
Oral glands in dipsadine “goo-eater” snakes: Morphology and histochemistry of the infralabial...
17
This article was published in an Elsevier journal. The attached copy is furnished to the author for non-commercial research and education use, including for instruction at the author’s institution, sharing with colleagues and providing to institution administration. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Oral glands in dipsadine “goo-eater” snakes: Morphology and histochemistry of the infralabial...

This article was published in an Elsevier journal. The attached copyis furnished to the author for non-commercial research and
education use, including for instruction at the author’s institution,sharing with colleagues and providing to institution administration.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Toxicon 51 (2008) 898–913
Oral glands in dipsadine ‘‘goo-eater’’ snakes: Morphology andhistochemistry of the infralabial glands in Atractus reticulatus,
Dipsas indica, and Sibynomorphus mikanii$
Leonardo de Oliveiraa, Carlos Jareda, Ana Lucia da Costa Prudenteb,Hussam Zaherc, Marta Maria Antoniazzia,�
aLaboratorio de Biologia Celular, Instituto Butantan, Av. Vital Brazil 1500, CEP 05503-900, Sao Paulo, SP, BrazilbMuseu Paraense Emılio Goeldi, Departamento de Zoologia, Herpetologia, Belem, PA, Brazil
cMuseu de Zoologia da Universidade de Sao Paulo, Sao Paulo, SP, Brazil
Received 21 August 2007; received in revised form 17 December 2007; accepted 18 December 2007
Available online 28 December 2007
Abstract
Although snake infralabial glands are generally constituted of mucous cells, among dipsadines, they are much more
developed and predominantly serous in nature, possibly due to the peculiar feeding habits of some species of this group,
the ‘‘goo-eaters’’, which feed on soft and viscous invertebrates. We compared the morphology and histochemistry of the
infralabial glands of three goo-eater species of Southeast Brazil, Atractus reticulatus, Dipsas indica and Sibynomorphus
mikanii. In A. reticulatus the glands are formed by mixed acini composed of mucous and seromucous cells and in D. indica,
they are composed of mucous tubules and seromucous acini. In S. mikanii the glands are organized in seromucous acini;
mucous cells are restricted to the gland anterior region and to the duct lining epithelium. Ultrastructurally, secretory
granule electron density varies from low to moderate, depending on their mucous or seromucous nature. The results
indicate a large morphological and histochemical variation in the infralabial glands, probably reflecting differences in the
secretion chemical composition and in feeding specialization among the three species. The protein content in the secretory
cells can be related with the presence of toxins that can be used in chemical prey immobilization or detaching of snails from
their shells.
r 2007 Elsevier Ltd. All rights reserved.
Keywords: Serpentes; Colubridae; Dipsadinae; Goo-eaters; Infralabial glands; Morphology
ARTICLE IN PRESS
www.elsevier.com/locate/toxicon
0041-0101/$ - see front matter r 2007 Elsevier Ltd. All rights reserved.
doi:10.1016/j.toxicon.2007.12.021
$Ethical statement: The authors warrant that the above manuscript is an original work, it has not been published before and it is not
submitted for publication anywhere. It contains no libelous or other unlawful statements, and it does not infringe on the rights of others.
The authors have no relationship with any manufactures or distributors of products used in this manuscript. This paper reflects our own
research and analysis and does so in a truthful and complete manner. All authors have contributed significantly to the execution, analysis
and writing of the study reported and all co-authors have agreed to submit the manuscript to Toxicon. Besides, the manuscript is
appropriately placed in the context of prior and existing research.�Corresponding author. Tel.: +55 11 3726 7222x2234; fax: +55 11 3726 1505.
E-mail address: [email protected] (M.M. Antoniazzi).

Author's personal copy
1. Introduction
In snakes, the infralabial and supralabial glands,the venom glands and Duvernoy’s or parotid glands(Taub, 1966) are among the best-known oral glands.Venom glands and Duvernoy’s glands, which arefound only within the derived superfamily Colubroi-dea, have been studied thoroughly due to their toxicproperties. Although being considered homologues(Kardong, 2002), these glands show many morpho-logical and functional differences among them. Whiletrue venom glands are found in members of thefamilies Elapidae, Viperidae and Atractaspididae,Duvernoy glands are exclusive (although not foundin all members) of the members of the family‘‘Colubridae’’, a probably paraphyletic assemblageof colubroid snakes that remains poorly resolvedphylogenetically (Underwood, 1967; Dowling andDuellman, 1978; McDowell, 1987; Kraus andBrown, 1998; Zaher, 1999; Kelly et al., 2003; Lawsonet al., 2005). Both types are associated to themaxillary teeth and are generally formed by serouscells (Kochva, 1987; Underwood, 1997; Vidal, 2002;Jackson, 2003). Differently from the venom andDuvernoy’s glands that are restricted to Colubroids,infralabial glands seem to occur in practically allsnakes that have been already studied (Haas, 1964;Taub, 1966; Underwood, 2002; Kley, 2006).
The most usual classification for reptile oralglands is based on the distinct types of secretorygranules they present, depending on their histo-chemical aspects, which reveal differences in theirchemical composition (Gabe and Saint-Girons,1969; Kochva, 1978; Underwood, 1997). Whileserous or seromucous cells, rich in protein material,are usually associated with the secretion of toxinsand are then expected to be present in venomglands, mucous cells are associated with mucoussecretion and lubrication (Underwood, 1997).
Although far less studied histologically, infra-labial glands have been traditionally regarded asbeing very similar to supralabial glands in produ-cing mucous secretion that plays an important rolein prey lubrication during ingestion (Kochva, 1978).However, distinctly from the pattern found in mostColubroids, a group of dipsadine snakes informallycalled ‘‘goo-eaters’’ by Cadle and Greene (1993)show distinctly developed and specialized infralabialglands that produce a secretion predominantlyserous in chemical nature, a fact that is probablyrelated to their peculiar feeding behaviour (Taub,1966; Gans, 1972; Zaher, 1996).
Dipsadines constitute a group of Colubrid snakeswith approximately 250 species, all restricted to theNew World, and with the larger diversity observedin Central America. The subgroup of goo-eaters feed only on soft and viscous invertebrates(generally molluscs as slugs and snails, and anne-lids) and include the genera Geophis, Atractus,Adelphicos, Ninia, Sibynomorphus, Sibon and Dipsas.Monophyly of the goo-eaters is still controversial,but it is usually accepted that they form two clades:Geophis, Atractus, Adelphicos and Ninia on the onehand, and Sibynomorphus, Sibon and Dipsas on theother hand (Ferrarezzi, 1994; Fernandes, 1995;Campbell and Smith, 1998; Zaher, 1999).
In southern Brazil, the goo-eaters are representedby three genera: Atractus, Dipsas and Sibynomorphus.The genus Atractus is characterized by the fossorialhabits, small size, short tail and head adapted toexcavation. Despite our poor knowledge on thebiology and natural history of the group, it isknown that Atractus feeds preferentially on earth-worms (Marques et al., 2001) whereas the generaDipsas and Sibynomorphus, which are associatedto both terrestrial and arboreal environments(Marques et al., 2001), feed exclusively on slugsand snails. These two genera present a number ofspecific adaptations enabling them to extractmolluscs from the shell without breaking it (Sazima,1989; Franco, 1994; Oliveira, 2001). According toSazima (1989), snakes from genus Dipsas feed byintroducing around one third of the mandible insidethe mollusc shell and making synchronic andindependent movements with the two mandiblehalves, while the maxilla externally holds the shell.In addition, the presence of serous cells in theinfralabial glands of Dipsas, as described by Taub(1966), together with the fact that the whole feedingprocess is very fast, taking only about 2minutes,suggests that the secretion of these glands can beinvolved in the chemical detaching of the molluscfrom the shell. Laporta-Ferreira (1985) and Lapor-ta-Ferreira and Salomao (1991), working withSibynomorphus neuwiedi, observed that extracts ofthe infralabial glands cause paralysis and death onslugs. In addition, complementary studies showedthat in Sibynomorphus mikanii the large amounts ofprotein found in the infralabial glands have aproteolytic effect on molluscs (Salomao andSawaya, 1992; Salomao and Laporta-Ferreira1994).
In snakes, several morphological and behaviouralspecializations are associated to the feeding habits
ARTICLE IN PRESSL. de Oliveira et al. / Toxicon 51 (2008) 898–913 899

Author's personal copy
(Pough, 1983; Pough and Groves, 1983; Voris andVoris, 1983). According to Savitzky (1983), thehabit of eating molluscs by extracting them fromtheir shells is restricted to a small number of speciesand may constitute one of the most radical changesin feeding strategies related to prey dealing amongsnakes. The ability of taking molluscs out of theirshells without breaking it can be observed in twoindependent lineages, represented by the Neotropi-cal dipsadines of genera Dipsas, Sibon and Sibyno-
morphus and by the Asian pareatines (subfamilyPareatinae) of genera Aplopeltura and Pareas
(Savitzky, 1983). In species of these genera manydifferent cranial morphological modifications re-lated to the feeding habits were observed, such asdiminishing of the number of palatomaxilar boneteeth, raise in the number of the dentary bone teeth,supratemporal bone reduction and quadrate boneenlargement (Brongersma, 1956; Gans, 1972).
