
Nickel localization in tissues of different hyperaccumulator species of Euphorbiaceae from...
8
Plant Soil (2007) 293:99–106 DOI 10.1007/s11104-007-9227-3 123 ORIGINAL PAPER Nickel localization in tissues of diVerent hyperaccumulator species of Euphorbiaceae from ultramaWc areas of Cuba Rosalina Berazaín · Vicenta de la Fuente · Lourdes Rufo · Nuria Rodríguez · Ricardo Amils · Blanca Díez-Garretas · Daniel Sánchez-Mata · Alfredo Asensi Received: 17 October 2006 / Accepted: 16 February 2007 / Published online: 14 March 2007 © Springer Science+Business Media B.V. 2007 Abstract Pantropical species of the genera Phyl- lanthus and Euphorbia and the Cuban endemic genus Leucocroton from the Euphorbiaceae fam- ily, were selected for nickel localization micro- analysis. Scanning Electron Microscopy coupled with Energy Dispersive X-ray Microanalysis (SEM-EDX) was used for qualitative detection of nickel in the selected Ni-hyperaccumulator species: Euphorbia helenae, Leucocroton linea- rifolius, L. Xavicans Phyllanthus orbicularis, P. discolor and P. xpallidus, all collected from Cuban ultramaWc soils. The leaves and stems from the Euphorbiaceae species analyzed were the organs with higher nickel accumulation. Elemen- tal mapping of leaves and stem tissues from these species have been compared. The highest Ni con- centrations were found in the laticifer tubes of stems and the epidermis tissues of leaves in all the analyzed species, suggesting a general pattern of the Euphorbiaceae family for nickel accumula- tion. The high nickel concentrations and its rather homogeneous distribution found in leaves of these Ni-hyperaccumulating plants suggest a pos- sible role in protection mechanisms against envi- ronmental stress, such as UV irradiation. Keywords Elemental mapping · Euphorbiaceae · Nickel · Nickel hyperaccumulators · SEM-EDX Introduction UltramaWc soils originate from the weathering of ultramaWc rocks and contain elevated concentra- tions of siderophilic elements including Mg, Fe, Mn, Ni, Cr, Co, V, Ti and some times Cu, some of them considered essential for plant growth at trace concentrations, but which are toxic at high con- centrations due to their interference with diVerent R. Berazaín Jardín Botánico Nacional, Universidad de La Habana, 19230 Ciudad Habana, Cuba V. de la Fuente (&) · L. Rufo Departamento de Biología, Facultad de Ciencias, Universidad Autónoma de Madrid, Cantoblanco, 28049 Madrid, Spain e-mail: [email protected] N. Rodríguez · R. Amils Centro de Astrobiología (INTA-CSIC), 28850 Torrejón de Ardoz, Madrid, Spain R. Amils Centro de Biología Molecular, Universidad Autónoma de Madrid, Cantoblanco, 28049 Madrid, Spain B. Díez-Garretas · A. Asensi Departamento de Biología Vegetal, Facultad de Ciencias, Universidad de Málaga, 29071 Málaga, Spain D. Sánchez-Mata Departamento de Biología Vegetal II, Facultad de Farmacia, Universidad Complutense de Madrid, 28040 Madrid, Spain
Transcript of Nickel localization in tissues of different hyperaccumulator species of Euphorbiaceae from...

Plant Soil (2007) 293:99–106
DOI 10.1007/s11104-007-9227-3ORIGINAL PAPER
Nickel localization in tissues of diVerent hyperaccumulator species of Euphorbiaceae from ultramaWc areas of Cuba
Rosalina Berazaín · Vicenta de la Fuente · Lourdes Rufo · Nuria Rodríguez · Ricardo Amils · Blanca Díez-Garretas · Daniel Sánchez-Mata · Alfredo Asensi
Received: 17 October 2006 / Accepted: 16 February 2007 / Published online: 14 March 2007© Springer Science+Business Media B.V. 2007
Abstract Pantropical species of the genera Phyl-lanthus and Euphorbia and the Cuban endemicgenus Leucocroton from the Euphorbiaceae fam-ily, were selected for nickel localization micro-analysis. Scanning Electron Microscopy coupledwith Energy Dispersive X-ray Microanalysis(SEM-EDX) was used for qualitative detectionof nickel in the selected Ni-hyperaccumulator
species: Euphorbia helenae, Leucocroton linea-rifolius, L. Xavicans Phyllanthus orbicularis, P.discolor and P. xpallidus, all collected fromCuban ultramaWc soils. The leaves and stems fromthe Euphorbiaceae species analyzed were theorgans with higher nickel accumulation. Elemen-tal mapping of leaves and stem tissues from thesespecies have been compared. The highest Ni con-centrations were found in the laticifer tubes ofstems and the epidermis tissues of leaves in all theanalyzed species, suggesting a general pattern ofthe Euphorbiaceae family for nickel accumula-tion. The high nickel concentrations and its ratherhomogeneous distribution found in leaves ofthese Ni-hyperaccumulating plants suggest a pos-sible role in protection mechanisms against envi-ronmental stress, such as UV irradiation.
