
Photoacclimation to Long-Term Ultraviolet Radiation Exposure of Natural Sub-Antarctic Phytoplankton...
13
Photochemistry and Photobiology, 2006, 82: 923–935 Symposium-in-Print: UV Effects on Aquatic and Coastal Ecosystems Photoacclimation to Long-Term Ultraviolet Radiation Exposure of Natural Sub-Antarctic Phytoplankton Communities: Fixed Surface Incubations Versus Mixed Mesocosms Marcelo Hernando* 1 , Irene Schloss 1,2 , Suzanne Roy 3 and Gustavo Ferreyra 2 1 CONICET, Argentina 2 Instituto Anta ´ rtico Argentino, Cerrito 1248, C1010AAZ, Buenos Aires, Argentina 3 Institut des Sciences de la Mer (ISMER), Universite ´ du Que ´bec a ` Rimouski, Rimouski, Que ´ bec, Canada Received 29 August 2005; accepted 24 May 2006; published online 26 May 2006 DOI: 10.1562/2005-08-29-RA-662 ABSTRACT Solar UVB radiation (280–320 nm) is known to have detri- mental effects on marine phytoplankton. Associated with the seasonal ozone hole in Antarctica, stratospheric ozone de- pletion occasionally influences the sub-Antarctic (Beagle Channel, Argentina) region, enhancing levels of UVB. The primary objective of this work was to study the effects of several (i.e. 6–10) days of exposure to UVB on the taxonomic composition and photosynthetic inhibition of local phyto- plankton communities. For different light treatments, fixed- depth incubations placed in an outdoors water tank were compared with incubations in 1900 L mesocosms, where vertical mixing was present. Phytoplankton growth was inhibited by UV radiation (UVR) in fixed-depth experiments but not in the mixed mesocosms. Under fixed and mixed conditions alike, photosynthesis was significantly inhibited by UVB at the beginning of the experiment but no longer after several days of exposure, suggesting that cells had acclimated to radiation conditions. There was a change in species com- position in response to UVR exposure in both experiments, which likely explained acclimation. In the community exposed to fixed conditions this change was from a phytoflagellate- dominated assemblage to a community with high relative abundance of diatoms after 6 days of exposure. UVA was re- sponsible for most of the observed growth inhibition; however, the reduction in photosynthesis was produced by UVB. The reasons behind this variability in responses to UVR are associated with species-specific sensitivity and acclimation, and the previous light history of cells. In the community ex- posed in mesocosms, an assemblage codominated by phyto- flagellates and diatoms was observed at the beginning of the experiments. After 10 days of exposure, green algae (Eutreptiella sp.) had increased, and phytoflagellates were the dominant group. The synthesis of mycosporine-like amino acids (MAAs), antioxidant enzymes and photosynthetic antenna pigments, in relation to repair and protection processes, may explain the reduced inhibition of both growth and photosynthesis that was observed in the phytoplankton community after several days of exposure. For environments such as the Beagle Channel seasonally exposed to the ozone hole, the results obtained from the fixed-depth experiments show that species can cope with UVR by means of MAA synthesis, while mixing would primarily promote a change in species composition and defense strategies. INTRODUCTION It has been shown that UVB and solar UVA radiation (320–400 nm) can have deleterious effects on phytoplankton. These radia- tions can, among other effects, reduce cell growth rates (1), induce DNA damage (2) and inhibit photosynthesis through effects on photosystem II (PSII) (3) and ribulose-1,5-biphosphate carboxylase/ oxygenase (RUBISCO) (4). UVB can penetrate up to ecologically relevant depths in the oceans (5–8). Under low ozone conditions, UVB flux could affect primary production below 15 m in the water column (9–11). However, Cullen and Lesser (12) and Neale et al. (13,14) suggested that the effects of UVB could be greatly attenuated by vertical mixing. The time of residence of cells at a certain depth (i.e. the light history of the cells [15,16]), is therefore influenced by the mixing regime prevailing in the water column. The degree of exposure as well as the balance between cell damage and repair processes will then be a function of both mixing velocity and depth (14). UVR effects have been studied especially in Antarctic waters due to the presence of the seasonal ozone thinning. Sub-Antarctic waters are, however, also exposed to the so-called ozone hole, which can periodically cover the southern tip of South America (17). Previous studies in ocean waters around Antarctica have shown that the tolerance to UVR varies among phytoplankton groups (18) and that a differential tolerance exists to either UVA or UVB (19). Phytoplankton can be protected from UVR- induced damage by the synthesis of mycosporine-like amino acids (MAAs), which absorb radiation in the UVR range (20–22). Antioxidant compounds can also be generated to reduce UVR damage (23). *Corresponding author email: [email protected] (Marcelo Hernando) Ó 2006 American Society for Photobiology 0031-8655/06 923
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Photoacclimation to Long-Term Ultraviolet Radiation Exposure of Natural Sub-Antarctic Phytoplankton...

Photochemistry and Photobiology, 2006, 82: 923–935
Symposium-in-Print: UV Effects on Aquatic and Coastal Ecosystems
Photoacclimation to Long-Term Ultraviolet Radiation Exposureof Natural Sub-Antarctic Phytoplankton Communities:Fixed Surface Incubations Versus Mixed Mesocosms
Marcelo Hernando*1, Irene Schloss1,2, Suzanne Roy3 and Gustavo Ferreyra2
1CONICET, Argentina2Instituto Antartico Argentino, Cerrito 1248, C1010AAZ, Buenos Aires, Argentina3Institut des Sciences de la Mer (ISMER), Universite du Quebec a Rimouski, Rimouski, Quebec, Canada
Received 29 August 2005; accepted 24 May 2006; published online 26 May 2006 DOI: 10.1562/2005-08-29-RA-662
ABSTRACT
Solar UVB radiation (280–320 nm) is known to have detri-mental effects on marine phytoplankton. Associated with theseasonal ozone hole in Antarctica, stratospheric ozone de-pletion occasionally influences the sub-Antarctic (BeagleChannel, Argentina) region, enhancing levels of UVB. Theprimary objective of this work was to study the effects ofseveral (i.e. 6–10) days of exposure to UVB on the taxonomiccomposition and photosynthetic inhibition of local phyto-plankton communities. For different light treatments, fixed-depth incubations placed in an outdoors water tank werecompared with incubations in 1900 L mesocosms, wherevertical mixing was present. Phytoplankton growth wasinhibited by UV radiation (UVR) in fixed-depth experimentsbut not in the mixed mesocosms. Under fixed and mixedconditions alike, photosynthesis was significantly inhibited byUVB at the beginning of the experiment but no longer afterseveral days of exposure, suggesting that cells had acclimatedto radiation conditions. There was a change in species com-position in response to UVR exposure in both experiments,which likely explained acclimation. In the community exposedto fixed conditions this change was from a phytoflagellate-dominated assemblage to a community with high relativeabundance of diatoms after 6 days of exposure. UVA was re-sponsible for most of the observed growth inhibition; however,the reduction in photosynthesis was produced by UVB.The reasons behind this variability in responses to UVR areassociated with species-specific sensitivity and acclimation,and the previous light history of cells. In the community ex-posed in mesocosms, an assemblage codominated by phyto-flagellates and diatoms was observed at the beginning ofthe experiments. After 10 days of exposure, green algae(Eutreptiella sp.) had increased, and phytoflagellates were thedominant group. The synthesis of mycosporine-like aminoacids (MAAs), antioxidant enzymes and photosynthetic
antenna pigments, in relation to repair and protectionprocesses, may explain the reduced inhibition of both growthand photosynthesis that was observed in the phytoplanktoncommunity after several days of exposure. For environmentssuch as the Beagle Channel seasonally exposed to the ozonehole, the results obtained from the fixed-depth experimentsshow that species can cope with UVR by means of MAAsynthesis, while mixing would primarily promote a changein species composition and defense strategies.
INTRODUCTION
It has been shown that UVB and solar UVA radiation (320–400
nm) can have deleterious effects on phytoplankton. These radia-
tions can, among other effects, reduce cell growth rates (1), induce
DNA damage (2) and inhibit photosynthesis through effects on
photosystem II (PSII) (3) and ribulose-1,5-biphosphate carboxylase/
oxygenase (RUBISCO) (4). UVB can penetrate up to ecologically
relevant depths in the oceans (5–8). Under low ozone conditions,
UVB flux could affect primary production below 15 m in the water
column (9–11). However, Cullen and Lesser (12) and Neale et al.(13,14) suggested that the effects of UVB could be greatly
attenuated by vertical mixing. The time of residence of cells at a
certain depth (i.e. the light history of the cells [15,16]), is therefore
influenced by the mixing regime prevailing in the water column.
The degree of exposure as well as the balance between cell damage
and repair processes will then be a function of both mixing velocity
and depth (14).
UVR effects have been studied especially in Antarctic waters
due to the presence of the seasonal ozone thinning. Sub-Antarctic
waters are, however, also exposed to the so-called ozone hole,
which can periodically cover the southern tip of South America
(17). Previous studies in ocean waters around Antarctica have
shown that the tolerance to UVR varies among phytoplankton
groups (18) and that a differential tolerance exists to either
UVA or UVB (19). Phytoplankton can be protected from UVR-
induced damage by the synthesis of mycosporine-like amino acids
(MAAs), which absorb radiation in the UVR range (20–22).
Antioxidant compounds can also be generated to reduce UVR
damage (23).
*Corresponding author email: [email protected](Marcelo Hernando)
� 2006 American Society for Photobiology 0031-8655/06
923
Fixed surface incubations (i.e. not vertically mixed) conditions
represent the worst-case scenario for phytoplankton. Samples from
the natural environment are exposed to such a radiation regime
only when remaining near the ocean surface. This situation may
be possible in the area from which the samples were obtained.
Navarino and Isla Grande de Tierra del Fuego act as barriers to
give this part of the Beagle Channel the character of a semienclosed
water body. Low surface salinity (2.5–2.7 PSU (practical salinity
units) [24]) from local inputs of great amounts of glacial melt water
(24) can produce a strong stratification, which will remain as long
as intense winds are absent (25).
Vertical mixing is one of the most important variables within
the water column (26) because it exposes planktonic organisms to
fluctuating (i.e. variable) radiation regimes (27). The simulation of
mixing and of more realistic environmental conditions is only
possible when a large water mass is studied. Several UVR studies
with natural phytoplankton communities have been performed
in recent years using outdoor mesocosms (e.g. 28–30). Although
this approach is different from the natural environment, it has
advantages primarily in terms of the more natural light environ-
ment and presence of planktonic communities resembling local
conditions. Most of the earlier studies were performed on a short-
term scale (hours). However, long-term (several days) studies are
required for a better understanding of the effect of UVR radia-
tion on the phytoplankton community.
The primary goal of this work was to study the effects of several-
day exposure to UVR on the taxonomic composition, growth and
photosynthetic inhibition of sub-Antarctic phytoplankton commu-
nities. The algae were exposed to constant light environments
(photosynthetic available radiation [PAR], 400–700 nm and UVR)
under different mixing regimes, comparing surface exposure with
those of mixed, large-volume mesocosms.