The specialized diet of the dipsadine goo-eaters,besides reflecting a series of morphological andbehavioural specializations (Dunn, 1951; Sazima,1989; Zaher, 1996), has also been associated to thecomposition of the secretion of their infralabialglands (Laporta-Ferreira et al., 1988; Salomao andLaporta-Ferreira, 1994).
The present paper is part of a wider morpholo-gical and histochemical study, which aims toinvestigate the oral glands and associated structuresof dipsadines, correlating their gland structure withsecretion biochemistry and with the natural historyand biology of the group. The present work focuseson the morphology and histochemistry and on thearrangement of the different types of cells within thesecretory epithelium of the infralabial glands ofthree representative dipsadine goo-eater speciesbelonging to the genera Sibynomorphus, Atractus
and Dipsas. Results were correlated to the feedinghabits of the species in an attempt to bring up datathat can be useful in future toxinological andphylogenetical studies on dipsadines.
2. Materials and methods
2.1. Animals
Adult individuals of Atractus reticulatus (n ¼ 3,Fig. 1A), Dipsas indica (n ¼ 3, Fig. 1B) andS. mikanii (n ¼ 4, Fig. 1C) from the State of SaoPaulo were used in this work. Specimens werefurnished by the Laboratory of Herpetology of theInstituto Butantan (IB).
The specimens were sacrificed with an intraper-itoneal overdose of sodic thiopental (30mg/kg). Theinfralabial glands were then dissected and processedfor morphological studies. The specimens werepreserved in formalin and deposited in the herpeto-logical collection of the IB and in the collection of
ARTICLE IN PRESS
Fig. 1. (A) Male of Atractus reticulatus; (B) female of Dipsas
indica; (C) male of Sibynomorphus mikanii.
L. de Oliveira et al. / Toxicon 51 (2008) 898–913900

Author's personal copy
the IB specific for the municipality of Sao Paulo(MSP) (A. reticulatus IB 72423, MSP 741, MSP 896;D. indica IB 72899, IB 73325, IB 73448; S. mikanii
IB 73326, MSP 637, MSP 935, MSP 936).
2.2. Histology
The glands were fixed in 4% paraformaldehyde inPBS 0.1M, pH 7.2 for 24 h, dehydrated in ethanoland embedded in historesin (glycol methacrylate,Leica).
Sections of 2 mm were obtained in a Microm HM340 microtome using glass knifes. For general studyof the tissues, the sections were stained withtoluidine blue-fuchsin (Junqueira, 1995).
2.3. Histochemistry
The sections were subjected to the followinghistochemical staining procedures (according toBancroft and Stevens, 1996): periodic acid-Schiff(PAS), alcian blue pH 2.5 and combined PAS andalcian blue pH 2.5 (Pearse, 1985; Kiernan, 2001), foridentification of neutral (PAS) and acid (alcian blue)mucosubstances, and bromophenol blue, for identi-fication of proteins.
2.4. Photomicrography
Photomicrographs were obtained with an OlympusBX51 microscope and in an Olympus SZ stereo-microscope equipped with a digital camera and withthe software Image-Pro Express (MediaCybernetics).
2.5. Transmission electron microscopy
Fragments of the glands with approximately1mm3 were fixed in Karnovsky solution pH 7.2(Karnovsky, 1965) for 24 h, post-fixed in 1%osmium tetroxide, contrasted in 2% uranyl acetate,dehydrated in ethanol and embedded in epoxy resin.Ultrathin sections (60 mm) were obtained in aSorvall MT 6000 ultramicrotome, contrasted in2% uranyl acetate and lead citrate, and examinedwith a LEO 906E, operating at 80 kV.
3. Results
3.1. General morphology
In all three species the infralabial glands arewhitish, thin and elongated, running laterally to the
mandible, from the anterior tip of the dentary tothe anterior portion of the compound bones(Figs. 2A–C). Each one, however, has particularcharacteristics that are described below. They all arebasically composed of a secretory portion and by anet of internal ducts. The whole gland is wrapped bya layer of connective tissue which penetrates into theglandular body in the form of many septa, dividingthe glandular body into lobules and involving theacini (or tubuli) and the ducts (Figs. 3A, 4Aand 5A). The acini are formed by a monolayered
ARTICLE IN PRESS
Fig. 2. (A–C) Schematic drawing of the skinned heads of
Atractus reticulatus (A), Dipsas indica (B) and Sibynomorphus
mikanii (C), showing the location of the infralabial gland (il) with
respect to the head muscles and mandible. Note the prominent
muscle levator anguli oris (lao) in D. indica. aep, muscle adductor
externus profundus (sensu Zaher, 1994); aes, muscle adductor
externus superficialis; n, nasal gland; pm, premaxillary gland; sl,
supralabial gland.
L. de Oliveira et al. / Toxicon 51 (2008) 898–913 901

Author's personal copy
epithelium constituted of mucous and seromucouscells, recognized by their histochemical character-istics (described in detail in the following section).The basic difference among the glands of the threespecies, besides their size in relation to the head, isthe relative number and disposition of mucous andseromucous cells.
In A. reticulatus, the infralabial gland is wide andshort, running along the internal lip margin, justunder the infralabial scales. It measures around6mm long (representing 40% of the total length ofthe head) and 2mm wide (Fig. 2A), and shows twoconstrictions, one in the anterior region and anotherin the posterior region. Mucous cells predominateover seromucous cells in the acini, and arecharacterized by flat basal nuclei and a cytoplasmfull of secretion granules. Two different types ofmucous cells are recognized, based on the tintorialproperties of their granules: one with light granulesand another with medium-stained granules(Fig. 3B). Although always present in the acini,the seromucous cells seem to occur in largernumbers in the periphery of the gland (Fig. 3A),being recognized by the presence of rounded basalnuclei and dense cytoplasm, full of small well-stained secretion granules (Fig. 3B). The ducts arelined by tall mucous cells, with flat basal nuclei andcytoplasm full of poorly stained secretion granules(Fig. 3A).
In D. indica, the infralabial gland is thin andelongated, is located ventrolaterally to the dentaryand compound bones, and is almost entirelycovered by the muscle levator anguli oris, which isespecially well-developed in this species. The glandmeasures about 16mm long and 1.7mm wide inadults, occupying approximately 70% of the totalhead length (Fig. 2B). It is divided into two distinctregions: a posterior and peripheral region consti-tuted exclusively by seromucous cells organized inacini, and an anterior and central region, consti-tuted of mucous cells organized in structuresresembling tubuli with wider lumina. Transitionzones are present between these two regions, wherethe two cellular types (mucous and seromucous) areobserved in the same secretory unit (Fig. 4A). Themucous cells are columnar, with flat basal nucleiand with a cytoplasm full of small secretion granules(Figs. 4A and B). The seromucous cells arepolygonal, with round basal nuclei. Their secretiongranules are larger and easily differentiated from themucous granules by their distinct affinities to thetoluidine blue-fuchsin (Figs. 4A and B). A third
cellular type, not very frequent, was observed, bothin the mucous tubuli and in the seromucous acini(mainly close to the transition zones), and wasidentified as another type of seromucous cell (sm2)that contrasts with the more abundant seromucouscell of type 1 (sm1). The seromucous cell sm2 issmaller than sm1, and shows more prominentsecretory granules containing heterogeneous mate-rial, which is more intensely stained by the toluidineblue-fuchsin when compared to sm1 (Fig. 4B).