Keywords Elemental mapping · Euphorbiaceae · Nickel · Nickel hyperaccumulators · SEM-EDX
Introduction
UltramaWc soils originate from the weathering ofultramaWc rocks and contain elevated concentra-tions of siderophilic elements including Mg, Fe,Mn, Ni, Cr, Co, V, Ti and some times Cu, some ofthem considered essential for plant growth at traceconcentrations, but which are toxic at high con-centrations due to their interference with diVerent
R. BerazaínJardín Botánico Nacional, Universidad de La Habana, 19230 Ciudad Habana, Cuba
V. de la Fuente (&) · L. RufoDepartamento de Biología, Facultad de Ciencias, Universidad Autónoma de Madrid, Cantoblanco, 28049 Madrid, Spaine-mail: [email protected]
N. Rodríguez · R. AmilsCentro de Astrobiología (INTA-CSIC), 28850 Torrejón de Ardoz, Madrid, Spain
R. AmilsCentro de Biología Molecular, Universidad Autónoma de Madrid, Cantoblanco, 28049 Madrid, Spain
B. Díez-Garretas · A. AsensiDepartamento de Biología Vegetal, Facultad de Ciencias, Universidad de Málaga, 29071 Málaga, Spain
D. Sánchez-MataDepartamento de Biología Vegetal II, Facultad de Farmacia, Universidad Complutense de Madrid, 28040 Madrid, Spain
123

100 Plant Soil (2007) 293:99–106
cellular functions (Reeves et al. 1999). Soils withhigh concentrations of these elements are usuallyhostile to plants, although they support highlyspecialized Xora, rich in endemics adapted to theseextreme conditions (Brooks 1998). In some cases,plant adaptation to soils with high content ofmetals is based on mechanisms of exclusion (Ernst1976). However, some plants actively absorb theseelements, including heavy metals, at very highconcentrations (hyperaccumulation), often exceed-ing those found in the soils (Brooks 1998). Theprocess of hyperaccumulation raises intriguingbiological questions such as the nature of themechanisms of accumulation (Callahan et al.2006), ways of avoiding toxicity (Kramer et al.1996; Kramer 2000; Seregin and Kozhevnikova2006), and the ecological signiWcance of such highlevels of some elements, mainly heavy metals,inside the plants (Davis and Boyd 2000). Biotech-nological applications of metal accumulatorplants, such as phytoremediation of contaminatedsoils have been successfully developed in therecent years (Brooks 1998; Brooks et al. 2001).
Many species strictly related to ultramaWc soilsbelong to the Euphorbiaceae family, such as vari-ous taxa of the genera Bonania, Euphorbia, Gym-nanthes, Leucocroton, Phyllanthus, Sapium, Savia,etc., and some are Ni hyperaccumulators, reachingconcentrations of over 1% of dry matter of this ele-ment. Reeves et al. (1996) published a long surveyon diverse Euphorbia, Leucocroton and Phyllan-thus taxa from Cuba, Puerto Rico and the Domini-can Republic. Recently it has been estimated that14% of the total Cuban Xora (920 out of 6375) areserpentine endemic (Reeves et al. 1999).
Euphorbia L. is a pantropical and holarticgenus with 13 species described from Cuba. Thesespecies are herbs and shrubs which can accumu-late high amounts of Ni in their laticifer tubes(over 3% in Euphorbia helenae s. l.).
