
Vibrio cortegadensis sp. nov., isolated from clams
27
Antonie van Leeuwenhoek Journal of Microbiology Vibrio cortegadensis sp. nov., isolated from clams --Manuscript Draft-- Manuscript Number: ANTO-D-13-00264R3 Full Title: Vibrio cortegadensis sp. nov., isolated from clams Article Type: Original Research Paper Keywords: Vibrio; V. cortegadensis sp. nov.; MLSA; DDH Corresponding Author: Jesús L. Romalde, Ph.D. Univ. Santiago de Compostela Santiago de Compostela, SPAIN Corresponding Author Secondary Information: Corresponding Author's Institution: Univ. Santiago de Compostela Corresponding Author's Secondary Institution: First Author: Aide Lasa, M.Sc. First Author Secondary Information: Order of Authors: Aide Lasa, M.Sc. Ana L Diéguez, M.Sc. Jesús L. Romalde, Ph.D. Order of Authors Secondary Information: Abstract: A group of four strains isolated from clams (Venerupis decussata and V. philippinarum) in Galicia (NW Spain) were subjected to a polyphasic characterization, based on the phenotypic characteristics, the analysis of chemotaxonomic features, the sequencing of the 16S rRNA and five housekeeping (atpA, pyrH, recA, rpoA and rpoD) genes, as well as DNA-DNA hybridization (DDH). The analysis of the phenotypic and chemotaxonomic characteristics and the results of a phylogenetic study, based on the 16S rRNA gene sequence analysis and multilocus sequence analysis (MLSA), clearly indicated that these strains belong to the genus Vibrio and were allocated between the Splendidus and Anguillarum clades showing a close relationship with the type strains of V. tapetis (98.8%), V. pomeroyi (98.0%) and V. crassostreae (97.9%). DDH results confirmed that these isolates constitute a new species. The name Vibrio cortegadensis sp. nov. is proposed with C 16.17T (= CECT 7227T = LMG 27474T) as the type strain. Powered by Editorial Manager® and ProduXion Manager® from Aries Systems Corporation
Transcript of Vibrio cortegadensis sp. nov., isolated from clams

Antonie van Leeuwenhoek Journal of Microbiology
Vibrio cortegadensis sp. nov., isolated from clams--Manuscript Draft--
Manuscript Number: ANTO-D-13-00264R3
Full Title: Vibrio cortegadensis sp. nov., isolated from clams
Article Type: Original Research Paper
Keywords: Vibrio; V. cortegadensis sp. nov.; MLSA; DDH
Corresponding Author: Jesús L. Romalde, Ph.D.Univ. Santiago de CompostelaSantiago de Compostela, SPAIN
Corresponding Author SecondaryInformation:
Corresponding Author's Institution: Univ. Santiago de Compostela
Corresponding Author's SecondaryInstitution:
First Author: Aide Lasa, M.Sc.
First Author Secondary Information:
Order of Authors: Aide Lasa, M.Sc.
Ana L Diéguez, M.Sc.
Jesús L. Romalde, Ph.D.
Order of Authors Secondary Information:
Abstract: A group of four strains isolated from clams (Venerupis decussata and V. philippinarum)in Galicia (NW Spain) were subjected to a polyphasic characterization, based on thephenotypic characteristics, the analysis of chemotaxonomic features, the sequencingof the 16S rRNA and five housekeeping (atpA, pyrH, recA, rpoA and rpoD) genes, aswell as DNA-DNA hybridization (DDH). The analysis of the phenotypic andchemotaxonomic characteristics and the results of a phylogenetic study, based on the16S rRNA gene sequence analysis and multilocus sequence analysis (MLSA), clearlyindicated that these strains belong to the genus Vibrio and were allocated between theSplendidus and Anguillarum clades showing a close relationship with the type strainsof V. tapetis (98.8%), V. pomeroyi (98.0%) and V. crassostreae (97.9%). DDH resultsconfirmed that these isolates constitute a new species. The name Vibrio cortegadensissp. nov. is proposed with C 16.17T (= CECT 7227T = LMG 27474T) as the type strain.
Powered by Editorial Manager® and ProduXion Manager® from Aries Systems Corporation

Dr. Elena Ivanova
Editor
Antonie van Leeuwenhoek Journal of Microbiology.
Santiago de Compostela, November 11, 2013
Dear Dr. Ivanova,
Thank you very much for the language edition on the ANTO-D-13-00264R2 by Lasa et
al., entitled " Vibrio cortegadensis sp. nov. isolated from clams. We are submitting a
new revised version, which was modified taking into account the suggestions of the
attached file sent.
Sincerely,
Dr. Jesus L. Romalde
*Cover LetterClick here to download Cover Letter: Coverletter.doc

Dr. Elena Ivanova
Editor
Antonie van Leeuwenhoek Journal of Microbiology.
Santiago de Compostela, November 7, 2013
Dear Dr. Ivanova,
Thank you very much for the language edition on the ANTO-D-13-
00264R2 by Lasa et al., entitled " Vibrio cortegadensis sp. nov. isolated
from clams. We are submitting a new revised version, which was modified
taking into account the suggestions of the attached file sent.
Yours sincerely,
Dr. Jesus l. Romalde
*Response to reviewers' commentsClick here to download Response to reviewers' comments: Letter to editor.docx

1
1
2
3
Vibrio cortegadensis sp. nov., isolated from clams 4
5
Aide Lasa, Ana L. Diéguez, and Jesús L. Romalde* 6
7
Departamento de Microbiología y Parasitología, CIBUS. Universidad de Santiago de 8
Compostela. Campus Sur s/n. 15782. Santiago de Compostela. Spain. 9
10
11
12
13
14
15
Running title: Vibrio cortegadensis sp. nov. 16
17
18
Submitted to: Antoine Van Leeuwenhoek, November 2013 19
Revised Ms. ANTO-D-13-00264R2 20
21
22
23
24
25
26 *Corresponding author: 27
Telf.: (+34) 981 563100 ext. 16961 28
Fax: (+34) 981 528085 29
E–mail: [email protected] 30
31
*ManuscriptClick here to download Manuscript: Lasa et al EI edits 9.11.13_rev.doc Click here to view linked References

2
ABSTRACT 32
A group of four strains isolated from clams (Venerupis decussata and V. philippinarum) 33
in Galicia (NW Spain) were subjected to a polyphasic characterization, based on the 34
phenotypic characteristics, the analysis of chemotaxonomic features, the sequencing of 35
the 16S rRNA and five housekeeping (atpA, pyrH, recA, rpoA and rpoD) genes, as 36
well as DNA–DNA hybridization (DDH). The analysis of the phenotypic and 37
chemotaxonomic characteristics and the results of a phylogenetic study, based on the 38
16S rRNA gene sequence analysis and multilocus sequence analysis (MLSA), clearly 39
indicated that these strains belong to the genus Vibrio and were allocated between the 40
Splendidus and Anguillarum clades showing a close relationship with the type strains of 41
V. tapetis (98.8%), V. pomeroyi (98.0%) and V. crassostreae (97.9%). DDH results 42
confirmed that these isolates constitute a new species. The name Vibrio cortegadensis 43
sp. nov. is proposed with C 16.17T (= CECT 7227
T = LMG 27474
T) as the type strain. 44
45
46
Keywords: Vibrio, V. cortegadensis sp. nov., MLSA, DDH 47
48
49
50
51
52
53
54
55
56
57
58

3
INTRODUCTION 59
60
The genus Vibrio comprises of a large number of species that are common inhabitants 61
of aquatic environments such as estuarine, coastal waters and sediments (Colwell 2006; 62
Thompson and Swings 2006). Several species of this genus have been associated with 63
marine eukaryotic organisms including fish, molluscs and crustaceans (Beaz–Hidalgo et 64
al. 2010) and, in addition, some of them have been described as pathogens to fish, 65
molluscs and crustaceans (Farto et al. 2003; Gay et al. 2004; Gómez–León et al. 2005; 66
Jensen et al. 2003; Kueh and Chan 1985; Lacoste et al. 2001; Le Roux et al. 2002; 67
Leano et al. 1998; Nicolas et al. 1996; Pujalte et al. 1993; Sugumar et al. 1998). 68
The increasing number of environmental studies and the introduction of molecular 69
techniques in bacterial taxonomy, such as DNA–DNA hybridization (DDH), multilocus 70
sequence analysis (MLSA) and amplified fragment length polymorphism (AFLP) 71
(Beaz–Hidalgo et al. 2008; Colwell 2006; Thompson and Swings 2006), have enhanced 72
the understanding of the family Vibrionaceae taxonomic structure and phylogeny. 73
Nowadays, there are 98 validly described species of the genus Vibrio, including two 74
subspecies (http://www.vibriobiology.net), that have been grouped into 14 clades 75
(Sawabe et al. 2007). The large number of the species described in the last 6 years, 76
together with the proposal of the new clades (i.g. Marisflavi and Comitans)(Pujalte, 77
2011), have led to an update of the intra-genus classification (Gomez-Gil, personnal 78
communication). 79
In a previous study on the diversity of vibrios conducted in 2004 and 2005, a collection 80
of isolates were obtained from reared clams, Manila clam (Venerupis philippinarum) 81
and carpet–shell clam (V. decussata), aquacultured in different geographical sites of the 82
coast of Galicia (NW Spain). A representative number of isolates of this collection were 83
analysed by AFLP and a group of four strains (cluster 68) could not be assigned to any 84
of the currently known species of the genus Vibrio (Beaz–Hidalgo et al. 2008). In the 85
present study, a polyphasic approach was employed for the characterization of the 86
cluster of four strains isolated from the clams. 87
88
89

