
Purification and characterization of NAD (P) H quinone reductase from the latex of Hevea...
11
PREPARATM BIOCHEMISTRY, 2s(1&2), s7 -67 (1995) PURIFICATION AND CIIARACTERIZATION OF AN NAD(P)H:QUINONE OXIDOREDUCTASE FROM GLYCINE MAX SEEDLINGS A. Rescigno, F. Sollai, S. Masala, M. C. Porcu, E. Sanjust, A. C. RinaHif, N. Curreli, D. Grifi and Augusto Rinaldi Istituto di Chimica Biologica, Università di Cagliari, Via della Pineta7l - 09125 Cagliari, Italy -f Dipartimento di Biochimica e Medicina sperimentale, università di Tor Vergata, Roma ABSTRACT An NAD(P)H:(quinone acceptor) oxidoreductase (EC 1.6.99.2) was purified fromGlycine max seedlings by means of chromatographic procedures. After 1371- fold purification, the enzyme showed a single band in IEF corresponding to an isoeléctric point of 6.1. A single band was also found in native-PAGE both by activity staining and Coomassie brilliant blue staining. The molecular mass determined in SDS-PAGE was 21900 Da, while in HPLC gel-filtration it was 61000 Da. The NAD(P)H:quinone oxidoreductase was able to use NADH or NADPH as the electron donor. Among the artificial quinoneswhich are reduced by this enzyme,6-hydroxydopa- and 6-hydroxydopamine-quinone are of particular interestbecause of their neurotoxiceffects. INTRODUCTION NAD(P)H:quinone oxidoreductase (EC 1.6-99.2) is a flavoprotein which shows diaphorase activity catalyzing the two-electron reduction of quinonic compounds to hydroquinones. Copyright @ 1995 by Marcel Dekker, Inc
Transcript of Purification and characterization of NAD (P) H quinone reductase from the latex of Hevea...

PREPARATM BIOCHEMISTRY, 2s(1&2), s7 -67 (1995)
PURIFICATION AND CIIARACTERIZATION OF AN
NAD(P)H:QUINONE OXIDOREDUCTASE FROM GLYCINE MAX
SEEDLINGS
A. Rescigno, F. Sollai, S. Masala, M. C. Porcu, E. Sanjust, A. C. RinaHif,
N. Curreli, D. Grifi and Augusto Rinaldi
Istituto di Chimica Biologica, Università di Cagliari, Via della Pineta7l - 09125
Cagliari, Italy-f
Dipartimento di Biochimica e Medicina sperimentale, università di Tor
Vergata, Roma
ABSTRACT
An NAD(P)H:(quinone acceptor) oxidoreductase (EC 1.6.99.2) was purified
fromGlycine max seedlings by means of chromatographic procedures. After 1371-
fold purification, the enzyme showed a single band in IEF corresponding to an
isoeléctric point of 6.1. A single band was also found in native-PAGE both by
activity staining and Coomassie brilliant blue staining. The molecular mass
determined in SDS-PAGE was 21900 Da, while in HPLC gel-filtration it was
61000 Da. The NAD(P)H:quinone oxidoreductase was able to use NADH or
NADPH as the electron donor. Among the artificial quinones which are reduced
by this enzyme, 6-hydroxydopa- and 6-hydroxydopamine-quinone are of particular
interest because of their neurotoxic effects.
INTRODUCTION
NAD(P)H:quinone oxidoreductase (EC 1.6-99.2) is a flavoprotein which
shows diaphorase activity catalyzing the two-electron reduction of quinonic
compounds to hydroquinones.
