
Ultrastructure of poison glands of South American frogs: A comparison betweenPhysalaemus albonotatus...
12
Ultrastructure of Poison Glands of South American frogs: A Comparison Between Physalaemus albonotatus and Leptodactylus chaquensis (Anura: Leptodactylidae) Blanca Beatriz Alvarez, 1 Giovanni Delfino, 2 Daniele Nosi, 3 * and Alessandro Terreni 4 1 Departamento de Biologı ´a, Universidad Nacional del Nordeste, 3400 Corrientes, Argentina 2 Dipartimento di Biologia Animale e Genetica dell’Universita ` , 50125 Firenze, Italy 3 Dipartimento di Anatomia, Istologia e Medicina legale dell’Universita ` , 50134 Firenze, Italy 4 Laboratorio Centrale di Analisi Biochimico-Cliniche, Azienda Ospedaliera di Careggi, 50134 Firenze, Italy ABSTRACT Serous (poison) cutaneous glands of the lep- todactylid species Physalaemus albonotatus and Lepto- dactylus chaquensis were compared using light and trans- mission electron microscopy. Glands in the two species share structural traits common in anurans, including the peripheral contractile sheath (myoepithelium) and the syncytial secretory unit that produces, stores, and modi- fies the poison. At the ultrastructural level, early steps of poison production are also similar and fit the usual path of proteosynthesis, involving rough endoplasmic reticulum (RER) and Golgi stacks (dictyosomes) in the peripheral syncytial cytoplasm. However, several differences are ob- vious during the maturational processes that lead post- Golgian products to their ultimate ultrastructural traits. In P. albonotatus, the dense product released from the dictyosomes acquires a thick repeating substructure, which, however, becomes looser in the inner portion of the syncytium. In L. chaquensis, serous maturation involves gradual condensation, and opaque, somewhat “vacuolized” granules are formed. These different maturational paths expressed during poison manufacturing in the two species agree with the polyphyletic origin of the family Leptodac- tylidae. On the other hand, data collected for P. albonota- tus fit previous findings from P. biligonigerus and stress the view that poisons produced by congeneric species share similar (or identical) ultrastructural features. J. Morphol. 263:247–258, 2005. © 2004 Wiley-Liss, Inc. KEY WORDS: ultrastructure; serous glands; skin; Lepto- dactylidae Poison cutaneous glands in anurans are referred to the main serous (or proteinaceous) cell line type occurring in the skin of anamniotic aquatic verte- brates (Quay, 1972). Poison glands are usually de- scribed as granular because of light microscopic fea- tures of their products, consisting of discrete structures, subspherical to elongate in shape, and characterized by various degrees of density and stain affinity. Based on transmission electron micro- scope criteria, these structures can, however, be as- cribed to various morphological categories (Delfino et al.,1992), so that the unifying concept of secretory granules seems inadequate to define such a hetero- geneous class of products. Actually, these “granules” represent the storage sites of various classes of ac- tive molecules (involved in both skin homeostasis and chemical defense, Daly et al., 1987) responsible for adaptation to terrestrial habitats (Toledo and Jared, 1995). They should be properly regarded as complex organelles that hold active molecules pro- duced within the inner cell membrane system (namely, rough endoplasmic reticulum (RER), smooth endoplasmic reticulum (SER), and Golgi ap- paratus: the biosynthetic machinery) or in the secre- tory unit hyaloplasm, to be later transferred into these storage structures (Delfino, 1979, 1991). The occurrence of different morphological classes of serous gland products, among anurans, contrasts with the common biosynthetic machinery shared by serous glands throughout the order. The different features of the serous products mostly depend on the post-Golgian (maturational) phase, which proceeds along specific pathways in different anuran families. Since a comparison between various genera revealed that the production of skin poisons with similar fea- tures is a consistent trait in species of the same genus (Table 1), we desired to extend our investiga- tions to different genera of the same family to verify any phylogenetic relevance of this morphological similarity. Accordingly, we studied the Argentine leptodactylid frogs Leptodactylus chaquensis and Physalaemus albonotatus (family Leptodactilyli- dae). Leptodactylidae is regarded as a paraphyletic taxon that includes hyloid frogs that do not share the derived traits usually indicative of the family status in anurans (Ford and Cannatella, 1993). The genera investigated are large groups, with broad spectra of diversity, representative of their peculiar *Correspondence to: Daniele Nosi, Dipartimento di Anatomia, Isto- logia e Medicina legale, Universita ` degli Studi di Firenze, Viale Mor- gagni 85, 50134 Firenze, Italy. E-mail: [email protected] Published online 21 December 2004 in Wiley InterScience (www.interscience.wiley.com) DOI: 10.1002/jmor.10301 JOURNAL OF MORPHOLOGY 263:247–258 (2005) © 2004 WILEY-LISS, INC.
Transcript of Ultrastructure of poison glands of South American frogs: A comparison betweenPhysalaemus albonotatus...

Ultrastructure of Poison Glands of South Americanfrogs: A Comparison Between Physalaemus albonotatusand Leptodactylus chaquensis (Anura: Leptodactylidae)Blanca Beatriz Alvarez,1 Giovanni Delfino,2 Daniele Nosi,3* and Alessandro Terreni4
1Departamento de Biologıa, Universidad Nacional del Nordeste, 3400 Corrientes, Argentina2Dipartimento di Biologia Animale e Genetica dell’Universita, 50125 Firenze, Italy3Dipartimento di Anatomia, Istologia e Medicina legale dell’Universita, 50134 Firenze, Italy4Laboratorio Centrale di Analisi Biochimico-Cliniche, Azienda Ospedaliera di Careggi, 50134 Firenze, Italy
ABSTRACT Serous (poison) cutaneous glands of the lep-todactylid species Physalaemus albonotatus and Lepto-dactylus chaquensis were compared using light and trans-mission electron microscopy. Glands in the two speciesshare structural traits common in anurans, including theperipheral contractile sheath (myoepithelium) and thesyncytial secretory unit that produces, stores, and modi-fies the poison. At the ultrastructural level, early steps ofpoison production are also similar and fit the usual path ofproteosynthesis, involving rough endoplasmic reticulum(RER) and Golgi stacks (dictyosomes) in the peripheralsyncytial cytoplasm. However, several differences are ob-vious during the maturational processes that lead post-Golgian products to their ultimate ultrastructural traits.In P. albonotatus, the dense product released from thedictyosomes acquires a thick repeating substructure,which, however, becomes looser in the inner portion of thesyncytium. In L. chaquensis, serous maturation involvesgradual condensation, and opaque, somewhat “vacuolized”granules are formed. These different maturational pathsexpressed during poison manufacturing in the two speciesagree with the polyphyletic origin of the family Leptodac-tylidae. On the other hand, data collected for P. albonota-tus fit previous findings from P. biligonigerus and stressthe view that poisons produced by congeneric speciesshare similar (or identical) ultrastructural features. J.Morphol. 263:247–258, 2005. © 2004 Wiley-Liss, Inc.