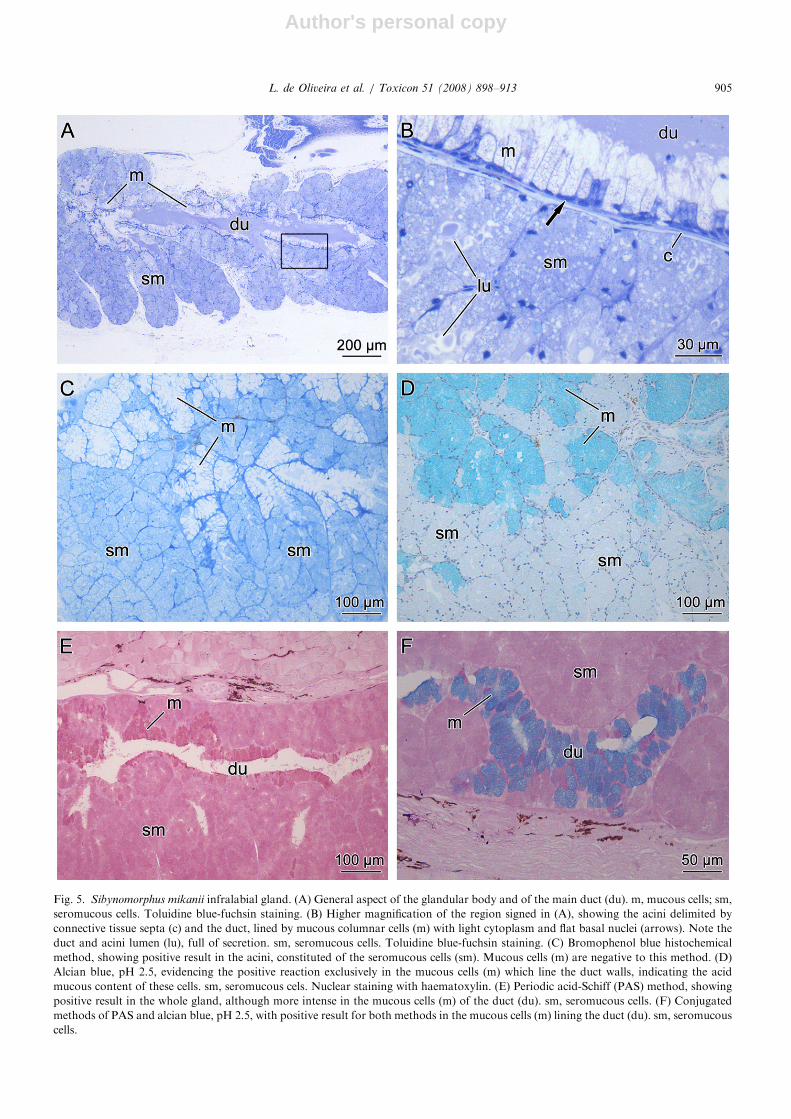
In S. mikanii, the infralabial gland is thin andelongated, running juxtaposed to the external faceof the dentary and compound bone. It measuresaround 7mm long and 1.5mm wide, occupying70% of the total length of the head (Fig. 2C). Theglandular body is composed of acini with muchreduced lumina, full of secretion, and formed bypolygonal seromucous cells with rounded basalnuclei and cytoplasm full of secretion granules(Figs. 5A and B). Some of these granules, whenstained by toluidine blue-fuchsin, show a well-stained core surrounded by a light region(Fig. 5B). The ducts are formed by mucous cellswith flat basal nuclei and cytoplasm full of granuleslightly stained by toluidine blue-fuchsin (Fig. 5B).
3.2. Histochemistry
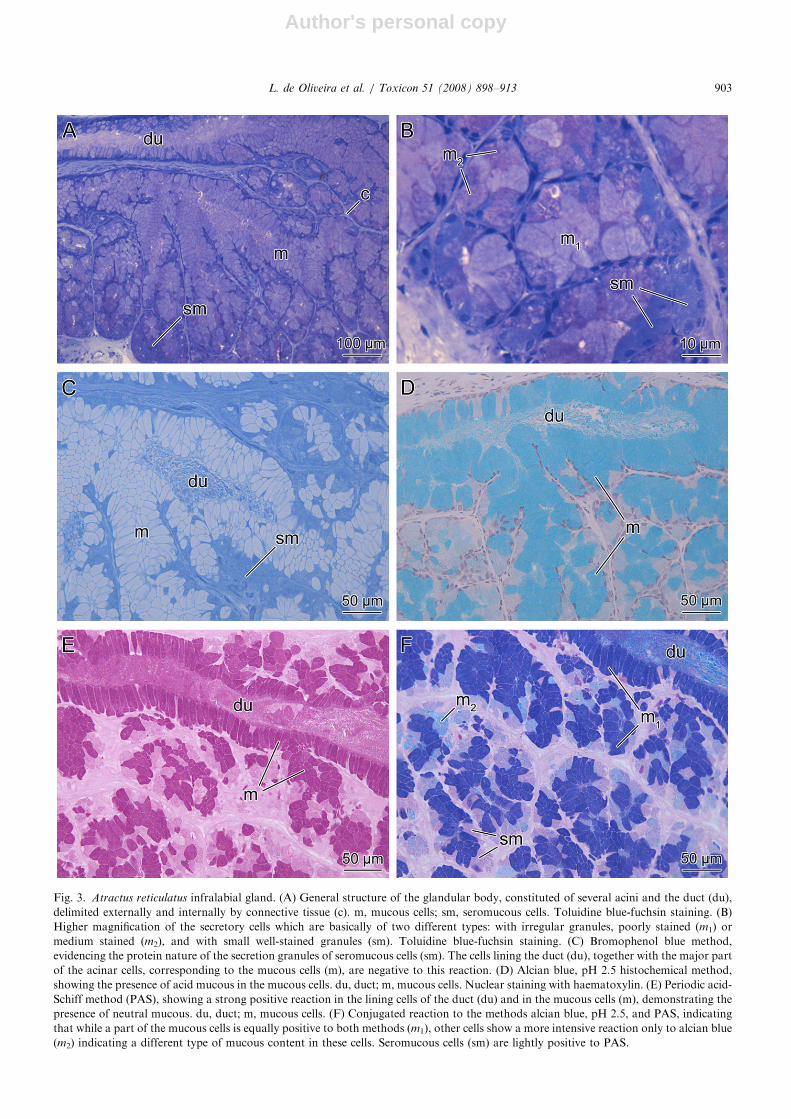
In A. reticulatus, most cells react positively toboth alcian blue (pH 2.5) and PAS, and negativelyto bromophenol blue (Figs. 3C–F), characterizingtheir mucous condition. Among these cells, twodistinct types are described as m1 and m2, dependingon their intensity of reaction to PAS or alcian blue(pH 2.5) well evidenced by the simultaneousapplication of both methods (Fig. 3F). The firsttype (m1), which is strongly reactive to both PASand alcian blue (pH 2.5) is the only one presentin the ducts and composes most cells of the acini(Figs. 3D–F). The second type (m2), more reactiveto alcian blue (pH 2.5), is present only in the acini(Fig. 3F). The seromucous cells, characterized bythe positive result to bromophenol blue, and a weakreaction to PAS, are present in smaller numbers inthe acini and seem to be more abundant in theperiphery of the gland (Figs. 3A, C and F).
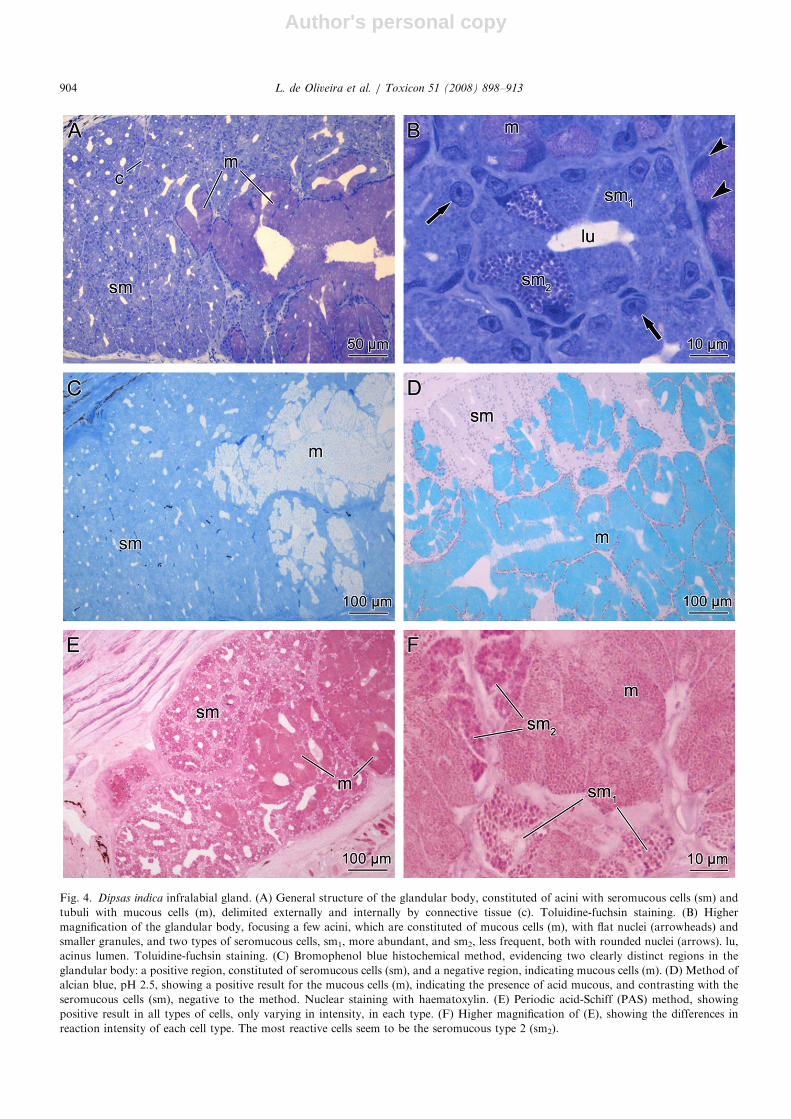
In D. indica the cells forming the acini are positiveto bromophenol blue and PAS (Figs. 4C and E,respectively) and negative to alcian blue, pH 2.5(Fig. 4D), indicating a seromucous condition.A second type of cell, with the same histochemi-cal characteristics but presenting a more intense
ARTICLE IN PRESSL. de Oliveira et al. / Toxicon 51 (2008) 898–913902
Author's personal copy
ARTICLE IN PRESS
Fig. 3. Atractus reticulatus infralabial gland. (A) General structure of the glandular body, constituted of several acini and the duct (du),
delimited externally and internally by connective tissue (c). m, mucous cells; sm, seromucous cells. Toluidine blue-fuchsin staining. (B)
Higher magnification of the secretory cells which are basically of two different types: with irregular granules, poorly stained (m1) or
medium stained (m2), and with small well-stained granules (sm). Toluidine blue-fuchsin staining. (C) Bromophenol blue method,
evidencing the protein nature of the secretion granules of seromucous cells (sm). The cells lining the duct (du), together with the major part
of the acinar cells, corresponding to the mucous cells (m), are negative to this reaction. (D) Alcian blue, pH 2.5 histochemical method,
showing the presence of acid mucous in the mucous cells. du, duct; m, mucous cells. Nuclear staining with haematoxylin. (E) Periodic acid-
Schiff method (PAS), showing a strong positive reaction in the lining cells of the duct (du) and in the mucous cells (m), demonstrating the
presence of neutral mucous. du, duct; m, mucous cells. (F) Conjugated reaction to the methods alcian blue, pH 2.5, and PAS, indicating
that while a part of the mucous cells is equally positive to both methods (m1), other cells show a more intensive reaction only to alcian blue
(m2) indicating a different type of mucous content in these cells. Seromucous cells (sm) are lightly positive to PAS.