Leucocroton Griseb. is a Caribbean genus.Most of the species are shrubs that are widely dis-tributed in Cuba, including 28 species, and withonly one species identiWed in Hispanola (Haiti).Most of the Cuban species of this genus are Ni-hyperaccumulators species which grow exclusivelyon ultramaWc (serpentine) or related substrates.L. Xavicans and L. linearifolius are the best char-acterized Cuban nickel hyperaccumulator species.
Phyllanthus L. is a pantropical and subtropicalgenus of herbs and shrub species. Over 600 spe-cies have been described, of which 53 taxons havebeen identiWed in Cuba. Approximately halfdevelop on ultramaWcs, the rest are not exclusiveto these substrates and can grow on other types ofsoils. Of the 45 taxons analyzed, 24 are Ni hyper-accumulators: 23 from Cuba and one from His-panola (Dominican Republic).
Two Phyllanthus species, P. orbicularis, P. dis-color and their natural hybrid: P. xpallidus havebeen extensively studied due to their Ni-hyperac-cumulation properties (Berazaín et al. 2007). Thenickel localization observed for these Phyllanthusspecies opened the question of whether it was ageneral pattern of the Euphorbiaceae family or adistribution peculiar to the members of thisgenus.
Table 1 shows the Ni content in leaves andstems of well characterized Ni-hyperaccumulatorspecies from the Cuban Euphorbiaceae family,corresponding to the genera Phyllanthus, Leuco-croton and Euphorbia. Nickel concentration inleaves exceeds 1% of dry weight for all the spe-cies. Phyllanthus xpallidus with a Ni contentbetween 2–6% dry weight in leaves shows themaximum concentration value found for thismetal in members of the Euphorbiaceae family(Berazaín et al. 2007). In the case of Euphorbiahelenae s. l. values of 3% have been measured inthe stem laticifer tubes (Reeves et al. 1996).
The aim of this work was to establish the pat-tern distribution of nickel in organs and tissues ofhyperaccumulator plants from the genera Phyl-lanthus, Leucocroton and Euphorbia grown intheir natural habitat, ultramaWc soils from Cuba.
Materials and methods
Plant samples
Plant samples of several Cuban Ni–hyperaccumu-lator species from Euphorbiaceae family werecollected from their natural ultramaWc environ-ments: Phyllanthus orbicularis Kunth (Cajálbana),Phyllanthus discolor Poepp. ex Spreng (Cajál-bana), Phyllanthus xpallidus Müll. Arg. (Cajál-bana), Euphorbia helenae Urb. s. l. (Holguín),
123

Plant Soil (2007) 293:99–106 101
Leucocronton Xavicans Müll. Arg., (Jardín Botá-nico de la Habana) and Leucocroton lineralifoliusBritt. (La Melba). Herbarium voucher specimensfrom the HAJB Herbarium (National BotanicGarden, La Habana University) were used forthis work. Three specimens of each of the selectedspecies were collected for further analyses. Theparts selected for the study were stems and leavesof the year. Leaves were collected near the bud ofstems. The dimensions of the collected sampleswere the following: Phyllanthus orbicularis,leaves of 0.5 £ 0.6 cm and stems of 1mm of diam-eter; Phyllanthus discolor, leaves of 1.9 £ 2.6 cmand stems of 1 mm of diameter; Phyllanthus xpal-lidus, leaves of 1.8 £ 2.2 cm and stems of 1mm ofdiameter; Euphorbia helenae, leaves of1.7 £ 5.2 cm and stems of 3 mm of diameter; Leu-cocronton Xavicans, leaves of 1.4 £ 7.3 cm and1 mm of diameter stems, and Leucocroton linera-lifolius, leaves of 0.4 £ 2.5 cm and stems of 3 mmof diameter.
A preliminary study was carried out with anickel accumulation test using Wlter paperimpregnated with dimethylglyoxime (1% solutionin ethanol) on dry fragments of plant tissues(Reeves et al. 1999). Of all tested specimens onlyleaves and stems gave strong positive results (redspots), thus stems and leaves were analyzed bySEM–EDX.