MATERIALS AND METHODS
Experimental set-up. Two series of experiments were performed in Ushuaia(548529S, 688189W, Tierra del Fuego, Argentina) with surface phytoplank-ton collected from the Beagle Channel. In the first experiment (fixed-depth),performed 11–17 January 1998, phytoplankton were exposed to either UVRor PAR in 500 mL vessels. Three irradiance treatments were included:PARþUVR (NUVB treatment), PARþUVA (WUVB treatment) and PAR
only (PAR treatment). This design allows the separation of the inhibitoryeffects of UVA (WUVB) and UVB (NUVB). Cylindrical UV-transparentvessels (Plexiglas UVT, GS 2458, Rohm and Haas, Darmstadt, Germany)were used to expose phytoplankton to full solar radiation. For the WUVBtreatment, the vessels were covered with Mylar (DuPont, 50% transmissionat 323 nm), which allows cells to be exposed to UVA þ PAR. The PARtreatment was performed with cylindrical Plexiglass vessels (UF3) (Rohmand Haas), which cuts off all UVR. Three replicates were performed foreach of the treatments. The vessels were stirred manually twice a day. Thealgae were exposed to natural sunlight during 6 days. The vessels wereplaced in a water tank, with flowing thermostatic cold tap water to main-tain in situ temperature (approximately 108C). Temperature was monitoreddaily with a mercury thermometer. To prevent nutrient depletion, culturemedium (f/2 concentration [31]) was added to the different treatments atthe beginning of the experiments.
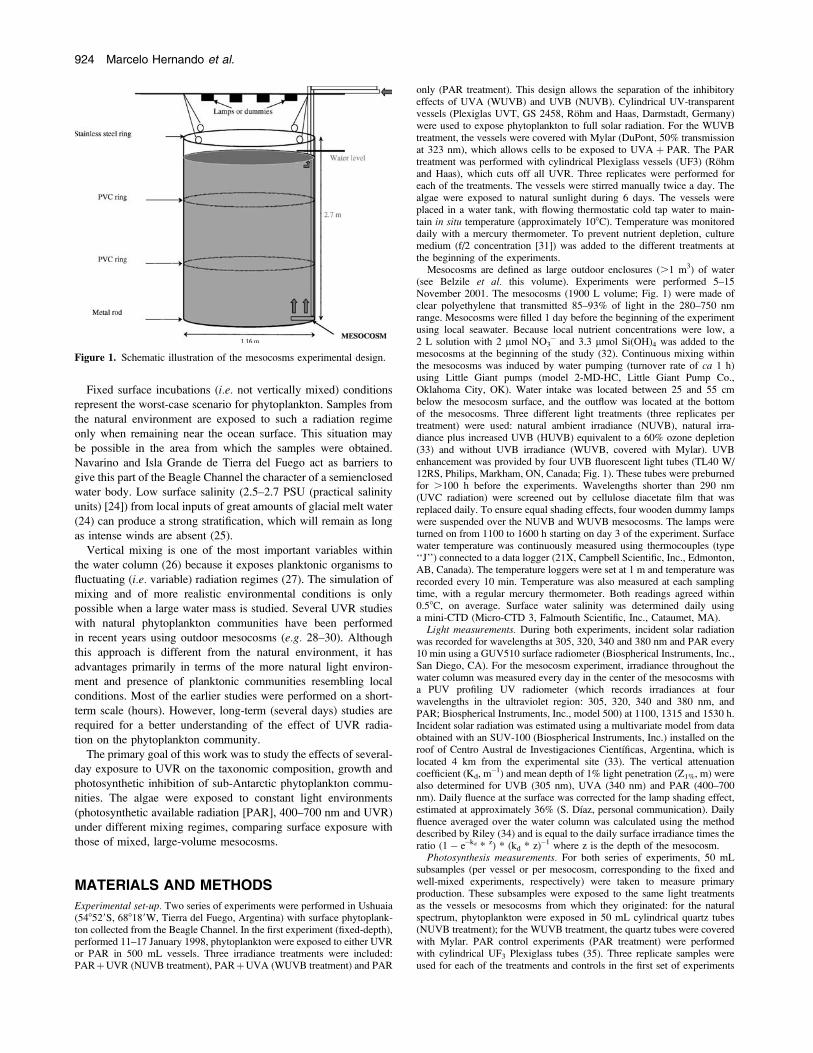
Mesocosms are defined as large outdoor enclosures (.1 m3) of water(see Belzile et al. this volume). Experiments were performed 5–15November 2001. The mesocosms (1900 L volume; Fig. 1) were made ofclear polyethylene that transmitted 85–93% of light in the 280–750 nmrange. Mesocosms were filled 1 day before the beginning of the experimentusing local seawater. Because local nutrient concentrations were low, a2 L solution with 2 lmol NO3
� and 3.3 lmol Si(OH)4 was added to themesocosms at the beginning of the study (32). Continuous mixing withinthe mesocosms was induced by water pumping (turnover rate of ca 1 h)using Little Giant pumps (model 2-MD-HC, Little Giant Pump Co.,Oklahoma City, OK). Water intake was located between 25 and 55 cmbelow the mesocosm surface, and the outflow was located at the bottomof the mesocosms. Three different light treatments (three replicates pertreatment) were used: natural ambient irradiance (NUVB), natural irra-diance plus increased UVB (HUVB) equivalent to a 60% ozone depletion(33) and without UVB irradiance (WUVB, covered with Mylar). UVBenhancement was provided by four UVB fluorescent light tubes (TL40 W/12RS, Philips, Markham, ON, Canada; Fig. 1). These tubes were preburnedfor .100 h before the experiments. Wavelengths shorter than 290 nm(UVC radiation) were screened out by cellulose diacetate film that wasreplaced daily. To ensure equal shading effects, four wooden dummy lampswere suspended over the NUVB and WUVB mesocosms. The lamps wereturned on from 1100 to 1600 h starting on day 3 of the experiment. Surfacewater temperature was continuously measured using thermocouples (type‘‘J’’) connected to a data logger (21X, Campbell Scientific, Inc., Edmonton,AB, Canada). The temperature loggers were set at 1 m and temperature wasrecorded every 10 min. Temperature was also measured at each samplingtime, with a regular mercury thermometer. Both readings agreed within0.58C, on average. Surface water salinity was determined daily usinga mini-CTD (Micro-CTD 3, Falmouth Scientific, Inc., Cataumet, MA).
Light measurements. During both experiments, incident solar radiationwas recorded for wavelengths at 305, 320, 340 and 380 nm and PAR every10 min using a GUV510 surface radiometer (Biospherical Instruments, Inc.,San Diego, CA). For the mesocosm experiment, irradiance throughout thewater column was measured every day in the center of the mesocosms witha PUV profiling UV radiometer (which records irradiances at fourwavelengths in the ultraviolet region: 305, 320, 340 and 380 nm, andPAR; Biospherical Instruments, Inc., model 500) at 1100, 1315 and 1530 h.Incident solar radiation was estimated using a multivariate model from dataobtained with an SUV-100 (Biospherical Instruments, Inc.) installed on theroof of Centro Austral de Investigaciones Cientıficas, Argentina, which islocated 4 km from the experimental site (33). The vertical attenuationcoefficient (Kd, m�1) and mean depth of 1% light penetration (Z1%, m) werealso determined for UVB (305 nm), UVA (340 nm) and PAR (400–700nm). Daily fluence at the surface was corrected for the lamp shading effect,estimated at approximately 36% (S. Dıaz, personal communication). Dailyfluence averaged over the water column was calculated using the methoddescribed by Riley (34) and is equal to the daily surface irradiance times theratio (1� e�kd * z) * (kd * z)�1 where z is the depth of the mesocosm.
Photosynthesis measurements. For both series of experiments, 50 mLsubsamples (per vessel or per mesocosm, corresponding to the fixed andwell-mixed experiments, respectively) were taken to measure primaryproduction. These subsamples were exposed to the same light treatmentsas the vessels or mesocosms from which they originated: for the naturalspectrum, phytoplankton were exposed in 50 mL cylindrical quartz tubes(NUVB treatment); for the WUVB treatment, the quartz tubes were coveredwith Mylar. PAR control experiments (PAR treatment) were performedwith cylindrical UF3 Plexiglass tubes (35). Three replicate samples wereused for each of the treatments and controls in the first set of experiments
Figure 1. Schematic illustration of the mesocosms experimental design.
924 Marcelo Hernando et al.
and one per mesocosm (six per light treatment) for the second set ofexperiments. Samples from both sets of experiments were incubated for 6 haround local noon (i.e. from 1000 to 1600 h) after inoculation of 5 lCi(0.185 MBq) of labeled (NaH14CO3) sodium bicarbonate (36). Followingincubation, the samples were filtered onto Whatman GF/F glass fiber filters(25 mm). The filters were then placed in 7 mL scintillation vials, exposed toHCl fumes overnight, dried, and counted using standard liquid scintillationtechniques (37) in a Packard Liquid Scintillation Analyzer Model 1600 TR(Canberra, Australia). For the fixed-depth experiment, 3 days were studied(i.e. days 0, 2 and 6). Mesocosm subsamples were studied daily, excepton day 4.
Algal pigment analyses and cell counts. In the fixed-depth experiments,80 mL samples were taken on days 0, 2 and 6 at 0900 h to determinechlorophyll a (chl a) concentration by fluorometry and another 20–30 mL,daily, for cell counts. Subsamples for cell counting were kept in darkbottles and fixed with formalin previously neutralized with sodium borate(final concentration 0.4%). Samples were counted with an invertedmicroscope using the Utermohl method (38). Chl a analyses (80 mLsamples for the fixed experiment, and 500 mL for the well-mixedexperiment) were extracted in 7 mL of absolute methanol (39) anddetermined by fluorometric analyses using a Turner fluorometer (model450). Chl a analyses in plankton .5 lm was performed as describedbefore, but after prefiltering the sample with Nuclepore Polycarbonatefilters (5 lm pore size). In the second series of experiments (mesocosms),algal pigments were additionally analyzed by chromatography. Thesesamples were filtered onto 25 mm GF/F filters (Whatman, Maidstone,UK), which were then frozen in liquid nitrogen and subsequently stored at�808C. Samples were sent from the experimental site to Rimouski forfurther analyses at the Institut des Sciences de la Mer de Rimouski.Methanol extracts of pigments present on the GF/F filters were analyzedby HPLC according to the method described by Zapata et al. (40).
Growth measurements. Instantaneous growth rates were determinedaccording to the following equation:
l ¼ lnðNn=Nn�1Þ ð1Þ
where l is the daily growth rate (d�1), Nn is the cell concentration on aspecific day and Nn�1 is the cell concentration on the day before.
To determine the degree of growth inhibition during the fixed ex-periments, instantaneous growth rate in phytoplankton exposed to UVR(WUVB and NUVB treatments) was compared to the average growth ofthose exposed to PAR.