L. de Oliveira et al. / Toxicon 51 (2008) 898–913 903
Author's personal copy
ARTICLE IN PRESS
Fig. 4. Dipsas indica infralabial gland. (A) General structure of the glandular body, constituted of acini with seromucous cells (sm) and
tubuli with mucous cells (m), delimited externally and internally by connective tissue (c). Toluidine-fuchsin staining. (B) Higher
magnification of the glandular body, focusing a few acini, which are constituted of mucous cells (m), with flat nuclei (arrowheads) and
smaller granules, and two types of seromucous cells, sm1, more abundant, and sm2, less frequent, both with rounded nuclei (arrows). lu,
acinus lumen. Toluidine-fuchsin staining. (C) Bromophenol blue histochemical method, evidencing two clearly distinct regions in the
glandular body: a positive region, constituted of seromucous cells (sm), and a negative region, indicating mucous cells (m). (D) Method of
alcian blue, pH 2.5, showing a positive result for the mucous cells (m), indicating the presence of acid mucous, and contrasting with the
seromucous cells (sm), negative to the method. Nuclear staining with haematoxylin. (E) Periodic acid-Schiff (PAS) method, showing
positive result in all types of cells, only varying in intensity, in each type. (F) Higher magnification of (E), showing the differences in
reaction intensity of each cell type. The most reactive cells seem to be the seromucous type 2 (sm2).
L. de Oliveira et al. / Toxicon 51 (2008) 898–913904
Author's personal copy
ARTICLE IN PRESS
Fig. 5. Sibynomorphus mikanii infralabial gland. (A) General aspect of the glandular body and of the main duct (du). m, mucous cells; sm,
seromucous cells. Toluidine blue-fuchsin staining. (B) Higher magnification of the region signed in (A), showing the acini delimited by
connective tissue septa (c) and the duct, lined by mucous columnar cells (m) with light cytoplasm and flat basal nuclei (arrows). Note the
duct and acini lumen (lu), full of secretion. sm, seromucous cells. Toluidine blue-fuchsin staining. (C) Bromophenol blue histochemical
method, showing positive result in the acini, constituted of the seromucous cells (sm). Mucous cells (m) are negative to this method. (D)
Alcian blue, pH 2.5, evidencing the positive reaction exclusively in the mucous cells (m) which line the duct walls, indicating the acid
mucous content of these cells. sm, seromucous cels. Nuclear staining with haematoxylin. (E) Periodic acid-Schiff (PAS) method, showing
positive result in the whole gland, although more intense in the mucous cells (m) of the duct (du). sm, seromucous cells. (F) Conjugated
methods of PAS and alcian blue, pH 2.5, with positive result for both methods in the mucous cells (m) lining the duct (du). sm, seromucous
cells.
L. de Oliveira et al. / Toxicon 51 (2008) 898–913 905

Author's personal copy
reaction to PAS (Fig. 4F), was observed both in theacini and in the tubuli, and was identified as thesecond type of seromucous cell (sm2). The columnarcells constituting the tubuli were negative tobromophenol blue (Fig. 4C) and strongly positiveto alcian blue, pH 2.5, (Fig. 4D) and PAS (Fig. 4E),indicating their mucous nature.
In S. mikanii the cells constituting the acini werepositive to bromophenol blue and to PAS (Figs. 5Cand E), and negative to alcian blue pH 2.5 (Fig. 5D)demonstrating the seromucous nature of theirsecretion. The columnar cells of the ducts werepositive to alcian blue, pH 2.5, intensely positive toPAS (Figs. 5D–F) and negative to bromophenolblue, indicating their mucous chemical nature(Fig. 5C).
3.3. Ultrastructure
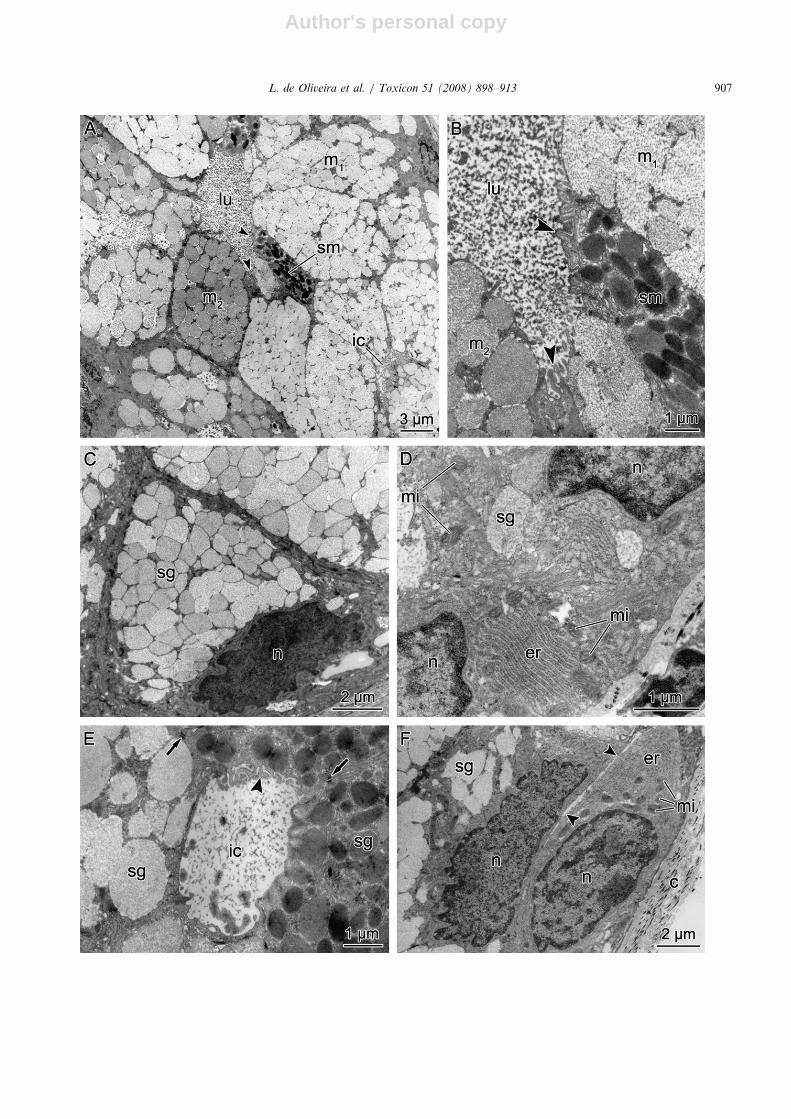
In A. reticulatus the acini are mainly composed ofpolygonal cells, with basal nuclei and cytoplasmpacked with low-electron-dense granules. Althoughbeing similar in aspect, these granules can showdifferences from one cell to another in their levelof electron density and packing in the cytoplasm(Fig. 6A). A few columnar cells, less voluminouswhen compared to the mucous cells (Figs. 6A–C),are present in the acini. These cells show well-individualized spherical or elliptical granules, whichare much more electron dense and heterogeneouswhen compared to the polygonal cells (Figs. 6B andC). The acinar lumen is usually full of granulousmaterial secreted by the cells from the apical regionfull of microvilli (Figs. 6A and B). In the perinuclearregion of both cell types a well-developed roughendoplasmic reticulum is observed, as well asmitochondria (Fig. 6D). Intercellular secretorycanaliculi (ISC) are very common among the acinarcells of both types (Figs. 6A and E). Myo-epithelialcells were observed in the acinar basis, between thebasal lamina and the secretory cells. These cells areelongated but irregular in shape, and the cytoplasm
is rich in mitochondria and rough endoplasmicreticulum (Fig. 6F).