KEY WORDS: ultrastructure; serous glands; skin; Lepto-dactylidae
Poison cutaneous glands in anurans are referredto the main serous (or proteinaceous) cell line typeoccurring in the skin of anamniotic aquatic verte-brates (Quay, 1972). Poison glands are usually de-scribed as granular because of light microscopic fea-tures of their products, consisting of discretestructures, subspherical to elongate in shape, andcharacterized by various degrees of density andstain affinity. Based on transmission electron micro-scope criteria, these structures can, however, be as-cribed to various morphological categories (Delfinoet al.,1992), so that the unifying concept of secretorygranules seems inadequate to define such a hetero-
geneous class of products. Actually, these “granules”represent the storage sites of various classes of ac-tive molecules (involved in both skin homeostasisand chemical defense, Daly et al., 1987) responsiblefor adaptation to terrestrial habitats (Toledo andJared, 1995). They should be properly regarded ascomplex organelles that hold active molecules pro-duced within the inner cell membrane system(namely, rough endoplasmic reticulum (RER),smooth endoplasmic reticulum (SER), and Golgi ap-paratus: the biosynthetic machinery) or in the secre-tory unit hyaloplasm, to be later transferred intothese storage structures (Delfino, 1979, 1991).
The occurrence of different morphological classesof serous gland products, among anurans, contrastswith the common biosynthetic machinery shared byserous glands throughout the order. The differentfeatures of the serous products mostly depend on thepost-Golgian (maturational) phase, which proceedsalong specific pathways in different anuran families.Since a comparison between various genera revealedthat the production of skin poisons with similar fea-tures is a consistent trait in species of the samegenus (Table 1), we desired to extend our investiga-tions to different genera of the same family to verifyany phylogenetic relevance of this morphologicalsimilarity. Accordingly, we studied the Argentineleptodactylid frogs Leptodactylus chaquensis andPhysalaemus albonotatus (family Leptodactilyli-dae). Leptodactylidae is regarded as a paraphyletictaxon that includes hyloid frogs that do not sharethe derived traits usually indicative of the familystatus in anurans (Ford and Cannatella, 1993). Thegenera investigated are large groups, with broadspectra of diversity, representative of their peculiar
*Correspondence to: Daniele Nosi, Dipartimento di Anatomia, Isto-logia e Medicina legale, Universita degli Studi di Firenze, Viale Mor-gagni 85, 50134 Firenze, Italy. E-mail: [email protected]
Published online 21 December 2004 inWiley InterScience (www.interscience.wiley.com)DOI: 10.1002/jmor.10301
JOURNAL OF MORPHOLOGY 263:247–258 (2005)
© 2004 WILEY-LISS, INC.
phyletic histories. These wide ranges of diversityreflect a single evolutionary line of adaptation inLeptodactylus but different evolutionary lines inPhysalaemus (Cei, 1980).
MATERIALS AND METHODS
Adult specimens of Physalaemus albonotatus and Leptodacty-lus chaquensis were collected near Resistencia (Chaco, RepublicaArgentina) and kept in the Laboratory of Comparative Anatomy
(Departamento de Biologia, UNNE, Corrientes). Skin strips (4mm2 in surface area) were removed from animals sacrificed with0.2% chlorobutanol and treated (4 h, 4°C) with an aldehyde mix-ture according to Karnovsky (1965). The tissue fragments werethen washed with 0.1 M, pH 7 cacodylate buffer (the same as theprefixative solution), and sent in 2–4 ml of this buffer (with theaddition of a drop of glutaraldehyde) to the Dipartimento diBiologia Animale e Genetica (Universita di Firenze, Italy). Here,the skin specimens were rinsed once more, reduced in size, andpostfixed in 1% OsO4 (90 min), again using the cacodylate buffer.After rinsing in this solution the samples were dehydrated in
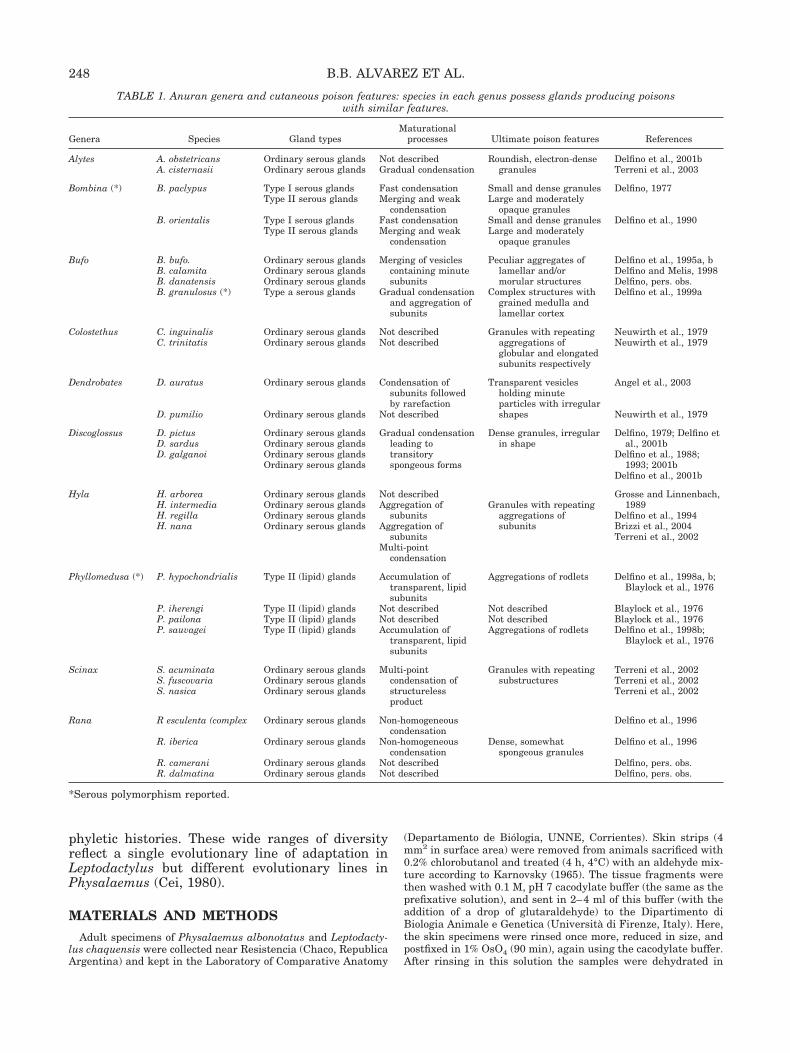
TABLE 1. Anuran genera and cutaneous poison features: species in each genus possess glands producing poisonswith similar features.