4
MATERIALS AND METHODS 90
91
Bacterial isolates 92
Bacterial strains C 16.17T (=CECT 7227
T= LMG 27474
T), CMJ 9.12 (=CECT 8125= 93
LMG 27475), CMJ 12.11 (=LMG 27477) and Rd 13.7(= LMG 27476) were 94
corresponded to the clam isolates of cluster 68 of Beaz–Hidalgo et al. (2008). Strain C 95
16.17T was isolated from Ria de Arousa (42º 37´3” N/ 8º 46´ 38” W), strains CMJ 9.12 96
and CMJ 12.11 were isolated from Ria de Camariñas (43º 8´16” N/ 9º 10´37” W) and 97
strain Rd 13.7 was isolated from Ria de Vigo (42º 18´27” N/ 8º 37´13” W). The four 98
isolates were characterized in comparison with the following reference strains obtained 99
from bacterial culture collections: V. atlanticus CECT 7223T, V. artabrorum CECT 100
7226T, V. celticus CECT 7224
T, V. chagasii LMG 21353
T, V. crassostreae CAIM 101
1405T, V. cyclitrophicus LMG 21359
T, V. gallaecicus CECT 7244
T, Vibrio gigantis 102
LMG 22741T, V. kanaloae LMG 20539
T, V. lentus CECT 5110
T, V. pomeroyi LMG 103
20537T, V. splendidus CECT 628
T, V. tasmaniensis LMG 20012
T, V. tapetis CECT 104
4600T
and V. anguillarum ATCC 19264T. All strains were cultured on plates of Marine 105
agar (MA, Difco) at 24±1 ºC for 24 h. Stock cultures were maintained frozen at –80 ºC 106
in Marine broth (MB, Pronadisa, Spain) supplemented with 15% of glycerol (v/v). 107
108
Phenotypical characterization 109
The four marine strains were subjected to the following phenotypic tests (MacFaddin 110
1993; Romalde and Toranzo 1991): cell morphology and motility, Gram stain, oxidase, 111
catalase, oxidation/fermentation test, fermentation and acid production from inositol, 112
mannitol and sucrose, gas and acid production from glucose, indole, methyl red, Voges–113
Proskauer reaction, utilization of citrate, arginine dihydrolase test (Moeller’s medium), 114
lysine and ornithine decarboxylation (Moeller’s medium), nitrate reduction, hydrolysis 115
of gelatin, Tween 80, amylase and aesculin. Salt tolerance test was performed on Basal 116
medium agar (BMA, neopeptone [4 g/l], yeast extract [1 g/l], bacteriological agar [15 117
g/l]) supplemented with 0, 0.5, 1, 3, 6, 8 and 10% NaCl. Growth at different 118
temperatures (4, 20, 25, 30, 37 and 44 ºC), pH (4-10), and on thiosulfate–citrate–bile 119
sucrose (TCBS) agar (Oxoid) were also determined. Sensitivity to the vibriostatic agent 120
O/129 (2,4–diamino–6,7–diisopropylpteridine) (150 µg per disc) was determined on 121
Müeller–Hinton (Oxoid) agar. All media were supplemented with 1% NaCl when 122
required. 123

5
Additional phenotypic characteristics were performed using API 20 NE, API 50CH and 124
API ZYM miniaturized systems (BioMerieux, France) using Saline solution (SS, 0.85% 125
NaCl) to prepare the bacterial suspensions. API 50CH was used with the slight 126
modifications described by Prado et al. (2005). Briefly, bacterial suspensions were 127
prepared in SS, adjusted to an OD580 of 1.0 and mixed (1:10, v/v) with ZOF medium 128
(without agar) (Lemos et al. 1985) for the inoculation of the strips. With the exception 129
of the growth at different temperatures, all phenotypic tests were performed at 24±1 ºC. 130
131
16S rRNA and housekeeping genes sequencing 132
Genomic DNA for sequencing was obtained as described previously (Osorio et al. 133
1999). Amplification and sequencing of the 16S rRNA gene and the housekeeping 134
genes atpA (ATP synthase alpha subunit gene), recA (recombinase A gene), pyrH 135
(uridine monophosphate kinase gene), rpoA (RNA polymerase alpha subunit gene) and 136
rpoD (RNA polymerase sigma factor gene) were performed according to Thompson et 137
al. (2004, 2005, 2007) and Pascual et al. (2010). For reference strains, sequences were 138
acquired from GenBank/EMBL/DDBJ. Sequence analyses were performed using the 139
DNASTAR Lasergene SEQMAN program. Sequence similarities of 16S rRNA and 140
housekeeping genes were determined using the EzTaxon-e server (www.eztaxon-141
e.ezbiocloud.net; Kim et al. 2012) and the BLASTN program respectively. Sequences 142
were aligned using CLUSTAL W tool (Larkin et al. 2007), and phylogenetic trees were 143
reconstructed using the neighbour-joining and maximum-likelihood algorithms (MEGA 144
version 5.05) (Tamura et al. 2011). Distance matrices were calculated by using 145
Kimura’s two-parameter correction and stability of the groupings was estimated by 146
bootstrap analysis (1000 replicates) using the MEGA version 5.0 (Tamura et al., 2011). 147
148
DNA–DNA hybridization (DDH) 149
Genomic DNA for DDH experiments was extracted using the commercial DNeasy 150
Blood & Tissue kit (QIAGEN), following the manufacturer´s protocol. DDH 151
experiments were undertaken between the strain C16.17T and the type strains of the 152
species with highest similarities in the 16S rRNA gene (V. tapetis, V. pomeroyi and V. 153
crassostreae) and a representative of the Anguillarum clade (V. anguillarum). DDH 154
experiments were performed with the hydroxyapatite/microtitre plate method (Ziemke 155
et al. 1998) using a hybridization temperature (Tm) of 60 ºC. Reciprocal reactions (i.e. 156

6
A×B and B×A) were performed and were generally within the limits of this method 157
(Goris et al. 1998). 158
159
Fatty acids analysis 160
Chemotaxonomic features were studied by the analyses of fatty acid methyl esters 161
(FAME). FAME were extracted and prepared from 24 h cultures on MA incubated at 162
24±1ºC as described by Sasser et al. (1990) according to the MIDI Microbial 163
Identifications System (MIDI, Newark, DE, USA). The two closest species in the 164
MLSA were analysed in parallel for comparison. 165
166
RESULTS AND DISCUSSION 167
168
The four marine strains were facultative anaerobic, motile, Gram–negative rods. 169
Positive for oxidase production and reduction of nitrates to nitrites, only one of the 170
strains (CMJ 12.11) was positive in the catalase test. They required salt for growth 171
(optimal range 1-3%), were able to grow at 4 ºC but not at 37 or 44ºC (optimal range 172
20-25ºC). Optimal pH was found to be in the range of 6 to 9. The strains C 16.17T and 173
CMJ 9.12 grew on thiosulfate–citrate–bile–sucrose agar (TCBS, Oxoid) but not the 174
strains CMJ 12.11 and Rd 13.7. They were sensitive to the vibriostatic agent O/129. 175
Differentiating phenotypic features for the four marine strains are shown in the Table 176
S1. 177
Sequence similarity of the 16S rRNA gene indicated that the isolates belonged to the 178
genus Vibrio. Isolate C16.17T showed highest sequence similarities with the species V. 179
tapetis (98.7%), V. pomeroyi (98.0%), and V. crassostreae (97.9%). Phylogenetic 180
analysis based on 16S rRNA gene sequences of the isolates, employing both NJ or ML 181
approaches, showed that the four marine strains present an intermediate position 182
between the representatives of Splendidus and Anguillarum clades (Figures 1 and S1). 183
Multilocus sequence analysis (MLSA) of housekeeping genes has been proposed as a 184
useful tool to define the phylogenetic relationships among microorganisms 185
(Stackebrandt and Ebers, 2006). In the genus Vibrio, several genes have been studied 186
for delineating new species, such as gyrB, atpA, recA, pyrH or dnaJ (Pascual et al. 187
2010; Sawabe et al. 2007; Thompson et al. 2004, 2005, 2007). In this study, sequences 188
of the genes atpA (1300 bp), pyrH (575 bp), recA (765 bp), rpoA (875 bp) and rpoD 189
(842 bp) were obtained for the clam isolates and compared with the closest relatives. 190

