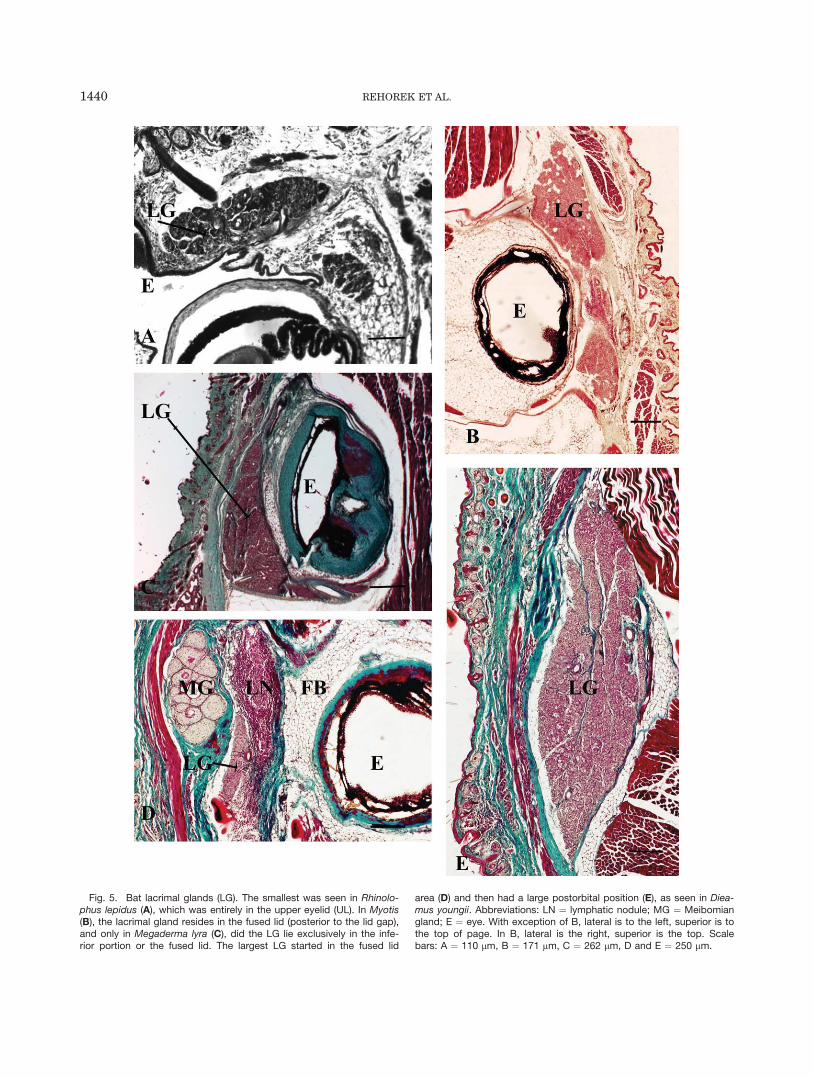
Fig. 5. Bat lacrimal glands (LG). The smallest was seen in Rhinolo-phus lepidus (A), which was entirely in the upper eyelid (UL). In Myotis(B), the lacrimal gland resides in the fused lid (posterior to the lid gap),and only in Megaderma lyra (C), did the LG lie exclusively in the infe-rior portion or the fused lid. The largest LG started in the fused lid
area (D) and then had a large postorbital position (E), as seen in Diea-mus youngii. Abbreviations: LN ¼ lymphatic nodule; MG ¼ Meibomiangland; E ¼ eye. With exception of B, lateral is to the left, superior is tothe top of page. In B, lateral is the right, superior is the top. Scalebars: A ¼ 110 lm, B ¼ 171 lm, C ¼ 262 lm, D and E ¼ 250 lm.
1440 REHOREK ET AL.
developed. Thus, enlarged apocrine sweat glands arefound in scattered species in each of the three main batlineages. In all remaining bats examined in this study,there appears to be no expansion of the facial apocrinesweat glands.
In contrast, clustered sebaceous glands appear to belargely a vespertilionid feature (see Haffner, 1995 forother references), although this study found differencesin the level of development among the genera examined.With the exception of Miniopterus, the sebaceous glandsof the other vespertilionids form large sebaceous clus-ters, which consist of 1-4 large lumina surrounded bysebaceous glands. This is an atypical arrangement of se-baceous tissue in mammals. Dalquest and Werner (1954)described such large sebaceous nests (clusters) in theNatalidae (related to the Vespertilionidae) and also inthe Emballonuridae, which are currently thought to be asister taxon to Phyllostomidae (Simmons, 1998). How-ever, these glands are reportedly absent, or present as
small clusters in the Molossidae (Dalquest and Werner,1954), another lineage closely related to the Vespertilio-nidae (MacDonald, 2006). Haffner (1995) suggested thatthe location and structure of these glands on the facialprotuberances may be ideal for distributing secretionsover the body with the head, since bat arms are modifiedfor flight. Thus, within bats’ sebaceous glands rangefrom simple glands supporting hair follicles (Rhinolphuslepidus) to the highly specialized scent glands of Myotis(Haffner, 2000).
Miniopterus is not only an extreme example of facialgland enlargement, but it also appears to be quite differ-ent to the condition seen in other vespertilionids. It isthe only genus in this study to possess both enlargedapocrine sweat and sebaceous glands. These sebaceousglands are arranged in a greatly expanded manner, notin the clusters seen in other vespertilionid bats. The se-baceous glands are arranged around a follicle (not alarge central lumen, as in other vespertilionids) and are
Fig. 6. Representative anterior ocular glands of bats. In Anoura geof-froyi (A) there is no Harderian gland (HG), with only a fat body (FB). InArtibeus jamaicensis (B) and Desmodus rotundus (C) there is a smallHG, situated inside the caruncle. In the case of A. jamaicensis (B), thereis also a sebaceous caruncular gland (CG). In Miniopterus australis (D),
there is a large HG, medial to the preorbital crevasse (C) and superome-dial to the suboccular crevasse. Abbreviations: LL ¼ lower lid; UL ¼upper lid. All sections are transverse, and with the exception of D, lateralis left and superior is the top of the page. In D, lateral is right and supe-rior is to the top. Scale bars: A, B, and D ¼ 250 lm, C ¼ 105 lm.