UVR-absorbing compounds. UVR-absorbing compounds (MAAs) weredetermined following the technique described in Helbling et al. (41).Samples from the fixed-depth experiments were extracted for 2 h in 7 mLof methanol at 88C. After extraction, samples were centrifuged and thesupernatant was scanned from 250 to 750 nm with a Shimadzu (model UV-1203) spectrophotometer. The peak height at 334 nm (optical depth,estimated as the height in millimeters of peak maximum from spectropho-tometric scan of methanol extract) was considered an estimate of the con-centration of UVR-absorbing compounds (42). For the mixed mesocosmsexperiments, MAAs were analyzed by chromatography. A two-step extrac-tion procedure was used (43) whereby labile MAAs were extracted first andkept aside, and the filters were re-extracted for less labile MAAs, a methoddeveloped to optimize the extraction efficiency of the diverse MAAassemblages. MAAs were then separated by reverse-phase isocratic HPLC,injecting 50 lL aliquots in a Phenosphere C-8 column (Phenomenex, 250 34.6 mm, 5 lm) protected by a Brownlee Spheri-5 RP-8 guard (AppliedBiosystems, 30 3 4.6 mm, 5 lm) in a mobile phase of 0.1% acetic acid in25% aqueous methanol (44). The MAAs were tentatively identified bycomparison of retention times and order of appearance with publishedresults, and from the absorption spectra of the various peaks. Peak areas andpublished extinction coefficients (45) were used to calculate the concen-tration of MAAs, which were then summed to give the concentration oftotal MAAs in order to compare with the spectrophotometric assay of thefixed-depth experiment.
Statistical analyses. Repeated measures analysis of variance (RMA-NOVA) were performed (Statistica, version 5.1) to determine the sig-nificance of the differences between treatments observed for the carbonassimilation rate, species abundance, total cell number, chl a and growthrate. Exposure days were the repeated factor. Normality was checked usinga one-sample Kolmogorov-Smirnov test, whereas the sphericity assumptionthat concerns variance homogeneity was checked using the Mauchley’stest. When significant differences were found, a post-hoc Tukey test was
performed to determine which factor differed. When interaction wassignificant or the assumptions of sphericity were not satisfied, a one-factoranalysis of variance (ANOVA) was performed (46).
The Levene’s test was used to check for homogeneity of variances (46)for temporal variability UVB, UVA and PAR irradiances.
RESULTS
Fixed-depth experiments
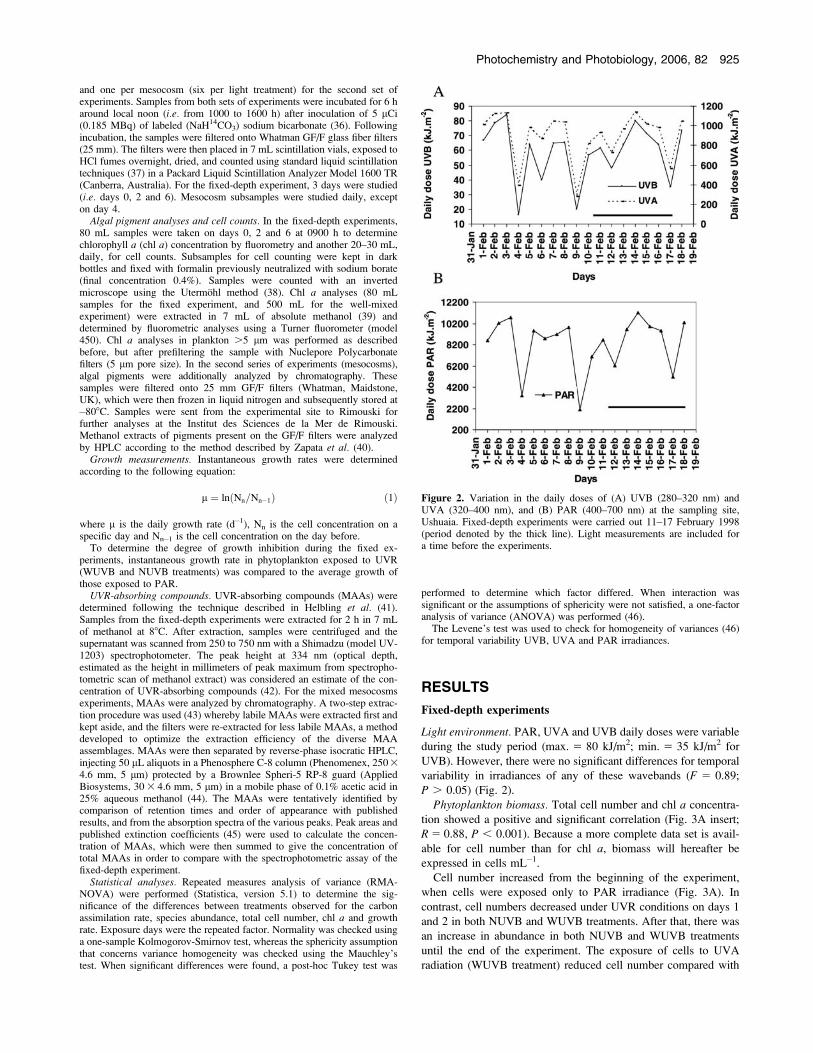
Light environment. PAR, UVA and UVB daily doses were variable
during the study period (max. 5 80 kJ/m2; min. 5 35 kJ/m2 for
UVB). However, there were no significant differences for temporal
variability in irradiances of any of these wavebands (F 5 0.89;
P . 0.05) (Fig. 2).
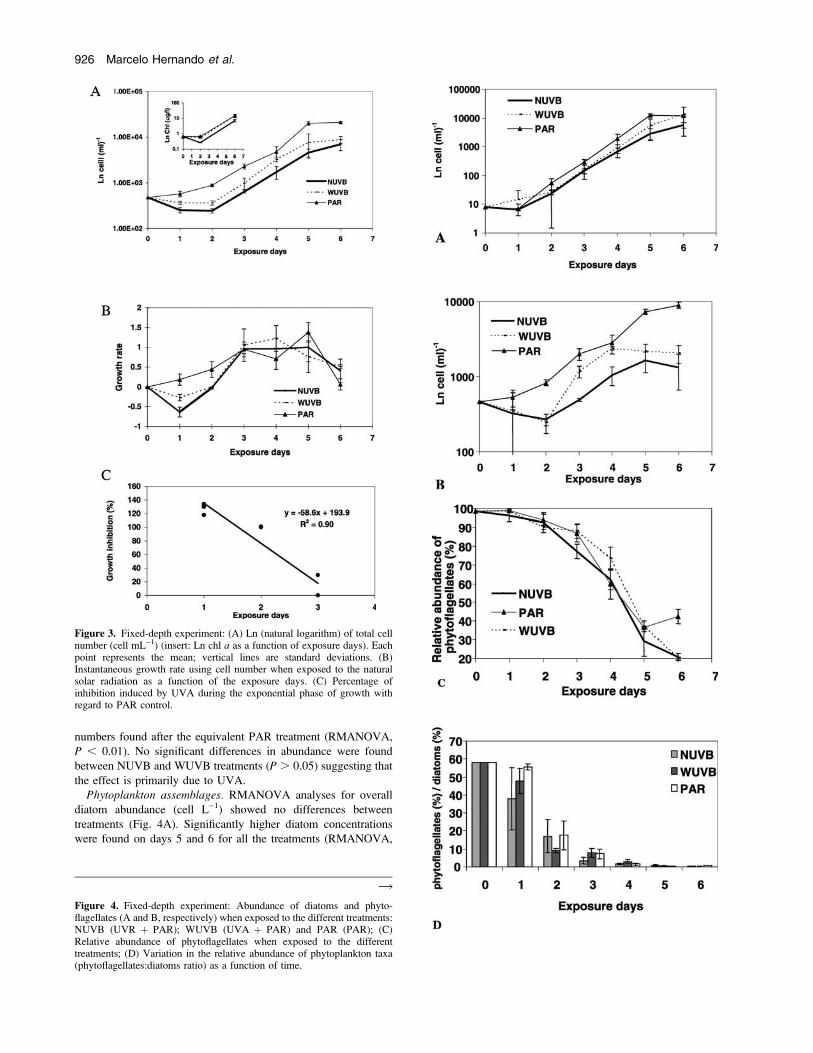
Phytoplankton biomass. Total cell number and chl a concentra-
tion showed a positive and significant correlation (Fig. 3A insert;
R 5 0.88, P , 0.001). Because a more complete data set is avail-
able for cell number than for chl a, biomass will hereafter be
expressed in cells mL�1.
Cell number increased from the beginning of the experiment,
when cells were exposed only to PAR irradiance (Fig. 3A). In
contrast, cell numbers decreased under UVR conditions on days 1
and 2 in both NUVB and WUVB treatments. After that, there was
an increase in abundance in both NUVB and WUVB treatments
until the end of the experiment. The exposure of cells to UVA
radiation (WUVB treatment) reduced cell number compared with
Figure 2. Variation in the daily doses of (A) UVB (280–320 nm) andUVA (320–400 nm), and (B) PAR (400–700 nm) at the sampling site,Ushuaia. Fixed-depth experiments were carried out 11–17 February 1998(period denoted by the thick line). Light measurements are included fora time before the experiments.
Photochemistry and Photobiology, 2006, 82 925
numbers found after the equivalent PAR treatment (RMANOVA,
P , 0.01). No significant differences in abundance were found
between NUVB and WUVB treatments (P . 0.05) suggesting that
the effect is primarily due to UVA.
Phytoplankton assemblages. RMANOVA analyses for overall
diatom abundance (cell L�1) showed no differences between
treatments (Fig. 4A). Significantly higher diatom concentrations
were found on days 5 and 6 for all the treatments (RMANOVA,
Figure 3. Fixed-depth experiment: (A) Ln (natural logarithm) of total cellnumber (cell mL�1) (insert: Ln chl a as a function of exposure days). Eachpoint represents the mean; vertical lines are standard deviations. (B)Instantaneous growth rate using cell number when exposed to the naturalsolar radiation as a function of the exposure days. (C) Percentage ofinhibition induced by UVA during the exponential phase of growth withregard to PAR control.
Figure 4. Fixed-depth experiment: Abundance of diatoms and phyto-flagellates (A and B, respectively) when exposed to the different treatments:NUVB (UVR þ PAR); WUVB (UVA þ PAR) and PAR (PAR); (C)Relative abundance of phytoflagellates when exposed to the differenttreatments; (D) Variation in the relative abundance of phytoplankton taxa(phytoflagellates:diatoms ratio) as a function of time.
!
