
The effect of clams ( Tapes philippinarum) on changes in microbial community structure in tidal flat...
12
Ecological Engineering 22 (2004) 185–196 The effect of clams (Tapes philippinarum) on changes in microbial community structure in tidal flat sediment mesocosms, based on quinone profiles Udin Hasanudin a,∗ , Masafumi Fujita b , Tadao Kunihiro c , Koichi Fujie a , Teruaki Suzuki d a Fujie-Goto Laboratory, Department of Ecological Engineering, Toyohashi University of Technology, Tempaku-cho, Toyohashi 441-8580, Japan b Department of Civil and Environmental Engineering, University of Yamanashi, 4-3-11 Takeda, Kofu, Yamanashi 400-8511, Japan c Faculty of Environmental and Symbiotic Sciences, Prefectural University of Kumamoto, 3-1-100 Tsukide, Kumamoto 862-8502, Japan d Aichi Fisheries Research Institute, 97, Wakamiya, Miya-cho, Gamagori 443-0021, Japan Received 4 November 2003; received in revised form 5 May 2004; accepted 18 May 2004 Abstract To elucidate the relationship between macrobiotics and microorganisms on pollutant degradation in tidal flat sediment, changes in water quality were monitored and shifts in microbial community structure were evaluated using the quinone profile method in mesocosms of tidal and subtidal flat sediment with and without clams. Dissolved organic carbon (DOC) and NH 4 -N concentrations in pore water of tidal sediment with clams were higher than in sediment without clams over a 24 h period. This is explained by the ability of clams to consume particulate organic matter and excrete DOC and NH 4 -N. Otherwise, the effect of clams was relatively low in subtidal sediments. The presence of clams and tidal conditions changed the predominant quinone species in both sediments. Ubiquinone with 10-isoprene unit (UQ-10) and menaquinone with 6-isoprene unit (MK-6) were predominant quinone species in tidal and subtidal sediment with clams, respectively. The effect of clams on changes in microbial community structure in tidal sediment was more rapid than in subtidal sediment. The differences in microbial community structure in tidal sediment with and without clams were mainly due to the profile changes in UQ-8, UQ-9, UQ-10, MK-7 (H 8 ) and MK-9 (H 2 )-containing bacteria. © 2004 Elsevier B.V. All rights reserved. Keywords: Clams; Dissimilarity; Microbial community structure; Principal component analysis; Quinone profile; Tidal flat 1. Introduction Tidal flat sediments are recognized as highly pro- ductive areas, that function as active buffer zones in the ∗ Corresponding author. Tel.: +81-532-44-6924; fax: +81-532-44-6914. E-mail address: [email protected] (U. Hasanudin). transport of nutrients between land and sea. Population increases, urbanization, and industrialization enhance nutrient loads to the environment, including to tidal flats. Imbalances between tidal flat sediments’ purifi- cation capacity and nutrient loads could turn the func- tion of tidal flat sediments from a purifier to a source of nutrients. To improve the function of tidal flat sedi- ments on seawater purification, the symbiotic function 0925-8574/$ – see front matter © 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.ecoleng.2004.05.003
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of The effect of clams ( Tapes philippinarum) on changes in microbial community structure in tidal flat...

Ecological Engineering 22 (2004) 185–196
The effect of clams (Tapes philippinarum) on changes in microbialcommunity structure in tidal flat sediment mesocosms,
based on quinone profiles
Udin Hasanudina,∗, Masafumi Fujitab, Tadao Kunihiroc,Koichi Fujiea, Teruaki Suzukid
a Fujie-Goto Laboratory, Department of Ecological Engineering, Toyohashi University of Technology, Tempaku-cho,Toyohashi 441-8580, Japan
b Department of Civil and Environmental Engineering, University of Yamanashi, 4-3-11 Takeda, Kofu, Yamanashi 400-8511, Japanc Faculty of Environmental and Symbiotic Sciences, Prefectural University of Kumamoto, 3-1-100 Tsukide, Kumamoto 862-8502, Japan
d Aichi Fisheries Research Institute, 97, Wakamiya, Miya-cho, Gamagori 443-0021, Japan
Received 4 November 2003; received in revised form 5 May 2004; accepted 18 May 2004
Abstract
To elucidate the relationship between macrobiotics and microorganisms on pollutant degradation in tidal flat sediment, changesin water quality were monitored and shifts in microbial community structure were evaluated using the quinone profile methodin mesocosms of tidal and subtidal flat sediment with and without clams.
Dissolved organic carbon (DOC) and NH4-N concentrations in pore water of tidal sediment with clams were higher thanin sediment without clams over a 24 h period. This is explained by the ability of clams to consume particulate organic matterand excrete DOC and NH4-N. Otherwise, the effect of clams was relatively low in subtidal sediments. The presence of clamsand tidal conditions changed the predominant quinone species in both sediments. Ubiquinone with 10-isoprene unit (UQ-10)and menaquinone with 6-isoprene unit (MK-6) were predominant quinone species in tidal and subtidal sediment with clams,respectively. The effect of clams on changes in microbial community structure in tidal sediment was more rapid than in subtidalsediment. The differences in microbial community structure in tidal sediment with and without clams were mainly due to theprofile changes in UQ-8, UQ-9, UQ-10, MK-7 (H8) and MK-9 (H2)-containing bacteria.© 2004 Elsevier B.V. All rights reserved.