BAT ORBITOFACIAL GLANDS 1441

found scattered throughout the skin (both above andbelow the orbital region). A similar situation occurs inshrews (Mary and Balakrishnan, 1991), in which such
an expanded nasal glandular region is thought to func-tion in scent marking.
Additionally, Miniopterus also has a suborbital cre-vasse, which increases the surface area of the skin,thereby increasing number of glands in that region.Though this crevasse has not been previously describedin bats, a similar structure has been described in someungulate mammals, which have a pre-orbital sac, linedby modified skin (like that of Miniopterus) whose integu-mentary structures have undergone varying levels ofdevelopment (Schaffer, 1940). In the Chinese muntjac(Muntiacus reevesi, Cervidae), the preobital fossa is linedby an epithelium with enlarged apocrine sweat
Fig. 7. In Eptesicus (A) and Diaemus youngii (B), there was no Hgobserved, but there was a glandular caruncle (C). In Anoura geoffroyi(C) there are sebaceous glands in the preorbital indent. Abbreviations:LL ¼ lower lid; UL ¼ upper lid. All sections are transverse, and withthe exception of (D), lateral is to the left and superior is the top of thepage. In D, lateral is right and superior is to the top. Scale bars: A ¼105 lm, B ¼ 262 lm, C ¼ 95 lm.
TABLE 3. Distribution of nasolacrimal duct,vomeronasal organ, orbital and facial glands in bats
Species HG NLD VNO
PteropodidRousettus leschenaulti þ 2 �Rhinolophus lepidus � 2 RHipposideros lankadiva � 2 RMegaderma lyra � 2 RRhinopoma microphyllum � 2 RMystacinidae/PhyllostomidaeMystacina tuberculata � � RDesmodus rotundus þ � þDiaemus youngii � þDiphylla ecaudata � ? þBrachyphylla cavernarum ? 3? RAnoura geoffroyi � � þMacrotus waterhousii � � þArtibeus jamaicensis þ 3 þVespertilionidaeEptesicus fuscus � 0 �Antrozous pallidus � 0 �Myotis lucifugus � 0 �Miniopterus (all 3 spp.) þ 2 þPhylogenetic order arranged according to Simmons, 2005.Abbreviations: þ ¼ present; � ¼ absent; HG ¼ Harderiangland; NLD ¼ nasolacrimal duct (0 ¼ NLD opens into floorof nostril; 1 ¼ NLD opens into middle part of inferior mea-tus; 2 ¼ NLD opens near nasoplatine duct and/or VNO; 3 ¼incomplete); VNO ¼ vomeronasal organ (R ¼ rudimentary).
Fig. 8. A basic cladogam of the three major bat lineages as deter-mined by Simmons (1998, 2005) and MacDonald (2006).
1442 REHOREK ET AL.

(arranged in lobules) and sebaceous glands (Rehoreket al., 2005b) not unlike those of Miniopterus. Cervuseldi thamin (Cervidae) uses its preorbital gland in scentmarking (Muller-Schwarze, 1975); perhaps Miniopterusdoes something similar. Based on the structural similar-ity, it would be worthwhile to investigate whether Mini-opterus has similar marking behaviors.
Lacrimal and Meibomian Glands
Most of the bats examined (except Antrozous pallidus)are typical mammals in that they possess both lacrimaland Meibomian glands. There appears to be no discerni-ble phylogenetic pattern in the relative development ofthe lacrimal gland among bats as each of the threegroups includes species with both very large and smallerlacrimal glands. Furthermore, there is interspecific vari-ation within Hipposideros. Hipposideros speoris pos-sesses a lacrimal gland with a large exorbital region(Korad and Joshi, 1992), whereas Hipposideros lanka-diva possesses only a lacrimal gland restricted to theeyelid (palpebral lacrimal gland). Hipposideros is a veryspecious genus of bats, which in modern systemic analy-ses is often embedded within Rhinolophidae, anotherspecious genus of bats (Nowak, 1998). Thus, based uponthe level of variation in lacrimal gland size between batfamilies, variation within a family is not unexpected.
Apart from providing fluid for lubrication, the bat lacri-mal gland is also a part of the eye-associated lymphatictissue. The presence of lymphatic tissue in the orbitalregion has been described in several amniote tetrapods(Aitken and Survashe, 1977; Montgomery and Maslin,1992; Pinard et al., 2003; Rehorek et al., 2005a, 2006). Asin several other tetrapods the main site for lymphatic tis-sue is the Harderian gland (Aitken and Survashe, 1977;Montgomery and Maslin, 1992; Rehorek et al., 2005a,2006), the absence of the Harderian gland in bats mayhave resulted in a shift of the immunogenic propertiestowards the conjunctiva and lacrimal glands. Humansalso lack a Harderian gland, and, like bats, the lymphatictissue is found spread out in the conjunctiva and the lacri-mal gland (Gillett et al., 1980; Knop and Knop, 2000).
Most bats examined possess Meibomian glands. Excep-tions are Rousettus leschenaulti and Brachyphylla caver-narum, but because only fetal specimens of these specieswere examined, the presence of Meibomian glands inadults cannot be ruled out. For the other bats, althoughthere is some variation in relative development of theseglands, some generalizations can nevertheless be made.Firstly, the structure of the Meibomian glands in five spe-cies (all within the Mystacinidae/Phyllostomidae lineage)differed from that of other mammals, including primates(Montagna and Machida, 1966; Gartner and Hiatt, 2007),lemmings (Rodentia: Dearden, 1959) and both possumsand bandicoots (Marsupiala: Green, 1963). These bat Mei-bomian glands comprise of one to four sebaceous clusterswith a single excretory pore per eyelid, whereas in theaforementioned mammals, the Meibomian glands are aseries of small sebaceous glands. Whether the clusteredMeibomian glands observed in these five species of batproduce more sebum than the rows of Meibomian glandsobserved in other mammals remains to be determined.
