
Structure and Ultrastructure of Leaf and Calyx Glands in Galphimia brasiliensis (Malpighiaceae
10
1935 American Journal of Botany 88(11): 1935–1944. 2001. STRUCTURE AND ULTRASTRUCTURE OF LEAF AND CALYX GLANDS IN GALPHIMIA BRASILIENSIS (MALPIGHIACEAE) 1 MARI ´ A A. CASTRO, 2,4 ANDREA S. VEGA, 3 AND MARI ´ A E. MU ´ LGURA 3 2 Laboratorio de Anatomı ´a Vegetal, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, 1428 Buenos Aires, Argentina; and 3 Instituto de Bota ´nica Darwinion, Labarde ´n 200, C.C. 22, 1642 Buenos Aires Argentina The present study describes the anatomical structure of calyx and leaf glands in Galphimia brasiliensis and analyzes the mechanism of secretion. The glands are marginal and suprabasal, cup-shaped, sessile, and scarcely visible with the naked eye. Light microscopy reveals the following features: a thin, smooth cuticle; unistratified secretory cells; subglandular parenchyma; and vascular bundle supply composed of phloem and xylem with abundant druses of calcium oxalate. Transmission electron microscopy reveals the presence of secretory cells with conspicuous nuclei, dense cytoplasm, lipid droplets, numerous vesicles, mitochondria, Golgi, rough endoplasmic reticulum (RER), and elongated plastids with osmiophilic contents. The secretion reaches the apoplastic space and accumulates beneath the cuticle. Finally, the viscous, translucent exudate is eliminated by mechanical rupture of the cuticle. Histochemical analysis confirms that lipids are the main constituent. Small amounts of poly- saccharides were also identified. Key words: anatomy; calyx glands; Galphimia brasiliensis; leaf glands; Malpighiaceae. The family Malpighiaceae includes ;66 genera and 1200 species and is distributed in tropical and subtropical regions of both hemispheres (Anderson, 1990). Nearly 950 species are endemic to the New World, northern South America being the major center of diversity (Anderson, 1979).The Malpighiaceae show an important variation in habitat preferences, types of fruits (Anderson, 1979), pollen morphology (Lobreau-Callen, 1984), and chromosome numbers (Foue ¨t, 1966), although flowers are very similar in morphology (‘‘floral conserva- tism’’; see Anderson, 1979) and in different aspects of their floral biology. The presence of calyx glands disposed in pairs constitutes a derived character of the Malpighiaceae (Anderson, 1990) and represents a character with diagnostic value in this family. Generally, the New World species have abaxial glands on the sepals that contain lipids (Vogel, 1974; Anderson, 1990) whose function is to attract pollinators, while leaf blades have two extrafloral nectaries in their lower third. Several authors (e.g., Hauman-Merck, 1912; Raw, 1979; Steiner, 1985; Sazima and Sazima, 1989; Vogel, 1990) have mentioned a close as- sociation between the Malpighiaceae and bees of Anthophor- idae subfamily, which collect lipids or pollen and lipids (Raw, 1979) with specialized organs. The structure and position of the oil-collecting organs are correlated with the types of oil glands used by different bees (Neff and Simpson, 1981). In the Malpighiaceae, bees of the tribe Centridini are intimately associated with plants that have glands with a layer of active secretory epithelial cells (Neff and Simpson, 1981). Hauman-Merck (1912) mentioned the presence of nectar in calyx glands of Stigmaphyllon Adr. Juss., but according to Lobreau-Callen (1989), while in Malpighiaceae species in the 1 Manuscript received 12 December 2000; revision accepted 3 April 2001. The authors thank Dr. William R. Anderson for his critical review of the manuscript and valuable suggestions and Graphic Designer Toma ´s E. Aversa. This work was supported by Consejo Nacional de Investigaciones Cientı ´ficas y Te ´cnicas (CONICET, PIP 0800/98). 4 Author for reprint requests (e-mail: [email protected]). New World the calyx glands produce lipids and traces of car- bohydrates, it is the opposite in Old World species. Vogel (1990) uses this argument to support his idea about the trans- formation of the secretions in calyx glands from nectar to lip- ids. The genus Galphimia Cav. belongs to the tribe Galphimieae of the subfamily Byrsonimoideae within the Malpighiaceae (Anderson, 1977). Galphimia includes approximately ten spe- cies in tropical and subtropical America, with a disjunct dis- tribution (Niedenzu, 1928). Most of the species grow in Cen- tral America; only one, Galphimia brasiliensis (L.) Adr. Juss., grows in South America (northeastern Argentina, southern Brazil, Bolivia, Paraguay, and Uruguay). In the genus Gal- phimia, calyx glands are greatly reduced or absent (Anderson, 1977), and G. brasiliensis has been considered to be in the group of species lacking them (Niedenzu, 1928; Lobreau-Cal- len, 1989). However, we observed that calyx glands are pre- sent in variable sizes, numbers, and positions within each flow- er. The objective of this research was to analyze the structural and ultrastructural anatomy and to identify the chemical com- position of the leaf and calyx glands in Galphimia brasiliensis in order to verify the homology of both structures. MATERIALS AND METHODS Plant materials—Both herbarium specimens and fresh material of Gal- phimia brasiliensis (L.) Adr. Juss. were studied, and voucher specimens were deposited in SI Herbarium (Instituto de Bota ´nica Darwinion). (See Table 1.) Anatomical studies—Leaves and flowers of G. brasiliensis were fixed in FAA (formalin : ethanol : acetic acid) and then stored in 70% ethanol. The material was dehydrated in an ethanol-alcohol series and embedded in paraffin after infiltration in a vacuum oven. Transverse and longitudinal sections 10 mm thick were cut on a rotary microtome and were double-stained with Saf- ranin-Fast Green (D’Ambrogio de Argu ¨eso, 1986). They were examined un- der a ZEISS (Zeiss, Oberkochen, Germany) Phomi III microscope with and without polarized light. Histochemical tests for light microscopy were used to detect the possible
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Structure and Ultrastructure of Leaf and Calyx Glands in Galphimia brasiliensis (Malpighiaceae

1935
American Journal of Botany 88(11): 1935–1944. 2001.