Keywords: Clams; Dissimilarity; Microbial community structure; Principal component analysis; Quinone profile; Tidal flat
1. Introduction
Tidal flat sediments are recognized as highly pro-ductive areas, that function as active buffer zones in the
∗ Corresponding author. Tel.:+81-532-44-6924;fax: +81-532-44-6914.
E-mail address: [email protected] (U. Hasanudin).
transport of nutrients between land and sea. Populationincreases, urbanization, and industrialization enhancenutrient loads to the environment, including to tidalflats. Imbalances between tidal flat sediments’ purifi-cation capacity and nutrient loads could turn the func-tion of tidal flat sediments from a purifier to a sourceof nutrients. To improve the function of tidal flat sedi-ments on seawater purification, the symbiotic function
0925-8574/$ – see front matter © 2004 Elsevier B.V. All rights reserved.doi:10.1016/j.ecoleng.2004.05.003

186 U. Hasanudin et al. / Ecological Engineering 22 (2004) 185–196
of microorganisms and macrobiotics on organic mat-ter decomposition should be optimized. Much atten-tion has been paid to the role and effect of tidal flatson the quality of seawater in tidal estuaries (Sohmaet al., 2001). The role of sediments in nutrient cy-cling has primarily been studied from the standpoint ofcarbon and nutrient turnover (Sundback et al., 2000).Most of the early transformation and degradation pro-cesses of organic matter in tidal flat sediments wererevealed to be mediated by microorganisms (Rutterset al., 2002). The influences of photosynthetic andheterotrophic organisms on nutrients and oxygen ex-changes between sediment and shallow water has alsobeen studied (Sundback et al., 2000). However, therelationship between microbial community structureand macrobiotic activity in tidal flat sediments has notyet been elucidated.
Clams are recognized as having filtering capabilitiesthrough consumption of particulate organic matter andexcretion of feces. Clams’ excretion activities increaseNH4-N, total Kjeldahl nitrogen and BOD concentra-tion (Zhu et al., 1999). Thus, the filtering function ofclams might produce carbon and nutrient sources formicroorganisms in tidal flat sediments.
Organic matter mineralization in sediments is a keycomponent of the carbon and nutrient cycles of manyaquatic ecosystems; furthermore, the metabolism oforganic matter is a major factor driving sedimentbiogeochemistry (King, 1986). Intertidal and subtidalestuarine sediments are recognized as important sitesfor nutrient transformation and sequestration via bio-geochemical cycling (Ogilvie et al., 1997; Megalhãeset al., 2003). Tidal flat purification functions aremainly determined by the capabilities of sedimentmicroorganisms to mineralize organic matters (King,1986; Lee et al., 1998).
Increased carbon and nutrient concentrations shouldincrease microbial growth in sediments. Therefore, thefilter function of clams can be used to increased micro-bial populations in tidal flat sediments. To determinethe contribution of clams to seawater purification, itis important to evaluate changes of water quality andthe characteristics of microbial community structurein tidal flat sediment with and without clams.
The quinone profile method has gained increasedrecognition as a simple and useful tool for the anal-ysis of changes in microbial population dynamics inmixed cultures. Microbial respiratory quinone is one
of the constituents of the bacterial respiratory chainand plays an important role as an electron trans-porter (Collins and Jones, 1981; Hu et al., 1999a).Quinones exist in almost all bacteria and in general,one species or genus of bacteria has only one domi-nant type of respiratory quinone. The quinone profile,which is usually represented as the mole fraction ofeach quinone type, should be specific for a microbialcommunity and can thus be used as an index to char-acterize the microbial community (Collins and Jones,1981; Hiraishi, 1999; Hu et al., 1996, 2001). Somechanges in a microbial population may not bringabout change in the quinone profile, because differ-ent bacteria can have the same dominant quinone(Hiraishi, 1988). However, any change in the quinoneprofile represents a change in the bacteria population(Hu et al., 1999b). It was demonstrated from the pre-vious study (Fujie et al., 1998) that the species with asmaller fractional content (usually as small as about0.001 in mole fraction) in an ecosystem could also bequantified with a higher reliability by the chemicalanalysis of microbial quinones. This made it possi-ble to evaluate the microbial community without aloss of information about the species with smallerfraction, which also play an important role in anecosystem (Hu et al., 1999b). The difference betweentwo quinone profiles, expressed as the mole fractionof each quinone type, can be quantified using the dis-similarity (D) index (Hiraishi, 1999; Hu et al., 1999a).Also, quinone content can be used as a biomass index(Hiraishi et al., 1998; Hu et al., 2001). Therefore, bothmicrobial community structure and biomass can besimultaneously quantified by analyzing quinones in amicrobial community (Hu et al., 2001). The quinoneprofile method has been successfully used to quan-tify the changes in microbial community in sediment(Hendrick and White, 1986), refractory industrialwastewater (Fujie et al., 1994; Hu et al., 1996, 1997),and the biological process for the treatments of domes-tic sewage (Hiraishi, 1988) and rivers (Kunihiro et al.,2002).
In this study, in order to evaluate the ability of clamsto enhance organic matter and nutrient transformationsin tidal and subtidal flat sediments, the diurnal varia-tion in pore water quality was investigated. Shifts inmicrobial community structure were evaluated usingquinone profiles in tidal and subtidal flat sedimentswith or without clams.