926 Marcelo Hernando et al.
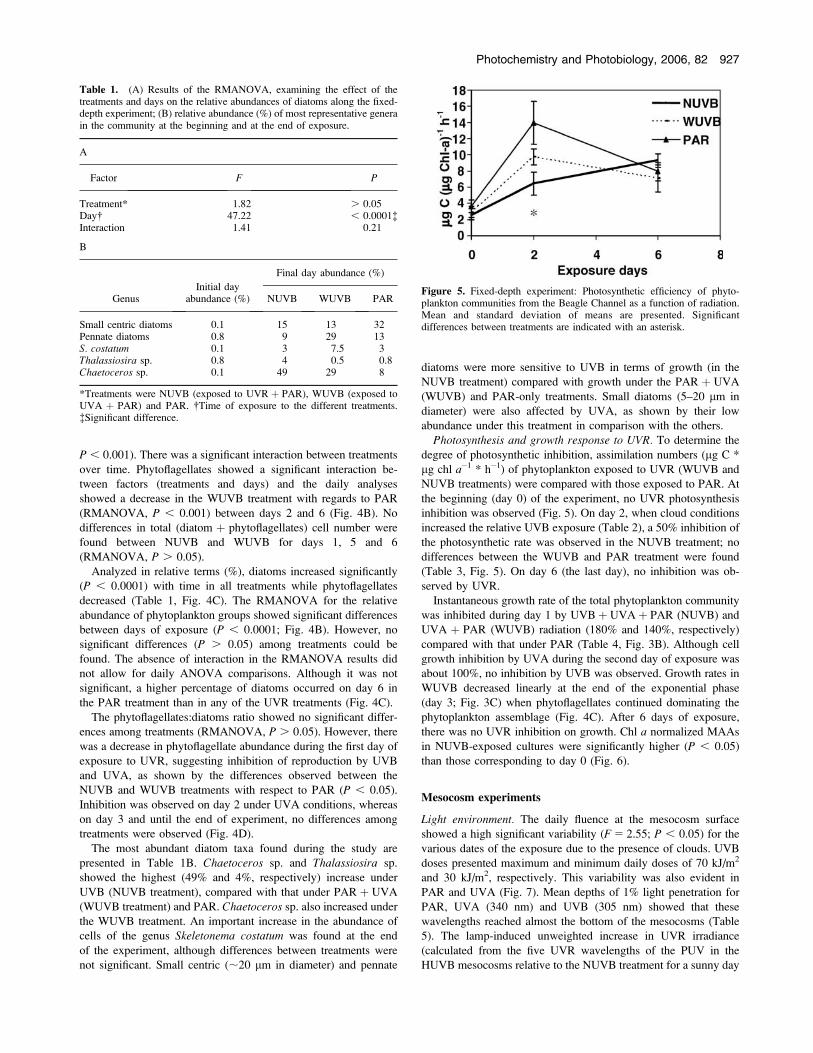
P , 0.001). There was a significant interaction between treatments
over time. Phytoflagellates showed a significant interaction be-
tween factors (treatments and days) and the daily analyses
showed a decrease in the WUVB treatment with regards to PAR
(RMANOVA, P , 0.001) between days 2 and 6 (Fig. 4B). No
differences in total (diatom þ phytoflagellates) cell number were
found between NUVB and WUVB for days 1, 5 and 6
(RMANOVA, P . 0.05).
Analyzed in relative terms (%), diatoms increased significantly
(P , 0.0001) with time in all treatments while phytoflagellates
decreased (Table 1, Fig. 4C). The RMANOVA for the relative
abundance of phytoplankton groups showed significant differences
between days of exposure (P , 0.0001; Fig. 4B). However, no
significant differences (P . 0.05) among treatments could be
found. The absence of interaction in the RMANOVA results did
not allow for daily ANOVA comparisons. Although it was not
significant, a higher percentage of diatoms occurred on day 6 in
the PAR treatment than in any of the UVR treatments (Fig. 4C).
The phytoflagellates:diatoms ratio showed no significant differ-
ences among treatments (RMANOVA, P . 0.05). However, there
was a decrease in phytoflagellate abundance during the first day of
exposure to UVR, suggesting inhibition of reproduction by UVB
and UVA, as shown by the differences observed between the
NUVB and WUVB treatments with respect to PAR (P , 0.05).
Inhibition was observed on day 2 under UVA conditions, whereas
on day 3 and until the end of experiment, no differences among
treatments were observed (Fig. 4D).
The most abundant diatom taxa found during the study are
presented in Table 1B. Chaetoceros sp. and Thalassiosira sp.
showed the highest (49% and 4%, respectively) increase under
UVB (NUVB treatment), compared with that under PAR þ UVA
(WUVB treatment) and PAR. Chaetoceros sp. also increased under
the WUVB treatment. An important increase in the abundance of
cells of the genus Skeletonema costatum was found at the end
of the experiment, although differences between treatments were
not significant. Small centric (;20 lm in diameter) and pennate
diatoms were more sensitive to UVB in terms of growth (in the
NUVB treatment) compared with growth under the PAR þ UVA
(WUVB) and PAR-only treatments. Small diatoms (5–20 lm in
diameter) were also affected by UVA, as shown by their low
abundance under this treatment in comparison with the others.
Photosynthesis and growth response to UVR. To determine the
degree of photosynthetic inhibition, assimilation numbers (lg C *
lg chl a�1 * h�1) of phytoplankton exposed to UVR (WUVB and
NUVB treatments) were compared with those exposed to PAR. At
the beginning (day 0) of the experiment, no UVR photosynthesis
inhibition was observed (Fig. 5). On day 2, when cloud conditions
increased the relative UVB exposure (Table 2), a 50% inhibition of
the photosynthetic rate was observed in the NUVB treatment; no
differences between the WUVB and PAR treatment were found
(Table 3, Fig. 5). On day 6 (the last day), no inhibition was ob-
served by UVR.
Instantaneous growth rate of the total phytoplankton community
was inhibited during day 1 by UVBþ UVAþ PAR (NUVB) and
UVA þ PAR (WUVB) radiation (180% and 140%, respectively)
compared with that under PAR (Table 4, Fig. 3B). Although cell
growth inhibition by UVA during the second day of exposure was
about 100%, no inhibition by UVB was observed. Growth rates in
WUVB decreased linearly at the end of the exponential phase
(day 3; Fig. 3C) when phytoflagellates continued dominating the
phytoplankton assemblage (Fig. 4C). After 6 days of exposure,
there was no UVR inhibition on growth. Chl a normalized MAAs
in NUVB-exposed cultures were significantly higher (P , 0.05)
than those corresponding to day 0 (Fig. 6).
Mesocosm experiments
Light environment. The daily fluence at the mesocosm surface
showed a high significant variability (F 5 2.55; P , 0.05) for the
various dates of the exposure due to the presence of clouds. UVB
doses presented maximum and minimum daily doses of 70 kJ/m2
and 30 kJ/m2, respectively. This variability was also evident in
PAR and UVA (Fig. 7). Mean depths of 1% light penetration for
PAR, UVA (340 nm) and UVB (305 nm) showed that these
wavelengths reached almost the bottom of the mesocosms (Table
5). The lamp-induced unweighted increase in UVR irradiance
(calculated from the five UVR wavelengths of the PUV in the
HUVB mesocosms relative to the NUVB treatment for a sunny day
Table 1. (A) Results of the RMANOVA, examining the effect of thetreatments and days on the relative abundances of diatoms along the fixed-depth experiment; (B) relative abundance (%) of most representative generain the community at the beginning and at the end of exposure.
A
Factor F P
Treatment* 1.82 . 0.05Day� 47.22 , 0.0001�Interaction 1.41 0.21
B
GenusInitial day
abundance (%)
Final day abundance (%)
NUVB WUVB PAR
Small centric diatoms 0.1 15 13 32Pennate diatoms 0.8 9 29 13S. costatum 0.1 3 7.5 3Thalassiosira sp. 0.8 4 0.5 0.8Chaetoceros sp. 0.1 49 29 8
*Treatments were NUVB (exposed to UVR þ PAR), WUVB (exposed toUVA þ PAR) and PAR. �Time of exposure to the different treatments.�Significant difference.
Figure 5. Fixed-depth experiment: Photosynthetic efficiency of phyto-plankton communities from the Beagle Channel as a function of radiation.Mean and standard deviation of means are presented. Significantdifferences between treatments are indicated with an asterisk.
Photochemistry and Photobiology, 2006, 82 927
at noon) was 1.72 times greater than natural irradiance. When
weighted with several biological weighting functions (BWFs) for
inhibition of photosynthesis, this relative increase was greater by
3.67-fold to 4.16-fold (47). These values are slightly high but in the
same range as some of the previous studies in mesocosms (48).
Phytoplankton assemblages. Microscopic results identified three
major groups: diatoms, small phytoflagellates and the eugleno-
phyte Eutreptiella sp. These groups correspond relatively well with
the pigment chemotaxonomy presented by Roy et al. (this issue).
The increase in diatom abundance coincided with the increase in
concentration of fucoxanthin (Roy et al., this issue). The highest
increase was observed in the abundance of ,3 lm phytoflagellates
and in the euglenophyte Eutreptiella sp. (11 lm wide 3 55 lm
long) in both HUVB and NUVB treatments (Fig. 8B,C). This
augmentation was higher in the HUVB treatment (Fig. 8B) than in
the NUVB treatment (Fig. 8C). In the latter treatment, the relative
abundance of Eutreptiella sp. was higher than in the other treat-
ments from day 5 to day 10.
Maximum phytoplankton abundance (as chl a concentration)
was observed on days 4–6, decreasing thereafter. No significant
differences between treatments were found (RMANOVA, P .
0.05; Fig. 9A).
The ratio of total chl a:chl a .5 lm showed an increase of algae
.5 lm between days 1 and 3 in all three treatments. In addition,
between days 3 and 5 the abundance of large-size (,5 lm) algae
increased in the HUVB treatment compared with abundances
after NUVB and WUVB treatments (Fig. 9B). On day 7, the ratio
increased without significant differences between treatments.
Because total chl a decreased on this day, the increase in the ratio
can be associated with a concomitant decrease in phytoflagellates
(Fig. 8).
Photosynthesis and growth response to UVR. No significant
differences between treatments (P . 0.05) were found in the in-
stantaneous growth rate of the phytoplankton community (calcu-
lated from chl a values) (RMANOVA, P . 0.05; Fig. 8A).
To determine photosynthetic inhibition by UVB, assimilation
numbers (lg C * lg chl a�1 * h�1) of phytoplankton exposed to
UVB (HUVB and NUVB treatments) were compared with phyto-
plankton exposed to WUVB treatment (Table 6). On day 1, ex-
posure to NUVB produced a 51% inhibition in photosynthesis with
regards to the WUVB treatment (Fig. 10). Higher assimilation
Table 3. (A) Results of the RMANOVA where the effect of thetreatments (NUVB, WUVB and PAR; see text) and time on algal carbonassimilation is evaluated during the fixed-depth experiment; (B) wheninteraction was significant (P , 0.05) each factor was analyzed by a one-factor ANOVA.
A
Factor F P
Treatment 10.59 , 0.01*Day 52.1 , 0.0001*Interaction 7.6 , 0.001*Day 0 1.57 . 0.05Day 2 8.59 , 0.01*Day 6 2.19 . 0.05
B
Average Homogeneity group�
Day 2, NUVB 0.25 ADay 2, WUVB 0.31 ADay 2, PAR 0.38 B
*Significant difference. �The same letter indicates that no significant dif-ferences (P . 0.05) were found (Tukey test).