STRUCTURE AND ULTRASTRUCTURE OF LEAF AND
CALYX GLANDS IN GALPHIMIA BRASILIENSIS
(MALPIGHIACEAE)1
MARIA A. CASTRO,2,4 ANDREA S. VEGA,3 AND MARIA E. MULGURA3
2Laboratorio de Anatomıa Vegetal, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires,1428 Buenos Aires, Argentina;
and 3Instituto de Botanica Darwinion, Labarden 200, C.C. 22, 1642 Buenos Aires Argentina
The present study describes the anatomical structure of calyx and leaf glands in Galphimia brasiliensis and analyzes themechanism of secretion. The glands are marginal and suprabasal, cup-shaped, sessile, and scarcely visible with the naked eye.Light microscopy reveals the following features: a thin, smooth cuticle; unistratified secretory cells; subglandular parenchyma;and vascular bundle supply composed of phloem and xylem with abundant druses of calcium oxalate. Transmission electronmicroscopy reveals the presence of secretory cells with conspicuous nuclei, dense cytoplasm, lipid droplets, numerous vesicles,mitochondria, Golgi, rough endoplasmic reticulum (RER), and elongated plastids with osmiophilic contents. The secretionreaches the apoplastic space and accumulates beneath the cuticle. Finally, the viscous, translucent exudate is eliminated bymechanical rupture of the cuticle. Histochemical analysis confirms that lipids are the main constituent. Small amounts of poly-saccharides were also identified.
Key words: anatomy; calyx glands; Galphimia brasiliensis; leaf glands; Malpighiaceae.
The family Malpighiaceae includes ;66 genera and 1200species and is distributed in tropical and subtropical regionsof both hemispheres (Anderson, 1990). Nearly 950 species areendemic to the New World, northern South America being themajor center of diversity (Anderson, 1979).The Malpighiaceaeshow an important variation in habitat preferences, types offruits (Anderson, 1979), pollen morphology (Lobreau-Callen,1984), and chromosome numbers (Fouet, 1966), althoughflowers are very similar in morphology (‘‘floral conserva-tism’’; see Anderson, 1979) and in different aspects of theirfloral biology.
The presence of calyx glands disposed in pairs constitutesa derived character of the Malpighiaceae (Anderson, 1990) andrepresents a character with diagnostic value in this family.Generally, the New World species have abaxial glands on thesepals that contain lipids (Vogel, 1974; Anderson, 1990)whose function is to attract pollinators, while leaf blades havetwo extrafloral nectaries in their lower third. Several authors(e.g., Hauman-Merck, 1912; Raw, 1979; Steiner, 1985; Sazimaand Sazima, 1989; Vogel, 1990) have mentioned a close as-sociation between the Malpighiaceae and bees of Anthophor-idae subfamily, which collect lipids or pollen and lipids (Raw,1979) with specialized organs. The structure and position ofthe oil-collecting organs are correlated with the types of oilglands used by different bees (Neff and Simpson, 1981). Inthe Malpighiaceae, bees of the tribe Centridini are intimatelyassociated with plants that have glands with a layer of activesecretory epithelial cells (Neff and Simpson, 1981).
Hauman-Merck (1912) mentioned the presence of nectar incalyx glands of Stigmaphyllon Adr. Juss., but according toLobreau-Callen (1989), while in Malpighiaceae species in the
1 Manuscript received 12 December 2000; revision accepted 3 April 2001.The authors thank Dr. William R. Anderson for his critical review of the
manuscript and valuable suggestions and Graphic Designer Tomas E. Aversa.This work was supported by Consejo Nacional de Investigaciones Cientıficasy Tecnicas (CONICET, PIP 0800/98).
4 Author for reprint requests (e-mail: [email protected]).
New World the calyx glands produce lipids and traces of car-bohydrates, it is the opposite in Old World species. Vogel(1990) uses this argument to support his idea about the trans-formation of the secretions in calyx glands from nectar to lip-ids.
The genus Galphimia Cav. belongs to the tribe Galphimieaeof the subfamily Byrsonimoideae within the Malpighiaceae(Anderson, 1977). Galphimia includes approximately ten spe-cies in tropical and subtropical America, with a disjunct dis-tribution (Niedenzu, 1928). Most of the species grow in Cen-tral America; only one, Galphimia brasiliensis (L.) Adr. Juss.,grows in South America (northeastern Argentina, southernBrazil, Bolivia, Paraguay, and Uruguay). In the genus Gal-phimia, calyx glands are greatly reduced or absent (Anderson,1977), and G. brasiliensis has been considered to be in thegroup of species lacking them (Niedenzu, 1928; Lobreau-Cal-len, 1989). However, we observed that calyx glands are pre-sent in variable sizes, numbers, and positions within each flow-er.
The objective of this research was to analyze the structuraland ultrastructural anatomy and to identify the chemical com-position of the leaf and calyx glands in Galphimia brasiliensisin order to verify the homology of both structures.
MATERIALS AND METHODS
Plant materials—Both herbarium specimens and fresh material of Gal-phimia brasiliensis (L.) Adr. Juss. were studied, and voucher specimens weredeposited in SI Herbarium (Instituto de Botanica Darwinion). (See Table 1.)
Anatomical studies—Leaves and flowers of G. brasiliensis were fixed inFAA (formalin : ethanol : acetic acid) and then stored in 70% ethanol. Thematerial was dehydrated in an ethanol-alcohol series and embedded in paraffinafter infiltration in a vacuum oven. Transverse and longitudinal sections 10mm thick were cut on a rotary microtome and were double-stained with Saf-ranin-Fast Green (D’Ambrogio de Argueso, 1986). They were examined un-der a ZEISS (Zeiss, Oberkochen, Germany) Phomi III microscope with andwithout polarized light.