U. Hasanudin et al. / Ecological Engineering 22 (2004) 185–196 187
2. Materials and methods
2.1. Research sites
All experimental plots were located close toMikawa Bay in Aichi Fishery Research Institute,Gamagori, Aichi, Japan. The experiment was con-ducted in four plots of mesocosms of tidal and subtidalsediments. Subtidal sediment was constructed 15 cmlower than tidal sediment to maintain the sedimentalways under immersion condition. The dimensionsof each plot were 2.0 m (W)× 2.5 m (L)× 0.4 m (D)with sand median grain size of 0.95 mm. Clams with1.9 cm average length and 1.2 g average whole wetweight (included shell) were added to two plots ofthese sediments corresponding to about 5000 clamsper plot on 23 July 2002. Fresh seawater was pumpedfrom about 200 m offshore and supplied to each tidein the mesocosms. To synchronize water level withnatural condition, the water level in Mikawa Bay wasonline measured and the data was used to arrangeflow rate of seawater to the mesocosms.
2.2. Sampling methods
One month after adding the clams, pore water qual-ity in all plots, i.e., DOC, NH4-N, NO3-N, NO2-Nand dissolved organic nitrogen (DON), were moni-tored for 24 h to investigate the effect of clams. About20 ml of pore water was taken from each plot of sed-iment using a syringe with a capillary tube equippedwith double layers of (1.2�m) filter paper (GF/Cfilter, Whatman). Seawater dissolved oxygen (DO)and temperature of seawater were measured in exper-imental field plot using an aqua-quality multi sensor(Model Clorotec ACL-1183-PDK, Alec ElectronicCo. Ltd.).
Sediment cores (n = 2) were collected from the0 to 10 cm depth of each plot using an acrylic tube(5 cm i.d.× 50 cm length) every month from July toOctober 2002. The samples obtained from all plotswere immediately cooled by ice and then stored at−20◦C before quinone analysis.
2.3. Analytical methods
Pore water DOC concentrations were measuredwith a TOC analyzer (TOC-5000A, Shimadzu). Ni-
trogen compounds (NH4-N, NO3-N, NO2-N anddissolved total nitrogen (DTN)) were analyzed withan automatic water analyzer (Model AACS-III,Bran + Luebbee). t-Test statistical analyses wereperformed to distinguish differences in pore waterquality between sediment with and without clams.Microbial quinones in the sediments were analyzedbased on the procedure previously reported (Hu et al.,1999a, 2001). Quinones were first extracted fromthe sediments by a mixture of chloroform–methanol(2:1, v/v) and then re-extracted by hexane. Thecrude extract was purified using solid extraction car-tridge (Sep-Pak® Plus Silica, Waters). The types andconcentrations of quinones were determined withHPLC equipped with an ODS column (Zorbax-ODS,4.6 mm i.d. × 250 mm, Shimadzu-Dupont) and amulti-channel UV detector (photodiode array detec-tor, model: SPD-M10A, Shimadzu). A mixture ofmethanol and di-isopropyl ether (9:2, v/v) was used asthe mobile phase at a flow rate of 1.0 ml min−1. Thetemperature of the column oven was maintained at35◦C. Quinone species were identified according tothe retention time on the column and the UV spectrumof each peak observed in the multi-channel UV de-tector. The linear relationship between the logarithmof the retention times of quinones and the equivalentnumber of isoprenoid unit (ENIU) was also used toidentify the quinone type (Hiraishi et al., 1991; Huet al., 1999a). Ubiquinone with 10 isoprenoid andvitamin K1 were used as the quantitative standardsfor ubiquinone and menaquinone, respectively. TheENIU can be approximated by the following equation:
ENIUk = a + b log
(ETk
ETstd
)+ c
[log
(ETk
ETstd
)]2
where ETk represents the elution time of a quinonespeciesk, and ETstd represents the elution time of stan-dard quinone. The constants are shown asa, b, andc,which are empirically obtained for each HPLC system(Tamaoka et al., 1983). The amounts of quinone werecalculated from the peak area based on the mole ab-sorption coefficients (ubiquinones: 14.4 mM−1 cm−1,menaquinones: 17.4 mM−1 cm−1 and plastoquinones:15.3 mM−1 cm−1) (Kroger, 1978). The quinone molefraction was calculated as a ratio of the quinone con-tent in the speciesk to the total quinone content.In this paper, the abbreviation of quinone types are

188 U. Hasanudin et al. / Ecological Engineering 22 (2004) 185–196
ubiquinone: UQ, menaquinone: MK, plastoquinone:PQ and vitamin K1: VK1.
The difference between microbial community struc-ture (quinone profiles) in the sediments with and with-out clams, expressed as the mole fraction of eachquinone type ini (with clams) andj (without clams)samples, can be quantified with dissimilarity index (D)defined by the equation ofHiraishi et al. (1991)asfollows:
D(i, j) = 1
2
n∑k=1
∣∣fki − fkj∣∣
where fki and fkj are the mole fractions of quinonespeciesk for i andj samples, respectively.D = 0 showsthat the two quinone profiles are identical (Hu et al.,1999a). On the basis of the fluctuations in dissimilarityof quinone profiles extracted from one soil, dissimilar-ities greater than 0.2 indicate a statistically significantdifference in the quinone profile between two samples(Fujie et al., 1998).
Principal component analysis (PCA) was performedto evaluate and interpret the shifts of microbial com-munity structure in tidal and subtidal sediments withand without clams. The shifts of quinone mole frac-tions at different sampling times were used to evaluatethe shifts of microbial community structure during the
Fig. 1. DOC concentrations in pore water of tidal and subtidal sediments with and without clams.
experiment. The mole fraction shifts of each quinonespecies were used to identify the changes of individ-ual quinone species in the sediment with and withoutclams. The software StatPartner Version 4 (O-ha Co.)was used for these purposes.