Table 2. Experimental doses received by the phytoplankton assemblagesduring the days corresponding to the productivity studies during thefixed-depth experiment.
Exposure day UVB (kJ/m2) UVA (kJ/m2) PAR (kJ/m2)
0 29 404 35462 37 417 53206 28 447 4322
Table 4. (A) Results of the RMANOVA examining the effect of thetreatments and days on the instantaneous growth rate of the totalcommunity along the fixed-depth experiment; (B) when the interactionwas significant (P , 0.05), each factor was analyzed by a one-factorANOVA.
A
Factor F P
Treatment* 30.37 , 0.01�Day� 61.82 , 0.001�Interaction 3.03 , 0.05�Day 1 35.1 , 0.001�Day 2 12.24 , 0.01�Day 3 0.15 . 0.05
B
Average Homogeneity group§
Day 1, NUVB �0.63 ADay 1, WUVB �0.27 BDay 1, PAR 0.18 CDay 2, NUVB �0.03 ADay 2, WUVB �0.01 ADay 2, PAR 0.44 B
*Treatments were NUVB (exposed to UVR þ PAR), WUVB (exposed toUVA þ PAR) and PAR. �Time of exposure to the different treatments.�Significant difference. §The same letter indicates that no significant dif-ferences (P . 0.05) were found (Tukey test).
Figure 6. Fixed-depth experiment: Concentration of photoprotectiveMAAs measured at 334 nm optical depth (estimated as the height in mmof peak maximum from the spectrophotometric scan of a methanol extract)at the beginning and the end of the experiment, for the different radiationtreatments, normalized to total chl a. Mean and standard deviation of meansare presented.
928 Marcelo Hernando et al.
numbers were evident on day 5 in the HUVB treatment. On day 6,
only NUVB samples were inhibited. At this time, the NUVB
samples were suddenly exposed to higher PAR and UVR levels
due to less cloudiness (Table 7). As in the previous fixed-depth
experiments, no differences were observed between treatments on
the last incubation days.
The chl a normalized total concentration of MAAs did not show
significant differences between treatments. The two treatments
exposed to UVB (NUVB and HUVB), showed higher values on
day 3. However, a decrease was observed between days 3 and 5,
after which concentrations remained low (Fig. 11). In contrast, the
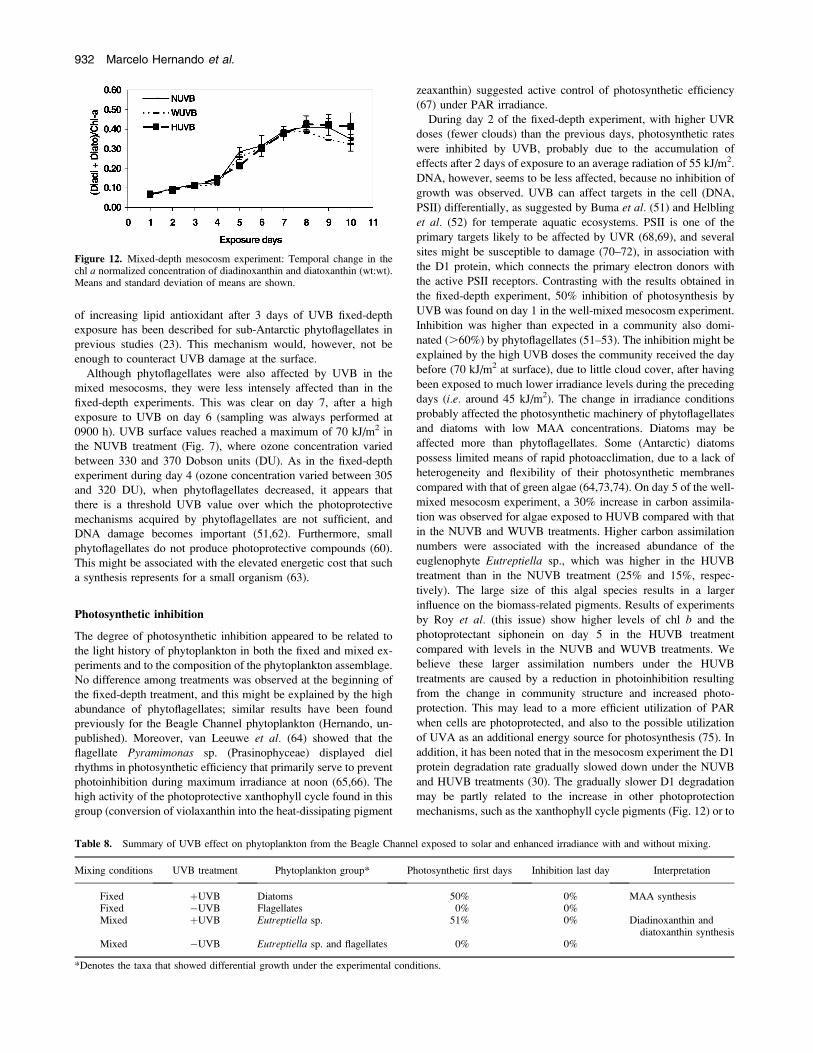
photoprotective pigments of the xanthophyll cycle, diadinoxanthin
and diatoxanthin, showed an increase in their chlorophyll-specific
concentration with time (Fig. 12), but no significant differences
between treatments was found.
DISCUSSION
Under both fixed and mixed conditions, a strong inhibition in
photosynthesis was observed at the beginning of the experiment.
This inhibition was no longer observed by the end of the experi-
ments (Table 8). Reasons for this change differ among experi-
ments. Moreover, phytoplankton composition changed during the
experiment, and the change in the dominant microalgae group in
each case contributes to explain the differential adaptation to UVB
observed. In the fixed experiments, the initial phytoflagellate-
dominated community changed toward one dominated by dia-
toms. In the mixed experiments, both small phytoflagellates and
the euglenophyte Eutreptiella sp. were present throughout the
experiment. Diatoms synthesize MAAs as a response to UVR;
Eutreptiella has the capacity to produce changes in the xanthophyll
pigments, and thereby defend itself from UVB damage (Table 8).
Changes in the community structure and physiologicaladaptations in response to UVR
Several days of exposure to high UVB may induce changes in the
dominant species, from less to more tolerant ones (49,50). The
colony-forming Thalassiosira sp. grew differentially at the end of
the fixed-depth experiment. This genus has been well studied in
Antarctic environments, where it has been shown that it is capable
of fast synthesis of photoprotective compounds (21). Other centric
diatoms were, however, more sensitive to UVB, especially the
smaller (,30 lm) ones. Small cells—with a relatively high surface
to volume ratio—are more resistant to photosynthesis inhibition but
more vulnerable to DNA damage (51–53). The balance between
these two processes will determine their abundance under both
UVA and UVB exposure. Genera such as Chaetoceros, changing
from 0.1% to 49% under NUVB, and only to 8% under PAR, show
a great capacity to adapt to these radiations, probably due to
morphological changes (54) and to a great DNA repair potential
(i.e. in 6 h).
A progressive decrease in the ratio of phytoflagellates to diatoms
under UVB treatment was observed in the fixed-depth experiments.
In unialgal cultures, Hernando and San Roman (35) showed that
Figure 7. Variation in daily doses of (A) UVB (280–320 nm) and UVA(320–400 nm), and (B) PAR (400–700 nm) at the sampling site, Ushuaia.Well-mixed experiments were carried out 5–15 November 2001 (denotedby black line). Light measurements are included for a time before theexperiments.
Table 5. PAR, UVA (340 nm) and UVB (305 nm) conditions for surfaceincubations.
Variable
Experimental day
9 November(day 4)
12 November(day 7)
Daily fluence at surface*
PAR (kJ/m2) 50.59 47.47UVA (340 nm) (kJ/m2) 5.8 5.6NUV-B (305 nm) (kJ/m2) 0.3 0.3HUV-B (305 nm) (kJ/m2) 2.4 2.5
Daily fluence averaged over the water column
PAR (kJ/m2) 26.72 27.60UVA (340 nm) (kJ/m2) 1.7 1.7NUV-B (305 nm) (kJ/m2) 0.06 0.06HUV-B (305 nm) (kJ/m2) 0.40 0.39
Kd (m�1)�PAR 0.6 0.5UVA (340 nm) 1.4 1.3UVB (305 nm) 1.9 1.8
Z1% (m)�PAR 7.7 9.2UVA (340 nm) 3.3 3.5UVB (305 nm) 2.4 2.6
*Daily fluence at surface was corrected for the lamp shading effect. Thevalues presented are not wavelength-integrated and correspond to eachwavelength indicated. �Kd is light attenuation coefficient. �Z1% is depthof the euphotic zone.
Photochemistry and Photobiology, 2006, 82 929
sub-Antarctic phytoflagellates are extremely sensitive to UVB, in
contrast to diatoms of the genus Navicula. Therefore, in addition to
a higher tolerance of diatoms to UVR, phytoflagellate diminution
left an open niche for diatoms to occupy.
MAA concentrations in the fixed-depth incubation experiments
were higher under NUVB than in the other treatments, coinciding
with an increase in diatom abundance (i.e. day 6). The same trend
was observed in the mixed mesocosms, where the HUVB treatment
showed a maximum in the MAA to chl a ratio for day 7 (Fig. 11),
coinciding with a relative decrease in phytoflagellate and diatom
abundance (Fig. 9B), whereas euglenophytes (Eutreptiella sp.)
increased.
Effects of UVR exposure on phytoplankton fromsurface waters
Fixed-depth experiments simulate the environmental conditions to
which microalgae are exposed in stratified surface waters. The
relative contribution of UVA and UVB to the instantaneous growth
rate of the community varied throughout the time of exposure.
UVA was responsible for most of the observed inhibition in the
fixed-depth experiment, except during the first day when the
inhibition was produced by UVB. In addition, we found that such
UVA inhibition decreased with time, suggesting that cells had
acclimated to the new radiation conditions. UVA inhibition of
growth can be observed in the fixed-depth experiment during day 2
as shown for some other regions (55–57). This can be due to
a larger amount of UVA reaching the Earth even though UVB
causes more damage per unit energy (58). Cell number decreased
in the WUVB treatments while it increased in the PAR treatments,
although this inhibition ceased on day 3, at the end of the
exponential growth phase. Then, the UVB increase due to the
reduced cloud cover caused damage to phytoflagellates, allowing
the better adapted diatoms (probably due to the synthesis of
MAAs) to proliferate. Since no photosynthesis inhibition (i.e.reduction in carbon incorporation) was observed during the first
exposure days, phytoflagellates of this community would seem to
be resistant to UVB. However, these organisms were susceptible to
DNA damage, as denoted by growth inhibition.
