
Phytoextraction of soil cobalt using hyperaccumulator plants
Upload
independentCategory
view
4download
0
DIVISION S-4-SOIL FERTILITY& PLANT NUTRITION
Zinc and Cadmium Uptake by Hyperaccumulator Thlaspi caerulescensGrown in Nutrient Solution
S. L. Brown,* R. L. Chancy, J. S. Angle, and A. J. M. Baker
ABSTRACTPhytoremediation of heavy-metal-contaminated soils can be an inex-
pensive means to remove hazardous metals from soil. Two metallo-phytes, Thlaspi caerulescens (J. & C. Presl, a Zn and Cd hyperaccumu-lator) from Prayon, Belgium, and a Zn-tolerant ecotype of bladdercampion [Silene vulgaris (Moench.) Garcke L.] from Palmerton, PA,were compared with tomato [Lycopersicon lycopersicum (L.) Karsten,metal intolerant] in nutrient solution to characterize Zn and Cd uptakeand tolerance. Zinc and Cd were added to solutions at a 50:1 molarratio to simulate concentrations often found on contaminated sites.Seven treatment concentrations were used, ranging (in half-log incre-ments) from 3.16 \aM Zn + 0.063 uM Cd to 10000 uM Zn + 200uMCd. Thlaspi caerulescens showed much greater tolerance to Zn/Cdtreatments than the other species, with toxicity stress only apparentat the 10000 uM Zn/200 uM Cd treatment. In this treatment, shootconcentrations of Zn and Cd were 33600 and 1140 nig kg~', respec-tively. Thlaspi caerulescens was also more effective at translocatingboth Zn and Cd from solution to shoots. Zinc concentration in shootsof T. caerulescens was higher than the other species at all Zn/Cdtreatments. Cadmium concentration in shoots of T. caerulescens weresignificantly higher than in bladder campion only at the 316 \aMZn/6.32 uM Cd treatment. This genotype of T. caerulescens maynot hyperaccumulate Cd. However, extreme Zn and Cd uptake andtolerance is evident in T. caerulescens, with >25000 mg Zn kg'1 and1000 mg Cd kg'1 before yield is reduced. Results suggest thatT. caerulescens may be a candidate for the phytoremediation of Zn-contaminated soils.
THE AMOUNT OF LAND contaminated with heavy met-als has increased during the last century due to
mining, smelting, and other industrial activities. At pres-ent, remediation technologies consist primarily of re-moval and replacement of contaminated soils (Geiger etal., 1993). An alternative soil remediation technology hasbeen proposed that would use rare, heavy-metal-tolerantplant species that are able to hyperaccumulate metals inplant shoots (Chaney, 1983; Baker and Brooks, 1989;Baker etal., 1991). Possibly, these tolerance mechanismscould be exploited to remove heavy metal pollutantsfrom soil (Chaney, 1983; Baker et al., 1991). Thisproposed technology, called phytoremediation, involvessuccessive croppings of hyperaccumulator plants to trans-locate polluting metals from soil to plant shoots. Shootsof some species accumulate as much as 4 g Ni or ZnS.L. Brown and J.S. Angle, Dep. of Agronomy, Univ. of Maryland,College Park, MD 20742; R.L. Chaney, USDA-ARS, EnvironmentalChemistry Lab., Beltsville, MD 20705; and A.J.M. Baker, Dep. of Animaland Plant Sciences, Sheffield Univ., Sheffield, UK S10 2UQ. Received25 Oct. 1993. *Corresponding author.
Published in Soil Sci. Soc. Am. J. 59:125-133 (1995).
kg ' in dry matter, and the plant ash is similar to low-grade metal ores (Baker and Walker, 1990). Harvestedplant shoot biomass could then be smelted to recycle theaccumulated metals. Before phytoremediation can bedeveloped for commercial uses, the behavior of hyperac-cumulator species must be more clearly understood. Anunderstanding of the patterns of heavy metal uptakeand the specificity of tolerance will enable growers tomaximize metal pollutant concentrations in plant shoots.
Plants able to tolerate high soil concentrations of partic-ular metals were first described several centuries ago(Cannon, 1960; Baker etal., 1988). Metal-tolerant plantsare currently used to revegetate sites denuded due toexcessive soil metal concentrations (e.g., Oyler, 1988).Recent studies investigated the evolution of tolerance(Antonovics et al., 1971; Baker et al., 1990), specificmechanisms of tolerance (Ernst, 1978; Baker, 1987;Baker and Walker, 1990; Cumming and Tomsett, 1992),and the specificity of metal tolerance (Baker, 1987; Cum-ming and Tomsett, 1992).