Genera Species Gland typesMaturational
processes Ultimate poison features References
Alytes A. obstetricansA. cisternasii
Ordinary serous glandsOrdinary serous glands
Not describedGradual condensation
Roundish, electron-densegranules
Delfino et al., 2001bTerreni et al., 2003
Bombina (*) B. paclypus Type I serous glands Fast condensation Small and dense granules Delfino, 1977Type II serous glands Merging and weak
condensationLarge and moderately
opaque granulesB. orientalis Type I serous glands Fast condensation Small and dense granules Delfino et al., 1990
Type II serous glands Merging and weakcondensation
Large and moderatelyopaque granules
Bufo B. bufo.B. calamitaB. danatensisB. granulosus (*)
Ordinary serous glandsOrdinary serous glandsOrdinary serous glandsType a serous glands
Merging of vesiclescontaining minutesubunits
Gradual condensationand aggregation ofsubunits
Peculiar aggregates oflamellar and/ormorular structures
Complex structures withgrained medulla andlamellar cortex
Delfino et al., 1995a, bDelfino and Melis, 1998Delfino, pers. obs.Delfino et al., 1999a
Colostethus C. inguinalisC. trinitatis
Ordinary serous glandsOrdinary serous glands
Not describedNot described
Granules with repeatingaggregations ofglobular and elongatedsubunits respectively
Neuwirth et al., 1979Neuwirth et al., 1979
Dendrobates D. auratus
D. pumilio
Ordinary serous glands
Ordinary serous glands
Condensation ofsubunits followedby rarefaction
Not described
Transparent vesiclesholding minuteparticles with irregularshapes
Angel et al., 2003
Neuwirth et al., 1979
Discoglossus D. pictusD. sardusD. galganoi
Ordinary serous glandsOrdinary serous glandsOrdinary serous glandsOrdinary serous glands
Gradual condensationleading totransitoryspongeous forms
Dense granules, irregularin shape
Delfino, 1979; Delfino etal., 2001b
Delfino et al., 1988;1993; 2001b
Delfino et al., 2001b
Hyla H. arboreaH. intermediaH. regillaH. nana
Ordinary serous glandsOrdinary serous glandsOrdinary serous glandsOrdinary serous glands
Not describedAggregation of
subunitsAggregation of
subunitsMulti-point
condensation
Granules with repeatingaggregations ofsubunits
Grosse and Linnenbach,1989
Delfino et al., 1994Brizzi et al., 2004Terreni et al., 2002
Phyllomedusa (*) P. hypochondrialis
P. iherengiP. pailonaP. sauvagei
Type II (lipid) glands
Type II (lipid) glandsType II (lipid) glandsType II (lipid) glands
Accumulation oftransparent, lipidsubunits
Not describedNot describedAccumulation of
transparent, lipidsubunits
Aggregations of rodlets
Not describedNot describedAggregations of rodlets
Delfino et al., 1998a, b;Blaylock et al., 1976
Blaylock et al., 1976Blaylock et al., 1976Delfino et al., 1998b;
Blaylock et al., 1976
Scinax S. acuminataS. fuscovariaS. nasica
Ordinary serous glandsOrdinary serous glandsOrdinary serous glands
Multi-pointcondensation ofstructurelessproduct
Granules with repeatingsubstructures
Terreni et al., 2002Terreni et al., 2002Terreni et al., 2002
Rana R esculenta (complex
R. iberica
R. cameraniR. dalmatina
Ordinary serous glands
Ordinary serous glands
Ordinary serous glandsOrdinary serous glands
Non-homogeneouscondensation
Non-homogeneouscondensation
Not describedNot described
Dense, somewhatspongeous granules
Delfino et al., 1996
Delfino et al., 1996
Delfino, pers. obs.Delfino, pers. obs.
*Serous polymorphism reported.
248 B.B. ALVAREZ ET AL.

graded ethanol, soaked in propyleneoxide, and infiltrated in Epon812 to obtain flat blocks by polymerization. Epon blocks were cutwith a NOVA LKB ultramicrotome into semithin (1–2 �m) andultrathin (yellow-white, interference color) sections. Semithinsections were stained with buffered Toluidine blue for prelimi-nary light microscope (LM) observations. Ultrathin sections werecollected on 300-mesh uncoated copper grids, then electron-densestained in sequence with hydroalcoholic uranyl acetate and alka-line lead citrate solutions (saturated and 2 mg/ml, respectively).Samples were observed (80 kV) under a Siemens 101 transmis-sion electron microscope (TEM).
RESULTSLight Microscope Observations
An LM comparison between serous glands in thespecies investigated disclosed identical structuraltraits along with obvious differences in secretoryproduct features. We first describe common featuresfollowed by contrasting morphofunctional traits.
The largest and most metabolically active compo-nent of the serous glands is the secretory unit (syn-cytial in structure and intradermal in position),which is provided with a muscular (myoepithelial)sheath (Fig. 1A,D,F). As usual in anurans, serousglands lack a proper lumen, which means that thepoison is stored within the serous syncytium (Fig.1A,D,F). The secretory units are roundish, althoughwhere glands crowd together they appear irregu-larly pear-shaped, with nuclei scattered in a singleperipheral row (Fig. 1A). Starting from the periph-ery, secretory activity proceeds towards the upperpole of the secretory unit, at the epidermis dermisboundary. Here the secretory units joins the glandneck (or intercalated tract), where the tips of themyoepithelial cells converge (Fig. 1B,C,E). The in-tercalated tract consists of a few layers of cells witha high nuclear–cytoplasmic ratio (Fig. 1A,B,E), andrepresents the regenerative compartment involvedin gland turnover. Although these stem cells mayinclude both adenoblasts and myoblasts, turnoverprocesses in adult specimens mostly involve secre-tory cytodifferentiation, as suggested by the occur-rence of single adenoblasts containing secretorygranules of various size and density (Fig. 1C). Thesesingle secretory cells will later merge into the syn-cytium. The gland neck possesses an inner compart-ment (the intercalated lumen, Fig. 1B,E), which in-cludes a slightly enlarged cavity extending downinto the secretory syncytium (Fig. 1D, insert). Thisextension—an extremely reduced gland lumen—iscompletely empty, because secretory granules col-lect around the limiting membrane that correspondsto the syncytium plasmalemma. The neck lumenexhibits a slender peripheral extension toward theepidermis, continuous with the gland duct (Fig.1B,E). According to functional criteria, the duct rep-resents the end portion of the gland and resembles atotally intraepidermal channel, lined by keratinizedcells (Fig. 1F). Because neck and duct are notaligned along the main gland axis, their respective
lumina usually appear in sequential sections (Fig.1A,B,E,F).
Secretory granules in serous glands of Physalae-mus albonotatus (Fig. 1A) are ellipsoidal to roundishin shape and are characterized by various degrees ofdensity. A prominent maturational gradient is de-tectable among serous aggregates, with smaller anddenser secretory granules at the periphery andlarger and paler granules toward the interior. Themost peripheral syncytial areas also contain small,moderately opaque granules. As later described inthe TEM results, this thin secretory material de-rives from early (post-Golgi) activity.