Sample preparation for scanning electron microscopy
To obtain micro-morphological scanning elec-tron microscope (m-mSEM) images of leaf and
stem cross sections, a conventional treatment wasgiven to the samples to enhance the visualizationof the diVerent plant tissues (Berazaín et al.2007). Dry leaf and stem samples were Wxed withformyl acetic alcohol (FAA). After washing witha 0.1 M phosphate buVer (pH 7.4) they weredehydrated and dried through a graded ethanolseries. Samples were cut with a sharp blade andmounted onto stubs.
For the qualitative SEM-EDX analysis themethodology described in Berazaín et al. (2007)was used. The leaf tissues analyzed were: cuticle,epidermis, mesophyll, vascular bundle and tric-homes (when present). The stem tissues analyzedwere: epidermis, cortex (including laticifers), cen-tral cylinder and pith. Dry herbarium sampleswere immersed in liquid nitrogen to preservestructural features, and to facilitate the genera-tion of cross sections of leaf and stem with a sharpblade. For each type of tissue four to Wve sampleswere analyzed from each species. In order to ana-lyze leaf surfaces, they were mounted Xat on thesurfaces of graphite stubs. Samples were mountedonto conductive graphite stubs and sputters andgold-coated in a BIO-RAD SC 502 apparatus forelectrical conductivity and to prevent chargingunder the electron beam. Samples were examinedwith a Hitachi S-3000N (Japan) scanning electronmicroscope (SEM) using an acceleration voltageof 20 kV and a working distance for analysis of15 mm. Analysis was performed at room tempera-ture. The qualitative element composition of sam-ples was determined by energy dispersive X-Raymicroanalysis using an INCAx-sight with a Si-LiDetector (Oxford, England), with a detection
Table 1 Reported nickel concentration (mg kg¡1 D.W.) of selected Ni-hy-peraccumulator species of the Euphorbiaceae family
Species Analyzed organs Ni concentration Reference
Leucocroton Xavicans Leaves 6710–15500 Reeves et al. (1996)Leucocroton linearifolius Leaves 13310–27240 Reeves et al. (1996)Euphorbia helenae s. l. Leaves 2000–9000 Reeves et al. (1996)
Stem (latex) 30900 Reeves et al. (1996)Phyllanthus orbicularis Leaves 4140–10950 Reeves et al. (1996)
Leaves 6380 Berazaín et al. (2007)Stem 3660 Berazaín et al. (2007)
Phyllanthus discolor Leaves 13670–31490 Reeves et al. (1996)Leaves 28110 Berazaín et al. (2007)Stem 14940 Berazaín et al. (2007)
Phyllanthus xpallidus Leaves 15390–60170 Reeves et al. (1996)Leaves 28480 Berazaín et al. (2007)Stem 17150 Berazaín et al. (2007)
123

102 Plant Soil (2007) 293:99–106
limit of 10% of the main element. This instrumentis able to detect the lighter elements, C, O and N,and the numerical data generated are referencedas default to the higher peak obtained in eachspectrum. Spectra were analyzed using INCAx-sight software (Oxford, England). Qualitative datawere extracted directly from spectra. Means werecalculated for at least Wve data. Obtained valueswere coded according to the following scale: (–)absence of nickel, (+) nickel concentrationbetween 0–1%, (++) nickel concentration between1–3%, and (+++) nickel concentration over 3%.
Mapping for the diVerent elements was dis-played using the Microanalysis Suite, INCA Suiteversion 3.04 (Oxford England) software. The gridfor microanalysis was set at ultra–Wne resolution.
Results
Leaves
To enhance the view of the diVerent tissues tofacilitate interpretation of the elemental analysis,micro-morphological scanning electron micros-copy was performed (m-mSEM) (Fig. 1). The leaftissues chosen for the analysis were: cuticle,epidermis, mesophyll (palisade and spongy paren-chyma) and vascular bundle. Figure 1A corre-sponds to a Euphorbia helenae s. l. leaf preparationshowing the characteristic morphology of leavesfrom the analyzed Euphorbiaceae species, thickepidermis with waxy coverts. Figure 1B shows atransverse section of Leucocroton linearifolius
leaf with trichomes in the abaxial face. Trichomesare only observed in Leucocroton species. Thesestructural features are absent in Euphorbia andPhyllanthus species. Compared to other hairyplants those from the Euphorbiaceae family areof organic nature.