Effects of UVR exposure in mixed mesocosms
Mixed situations are also common in Patagonian coasts, where
a shallow pycnocline can be disrupted by high-intensity winds. In
Figure 8. Mixed-depth mesocosm experiment: (A) Instantaneous growthrate using chl a L�1 when exposed to the natural solar radiation as a functionof the exposure days. (B) Total cell number of centric diatoms, Eutreptiellasp. and phytoflagellates in Ln (natural logarithm) cell number mL�1 forHUVB treatment. (C) Total cell number of centric diatoms, Eutreptiella sp.and phytoflagellates in Ln cell number mL�1 for NUVB treatment. Eachpoint represents the mean and vertical bars are standard deviation.
Figure 9. Mixed-depth mesocosm experiment: (A) Relative abundance ofphytoplankton communities using chl a concentration by L�1 when exposedto the different treatments: NUVB (UVRþ PAR); WUVB (UVAþ PAR)and PAR (PAR); (B) Variation in the relative abundance (ratio of chl atotal:chl a .5 um) as a function of time.
930 Marcelo Hernando et al.
contrast with the results obtained for fixed-depth incubations, the
relative abundance of diatoms and phytoflagellates in the mixed
mesocosms did not show significant differences among treatments,
although lower abundance in phytoflagellates can be observed
when comparing the initial and final values. However, an increase
in Eutreptiella sp. is evident in the HUVB treatment. The presence
of Eutreptiella sp. can additionally be traced by the pigment
siphonein (Roy et al. in this issue). This is a derivative of b-
carotene, which acts as a pigment antenna in the photosynthesis
process (23) and is an antioxidant, protecting cells against
oxidative damage initiated by singlet oxygen (59).
Furthermore, the persistence of phytoflagellates in mesocosms
has an explanation in that when cells are mixed below the UVR
damaging zone, they can reduce damage either by activating DNA
and photosynthesis repair, and induce various photoprotective
mechanisms. Active repair of DNA damage seems to be effective
in phytoplankton assemblages from Patagonia, as seen in studies
carried out by Buma et al. (51) and Helbling et al. (52). Helbling
et al. (60) have shown the size dependence of DNA damage in
phytoplankton from Patagonian coastal waters, with small cells
(,2 um) being more sensitive to UVR than large cells. This is in
agreement with other studies carried out by Boelen et al. (61) and
Karentz et al. (54), which also suggest that DNA damage is
associated with the size structure of the community. Helbling et al.(52) showed that cyclobutane pyrimidine dimers (CPDs), an index
of DNA damage, accumulated in the surface (0.5 m), whereas no
accumulation was observed at depths of 3 and 6 m, in accordance
with the strong attenuation of DNA-effective UVR. The presence
Table 6. (A) Results of the RMANOVA where the effect of thetreatments (NUVB, WUVB and PAR; see text) and time on algal carbonassimilation for the different days is evaluated during the mixed mesocosmsexperiments; (B and C) when interaction was significant (P , 0.05) eachfactor was analyzed by a one factor ANOVA.
A
Factor F P
Treatment 1.5 . 0.05Day 87.57 , 0.000001*Interaction 2.55 , 0.01*
B
Day 1 11.98 , 0.01*Day 2 0.16 . 0.05Day 3 0.08 . 0.05Day 5 7.66 , 0.01*Day 6 8.74 , 0.01*Day 7 3.49 . 0.05Day 8 3.5 . 0.05Day 9 0.46 . 0.05Day 10 0.64 . 0.05
C
Average Homogeneity group�
Day 1
WUVB 2.76 BNUVB 1.35 A
Day 5
WUVB 1.66 ACNUVB 1.96 ACHUVB 2.34 AB
Day 6WUVB 1.03 BNUVB 0.71 AHUVB 1.09 Bc
*Significant difference. �The same letter indicates that no significant dif-ferences (P . 0.05) were found (Tukey test).
Figure 10. Mixed-depth mesocosm experiment: Photosynthetic efficiencyof phytoplankton communities from the Beagle Channel as a function ofradiation. Mean and standard deviation of means are represented.Significant differences between treatments are indicated with an asterisk.HUVB treatment started on day 3.
Table 7. Experimental doses (0.2 m depth and 6 h integrated) thatphytoplankton assemblages received during the days corresponding to theproductivity studies in the well-mixed mesocosm experiment.
Exposure day UVB (kJ/m2) UVA (kJ/m2) PAR (kJ/m2)
1 19 444 41972 22 441 41443 31 660 67605 22 419 40786 45 843 93477 43 796 87308 33 659 68829 22 474 4690
10 26 508 4974
Figure 11. Mixed-depth mesocosm experiment: Temporal change in thechl a normalized total concentration of MAAs (lg MAAs * lg chl a�1).Mean and standard deviation of means are shown.
Photochemistry and Photobiology, 2006, 82 931
of increasing lipid antioxidant after 3 days of UVB fixed-depth
exposure has been described for sub-Antarctic phytoflagellates in
previous studies (23). This mechanism would, however, not be
enough to counteract UVB damage at the surface.
Although phytoflagellates were also affected by UVB in the
mixed mesocosms, they were less intensely affected than in the
fixed-depth experiments. This was clear on day 7, after a high
exposure to UVB on day 6 (sampling was always performed at
0900 h). UVB surface values reached a maximum of 70 kJ/m2 in
the NUVB treatment (Fig. 7), where ozone concentration varied
between 330 and 370 Dobson units (DU). As in the fixed-depth
experiment during day 4 (ozone concentration varied between 305
and 320 DU), when phytoflagellates decreased, it appears that
there is a threshold UVB value over which the photoprotective
mechanisms acquired by phytoflagellates are not sufficient, and
DNA damage becomes important (51,62). Furthermore, small
phytoflagellates do not produce photoprotective compounds (60).
This might be associated with the elevated energetic cost that such
a synthesis represents for a small organism (63).
Photosynthetic inhibition
The degree of photosynthetic inhibition appeared to be related to
the light history of phytoplankton in both the fixed and mixed ex-
periments and to the composition of the phytoplankton assemblage.
No difference among treatments was observed at the beginning of
the fixed-depth treatment, and this might be explained by the high
abundance of phytoflagellates; similar results have been found
previously for the Beagle Channel phytoplankton (Hernando, un-
published). Moreover, van Leeuwe et al. (64) showed that the
flagellate Pyramimonas sp. (Prasinophyceae) displayed diel
rhythms in photosynthetic efficiency that primarily serve to prevent
photoinhibition during maximum irradiance at noon (65,66). The
high activity of the photoprotective xanthophyll cycle found in this
group (conversion of violaxanthin into the heat-dissipating pigment
zeaxanthin) suggested active control of photosynthetic efficiency
(67) under PAR irradiance.
During day 2 of the fixed-depth experiment, with higher UVR
doses (fewer clouds) than the previous days, photosynthetic rates
were inhibited by UVB, probably due to the accumulation of
effects after 2 days of exposure to an average radiation of 55 kJ/m2.
DNA, however, seems to be less affected, because no inhibition of
growth was observed. UVB can affect targets in the cell (DNA,
PSII) differentially, as suggested by Buma et al. (51) and Helbling
et al. (52) for temperate aquatic ecosystems. PSII is one of the
primary targets likely to be affected by UVR (68,69), and several
sites might be susceptible to damage (70–72), in association with
the D1 protein, which connects the primary electron donors with
the active PSII receptors. Contrasting with the results obtained in
the fixed-depth experiment, 50% inhibition of photosynthesis by
UVB was found on day 1 in the well-mixed mesocosm experiment.
Inhibition was higher than expected in a community also domi-
nated (.60%) by phytoflagellates (51–53). The inhibition might be
explained by the high UVB doses the community received the day
before (70 kJ/m2 at surface), due to little cloud cover, after having
been exposed to much lower irradiance levels during the preceding
days (i.e. around 45 kJ/m2). The change in irradiance conditions
probably affected the photosynthetic machinery of phytoflagellates
and diatoms with low MAA concentrations. Diatoms may be
affected more than phytoflagellates. Some (Antarctic) diatoms
possess limited means of rapid photoacclimation, due to a lack of
heterogeneity and flexibility of their photosynthetic membranes
compared with that of green algae (64,73,74). On day 5 of the well-
mixed mesocosm experiment, a 30% increase in carbon assimila-
tion was observed for algae exposed to HUVB compared with that
in the NUVB and WUVB treatments. Higher carbon assimilation
numbers were associated with the increased abundance of the
euglenophyte Eutreptiella sp., which was higher in the HUVB
treatment than in the NUVB treatment (25% and 15%, respec-
tively). The large size of this algal species results in a larger
influence on the biomass-related pigments. Results of experiments
by Roy et al. (this issue) show higher levels of chl b and the
photoprotectant siphonein on day 5 in the HUVB treatment
compared with levels in the NUVB and WUVB treatments. We
believe these larger assimilation numbers under the HUVB
treatments are caused by a reduction in photoinhibition resulting
from the change in community structure and increased photo-
protection. This may lead to a more efficient utilization of PAR
when cells are photoprotected, and also to the possible utilization
of UVA as an additional energy source for photosynthesis (75). In
addition, it has been noted that in the mesocosm experiment the D1
protein degradation rate gradually slowed down under the NUVB
and HUVB treatments (30). The gradually slower D1 degradation
may be partly related to the increase in other photoprotection
mechanisms, such as the xanthophyll cycle pigments (Fig. 12) or to
Figure 12. Mixed-depth mesocosm experiment: Temporal change in thechl a normalized concentration of diadinoxanthin and diatoxanthin (wt:wt).Means and standard deviation of means are shown.
Table 8. Summary of UVB effect on phytoplankton from the Beagle Channel exposed to solar and enhanced irradiance with and without mixing.
Mixing conditions UVB treatment Phytoplankton group* Photosynthetic first days Inhibition last day Interpretation
Fixed þUVB Diatoms 50% 0% MAA synthesisFixed �UVB Flagellates 0% 0%Mixed þUVB Eutreptiella sp. 51% 0% Diadinoxanthin and
diatoxanthin synthesisMixed �UVB Eutreptiella sp. and flagellates 0% 0%
*Denotes the taxa that showed differential growth under the experimental conditions.
932 Marcelo Hernando et al.

the presence of siphonein, as mentioned above. The 30% inhibition
observed on day 6 in the NUVB treatment compared with that of
the other two treatments coincided with the highest UVB value
measured (45 kJ/m2, twice as much as in the previous day) due to
a reduced cloud cover.
By day 7, Eutreptiella sp. abundance doubled and phytoflagel-
lates decreased. Eutreptiella sp., already protected from UVR,
continued growing and showed no photosynthetic inhibition under
a high UVR dose, similar to that of day 6. Because the xanthophyll
cycle of euglenophytes uses diadinoxanthin and diatoxanthin, in
contrast to other chl b–containing algae (76), the high level of these
photoprotective pigments in the HUVB treatment (Fig. 12) con-
firms the role of photoprotection.