3. Results and discussion
3.1. Diurnal variation of DOC concentration
The diurnal variation of DOC concentration in porewater of tidal and subtidal sediments with and withoutclams is shown inFig. 1. Concentration of DOC intidal sediment with clams was about 2 mg l−1 higherthan that in the sediment without clams. Clams con-sumed particulate organic matters (POM) and excretedtheir feces as a source of DOC. The concentrationincreased with decreasing water level. During emer-sion, the penetration of oxygen into sediments mayincrease (Durand et al., 2002; Kuwae et al., 2003) andlead to higher production of DOC due to clam activi-ties. In addition, during daytime low water level, lightintensity was highest. Photosynthetic bacteria activ-ity might be stimulated to produce more oxygen. DOconcentration of overlying water at around 13.00 a.m.was about 7 mg l−1, while at around 6.00 a.m. it was

U. Hasanudin et al. / Ecological Engineering 22 (2004) 185–196 189
only about 5 mg l−1. Due to higher DO concentration,the production of DOC from clams also higher at lowwater level.
A different pattern was found in subtidal sediment.t-Test analysis showed that DOC concentrations in thesediment with and without clams were similar. Sincesubtidal sediment is always under immersion condi-tions, it is expected that DO concentration should below, lowering the activity level of clams. Therefore,there was no significant difference in DOC concentra-tion in the sediment with and without clams.
3.2. Diurnal variation of NH4-N, NO3-N and NO2-Nconcentrations
As shown inFig. 2, the concentration of NH4-N intidal sediment with clams was higher than that in thesediment without clams. The content of NH4-N in thissediment was about 90% of dissolved inorganic nitro-gen (DIN) or about 80% of DTN. This result is con-sistent with the results of previous research.Zhu et al.(1999)has reported that the excretion rate ofT. philip-pinarum is about 50 and 80 mg (kg clam)−1 day−1 forammonia nitrogen and total Kjeldahl nitrogen (TKN),respectively, indicating that NH4-N is the primaryproduct of clam excretion. NH4-N concentrationswere also higher at low water levels. A differentpattern occurred in subtidal sediment. Low concen-
Fig. 2. Concentrations of NH4-N, NO3-N, NO2-N and DON in pore water of tidal and subtidal sediments with and without clams.
trations of DO and organic matter in the sedimentcaused a reduction in clam activity. Therefore, NH4-Nconcentration in subtidal sediment with and with-out clams was relatively low and similar each other.A similar pattern was observed in changes of DOCconcentrations. We propose that DOC and NH4-Nconcentrations can be used as indicators of clam ac-tivities in tidal flat sediment. The concentrations ofNO3-N and NO2-N in tidal sediment with and with-out clams were relatively low and stable over a 24 hperiod. This indicated that these concentrations haveno direct relationship to clam activities. NO3-N wasmainly produced from NH4-N oxidation by nitrifyingbacteria (Usui et al., 2001). While, NO2-N producedvia ammonium oxidation and during denitrification(Megalhães et al., 2003).
The diurnal variation of pore water quality indicatedthat clams could be used to improve the transformationof organic matter and nutrients in tidal flat sediment.Finally, changes in the chemical environment mightlead to changes in microbial community structure intidal flat sediment. To assess this, quinone profileswere performed.
3.3. Quinone composition
Quinone contents in the sediment with and with-out clams at different sampling times are shown in

190 U. Hasanudin et al. / Ecological Engineering 22 (2004) 185–196
Fig. 3. Quinone content in tidal and subtidal sediments with and without clams.
Fig. 3. Clams had almost no effect on plastoquinone-9(PQ-9) and VK1 contents. However, PQ-9 and VK1contents in tidal sediment were higher than in subtidalsediment. PQ-9 and VK1-containing microorganismsare light regulated photosynthetic groups. Higher lightintensity in tidal sediment led to increased PQ-9 andVK1 content.
The effects of clams on UQ and MK content var-ied with sampling time. Changes in environmentalconditions, such as temperature and DO, affected theactivity level of clams (Zhu et al., 1999) and mi-croorganisms in the sediment. Temperature and DOchanges during the experiment are shown inFig. 4. In
Fig. 4. Changes in temperature and DO of seawater during theexperiment.
August, mean temperature of seawater was 27.4◦C.At this temperature, clam activities are high (Suzukiet al., 1998). Thus, production of dissolved organicmatter was expected to be higher, but clam activitiesalso increased DO consumption rates in the sediment.Due to DO limitation, UQ content in tidal sedimentwith clams was lower than in the sediment withoutclams.Hiraishi (1999)reported that UQ occur mainlyin strictly aerobic microbes. MK content in the sed-iment with clams was higher than in the sedimentwithout clams. MK is mainly produced by strictlyanaerobic respiratory bacteria (Hiraishi, 1999). Lim-itations of DO concentration in the sediment withclams stimulated the growth of MK-containing bacte-ria. A different pattern occurred in subtidal sediment.Changes of UQ and MK contents in subtidal sedi-ment with and without clams showed similar trends,indicating that the effect of clams on microbial com-munity structure was relatively low.