The micro-morphological SEM-EDX analysisof leaves from the selected species of the Euphor-biaceae family showed that the highest Ni concen-tration was found in the epidermal tissues of allthe analyzed samples. Mesophyll and vascularbundle had a similar but always lower Ni concen-tration than the epidermis (Table 2).
The high resolution EDX-dot map analysis of atransverse section of Leucocroton linearifoliusconWrmed the presence of nickel in the interior ofthe epidermal cells in a clear, sharp, compactform (Fig. 2A and 2B). A lower, although homo-geneous, concentration of nickel was observed inthe transition zone between the palisade paren-chyma and mesophyll cells (Fig. 2B).
Stem
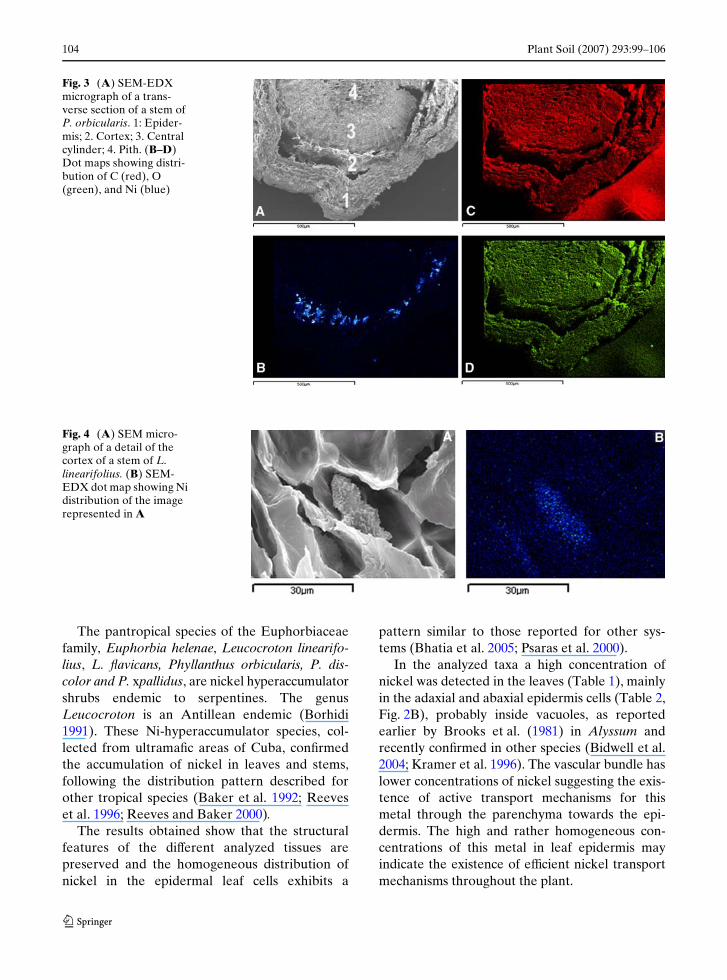
Epidermis, cortex, laticifers, central cylinder andpith were the stem tissues analyzed by SEM–EDX. Latex contained inside the laticifer tubesshowed the highest Ni concentration (Table 2).The high resolution EDX-dot map analysis of atransverse section of P. orbicularis stem con-Wrmed the presence of high nickel concentrationsin these structures (Fig. 3A and 3B). It is proba-ble that the articulated laticifer tubes, typical ofthe Euphorbiaceae, are responsible for the
Fig. 1 (A) m-mSEM micrograph of a transverse section ofa leaf–blade of E. helenae s. l.. 1: Adaxial epidermis; 2:Vascular bundle. 3: Abaxial epidermis. (B) m-mSEMmicrograph of a transverse section of a leaf–blade of
L. linearifolius. 1: Adaxial epidermis; 2: Mesophyll (2apalisade parenchyma and 2b spongy parenchyma); 3:Vascular bundle; 4: Base of trichomes
123

Plant Soil (2007) 293:99–106 103
occasional heterogeneous distribution of nickel tothe diVerent tissues, because this element is foundexclusively in the laticifer tubes originated fromthe laticifer cells (Metcalfe 1983).
The Ni distribution throughout the rest of thestem tissues was relatively homogeneous,although the cortex seemed to have a higher con-centration. Figure 4 shows a detail of the cortex ofL. linearifolius in which cells from the paren-chyma show an accumulation of material giving astrong speciWc signal for nickel.