Secondly, in the Pteropodid group, as well as in Des-modus rotundus and the Vespertilionidae, it appearsthat the Meibomian glands are either normal sized (sim-
ple branched acinar glands) or slightly enlarged seba-ceous glands. The single gland units observed in thesebats are morphologically similar to those single glandunits described in primates (Miraglia and Gomes, 1969;Stephens et al., 1989) and shrews (Mary and Balak-rishnan, 1991).
Thirdly, the divergence of Desmodus rotundus (withsmall Meibomian glands) and Diaemus youngii (withclustered Meibomian glands) may be interpreted as ei-ther glandular plasticity or bat variation at the inter-generic level. Similar variation was also noted in theprecise structure of the Meibomian glands of lemmings(Dearden, 1959).
Harderian and Caruncular Sebaceous Glands
There are two identifiably different glands in the ante-rior part of the orbit in some bat species. The caruncularsebaceous gland is a simple branched acinar gland, asso-ciated with a hair follicle in the region of the caruncle,which used a holocrine secretory mechanism. Thus it istermed the caruncular sebaceous gland. The distributionof this gland among bats follows no obvious phylogeneticpattern. It is present in only a few species, once againfrom each of the three groups of bats: Hipposideros lan-kadiva (allied with the pteropodid bats), Diaemusyoungii (in the more derived group of phyllostomid andtheir near relatives) and Eptesicus (vespertilionid).
In contrast, the Harderian gland is a tubuloacinargland using a merocrine secretory mechanism (Payne,1994) of variable size but larger than that of the seba-ceous glands, and is usually found deeper in the orbit.This distinction is clearly seen in Artibeus jamaicensis(Fig. 6B), which is thus far the only species in whichboth glands are present. As the Harderian gland is pres-ent in most terrestrial vertebrates, and thus a primitivecharacteristic for amniotes (see Payne, 1994 for review),the absence of this gland in any amniote would be aderived characteristic. However, the distribution of thisgland among bats does not neatly follow the currentview of phylogeny (see Simmons, 2005 for bat phylog-eny). It is present, and well developed, in members oftwo very different lineages, the basal Pteropodidae(Fruit bats: Rousettus leschenaulti) and the more derivedvespertilionids (Miniopterus spp.). In contrast it is rudi-mentary in two more distantly related Phyllostomids(Desmodus rotundus and Artibeus jamaicensis). How-ever, what is most striking about this distribution isthat none of the closest relatives of either Miniopterus(including other Vespertilionids such as Antrozous pal-lidus, Myotis lucifugus and Eptesicus fuscus) or Desmo-dus rotundus (including Diaemus youngii or any of theother Phyllostomids such as Anoura spp. and Macrotuswaterhousii) possess a Harderian gland. This gland isalso absent among bats allied with the Pteropodidae(e.g.: Rhinopoma microphyllum, Megaderma lyra, Rhino-lophus lepidus and Hipposideros lankadiva).
These observations indicate that Paule’s (1957) conclu-sion that bats have no Harderian glands (based on a sin-gle vespertilionid Myotis and a single PhyllostomidDesmodus rotundus) is incorrect on at least two levels.Firstly, based upon the aforementioned variation in batmorphology, generalizations based on two specimens(Paule, 1957) will not give accurate results. Secondly, thecontradiction between the observations of Paule (1957)
BAT ORBITOFACIAL GLANDS 1443

and this study, regarding the presence of a Harderiangland in Desmodus rotundus, may be due to samplingtechnique. Although the precise technique used by Paule(1957) is not known, it has been shown that serial sectionsare the best way to determine the presence of a Harderiangland, especially if it is small or rudimentary (Rehorekand Smith, 2006). There is a further complication withinthe Desmodontinae, one of which (Desmodus rotundus)possesses a Harderian gland whereas the others possesseither a caruncular sebaceous gland (Diaemus youngii) ornothing at all (Diphylla ecaudata, though our specimenwas a fetus, perhaps it had as yet to develop).
Generalizations can be made about the nature of thesecretant produced by the caruncular sebaceous andHarderian glands. The secretant of the caruncular seba-ceous glands usually consists of sebum and is an oilymixture of various lipids and cellular debris. The usualrole of sebum lies in maintaining skin and hair flexibil-ity (Gartner and Hiatt, 2007). Sebaceous glands mayalso play a role in scent marking and grooming (Haffner,2000). However, the secretory nature of the Harderiangland is more difficult to determine. The secretory na-ture of the bat Harderian gland cannot be determinedwithout further histochemical analysis. Numerous func-tions have been ascribed to the mammalian Harderiangland including grooming and production of melatonin(see Payne, 1994 for review).
GENERAL DISCUSSION
The relative development of the orbitofacial glands ofbats appears to show little correlation with phylogeny.However, a correlation is apparent between the presenceof at least one enlarged potentially scent-producinggland (facial and/or Harderian/caruncular sebaceous)
and the level of social organization in the specific batspecies examined (see Table 4). Bats exhibit many differ-ent types of social arrangements, from solitary to short-term aggregations to long-term aggregations leading tothe possible formation of a ‘‘society’’ (Fenton, 2001). BothRhinopoma (Sharifi and Hemmati, 2002) and Rhinolo-phus lepidus possess expanded sweat glands and theyalso exhibit sexual segregation (Nowak, 1998; Sedgeley,2003). In contrast, all vespertilionids, which possessenlarged sebaceous glands with multiple lobules, lobu-lated sweat glands and only moderately developed Mei-bomian glands (Harrison and Davies, 1949; Dalquestand Werner, 1954; Haffner, 1995, 2000; this study) areall highly gregarious and form nursing and or maternal/birthing colonies (Thomas et al., 1979; Nowak, 1998;Whitaker, 1998). The more complicated the form of socialarrangement, the more likely that the animals havedeveloped suitable communication mechanisms.
Bats use many channels for communication, but chem-ical and vocal communications are the main ones. Inaddition, the more complex the behaviour is (e.g., sexattraction is relatively simple whereas recognition sig-nals are more complex) the more complex the chemicalcompound that is produced to facilitate it (Alberts,1992). Scent-producing glands are usually the source ofsuch secretions. Thus, the morphological results need tobe compared to known observations of the chemicalsenses. The two best described are the vomeronasal andthe olfactory senses.