CONCLUSIONS
Fixed and mixed conditions represent two possible scenarios in
coastal sub-Antarctic waters seasonally exposed to the ozone hole,
where coastal waters can be both stratified or mixed, and therefore
present different UVR environments for phytoplankton. As shown
in Tables 3, 4 and 6, local microalgae can acclimate to high
irradiance conditions, for both growth rate and photosynthesis, in
agreement with other studies from this region (18,77). The struc-
ture of the phytoplankton community seems to play a central role
in this response, both under fixed-depth and mixed conditions.
Phytoflagellates, diatoms and other groups can grow differentially
in either scenario, developing different (group-specific) defense
and protection mechanisms. Furthermore, the contrast between
fixed-depth and mixed experiments showed that species can cope
with UVR. Under fixed light, MAA synthesis seems the most
effective photoprotective mechanism. Under variable light con-
ditions, Eutreptiella sp. was successfully protected by the synthesis
of diadinoxanthin and diatoxanthin.
Acknowledgements—This research was supported by a team grant fromthe Inter American Institute for Global Change (M. Vernet, lead principalinvestigator), by grants from the Natural Sciences and Engineering Coun-cil of Canada (NSERC) to Demers et al. (Special Research Opportunity)and partially by student stipends from CONICET, Argentina. The scintil-lation counter was provided by Universidad de Buenos Aires. We thankthe technical teams, Oscar Gonzales for his help with the incubations inmesocosms and Miguel Barbagallo for technical assistance with data col-lection during fixed-depth experiments and Leonardo Cantoni for lightdata processing. We especially thank Dr. San Roman for the use of hislaboratory installations and equipment.
REFERENCES
1. Villafane, V. E., K. Sundback, F. L. Figueroa and E. W. Helbling(2003) Photosynthesis in the aquatic environment as affected by UVR.In The Royal Society of Chemistry, Cambridge (Edited by E. W.Helbling and H. E. Zagarese), pp. 357–397. Cambridge Press, UK.
2. Buma, A. G. J., P. Boelen and W. H. Jeffrey (2003) UVR- inducedDNA damage in aquatic organisms. In The Royal Society of Chemistry,Cambridge (Edited by E. W. Helbling and H. E. Zagarese), pp.291–327. Cambridge Press, UK.
3. Franklin, L. A., I. Yakovleva, U. Karsten and K. Luning (1999)Synthesis of mycosporine-like amino acids in Chondrus crispus(Florideophyceae) and the consequences for sensitivity to ultravioletB radiation. J. Phycol. 35(4), 682–693.
4. Lesser, M. P. (1996) Acclimation of phytoplankton to UVB radiation:Oxidative stress and photoinhibition of photosynthesis are notprevented by UV absorbing compounds in the dinoflagellateProrocentrum micans. Mar. Ecol. Prog. Ser. 132, 287–297.
5. Smith, R. C. and K. S. Baker (1979) Penetration of UV-B andbiologically effective dose-rates in natural waters. Photochem. Photo-biol. 29, 311–323.
6. Smith, R. C. and K. S. Backer (1981) Optical properties of the clearestnatural waters (200–800 mm). Appl. Opt. 20, 177–184.
7. Mitchell, B. G. (1990) Action spectra of ultraviolet photoinhibition ofAntarctic phytoplankton and a model of spectral diffuse attenuationcoefficients. In Response of Marine Phytoplankton to NaturalVariations in UVBR Flux (Edited by B. G. Mitchel, O. Holm-Hansenand I. Sobolev), pp. 1–10. Scripps Institution of Oceanography, LaJolla, CA.
8. Holm-Hansen, O., E. W. Helbling, and D. Lubin (1993) Ultravioletradiation in Antarctica: Inhibition of primary production. Photochem.Photobiol. 58, 567–570.
9. Smith, R. C., K. S. Backer, O. Holm-Hansen and R. Olson (1980)Photoinhibition of photosynthesis in natural waters. Photochem.Photobiol. 31, 585–592.
10. Holm-Hansen, O., B. G. Mitchell and M. Vernet (1989) Ultravioletradiation in Antarctic waters: Effects on rates of primary production.Antarct. J. U. S. 24, 177–178.
11. Smith, R. C., B. B. Prezelin, K. S. Baker, R. R. Bidigare, N. P.Boucher, T. Coley, D. Karentz, S. MacIntyre, H. A. Matlick, D.Menzies, M. Ondrusek, Z. Wan and K. J. Waters (1992) Ozonedepletion: Ultraviolet radiation and phytoplankton biology in Antarcticwaters. Science 255, 952–959.
12. Cullen, J. J. and M. P. Lesser (1991) Inhibition of photosynthesis byultraviolet radiation as a function of dose and dosage rate: Resultsfor a marine diatom. Mar. Biol. 111, 183–190.
13. Neale, P. J., A. T. Banaszak and C. R. Jarriel (1998) Ultravioletsunscreens in Gymnodinium sanguineum (Dinophyceae) mycosporine-like amino acids protect against inhibition of photosynthesis. J. Phycol.34, 928–938.
14. Neale, P. J., E. W. Helbling and H. E. Zagarese (2002) Modulation ofUVR exposure and effects by vertical mixing and advection. InEuropean Society for Photobiology, Comprehensive Series in Photo-sciences, Vol. 1 (Edited by D.-P. Hader and G. Jori), pp. 108–134. TheRoyal Society of Chemistry, Cambridge, UK.
15. Ivanov, A. G., E. Miskiewicz, A. K. Clarke, B. M. Greenberg, andN. P. A. Huner (2000) Protection of photosystem II against UV-A andUV-B radiation in the cyanobacterium Plectonema boryanum: The roleof growth temperature and growth irradiance. Photochem. Photobiol.72, 772–779.
16. MacDonald, T. M., L. Dubois, L. C. Smith and D. A. Campbell (2003)Sensitivity of cyanobacterial antenna, reaction centre and CO2
assimilation transcripts and proteins to moderate UVB: Lightacclimation potentiates resistance to UVB. Photochem. Photobiol.77, 405–412.
17. Orce, L. V., N. A. San Roman, A. Paladini, C. Labraga and E. W.Helbling (1997) Multiple regression fit. Latitudinal UVR-PARmeasurements in Argentina: Extent of the ‘ozone hole’. GlobalPlanetary Change 15, 113–121.
18. Hader, D.-P., H. D. Kumar, R. C. Smith and R. C. Worrest (2003)Aquatic ecosystems: Effects of solar ultraviolet radiation andinteractions with other climatic change factors. Photochem. Photobiol.Sci. 2, 39–50.
19. Villafane, V. E., E. W. Hebling, O. Holm-Hansen and B. E. Chalker(1995) Acclimatization of Antarctic natural phytoplankton assemblageswhen exposed to solar ultraviolet radiation. J. Plankton Res. 17,2295–2306.
20. Carreto, J. I., M. O. Carignan, G. Daleo and S. G. De Marco (1990)Occurrence of mycosporine-like amino acids in the red-tide dinofla-gellate Alexandrium excavatum: UV-photoprotective compounds?J. Plankton Res. 12, 909–921.
21. Karsten, U., K. Bischof, D. Hanelt, H. Tug and C. Wiencke (1999) Theeffect of UV radiation on photosynthesis and UV-absorbing substancesin the endemic Arctic macroalga Develaraea ramentacea (Rhodophyta).Physiol. Plant 105, 58–66.
22. Hernando, M. P., J. I. Carreto, M. O. Carginan, G. A. Ferryra andC. Grob (2002) Effects of solar radiation on growth and mycosporine-like amino acids content in an Antarctic diatom. Polar Biol. 25, 12–20.
23. Hernando, M., G. Malanga and G. A. Ferreyra (2005) Oxidative stressand antioxidant defenses due UV radiation in sub-Antarctic marinephytoplankton. Sci. Mar. 69 (Suppl. 2), 287–295.
24. Mostajir, B., S. Demers, S. de Mora, C. Belzile, J.-P. Chanut, M.Gosselin, S. Roy, P. Z. Villegas, J. Fauchot, J. Bouchard, D. Bird, P.Monfort and M. Levasseur (1999) Experimental test of the effect of
Photochemistry and Photobiology, 2006, 82 933

ultraviolet-B radiation in a planktonic community. Limnol. Oceanogr.44, 586–596.
25. Kloser, H. (1996) Cruise report of the Joint Chilean-German-ItalianMagellan ‘‘Victor Hensen’’ Campaign in 1994. In Berichte ZurPolarforschung, Vol. 190 (Edited by W. Arntz and M. Gorny), pp. 18–19. Alfred Wegener Institute—Bremerhaven, Germany.
26. Villafane, V. E., E. W. Helbling and H. E. Zagarese (2001) Solarultraviolet radiation and its impact on aquatic systems of Patagonia,South America. Ambio 30, 112–117.
27. Helbling, E. W., V. E. Villafane and O. Holm-Hansen (1994) Effectsof ultraviolet radiation on Antarctic marine phytoplankton photosyn-thesis with particular attention to the influence of mixing. In Ultra-violet Radiation in Antarctica: Measurements and Biological Effects,American Geophysical Union, Antarctic Research Series, Vol. 62(Edited by C. S. Weiler and P. Penhale), pp. 207–227. Scripps Instituteof Oceanography, San Diego, CA.
28. Belzile, C., S. Demers, D. R. S. Lean, B. Mostajir, S. Roy, S. De Mora,D. Bird, M. Gosselin, J. P. Chanut and M. Levasseur (1998) Anexperimental tool to study the effects of ultraviolet radiation onplanktonic communities: A mesocosm approach. Environ. Technol.19, 667–682.
29. Wangberg, S.-A., K. Garde, K. Gustavson and J.-S. Selmer (1999)Effects of UVB radiation on marine phytoplankton communities.J. Plankton Res. 21, 147–166.
30. Bergmann, T., T. L. Richardson, H. W. Paerl, J. L. Pinckney andO. Schofield (2002) Synergy of light and nutrients on the photosyn-thetic efficiency of phytoplankton populations from the Neuse RiverEstuary, North Carolina. J. Plankton Res. 24, 923–933.
31. Guillard, R. L. (1975) Culture of phytoplankton for feeding marineinvertebrates. In Culture of Marine Invertebrates Animals (Editedby W. L. Smith and M. H. Chanley), pp. 29–60. Plenum Publishing,New York.
32. Bouchard, J. N., D. A. Campbell and S. Roy (2005) Effects of UVBradiation on the D1 protein repair cycle of natural phytoplanktoncommunities from three latitudes (Canada, Brazil, and Argentina).J. Phycol. 41, 273–286.
33. Dıaz, S., C. Camilion, K. Lacoste, J. Escobar, S. Demers, S. Gianesella,S. Roy and M. Vernet (2003) Simulation of increasing UV radiation asa consequence of ozone depletion. In Proceedings SPIE’s 48th AnnualMeeting. Ultraviolet Ground- and Space-based Measurements, Modelsand Effects III. (Ed. by J. Slusser, J. Herman and Wei Gao), pp. 216–227. SPIE The International Society for Optical Engineering.San Diego.