Histochemical tests for light microscopy were used to detect the possible
1936 [Vol. 88AMERICAN JOURNAL OF BOTANY
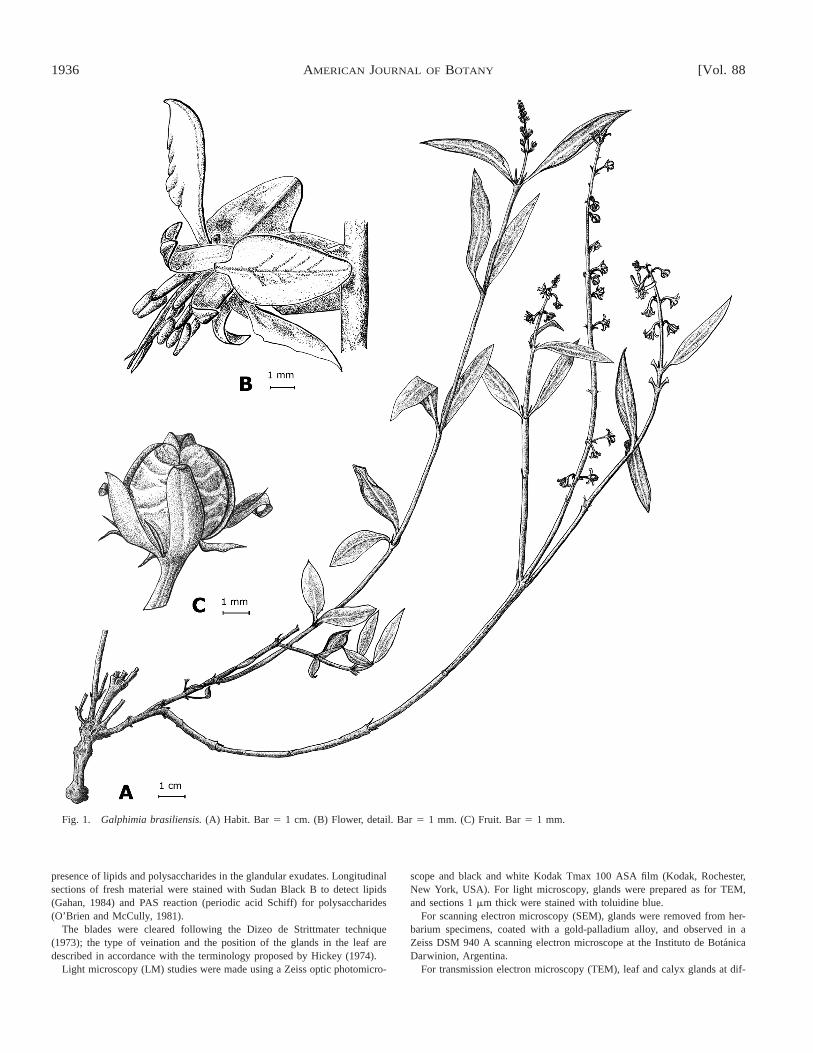
Fig. 1. Galphimia brasiliensis. (A) Habit. Bar 5 1 cm. (B) Flower, detail. Bar 5 1 mm. (C) Fruit. Bar 5 1 mm.
presence of lipids and polysaccharides in the glandular exudates. Longitudinalsections of fresh material were stained with Sudan Black B to detect lipids(Gahan, 1984) and PAS reaction (periodic acid Schiff) for polysaccharides(O’Brien and McCully, 1981).
The blades were cleared following the Dizeo de Strittmater technique(1973); the type of veination and the position of the glands in the leaf aredescribed in accordance with the terminology proposed by Hickey (1974).
Light microscopy (LM) studies were made using a Zeiss optic photomicro-
scope and black and white Kodak Tmax 100 ASA film (Kodak, Rochester,New York, USA). For light microscopy, glands were prepared as for TEM,and sections 1 mm thick were stained with toluidine blue.
For scanning electron microscopy (SEM), glands were removed from her-barium specimens, coated with a gold-palladium alloy, and observed in aZeiss DSM 940 A scanning electron microscope at the Instituto de BotanicaDarwinion, Argentina.
For transmission electron microscopy (TEM), leaf and calyx glands at dif-
November 2001] 1937CASTRO ET AL.—LEAF AND CALYX GLANDS IN GALPHIMIA BRASILIENSIS
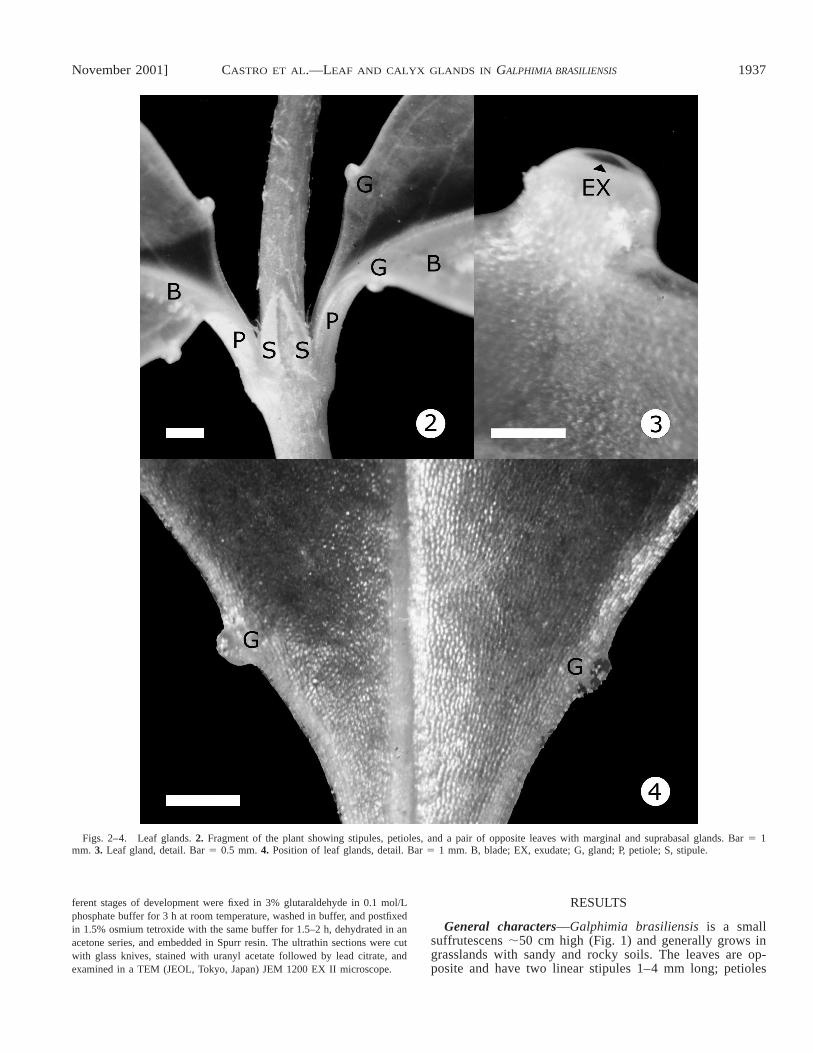
Figs. 2–4. Leaf glands. 2. Fragment of the plant showing stipules, petioles, and a pair of opposite leaves with marginal and suprabasal glands. Bar 5 1mm. 3. Leaf gland, detail. Bar 5 0.5 mm. 4. Position of leaf glands, detail. Bar 5 1 mm. B, blade; EX, exudate; G, gland; P, petiole; S, stipule.