Copyright @ 1995 by Marcel Dekker, Inc

58 RESCIGNO ET AL.
The proposed scheme of the reaction is:
Q + NAD(P)H + H+ -r QHZ + NAD(P)+where Q is a quinone. This feature of the enzyme appears to be unique amongNADP(H)-oxidizing flavoproteins, which generally are one-electron transferringenzymesl, and therefore give rise to semiquinones that are readily autoxidizedwith the formation of the superoxide radical 02-. NAD(p)H:quinoneoxidoreductase (QR), reacting with both NADH and NADpH is widely spreadthroughout the animal kingdom, plants, bacteria and fungi, and has been isolatedfrom a number of sources including human tissues2 and plants3,+. In animal tissuesthe role of the enzyme is thought to constitute a cellular device controllingsemiquinone and superoxide radical formations. In fact, quinone reductase mightact in the cellular detoxification of quinones by shunting these substances awayfrom the redox cycling which is inherent to the one-electron reduction processes6.It has been proposed that in plants the quinone reductases may play a role in theenzymic mechanism for polymerization and depolymerization of lignin. In fact,
QR interacted with laccase and peroxidases, thereby increasing thedecarboxylation of the lignin model compound vanillic acid7. Moreover, theligninolytic basidiomycete Phanerochaete chrysosporium was seen to produceseveral quinone oxidoreductases8. other authors have proposed that eR might bea relevant membrane-peripheral redox system permitting an overproduction ofcarotenoids by regulating the redox state of quinonesa. on the other hand, thesignificance of the presence of QR in both roots and seedlings during the earlystages of germination still remains unclear. In this paper we report the purificationof QR from Glycine max seedlings and some properties are given. Crudehomogenate activity staining showed six bands, of which the largest was purified.
MATERIALS AND METHODS
Chemicals and instrumentation. Agarose Cibacron Blue-3GA, AH-Sepharose 48, NADH and NADPH were purchased from Sigma. 2,6-Dichlorophenol-indophenol was from Fluka. cM-cellulose was obtained fromwhatman International, and sephacryl s-200 HR from pharmacia. All otherreagents were of the highest grade available and used without further purification.SDS- and native-PAGE reagents, Mini-Protean ready gels and both molecularweight and isoelectric focusing standards were purchased from Bio-Rad. IEF was

NAD(P)H :QUINONE OXIDOREDUCTASE
carried out on a LKB-Multiphor tr electrophoresis unit. The molecular weight
determination was also carried out on a Beckman System Gold HPLC unit
equipped with a TSK-300 column, using standard gel-filtration from Bio-Rad.
Glycine max (L.) Merr seeds (1 Kg) were soaked in tap water for 3 hr.
Seedlings were grown for 7 days in the dark at25'C on wet vermiculite'
Purification. Soybean seedlings (630 g) were homogenized in 1500 ml of 50
mM Tris-HCl buffer (pH 7.5) for 3 min in a Waring Blender and the resulting
suspension was filtered through a nylon cloth. The filtered suspension was
centrifuged at26}00 x g for 40 min in a refrigerated centrifuge and the pellet then
discarded (Step l). The supernatant was applied to an Agarose Cibacron Blue-
3GA column (5x7 cm) previously equilibrated with 100 mM Tris-HCl buffer (pH
7.5). In these conditions QR was bound to the agarose. The resin was washed with
100 mM Tris-HCl buffer (pH 7.5) containing 100 mM NaCl until the absorbance
at 280 nm was below 0.01. Elution of QR took place at a flow-rate of 1 ml min-l
with the same buffer containing 200 mM NaCl (Step 2). The enzyme solution
from step 2 was concentrated by ultrafiltration and dialyzed against l5 I of 30 mM
Tris-HCl buffer (pH 7 .5); it was then applied to an AH-Sepharose 48 column (1x5
cm) equilibrated with 30 mM Tris-HCl buffer (pH 7.5) and washed with the same
buffer until A2gg was < 0.01. The enzyme was eluted with 60 mM of the same
buffer (Step 3). The pooled fractions from step 3 were concentrated and twice
dialyzed against 15 I of 30 mM potassium phosphate buffer (pH 6.5). The next
step was performed on a CM-Cellulose column (2.5x5 cm) equilibrated with the
same buffer as above. In these conditions the enzyme did not bind to the resin and
was therefore recovered in the washing, which was collected until A2gg was <0.01
(Step 4). The solution from step 4 was concentrated by ultrafiltration and dialyzed
against 15 I of 50 mM Tris-HCl buffer (pH 7.5). Finally, gel-filtration
chromatography was carried out on a Sephacryl 5-200 HR column (2.5x80 cm)
equilibrared with 50 mM Tris-HCl buffer (pH 7.5) and elution was performed
with the same buffer. The fractions with the highest specific activity were pooled
and concentrated by ultrafiltration (Step 5). The purification procedure is
summarized in Table l. All purification steps were performed at 4". The overalì
purification achieved was 1371 fold with a recovery of about 5lvo.The purified
enzyme was yellow in solution like other flavoproteins.