In September, UQ and MK contents in tidal sedi-ment with clams were higher than in sediment withoutclams. Temperature of seawater decreased to about24◦C, causing a decrease in clam activities. There-fore, DO concentration in the sediment increased.High concentration of dissolved organic matter fromclam activities and the increase of DO concentrationenhanced the growth rate of UQ-containing microor-ganism in the sediment with clams. From August to
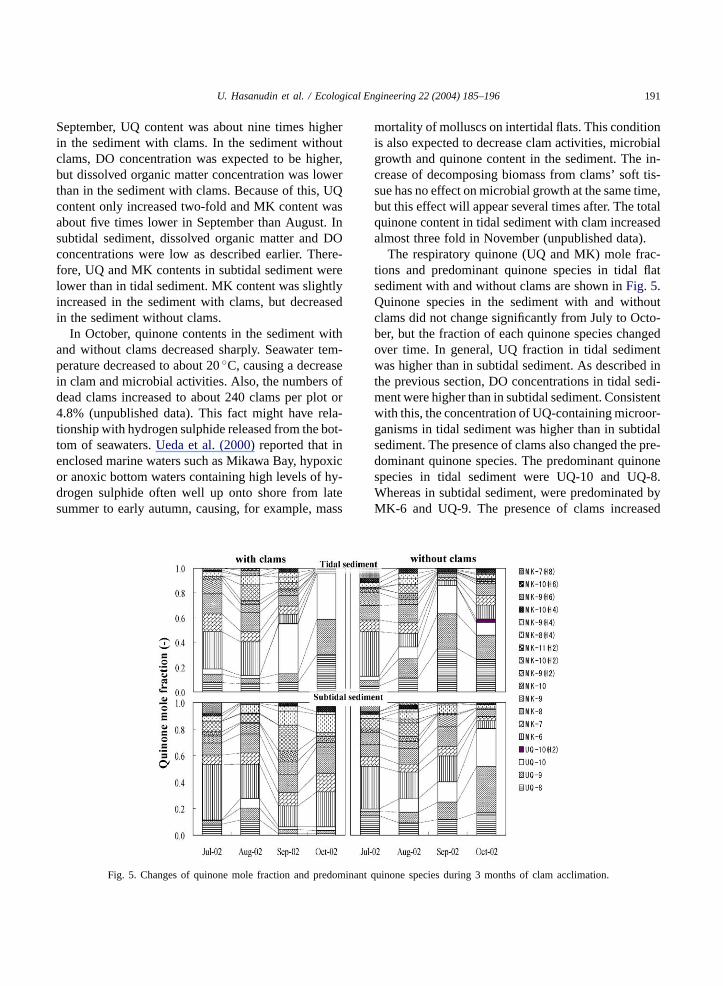
U. Hasanudin et al. / Ecological Engineering 22 (2004) 185–196 191
September, UQ content was about nine times higherin the sediment with clams. In the sediment withoutclams, DO concentration was expected to be higher,but dissolved organic matter concentration was lowerthan in the sediment with clams. Because of this, UQcontent only increased two-fold and MK content wasabout five times lower in September than August. Insubtidal sediment, dissolved organic matter and DOconcentrations were low as described earlier. There-fore, UQ and MK contents in subtidal sediment werelower than in tidal sediment. MK content was slightlyincreased in the sediment with clams, but decreasedin the sediment without clams.
In October, quinone contents in the sediment withand without clams decreased sharply. Seawater tem-perature decreased to about 20◦C, causing a decreasein clam and microbial activities. Also, the numbers ofdead clams increased to about 240 clams per plot or4.8% (unpublished data). This fact might have rela-tionship with hydrogen sulphide released from the bot-tom of seawaters.Ueda et al. (2000)reported that inenclosed marine waters such as Mikawa Bay, hypoxicor anoxic bottom waters containing high levels of hy-drogen sulphide often well up onto shore from latesummer to early autumn, causing, for example, mass
Fig. 5. Changes of quinone mole fraction and predominant quinone species during 3 months of clam acclimation.
mortality of molluscs on intertidal flats. This conditionis also expected to decrease clam activities, microbialgrowth and quinone content in the sediment. The in-crease of decomposing biomass from clams’ soft tis-sue has no effect on microbial growth at the same time,but this effect will appear several times after. The totalquinone content in tidal sediment with clam increasedalmost three fold in November (unpublished data).
The respiratory quinone (UQ and MK) mole frac-tions and predominant quinone species in tidal flatsediment with and without clams are shown inFig. 5.Quinone species in the sediment with and withoutclams did not change significantly from July to Octo-ber, but the fraction of each quinone species changedover time. In general, UQ fraction in tidal sedimentwas higher than in subtidal sediment. As described inthe previous section, DO concentrations in tidal sedi-ment were higher than in subtidal sediment. Consistentwith this, the concentration of UQ-containing microor-ganisms in tidal sediment was higher than in subtidalsediment. The presence of clams also changed the pre-dominant quinone species. The predominant quinonespecies in tidal sediment were UQ-10 and UQ-8.Whereas in subtidal sediment, were predominated byMK-6 and UQ-9. The presence of clams increased
192 U. Hasanudin et al. / Ecological Engineering 22 (2004) 185–196
Fig. 6. Dissimilarity index of respiratory quinone (UQ and MK)profiles between plots with and without clams in tidal and subtidalsediments.
DOC and NH4-N concentrations in tidal and subti-dal sediment as described previously. Clam activitiesalso increased DO consumption rate in the sediment.Those conditions might lead to change in quinonemole fraction and predominant quinone species.