ferent stages of development were fixed in 3% glutaraldehyde in 0.1 mol/Lphosphate buffer for 3 h at room temperature, washed in buffer, and postfixedin 1.5% osmium tetroxide with the same buffer for 1.5–2 h, dehydrated in anacetone series, and embedded in Spurr resin. The ultrathin sections were cutwith glass knives, stained with uranyl acetate followed by lead citrate, andexamined in a TEM (JEOL, Tokyo, Japan) JEM 1200 EX II microscope.
RESULTS
General characters—Galphimia brasiliensis is a smallsuffrutescens ;50 cm high (Fig. 1) and generally grows ingrasslands with sandy and rocky soils. The leaves are op-posite and have two linear stipules 1–4 mm long; petioles
1938 [Vol. 88AMERICAN JOURNAL OF BOTANY
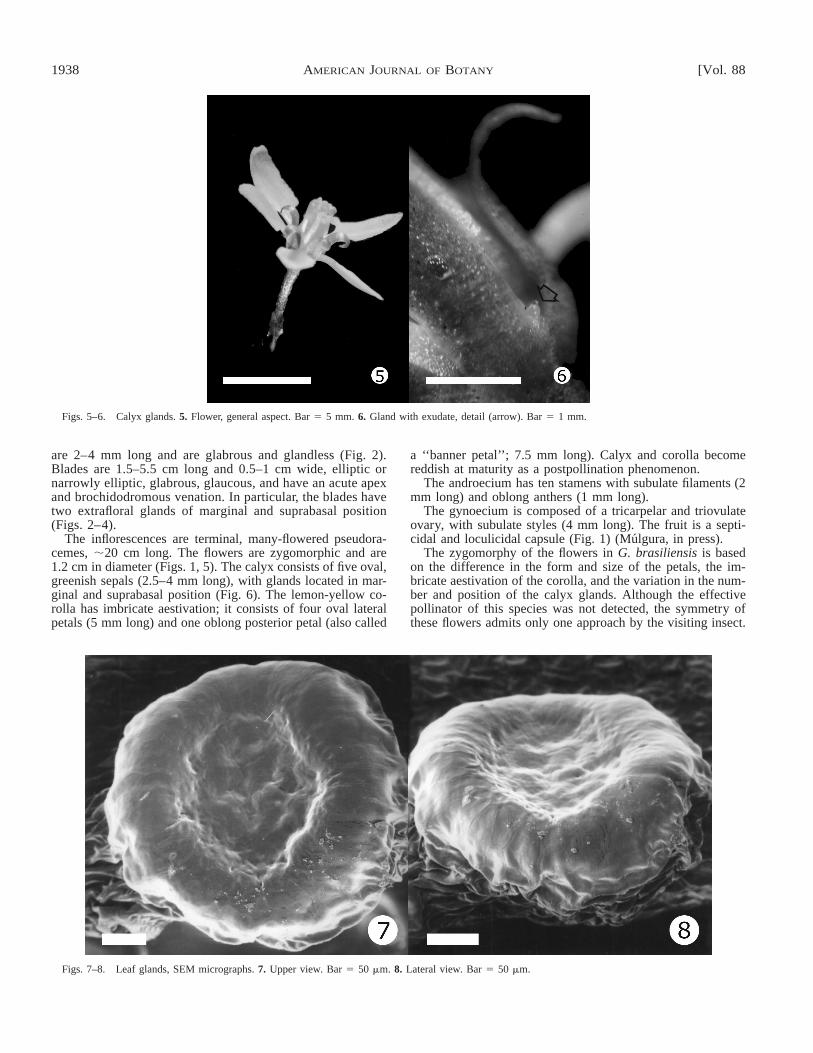
Figs. 5–6. Calyx glands. 5. Flower, general aspect. Bar 5 5 mm. 6. Gland with exudate, detail (arrow). Bar 5 1 mm.
Figs. 7–8. Leaf glands, SEM micrographs. 7. Upper view. Bar 5 50 mm. 8. Lateral view. Bar 5 50 mm.
are 2–4 mm long and are glabrous and glandless (Fig. 2).Blades are 1.5–5.5 cm long and 0.5–1 cm wide, elliptic ornarrowly elliptic, glabrous, glaucous, and have an acute apexand brochidodromous venation. In particular, the blades havetwo extrafloral glands of marginal and suprabasal position(Figs. 2–4).