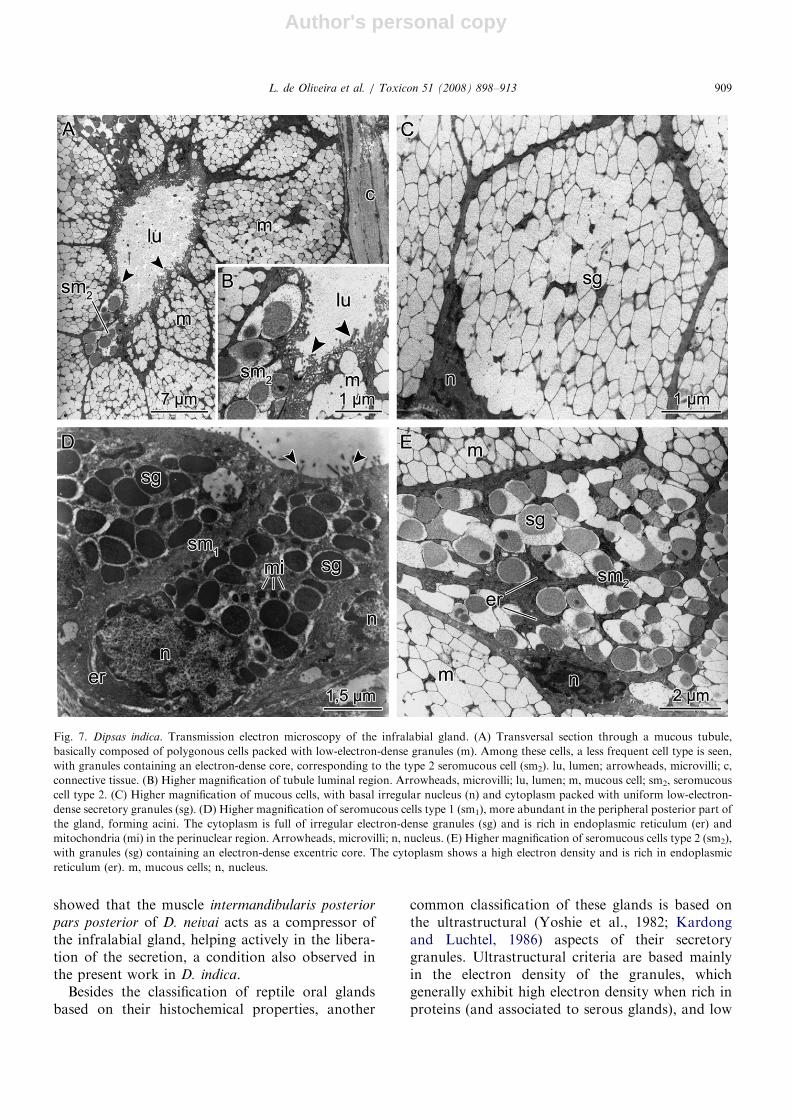
In D. indica the acini are mostly constituted ofpolygonal cells full of elliptical and electron-densesecretory granules (Fig. 7D). The cytoplasm of theperinuclear region of these cells is rich in endoplas-mic reticulum (Fig. 7D). The anterior and centraltubular region of the gland is constituted ofpolygonal cells packed with low-electron-densegranules (Figs. 7A and C). Another type of cell,less frequent and less voluminous, was identifiedboth in the acini and in the tubuli. These cells aresmaller, with the cytoplasm full of granules,heterogeneous in size, composed of a dense excen-trical core surrounded by a low-density halo(Figs. 7A, B and E). The apical region of the cellsshows a large number of microvilli facing the lumenin which the secretory granules are secreted byexocytosis (Figs. 7A, B and D).
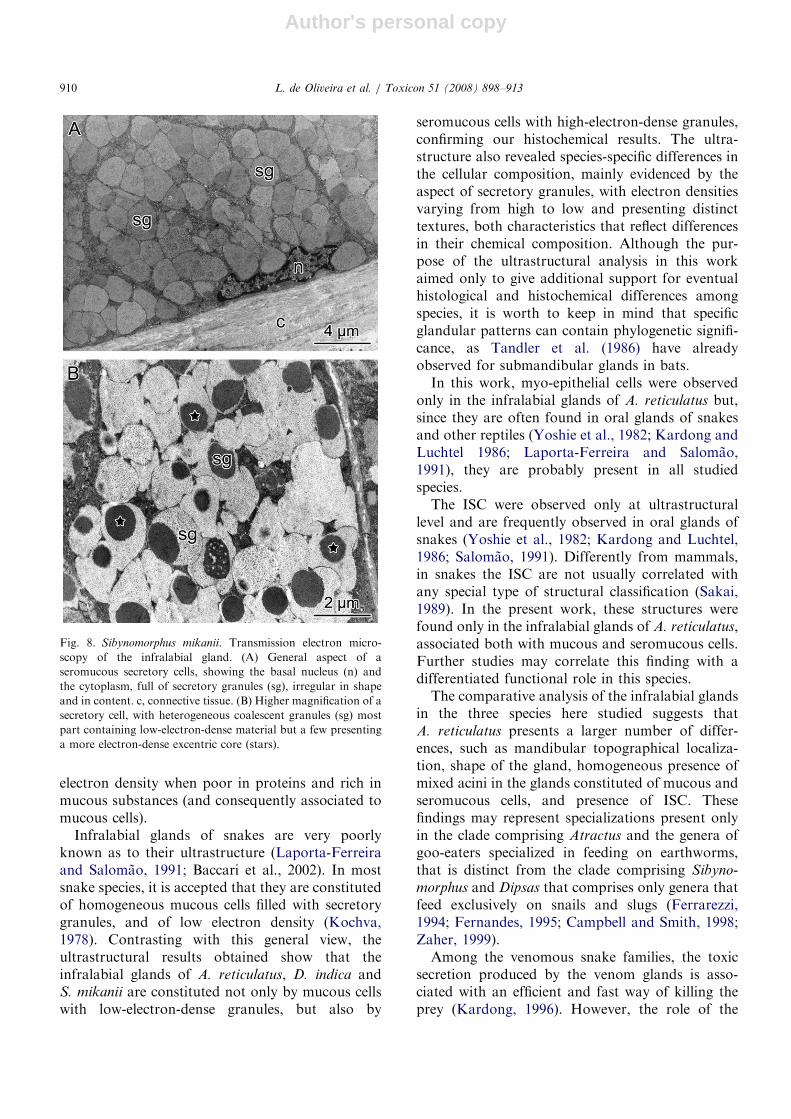
In S. mikanii, the cells composing the acini arepolygonal, with flat and irregular basal nuclei. Thecytoplasm is packed with coalescent secretorygranules, rounded or polygonal in shape. Thegranule content is of low electron density and quitehomogeneous (Fig. 8A), although some granulescan show an excentrical core of higher electrondensity (Fig. 8B).
4. Discussion
Despite the infralabial glands being so commonamong snakes, information about the morphologyand function of these glands is very scarce. Mostpapers are restricted to simple morphologicaldescriptions, without any functional and phylogene-tical association (Zago, 1971; Baccari et al., 2002).Underwood (2002), studying the morphology of theoral region of some snake species, found a numberof structural variations, including in the supralabialand infralabial glands, concluding that detailedinvestigations about these structures are in needand will potentially contribute to clarify the
ARTICLE IN PRESS
Fig. 6. Atractus reticulatus. Transmission electron microscopy of the infralabial gland. (A) An acinus, which is constituted in most part by
polygonous cells, packed with low (m1) to medium (m2) electron-dense heterogeneous granules. The other type of cell present in the acinus,
less frequent, is characterized by elliptical electron-dense granules (sm). The lumen (lu) is full of secretion. ic, intercellular canaliculus. (B)
Higher magnification of the internal portion of an acinus showing microvilli (arrowheads) in the apical membrane of all types of secretory
cells (m1, m2 and sm). (C) Higher magnification of mucous cells with basal irregular nucleus (n) and cytoplasm packed with low-electron-
dense granules (sg). (D) Detail of the basal cytoplasm of the mucous cells where organelles such as mitochondria (mi) and endoplasmic
reticulum (er) are more prominent. n, nuclei; sg, secretory granules. (E) Intercellular canaliculus (ic) among neighbouring acinar cells.
Arrows, desmosomes; arrowheads, microvilli; sg, secretory granules. (F) Myo-epithelial cell, with cytoplasm rich in mitochondria (mi) and
endoplasmic reticulum (er), juxtaposed to the basis of a secretory cell. Arrowheads, cell limit; c, connective tissue; n, nuclei; sg, secretory
granule.
L. de Oliveira et al. / Toxicon 51 (2008) 898–913906
Author's personal copy
ARTICLE IN PRESSL. de Oliveira et al. / Toxicon 51 (2008) 898–913 907

Author's personal copy
relations between morphological and functionalaspects of snake’s venomous systems. The morpho-logical and histochemical analysis conducted on thethree species studied herein, clearly shows theexistence of important anatomical, microscopicaland histochemical variations among goo-eaterdipsadine snakes. From the anatomical viewpoint,the infralabial glands of A. reticulatus are signifi-cantly different from the other two species, both interms of their anatomy as to their topography withrespect to the mandible. Differently from the othertwo species, D. indica presents a much moredeveloped muscle levator anguli oris, which runsalong the whole length of the mandible, a differencealready noted by Gans (1972).