Discussion
The capacity of some plants to bind metals is aform of tolerating and storing heavy metals whosephysiological function is not well understood. Theprocess of hyperaccumulation seems to dependon the characteristic patterns of metal transloca-tion and distribution, while their concomitant tol-erance for excess metal relies on diVerentdetoxiWcation mechanisms (Seregin and Kozhev-nikova 2006).
Table 2 Qualitative nickel concentration of the selectedspecies of Euphorbiaceae family detected by SEM-EDX.Mesophyll includes palisade and spongy parenchyma.
Cortex includes laticifers. Code: –, absence of nickel; +,nickel concentration between 0–1%; ++, nickel concentra-tion between 1–3%; +++, nickel concentration over 3%
Analyzed tissues E. helenae P. orbicularis P. discolor P. xpallidus L. linearifolius L. Xavicans
LeavesCuticle:Adaxial + ++ ++ ++ ++ +Abaxial + ++ ++ ++ + +Epidermis:Adaxial ++ ++ ++ +++ +++ +++Abaxial + ++ ++ +++ +++ ++Mesophyll + + + ++ ++ +Vascular bundle + + ++ ++ + +Trichomes (base) ++ ++StemsEpidermis ++ + + ++ ++ +Cortex +++ +++ +++ +++ +++ +++Central cylinder ++ + +++ + ++ +Pith + + ++ ++ ++ +
Fig. 2 (A) SEM-EDX micrograph of a trans-verse section of a leaf–blade of L. linearifolius. (B–D) Dot maps showing distribution of C (red), O (green) and Ni (blue)
123
104 Plant Soil (2007) 293:99–106
The pantropical species of the Euphorbiaceaefamily, Euphorbia helenae, Leucocroton linearifo-lius, L. Xavicans, Phyllanthus orbicularis, P. dis-color and P. xpallidus, are nickel hyperaccumulatorshrubs endemic to serpentines. The genusLeucocroton is an Antillean endemic (Borhidi1991). These Ni-hyperaccumulator species, col-lected from ultramaWc areas of Cuba, conWrmedthe accumulation of nickel in leaves and stems,following the distribution pattern described forother tropical species (Baker et al. 1992; Reeveset al. 1996; Reeves and Baker 2000).
The results obtained show that the structuralfeatures of the diVerent analyzed tissues arepreserved and the homogeneous distribution ofnickel in the epidermal leaf cells exhibits a
pattern similar to those reported for other sys-tems (Bhatia et al. 2005; Psaras et al. 2000).
In the analyzed taxa a high concentration ofnickel was detected in the leaves (Table 1), mainlyin the adaxial and abaxial epidermis cells (Table 2,Fig. 2B), probably inside vacuoles, as reportedearlier by Brooks et al. (1981) in Alyssum andrecently conWrmed in other species (Bidwell et al.2004; Kramer et al. 1996). The vascular bundle haslower concentrations of nickel suggesting the exis-tence of active transport mechanisms for thismetal through the parenchyma towards the epi-dermis. The high and rather homogeneous con-centrations of this metal in leaf epidermis mayindicate the existence of eYcient nickel transportmechanisms throughout the plant.
Fig. 3 (A) SEM-EDX micrograph of a trans-verse section of a stem of P. orbicularis. 1: Epider-mis; 2. Cortex; 3. Central cylinder; 4. Pith. (B–D) Dot maps showing distri-bution of C (red), O (green), and Ni (blue)
Fig. 4 (A) SEM micro-graph of a detail of the cortex of a stem of L. linearifolius. (B) SEM-EDX dot map showing Ni distribution of the image represented in A
123

Plant Soil (2007) 293:99–106 105
The highest nickel concentrations for the stud-ied species were found in the stem laticifer tubes,similar to those observed for other Euphorbia-ceae species (P. palawanensis (Baker et al. 1992)and P. serpentinus (Kersten et al. 1980)) and spe-cies from other families (Kersten et al. 1980; Sag-ner et al. 1998). Although the highest Niconcentration was found in laticifer tubes, the leafepidermis tissues show the highest Ni mass valuesin comparison to the rest of the plant tissues(Table 2).