3.4. Changes of microbial community structure
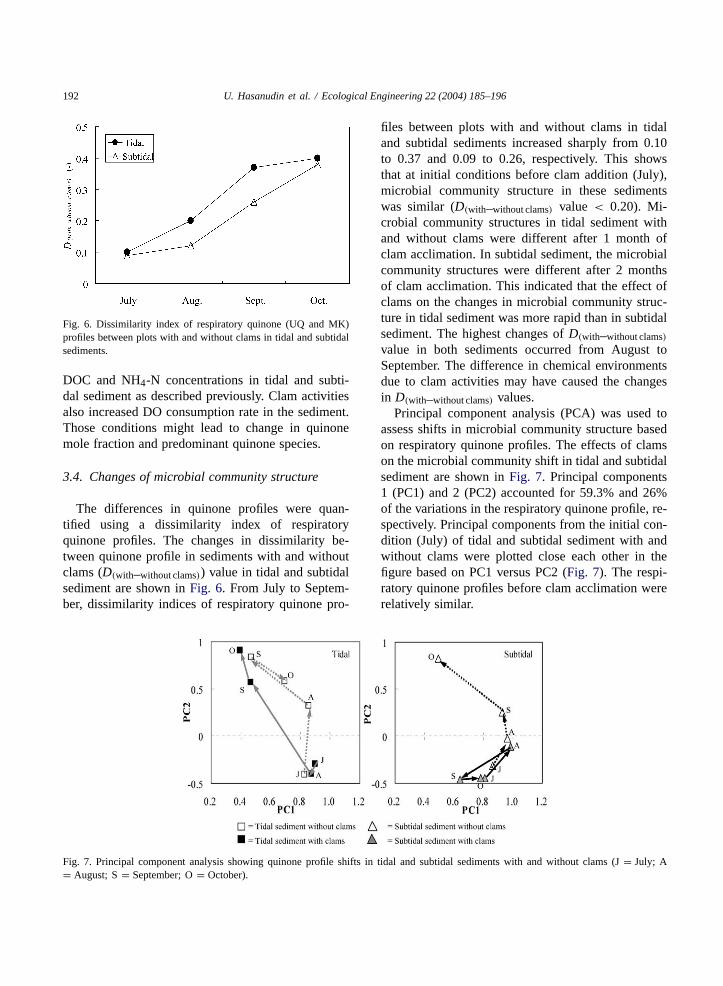
The differences in quinone profiles were quan-tified using a dissimilarity index of respiratoryquinone profiles. The changes in dissimilarity be-tween quinone profile in sediments with and withoutclams (D(with–without clams)) value in tidal and subtidalsediment are shown inFig. 6. From July to Septem-ber, dissimilarity indices of respiratory quinone pro-
Fig. 7. Principal component analysis showing quinone profile shifts in tidal and subtidal sediments with and without clams (J= July; A= August; S= September; O= October).
files between plots with and without clams in tidaland subtidal sediments increased sharply from 0.10to 0.37 and 0.09 to 0.26, respectively. This showsthat at initial conditions before clam addition (July),microbial community structure in these sedimentswas similar (D(with–without clams) value < 0.20). Mi-crobial community structures in tidal sediment withand without clams were different after 1 month ofclam acclimation. In subtidal sediment, the microbialcommunity structures were different after 2 monthsof clam acclimation. This indicated that the effect ofclams on the changes in microbial community struc-ture in tidal sediment was more rapid than in subtidalsediment. The highest changes ofD(with–without clams)value in both sediments occurred from August toSeptember. The difference in chemical environmentsdue to clam activities may have caused the changesin D(with–without clams) values.
Principal component analysis (PCA) was used toassess shifts in microbial community structure basedon respiratory quinone profiles. The effects of clamson the microbial community shift in tidal and subtidalsediment are shown inFig. 7. Principal components1 (PC1) and 2 (PC2) accounted for 59.3% and 26%of the variations in the respiratory quinone profile, re-spectively. Principal components from the initial con-dition (July) of tidal and subtidal sediment with andwithout clams were plotted close each other in thefigure based on PC1 versus PC2 (Fig. 7). The respi-ratory quinone profiles before clam acclimation wererelatively similar.

U. Hasanudin et al. / Ecological Engineering 22 (2004) 185–196 193
In July, quinone profiles in tidal sediment withand without clams had high positive correlation withPC1 and negative correlation with PC2. In August,quinone profiles in the sediment with clams still hadhigh positive correlation with PC1 (0.87) and neg-ative correlation with PC2 (−0.4). However, in thesediment without clams there was high positive cor-relation with PC1 (0.86) and low positive correlationwith PC2 (0.32). This provides evidence that clamsinfluenced the changes of quinone profiles in tidalsediment. D(with–without clams) value increased from0.10 to 0.20 in this period (Fig. 6). In September,quinone profiles in tidal sediment with clams changedfrom high positive correlation with PC1 (0.87) andnegative correlation with PC2 (−0.4) to low positivecorrelation with PC1 (0.47) and PC2 (0.57). In thesediment without clams, quinone profiles changed tolow positive correlation with PC1 (0.47) and highpositive correlation with PC2 (0.84). The highestincrease ofD(with–without clams) value occurred fromAugust to September. In October, quinone profilesin the sediment with clams had low positive (0.4)and high positive (0.91) correlation with PC1 andPC2, respectively. However, in the sediment with-
Fig. 8. Principal component analysis showing differences in loading values for individual quinone species in tidal sediment with (A) andwithout (B) clams.
out clams, quinone profiles had positive correlationwith PC1 (0.69) and PC2 (0.58). In this month, theD(with–without clams) value increased slightly to 0.40.This indicates that the effect of clams on the changesof microbial community structure was influenced byenvironmental condition such as temperature, DO andorganic matter concentration.