Although it has been a common practice toclassify snake’s oral glands according to tradition-ally pre-existing schemes, it is prudent to take intoaccount that the interpretation of histologicalsections is not trivial and can result in somecontroversy. More specifically, infralabial glandsin snakes have been recurrently classified as acinous,tubulous or tubule-acinous (Smith and Bellairs,1947; Lopes et al., 1981; Gopalakrishnakone andKochva, 1993). If one follows such classification forthe three species studied herein, the arrangement ofseromucous cells and mucous cells in the infralabialglands of S. mikanii, A. reticulatus and D. indica willsuggest that these glands are acinous in the formertwo species whereas they are both acinous andtubulous on the latter species.
Our results indicate that the infralabial glands ofthe three studied species are well developed andconstituted of mucous and seromucous cells, ac-cording to the classification of Gabe and Saint-Girons (1969). In A. reticulatus, the infralabialgland showed both types of cells within thesame acinus. A similar situation was observed inA. zebrinus (Antoniazzi et al., 2005), Micrurus
corallinus (Lopes et al., 1972) and Philodryas
patagoniensis (Lopes et al., 1982). In D. indica, theinfralabial gland showed two distinct regions,recognized by the morphological and histochemicalcharacteristics, with mucous tubules in the centraland anterior regions of the gland, and seromucousacini distributed in the posterior region and on theperiphery of the gland, around the mucous tubules.In contrast, the results obtained for the infralabialgland of Dipsas albifrons, indicate that it seems to becomposed in great part by seromucous cells, withmucous lobules present only in the anterior region(Antoniazzi et al., 2005). In S. mikanii, the infra-
labial gland showed to be composed essentially by asimpler secretory epithelium, with seromucous cellsorganized in acini, as previously observed byContrera et al. (1983). Mucous cells in the glandsof this species were restricted to the epithelium ofthe ducts and to the anterior region of the glandularbody, which is probably directly connected to theducts. The same distribution pattern of mucous cellsrestricted to the anterior region and duct of thegland has been described for the Duvernoy’s glandsof many colubrid species (Taub, 1966, 1967; Yoshieet al., 1982; Zalisko and Kardong, 1992), in whichthe ducts not only transport the secretion producedin the secretory units, but are also responsible formucous secretion.
Underwood (1997) has pointed out to thedifficulty in observing the ductal system in infra-labial glands. The same difficulty was noted in thepresent work, which was conducted with glandsisolated from the mandible and, as a consequence,was more focused in the observation of theglandular body. Even after serial sectioning of thesamples, the observations here presented remainedrestricted only to the ductal portion inside theglands. In the three species studied, the internalducts are constituted of a monolayer of columnarcells, full of mucous secretory granules, similar tothe pattern observed in Duvernoy’s glands (Taub,1966, 1967; Yoshie et al., 1982; Zalisko andKardong, 1992). On the other hand, differentlyfrom what is observed in the Duvernoy’s glands ofmany colubrids, where the ductal system is morecomplex and involves several structures (Zaliskoand Kardong, 1992), the infralabial glands areusually interpreted as a simple polystomaticstructure (Smith and Bellairs, 1947; Zago, 1971;Kochva, 1978; Underwood, 2002). Studies ofLaporta-Ferreira and Salomao (1991) on S. neuwiedi
contrasted with these descriptions, as they observedthe presence of a single duct in the infralabial glandprobably opening in the anterior region of the oralcavity. In the same way, Zaher (1996) has describedanterior hypertrophied ducts directed towards therostral region in the infralabial glands of Dipsas
neivai. In both papers the authors correlate theirmorphological observations with the peculiar snailfeeding habits of the studied snakes, suggesting that‘‘venomous’’ contents from the infralabial glandswould act inside the mollusc shell helping itsdetaching and extraction. This supposition matchesthe observations by Sazima (1989) on the beha-vioural feeding of D. indica. Moreover, Zaher (1996)
ARTICLE IN PRESSL. de Oliveira et al. / Toxicon 51 (2008) 898–913908
Author's personal copy
showed that the muscle intermandibularis posterior
pars posterior of D. neivai acts as a compressor ofthe infralabial gland, helping actively in the libera-tion of the secretion, a condition also observed inthe present work in D. indica.
Besides the classification of reptile oral glandsbased on their histochemical properties, another
common classification of these glands is based onthe ultrastructural (Yoshie et al., 1982; Kardongand Luchtel, 1986) aspects of their secretorygranules. Ultrastructural criteria are based mainlyin the electron density of the granules, whichgenerally exhibit high electron density when rich inproteins (and associated to serous glands), and low
ARTICLE IN PRESS
Fig. 7. Dipsas indica. Transmission electron microscopy of the infralabial gland. (A) Transversal section through a mucous tubule,
basically composed of polygonous cells packed with low-electron-dense granules (m). Among these cells, a less frequent cell type is seen,
with granules containing an electron-dense core, corresponding to the type 2 seromucous cell (sm2). lu, lumen; arrowheads, microvilli; c,
connective tissue. (B) Higher magnification of tubule luminal region. Arrowheads, microvilli; lu, lumen; m, mucous cell; sm2, seromucous
cell type 2. (C) Higher magnification of mucous cells, with basal irregular nucleus (n) and cytoplasm packed with uniform low-electron-
dense secretory granules (sg). (D) Higher magnification of seromucous cells type 1 (sm1), more abundant in the peripheral posterior part of
the gland, forming acini. The cytoplasm is full of irregular electron-dense granules (sg) and is rich in endoplasmic reticulum (er) and
mitochondria (mi) in the perinuclear region. Arrowheads, microvilli; n, nucleus. (E) Higher magnification of seromucous cells type 2 (sm2),
with granules (sg) containing an electron-dense excentric core. The cytoplasm shows a high electron density and is rich in endoplasmic
reticulum (er). m, mucous cells; n, nucleus.
L. de Oliveira et al. / Toxicon 51 (2008) 898–913 909
Author's personal copy
electron density when poor in proteins and rich inmucous substances (and consequently associated tomucous cells).
Infralabial glands of snakes are very poorlyknown as to their ultrastructure (Laporta-Ferreiraand Salomao, 1991; Baccari et al., 2002). In mostsnake species, it is accepted that they are constitutedof homogeneous mucous cells filled with secretorygranules, and of low electron density (Kochva,1978). Contrasting with this general view, theultrastructural results obtained show that theinfralabial glands of A. reticulatus, D. indica andS. mikanii are constituted not only by mucous cellswith low-electron-dense granules, but also by
seromucous cells with high-electron-dense granules,confirming our histochemical results. The ultra-structure also revealed species-specific differences inthe cellular composition, mainly evidenced by theaspect of secretory granules, with electron densitiesvarying from high to low and presenting distincttextures, both characteristics that reflect differencesin their chemical composition. Although the pur-pose of the ultrastructural analysis in this workaimed only to give additional support for eventualhistological and histochemical differences amongspecies, it is worth to keep in mind that specificglandular patterns can contain phylogenetic signifi-cance, as Tandler et al. (1986) have alreadyobserved for submandibular glands in bats.
In this work, myo-epithelial cells were observedonly in the infralabial glands of A. reticulatus but,since they are often found in oral glands of snakesand other reptiles (Yoshie et al., 1982; Kardong andLuchtel 1986; Laporta-Ferreira and Salomao,1991), they are probably present in all studiedspecies.
The ISC were observed only at ultrastructurallevel and are frequently observed in oral glands ofsnakes (Yoshie et al., 1982; Kardong and Luchtel,1986; Salomao, 1991). Differently from mammals,in snakes the ISC are not usually correlated withany special type of structural classification (Sakai,1989). In the present work, these structures werefound only in the infralabial glands of A. reticulatus,associated both with mucous and seromucous cells.Further studies may correlate this finding with adifferentiated functional role in this species.
The comparative analysis of the infralabial glandsin the three species here studied suggests thatA. reticulatus presents a larger number of differ-ences, such as mandibular topographical localiza-tion, shape of the gland, homogeneous presence ofmixed acini in the glands constituted of mucous andseromucous cells, and presence of ISC. Thesefindings may represent specializations present onlyin the clade comprising Atractus and the genera ofgoo-eaters specialized in feeding on earthworms,that is distinct from the clade comprising Sibyno-
morphus and Dipsas that comprises only genera thatfeed exclusively on snails and slugs (Ferrarezzi,1994; Fernandes, 1995; Campbell and Smith, 1998;Zaher, 1999).
Among the venomous snake families, the toxicsecretion produced by the venom glands is asso-ciated with an efficient and fast way of killing theprey (Kardong, 1996). However, the role of the
ARTICLE IN PRESS
Fig. 8. Sibynomorphus mikanii. Transmission electron micro-
scopy of the infralabial gland. (A) General aspect of a
seromucous secretory cells, showing the basal nucleus (n) and
the cytoplasm, full of secretory granules (sg), irregular in shape
and in content. c, connective tissue. (B) Higher magnification of a
secretory cell, with heterogeneous coalescent granules (sg) most
part containing low-electron-dense material but a few presenting
a more electron-dense excentric core (stars).