An important question regarding nickel hyper-accumulation pertains to the form in which themetal accumulates in the plant. It has beenreported that organic dicarboxilic acids, such ascitrate and malate, can produce nickel complexesinside vacuoles of leaf epidermal cells (Kerstenet al. 1980) or in the laticifer tubes (Sagner et al.1998; Perrier et al. 2004). Chelating eYciencyseems to depend on the plant species. (Sereginand Kozhevnokova 2006). In addition to organicacid complexes, several nickel hyperaccumulatorplant species comprise metal aquacomplexes(Ni(H2O)6
2+) or metal complexes with histidine(Kersten et al. 1980; Kramer et al. 1996; Lee et al1977; Persans et al. 1999). Nickel and other metalshave been detected associated to cell walls aswell, which might be due to the aYnity of ionicmetal for some components of the cell wall suchas polygalacturonic acid or pectins (Merce et al.2001). In our case the mechanism by which nickelaccumulates in the diVerent Euphorbiaceae spe-cies is not yet clear, although the similarity in themetal localization pattern detected suggests com-parable mechanisms.
The hypothesis that nickel hyperaccumulationis a protection strategy against microbial patho-genesis or predation seems reasonable (Boyd andMoar 1999), although the homogeneous distribu-tion of nickel in leaf epidermis of Euphorbiaceaespecies living in extreme xeric habitats points toalternative functional interpretations. We pro-pose, in the case of the reported Euphorbiaceaespecies, that the high concentration of nickelfound in their leaves could, through the UV radi-ation shielding exerted by this metal at speciWcregions of the ultraviolet spectra, prevent the oxi-dative stress that results from exposure to highradiation intensities (Berazaín et al. 2007). This
shielding has been recently described for algaegrowing in high concentration of ferric iron solu-tions (Gómez et al. 2004). Work to evaluate thisadditional function for nickel in hyperaccumula-tor plants species grown in xero-edaphic climaticconditions is currently in progress.
Acknowledgements This work was supported by grantsBOS2002–02148 and CGL2006–02534 and institutionalgrant from the Fundación Areces to the Centro de BiologíaMolecular. L. R. is a fellow from the Comunidad Autóno-ma de Madrid. We thank both the Universidad Autónomade Madrid and the Universidad Complutense for theirWnancial support (International Programs) that enabledVF and DSM to attend all the events of the IVth Interna-tional Conference on Serpentine Ecology held in Havana(Cuba) in April, 2003, which was the seed of this study.
References
Baker AJM, Proctor J, Van Balgooy MMJ, Reeves RD(1992) Hyperaccumulation of nickel by the Xora of theultramaWcs of Palawan, Republic of the Philippines.In: Baker AJM, Proctor J, Reeves RD (eds) The Veg-etation of UltramaWc (Serpentine) Soils. InterceptLtd. Andover, UK, pp 291–304
Berazaín R, Fuente V, Sanchez-Mata D, Rufo L, Rodrí-guez N, Amils R (2007) Nickel localization on tissuesof hyperaccumulator species of Phyllanthus L. (Eu-phorbiaceae) from ultramaWc areas of Cuba. BiolTrace Elem Res 115:67–86
Bhatia NP, Baker AJM, Walsh KB, Midmore DJ (2005) Arole for nickel in osmotic adjustment in drought-stressed plants of the nickel hyperaccumulator Stack-ousia tryonii Bailey. Planta 223:134–139
Bidwell SD, Crawford SA, Woodrow IE, Sommer-Knud-sen J, Marshall AT (2004) Sub-cellular localization ofNi in the hyperaccumulator, Hybanthus Xoribundus(Lindley) F. Muell. Plant Cell and Environ 27:705–716
Borhidi A (1991) Phytogeography and Vegetation Ecologyof Cuba. Akadémiai Kiadó, Budapest, HG