7
Each housekeeping gene pointed different species of the genus Vibrio as the closest 191
relative, with similarity values lower than 93% in all cases (Figure S2). Phylogenetic 192
trees based on each housekeeping gene and on concatenated sequences of the five 193
housekeeping genes, not only showed that the four clam isolates form a tight group, but 194
also suggested a closer relationship with the Splendidus clade (Figures 2, S2 and S3). 195
Further studies will confirm the inclusion of this group of isolates in a specific clade of 196
the genus. The GenBank accession numbers for the 16S rRNA, atpA, pyrH, recA, rpoA 197
and rpoD gene sequences obtained for the four clam strains are listed in Supplementary 198
Table S2. 199
The type strain C16.17T showed levels of DNA relatedness of 53% (reciprocal 45%) 200
with V. tapetis CECT 4600T, 41% (38%) with V. pomeroyi LMG 20537
T, 41% (44%) 201
with V. crassostreae CAIM 1405T, and 35% (39%) with V. anguillarum ATCC 19264
T. 202
All these wDDH values were below 70%, threshold to delimit species. On the other 203
hand, the four clam isolates showed DDH values of at least 80% (data not shown). 204
These results demonstrated that the four clam strains represent a novel species within 205
the genus Vibrio. 206
The isolates from cultured clam can be differentiated from the phylogenetically related 207
species of the genus Vibrio, by several phenotypic features (Table 1). Strains can be 208
distinguished from V. tapetis by their ability in the arginine dyhydrolsis, the 209
fermentation of glycerol and D–maltose, and the lack of the fermentation of D–mannitol 210
and amygdalin. At the same time, the clam isolates can be differentiated from V. 211
pomeroyi by their ability in fermentation of glycerol and inability in the fermentation of 212
D–mannitol. These strains can be differentiated from V. crassostreae by their inability 213
to grow at 6% of NaCl, the fermentation of D–maltose and inability in the fermentation 214
of D–mannitol and amygdalin. In addition, the analyses of FAMEs of the type strain 215
C16.17T
showed its distinct FA profile (Table S3). 216
217
The analysis of the polyphasic study clearly indicated that strains of the cluster 68 218
represent a new taxon within the genus Vibrio. The name Vibrio cortegadensis sp. nov. 219
is proposed for this new species. 220
221

8
222
Description of Vibrio cortegadensis sp. nov. 223
Vibrio cortegadensis [cor.te.ga.den´.sis. N.L. gen. n. cortegadensis intended to mean 224
that the type strain was isolated from Cortegada Island in Carril (Galicia, Spain)] 225
226
Gram-negative motile rods with facultative anaerobic metabolism. All strains are 227
sensitive to the vibriostatic agent O/129, positive for arginine dihydrolase (Moeller’s 228
medium), indole reaction, production of oxidase, lipase and amylase, and for the 229
reduction of nitrates to nitrites. They are negative for the decarboxylation of lysine and 230
ornithine, Voges–Proskauer reaction, utilization of citrate, and the hydrolysis of 231
aesculin and urea. Variable reaction was observed for the catalase test, showing only 232
CMJ 9.12 strain a positive reaction. Strain Rd 13.7 is positive for ONPG test. The 233
ability to grow on TCBS is variable, being strains C16.17T and CMJ 9.12 able to grow 234
on this medium as green colonies (sucrose negative). The four isolates showed growth 235
from 1 to 3% NaCl, but not in the absence of NaCl or at salinities higher than 6% NaCl. 236
The strains are able to grow from 4 to 30ºC, but not at 37 ºC and 44 ºC. 237
All strains produce fermentation of D–glucose, D–fructose, N–acetylglucosamine, 238
aesculin, D–maltose, D–trehalose, starch, glucogen and potassium 2–ketogluconate 239
(weakly reaction), but not of erythritol, D–arabinose, L–xylose, D–adonitol, methyl–240
βD–xylopyranoside, L–sorbose, L–rhamnose, dulcytol, inositol, D–sorbitol, methyl–241
αD–mannopyranoside, methyl–αD–glucopyranoside, amygdalin, arbutin, salicin, D–242
cellobiose, D–lactose, D–melibiose, D–sucrose, inuline, D–melezitose, D–raffinose, 243
xylitol, gentiobiose, D–turanose, D–lyxose, D–tagatose, D–fucose, L–fucose, D–244
arabitol, L–arabitol, potassium gluconate and potassium 5–ketogluconate. In the API 245
ZYMsystem, all strains show positive reactions for alkaline phosphatase, esterase, 246
esterase lipase, leucine arylamidase, valine arylamidase, acid phosphatase, naftol–AS–247
BI–phosphohydrolase. The major fatty acids of the type strain C16.17T are C12:0 3OH 248
(4.3%), C16:0 (22.6%), summed feature in 3 (comprising C16:1 ω7c and/or C16:1 ω6c) 249
(47.9%), and summed feature 8 (comprising C18:1 ω7c and/or C18:1 ω6c)(9.9%). 250
The type strain C16.17T (=CECT 7227
T= LMG 27474
T) was isolated in the north–251
western coast of Spain (Galicia), from healthy cultured clams, Venerupis decussata. 252
Isolates CMJ 9.12 (=CECT 8125= LMG 27475), CMJ 12.11 (=LMG 27427) and Rd 253
13.7(= LMG 27476) were also deposited at culture collections as reference strains of the 254
species. 255

9
256
Acknowledgements 257
This work was supported in part by grant AGL–2010–18438 from the Ministerio de 258
Ciencia e Innovación (Ministry of Science and Innovation) (Spain). A. L. acknowledges 259
the Ministerio de Economía y Competitividad (Ministry of Economy and 260
Competitiveness) (Spain) for a research fellowship. 261
262
Conflict of interest 263
The authors declare that they have no conflict of interest. 264
265
266

10
REFERENCES 267
268
Beaz Hidalgo R, Cleenwerck I, Balboa S, De Wachter M, Thompson FL, Swings J, de 269
Vos P, Romalde JL (2008) Diversity of Vibrios associated with reared clams in 270
Galicia (NW Spain). Syst Appl Microbiol 3:215–222. 271
Beaz–Hidalgo R, Balboa S, Romalde JL, Figueras MJ (2010a). Diversity and 272
pathogenicity of Vibrio species in cultured bivalve molluscs. Environ Microbiol 273
Reports 2:34–43. 274
Colwell RR (2006) A global and historical perspective of the genus Vibrio. In: 275
Thompson FL, Austin B, Swings J. (Eds.), The Biology of Vibrios. American 276
Society for Microbiology, Washington, DC, pp. 3–11. 277
Farto R, Armada SP, Montes M, Guisande JA, Pérez MJ, Nieto TP (2003) Vibrio lentus 278
associated with diseased wild octopus (Octopus vulgaris). J Invertebr Pathol 83:149–279
156. 280
Gay M, Renault T, Pons AM, Le Roux F (2004) Two Vibrio splendidus related strains 281
collaborate to kill Crassostrea gigas: taxonomy and host alterations. Dis Aquat Org 282
62:65–74. 283
Gómez–León J, Villamil L, Lemos ML, Novoa B, Figueras A (2005) Isolation of Vibrio 284
alginolyticus and Vibrio splendidus from aquacultured carpet shell clam (Ruditapes 285
decussatus) larvae associated with mass mortalities. Appl Environ Microbiol 71:98–286
104 287
Goris J, Suzuki K, de Vos P, Nakase T, Kersters K (1998) Evaluation of a microplate 288
DNA–DNA hybridization method compared with the initial renaturation method. 289
Can J Microbiol 44:1148–1153. 290
Jensen S, Samuelsen OB, Andersen K, Torkildsen L, Lambert C, Choquet G, Paillard C 291
Bergh O (2003) Characterization of strains Vibrio splendidus and V. tapetis isolated 292
from corkwing wrasse Symphodus melops suffering vibriosis. Dis Aquat Org 53:25–293
31. 294
Kim OS, Cho YJ, Lee K, Yoon SH, Kim M, Na H, Park SC, Jeon YS, Lee JH, Yi H, 295
Won S, Chun J (2012). Introducing EzTaxon-e: a prokaryotic 16S rRNA Gene 296
sequence database with phylotypes that represent uncultured species. Int J Syst Evol 297
Microbiol 62:716-721. 298
Kueh CSW, Chan KY (1985) Bacteria in bivalve shellfish with special reference to the 299
oyster. J Appl Bacteriol 59:41–47. 300