L. de Oliveira et al. / Toxicon 51 (2008) 898–913910

Author's personal copy
Duvernoy’s glands in colubrids is still controversialand many possible functions, such as prey immobi-lization, food lubrication and defensive use, havebeen attributed to them (Kardong, 2002). On theother hand, infralabial glands have been generallyassociated only with mucous production (Kochva,1978). Only a few reports of venomous function forsuch glands have been made in lizard species fromgenera Heloderma, Pogona and Varanus (Gabe andSaint-Girons, 1969; Kochva, 1978; Fry et al., 2006).Among snakes, as far as it has been reported,venom secretion has been always associated to thesuperior maxillary units (Underwood, 1997; Vidal,2002; Jackson, 2003). Contrasting with othersnakes, Duvernoy’s glands of goo-eater dipsadinesare very small, while their infralabial glands are welldeveloped and composed of a large number ofserous cells which have been associated to thepresence of toxins. In fact, Laporta-Ferreira (1985)and Laporta-Ferreira and Salomao (1991) reportedthat the infralabial glands of S. neuwiedi have moreprotein content than their own Duvernoy’s glands.These authors also observed that biological essaystesting extracts of the infralabial glands ofS. neuwiedi on slugs caused paralysis and death ofthe molluscs, suggesting other functions for theseglands besides the simple food lubrication. Salomaoand Sawaya (1992) and Salomao and Laporta-Ferreira (1994) studied the biological activity of theoral glands of S. mikanii and found that, althoughthe three glandular types demonstrated equivalentproteolytical activity, infralabial glands carriedlarger amounts of protein when compared tosupralabial and Duvernoy’s glands. Salomao andLaporta-Ferreira (1994) suggested that, althoughbiological essays demonstrated that the extract ofthe three types of glands of S. mikanii caused intensemucous liberation, body contortions and immobili-zation when applied on slugs, the effect of paralysiswas probably related to Duvernoy’s glands whilesupralabial and infralabial glands acted moreintensely in the digestive process. Similarly, thesecretion of the infralabial glands of A. reticulatus
and D. indica may have equivalent functions. Inaddition, it is important to consider that theinvertebrate goo-eater snakes feed on are naturallyviscous and, in many cases, lubrication may not benecessary to help prey ingestion. For this reason,different functions for these glands might beexpected, as for example, the production of toxinscapable of acting specifically on the preys ingestedby each species of snake.
This work is expected to be the first one of a seriesrelated to the goo-eater snake oral glands. Furtherstudies focused on histological serial sectios ofwhole heads, as well as on the biochemical analysisof infralabial gland secretion may clarify manyaspects related to duct topography and sites ofsecretion release, secretory composition and itseffect on preys.
Acknowledgements
This project was supported by research grantsfrom the Fundac- ao de Amparo a Pesquisa doEstado de Sao Paulo, Programme BIOTA/FAPESP(Grant no. 02/13602-4) and Conselho Nacional deDesenvolvimento Cientıfico de Tecnologico (Grantno. 47521/2004-4). We also thank CAPES forproviding a grant to Leonardo de Oliveira andCNPq for grants to Hussam Zaher (306158/2004-3)and Carlos Jared (300170/96-3). Thanks are due toValdir J. Germano and Maria Helena Ferreira fortheir efficient technical assistance. We thank DarlanTavares Feitosa for the schematic draw of theskinned heads of specimens. IBAMA providedanimal collection permits nos. 02001007052/2001(Hussam Zaher) and 02027001353/2006-58 (Leonardode Oliveira).
References
Antoniazzi, M., Oliveira, L., Zaher, H., Jared, C., 2005.
Morphological characterization of the infralabial glands of
‘‘goo-eater’’ snakes (Colubridae: Dipsadinae). IN: Abstracts
of Fifth World Congress of Herpetology, Stellemboch, South
Africa, p. 119.
Baccari, G.C., Ferrara, D., Di Matteo, L., Minucci, S., 2002.
Morphology of the salivary glands of three Squamata species:
Podarcis sicula sicula, Tarentola mauritanica and Coluber
viridiflavus. Acta Zool. (Stockholm) 83, 117–124.
Bancroft, J.D., Stevens, A., 1996. Theory and Practice of
Histological Techniques, fourth ed. Churchill Livingstone,
New York, 766pp.
Brongersma, 1956. The palato-maxillary arch in some Asiatic
Dipsadinae (Serpentes). Proc. Kon. Ned. Akad. Wet.,
Amsterdam, Ser. C 59, 439–446.
Cadle, J.E., Greene, H.W., 1993. Phylogenetic patterns, biogeo-
graphy, and the ecological structure of Neotropical snake
assemblages. In: Ricklefs, R.E., Schluter, D. (Eds.), Species
Diversity in Ecological Communities: Historical and Geo-
graphical Perspective. University of Chicago Press, Chicago,
pp. 281–293.
Campbell, J.A., Smith, E.N., 1998. A new genus and species of
colubrid snake from the Sierra de Las Minas of Gratemala.
Herpetologica 54 (2), 207–220.
ARTICLE IN PRESSL. de Oliveira et al. / Toxicon 51 (2008) 898–913 911

Author's personal copy
Contrera, M.G.D., Lopes, R.A., Costa, J.R.V., Petenusci, S.O.,
Lima-Verde, J.S., 1983. The histology of salivary glands in the
colubrid snake Sibynomorphus mikanii (Schlegel, 1837). Can.
J. Zool. 61, 936–941.
Dowling, H.G., Duellman, W.E., 1978. Systematic Herpetology:
A Synopsis of Families and Higher Categories. HISS
Publications, New York.
Dunn, E.R., 1951. The status of the snake genera Dipsas and
Sibon, a problem for ‘‘quantum evolution’’. Evolution 5,
355–358.
Fernandes, R., 1995. Phylogeny of the dipsadine snakes. Thesis
(Doctor of Philosophy), The University of Texas at Arling-
ton, 115pp.
Ferrarezzi, H., 1994. Uma sinopse dos generos e classificac- ao das
serpentes ‘‘Squamata’’. II. Famılia Colubridae. In: Nasci-
mento, L.B., Bernardes, A.T., Cotta, G.A. (Eds.), Herpeto-
logia no Brasil 1. Editora PUC-MG, Fundac- ao Esequiel Dias,
Fundac- ao Biodiversitas, Belo Horizonte, pp. 81–91.
Franco, F.L., 1994. O genero Sibynomorphus Fitzinger, 1843, no
Brasil (Colubridae: Xenodontinae, Dipsadini). Dissertac- ao
(Mestrado em Zoologia), Pontifıcia Universidade Catolica do
Rio Grande do Sul, Porto Alegre.
Fry, B.G., Vidal, N., Norman, J.A., Vonk, F.J., Scheib, H.,
Ramjan, S.F.R., Kuruppu, S., Fung, K., Hedges, S.B.,
Richardson, M.K., Hodgson, W.C., Ignjatovic, V., Summer-
hayes, R., Kochva, E., 2006. Early evolution of the venom
system in lizards and snakes. Nature 439, 584–589.
Gabe, M., Saint-Girons, H., 1969. Donnees histologiques sur les
glandes salivaires des lepidosauriens. Mem. Mus. Nat. Hist.
Nat. A 58 (1), 3–116.
Gans, C., 1972. Feeding in Dipsas indica and Dunn’s paradox.
Am. Zool. 12 (4), 730.
Gopalakrishnakone, P., Kochva, E., 1993. Histological features
of the venom apparatus of sea snake Lapemis curtus. Snake
25, 27–37.
Haas, 1964. Anatomical observations on the head of Liotyphlops
albirostris (Typhlopidae, Ophidia). Acta Zool. 45, 1–62.
Jackson, K., 2003. The evolution of venom-delivery systems in
snakes. Zool. J. Linn. Soc. 137, 227–354.
Junqueira, L.C.U., 1995. Histology revisited—technical improve-
ment promoted by the use of hydrophilic resin embedding.
Rev. Cien. Cult. 47, 92–95.
Kardong, K.V., 1996. Snake toxins and venoms: an evolutionary
perspective. Herpetologica 52 (1), 36–46.
Kardong, K.V., 2002. Colubrid snakes and Duvernoy’s ‘‘venom’’
glands. J. Toxicol.—Toxin Rev. 21 (1&2), 1–19.
Kardong, K.V., Luchtel, D.L., 1986. Ultrastructure of Duver-
noy’s gland from the wandering garter snake, Thamnophis
elegans vagrans (Serpentes, Colubridae). J. Morphol. 188,
1–13.
Karnovsky, M.J., 1965. A formaldehyde–glutaraldehyde fixative
of high osmolality for use in electron microscopy. J. Cell Biol.
27A, 137–138.
Kelly, C.M.R., Barker, N.P., Villet, M.H., 2003. Phylogenetics of
advanced snakes (Caenophidia) based on four mitochondrial
genes. Syst. Biol. 52, 439–459.
Kiernan, J.A., 2001. Histological and Histochemical Methods—
Theory and Practice, third ed. Oxford University Press,
London, 213–220.