Role of Glands As Cues for theVomeronasal System
It has been proposed (Hillenius and Rehorek, 2005)that primitively, the Harderian gland secretions may
TABLE 4. Association of potential scent-producing gland
Species Scent gland HG SocialityRelative olfactory
bulb size
PteropodidRousettus leschenaulti (fetus) ?HG Loose crowding (Nowak, 1998) 42.8Rhinolophus lepidus FG MR (Nowak, 1998) 28.5Hipposideros lankadiva CG No SS (Sapkal and Bhandarkar, 1984) –Megaderma lyra MG MR (Nowak, 1998) 30.7Rhinopoma microphyllum FG MR (Sharifi and Hemmati, 2002) 37.5Mystacinidae/PhyllostomidaeMystacina tuberculata MG SS (Sedgely, 2003) –Desmodus rotundus FG HG SO (Nowak, 1998) 45.5Diaemus youngii CG, MG SO (Schutt et al, 1999) –Diphylla ecaudata (fetus) – –Brachyphylla cavernarum (fetus) ?HG – –Anoura geoffroyi MG, PI SS (Nowak, 1998) –Macrotus waterhousii MG MR (Nowak, 1998) 40.0Artibeus jamaicensis CG, FG,
MGHG SO (Kunz et al, 1983;
Ortega and Arita, 1999)50.0
VespertilionidaeEptesicus fuscus CG, FG MR þ NC (Nowak, 1998) –Antrozous pallidus FG MR þ NC (Nowak, 1998) 35.3Myotis lucifugus FG MR þ NC (Nowak, 1998) 42.8Miniopterus (all 3 spp.) FG HG MR þ NC (Nowak, 1998) –
Association of potential scent-producing gland (either sweat or sebaceous glands: Caruncular sebaceous gland (CG);enlarged Facial gland ¼ FG; enlarged Meibomian glands ¼ MG, Preorbital indent ¼ PI) Harderian gland (HG) and knownsociality (MR ¼ maternal/birthing roosts; NC ¼ nursing colonies; SO ¼ Social organization; SS ¼ sexual segregation; – ¼unknown) and relative olfactory bulb size (calculated ratio of olfactory bulb diameter to cerebral hemisphere diameter,expressed as a ratio, taken from Bhatnagar and Kallen, 1974).
1444 REHOREK ET AL.

have led into the VNO and served some function in thevomeronsal sense. This suggestion was based upon theconnection between the Harderian gland and the VNOvia the NLD observed in many amphibians and reptiles.However, in mammals, the VNO is rarely closely associ-ated with the NLD (Hillenius and Rehorek, 2005; Rossieand Smith, 2007). This may be due to the reduction orloss of the septomaxillary bone, which was associatedwith the rostral terminus of the NLD in primitive tetra-pods (Hillenius, 2000). In bats, however, there is muchvariation in the presence and relative development ofboth the NLD and the VNO (see Table 3; Gobbel, 2002).This, once again, however, appears to have little clearcongruence with bat phylogeny (see Table 3). The clear-est pattern is observed in the pteropodid group, in whomall the species examined had a shortened NLD. Membersof the Pteropodidae itself lack a VNO altogether (Bhat-nagar et al., 1996) and the VNO is rudimentary in mostof the remaining taxa allied with the pteropodids(Cooper and Bhatnagar, 1976). Thus, although at leastone member of this group, Rousettus leschenaulti,retains a Harderian gland, its orbital secretions cannotplay a role in the VNO function, since the VNO isabsent. This condition parallels that of archosaurs, inwhich a well-developed Harderian gland and NLD, butno VNO, have been described (Rehorek et al., 2005a). Inthat case, it has been suggested that the Harderiangland in these animals may play a role in the mucosalimmune system, as suggested by the large lymphaticaggregations present in the stroma (Rehorek et al.,2005a). The Harderian gland has apparently been lost inother species in the pteropodid group. The loss of theHarderian gland in these bats may be associated withthe reduction of the VNO, but further investigation isneeded to confirm this possibility.
The pattern is less clear among the remaining bats. Inthe Mystacinidae and Phyllostomidae, which appear toshare a common ancestor, the NLD is either lacking orrudimentary. The VNO, on the other hand, is usuallywell developed (it is rudimentary in Brachyphylla). Inthis group, several unrelated species possessed Harder-ian glands (Desmodus rotundus, Artibeus jamaicensisand possibly Brachyphylla cavernosum). Once again,this condition is not unique. In turtles, the Harderiangland has also been disconnected from the VNO throughthe loss of the NLD (Hillenius and Rehorek, 2005). Inmost turtles the Harderian gland is thought to functionin orbital lubrication, but it has been suggested that toplay a role in osmoregulation in several aquatic turtles(Cowan, 1971). It is not known whether the Harderianglands have an osmoregulatory function similar to thoseof turtles. In any case, the Harderian gland, small as itis in these three species of bats, could play a functionalrole in the orbital region.
With the exception of Miniopterus, the vespertilionidbats possess no VNO and have a variably developedNLD. The NLD opens either into the floor of the nostril(Eptesicus fuscus, Antrozous pallidus, Myotis lucifugus)or near the nasopalatine duct/VNO (Miniopterus). In ei-ther case, secretions from the orbital region pass intothe nasal cavity. The condition in Miniopterus is remi-niscent of the primitive tetrapod condition (wherein theNLD open in the vicinity or the VNO, Hillenius andRehorek, 2005), but that of the other three vespertilio-nids is reminiscent of the typical mammalian condition.
This is exemplified by rodents, including the mongoliangerbil, in which the secretions for the orbital region passthrough the NLD and exit at the nares, only to besmeared over the body (Thiessen, 1992). However, unlikerodents, these vespertilionid bats lack a Harderiangland.