The inflorescences are terminal, many-flowered pseudora-cemes, ;20 cm long. The flowers are zygomorphic and are1.2 cm in diameter (Figs. 1, 5). The calyx consists of five oval,greenish sepals (2.5–4 mm long), with glands located in mar-ginal and suprabasal position (Fig. 6). The lemon-yellow co-rolla has imbricate aestivation; it consists of four oval lateralpetals (5 mm long) and one oblong posterior petal (also called
a ‘‘banner petal’’; 7.5 mm long). Calyx and corolla becomereddish at maturity as a postpollination phenomenon.
The androecium has ten stamens with subulate filaments (2mm long) and oblong anthers (1 mm long).
The gynoecium is composed of a tricarpelar and triovulateovary, with subulate styles (4 mm long). The fruit is a septi-cidal and loculicidal capsule (Fig. 1) (Mulgura, in press).
The zygomorphy of the flowers in G. brasiliensis is basedon the difference in the form and size of the petals, the im-bricate aestivation of the corolla, and the variation in the num-ber and position of the calyx glands. Although the effectivepollinator of this species was not detected, the symmetry ofthese flowers admits only one approach by the visiting insect.
November 2001] 1939CASTRO ET AL.—LEAF AND CALYX GLANDS IN GALPHIMIA BRASILIENSIS
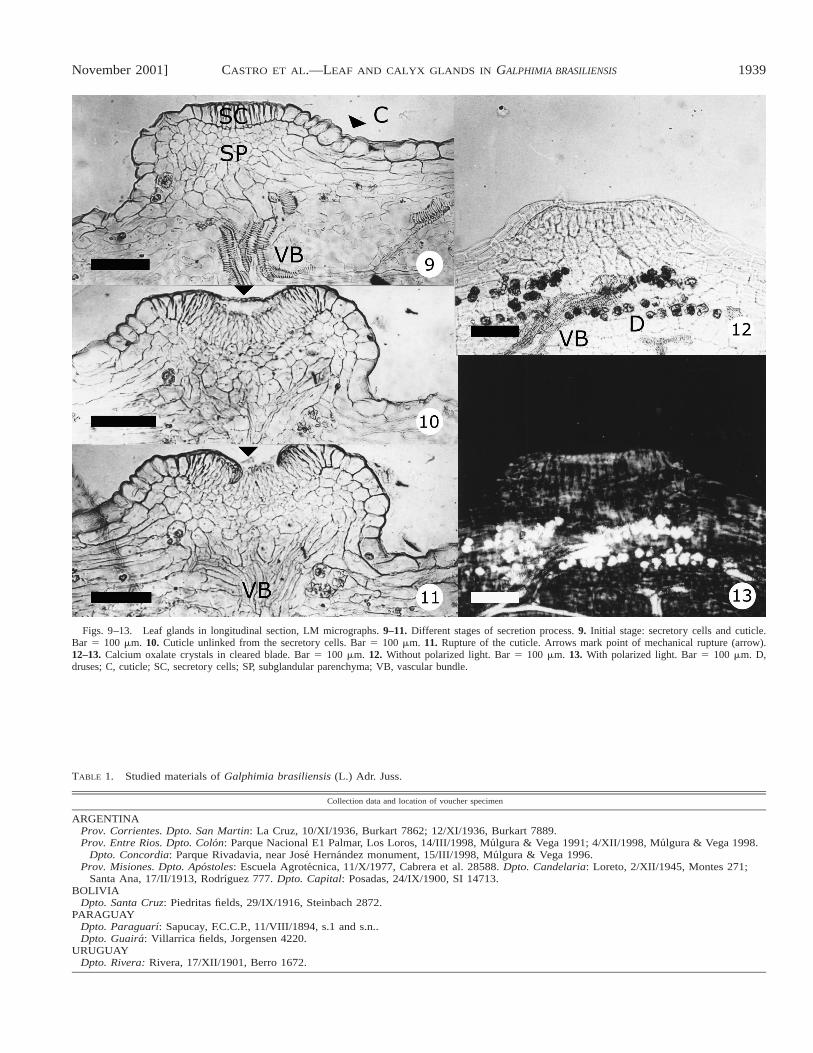
Figs. 9–13. Leaf glands in longitudinal section, LM micrographs. 9–11. Different stages of secretion process. 9. Initial stage: secretory cells and cuticle.Bar 5 100 mm. 10. Cuticle unlinked from the secretory cells. Bar 5 100 mm. 11. Rupture of the cuticle. Arrows mark point of mechanical rupture (arrow).12–13. Calcium oxalate crystals in cleared blade. Bar 5 100 mm. 12. Without polarized light. Bar 5 100 mm. 13. With polarized light. Bar 5 100 mm. D,druses; C, cuticle; SC, secretory cells; SP, subglandular parenchyma; VB, vascular bundle.
TABLE 1. Studied materials of Galphimia brasiliensis (L.) Adr. Juss.
Collection data and location of voucher specimen
ARGENTINAProv. Corrientes. Dpto. San Martin: La Cruz, 10/XI/1936, Burkart 7862; 12/XI/1936, Burkart 7889.Prov. Entre Rios. Dpto. Colon: Parque Nacional E1 Palmar, Los Loros, 14/III/1998, Mulgura & Vega 1991; 4/XII/1998, Mulgura & Vega 1998.
Dpto. Concordia: Parque Rivadavia, near Jose Hernandez monument, 15/III/1998, Mulgura & Vega 1996.Prov. Misiones. Dpto. Apostoles: Escuela Agrotecnica, 11/X/1977, Cabrera et al. 28588. Dpto. Candelaria: Loreto, 2/XII/1945, Montes 271;
Santa Ana, 17/II/1913, Rodrıguez 777. Dpto. Capital: Posadas, 24/IX/1900, SI 14713.BOLIVIA
Dpto. Santa Cruz: Piedritas fields, 29/IX/1916, Steinbach 2872.PARAGUAY
Dpto. Paraguarı: Sapucay, F.C.C.P., 11/VIII/1894, s.1 and s.n..Dpto. Guaira: Villarrica fields, Jorgensen 4220.