In contrast to the above findings, serous glands ofLeptodactylus chaquensis contain secretory gran-ules with homogenous features: these are dense,roundish particles, provided with an evident halo,that does not exhibit any obvious maturational gra-dient (Fig. 1D, insert, E,F).
Smaller glands are scattered between the serousunits in both species; these are mucous glands andpossess an obvious lumen, its width depending onthe functional stage of the encircling mucocytes (Fig.1A,D,F). The mucous product consists of discrete,dense granules when intracytoplasmic (Fig,1A,D,F), whereas it is more transparent and struc-tureless when intraluminal (Fig. 1D,F).
Electron Microscope ObservationsPhysalaemus albonotatus. Because of the centrip-
etal, functional gradient usual in serous glands, un-der TEM the most attention was paid to the periph-eral areas of the secretory syncytium, wherebiosynthesis and early maturation are interlaced.The peripheral cytoplasm of the secretory unit isenveloped by a continuous layer of thin myoepithe-lial cells (Figs. 2A, 3B), which exhibit dense plaquesamong myofilaments as peculiar components oftheir cytoskeleton (Fig. 2B). Further typical featuresin these contractile cells are scattered profiles ofsarcoplasmic reticulum (SR, Fig. 2C) and minute,spherical inpocketings (caveolae) from the plasmamembrane. In tangential sections, these caveolaeform a somewhat honeycomb pattern due to theirthick arrangement (Fig. 2D). In some instances, sin-gle caveolae can be seen in close contiguity to com-plements of the sarcoplasmic reticulum (Fig. 2E).The narrow space between secretory and contractilecompartments contains thin cell processes; that arecylindrical in shape and contain aligned microtu-bules: thus, they resemble axonal processes (Fig.2B). A remarkable functional trait in adult glands isthe sharp contrast between large amounts of maturegranules and resting biosynthesis apparatus (Figs.2A, 3B,F). Biosynthetic organelles consist of roughendoplasmic reticulum (RER, Fig. 2A,B,H) andGolgi stacks (Fig. 2F,G). RER cisterns are flat (Fig.2A) and contain a slightly opaque product (Fig. 2B),whereas Golgi saccules are mostly dilated and irreg-
249POISON GLAND ULTRASTRUCTURE IN LEPTODACTYLIDAE

ular in profile (Fig. 2F). Slender saccules are, how-ever, provided with enlarged ends, where a struc-tureless material tends to aggregate in denseparticles (Fig. 2G). Once released as vesicles fromthe Golgi apparatus, this product forms small gran-ules exclusively found in the most peripheral cyto-plasm (Fig. 2F). Both the amorphous product anddense granules are contained within post-Golgi se-cretory vesicles engaged in merging processes (Fig.2H). Larger vesicles thus arise, where the secretory
material becomes rearranged in a labyrinthine pat-tern, resulting from alternating dense and paleproducts (Fig. 3A). Secretory vesicles gradually turninto secretory pregranules, while the labyrinthinepattern changes into a loose aggregation of subunits(Fig. 3A). As expected from LM microscope observa-tions, under TEM the ultimate secretory granulesexhibit an obvious range of ultrastructural featureseven in the peripheral cytoplasm, where they areirregular in shape and vary in density (Fig. 3B).
Fig. 1. Features of serous cutaneous glands in Physalaemus albonotatus (A–C, serial section) and Leptodactylus chaquensis (D–F).LM. A: Main architecture of serous glands including duct, intercalary tract, secretory unit, and myoepithelium. Serous product isstored within the secretory unit syncytium and exhibits an obvious maturational gradient; arrow points to the duct lumen.B: Transitional region between intercalary tract (neck) and duct (compare with E); notice myoepithelial cells converging toward thegland neck; arrowhead and arrow point to the lumina of intercalary and duct, respectively. C: Peripheral region of the gland neck:notice adenoblast differentiating from the stem cell pool. D: Low- power magnification showing differences between serous and mucousglands: notice obvious lumina in mucous glands (closed and open, large asterisks) and the solid serous unit (left). Also notice theexiguous sub-intercalary lumen in serous gland (small, closed asterisk in the insert). E: Transition region between neck and duct(compare with B), as shown by continuous lumina: arrowhead points to the neck lumen and arrow to the duct lumen. F: Mainorganization of serous gland; the intercalary tract appears in tangential section since the functional axis of the gland is not alignedwith the neck axis. Arrow points to the duct lumen. d, gland duct; it, intercalary tract; mec, myoepithelial cells; mg, mucous gland;SIII-VI, maturational stages of serous product; sc, differentiating secretory cell (adenoblast).
250 B.B. ALVAREZ ET AL.
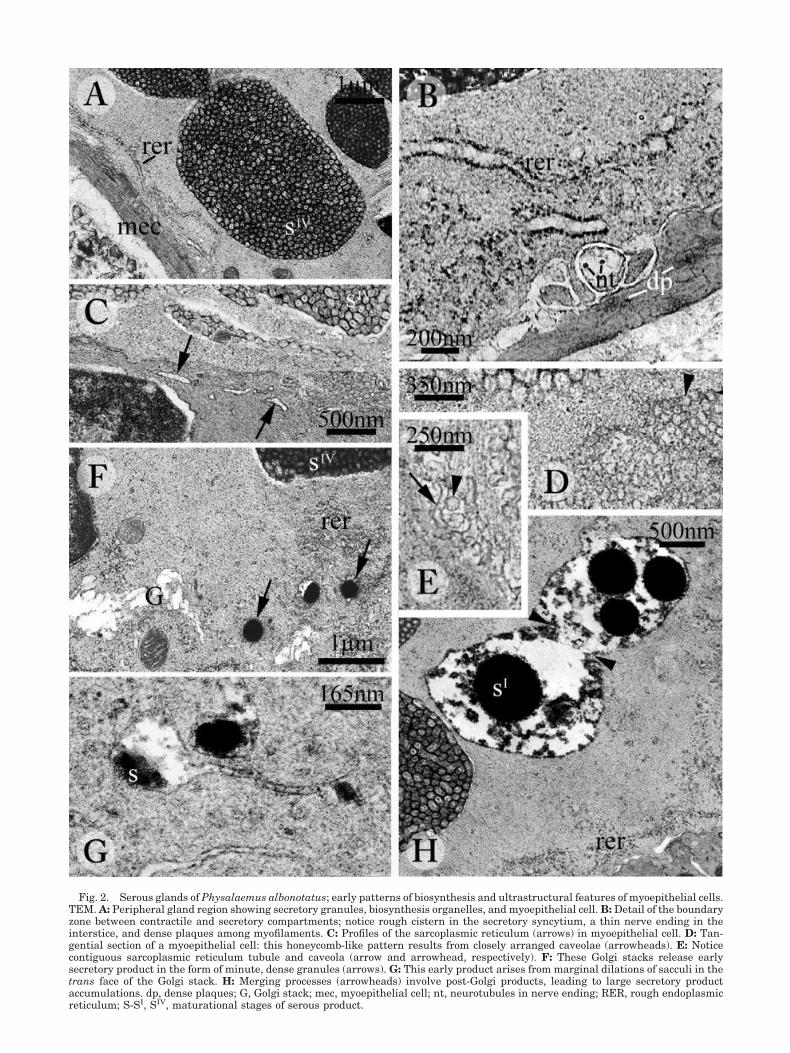
Fig. 2. Serous glands of Physalaemus albonotatus; early patterns of biosynthesis and ultrastructural features of myoepithelial cells.TEM. A: Peripheral gland region showing secretory granules, biosynthesis organelles, and myoepithelial cell. B: Detail of the boundaryzone between contractile and secretory compartments; notice rough cistern in the secretory syncytium, a thin nerve ending in theinterstice, and dense plaques among myofilaments. C: Profiles of the sarcoplasmic reticulum (arrows) in myoepithelial cell. D: Tan-gential section of a myoepithelial cell: this honeycomb-like pattern results from closely arranged caveolae (arrowheads). E: Noticecontiguous sarcoplasmic reticulum tubule and caveola (arrow and arrowhead, respectively). F: These Golgi stacks release earlysecretory product in the form of minute, dense granules (arrows). G: This early product arises from marginal dilations of sacculi in thetrans face of the Golgi stack. H: Merging processes (arrowheads) involve post-Golgi products, leading to large secretory productaccumulations. dp, dense plaques; G, Golgi stack; mec, myoepithelial cell; nt, neurotubules in nerve ending; RER, rough endoplasmicreticulum; S-SI, SIV, maturational stages of serous product.