Hyperaccumulator species are defined as those whoseleaves contain MOO mg Cd kg'1, 1000 mg Ni and Cukg"1, or >10000 mg Zn and Mn kg"1 (dry weight) whengrown in metal-rich soils (Baker and Brooks, 1989;Baker et al., 1994). Species also accumulate metals whengrown in conventional potting media (Reeves andBrooks, 1983). Possibly, hyperaccumulator plants havea higher requirement for metals such as Zn, which areessential micronutrients, and show a positive response toincreased soil or solution concentrations of these elements(Hajar, 1987).
Thlaspi caerulescens and bladder campion are bothmetallophytes. Thlaspi caerulescens can be considered anendemic Zn hyperaccumulator (Baker, 1987). Endemicmetallophytes are defined as ancient colonizing speciesthat are only competitive on contaminated sites. Thlaspicaerulescens grows as a small basal rosette up to 15 cmhigh. Its habitat is generally confined to metal-contaminated sites. Thlaspi caerulescens is related to theperennial weed pennycress (Thlaspi arvense L.). Bladdercampion has ecotypes tolerant of a variety of metals(Baker, 1978; Verkleij et al., 1990; Schat and Kalff,1992). It can grow on unpolluted as well as contaminatedsites. Both species may prove useful in the remediationof contaminated sites, T. caerulescens through phyto-remediation and bladder campion through revegetation.
Previous work with T. caerulescens has concentratedon defining metal concentrations and growth patterns ofplants found on contaminated sites (Rascio, 1977; Reeves
125

126 SOIL SCI. SOC. AM. J., VOL. 59, JANUARY-FEBRUARY 1995
and Brooks, 1983; Reeves et al., 1983). Limits of toler-ance and cotolerance have also been addressed by someresearchers (Reeves and Baker, 1984; Ingrouille andSmirnoff, 1986; Hajar, 1987; Baker et al., 1994). Rascio(1977) measured the Zn content of Thlaspi rotundifoliumssp. cepaeifolium (Wulfen) Rouy & Foucaud roots,shoots, and leaves from a mine site at different intervalsduring a 1-yr period. Leaf concentrations were consis-tently higher than root or stem concentrations.
For phytoremediation to prove effective, it is necessaryto delineate patterns of uptake and limits of toleranceof potential phytoremediation plant species. It is alsoimportant to define accumulation patterns at lower con-centrations of soil metals to determine if phytoremedia-tion efficiency will decline as soils approach natural metalconcentrations. This study was conducted using nutrientsolution to observe Zn and Cd uptake across a widerange of solution metal concentrations. Zinc and Cduptake and distribution within the three plant specieswere compared to develop more specific insight into thefunctioning of a hyperaccumulator species (T. caerules-cens) in comparison with a tolerant species (bladdercampion) and a sensitive species (tomato). Mineral nutri-ent concentrations in T. caerulescens were also measuredto determine the effect of high Zn concentrations on thedistribution and uptake of necessary plant nutrients bya hyperaccumulator.
MATERIALS AND METHODSA nutrient solution study was conducted to define patterns
of Zn and Cd uptake by two Zn- and Cd-tolerant races ofbladder campion and T. caerulescens compared with a nontoler-ant species ('Rutgers' tomato) at varying solution concentrationsof Zn and Cd. Seed for bladder campion was collected fromplants growing on the mountainside < 1.6 km directly down-wind from a Zn smelter in Palmerton, PA. Seed for T. caerules-cens was obtained from plants growing near a Zn/Cd smelterin Prayon, Belgium (Vazquez et al., 1992). The study wasconducted in an environmental growth chamber in which tem-perature was maintained at 25 °C day and 19°C night, andrelative humidity was set at 70%. The day period was 16 h,with >400 umol photon m~2 s~' photosynthetically activeradiation from a combination of cool-white fluorescent andincandescent lamps.
Thlaspi caerulescens was germinated by placing seeds inan aerated dilute 0.5 concentrated Hoagland solution (2.5 mMCaNO3 and KNO3; 10 nMFe as FeEDDHA; 0.1 mMK2HPO4;75 nM KC1; 10 uJtf H3BO3; 1 mM Mg as MgSO4; 4 \iM Mnas MnCl2-4H2O; 0.5 uAf Cu as CuSO4-5H2O; 1 uAf Zn asZnSO4 • 7H2O; and 0.2 \iM Mo as Na2MoO4 • 2H2O). After 2 d,radicals and two primary leaves emerged and seedlings weretransplanted to standard seed germination papers. The paperswere kept moist in a germination solution containing 1 mMMgSO4, 0.1 mM K2HPO4, 2.5 mM Ca(NO3)2, and 2.5 mMKNO3. After 14 d, T. caerulescens was transferred from theseed germination papers into 8-L buckets containing the samenutrient solution described above. Seedlings were maintainedin these buckets for an additional 14 d before treatments wereinitiated.