URUGUAYDpto. Rivera: Rivera, 17/XII/1901, Berro 1672.

1940 [Vol. 88AMERICAN JOURNAL OF BOTANY
Figs. 14–15. Active leaf glands, sections 1 mm thick, LM micrographs. 14. Initial stage: smooth cuticle, secretory cells, subglandular parenchyma. Bar 550 mm. 15. Exudate beneath the cuticle. Bar 5 50 mm. C, cuticle; EX, exudate; SC, secretory cells; SP, subglandular parenchyma.
Figs. 16–17. Active calyx glands in longitudinal sections, LM micrographs. 16. Initial stage. Bar 5 100 mm. 17. Secretory stage. Bar 5 50 mm. C, cuticle;SC, secretory cells; SP, subglandular parenchyma; VS, vascular bundles.
→
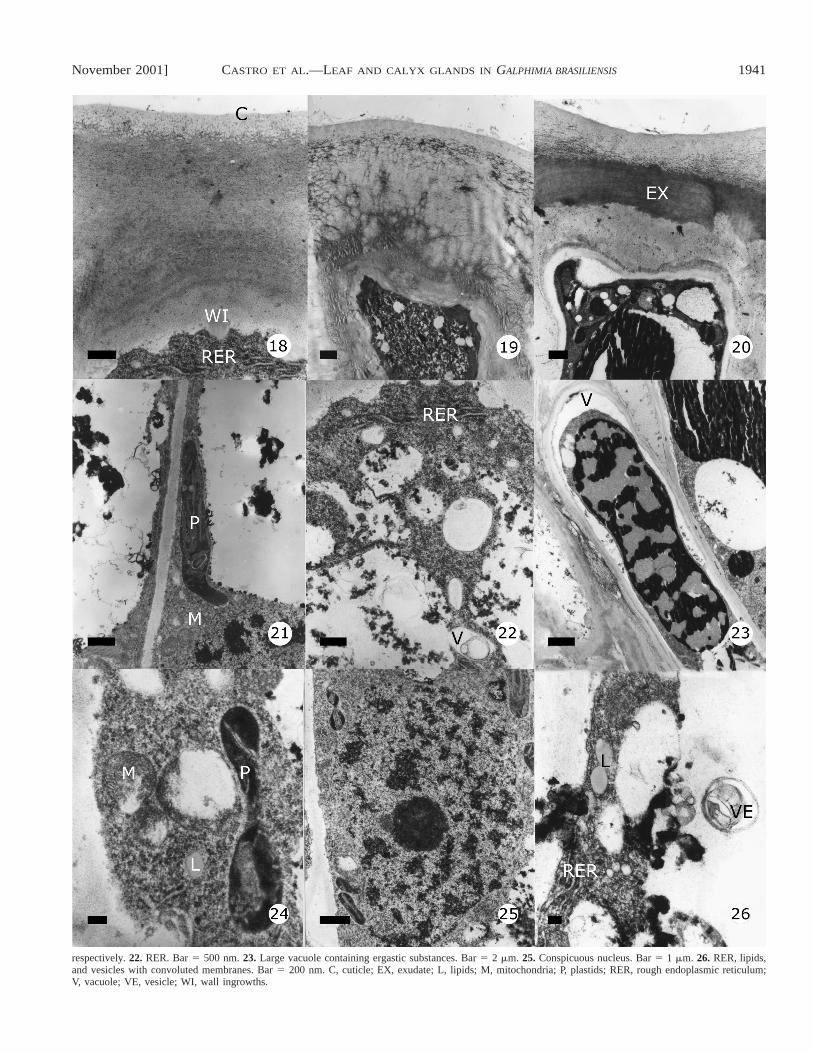
Figs. 18–26. Secretory cells in active leaf glands, TEM micrographs. 18. Outer tangential cell wall and slight ingrowth. Bar 5 500 nm. 19–20. Exudatecrossing the wall: note, in Fig. 20 the exudate is reaching the subcuticular space. Bars 5 1 mm. 21, 24. Plastids and mitochondria. Bars 5 1 mm and 200 nm,
Morphology and anatomy of glands—Leaf and calyxglands exhibit similar structural and ultrastructural organiza-tion. Glands are 0.2–0.3 mm high and 0.35–0.4 mm broad,sessile, cup-shaped, with a central slight concavity (Figs. 3,6–8) are scarcely visible with the naked eye except for theslightly pedicellate leaf and calyx glands observed in the spec-imen Burkart 7889 (SI).
Light microscopy of longitudinal sections of glands showsthese features, from the outside in: cuticle, secretory tissue, acentral core of subglandular parenchyma cells, and vascularsupply bundles (Figs. 9–11, 14–17). The cuticle is smooth andthin (Figs. 7, 8, 14, 16). The unistratified secretory tissue iscomposed of tightly packed elongated cells, which form a pal-isade layer involved in the synthesis and secretion of the ex-
November 2001] 1941CASTRO ET AL.—LEAF AND CALYX GLANDS IN GALPHIMIA BRASILIENSIS
respectively. 22. RER. Bar 5 500 nm. 23. Large vacuole containing ergastic substances. Bar 5 2 mm. 25. Conspicuous nucleus. Bar 5 1 mm. 26. RER, lipids,and vesicles with convoluted membranes. Bar 5 200 nm. C, cuticle; EX, exudate; L, lipids; M, mitochondria; P, plastids; RER, rough endoplasmic reticulum;V, vacuole; VE, vesicle; WI, wall ingrowths.