34. Riley, G. A. (1957) Phytoplankton of the North central Sargasso Sea.Limnol. Oceanogr. 2, 252–270.
35. Hernando, M. and N. San Roman (1999) Preliminary data on chroniceffects of ultraviolet radiation on the growth of some phytoplanktonspecies of the Beagle Channel, Argentina. Sci. Mar. 63 (Suppl. 1),81–88.
36. Steeman, N. (1952) The use of radiocarbon (14C) for measuring or-ganic production in the sea. J. Cons. Int. Explor. Mer. 18, 177–180.
37. Holm-Hansen, O. and E. W. Helbling (1995) Tecnicas para la medicionde la productividad primaria en el fitoplancton. In Manual de metodosficologicos (Edited by K. Alveal, M. E. Ferrario, E. C. Oliveira andE. Sar), pp. 329–350. Universidad de Concepcion, Concepcion, Chile.
38. Utermohl, H. (1958) Zur Vervollkommnung der quantitativer Phyto-plankton-methodik. Mittelbach Internationale Veriningung fur Theor-etische und Angewandte Limnologie 9, 1–38.
39. Holm-Hansen, O. and B. Riemann (1978) Chlorophyll a determination:Improvements in methodology. Oikos 30, 438–447.
40. Zapata, M., F. Rodrıguez and J. L. Garrido (2000) Separation ofchlorophylls and carotenoids from marine phytoplankton: A newHPLC method using a reversed phase C8 column and pyridine-containing mobile phases. Mar. Ecol. Prog. Ser. 195, 29–45.
41. Helbling, E. W., B. E. Chalker, W. C. Dunlap, O. Holm-Hansen andV. E. Villafane (1996) Photoacclimation of Antarctic marine diatomsto solar ultraviolet radiation. J. Exp. Mar. Biol. Ecol. 204, 85–101.
42. Dunlap, W. C., G. A. Rae, E. W. Helbling, V. E. Villafane and O.Holm-Hansen (1995) Ultraviolet-absorbing compounds in natural as-semblages of Antarctic phytoplankton, Antarct. J. U. S. 30, 323–326.
43. Laurion, I., F. Blouin and S. Roy (2003) The quantitative filtertechnique for measuring phytoplankton absorption: Interference byMAAs in the UV waveband. Limnol. Oceanogr. Methods 1, 1–9.
44. Shick, J. M., S. Romaine-Lioud, C. Ferrier-Pages and J.-P. Gattuso(1999) Ultraviolet-B radiation stimulates shikimate pathway-dependentaccumulation of mycosporine-like amino acids in the coral Stylophorapistillata despite decreases in its population of symbiotic dinoflagel-lates. Limnol. Oceanogr. 44, 1667–1681.
45. Tsujino, Y., K. Yabe and Y. Sekikawa (1980) Isolation and structureof a new amino acid, shinorine, from the red alga Chondrus yendoiYamada et Mikami. Bot. Mar. 23, 65–67.
46. Scheiner, S. M. and J. Gurevitch (2001) Design and Analysis ofEcological Experiments, 2nd ed. Oxford University Press, New York.
47. Longhi, M. L., G. Ferreyra, I. Schloss and S. Roy (2005) Variablephytoplankton response to enhanced uv-b and nitrate addition inmesocosm experiments at three latitudes (Canada, Brazil andArgentina). Mar. Ecol. Prog. Ser. 313, 57–72.
48. Mostajir, B., S. Demers, S. de Mora, C. Belzile, J.-P. Chanut, M.Gosselin, S. Roy, P. Z. Villegas, J. Fauchot, J. Bouchard, D. Bird, P.Monfort and M. Levasseur (1999) Experimental test of the effect ofultraviolet-B radiation in a planktonic community. Limnol. Oceanogr.44, 586–596.
49. Worrest, R. C., K. U. Wolniakowski, J. D. Scott, D. L. Brooker, B. E.Thomson and H. Van Dyke (1981) Sensitivity of marine phytoplanktonto UVB radiation: Impact upon a model ecosystem. Photochem.Photobiol. 33, 223–227.
50. Vernet, M. (2000) Effects of UV radiation on the physiology andecology of marine phytoplankton. In The Effects of UV Radiation inthe Marine Environment, Cambridge Environmental ChemistrySeries. (Edited by S. de Mora, S. Demers and M. Vernet),pp. 237–278. Cambridge University Press, UK.
51. Buma, A. G. J., E. W. Helbling, M. K. de Boer and V. E. Villafane(2001) DNA damage patterns in temperate South-Atlantic picophyto-plankton assemblages exposed to solar ultraviolet radiation. J.Photochem. Photobiol. B Biol. 62, 9–18.
52. Helbling, E. W., A. G. J. Buma, M. K. de Boer and V. E. Villafane(2001) In situ impact of solar ultraviolet radiation on photosynthesisand DNA in temperate marine phytoplankton. Mar. Ecol. Prog. Ser.211, 43–49.
53. Helbling, E. W., V. E. Villafane and E. S. Barbieri (2001) Sensitivityof winter phytoplankton communities from Andean lakes to artificialultraviolet-B radiation. Rev. Chil. Hist. Nat. 74, 273–282.
54. Karentz, D., J. E. Cleaver and D. L. Mitchell (1991) Cell survivalcharacteristics and molecular responses of Antarctic phytoplankton toultraviolet-B radiation. J. Phycol. 27, 326–341.
55. Kim, D. S. and Y. Watanabe (1993) The effect of long-wave ultravioletradiation (UV-A) on the photosynthetic activity of natural populationof freshwater phytoplankton. Ecol. Res. 8, 225–234.
56. Holm-Hansen, O., V. E. Villafane and E. W. Helbling (1997) Effects ofsolar ultraviolet radiation on primary production in Antarctic waters. InAntarctic Communities: Species, Structure and Survival (Edited by B.Battaglia, J. Valencia and D. W. H. Walton), pp. 375–380. CambridgeUniversity Press, Cambridge, UK.
57. Villafane, V. E., M. Andrade, V. Lairana, F. Zaratti and E. W. Helbling(1999) Inhibition of phytoplankton photosynthesis by solar ultravioletradiation: Studies in Lake Titicaca, Bolivia. Freshw. Biol. 42,215–224.
58. Frederick, J. E., H. F. Snell and E. K. Haywood (1989) Solar ultra-violet radiation at the earth’s surface. Photochem. Photobiol 50,443–450.
59. Egeland, E. S., R. R. L. Guillard and S. Liaaen-Jensen (1997)Additional carotenoid prototype representatives and a general chemo-systematic evaluation of carotenoids in prasinophyceae (chlorophyta).Phytochemistry 44(6), 1087–1097.
60. Helbling, E. W., E. S. Barbieri, M. A. Marcoval, R. J. Goncalves andV. E. Villafane (2005) Impact of solar ultraviolet radiation on marinephytoplankton of Patagonia, Argentina. Photochem. Photobiol. 81,807–818.
61. Boelen, P., I. Obernosterer, A. A. Vink and A. G. J. Buma (1999)Attenuation of biologically effective UV radiation in tropical Atlanticwaters measured with a biochemical DNA dosimeter. Photochem.Photobiol. 69, 34–40.
62. Helbling, E. W., V. E. Villafane, A. G. J. Buma, M. Andrade and F.Zaratti (2001) DNA damage and photosynthetic inhibition induced bysolar UVR in tropical phytoplankton (Lake Titicaca, Bolivia). Eur. J.Phycol. 36, 157–166.
934 Marcelo Hernando et al.

63. Garcia-Pichel, F. (1994) A model for shelf-shading in planktonicorganisms and its implications for the usefulness of ultraviolet sun-screens. Limnol. Oceanogr. 39, 1704–1717.
64. Van Leeuwe, M. A., B. van Sikkelerus, W. W. C. Gieskes and J.Stefels (2005) Taxon-specific differences in photoacclimation tofluctuating irradiance in an Antarctic diatom and a green flagellate.Mar. Ecol. Prog. Ser. 288, 9–19.
65. Harding, L. W., T. R. Fisher Jr. and M. A. Tyler (1987) Adaptiveresponses of photosynthesis in phytoplankton: Specificity to time-scaleof change in light. Biol. Oceanogr. 4, 403–437.
66. Prezelin, B. B. (1992) Diel periodicity in phytoplankton productivity.Hydrobiologia 238, 1–35.
67. Demmig-Adams, B. and W. W. Adams (1996) Chlorophyll andcarotenoid composition in leaves of Euonymus kiautschovicusacclimated to different degrees of light stress in the field. Aust. J.Plant Physiol. 23, 649–659.
68. Melis, A., J. A. Nemson and M. A. Harrison (1992) Damage tofunctional components and partial degradation of photosystem IIreaction center proteins upon chloroplast exposure to ultraviolet-Bradiation. Biochim. Biophys. Acta 1109, 312–320.
69. Adir, N., Z. Hagit, S. Shochat and I. Ohad (2003) Photoinhibition—Ahistorical perspective. Photosynth. Res. 76, 343–370.
70. Vass, I., S. Styring, T. Hundal, A. Koivuniemi, E.-M. Aro and B.Andersson (1992) Reversible and irreversible intermediates duringphotoinhibition of photosystem II: Stable reduced QA species promotechlorophyll triplet formation. Proc. Natl. Acad. Sci. U. S. A. 89,1408–1412.
71. Bumann, D. and D. Oesterhelt (1995) Destruction of a single
chlorophyll is correlated with the photoinhibition of photosystem II
with a transiently inactive donor side. Proc. Natl. Acad. Sci. U. S. A.
92, 12195–12199.72. Andersson, J. M., Y. Park and W. S. Chow (1997) Photoinactivation
and photoprotection of photosystem II in nature. Physiol. Plant 100,
214–233.73. Briantais, J. M., C. Vernotte and B. Maison (1982) Influence of
stacking on the distribution of light energy in the photosynthetic
apparatus. Physiol. Veg. 20, 111–122.74. Anderson, J. M., W. S. Chow and Y. I. Park (1995) The grand design
of photosynthesis: Acclimation of the photosynthetic apparatus to
environmental cues. Photosynth. Res. 46, 129–139.75. Neori, A., M. Vernet, O. Holm-Hansen and F. T. Haxo (1988)
Comparison of chlorophyll far-red and red fluorescence excitation
spectra with photosynthetic oxygen action spectra for photosystem II in
algae. Mar. Ecol. Prog. Ser. 44, 297–302.76. Hager, A. and H. Stransky (1970) Das Carotinoidmuster und die
Verbreitung des lichtinduzierten Xanthophyllcyclus in verschiedenen
Algenklassen. V. Einzelne Vertreter der Cryptophyceae, Euglenophy-
ceae, Bacillariophyceae, Chrysophyceae und Phaeophyceae. Arch.
Mikrobiol. 73, 77–89.77. Davidson, A. T., H. J. Marchant and W. K. de la Mare (1996) Natural
UVB exposure changes the species composition of Antarctic
phytoplankton in mixed cultures. Aquat. Microb. Ecol. 10, 299–305.
Photochemistry and Photobiology, 2006, 82 935




