59

RESCIGNO ET AL.60
O Òt oo F--O l-- \O rl r)
=<D ra
V < - Èt-- \O tr) t--
$ \O cr)O =
()
o
X
0)
t
a-l- z
i l E- d J
()
(l
ùs>'
r \
I
' -
O \ N O \ \ O @
h F € 3 +
8 e 8 S pci c.i c.; oi --.;
$
s o o o o Ò ì= c . ) $ ò l
>r
È >F O
.9 >.' 6 >
ú a d
i : ( )^ =- È
oU)
r lc.r(.)
= MM H
m È
9 E +: o : =b u ; , _ . è (l O F È y , ,E = E € u )= v ! = =- c H * b h9 2 , ; 9 Èe ù rU

NAD(P)H : QUINONE OXIDOREDUCTASE
Enzyme assay. NAD(P)H:quinone reductase activity was measured with
NADH as the electron donor and 2,6-dichlorophenol-indophenol (DCPIP) as the
electron acceptor. The standard assay system contained 100 mM Tris-HCl buffer
(pH 7.5), 0.08% Triton X-100,0.5 mM NADH, and 40 pM DCPIP. The reaction
was started by the addition of 100 pl of a suitable diluted enzyme solution in a
final vol of 1 ml. The reduction of DCPIP was followed at l,6g6n- (e = 18 mM-l
"ttr-l; at 25'C. With the various other quinones used as acceptors, the enzymic
activity was followed by measuring the decrease of NADH at 340 nm. The effect
of temperature was analyzed by measuring the latter directly in the cuvette as soon
as the reaction was stopped. As for activity staining, after native-PAGE, the gel
was stained by immersing in 50 ml of 0.8 mM DCPIP dissolved in 100 mM Tris-
HCI buffer (pH 7.5) for 5 min. The excess of dye was removed and the gel was
washed twice with 100 ml of 100 mM Tris-HCl buffer (pH 1.5). A 2 mM NADH
solution was then applied to the stained gel. After 5-10 minutes, according to the
enzyme concentration, the QR activity showed up as bleached bands on a blue
background.
Protein analysis. Proteins were estimated by the Bradford method using
bovine serum albumin as the standarde. SDS- and native-PAGE were performed in
accordance with the Bio-Rad Mini-Protean tr ready gels Instruction Manual.
Isoelectric focusing was carried out as described by Righettir0.
RESULTS AND DISCUSSION
Criteria of purity. Quinone reductase, obtained with the purification
procedure summarized in TABLE I, was checked for homogeneity by PAGE
under various conditions. Only one band was detected by activity staining of the
gel both on non-denaturing PAGE and isoelectric focusing. Similarly, one band
was detected by Coomassie brilliant blue staining. The isoelectric point of QR was
6.1. Only one band was noted in SDS-PAGE in the presence and absence of
mercaptoethanol.
Molecular weight determination. The MW determination was carried out by
SDS-PAGE and HPLC gel-filtration. The MW determined with SDS-PAGE was
21900, both in the presence and the absence of mercaptoethanol (fg. l). On HPLC
6 l

62 RESCIGNO ET AL.
FIGURE 1
SDS-polyacrylamide gel electrophoresis of purified NAD(P)H:quinoneoxidoreductase. The first lane indicates the molecular weight standard proteins;the second lane, the purified quinone reductase. Standard proteins in MWdecreasing order: phosphorylase b, bovine serum albumin, ovalbumin, bovinecarboanhydrase, soybean trypsin inhibitor, hen egg lysozyme.
gel-filtration chromatography a single peak, corresponding to a MW of 616fi) +
1000 SD, was eluted. The difference between these two determinations may be
due to the fact that the molecular mass determined by gel-filtration depends on the
shape of the protein and that the QR purified from Glycine rnuc may consist of
two identical monomers, as reported for most quinone reductases6. From the
flavine content, in accordance with the method described by Mahlerll, the
ininimum molecular weight was estimated to be about 20000.