Figure 3.
252 B.B. ALVAREZ ET AL.

Higher magnification of these secretory deposits dis-closes a consistent repeating substructure, withminute subunits arranged in a thick pattern (Figs.2A, 3A,C,H). Just distally from the myoepithelium-secretory syncytium boundary, secretory granulesare more homogenous in shape as well as density;they are densely packed, their flat surfaces facingeach other (Fig. 3D), so that their circular profilesappear somewhat polygonal in section (Fig. 3C). Asa result of crowding, extremely thin cytoplasm lay-ers separate contiguous granules (Fig. 3D) and fol-low the complex membrane relationships that leadto merging processes (Fig. 3E). A characteristic traitof maturational processes is the relationship be-tween central granules and the secretory syncytium.The slender halo that encircles the serous productsextends locally toward the cytoplasm, isolatingsmall areas that are circular to elliptical in sectionand exhibit complex patterns of parallel membranes(Fig. 3F). These membranes may belong to the cellinner membrane system (e.g., the endoplasmic retic-ulum) or originate from invagination of the granulelimiting membrane. In some instances, these smallcytoplasmic portions are regularly aligned aroundthe granule periphery in a recurrent pattern (Fig.3G). Proceeding toward the inner region of the syn-cytium, the thickly arranged subunits of the serousdeposits gradually vanish (Fig. 3H), explaining thepale granules detected in the inner syncytium underLM.
Leptodactylus chaquensis. Low-power TEM mi-crographs confirm the basic morphological traitsin serous glands of this species, including secre-tory granule features. Secretory granules mayvary in shape, but exhibit rather homogenouselectron-opacities (Fig. 4A). Since they are pro-vided with wide halos, the inner portion of thesecretory syncytium contains an obvious cavitarysystem and sharply contrasts with the peripheralcytoplasm, which has a solid background and ac-commodates a single row of nuclei (Fig. 4A). Be-cause the dilated perigranular halos merge to-gether, the inner region of the secretory unitseems to contain a lumen with an irregular profile,in which serous product is stored (Fig. 4A). How-ever, at higher magnification the peripheral syn-
cytium appears to be continuous with the cyto-plasmic partitions inserted between the granulesin the inner syncytium (Fig. 4B), demonstratingthat the “lumen” is actually a complex of contigu-ous compartments. As in the former species, thespace between secretory unit and contractilesheath is exiguous and holds small neurite end-ings with dense-cored synaptic vesicles (Fig. 4C).Contractile sheath and secretory unit can be eas-ily distinguished by their different cytoplasmiccontents: the myoepithelial cells hold denseplaques along the myofilament bundles (Fig. 4D),together with complements of the sarcoplasmicreticulum, both vesicular (Fig. 4C) and tubular inshape (Fig. 4E); the secretory syncytium containsslender cisterns of the RER (Fig. 4C). Further-more, there are no plasma membranes interveningbetween nuclei in the peripheral syncytialcytoplasm (Fig. 4B), whereas the contractile com-partment consists of discrete smooth muscle cells,linked by desmosomes that alternate with periph-eralcaveolae (Fig. 4F). In addition to flattenedRER profiles, containing moderately opaque ma-terial, the biosynthetic apparatus of the secretorysyncytium includes stacked saccules of the Golgiapparatus (Figs. 4G, 5A), involved in condensationof serous product (Fig. 4H).
In comparison with Leptodactylus chaquensis,poison biosynthesis and maturation in Physalae-mus albonotatus proceeds according to a less ob-vious sequential development. Secretory granulesderive from homogeneous thickening of the prod-uct contained in Golgi vesicles (Fig. 4H). Conden-sation may be either restricted to foci or diffuse, sothat secretory granules show patterns of alternat-ing paler and denser areas, or homogenous opacity(Fig. 5A). Advanced maturation leads to spongy-like granules alternating with granules of compactstructure (Fig. 4A). Small, opaque particles areseldom detected in large vesicles that also containa finely dispersed material (Fig. 5B). The widehalo around secretory granules contains a net-work of minute structures (Fig. 5A); these are thincytoplasmic outgrowths emanating from the syn-cytium (Fig. 5C) and arranged in a branched pat-tern (Fig. 5D).
Fig. 3. Maturational processes affecting the secretory product in serous cutaneous glands of Physalaemus albonotatus. TEM.A: The dense, discrete secretory aggregates contained in the post-Golgi vesicles tend to change into a labyrinthine-shaped material,due to random distribution of opaque and light products (SII). In later granules (SIII), discrete subunits appear, which acquire a thickerarrangement, with an obvious repeating substructure (SIV). B: As a result of this secondary condensation, the secretory depositsaccumulating in the syncytium tend to resemble real granules, namely, opaque storage bodies. Secretory granules crowd in theperipheral cytoplasm and mask the maturational gradient; this low-magnification picture does not resolve any substructure insecretory granules. C: Higher magnification shows that the recurrent aggregation of subunits represents a common trait in theseperipheral granules, which face each other with flat surfaces (opposite arrows). D,E: Thin cytoplasmic screens are interposed betweencontiguous serous deposits, molding their profiles and following granule gear-like relationships (arrows in E); notice the narrow halosencircling serous aggregates. F: These narrow compartments may evaginate into the syncytium, encircling small cytoplasmic portions,provided with inner membrane systems. G: In some instances, these small cytoplasmic portions exhibit a regular arrangement aroundthe granules. H: As described under LM, the thick subunit arrangement tends to be progressively weaker in the inner syncytium(SIV-SVI). h, perigranular halo; mec, myoepithelial cell; RER, rough endoplasmic reticulum.