11
Lacoste A, Jalabert F, Malham S, Cueff A, Gélébart F, Cordevant C, Lange M, Poulet 301
SA (2001) A Vibrio splendidus strain is associated with summer mortality of juvenile 302
oysters Crassotsrea gigas in the Bay of Morlaix (North Britanny, France). Dis Aquat 303
Org 46:139–145. 304
Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, 305
Valentin F, Wallace IM, Wilm A, Lopez R, Thompson JD, Gibson TJ, Higgins DG 306
(2007) Clustal W and Clustal X version 2.0. Bioinformatics 23:2947-2948. 307
Le Roux F, Goubet A, Thompson FL, Faury N, Gay M, Swings J, Saulnier D (2005) 308
Vibrio gigantis sp. nov., isolated from the haemolymph of cultured oysters 309
(Crassostrea gigas). Int J Syst Evol Microbiol 55:2251–2255. 310
Leano EM, Lavilla–Pitogo CR, Paner MG (1998) Bacterial flora in the hepatopancreas 311
of pond reared Penaeus monodon juveniles with luminous vibriosis. Aquaculture 312
164:367–374. 313
Lemos ML, Toranzo AE, Barja JL (1985) Modified medium for the oxidation–314
fermentation test in the identification of marine bacteria. Appl Environ Microbiol 315
49:1541–1543. 316
MacFaddin JF (1993) Pruebas bioquímicas para la Identificación de Bacterias de 317
Importancia Clínica (translation by Médica Panamericana SA). Williams & Wilkins, 318
Baltimore, MD (In Spanish). 319
Nicolas JL, Corre S, Gauthier G, Robert R, Ansquer D (1996) Bacterial problems 320
associated with scallop Pecten maximus larval culture. Dis Aquatic Org 27:67–76. 321
Osorio CR, Collins MD, Toranzo AE, Romalde JL (1999) 16S rRNA gene sequence 322
analysis of Photobacterium damselae and nested–PCR method for rapid detection of 323
fish pasteurellosis. Appl Environ Microbiol 65: 2942–2946. 324
Pascual J, Macián MC, Arahal DR, Garay E, Pujalte MJ (2010) Multilocus sequence 325
analysis of the central clade of the genus Vibrio by using the 16S rRNA, recA, pyrH, 326
rpoD, gyrB, rctB and toxR genes. Int J Syst Evol Microbiol 60:154–165. 327
Prado S, Romalde JL, Montes J, Barja JL (2005). Pathogenic bacteria isolated from 328
diseased outbreaks in shellfish hatcheries. First description of Vibrio neptunius as an 329
oyster pathogen. Dis Aquat Org 67:209-215. 330
Pujalte MJ (2011). La familia Vibrionaceae: una actualización taxonómica. XXIII 331
Congreso Nacional de la Sociedad Española de Microbiología . Salamanca 332
Pujalte MJ, Ortigosa M, Urdaci MC, Garay E, Grimont PAD (1993) Vibrio mytili sp. 333
nov., from mussels. Int Syst Bacteriol 43:358–362. 334

12
Romalde JL, Toranzo AE (1991) Evaluation of the API 20E system for the routine 335
diagnosis of the enteric redmouth disease. Bull Eur Assoc Fish Pathol 11:147–149. 336
Sasser M (1990) Identification of bacteria by gas chromatography of cellular fatty acids. 337
Microbial ID, Inc., Newark, Delaware. 338
Sawabe T, Kita–Tsukamoto K, Thompson FL (2007) Inferring the evolutionary history 339
of vibrios by means of multilocus sequence analysis. J Bacteriol 189:7932–7936. 340
Stackebrandt E Ebers J (2006) Taxonomic parameters revisited: tranished gold 341
standards. Microbiol Today 33:152-155. 342
Sugumar G, Nakai T, Hirata Y, Matsubara D, Helinski DR (1998) Vibrio splendidus 343
biovar II as the causative agent of bacillary necrosis of Japanese oyster Crassostrea 344
gigas larvae. Dis Aquat Organ 33:111–118. 345
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011) MEGA5: 346
molecular evolutionary genetics analysis using maximum likelihood, evolutionary 347
distance, and maximum parsimony methods. Mol Biol Evol 28:2731–2739. 348
Thompson CC, Thompson FL, Vandemeulebroecke K, Hoste B, Dawyndt P, Swings J 349
(2004) Use of recA as an alternative phylogenetic marker in the family 350
Vibrionaceae. Int J Syst Evol Microbiol 54:919–924. 351
Thompson CC, Thompson FL, Vicente AC, Swings J (2007) Phylogenetic analysis of 352
vibrios and related species by means of atpA gene sequences. Int J Syst Evol 353
Microbiol 57:2480–2484. 354
Thompson FL, Gevers D, Thompson CC, Dawyndt P, Naser S, Hoste B, Munn CB, 355
Swings J (2005) Phylogeny and molecular identification of Vibrios on the basis of 356
multilocus sequence analysis. Appl Environ Microbiol 71:5107–5115. 357
Thompson FL, Swings J (2006) Taxonomy of the vibrios. In: Thompson, F.L., Austin, 358
B., Swings, J. (Eds.), The Biology of Vibrios. American Society for Microbiology, 359
Washington, DC, pp. 29–43. 360
Ziemke F, Höfle MG, Lalucat J, Rosselló–Mora R. (1998) Reclassification of 361
Shewanella putrefaciens Owen's genomic group II as Shewanella baltica sp. nov. Int 362
J Syst Bacteriol 48:179–186. 363
364
365
366
367 368

13
369 Figure legends 370
371 Figure 1. Phylogenetic position of the four isolates according to 16S rRNA gene 372
sequence analysis. The tree is a NJ tree; Vibrio cholera was used as an outgroup. 373
GeneBank sequence accession numbers are given in parentheses. Numbers at the 374
nodes show the percentage bootstrap values (only values higher than 50% are 375
shown). Bar, 0.002 substitutions per nucleotide position. 376
377
378
Figure 2. Phylogenetic position of the four isolates according to MLSA of the five 379
housekeeping genes atpA, pyrH, recA, rpoA and rpoD, and the 16S rRNA gene. The 380
tree is a NJ tree. Numbers at the nodes show the percentage bootstrap values (only 381
values higher than 50% are shown). Bar, 0.02 substitutions per nucleotide position. 382
383 384 385
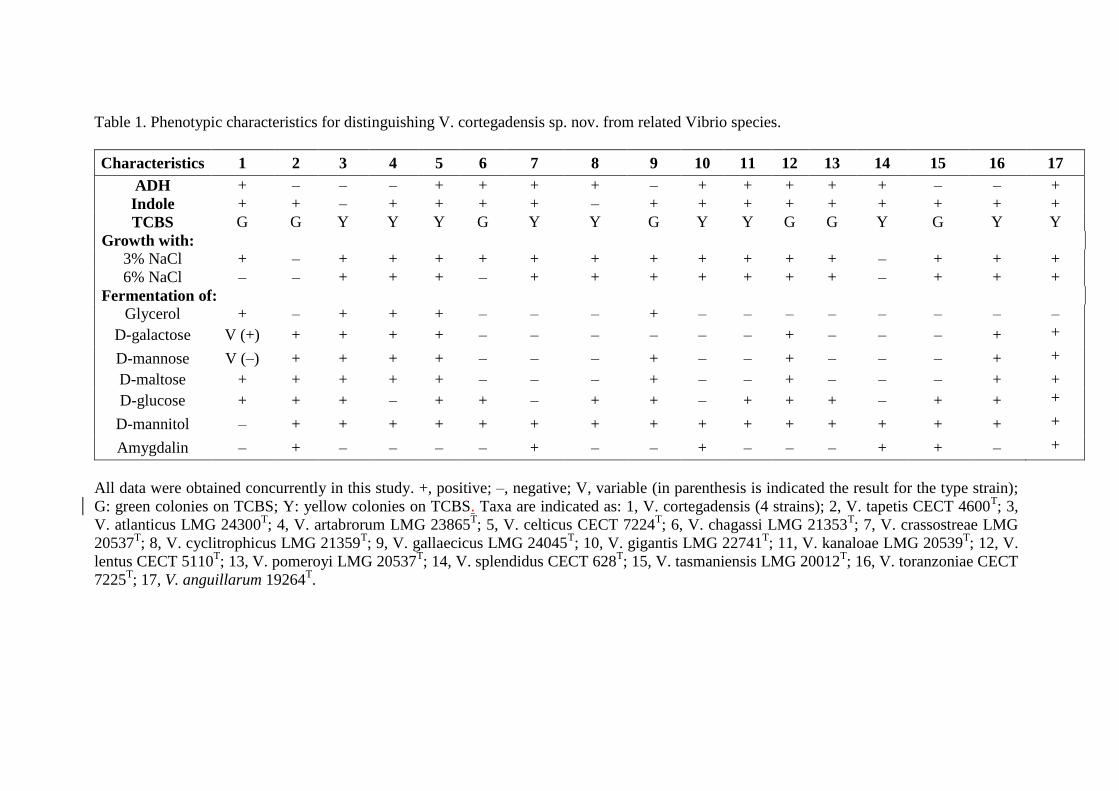
Table 1. Phenotypic characteristics for distinguishing V. cortegadensis sp. nov. from related Vibrio species.
Characteristics 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
ADH + – – – + + + + – + + + + + – – +
Indole + + – + + + + – + + + + + + + + +
TCBS G G Y Y Y G Y Y G Y Y G G Y G Y Y
Growth with:
3% NaCl + – + + + + + + + + + + + – + + +
6% NaCl – – + + + – + + + + + + + – + + +
Fermentation of:
Glycerol + – + + + – – – + – – – – – – – –
D-galactose V (+) + + + + – – – – – – + – – – + +
D-mannose V (–) + + + + – – – + – – + – – – + +
D-maltose + + + + + – – – + – – + – – – + +
D-glucose + + + – + + – + + – + + + – + + +
D-mannitol – + + + + + + + + + + + + + + + +
Amygdalin – + – – – – + – – + – – – + + – +
All data were obtained concurrently in this study. +, positive; –, negative; V, variable (in parenthesis is indicated the result for the type strain);
G: green colonies on TCBS; Y: yellow colonies on TCBS. Taxa are indicated as: 1, V. cortegadensis (4 strains); 2, V. tapetis CECT 4600T; 3,
V. atlanticus LMG 24300T; 4, V. artabrorum LMG 23865
T; 5, V. celticus CECT 7224
T; 6, V. chagassi LMG 21353
T; 7, V. crassostreae LMG
20537T; 8, V. cyclitrophicus LMG 21359
T; 9, V. gallaecicus LMG 24045
T; 10, V. gigantis LMG 22741
T; 11, V. kanaloae LMG 20539
T; 12, V.
lentus CECT 5110T; 13, V. pomeroyi LMG 20537
T; 14, V. splendidus CECT 628
T; 15, V. tasmaniensis LMG 20012
T; 16, V. toranzoniae CECT
7225T; 17, V. anguillarum 19264
T.