Miniopterus deserves special attention. It is the onlybat in this study to possess a Harderian gland, a VNOand a NLD that opens in the vicinity of the VNO, and isreminiscent of the primitive tetrapod condition (Hille-nius and Rehorek, 2005). However, Miniopterinae (Mini-opterus) are currently not thought to be the mostprimitive subfamily of Vespertilionidae. It is currentlyplaced between the Myotinae (Myotis)/ Vespertilioninae(Eptesicus) lineage and the Antrozoinae (Antrozous) line-age (Simmons, 1998). However, all three of these species(Myotis, Eptesicus and Antrozous) lack both a VNO anda Harderian gland (although they possess a full lengthNLD), and thus appear to be more derived than Miniop-terus. These anatomical observations are difficult to rec-oncile with the phylogenetic position of Miniopterus inthe Vespertilionidae and thus needs to be revisitied.With the exception of Miniopterus, it may be concludedthat there is no consistent pattern between the presenceof a Harderian gland and the relative development ofthe VNO and NLD in bats. Instead, there is a lot morevariation in bat HG-NLD-VNO morphology than in mostother described vertebrates.
The condition in Antrozous pallidus is unusual. Thisspecies lacks a Harderian gland, a lacrimal gland, and aVNO, although it retains a full-length nasolacrimal duct.If the primary role of the nasolacrimal duct is to drainorbital fluid into the nose, then the role of this structurein Antrozous pallidus needs to be re-examined. The pres-ence of such a well developed duct in the absence oflarge glands (except a pair of enlarged tarsal glands)raises questions about the presumed drainage role forthis structure and the source of orbital lubricants needsto be determined for this taxon. Lymphatic tissue isspread throughout the conjunctiva.
Thus, in conclusion, the results of this study implythat the relative development of the orbitofacial glandsof bats is not correlated with either that of the NLD orthe VNO. As it has been suggested that the HG-NLD-VNO axis may be the primitive condition (Hillenius andRehorek, 2005), the loss of any of these component maythus be presumed to be indicative of a derived condition.This has been described in turtles (which lack a NLD:Saint Girons, 1988), archosaurs (birds and crocodilians:which lack a VNO: Rehorek et al., 2005a) and severalprimates (which lack a HG and possibly a VNO: Rehorekand Smith, 2006). Bats are unique in that they not onlyare indicative of the derived condition, but also thatthey exhibit more variation in the HG-NLD-VNO axisthan all the other tetrapods combined.
Role of Glands As Cues for theOlfactory System
The size of these potential scent-producing glands andthe level of communality can also be correlated to relativedevelopment of olfactory system (see Table 4). One indi-rect measurement of olfactory acuity may be the ratio ofolfactory bulb diameter to cerebral hemisphere diameter.Although we acknowledge that ratios may be misleading
BAT ORBITOFACIAL GLANDS 1445

(Smith and Bhatnagar, 2004) other means of assessingrelative size (such as calculation of residuals) are imprac-tical due to the small sample size. Based on these ratios,the proportionally largest olfactory bulbs in bats has beenrecorded in Artibeus jamaicensis (Bhatnagar and Kallen,1974a,b). Not only does this species exhibit more anatomi-cal development of both the olfactory and visual struc-tures (Bhatnagar, 1975), but it also possesses lobulatedapocrine sweat glands. The ducts of these glands are asso-ciated with hair follicles, which is typical for glands thatplay a role in scent production (Gartner and Hiatt, 2007).Enlarged apocrine sweat glands have also been describedin lagomorphs (Lyne et al., 1964) and rodents (Rauschand Bridgen, 1989) in which they are presumed to play arole in scent marking (objects and individuals). Thus, thepresence of lobulated apocrine sweat glands is consistentwith the contention that Artibeus jamaicensis uses olfac-tion to a greater degree than most other bats. The olfac-tory system in Artibeus jamaicensis may not onlyfunction in finding food (frugivorous bats appear to havea greater olfactory acuity than nonfrugivorous bats:Bhatnagar and Kallen, 1974a,b), but may also beexplained by their level of social organization. Thesebats form complex social units, namely harems (Kunzet al., 1983; Ortega and Arita, 1999) and individual iden-tification is apparently very important. Such informationmay be passes along chemically using the lobulatedapocrine sweat glands.
The second proportionally largest olfactory acuity hasbeen recorded in Desmodus rotundus (Bhatnagar andKallen, 1974b; Bhatnagar, 2008). This sanguivorous batalso possesses lobulated apocrine sweat glands. Desmo-dus rotundus forms some recognizable social units(Nowak, 1998). In this case, the relatively well developedolfactory system of these bats may be explained by acombination of social structure and dietary require-ments. In Diaemus youngii (also sanguivorous, and aclose relative of Desmodus rotundus) there are addition-ally some recognizable social units, although there is lit-tle evidence of enhanced facial glands. Though therelative development of the olfactory system of Diameusyoungii is unknown, it does not appear to use chemicalsignaling during threat behavior, e.g.: the exposure of apair of large oral glands of the newcomer male and theinsertion of the tongue onto these glands by the domi-nant male (Schutt et al., 1999). Further investigation ofthe olfactory acuity of Desmodontinae is thus needed.
Though not as large as either Artibeus jamaicensis orDesmodus rotundus in size, the insectivourous vesperti-lionid bats still have a relatively well developed olfactorysystem (Bhatnagar and Kallen, 1974a,b; Bhatnagar,1975). Though the relative development of the olfactorysystem in vespertilionids may be correlated to their feed-ing lifestyle (Bhatnagar and Kallen, 1974a,b), it does notseem to explain the level of facial glandular developmentseen in these bats. At this stage, it become necessary toseparate out Miniopterus from the other vespertilionids,as the structure of their facial glands is significantly dif-ferent from that of the others. Vespertilionids, other thanMiniopterus, possess large facial glands, but unlike thatof either Artibeus jamaicensis or Desmodus rotundus,they are sebaceous glands, with relatively underdevel-oped associated sweat glands. Such a combination ofglands was also described in lagomorphs (Mykytowycz,1966). Further chemical analysis of this chemical mixture
led Goodrich and Mykytowycz (1972) to conclude that thesebaceous portion of the secretant may function as a fix-ing medium for the apocrine sweat secretions (which werereported to have a ‘‘rabbity’’ smell). The clustered seba-ceous glands may have a similar function in vespertilionidbats: the copious amount of sebum produced by these clus-tered sebaceous glands may act to keep the secretions ofsweat glands on the body or substrate. Since vespertilio-nids are gregarious and highly social, this mixture of se-baceous and sweat secretions may be smeared uponindividuals. This mixture would then function more as amore permanent identifier (with the sebum functioning asglue for the sudoriparous apocrine secretions) than justsweat secretions alone (which are volatile and prone todissipation when the bat searches for food).