253POISON GLAND ULTRASTRUCTURE IN LEPTODACTYLIDAE
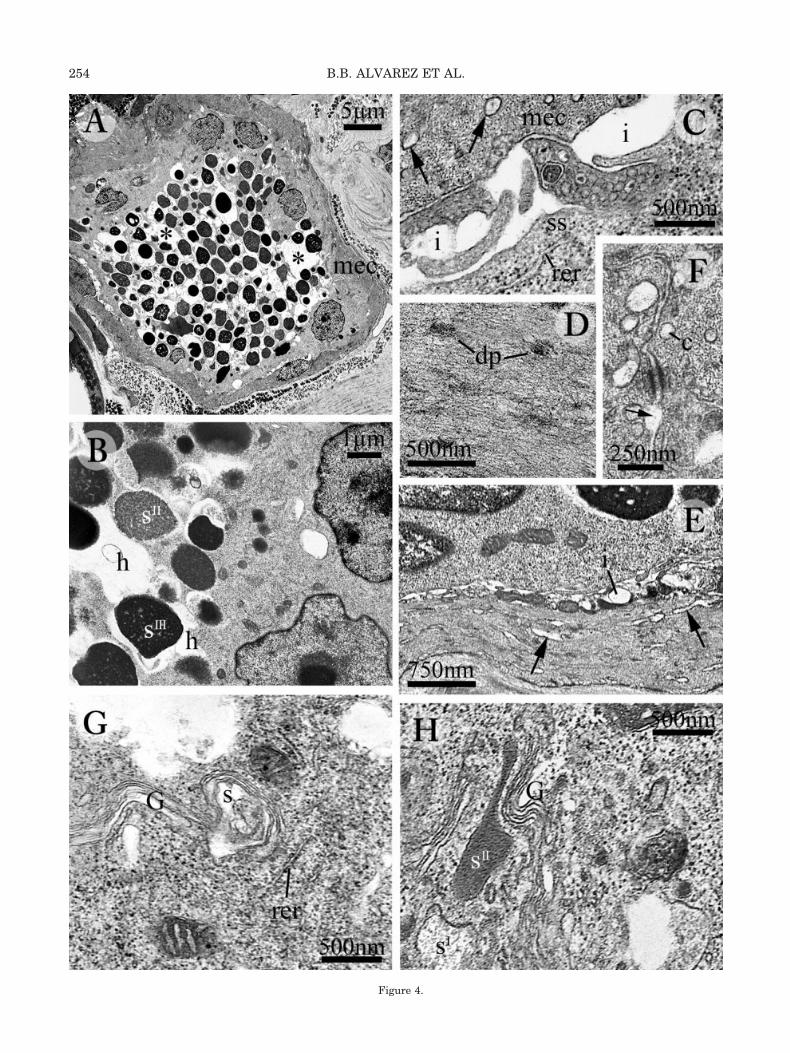
Figure 4.
254 B.B. ALVAREZ ET AL.

DISCUSSIONThe ultrastructural traits we have detected reveal
that consistent differences exist between the secre-tory compartments of the two leptodactylid speciesunder comparison. By contrast, myoepithelial cellsin Leptodactylus chaquensis and Physalaemus albo-notatus exhibit identical subcellular features (caveo-lar invaginations, sarcoplasmic reticulum, and
dense plaques). These traits fit the type II patterndescribed in cutaneous glands of several amphibi-ans, referred to smooth contractile cells providedwith phasic activity (Bani, 1976).
Different traits in the syncytia mostly deal withserous products and post-Golgi (maturational)phases of the secretory cycle, because in both speciessecretory activity involves the same organelle ma-
Fig. 4. Main ultrastructural features in serous glands of Leptodactylus chaquensis: secretory unit (A–C,E,G,H) and myoepithelium(C–E). A: Low-power magnification of serous gland, showing homogeneous features of secretory granules and wide cavities (asterisks)in the cytoplasm. B: These cavities are dilated perigranular halos; notice contiguous nuclei in the secretory unit, with no interveningmembrane. C: Secretory and contractile compartments are separated by slender interstices holding thin neurite endings; notice thatdense-cored synaptic vesicles are distributed on both sides of the interstice, secretory and contractile as well. Arrows point to vesicularprofiles of the sarcoplasmic reticulum. D: Myoepithelial cell cytoskeleton includes dense bodies interspersed among myofilaments.E: Notice elongate profiles of the sarcoplasmic reticulum, oriented along the main axis of this myoepithelial cell (arrows). F: Desmo-some-like junctions occur between myoepithelial cells; the small vesicles close to the myoepithelial cell plasma membranes arepermanent caveolar inpocketings continuous with the interstices (arrow). G: Slender RER complements and Golgi stack; the Golgi isreleasing amorphous, secretory material. H: Early stages of post-Golgi condensation affecting the secretory product. c, caveola; dp,dense plaques in myoepithelial cell; G, Golgi stack; h, perigranular halo; i, interstice between secretory and contractile compartments;RER, rough endoplasmic reticulum; S-SII, stages of serous synthesis and maturation; ss, secretory syncytium.
Fig. 5. Maturational processes affecting the secretory product in serous cutaneous glands of Leptodactylus chaquensis. TEM.A: Higher-power magnification discloses differences between serous deposits, which are mostly related to their densities. B: Peculiarfeature of serous product, consisting of an electron-opaque core inserted in a compartment holding finely dispersed material. C: Thecytoplasm that borders the perigranular halo evaginates, forming thin, microvillous outgrowths. D: These minute cytoplasmicprocesses form a three-dimensional net; arrows point to branching points. G, Golgi stack; SII, SII/III, SIII, stages of serous maturation.
255POISON GLAND ULTRASTRUCTURE IN LEPTODACTYLIDAE

chinery pursuing a common proteosynthetic path-way and their early products share similar features.In both species, the early secretory material con-tained in the rough cisterns shows a weak opacity,but later the Golgi apparatus performs an obviouscondensing activity, because the serous product re-leased from the sacculi of the trans side is moreopaque. Starting from the Golgi step, serous matu-ration processes continue during the intracytoplas-mic storage of the poison deposits (granules), andfollow the differential pathways summarized in Fig-ure 6, in accordance with the centripetal matura-tional gradient characteristic of serous anuranglands (Delfino, 1991). As a result of these matura-
tional processes, the skin poisons acquire the pecu-liar traits of the species observed.