Figure 1Click here to download high resolution image
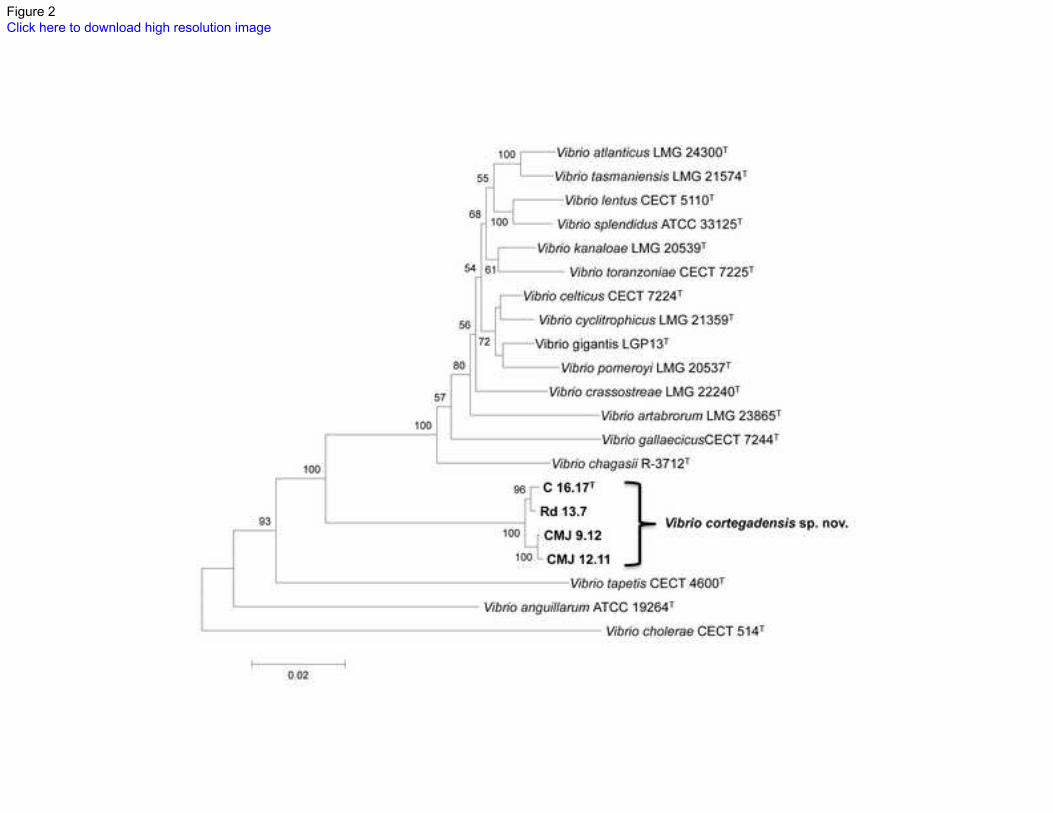
Figure 2Click here to download high resolution image

Description of the new species Vibrio cortegadensis sp. nov., isolated from clams
Aide Lasa, Ana L. Diéguez, and Jesús L. Romalde *
.
Supplementary Material
Supplementary MaterialClick here to download attachment to manuscript: Supplementary material.doc