NAD(P)H : QUINONE OXIDOREDUCTASE
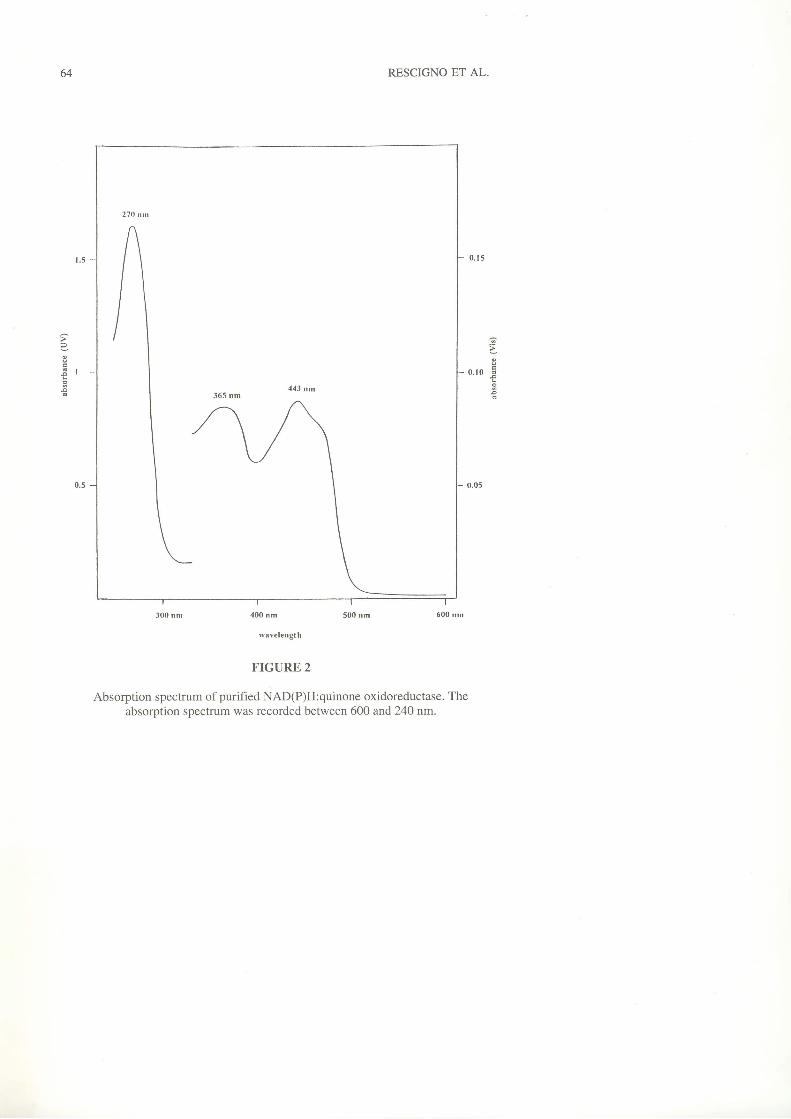
Spectroscopic properties. In the UV-VIS region the purified QR, in its
oxidized form, exhibited the typical absorption spectrum of flavoproteins with
absorption maxima at270,365, and 443 nm (ftq.2).