In Pysalaemus albonotatus the dense material re-leased by the Golgi apparatus reaches a thick re-peating substructure through sequential, intermedi-ate features, consisting of alternations of dense andpale areas and labyrinthine patterns. This matura-tional sequence has also been described by Terreniet al. (2002) in hylid and pseudid serous glands,genera Hyla and Scinax, and Pseudis, respectively.Maturational changes detected in P. albonotatus areshort-cuts of the post-Golgi processes occurring intadpoles of the congeneric P. biligonigeros (Delfinoet al., 2001a) and the hylid Scinax nasica (Terreni et
Fig. 6. Schematic, 3D representation (not to scale) of activity in serous glands of Physalaemus albonotatus (A–C), and Leptodac-tylus chaquensis (AI–CI). Based on present findings and data from Delfino et al. (2001). This comparison shows the main arrangementof the secretory units (A and AI), serous maturation (B and BI), and cytoplasm-secretory product relationships (C and CI). S-SV/VI,stages of serous maturation in both species.
256 B.B. ALVAREZ ET AL.

al., 2003). Further maturational steps affectinggranules of P. albonotatus lead to the repeating sub-structures gradually vanishing, as also observed in“ordinary” serous glands of P. biligonigeros (Delfinoet al., 1999b). In the latter species, on the contrary,large serous units have been described (“inguinalglands”; Cei, 1980) where serous maturation in-volves extreme thickening of the repetitive subunitarrangement, until extremely opaque, structurelessgranules are formed. This process allows remark-able amounts of poison to be stored in a condensed(possibly inactive) state (Delfino et al., 1999b). Theserous glands we have described in P. albonotatusbelong to the ordinary type, because, in these spe-cies, inguinal glands are either extremely small orabsent (Lynch, 1970).
When compared with Pysalaemus albonotatusspecies, maturational processes in serous glands ofLeptodactylus chaquensis display a less obvious se-quential development. However, a fundamentaltrend is apparent, roughly along the centripetal gra-dient that leads to the formation of granules pro-vided with a somewhat spongious substructure.Similar “vacuolized” granules have been describedin serous glands of species pertaining to the familiesDiscoglossidae and Ranidae (see Table 1): Discoglos-sus pictus (Delfino, 1979; Delfino et al., 2001b) andD. sardus (Delfino et al., 1993); Rana esculenta(Barni et al., 1987; Delfino et al., 1996), R. iberica(Delfino et al., 1996), R. camerani and R. dalmatina(Delfino, pers. obs.). This peculiar substructureseems to be related to the secretory product matu-ration. Among ultrastructural aspects of gland ac-tivity, which could better describe maturationalchanges, we detected enhanced serous product-syncytial cytoplasm relationships, linked to two gen-eral mechanisms consistent in several anuran spe-cies (Delfino et al., 2001b). These are: 1) evaginationof the compartment (halo) encircling the serous ma-terial into the cytoplasm, and 2) the development ofa network of branched and slender cytoplasmic pro-cesses (resembling minute microvilli) from the syn-cytium. Although both of these devices, which am-plify the surface area of the granule-limitingmembrane, may coexist in the same glands, it ap-pears that evaginations of the halo prevail in gran-ules with ordered substructures, whereas microvil-lous outgrowths predominate in glands producingspongy-like granules (Delfino et al., 2001b). An in-crease in the surface areas involved in exchangebetween cytoplasm and granules may be related toin/outflows of substances: some low molecularweight poison fractions are produced in the secre-tory syncytium cytosol to be later transferred intogranules (Delfino, 1979, 1991). Furthermore, con-densation of the serous product involves transport ofwatery fractions towards the cytoplasmic compart-ment. In dendrobatid frogs serous granules storeproducts from various sites, both endo- and/or exog-enous in origin, including dietary arthropods (Daly,
1998; Smith et al., 2001) and symbiotic microorgan-isms (Daly et al., 1997).
Biochemical data on the composition of anuranskin poisons from a wide taxonomic distribution in-dicate several differences between Physalaemus andLeptodactylus, concerning biogenic amines and pep-tides (Daly et al., 1987). Nonetheless, major classesof active molecules produced by serous glands arewidely distributed among anurans regardless oftheir taxonomic position. Furthermore, ordinary ul-trastructural analysis can only provide informationon the biosynthesis of large molecules, occurring inserous skin products, such as proteins (involvingRER) or steroids (SER), whereas small active mole-cules cannot be detected because they are producedin the cytosol (Delfino, 1991). Despite the sites oforigin of the active poison molecules, it should beremembered that the different maturational pro-cesses observed in P. albonotatus and L. chaquensisare related to changes in the physical state of theproducts, rather than to chemical composition.
The remarkable differences between cutaneouspoisons in the two genera under comparison agreewith the possible non-monophyletic status of thefamily Leptodactylidae (Ford and Cannatella, 1993).At the generic level, it appears that poison biosyn-thesis in Physalaemus albonotatus closely resemblesthat of P. biligonigerus (Delfino et al. 1999b, 2001a;Terreni et al., 2003), which, in turn, support thedata listed in Table 1. Results obtained from Phys-alaemus in the present study are even more relevantbecause we have described identical features in spe-cies pertaining to different groups of this genus: thesmaller biligonigerus tribe and the larger cuvieritribe including albonotatus (Cei, 1980).
LITERATURE CITED
Angel R, Delfino G, Parra GJ. 2003. Ultrastructural patterns ofsecretory activity in poison cutaneous glands of larval andjuvenile Dendrobates auratus (Amphibia, Anura). Toxicon 41:29–39.
Bani G. 1976. Cellule mioepiteliali accluse alle ghiandole cutaneedi alcuni Anfibi. Archo Ital Anat Embriol 81:133–184.
Barni S, Bernocchi G, Bottiroli G. 1987. Histochemistry and mor-phology of the secretory granules of skin venom glands of Ranaesculenta during the active and hibernating period. Arch Biol98:391–406.
Blaylock LA, Ruibal R, Platt-Aloia K. 1976. Skin structure andwiping behaviour of phyllomedusine frogs. Copeia 1976:283–295.
Brizzi R, Corti C, Delfino G, Drewes R. 2004. Ultrastructuralpatterns of secretory activity in serous cutaneous glands of thePacific tree-frog Hyla regilla (Anura, Hylidae). Ital J Zool 71(suppl. 2).
Cei JM. 1980. Amphibians of Argentina. Monit Zool Ital (NuovaSerie), Monografia 2.
Daly JW. 1998. Thirty years of discovering arthropod alkaloids inamphibian skin. J Nat Toxins 61:162–172.