Miniopterus, once again, is an extreme example. Aloneamong the vespertilionids, this genus has a large Har-derian gland as well as a suborbital crevasse lined bylobulated sweat and sebaceous glands. This anatomicalarrangement is reminiscent of that in ungulates. Indeer, it has been suggested that secretions from the Har-derian gland may flow into the preorbital fossa (Rehoreket al., 2005b), which would further enhance the chemicalmixture in the sac. It is this solution that is then usedfor chemical communication. Something similar could behappening in Miniopterus. Whereas all vespertilionidsform maternal/birthing or nursing colonies, Miniopterusforms the largest nurseries (Miniopterus nursery colonycan contain up to 110,000 juveniles, one female per juve-nile, compare to Eptesicus fuscus: 10–14 females, Antro-zous pallidus: 12–100 females and Myotis lucifugus: 15–1,110 females: Nowak, 1998). The young are deposited inthese very large nurseries (under the care of other bats)whilst the mother goes out foraging (Nowak, 1998).Thus, an efficient and specific form of recognition wouldbe vital to locate the baby bat. This may explain whyMiniopterus have such well developed glands.
Enlarged sweat glands were also observed in Rhinol-phus lepidus and lobulated sweat gland in Rhinopomakinneari. The olfactory system is either less (Rhinolo-phus) or equally (Rhinopoma) well developed as in ves-pertilionid bats (Bhatnagar and Kallen, 1974a,b). Therabbit chin glands, also lobulated sweat glands (Lynneet al., 1964), lack any actual smell (Goodrich and Myky-towycz, 1972). Both these bats (Rhinolophus and Rhino-poma) and the rabbit chin gland possess normallydeveloped sebaceous glands. Thus not all sweat glandsmay function in scent marking or social communication.
The relative development of facial glands in adult speci-mens of Rousettus leschenaulti (Pteropodidae) and Bra-chyphylla cavernarum (Phyllostomidae) remains to bedescribed. The fetal specimens of both species possesseither a Harderian gland (Rousettus leschenaulti) or aprimordium that may develop into a Harderian gland(Brachyphylla cavernarum). Neither of these speciesexhibits much in the way of a social organization, thoughRousettus does have marked sexual dimorphism (Nowak,1998). Thus, alternate functions for the Harderian glandneed to be considered. It is unlikely to function in thevomeronasal sense, since the VNO is either absent (Rou-settus) or rudimentary with an incomplete NLD (Brachy-phylla cavernarum). In the case of Rousettus leschenaulti,the presence of a well developed NLD, opening into thenasal cavity, is reminiscent of the archosauran condition(Rehorek et al., 2005a), in which, the presence of
1446 REHOREK ET AL.

lymphatic tissue indicates a role in the immune system. Asimilar function may occur in Rousettus, though furtherinvestigation using adult specimens is required.
The condition in Brachyphylla cavernarum dependsupon what the primordial bud develops into. If it is a Har-derian gland, then it will be the third phyllostomid with aHarderian gland, but the only one without a VNO. Thus,this will mean that there are at least two departures fromthe phyllostomid condition (VNO with no or rudimentaryNLD). On the other hand, if the primordium ends upbeing a caruncular sebaceous gland (as seen in the Des-modontinae), then Brachyphylla will be a more typicalphyllostomid bat with a rudimentary VNO as a stand outfeature. Further examination of an adult Brachyphyllacavernarum would thus be more instructive.
The comparative structure of bat facial and orbitalglands is highly diverse, and few clear patterns, whetherphylogenetic or function, are apparent. What can beunequivocally concluded is that Paule’s (1957) assertionthat bats have no Harderian glands is incorrect. Thefunction of these glands, however, remains largelyunknown, although future examination and correlationwith social behavior may prove instructive. The role ofthe other orbital glands may involve immunological andlubricatory aspects. With respect to the facial glands,the level of development of these glands may be corre-lated to the relative development of the olfactory systemand social structure, though it should be noted that notall glands function in the same manner. Chemical analy-sis of these glands is needed to further elucidate anypotential role that they may have.
ACKNOWLEDGMENTS
The authors thank W. Hillenius for criticism of this ar-ticle. The specimens used in this study were all eitherreceived from colleagues, collected in the wild by KPB,or obtained from museums for a long-term, ongoing,study on the vomeronasal organ of bats. Appropriateacknowledgments have been made to these sources inthe publications, some of which are cited in this work.For Mystacina tuberculata, the authors are grateful toDr. James Dale Smith. The authors acknowledge thegenerous help of Kamar Karim (Rousettus), the late Wil-liam A. Wimsatt (Desmodus, Artibeus), David Nellis(Brachyphylla), Paul Heideman (Anoura geoffroyi), JohnWible, Carnegie Museum of Natural History (Anouracultrata), and Heinz Stephan (Miniopterus magneter).The serially sectioned heads of over fifty species of bats,collected over some 40 years is now housed at SlipperyRock University under the care of Timothy Smith foruse by bat researchers. The preserved specimens (whole,dissections, skeletal preparations) are housed at theDepartment of Biology, University of Louisville, underthe curatorial supervision of Jennifer Jones.
LITERATURE CITED
Aitken ID, Survashe BD. 1977. Plasma cells in vertebrate paraoccu-lar glands. Int Arch Allergy Appl Immun 53:62–67.
Alberts AC. 1992. Constraints on the design of chemical communi-cation systems in terrestrial vertebrates. Am Nat 139:S62–S89.
Bhatnagar KP. 1975. Olfaction in Artibeus jamaicensis and Myotislucifugus in the context of vision and echolocation. Experientia31:856.