1942 [Vol. 88AMERICAN JOURNAL OF BOTANY
Figs. 27–32. Secretory cells in active calyx glands, TEM micrographs. 27. Exudate crossing the wall (arrow). Bar 5 500 nm. 28. Cytoplasm with mito-chondria, wall and exudate in the apoplastic space. Bar 5 500 nm. 29. Mitochondria and plastids. Note drops of exudate in the apoplastic space (arrow). Bar5 200 nm. 30. Mitochondria, plastids, and Golgi. Bar 5 500 nm. 31. Conspicuous nuclei and dense cytoplasm. Bar 5 1 mm. 32. Radial cell wall between twoconsecutive secretory cells and unusually shaped plastids. Bar 5 500 nm. C, cytoplasm; EX, exudate; G, Golgi; M, mitochondria; N, nuclei; P, plastid; V,vacuole.
udate. The secretory cells are high and narrow (45.5–71.5 mmlong and 7.8–13 mm wide), thin-walled (Fig. 9), and charac-terized by a densely staining cytoplasm and a relatively largenucleus (Figs. 14, 17). The subglandular parenchyma consistsof six to seven layers of isodiametric cells with reduced inter-cellular spaces. Vascular supply of xylem and phloem reachesthe periphery of the subglandular tissue (Figs. 16, 17). Highconcentrations of calcium oxalate druses (Figs. 12, 13) and afew cubic crystals were observed in this area. The latter arenot birefringent in polarized light.
Transmission electron microscopy observations reveal thatthe dense appearance of the cytoplasm results from an abun-dance of ribosomes, rough endoplasmic reticulum (RER), mi-tochondria, Golgi, numerous small and translucent vesicles,plastids, and a large vacuole containing ergastic substances(Figs. 21, 25, 30, 31). The RER is composed of narrow cis-ternae dispersed in cytoplasm or closely stacked near the plas-ma membrane (Figs. 18, 22, 26). Mitochondria are elliptic orglobular, occur in great numbers and possess many well-de-veloped cristae. They are dispersed or aggregated in the cy-
toplasm (Figs. 21, 28–30). Golgi are particularly present inactive secretory cells and are abundant in the presecretorystage. Numerous vesicles containing convoluted membranescan be seen and probably represent deposits of membrane ma-terial synthesized by the active secretory cells. Vesicles fusedwith the tonoplast are observed during the synthesis of thesecretory products (Figs. 22, 26). Plastids are frequent, en-larged, and unusually shaped, with starch and dense zones(Figs. 21, 24, 29, 30, 32). Large vacuoles containing denseergastic substances mixed with other more translucent prod-ucts were observed (Fig. 23). Some lipid droplets are presentin the cytoplasm (Figs. 24, 26). The inner side of the outerpericlinal secretory cell wall presents slight ingrowths asso-ciated with plasma membrane invagination (Fig. 18).
In the active stage and with the onset of secretion, the ex-udate released into the apoplastic space flows toward the walland begins to accumulate beneath the cuticle (Figs. 18–20, 27–29). The cuticle is smooth; neither pores nor pore-like struc-tures were observed. Its mechanical rupture results from pres-

November 2001] 1943CASTRO ET AL.—LEAF AND CALYX GLANDS IN GALPHIMIA BRASILIENSIS
sure of the accumulated exudate or the action of insect visitors(Figs. 15, 19, 20, 27).
The exudate of both glands is translucent and slightly vis-cous, composed mainly of lipids with small amounts of sugars(Figs. 3, 6). Histochemical tests with Sudan Black B confirmthe presence of abundant lipids through an intensive dark bluestain. The PAS reaction corroborates the presence of traces ofpolysaccharides.
DISCUSSION
The present study was undertaken to elucidate the structuraland ultrastructural features related to the anatomy and secre-tion mechanism of calyx and leaf glands in Galphimia brasi-liensis.
Flowers of G. brasiliensis have been considered actinomor-phic by Lobreau-Callen (1989); however, the imbricate aesti-vation of the corolla and the variation in number and dispo-sition of calyx glands are characters that determine the flower’szygomorphy. The number, size, and position of the calyxglands are variable among flowers of the same inflorescenceand among inflorescences of the same plant. Although thisspecies has been included in the group with eglandular flowers(Niedenzu, 1928; Lobreau-Callen, 1989), there may be two tozero calyx glands on each sepal. Vogel (1990) has interpretedthe presence of single glands as the result of fusion of a pairof glands; however, our observation of some flowers with pairsof glands in which one of the glands was notably reducedsuggests that single glands result from loss, not fusion.
Glands on leaves and sepals exhibit similar structural andultrastructural organization. Light microscopy study of longi-tudinal sections reveals these features from the outside in: cu-ticle, secretory tissue, a central core of subglandular paren-chyma cells, and vascular supply bundles. This anatomical pat-tern of organization is the same as that observed by Cocucci,Holgado, and Anton, (1996) in Dinemandra ericoides Adr.Juss., but in this species the secretory cells are clavate shapedand unlinked for more than 90% of their length. In G. brasi-liensis, the secretory cells are elongated, tightly packed, andlinked throughout their length.
The TEM observations showed that several ultrastructuralchanges occur with the onset of secretion. Active secretorycells exhibit a conspicuous nucleus, dense cytoplasm, lipiddroplets, numerous vesicles, mitochondria, Golgi, RER, andelongated plastids with osmiophilic contents. The secretionreaches the apoplastic space and accumulates beneath the cu-ticle. Finally, in both types of glands, the scarce translucentexudate is eliminated by mechanical rupture of the cuticle.
Due to the reduced size of glands and the scarce exudate,only histochemical tests were made. Presence of lipids wasdetected with Sudan Black B. These results do not support theconclusions provided by several authors (Lobreau-Callen,1989; Anderson, 1990; Vogel, 1990) who agree that in Mal-pighiaceae leaf glands are nectaries that produce sugars butnot oils.
Calyx glands in Old World species are reported to be nec-taries while these in New World species are lipophilic glands(Faegri and van der Pijl, 1979). Baker (1978) analyzed thelipophilic secretion of Malpighiaceae and found small amountsof sugars and amino acids. Vinson et al. (1997) analyzed thefloral chemistry of Byrsonima crassifolia (L.) H.B.K. andfound two types of floral oils: the most common type con-tained a trace of carbohydrate; the other type had large
amounts of an unknown lipid that was more polar than thestandards.