Kley, N.J., 2006. Morphology of the lower jaw and suspensorium
in the Texas blindsnake, Leptotyphlops dulcis (Scolecophidia:
Leptotyphlopidae). J. Morphol. 267, 494–515.
Kochva, E., 1978. Oral glands of the reptilia. In: Gans, C. (Ed.),
Biology of the reptilia, vol. 8. Academic Press, London,
New York, 782pp.
Kochva, E., 1987. The origin of snakes and evolution of the
venom apparatus. Toxicon 25 (1), 65–106.
Kraus, F., Brown, W.M., 1998. Phylogenetic relationships of
colubroid snakes based on mitochondrial DNA sequences.
Zool. J. Linn. Soc. 122, 455–487.
Laporta-Ferreira, I.L., 1985. Fisioecologia da serpente molusco-
faga—Sibynomorphus neuwiedi—(Colubridae, Dipsadinae).
Dissertac- ao (Mestrado em Fisiologia)-Instituto de Biocien-
cias, Universidade de Sao Paulo, Sao Paulo.
Laporta-Ferreira, I.L., Salomao, M.G., 1991. Morphology,
physiology and toxicology of the oral glands of a tropical
cochleophagous snake, Sibynomorphus neuwiedi (Colubri-
dae—Dipsadinae). Zool. Anz. 227, 198–208.
Laporta-Ferreira, I.L., Salomao, M.G., Sawaya, P., Puorto, G.,
1988. Mecanismo de tomada de alimento por serpentes
tropicais moluscofagas (Sibynomorphus neuwiedi e Sibyno-
morphus mikanii) adaptac- oes morfofisiologicas do esqueleto
cefalico. Bol. Fisiol. Anim. Sao Paulo 12, 81–88.
Lawson, R., Slowinski, J.B., Crother, B.I., Burbrink, F.T., 2005.
Phylogeny of the Colubroidea (Serpentes): new evidence from
mitochondrial and nuclear genes. Mol. Phylogenet. Evol. 37,
581–601.
Lopes, R.A., Valeri, V., Campos, G.M., Faria, R.M., 1972.
Estudo morfologico e histoquımico de mucopolissacarıdeos
em glandulas cefalicas de Micrurus c. corallinus (Ophidia,
Elapidae). Rev. Cien. Cult. 24, 214.
Lopes, R.A., Costa, J.R.V., Petenusci, S.O., Contrera, M.G.D.,
1981. The salivary glands of Leimadophis viridis (Gunther,
1862) (Ophidea Colubridae): a morphological, morphometric
and histochemical study. Monit. Zool. Ital. (N.S.) 15,
249–261.
Lopes, R.A., Contrera, M.G.D., Costa, J.R.V., Petenusci, S.O.,
Lima-Verde, J.S., 1982. Les glandes salivaires de Philodryas
patagoniensis Girard, 1857 (Serpentes Colubridae). Etude
morphologique, morphometrique et histochimique. Arch.
Anat. Microsc. 71 (3), 175–182.
Marques, O.A.V., Eterovic, A., Sazima, I., 2001. Serpentes da
Mata Atlantica. Guia ilustrado para a Serra do Mar. Holos,
Ribeirao Preto, 184pp.
McDowell, S.B., 1987. Systematics. In: Seigel, R.A., Collins, J.T.,
Novak, S.S. (Eds.), Snakes: Ecology and Evolutionary
Biology. Macmillan Publishing, New York, NY, pp. 3–50.
Oliveira, J.L., 2001. Ecologia de tres especies de dormideira,
Sibynomorphus (Serpentes: Colubridae). Dissertac- ao (Mestra-
do em Ecologia)-Instituto de Biociencias, Universidade de
Sao Paulo, Sao Paulo.
Pearse, A.G., 1985. Histochemistry: Theoretical and Applied,
vol. 2, fourth ed. Churchill Livingstone, Edinburg, 684–687.
Pough, F.H., 1983. Feeding mechanisms, body size, and the
ecology and evolution of snakes. Am. Zool. 23, 339–342.
Pough, F.H., Groves, J.D., 1983. Specializations of the body
form and food habits of snakes. Am. Zool. 23, 443–454.
Sakai, T., 1989. Major ocular glands (Harderian gland and
lacrimal gland) of the musk shrew (Suncus murinus) with a
review on the comparative anatomy and histology of the
mammalian lacrimal glands. J. Morphol. 2001, 39–57.
Salomao, M.G., 1991. Estrutura e secrec- ao das glandulas de
Duvernoy de Sibynomorphus mikanii (Colubridae: Dipsadi-
nae) e Philodryas olfersii (Colubridae: Xenodontinae), e das
ARTICLE IN PRESSL. de Oliveira et al. / Toxicon 51 (2008) 898–913912

Author's personal copy
glandulas de veneno de Bothrops jararaca (Viperidae:
Crotalinae) e Micrurus frontalis (Elapidae: Elapinae) e sua
influencia dos estados de alimentac- ao e jejum. Tese (doutor-
ado em Fisiologia)—Instituto de Biociencias, Universidade de
Sao Paulo, Sao Paulo, 122pp.
Salomao, M.G., Laporta-Ferreira, I.L., 1994. The role of
secretions from the supralabial, infralabial, and Duvernoy’s
glands of the slug-eating snake Sibynomorphus mikanii
(Colubridae: Dipsadinae) in the immobilization of molluscan
prey. J. Herpetol. 28, 369–371.
Salomao, M.G., Sawaya, P., 1992. Propriedades toxicas da saliva
de serpentes aglifas: Sibynomorphus mikani (Colubridae:
Dipsadinae). Ann. Acad. Bras. Ci. 64 (2).
Savitzky, A.H., 1983. Coadapted character complexes among
snakes: fossoriality, piscivory, and durophagy. Amer. Zool.
23, 397–409.
Sazima, I., 1989. Feeding behavior of the snail-eating snake,
Dipsas indica. J. Herpetol. 23, 464–468.
Smith, M., Bellairs, A.A., 1947. The head glands of snakes, with
remarks on the evolution of the parotid gland and teeth of the
opisthoglypha. J. Linn. Soc. (Zool.) (London) 41, 353–368.
Tandler, B., Nagato, T., Phillips, C.J., 1986. Systematic implica-
tions of comparative ultrastructure of secretory acini in the
submandibular salivary gland in Artibeus (Chiroptera: Phyl-
lostomidae). J. Mammal. 67 (1), 81–90.
Taub, A.M., 1966. Ophidian cephalic glands. J. Morphol. 118,
529–542.
Taub, A.M., 1967. Comparative studies on Duvernoy’s gland of
colubrid snakes. Bull. Am. Mus. Nat. Hist. 138, 1–50.
Underwood, G., 1967. A Contribution to the Classification of
Snakes. British Museum (Natural History), London, UK.
Underwood, G., 1997. An overview of venomous snake evolu-
tion. Symp. Zool. Soc. Lond. 70: 1-13. In: Thorpe, R.S.,
Wuster, W., Malhotra, A. (Eds.), Venomous Snake. Ecology,
Evolution and Snakebite. The Zoological Society of London.
Clarendon Press, Oxford.
Underwood, G., 2002. On the rictal structures of some snakes.
Herpetologica 58 (1), 1–17.
Vidal, N., 2002. Colubroid systematics: evidence for an early
appearance of the venom apparatus followed by extensive
evolutionary timkering. J. Toxicol.—Toxin Rev. 21 (1&2),
21–41.
Voris, H.K., Voris, H.H., 1983. Feeding strategies in marine
snakes: an analysis of evolutionary, morphological, behavior-
al and ecological relationships. Am. Zool. 23, 411–425.
Yoshie, S., Ishiyama, M., Ogawa, T., 1982. Fine structure of
Duvernoy’s gland of the japanese colubrid snake, Rhabdophis
tigrinus. Arch. Histol. Jap. 45 (4), 375–384.
Zago, D.A., 1971. Estudo morfologico e histoquımico de
glandulas salivares relacionadas com a evoluc- ao da func- ao
venenosa nos ofıdios. Tese (doutorado), Universidade de Sao
Paulo, Sao Paulo.
Zaher, H., 1994. Comments on the evolution of the jaw adductor
musculature of snakes. Zool. Lin. Soc. 111, 339–384.
Zaher, H., 1996. A musculatura associada a glandula infralabial
de Dipsas neivai: um novo sistema de inoculac- ao de veneno
relacionado a malacofagia (Serpentes, Dipsadinae). In:
Resumos do XXI Congresso Brasileiro de Zoologia,
vol. 940, p. 201.
Zaher, H., 1999. Hemipenial morphology of the South American
Xenodontine snakes, with a proposal for a monophyletic
Xenodontinae and a reappraisal of Colubroid hemipenes.
Bull. Am. Mus. Nat. Hist. 240, 168pp.
Zalisko, E.J., Kardong, K., 1992. Histology and histochemistry
of the Duvernoy’s gland of the brown snake Boiga irregularis
(Colubridae). Copeia 1992 (3), 791–799.
ARTICLE IN PRESSL. de Oliveira et al. / Toxicon 51 (2008) 898–913 913




