Bladder campion seed was placed in standard seed germina-tion papers kept moist with the same germination solutiondescribed above. Seeds germinated within 6 d. Bladder campionwas transferred to 8-L buckets containing the nutrient solution
described above 2 wk after seeds germinated and was main-tained in the growth medium for 7 d until metal treatmentswere initiated. Tomato seeds were also germinated in seedgermination papers for 10 d. Seedlings were transplanted into8-L buckets containing the above nutrient solution 4 d beforemetal treatments were begun.
Two T. caerulescens, three bladder campion, and two tomatoplants were placed into each of the 2-L polyethylene beakersused for the experiment. The continuosly aerated beakers werearranged in a randomized complete block design with threereplications. Beakers were filled to 2 L with the previouslydescribed nutrient solution minus Zn. Seven treatments withZn and Cd added at a 50:1 molar ratio were then applied. Zincwas supplied as ZnCl2 and Cd was added as 3CdSO4-8H2O.Treatments were 3.16/0.063, 31.6/0.632, 100/2, 316/6.32,1000/20, 3160/63.2, and 10000/200 uM Zn/Cd. Normal Znconcentrations for nutrient solutions are 1 jxM Zn. In mostuncontaminated soils, water-soluble Zn and Cd concentrationsare <1 |xM. In the soil directly behind the smelter in Palmerton,total metals were 0.738 mol Zn kg'1 and 9.1 mmol Cd kg"'.Water-extractable (5 g soil per 25 mL H2O) concentrations(which may have some relationship to soil solution concentra-tions) were 2.74 mM Zn and 32.9 uM Cd (Brown et al.,1994).
The activities of free Zn and Cd as well as other nutrientsin the nutrient solutions were calculated using GEOCHEM-PC(Parker et al., 1995). Levels of Zn and Cd treatments weretoo high to consider using chelator-buffer techniques to buffermetal ion activities in the nutrient solution. Differences inconcentration and calculated activity are reported in Table 1.Solution pH was maintained near 6.0 by the addition of 2 mMMES (2-morpholinoethanesulfonic acid) buffer and by ad-justing pH as necessary by the addition of KOH. This wasdone to control the precipitation of Zn phosphate across allunits. All plants were equally replicated in all treatments, exceptfor the 10000 uM treatment in which only T. caerulescens wasincluded because earlier studies showed that the other specieswould not survive this treatment. Within each beaker, plantroots were separated daily to prevent interspecies contamina-tion. Each day, 10 uAf K2HPO4, 0.333 uM CaNO3, 0.024mM MgSCX, 106 nM Mn, and 269 nM B were added to eachbucket to maintain nutrient supply. Deionized water was addedto buckets to maintain solution volume. Sixteen days afterinitiation of treatment, all solutions were completely replaced.
Table 1. Total Zn and Cd concentrations and calculated activity(using GEOCHEM-PC) of free Zn2+ and Cd2+ for seven Zn/Cdtreatment additions (Cd/Zn molar ratio 1:50) to 0.5 strengthHoagland solution.
Treatment|iM
Zinc3.1631.61003161000316010000
Cadmium0.0630.6322.006.3220.063.2200
Concentration————————— — log mol/L —
5.504.504.003.503.002.502.00
7.206.205.705.204.704.203.70
Calculatedactivity
5.764.764.263.843.332.822.37
7.476.485.985.495.024.624.34

BROWN ET AL.: ZINC AND CADMIUM UPTAKE BY A HYPERACCUMULATOR 127
HarvestEight days after the application of treatments, tomato plants
in the two highest Zn treatments were harvested. All tomatoplants in these treatments had died. Death, or senescence, wasdenned as the loss of viability of the terminal bud. Bladdercampion in the 3160 \iM Zn/63.2 \\.M Cd treatment was har-vested 13 d into the experiment. Fifteen days after treatmentwas initiated, the remaining tomato plants were harvested.Tomato plants in the 3.16 uM Zn/0.063 \iM Cd treatmentwere starting to shade the other species. All remaining bladdercampion were harvested at 22 d. At this time, the plants inthe 1000 uAf Zn/20 uAf Cd treatment had also senesced.Bladder campion in all remaining treatments had entered thereproductive growth stage. Thlaspi caerulescens was harvested28 d after beginning the treatments.