Substrate specificity and inhibitors. Glycine max qtinone reductase is able
to reduce a number of compounds in the presence of NADH and NADPH as the
electron donor. Quinones, substituted naphthoquinones (including natural
occurring compounds as juglone and lawsone), dyes and other compounds were
all tested for their ability to act as an electron acceptor or as an inhibitor. TABLE
II shows the substrate specificity of the enzyme. Every tested compound showed a
lower relative activity in respect to DCPIP. It is noteworthy that QR is able to
catalyze the reduction of 6-hydroxydopa-quinone (6-OHDOPA-quinone) and 6-
hydroxydopamine-quinone (6-OHDA-quinone). Both these compounds have a
particular interest both because oftheir neurotoxicityl2'I3, and the discovery that 6-
OHDOPA-quinone is the cofactor at the active site of bovine serum amine
oxidasela. 6-OHDOPA-quinone and 6-OHDA-quinone were obtained by
autoxidation of the corresponding reduced forms. In fact, 6-hydroxydopa (6-
OHDOPA) and 6-hydroxydopamine (6-OHDA) autoxidized in 100 mM Tris-HCl
buffer (pH 7.5) when in the presence of molecular oxygen. Autoxidation of these
compounds was followed at 485 nm (maximum absorption of the oxidized form)
and was complete after 10 min. Quinone reductase (0.2 pg rn1-l; and 0.5 mM
NADH were incubated in 100 mM Tris-HCI buffer (pH 7.5) containing 40 pM 6-
OHDOPA- or 6-OHDA-quinone in 1 ml volume. The relative activity of these
compounds was not referred to DCPIP activity since, during the reaction, an
equilibrium is established between 6-OHDOPA-quinone (or 6-OHDA-quinone),
which undergoes reduction, and 6-OH-DOPA (or 6-OH-dopamine) which is
rapidly oxidized by molecular oxygen. As a consequence of the above, NADH
consumption was greater than the stoichiometric amount needed to reduce 40 pM
quinone solution. 6-OHDA-quinone's relative activity was then calculated to be
30Vo of the value found for 6-OHDOPA-quinone.
We have noted that QR showed an action opposite to that of peroxidases
and laccases, the two enzymes involved in the metabolism of lignin. For example,
horseradish peroxidase catalyzes oxidation of synthetic compounds like
syringaldazine and o-dianisidine to the corresponding quinone derivatives in the
presence oî H2O2. In the same way, laccasesls and peroxidasesl6 use 2,4,5-
63
64 RESCIGNO ET AL.
300 nm 400 nm 500 nm ó00 nnr
rv{velengt h
FIGURE 2
Absorption spectrum of purified NAD(P)H:quinone oxidoreductase. Theabsorption spectrum was recorded between 600 and 24O nm.
F
5 l o r o E- - _ - o

NAD(P)H:QUINONE OXIDOREDUCTASE
TABLE II
Relative activity of the Glycine nax NAD(P)H:quinone oxidoreductase (QR) with
different electron acceptors. Activities are reported in relation to the standard QRassay with DCPIP. Quinone reductase was 0.04 mg ml-I. Acceptors concentration
was 40 pM, NADH was 0.5 mM' Acceptors reduction was measured as NADH
oxidation at 340 nm. N.D., not detectable.
65
Electron acceptor
DCPIPJugloneLawsonePotassium ferricyanide5,8-Dihydroxynaphthoquinone6-OHDOPA-quinone (*)6-OHDA-quinone (*)
Relativeactivity
(7o)
10058
N.D.44t9
10030
(*) Details about relative activities of 6-hydroxydopa-quinone and 6-
hydroxydopamine-quinone are given in the Results and Discussion section.
trihydroxyphenethylamine (6-OHDA) and 2,4,5ìrihydroxyphenylalanine (6-
OHDOPA) as substrates. Glycine max QR reduces the quinones generated by
peroxidase and laccase to the corresponding hydroquinones using NADH and/or
NADPH as a substrate. In the presence of DCPIP, QR showed a K- value for
NADH 341 pM, while Kn' for NADPH was 190 pM.
We have tested the inhibitory effect of a number of compounds on the QR
activity. TABLE III shows the more significant results. The enzyme was totally
inhibited by a0 pM dicumarol, a typical inhibitor of quinone reductases.
Effect of pH and temperature. Effect of pH on QR activity was tested in 100
mM Tris-HCl buffer using DCPIP as the electron acceptor. The pH curve showed
an optimum at 7.5. Over 6O7o of activity still remained at pH 9.
The effect of temperature on the rate of the reaction was studied over the
range 7 -57"C under standard assay conditions. Over the tange 7 -3O" a straight line
was obtained in the Arrhenius plot with an activation energy of 6 kcal mol-l.