Bhatnagar KP. 2008. The brain of the common vampire bat, Desmo-dus rotundus murinus (Wagner, 1840): a cytoarchitectural atlas.Braz J Biol 68:583–599.
Bhatnagar KP, Kallen FC. 1974a. Morphology of the nasal cavitiesand associated structures in Artibeus jamaicensis and Myotis luci-fugus. Am J Anat 139:167–190.
Bhatnagar KP, Kallen FC. 1974b. Cribiform plate of ethmoid, olfac-tory bulb and olfactory acuity in forty species of bats. J Morph142:71–90.
Bhatnagar KP, Wible JR, Karim KB. 1996. Development of the vom-eronasal organ in Rousettus leschenaulti (Megachiroptera, Ptero-podidae). J Anat 188:129–135.
Caspers B, Wibbelt G, Voigt CC. 2009. Histological examinations offacial glands in Saccopteryx bilineata (Chiroptera, Emballonuri-dae), and their potential use in territorial marking. Zoomorphol-ogy 128:37–43.
Castaneda Mde LA, Martınez-Gomez M, Guevara-Guzman R, Hud-son R. 2007. Chemical communication in domestic mammals. VetMex 38:105–123.
Cooper JG, Bhatnagar KP. 1976. Comparative anatomy of the vom-eronasal organ complex in bats. J Anat 122:571–601.
Cowan FBM. 1971. The ultrastructure of the lachrymal ‘‘salt’’glands and the Harderian gland in the euryhaline Malclemys andsome closely related stenohalne emydines. Can J Zool 49:691–697.
Dalquest WW, Werner HJ. 1954. Histological aspects of the faces ofNorth American bats. J Mamm 35:147–160.
Deardon LC. 1959. Meibomian glands in Lagurus. J Mamm 40:20–25.
Fenton MB. 2001. Bats. New York: Checkmark Books.Gartner LP, Hiatt JL. 2007. Color Textbook of Histology, 3rd ed.Philadelphia: Saunders.
Gillette TE, Allansmith MR, Greiner JV, Janusz M. 1980. Histologicand immunohistologic comparison of main and accessory lacrimaltissue. Am J Opthal 89:724–730.
Gobbel L, 2002. Ontogenetic and phylogenetic transformations ofthe lacrimal-conducting apparatus among Microchiroptera. Mam-mal Biol 67:338–357.
Goodrich BS, Mykytowycz R. 1972. Individual and sex differencesin the chemical composition of pheromore-like substances fromthe skin glands of the rabbit, Oryctolagus cuniculus. J Mamm53:540–548.
Green LMA. 1963. Distribution and comparative histology of cuta-neous glands in certain marsupials. Aust J Zool 11:250–272.
Haffner A. 1995. The possibilites of scent marking in the mouse-eared bat Myotis myotis (Borkhausen, 1797) and the noctule batNyctalus noctula (Schreber, 1774) (Mammalia, Chiroptera). ZSaugetierkunde 60:112–118.
Haffner M. 2000. Structure and function of pilosebaceous units onthe heads of small mammals (Insectivora, Chiroptera, Rodentia).Acta Zool 81:195–203.
Harrison Dl, Davies DV. 1949. A note on some epithelial structuresin Microchiroptera. Proc Zool Soc 119:351–357.
Hillenius WJ. 2000. Septomaxilla of nonmammalian synapsids: soft-tissue correlates and a new functional interpretation. J Morph245:29–50.
Hillenius WJ, Watrobski LK, Rehorek SJ. 2001. Passage of tearduct fluids through the nasal cavity of frogs. J Herp 35:701–704.
Hillenius WJ, Rehorek SJ. 2005. From the eye to the nose: ancientorbital to vomeronasal communication in tetrapods? In: MasonRT, Le Master MP, Muller-Schwarze D, editors. Chemical signalsin vertebrates X. New York: Springer. p 228–241.
Knop N, Knop E. 2000. Conjunctiva-associated lymphatic tissue inthe human eye. Invest Opthalmol Vis Sci 41:1270–1279.
Korad VS, Joshi PV. 1992. Anatomical, histological and histochemi-cal studies of the lacrimal gland in Schneider’s leaf nosed batHipposideros speoris (Schneider) (Rhinolophidae, Microchirop-tera). J Anim Morphol Physiol 39:71–83.
Kulzer VE, Helmy I, Necker G. 1985. Untersuchungen uber dieDrusen der gesichtregion der agyptischen Mausschwanz-Fleder-maus Rhinopoma hardwickei cystops Thomas, 1903. Z Saugetier-kunde 50:57–68.
BAT ORBITOFACIAL GLANDS 1447

Kunz TH, August PV, Burnett CD. 1983. Harem Social organizationin cave roosting Artibeus jamaicensis (Chiroptera: Phyllostomi-dae). Biotropica 15:133–138.
Locke R. 2008. The amazing vampire bat. Bats 26:10–11.Lyne AG, Molyneux GS, Mykytowycz R, Parakkal PF. 1964. The de-velopment, structure and function of the submandibular cutane-ous (chin) glands in the rabbit. Aust J Zool 12:340–348.
MacDonald DW, editor. 2006. The encyclopedia of mammals. NewYork: Facts on File Inc.
Madkour GA. 1961. The structure of the facial area in the mouse-tailed bat, Rhinopoma hardwickei cystops, Thomas. Bull Zool SocEgypt 16:50–54.
Mary E, Balakrishnan M. 1991. Specialized cephalic glands of Sun-cus murinus viridescens. Acta Ther 36:187–192.
Miraglia T, Gomes NF. 1969. The Meibomian glands of the marmo-set (Callithrix jacchus). Acta Anat 74:104–113.
Montagna W, Machida H. 1966. The skin of primates XXXII. ThePhilippine tarsier (Tarsius syrichta). Am J Phys Anthrop 25:71–84.
Montgomery RD, Maslin WR. 1992. A comparison of the gland ofHarder response and head-associated lymphoid tissue (HALT)morphology in chickens and turkeys. Avian Dis 36:755–759.
Muller-Schwarze D. 1975. A note on the use of the antorbital glandin marking by Eld’s deer. Appl Anim Ethol 1:301–303.