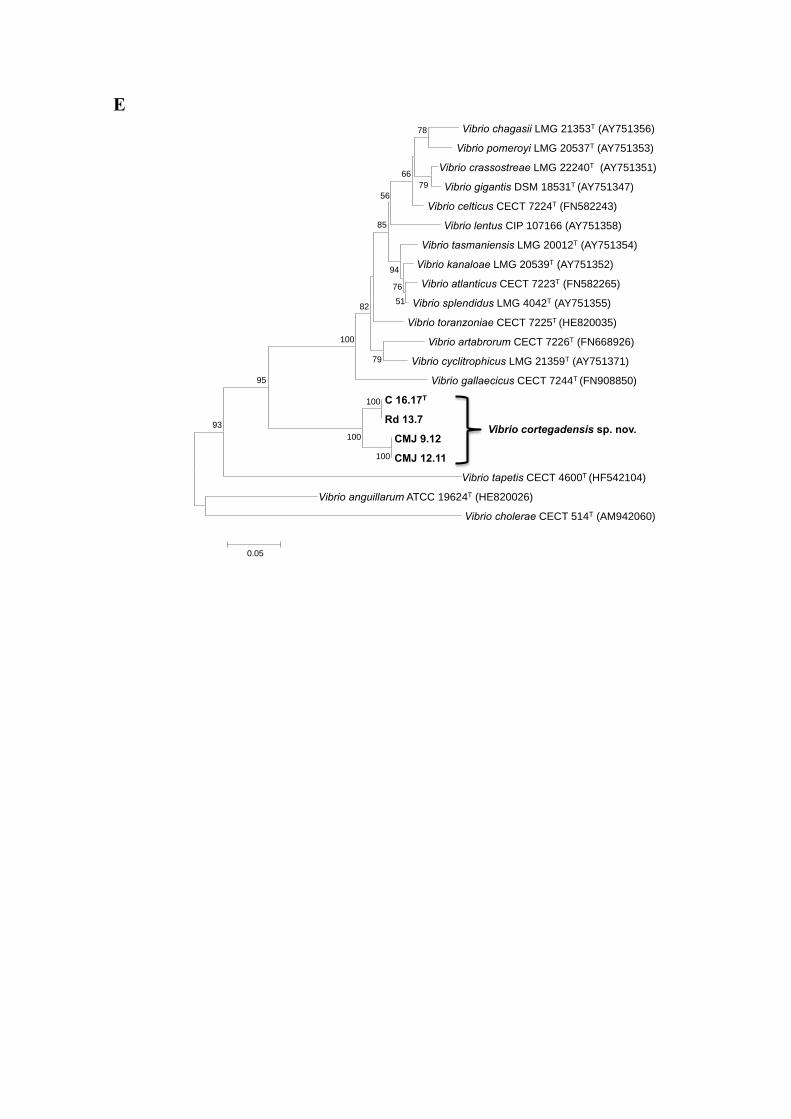
Figure S1. Phylogenetic tree based on partial 16S rRNA gene sequences, obtained by
the Maximum Likelihood method with the K2+G+I model. GeneBank sequence
accession numbers are given in parentheses. Numbers at the nodes show the percentages
bootstrap values (only values higher than 50% are shown). Bar, 0.01 substitutions per
nucleotide position.
Vibrio kanaloae LMG 20539T (AM162657)
Vibrio lentus CECT 5110T (NR 028926)
Vibrio hemicentroti DSM 26178T (JX204734)
Vibrio atlanticus LMG 24300T (EF599163)
Vibrio tasmaniensis LMG 21574T (AJ514912)
Vibrio cyclitrophicus LMG 21359T (AM162656)
Vibrio splendidus ATCC 33125T (X74724) Vibrio gallaecicus CECT 7244T (NR 044520)
Vibrio gigantis LGP 13T (AJ582810)
Vibrio celticus LMG 23850T (EF599162)
Vibrio artabrorum LMG 23865T (EF599164)
Vibrio toranzoniae CECT 7225T (HE978310)
Vibrio pomeroyi LMG 20537T (AJ491290)
Vibrio chagasii R-3712T (AJ316199)
Vibrio crassostreae CAIM 1405T (EF094887)
Vibrio tapetis CECT 4600T (HE795129)
CMJ 9.12 (HF955038)
Rd 13.7 (HF955040)
C 16.17T (HF955037)
CMJ 12.11 (HF955039)
Vibrio pacinii LMG 19999T (AJ316194)
Vibrio ordalii ATCC 33509T (AEZC01000173)
Vibrio anguillarum ATCC 19264T (X16895)
Vibrio aestuarianus ATCC 35048T (X74689)
Vibrio mediterranei LMG11258T (HM771351) Vibrio shilonii AK1T (ABCH01000080)
Vibrio scophthalmi LMG 19158T (HM771340)
Vibrio diazotrophicus LMG7893T (X56577) Vibrio plantisponsor MSSRF60T (GQ352641)
Vibrio vulnificus LMG13545T (X76333) Vibrio azureus LC2-005T (AB428897)
Vibrio sagamiensis LC2-047TT (AB428909)
Vibrio jasicida TCFB 0772T (AB562589)
Vibrio owensii DY05T (GU018180)
Vibrio corallilyticus LMG 20984T (AJ440005)
Vibrio nereis ATCC 25917T (HQ890463) Vibrio harveyi LMG4044T (AY750575)
Vibrio variabilis R-40492T (GU929924)
Vibrio maritimus R-40493T (GU929925)
Vibrio nigripulchritudo LMG 3896T (HM771352) Vibrio halioticoli LMG 18542T (AB000390)
Vibrio cholerae CECT 514T (NR 044853)
62
100
87
54
80
90
99
67
55
0.01
Vibrio cortegadensis sp. nov.
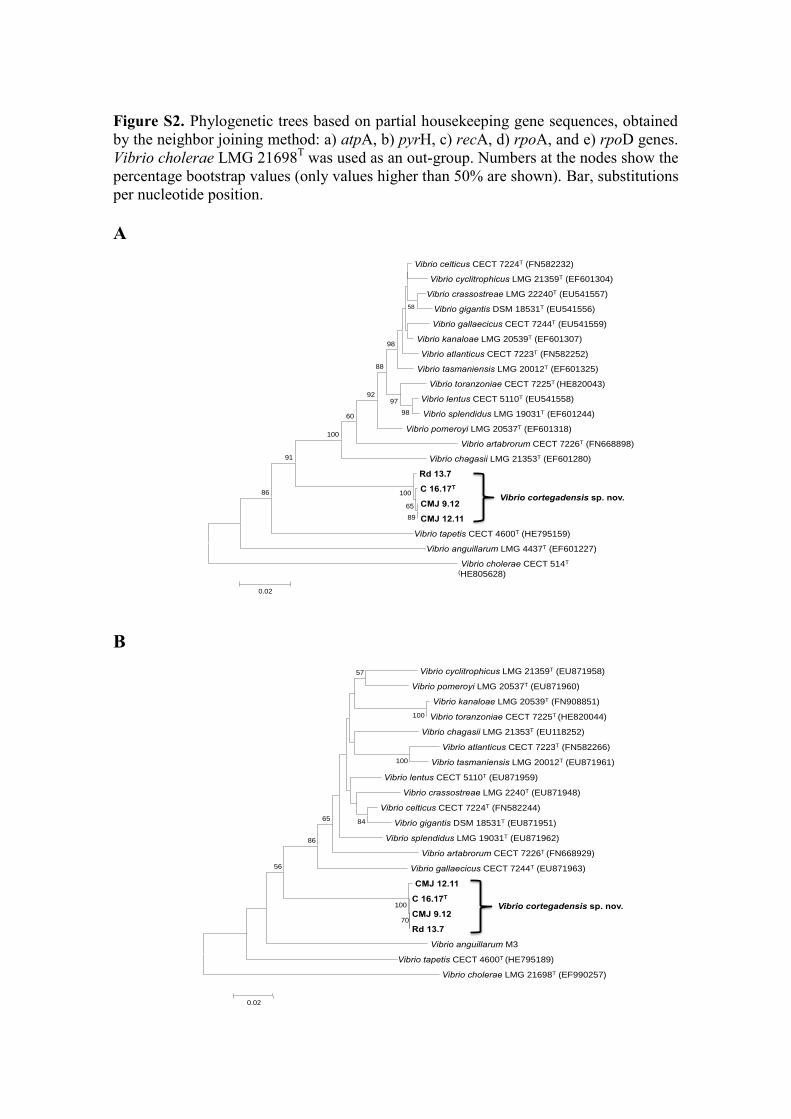
Figure S2. Phylogenetic trees based on partial housekeeping gene sequences, obtained
by the neighbor joining method: a) atpA, b) pyrH, c) recA, d) rpoA, and e) rpoD genes.
Vibrio cholerae LMG 21698T was used as an out-group. Numbers at the nodes show the
percentage bootstrap values (only values higher than 50% are shown). Bar, substitutions
per nucleotide position.
A
Vibrio celticus CECT 7224T (FN582232) Vibrio cyclitrophicus LMG 21359T (EF601304)
Vibrio crassostreae LMG 22240T (EU541557) Vibrio gigantis DSM 18531T (EU541556) Vibrio gallaecicus CECT 7244T (EU541559)
Vibrio kanaloae LMG 20539T (EF601307) Vibrio atlanticus CECT 7223T (FN582252)
Vibrio tasmaniensis LMG 20012T (EF601325) Vibrio toranzoniae CECT 7225T (HE820043)
Vibrio lentus CECT 5110T (EU541558) Vibrio splendidus LMG 19031T (EF601244)
Vibrio pomeroyi LMG 20537T (EF601318) Vibrio artabrorum CECT 7226T (FN668898)
Vibrio chagasii LMG 21353T (EF601280) Rd 13.7 C 16.17T CMJ 9.12 CMJ 12.11