66 RESCIGNO ET AL.
TABLE III
Relative inhibition of the G.max NAD(P)H:quinone oxidoreductase. Inhibition isgiven in relation to the standard assay with DCPIP taken as l00%o of activify. Allinhibitors and DCPIP were present in the concentration of 40 pM; NADH was 0.5mM. The inhibiting effect was measured as NADH oxidation at 340 nm.
Inhibitor Relative inhibition ( 7o )
DicumarolCibacron Blue F3G-AProcion Blue MX-RAlizarin Red S
Between 30' and 43'C, the enzyme activity was unaffected by the
temperature, the graph showing a plateau; over 43oC it began to decrease. This
behaviour could not be compared with other quinone reductases because of the
general lack of data in this regard.
CONCLUSIONS
We have reported the purification of QR ftom Glycine max seedlings.
The yield obtained and the high specific enzymic activity achieved make
this purification procedure useful for obtaining good amounts of QR, sufficient for
further studies. In fact, QR is not yet commercially available and, therefore,
Glycine max seedlings seem to constitute an inexpensive and easily accessible
source.
As for the role this enzyme plays in lignin metabolism of plants, it is
interesting to point out its behaviour opposite to that of laccase and peroxidase
activities. On the basis of our present knowledge, it is not yet possible clearly to
understand to what extent QR is involved in the early stages of seed germination.
Finally, the purified enzyme can be seen to be a useful tool in the study of
the relationship between the brain's NAD(P)H:quinone oxidoreductase and the
neurotoxic quinone derivatives of cathecholamines. In this regard, further studies
are in progress in our laboratories.
100t )
7538

NAD(P)H : QUINONE OXIDOREDUCTASE
REFERENCES
1. T.Iyanagi, Chem. Scr.,27A,3l (1981).
2. D. Smith, L. F. Martin and R. V/allin, Cancer Lett.,42, 103- 110 (1988).
3. F. Guerrini, V. Valenti and P. Pupillo, Plant Physiol., 85, 828-834 (1987).
4. M. P. Mayer, U. Nievelstein and P. Beter, Plant Physiol. Biochem., 30 (4),
389-398 (1992).
5. C. Lind, P. Hochstein and L. Ernster, Arch. Biochem. Biophys..216, ll8-
18s (1982).
6. C. Bryant and W. D. McElroy, Nitroreductases. In "Chemistry and
Biochemistry of Flavoenzymes" Vol.2, F. Mùller ed., CRC Press, Boca
Raton, FL, 1991, pp. 295-297.
7. P. Ander, M. Chittra, R. L. Farrel and K. E. Eriksson, J. Biotechnol. 13,
189-198 ( r990) .
8. D. Constam, A. Muheim, W. Zimmermann and A. Fiechter, J. General
Microbiol. l3l, 2209-2214 (1991).
9. M. M. Bradford, Anal. Biochem. 72,248-254 (1976).
10. P. G. Righetti, "Isoelectric focusing: theory, methodology and applications"
Work T.S. and Burdon R. H. ed., Elsevier Press, London, UK, (1986).
11. H. R. Malher, Methods of Enzymology,2,7O7-7ll (1955).
12. G. R. Breese and T. D. Traylor, J. Pharmacol. Exp. Ther. 174,413-420
(r91O).13. J. W. Olney, C. F. Zorumski, G. R. Stewart, M. T. Price, G. Wang and J.
Labruyere, Exp. Neurol., 108,269-272 (1990).
14. S.M. Janes, D. Mu, D. Wemmer, A. J. Smith, S. Kaur, D. Maltby, A. L.
Burlingame and J. P. Klinman, Science 248,981-981 (1990).
15. A. Padiglia, R. Medda, A. Rescigno and G. Floris, Italian J. Biochem. 43
(r),24-28 (1994).
16. A. Padiglia, A. Rescigno, R. Medda and G. Floris, Anal. Lett. 27 (3),523-
s3o (1994).
67




