At harvest, roots and shoots of all species were separated.Roots were rinsed in deionized water. Thlaspi caerulescensshoots were further separated into leaf and stem tissue. Allsamples were dried at 70°C.
Sample AnalysisDry plant samples were weighed and placed in a 480°C
oven for 16 h. Ash was digested with concentrated HNO3 andtaken to dryness, and the residue was dissolved with 3 M HC1.Samples were brought to 25 mL in 1 M HC1. Necessarydilutions were made in 1 M HC1 to maintain constant viscosity.The accuracy of dilutions was checked by diluting knownstandards. Zinc and Cd concentrations of plants were deter-mined using a flame atomic absorption spectrometer. Thlaspicaerulescens samples were also analyzed for Ca, Mg, Mn,Cu, P, Mo, Fe, and K using an inductively coupled plasmaspectrometer. Standards were prepared with Zn and Cd concen-trations approximately equal to plant sample concentrations toassure that these high concentrations would not interfere withreadings of other elements. Cobalt was added to all standardsand samples (40.0 mg Co L~' reference standard) as an internalstandard for inductively coupled plasma analysis.
Statistical AnalysisData was analyzed using SAS-PC (SAS Institute, 1989). Data
required logarithmic transformation to attain homogeneity.Log-transformed treatment means were separated using theWaller-Duncan K-Tatio f-test after it was determined that therewas a significant (P < 0.05) treatment effect using the GLMprocedure. Data presented are arithmetic means of untrans-formed data. All figures show standard errors calculated fromthis data. Root mass from bladder campion in the 3160 \i,MZn/63.2 \iM Cd and tomato in the 1000 \iM Zn/20 \iM Cdtreatments were combined across all replicates due to lowyield. Because of this, standard errors were not calculable.
RESULTS AND DISCUSSIONVisual Symptoms
Two days after the initiation of treatments, both bladdercampion and tomato leaves receiving the 3160 |o,M Zn/63.2 \iM Cd treatment became purple. This may befrom a Zn-induced P deficiency. Tomato leaves in thistreatment also had necrotic spots. Thlaspi caerulescenswas chlorotic in all treatments at the start of treatments.This chlorosis may be related to an abnormally high Znrequirement that was not met by the concentration ofZn in the dilute 0.5 concentrated Hoagland solution in
which the plants were maintained prior to the beginningof the experiment (Hajar, 1987). On the fourth day ofthe trial, tomato plants in the 3160 \iM Zn/63.2 \iM Cdtreatment were dead and those in the 1000 uM Zn/20uM Cd treatments were also showing signs of toxicity.Bladder campion plants in the 1000 nM/20 \aM Cd treat-ment were showing some purpling in older leaves.Thlaspi caerulescens remained chlorotic in all treatments.
Seven days after treatment initiation, tomato remainedvigorous only in the 3.16 \iMZn/0.063 |iMCd treatment.Chlorosis and lateral root development were apparenton all higher treatments, with growth suppression sig-nificant (P = 0.01) in the 31.6 \iM Zn/0.632 \iM Cdtreatment (Fig. 1). All bladder campion seedlings receiv-ing the 3160 u^f Zn/63.2 nMCd treatment died. Bladdercampion supplied with 1000 \iM Zn/20 \iM Cd exhibitedsevere toxicity. At 316 \iM Zn/6.32 uAf Cd, bladdercampion plants showed some purpling of older leavesbut otherwise appeared vigorous. Thlaspi caerulescensremained chlorotic, but showed no growth effects or lossof vigor in all treatments.