Vibrio tapetis CECT 4600T (HE795159)
Vibrio anguillarum LMG 4437T (EF601227)
Vibrio cholerae CECT 514T
(HE805628)
89 65
100 86
91
100
60 98 97 92
88
98
58
0.02
Vibrio cortegadensis sp. nov.
B
Vibrio cyclitrophicus LMG 21359T (EU871958) Vibrio pomeroyi LMG 20537T (EU871960)
Vibrio kanaloae LMG 20539T (FN908851) Vibrio toranzoniae CECT 7225T (HE820044)
Vibrio chagasii LMG 21353T (EU118252) Vibrio atlanticus CECT 7223T (FN582266)
Vibrio tasmaniensis LMG 20012T (EU871961) Vibrio lentus CECT 5110T (EU871959)
Vibrio crassostreae LMG 2240T (EU871948) Vibrio celticus CECT 7224T (FN582244)
Vibrio gigantis DSM 18531T (EU871951) Vibrio splendidus LMG 19031T (EU871962)
Vibrio artabrorum CECT 7226T (FN668929) Vibrio gallaecicus CECT 7244T (EU871963) CMJ 12.11
C 16.17T CMJ 9.12 Rd 13.7
Vibrio anguillarum M3 Vibrio tapetis CECT 4600T (HE795189)
Vibrio cholerae LMG 21698T (EF990257)
70 100
100
100
56
86
84
57
65
0.02
Vibrio cortegadensis sp. nov.
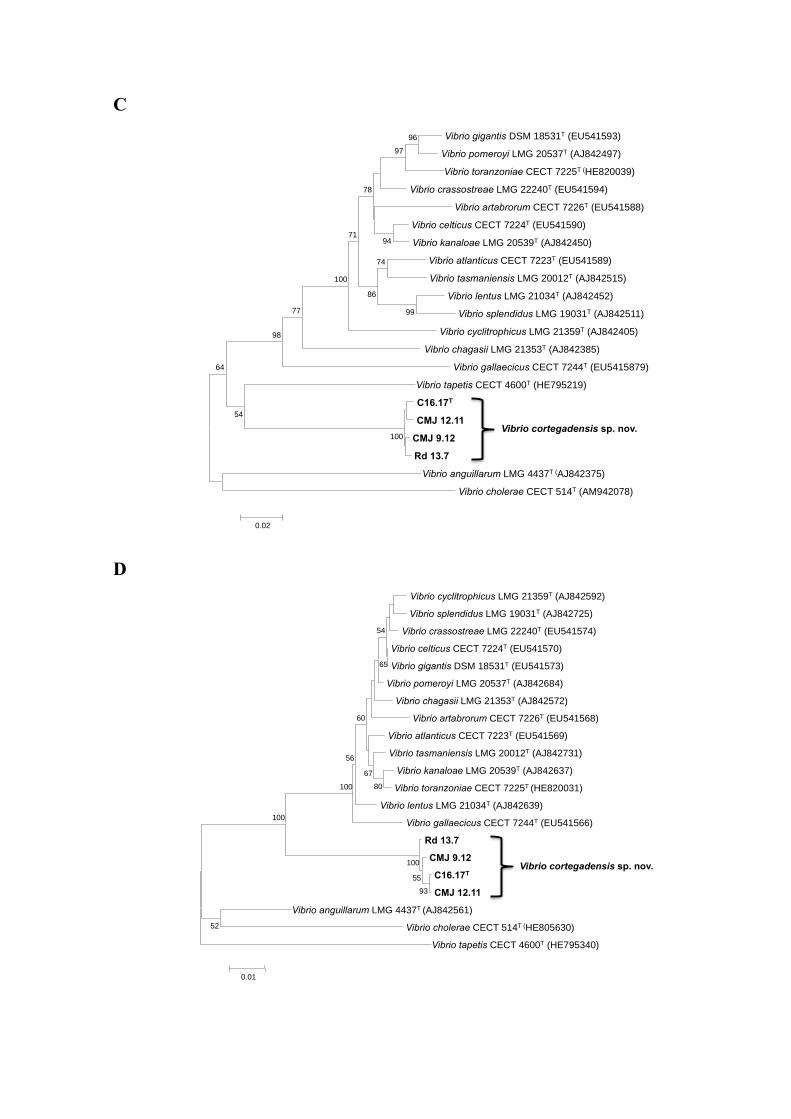
C
Vibrio gigantis DSM 18531T (EU541593) Vibrio pomeroyi LMG 20537T (AJ842497)
Vibrio toranzoniae CECT 7225T (HE820039) Vibrio crassostreae LMG 22240T (EU541594)
Vibrio artabrorum CECT 7226T (EU541588) Vibrio celticus CECT 7224T (EU541590) Vibrio kanaloae LMG 20539T (AJ842450)
Vibrio atlanticus CECT 7223T (EU541589) Vibrio tasmaniensis LMG 20012T (AJ842515)
Vibrio lentus LMG 21034T (AJ842452) Vibrio splendidus LMG 19031T (AJ842511)
Vibrio cyclitrophicus LMG 21359T (AJ842405) Vibrio chagasii LMG 21353T (AJ842385)
Vibrio gallaecicus CECT 7244T (EU5415879) Vibrio tapetis CECT 4600T (HE795219)
C16.17T CMJ 12.11
CMJ 9.12 Rd 13.7
Vibrio anguillarum LMG 4437T (AJ842375)
Vibrio cholerae CECT 514T (AM942078)
100
54
64
98
77 99
96 97
74
86
94
100
71
78
0.02
Vibrio cortegadensis sp. nov.
D
Vibrio cyclitrophicus LMG 21359T (AJ842592) Vibrio splendidus LMG 19031T (AJ842725)
Vibrio crassostreae LMG 22240T (EU541574) Vibrio celticus CECT 7224T (EU541570) Vibrio gigantis DSM 18531T (EU541573)
Vibrio pomeroyi LMG 20537T (AJ842684) Vibrio chagasii LMG 21353T (AJ842572)
Vibrio artabrorum CECT 7226T (EU541568) Vibrio atlanticus CECT 7223T (EU541569) Vibrio tasmaniensis LMG 20012T (AJ842731)
Vibrio kanaloae LMG 20539T (AJ842637) Vibrio toranzoniae CECT 7225T (HE820031)
Vibrio lentus LMG 21034T (AJ842639) Vibrio gallaecicus CECT 7244T (EU541566)
Rd 13.7 CMJ 9.12 C16.17T CMJ 12.11
Vibrio anguillarum LMG 4437T (AJ842561)
Vibrio cholerae CECT 514T (HE805630) Vibrio tapetis CECT 4600T (HE795340)
93 55
100
52
100
80 100
56 67
60
65
54
0.01
Vibrio cortegadensis sp. nov.
E
Vibrio chagasii LMG 21353T (AY751356) Vibrio pomeroyi LMG 20537T (AY751353)
Vibrio crassostreae LMG 22240T (AY751351) Vibrio gigantis DSM 18531T (AY751347)
Vibrio celticus CECT 7224T (FN582243) Vibrio lentus CIP 107166 (AY751358)
Vibrio tasmaniensis LMG 20012T (AY751354) Vibrio kanaloae LMG 20539T (AY751352) Vibrio atlanticus CECT 7223T (FN582265)
Vibrio splendidus LMG 4042T (AY751355) Vibrio toranzoniae CECT 7225T (HE820035)