Mykytowycz R. 1966. Observations on odoriferous and other glandsin the Australian wild rabbit, Oryctolagus cuniculus (L.) and thehare, Lepus europaeus P. CSIRO Wildlife Res 11:49–64.
Nowak RM. 1998. Walker’s mammals of the world, 6th ed. Balti-more: Johns Hopkins University press. Vol. 1.
Ortega J, Arita HT. 1999. Structure and social dynamics of haremgroups of Artibeus jamaicensis (Chiroptera: phyllostomidae).J Mamm 80:1173–1185.
Paule WJ. 1957. The comparative histochemistry of the Harderiangland. Ph.D. dissertation. Columbus, Ohio: Ohio State University.
Payne AP. 1994. The Harderian gland: a tercentennial review.J Anat 185:1–49.
Pinard CL, Weiss ML, Brightman AH, Fenwick BW, Davidson HJ.2003. Evaluation of lysozyme and lactoferrin in lacrimal andother ocular glands of bison and cattle and in tears of bison. AmJ Vet Res 64:104–108.
Quay WB. 1970. Integument and derivatives. In: Wimsatt WA, edi-tor. Biology of bats. New York: Academic press. Vol. 2: p 1–56.
Rausch RL, Bridgens JG. 1989. Structure and function of sudorifer-ous facial glands in nearctic marmots, Marmota spp. (Rodentia:Sciuridae). Zool Anz 223S:265–282.
Rehorek SJ, Baker JJ, Hutchinson MN, Firth BT. 2006. The Har-derian gland of two species of skink (Tiliqua rugosa and Hemier-gis decersiensis): a discussion of the significance of lymphatictissue in the squamate Harderian gland. Can J Zool 84:706–714.
Rehorek SJ, Hillenius WJ, Kennaugh J, Chapman N. 2005b. Thedeer, the gland and the fossa: on the trail of scent. In: Mason RT,Le Master MP, Muller-Schwarze D, editors. Chemical signals invertebrates X. New York: Springer. p 152–158.
Rehorek SJ, Hillenius WJ, Quan W, Halpern M. 2000. Passage ofthe Harderian gland secretions to the vomeronasal organ of thesnake, Thamnophis sirtalis. Can J Zool 78:1284–1288.
Rehorek SJ, Legenzoff, EJ, Carmody K, Smith TD, Sedlmayr JC.2005a. Alligator tears: a re-evaluation of the lacrimal apparatusof the crocodilians. J Morphol 266:298–308.
Rehorek SJ, Smith TD. 2006. The primate Harderian gland: does itreally exist? Ann Anat 188:319–327.
Rossie JB, Smith TD. 2007. Ontogeny of the nasolacrimal duct inprimates: functional and phylogenetic implications. J Anat210:195–208.
Sakai T. 1981. The mammalian Harderian gland: morphology, bio-chemistry, function and phylogeny. Arch Hist Jpn 44:299–333.
Saint Girons H. 1988. Les glandes cephaliques exocrines des Rep-tiles 1. Donnees anatomiques et histologiques. Ann Sci Nat ZoolParis 13e Serie 9:221–255.
Sapkal VM, Bhandarkar WR. 1984. Breeding habits and associatedphenomena in some Indian bats. Part IX Hipposideros lankadiva(Kelaart)—Hipposideridae. J Bombay Nat Hist Soc 81:380–386.
Schaffer J. 1940. Die Hautdrusenorgane der Saugetiere. Berlin,Germany: Urban und Schwarzenberg.
Schutt WA, Muradali F Jr, Mondol N, Joseph K, Brockmann K.1999. Behavior and maintenance of captive white-winged vampirebats, Diaemus youngi. J Mamm 80:71–81.
Sedgeley JA. 2003. Roost site selection and roosting behaviour inlesser short-tailed bats (Mystacina tuberculata) in comparisonwith long-tailed bats (Chalinolobus tuberculatus) in Nothofagusforest, Fiordland. N Z J Zool 30:227–241.
Sharifi M, Hemmati Z. 2002. Variation in the diet of the greatermouse-tailed bat, Rhinipoma microphyllum (Chiroptera; Rhinopo-matidae) in South-Western Iran. Zool Middle East 26:65–70.
Simmons NB. 1998 A re-appraisal of interfamily relationships ofbats. In: Kunz TH, Racey PA, editors. Bat biology and conserva-tion. Washington: Smithsonian Institution Press. p 3–26.
Simmons NB. 2005. Order Chiroptera. In: Wilson DE, Reeder DM,editors. Mammal species of the world: a taxonomic and geo-graphic reference, 3rd ed. Baltimore: The Johns Hopkins Univer-sity Press. p 312–529.
Smith TD, Bhatnagar KP. 2004. Microsmatic primates: reconsider-ing how and when size matters. Anat Rec 279B:24–31.
Stephens LC, Schultheiss TE, Vargas KJ, Cromeens DM, Gray KN,Ang KK. 1989. Glands of the eyelids of rhesus monkeys (Macacamulatta). J Med Primatol 18:383–396.
Theissen D. 1992. The function of the Harderian gland in the Mon-golian gerbil, Meriones unguiculatus. In: Webb SM, Hoffman RA,Puig-Domingo ML, Reiter RJ, editors. Harderian glands: porphy-rin metabolism, behavioral and endocrine effects. Berlin:Springer. p 127–140.
Thomas DW, Fenton MB, Barclay RMR. 1979. Social behavior ofthe little brown bat, Myotis lucifugus. Behav Ecol Sociobiol 6:129–136.
Wake MH. 1985. The comparative morphology and evolution of theeye of caecilians (Amphibia: Gymnophiona). Zoomorphology 105:227–295.
Whitaker JO Jr. 1998. Life history and roost switching in sixsummer colonies of eastern pippistrelles in buildings. J Mammol79:651–659.
Wible JR, Bhatnagar, KP. 1996. Chiropteran vomeronasal complexand the interfamilial relationships of bats. J Mamm Evol 3:285–314.
1448 REHOREK ET AL.




