Boyd RS, Moar WJ (1999) The defensive function of Ni inplants: response of the polyphagos herbivore Spodop-tera exigua (Lepidoptera: Noctuidae) to hyperaccumu-lator and accumulator species of Streptanthus(Brassicaceae). Oecologia 118:218–224
Brooks RR (1998) Plants that hyperaccumulate heavy met-als: their role in phytoremediation, microbiology,archaeology, mineral exploration and phytomining.CAB International, Wallingford, Oxon. 380 pp
Brooks RR, Shaw S, Asensi A (1981) The chemical formand physiological function of nickel in some IberianAlyssum species. Physiol Plantarum 51:167–170
Brooks RR, Robinson BH, Howes AW, Chiarucci A(2001) An evaluation of Berkheya coddii Roessler andAlyssum bertolonii Desv for phytoremediation andphytomining of nickel. S Afr J Sci 97:558–560
123

106 Plant Soil (2007) 293:99–106
Callahan DL, Baker AJM, Kolev SD, Wedd AG (2006)Metal ion ligands in hyperaccumulating plants. J BiolInorg Chem 11:2–12
Davis MA, Boyd RS (2000) Dynamics of Ni-based defenceand organic defences in the Ni hyperaccumulator,Streptanthus polygaloides (Brassicaceae). New Phytol146:211–217
Ernst WHO (1976) Physiological and biochemical aspectsof metal tolerance. In: Mansfeld TA (ed) EVects of airpollutants on plants. Cambridge University Press,Cambridge, UK, pp 115–133
Gómez F, Grau A, Vázquez L, Amils R (2004) UV radia-tion eVects over microorganisms and study of protec-tive agents. ESA SP-545:21–25
Kersten WJ, Brooks RR, Reeves RD, JaVré T (1980)Nature of nickel complexes in Psychotria dourrei andother nickel-accumulating plants. Phytochemistry19:1963–1965
Krämer U (2000) Cadmium for all meals – plants with anunusual appetite. New Phytol 45:1–5
Krämer U, Cotterhowells JD, Charnock JM, Baker AJM,Smith JAC (1996) Free histidine as a metal chelator inplants that accumulate nickel. Nature 379:635–638
Lee J, Reeves RD, Brooks RR, JaVre T (1977) Isolationand identiWcation of a citrate-complex of nickel fromnickel-accumulating plants. Phytochemistry 16:1503–1505
Merce ALR, Landaluze JS, Mangrich AS, Szpoganicz B,Sierakowski MR (2001) Complexes of arabinogalac-tan of Pereskia aculeate and Co2+, Cu2+, Mn2+ and Ni2+
. Biores Technol 76:29–37Metcalfe CR (1983) Laticifers and latex . In Anatomy of
the dicotyledons. Metclafe CR, Chalk L (eds) Vol. II.Claredon press, Oxford, pp 70–81
Perrier N, Colin F, JaVré T, Ambrosi JP, Rose J, BotteroJY (2004) Nickel speciation in Sebertia acuminata, aplant growing on a lateritic soil of New Caledonia. CRGeosci 336:567–577
Persans MW, Yan X, Patnoe JM, Kramer V, Salt DE(1999) Molecular dissection of the role of histidine innickel hyperaccumulation in Thlaspi goesingense (Ha-lacsy). Plant Physiol 121:1117–1126
Psaras GK, Constantinidis TH, Cotsopoulos B, Manetas Y(2000) Relative abundance of nickel in the leafepidermis of eight hyperaccumulators: evidence thatthe metal is excluded from both guard cells and tric-homes. Ann Bot (London) 86:73–78
Reeves RD, Baker AJM (2000) Metal-accumulatingplants. In: Raskin I, Ensley BD (eds) Phytoremedia-tion of toxic metals: using plants to clean up the envi-ronment. John Wiley and sons, New York, USA, pp193–229
Reeves RD, Baker AJM, Borhidi A, Berazaín R (1996)Nickel-accumulating plants from the ancient serpen-tine soils of Cuba. New Phytol 133:217–224
Reeves RD, Baker AJM, Borhidi A, Berazaín R (1999)Nickel hyperaccumulation in the serpentine Xora ofCuba. Ann Bot (London) 83:29–38
Sagner S, Kneer R, Wanner G, Cosson JP, Deus-NeumannB, Zenk MH (1998) Hyperaccumulator, complexationand distribution of Ni in Sebertia acuminata. Phyto-chemistry 47(3):339–47
Seregin IV, Kozhevnikova AD (2006) Physiological role ofnickel and its toxic eVects on higher plants. Russ JPlant Physiol 53(2):257–308
123




