In both calyx and leaf glands of G. brasiliensis, the presenceof abundant lipids and small amounts of sugars probably in-dicates that the lipophilic glands derive from nectaries. Vogel(1990) suggested that the occurrence in some nectars of smallamounts of lipids and sugar traces in some floral oils supportsthe view that these change were feasible.
Finally, the present study confirms that leaf and calyx glandsare homologous structures. According to W. and Ch. Anderson(University of Michigan, personal communication) it seemsquite possible that the glands on the sepals are simply a con-tinuation of the expression of the genes from the leaves, andthat the real function of the glands, assuming there is one, hasto do with their role on the leaves. In Galphimia the calyxglands represent an independent evolution from leaf glands,providing a model that may explain the origin of the largecalyx glands found in many other Malpighiaceae from theglands on the abaxial surface of the leaves in ancestral plants.
LITERATURE CITED
ANDERSON, W. R. 1977. Byrsonimoideae, a new subfamily of the Malpigh-iaceae. Leandra 7: 5–18.
ANDERSON, W. R. 1979. Floral conservatism in Neotropical Malpighiaceae.Biotropica 11: 219–233.
ANDERSON, W. R. 1990. The origin of the Malpighiaceae—the evidence frommorphology. Memoirs of the New York Botanical Garden 64: 210–224.
BAKER, H. G. 1978. Chemical aspects of the pollination biology of woodyplants in the tropics. In P. B. Tomlinson and M. H. Zimmermann [eds.],Tropical trees as living systems, 57–82. University Press, Cambridge,UK.
COCUCCI, A. A., A. M. HOLGADO, AND A. M. ANTON. 1996. Estudio mor-fologico y anatomico de los eleoforos pedicelados de Dinemandra eri-coides, Malpigiacea endemica del desierto de Atacama, Chile. Darwin-iana 34: 183–192.
D’ AMBROGIO DE ARGUESO, A. 1986. Manual de tecnicas en histologıa veg-etal. Hemisferio Sur Sociedad Anonima, Buenos Aires, Argentina.
DIZEO DE STRITTMATER, C. 1973. Nueva tecnica de diafanizacion. Boletınde la Sociedad Argentina de Botanica 15: 126–129.
FAEGRI, K., AND L. VAN DER PIJL. 1979. The principles of pollination ecol-ogy, 3rd ed. Pergamon Press, Oxford, UK.
FOUET, M. 1966. Contribution a l’etude cyto-taxonomique des Malpighiacees.Adansonia 6: 457–505.
GAHAN, P. B. 1984. Plant histochemistry and cytochemistry: an introduction.Academic Press, New York, New York, USA.
HAUMAN-MERCK, L. 1912. Sobre la polinizacion de una Malpighiacea delgenero Stigmaphyllon. Physis (Buenos Aires) 1: 81–87.
HICKEY, L. J. 1974. Clasificacion de la arquitectura de las hojas de las di-cotiledoneas. Boletın de la Sociedad Argentina de Botanica 16: 1–26.
LOBREAU-CALLEN, D. 1984. Pollen et paleobotanique des Malpighiaceae. Re-vue de Paleobiologie, volumen special: 131–138.
LOBREAU-CALLEN, D. 1989. Les Malpighiaceae et leurs pollinisateurs. Co-adaptation ou coevolution. Bulletin du museum national d’histoire na-turelle. Section B, Adansonia:, Botanique phytochimie 11: 79–94.
MULGURA, M. E. In press. Malpighiaceae. In N. Bacigalupo [ed.], Flora il-ustrada de Entre Rıos. Coleccion Cientıfica del Instituto Nacional deTecnologıa Agropecuaria 6 (4). Instituto Nacional de Tecnologıa Agro-pecuaria (INTA), Buenos Aires, Argentina.
NEFF, J. L., AND B. B. SIMPSON. 1981. Oil-collecting structures in the An-thophoridae (Hymenoptera): morphology, function, and use in system-atics. Journal of the Kansas Entomological Society 54: 95–123.
NIEDENZU, F. 1928. Malpighiaceae. In A. Engler [ed.], Das Pflanzenreich,141 (1-3): 1–870. Wilhelm Engelmann, Leipzig, Germany.
O’BRIEN, T. P., AND M. E. MCCULLY. 1981. The study of plant structure:principles and selected methods. Termarcarphi PTY, Melbourne, Austra-lia.
RAW, A. 1979. Centris dirrhoda (Anthophoridae), the bee visiting West In-dian cherry flowers (Malpighia punicifolia). Revista de Biologıa Tropical27: 203–205.

1944 [Vol. 88AMERICAN JOURNAL OF BOTANY
SAZIMA, M., AND I. SAZIMA. 1989. Oil-gathering bees visit flowers of eglan-dular morphs of the oil-producing Malpighiaceae. Botanica Acta 102:106–111.
STEINER, K. E. 1985. Functional dioecism in the Malpighiaceae: the breedingsystem of Spachea membranacea Cuatr. American Journal of Botany 72:1537–1543.
VINSON, S. B., H. J. WILLIAMS, G. W. FRANKIE, AND G. SHRUM. 1997. Floral
lipid chemistry of Byrsonima crassifolia (Malpighiaceae) and a use offloral lipids by Centris bees (Hymenoptera: Apidae). Biotropica 29: 76–83.
VOGEL, S. 1974. Olblumen und olsammelnde Bienen. Tropische und Subtro-pische Pflanzenwelt 7: 1–267.
VOGEL, S. 1990. History of the Malpighiaceae in the light of pollinationecology. Memoirs of the New York Botanical Garden 55: 130–142.


![Effect of different electrolytes on the swelling properties of calyx[4]pyrrole-containing polyacrylamide membranes](https://static.fdokumen.com/doc/165x107/631f4fc8d10f1687490fbd44/effect-of-different-electrolytes-on-the-swelling-properties-of-calyx4pyrrole-containing.jpg)

