Vibrio artabrorum CECT 7226T (FN668926) Vibrio cyclitrophicus LMG 21359T (AY751371)
Vibrio gallaecicus CECT 7244T (FN908850) C 16.17T
Rd 13.7 CMJ 9.12 CMJ 12.11
Vibrio tapetis CECT 4600T (HF542104)
Vibrio anguillarum ATCC 19624T (HE820026)
Vibrio cholerae CECT 514T (AM942060)
100
100
100 93
95
78
100 79
79 66
82
56
85
94
51 76
0.05
Vibrio cortegadensis sp. nov.
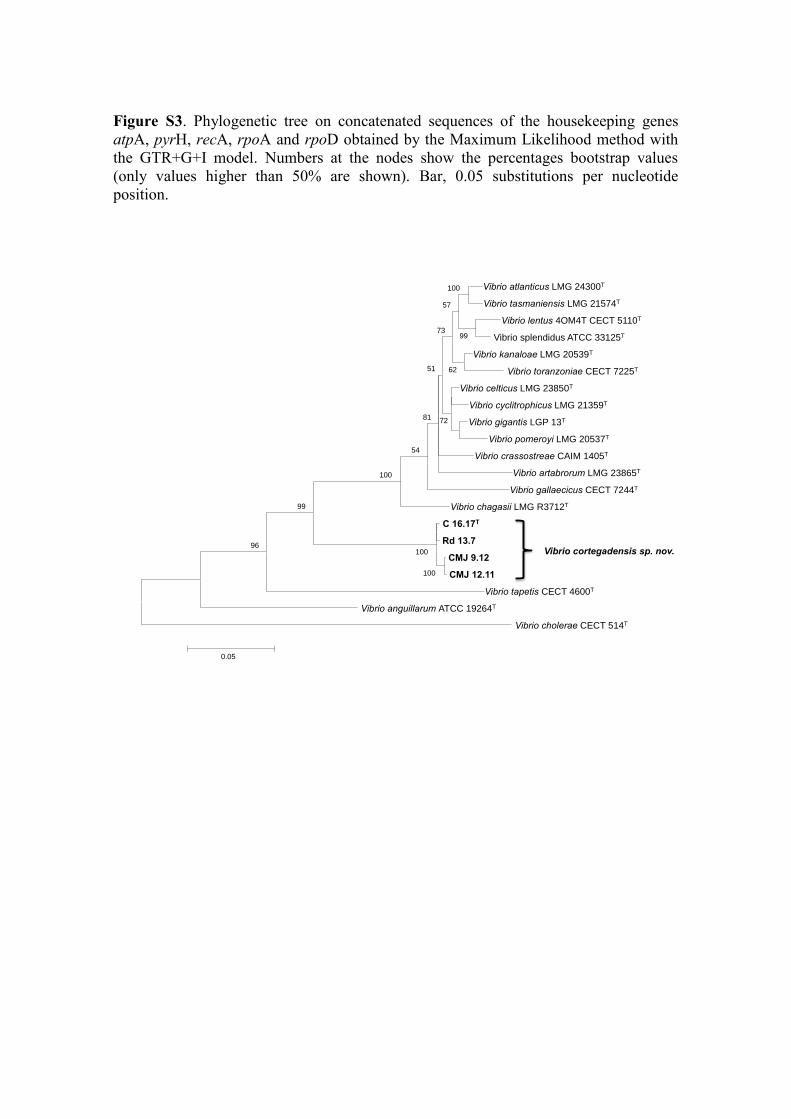
Figure S3. Phylogenetic tree on concatenated sequences of the housekeeping genes
atpA, pyrH, recA, rpoA and rpoD obtained by the Maximum Likelihood method with
the GTR+G+I model. Numbers at the nodes show the percentages bootstrap values
(only values higher than 50% are shown). Bar, 0.05 substitutions per nucleotide
position.
Vibrio atlanticus LMG 24300T
Vibrio tasmaniensis LMG 21574T
Vibrio lentus 4OM4T CECT 5110T
Vibrio splendidus ATCC 33125T Vibrio kanaloae LMG 20539T
Vibrio toranzoniae CECT 7225T
Vibrio celticus LMG 23850T Vibrio cyclitrophicus LMG 21359T
Vibrio gigantis LGP 13T Vibrio pomeroyi LMG 20537T
Vibrio crassostreae CAIM 1405T Vibrio artabrorum LMG 23865T
Vibrio gallaecicus CECT 7244T
Vibrio chagasii LMG R3712T
C 16.17T Rd 13.7
CMJ 9.12 CMJ 12.11
Vibrio tapetis CECT 4600T Vibrio anguillarum ATCC 19264T
Vibrio cholerae CECT 514T
100
100 96
99
100
100
99
54
62
72
57
81
73
51
0.05
Vibrio cortegadensis sp. nov.
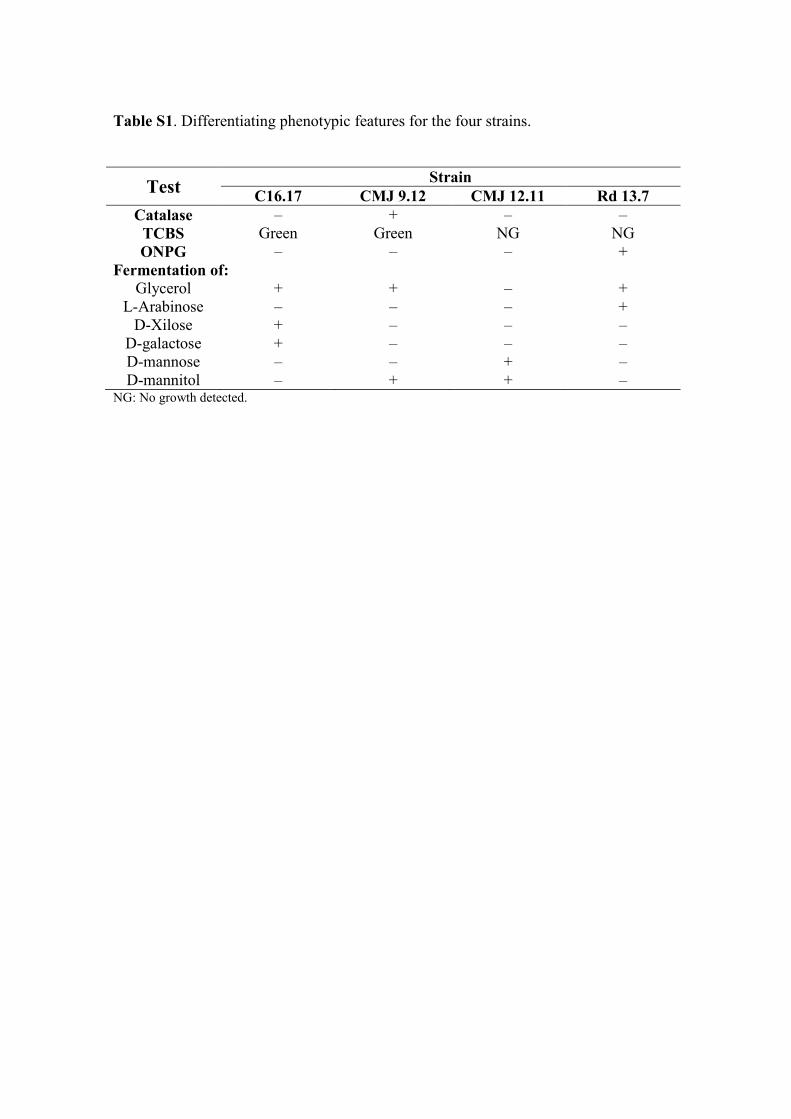
Table S1. Differentiating phenotypic features for the four strains.
Test Strain
C16.17 CMJ 9.12 CMJ 12.11 Rd 13.7 Catalase – + – –
TCBS Green Green NG NG
ONPG – – – +
Fermentation of:
Glycerol + + – +
L-Arabinose – – – +
D-Xilose + – – –
D-galactose + – – –
D-mannose – – + –
D-mannitol – + + – NG: No growth detected.
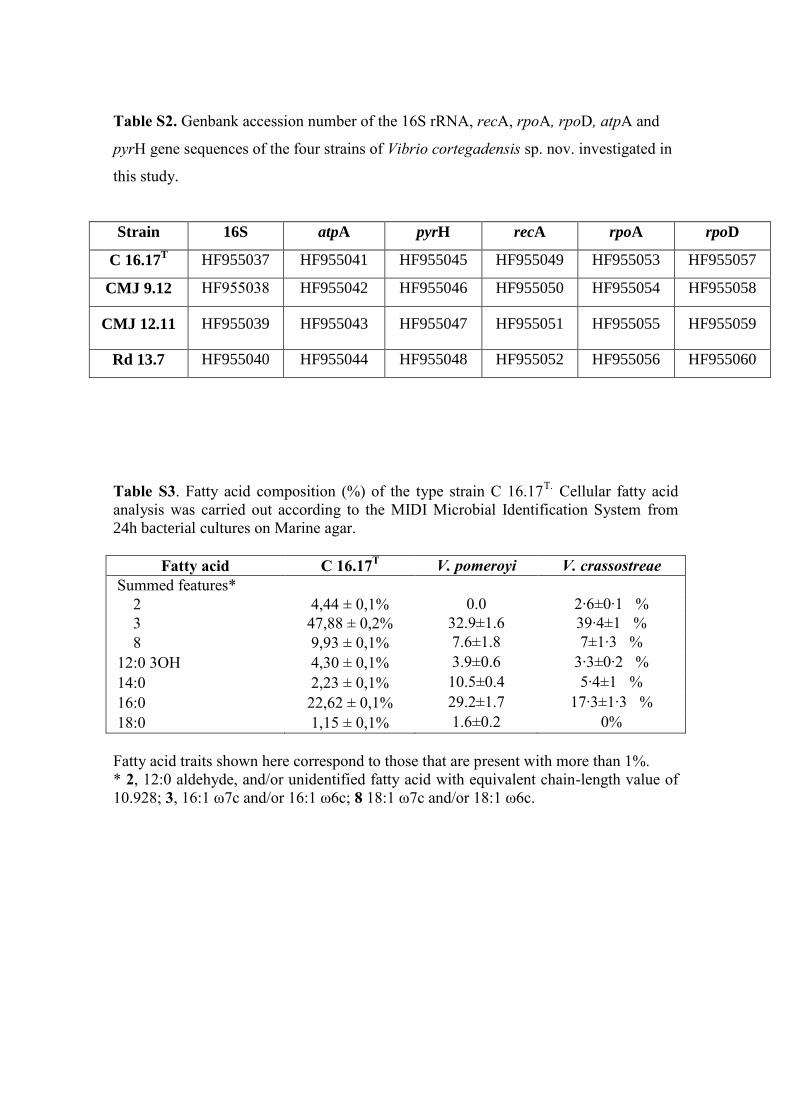
Table S2. Genbank accession number of the 16S rRNA, recA, rpoA, rpoD, atpA and
pyrH gene sequences of the four strains of Vibrio cortegadensis sp. nov. investigated in
this study.
Table S3. Fatty acid composition (%) of the type strain C 16.17T. Cellular fatty acid
analysis was carried out according to the MIDI Microbial Identification System from
24h bacterial cultures on Marine agar.
Fatty acid C 16.17T V. pomeroyi V. crassostreae
Summed features*
2 4,44 ± 0,1% 0.0 2·6±0·1 %
3 47,88 ± 0,2% 32.9±1.6 39·4±1 %
8 9,93 ± 0,1% 7.6±1.8 7±1·3 %
12:0 3OH 4,30 ± 0,1% 3.9±0.6 3·3±0·2 %
14:0 2,23 ± 0,1% 10.5±0.4 5·4±1 %
16:0 22,62 ± 0,1% 29.2±1.7 17·3±1·3 %
18:0 1,15 ± 0,1% 1.6±0.2 0%
Fatty acid traits shown here correspond to those that are present with more than 1%.
* 2, 12:0 aldehyde, and/or unidentified fatty acid with equivalent chain-length value of
10.928; 3, 16:1 ω7c and/or 16:1 ω6c; 8 18:1 ω7c and/or 18:1 ω6c.
Strain 16S atpA pyrH recA rpoA rpoD
C 16.17T HF955037 HF955041 HF955045 HF955049 HF955053 HF955057
CMJ 9.12 HF955038 HF955042 HF955046 HF955050 HF955054 HF955058
CMJ 12.11 HF955039 HF955043 HF955047 HF955051 HF955055 HF955059
Rd 13.7 HF955040 HF955044 HF955048 HF955052 HF955056 HF955060




