In subtidal sediment from July to August, changesin microbial community structure had the same patternwith and without clams. This indicates that the effectof clams on changes in microbial community structurewere relatively low during this period. This patternwas consistent with the changes inD(with–without clams)value. TheD(with–without clams) value only increasedfrom 0.09 to 0.12. The sharp change in pattern oc-curred from August to October. In September, quinoneprofiles in the sediment with clams had high posi-tive (0.64) and low negative (−0.46) correlations withPC1 and PC2, respectively. In the sediment withoutclams, the quinone profiles had high and low posi-tive correlations with PC1 (0.93) and PC2 (0.25), re-spectively. From September to October, the changesof principal component value in the sediment with andwithout clams were more evident. Quinone profiles

194 U. Hasanudin et al. / Ecological Engineering 22 (2004) 185–196
Fig. 9. Principal component analysis showing differences in loading values for individual quinone species in subtidal sediment with (C)and without (D) clams.
in the sediment with clams had high positive correla-tion with PC1 (0.78) and low negative correlation withPC2 (−0.44). While in the sediment without clams,quinone profiles had low and high positive correlationwith PC1 (0.49) and PC2 (0.82), respectively. Thisresult consistent with theD(with–without clams) value insubtidal sediment, which also increased sharply fromAugust to September and from September to October(Fig. 6).
From an engineering standpoint, these results arevery important for optimizing the contribution ofclams to tidal flat waters purification functions. Thecontribution of clams is greatest in tidal sediment atthe end of summer (September).
Figs. 8 and 9show the differences of loading val-ues of PCA for individual quinone species of tidal andsubtidal sediment with and without clams. We proposethat the distant between loading values for individ-ual quinone species in the sediment with and withoutclams in PC1–PC2 plots can be used as indicator of thedifferences between quinone species profile. Based onthe distant, the microbial community changes in tidalsediment were mainly due to changes in UQ-10, UQ-9,UQ-8, MK-7 (H8) and MK-9 (H2) profiles. Whilein subtidal sediment were mainly due to changes in
UQ-9, UQ-10, UQ-8, MK-9 (H2) and MK-10 profiles.This indicates that the changes of quinone profiles intidal and subtidal sediments were caused by almostsimilar species. Its mean that the effects of clam onchange in microbial community structure in tidal andsubtidal sediments were might be only different in themagnitude of effect.
4. Conclusions
Clams appear to be suitable organisms to acceler-ate the decomposition of organic matter in tidal flatsediment. Clam activities increased DOC and NH4-Nconcentration in tidal sediment. However, the effectof clams in subtidal flat sediment was relatively low;DOC and NH4-N concentrations in the sediment werealmost identical.
The effect of clams on changes in microbialcommunity structure in tidal sediment was higherthan in subtidal sediment. Microbial communityshift in tidal sediment was greater than in subtidalsediment. The differences in microbial communitystructure in tidal sediment with and without clamswere mainly due to changes in UQ-10, UQ-9, UQ-8,

U. Hasanudin et al. / Ecological Engineering 22 (2004) 185–196 195
MK-7 (H8) and MK-9 (H2)-containing bacterial pro-files. Changes in UQ-9, UQ-10, UQ-8, MK-9 (H2)and MK-10-containing bacterial profiles dominatedchanges in microbial community structure in subtidalsediment.
The utilization of clams to improve tidal flat purifi-cation function needed consideration on environmen-tal conditions. Temperature, DO, immersion–emersioncycle and concentration of biodegradable organic mat-ter in the sediment are potential factors to enhanceclam and microbial activities.
Acknowledgements
The authors thank Mr. Kazuya Takeda and Mr.Kazuhiro Shimada for technical assistance and advice.This study was supported financially by 21st CenturyCOE (Center of Excellence) Project, “Ecological En-gineering for Homeostatic Human Activities”, fundedby the Ministry of Education, Culture, Sport, Scienceand Technology, Japan.
References
Collins, D.M., Jones, D., 1981. Distribution of isoprenoid quinonestructural types in bacteria and their taxonomic implications.Microbiol. Rev. 45 (2), 316–354.
Durand, F., Peters, L.D., Livingstone, D.R., 2002. Effect ofintertidal compared to subtidal exposure on the uptake, loss,and oxidative toxicity of water born benzo[a]pyrene in themantle and whole tissues of musselMytilus edulis L. Mar.Environ. Res. 54, 271–274.
Fujie, K., Hu, H.Y., Tanaka, H., Urano, K., 1994. Ecological studiesof aerobic submerged biofilter on the basis of respiratoryquinone profiles. Water Sci. Technol. 29, 373–376.
Fujie, K., Hu, H.Y., Tanaka, H., Urano, K., Saito, K., Katayama,A., 1998. Analysis of respiratory quinone profile in soil forcharacterization of microbiota. Soil Sci. Plant Nutr. 44, 393–404.
Hendrick, I.A., White, D.C., 1986. Microbial respiratory quinonesin the environment. J. Microbiol. Meth. 5, 243–254.
Hiraishi, A., 1988. Respiratory quinone profiles as a tools foridentifying different bacterial populations in activated sludge.J. Gen. Appl. Microbiol. 34, 39–56.