Daly JW, Myers CW, Whittaker N. 1987. Further classification ofskin alkaloids from Neotropical poison frogs (Dendrobatidae),with a general survey of toxic/noxious substances in the Am-phibia. Toxicon 25:1023–1095.
257POISON GLAND ULTRASTRUCTURE IN LEPTODACTYLIDAE

Daly JW, Padgett WL, Saunders RL, Cover JF. 1997. Absence oftetrodotoxins in a captive-raised riparian frog Atelopus varius.Toxicon 35:705–709.
Delfino G. 1977. Il differenziamento delle ghiandole granulosecutanee in larve di Bombina variegata pachypus (Bonaparte)(Anfibio: Anuro: Discoglosside). Ricerca al microscopio ottico eal microscopio elettronico. Archo Ital Anat Embriol 82:337–363.
Delfino G. 1979. Le ghiandole granulose cutanee di Alytes cister-nasii Bosca e Discoglossus pictus Otth (Anfibi, Anuri, Discoglos-sidi): Struttura, ultrastruttura e alcuni dati citochimici. ArchoItal Anat Embriol 84:81–106.
Delfino G. 1991. Ultrastructural aspects of venom secretion inanuran cutaneous glands. In: Tu AT, editor. Handbook of nat-ural toxins, vol. 5. Reptile venoms and toxins. New York: Mar-cel Dekker. p 777–802.
Delfino G, Melis G. 1998. Serous cutaneous glands in the natter-jack Bufo calamita (Anura, Bufonidae): the fundamental role ofaggregation processes during poison maturation. Zoology 101:53–66.
Delfino G, Brizzi R, Borrelli G. 1988. Cutaneous glands in an-urans: differentiation of the secretory syncytium in serous An-lagen. Zool Jahrbuch (Anat) 117:255–275.
Delfino G, Brizzi R, Calloni C. 1990. A morpho-functional char-acterization of the serous cutaneous glands in Bombina orien-talis (Anura:Discoglossidae). Zool Anz 225:295–310.
Delfino G, Brizzi R, De Santis R, Melosi M. 1992. Serous cutane-ous glands of the western spade-foot Pelobates cultripes (Am-phibia: Anura): an ultrastructural study on adults and juve-niles. Archo Ital Anat Embriol 97:109–120.
Delfino G, Brizzi R, Jantra S, Streitberger M. 1993. Cutaneousvenom glands in the Tyrrhenian painted frog Discoglossus sar-dus (Tschudi), ontogenetic evolution of the biosynthesis appa-ratus. Acta Biol Benrodis 5:129–139.
Delfino G, Brizzi R, Calloni C. 1994. Serous cutaneous glands inthe tree-frog Hyla arborea (L.): origin, ontogenetic evolution,and possible functional implications of the secretory granulesubstructure. Acta Zool (Stockh) 75:27–36.
Delfino G, Brizzi R, Jantra S, Feri L. 1995a. Post-Golgian matu-rative processes during the biosynthesis of poison secretion incutaneous glands of the European common toad Bufo bufo. JNat Toxins 4:97–113.
Delfino G, Brizzi R, Feri L. 1995b. Chemical skin defence in Bufobufo: an ultrastructural study during ontogenesis. Zool Anz234:101–111.
Delfino G, Brizzi R, Melis G. 1996. Merocrine secretion fromserous cutaneous glands in Rana esculenta complex and Ranaiberica. Alytes 13:179–192.
Delfino G, Brizzi R, Alvarez BB, Kracke-Berndorff R. 1998a.Serous cutaneous glands in Phyllomedusa hypochondrialis(Anura, Hylidae): secretory patterns during ontogenesis. TissueCell 30:30–40.
Delfino G, Alvarez BB, Brizzi R, Cespedez JA. 1998b. Serouscutaneous glands of Argentine Phyllomedusa Wagler 1830(Anura Hylidae): secretory polymorphism and adaptive plastic-ity. Trop Zool 11:333–351.
Delfino G, Brizzi R, Alvarez BB, Taddei L. 1999a. Secretorypolymorphism and serous cutaneous glands heterogeneity inBufo granulosus (Amphibia, Anura). Toxicon 37:1281–1296.
Delfino G, Brizzi R, Alvarez BB, Gentili M. 1999b. Granularcutaneous glands in the frog Physalaemus biligonigerus(Anura, Leptodactylidae): comparison between ordinary serousand “inguinal” glands. Tissue Cell 31:576–586.
Delfino G, Nosi D, Brizzi R, Alvarez BB. 2001a. Serous cutaneousglands in the paludiculine frog Physalaemus biligonigerus(Anura, Leptodactylidae): patterns of cytodifferentiation andsecretory activity in prematomorphic specimens. Acta Zool(Stockh) 82:149–158.
Delfino G, Nosi D, Giachi F. 2001b. Secretory granule-cytoplasmrelationships in serous glands of anurans: ultrastructural evi-dence and possible functional role. Toxicon 39:1161–1171.
Ford LS, Cannatella DC. 1993. The major clades of frogs. Herpe-tol Monogr 7:94–117.
Grosse WR, Linnenbach M. 1989. Feinbau der Hautdrusen undEntwicklung des Melanophoren-Musters bei Laubfrosch, Hylaarborea L. (Amphibia, Anura). Zool Anz 223:211–219.
Karnovsky MJ. 1965. A formaldehyde-glutaraldehyde fixative ofhigh osmolarity for use in electron microscopy. J Cell Biol27:137A.
Lynch JD. 1970. Systematic status of the American leptodactylidfrog genera Engystomops, Eupemphix, and Physalaemus. Co-peia 1970:488–496.
Neuwirth M, Daly JW, Myers CW, Tice LW. 1979. Morphology ofthe granular secretory glands in skin of poison-dart frogs (Den-drobatidae). Tissue Cell 11:755–771.
Quay WB. 1972. Integument and environment: glandular compo-sition, function, evolution. Am Zool 12:95–108.
Smith BP, Tyler MJ, Kanekko T, Garraffio M, Spande TF, DalyJW. 2001. Evidence for biosynthesis of pseudorhynamine alka-loids by an Australian myobathrachid frog (Pseudophryne) andfor sequestration of dietary pumiliotoxins. J Nat Toxins 65:439–447.
Terreni A, Nosi D, Delfino G. 2002. Cutaneous serous glands inSouth-American anurans, an ultrastructural comparison be-tween hylid and pseudid species. Ital J Zool 69:115–123.
Terreni A, Nosi D, Greven H, Delfino G. 2003. Development ofserous cutaneous glands in Scinax nasica (Anura, Hylidae):patterns of poison biosynthesis and maturation in comparisonwith larval glands in specimens of other families. Tissue Cell35:274–87.
Toledo RC, Jared C. 1995. Cutaneous granular glands and am-phibian venoms. Comp Biochem Physiol 111A:11–29.
258 B.B. ALVAREZ ET AL.




