Hiraishi, A., Morishima, Y., Takeuchi, J., 1991. Numerical analysisof lipoquinone pattern in monitoring bacterial in wastewatertreatment systems. J. Gen. Appl. Microbiol. 37, 57–70.
Hiraishi, A., Ueda, Y., Ishihara, J., 1998. Quinone profiling ofbacterial communities in natural and synthetic sewage activated
sludge for enhanced phosphate removal. Appl. Environ.Microbiol. 64, 992–998.
Hiraishi, A., 1999. Isoprenoid quinones as biomarkers of microbialpopulations in the environment. J. Biosci. Bioeng. 88, 449–460.
Hu, H.Y., Nozawa, M., Fujie, K., Makabe, T., Urano, K., 1996.Studies of microbial acclimation to hard chemicals on the basisof respiratory quinone profiles and kinetic analyses. Water Sci.Technol. 34, 249–256.
Hu, H.Y., Fujie, K., Tanaka, H., Makabe, T., Urano, K., 1997.Respiratory quinone profile as a tool for refractory chemicalbiodegradation study. Water Sci. Technol. 35, 103–110.
Hu, H.Y., Fujie, K., Urano, K., 1999a. Development of a novelsolid phase extraction method for analysis of bacteria quinonesin activated sludge with higher reliability. J. Biosci. Bioeng.87, 378–382.
Hu, H.Y., Fujie, K., Nakagome, H., Urano, K., Katayama, A.,1999b. Quantitative analyses of the change in microbialdiversity in a bioreactor for wastewater treatment based onrespiratory quinones. Water Res. 33, 3263–3270.
Hu, H.Y., Lim, B.R., Goto, N., Fujie, K., 2001. Analytical precisionand repeatability of respiratory quinones for quantitative studyof microbial community structure in environmental samples.J. Microbiol. Meth. 47, 17–24.
King, M.G., 1986. Characterization of�-glucosidase activity inintertidal marine sediment. Appl. Environ. Microbiol. 51, 373–380.
Kroger, A., 1978. Determination of contents and redox statesof ubiquinone and menaquinone. Meth. Enzymol. 53, 579–591.
Kunihiro, T., Hu, H.Y., Lim, B.R., Goto, N., Fujie, K., 2002.Analysis of the differences in microbial community structuresbetween suspended and sessile microorganisms in rivers basedon quinone profile. J. Gen. Appl. Microbiol. 48, 35–41.
Kuwae, T., Kibe, E., Nakamura, Y., 2003. Effect of emersion andimmersion on the porewater nutrient dynamics of an intertidalsandflat in Tokyo Bay. Est. Coast. Shelf Sci. 57, 929–940.
Lee, J.G., Nishijima, W., Mukai, T., Takimoto, K., Seiki, T.,Hiraoka, K., Okada, M., 1998. Factors to determine thefunctions and structures in natural and constructed tidal flats.Water Res. 32, 2601–2606.
Megalhães, M.C., Bordalo, A.A., Wiebe, J.W., 2003. Intertidalbiofilms on rocky substratum can play a major role in estuarinecarbon and nutrient dynamics. Mar. Ecol. Prog. Ser. 258, 275–281.
Ogilvie, B., Nedwell, D.B., Harrison, R.M., Robinson, A., Sage,A., 1997. High nitrate, muddy estuaries as nitrogen sinks: thenitrogen budget of the River Colne estuary (United Kingdom).Mar. Ecol. Prog. Ser. 150, 217–228.
Rutters, H., Sass, H., Cypioka, H., Rullkotter, J., 2002. Microbialcommunities in Wadden Sea sediment core-clues from analysesof intact glyceride lipids, and released fatty acids. Org.Geochem. 33, 803–816.
Sohma, A., Sekiguchi, Y., Yamada, H., Sato, T., Nakata, K., 2001.A new coastal marine ecosystem model study coupled withhydrodynamics and tidal flat ecosystem effect. Mar. Poll. Bull.43, 187–208.

196 U. Hasanudin et al. / Ecological Engineering 22 (2004) 185–196
Sundback, K., Miles, A., Goransson, A., 2000. Nitrogen fluxes,denitrification and the role of microphytobenthos in microtidalshallow-water sediments: an annual study. Mar. Ecol. Prog.Ser. 200, 59–76.
Suzuki, T., Aoyama, H., Kai, M., 1998. The effect of dissolvedoxygen deficiency on dead clam (Ruditapes philippinarum)formation in Mikawa bay. J. Adv. Mar. Sci. Tech. Soc. 4, 35–40 (in Japanese).
Tamaoka, J., Katayama-Fujimura, Y., Kuraishi, H., 1983. Analysisof bacterial menaquinone mixture by high performancechromatography. J. Appl. Bacteriol. 54, 31–36.
Ueda, N., Tsutsumi, H., Yamada, M., Hanamoto, K., 2000. Impactsof oxygen-deficient water on the macrobenthic fauna of DokaiBay and on adjacent intertidal flats, in Kitakyushu. Jpn. Mar.Poll. Bull. 40, 906–913.
Usui, T., Koike, I., Ogura, N., 2001. N2O production, nitrificationand denitrification in an estuarine sediment. Est. Coast. ShelfSci. 52, 769–781.
Zhu, S., Saucier, B., Durfey, J., Chen, S., Dewey, B., 1999. Wasteexcretion characteristics of Manila clams (Tapes philippinarum)under different temperature conditions. Aquacult. Eng. 20, 231–244.




