At 15 d, tomato was harvested for all remaining treat-ments. At 21 d, T. caerulescens at 3160, 1000, 100,and 31.6 uJlfZn/63.2,20,2, and 0.632 nMCd treatmentswere no longer chlorotic. Thlaspi caerulescens in the10000 M-A/ Zn/200 \iM Cd treatment was severely chlo-rotic, with root and shoot growth inhibited. Thlaspicaerulescens in both the 316 \iM Zn/6.32 \iM Cd and 3.16HMZn/0.0632 n,MCd treatments remained chlorotic and
u.u
en
h-I 4.0OLJ5I—Og 2.0C/l
0.0
1.5
-xCTI—
1.0EJiJ>•>-
D 0.53£
n n
•- - • Bladder Campion»— — A Tomato• —— • T. caerulescens
' • I
Y~~\ ! s^i•,^K\ X;X/ \ -
'"1 X \*""•--.. , v \a ~i'----.. " •
"""*••— — . ,f- - -. *1 0/0.2 1 00/2 1 000/20 1 0 000/200
•- — • Bladder Campion* -—-* Tomato
T • — • T. caerulescens
- i\T \
' "X->i A -r— - ^'\: \'~4. • \b . . . . . . . . . . . . . . . ;'"">v^__.._,...\
10/0.2 100/2 1000/20
Zn/Cd TREATMENT
10000/200
Fig. 1. Dry yield of (a) shoots and (b) roots of Thlaspi caerulescens(hyperaccumulator), bladder campion (indicator), and tomato (sus-ceptible species) grown in nutrient solution with Zn added at a 50:1ratio to Cd across seven Zn/Cd treatments. Bars represent standarderrors of values averaged across replications.
128 SOIL SCI. SOC. AM. J. , VOL. 59, JANUARY-FEBRUARY 1995
plants were smaller than the other treatments. While thischlorosis may be related to a Zn deficiency in the 3.16\nM Zn/0.063 u.M Cd and 31.6 \iM Zn/0.632 \iM Cdtreatments, there is no apparent explanation for the visualsymptoms in the 316 pM Zn/6.32 U.M Cd treatment.Bladder campion receiving the 1000 \iM Zn/20 \iM Cdtreatment died but at all lower Zn/Cd treatments hadentered the reproductive growth phase. The effect ofplant growth cycle on metal uptake is not known. Bladdercampion was harvested at 22 d. At 28 d, all T. caerules-cens were harvested; plants in the highest treatment wereshowing signs of toxicity (chlorosis and significant [P <0.01] yield reduction) although they maintained theirturgor. Plant roots and shoots in the 3.16 \iM Zn/0.063u,M Cd and 316 \aM Zn/6.32 \iM Cd treatments weresmaller across all replicates compared with other treat-ments (P < 0.05).
Concentration of Zinc and Cadmiumin Plant Tissues
Bladder campion had significantly higher root concen-trations of Cd than T. caerulescens and tomato in thelower Zn/Cd treatments (P < 0.05) (Fig. 2). Root Cdconcentrations in tomato were also significantly higherthan T. caerulescens (P < 0.05). At treatments higherthan 316 fj.MZn/20 \iMCd, this relationship for bladder
campion and T. caerulescens changed, with root Cdconcentrations no longer significantly different betweenthe two species. Also for these treatments, bladder cam-pion showed severe phytotoxic leaf symptoms. BecauseT. caerulescens was more tolerant of Zn and Cd, muchhigher root Cd concentrations were reached in T. caer-ulescens than in tomato or bladder campion before yieldwas reduced significantly.
Shoot Cd concentrations were similar for all speciesat the two lowest treatments. At the 2 |o,M Cd and higher,concentrations of Cd in T. caerulescens shoots was sig-nificantly higher than in the other species. The differenceincreased as Cd concentration in solution increased. Con-centrations of Cd in shoot tissue of T. caerulescensreached 1290 mg kg"1 in the 3160 \aM Zn treatmentwith no visible symptoms of Cd or Zn toxicity.
Zinc concentration in plants followed a different pat-tern than Cd concentration (Fig. 3). Thlaspi caerulescenshad significantly lower concentrations of Zn in rootsthan the other species to 316 U.M Zn/6.32 |iM Cd. Abovethat level, root concentrations of Zn were not significantlydifferent for bladder campion, T. caerulescens, and to-mato. Shoot concentrations of Zn were significantlyhigher for T. caerulescens in all treatments (P < 0.001).The shoot Zn concentrations of T. caerulescens reached26000 mg kg"1 in the 3160 \aM Zn treatment withoutany visible signs of Zn toxicity.
tuwu
'en 3000
CJl
E•^ 2000"DCJ-I-'
0 1000QL
0
1500
I 120001V
01
Jl 900
o0 6000
in300
n
K\N Bladder Campion1 1 TomatoKXX T. caerulescens
T
|
a _ ^ n _ it0.063 0.632 2
N\N Bladder Campioni i TomatoKXX T. caerulescens
b ___ nn|
^ -
:
P M vH vH v
5.32 20
iN Nr1
Sn: HI
I
63
I
\1
I
I
2 2
1
J
:
-
-
00
•
120 000 -
0.063 0.632 6.32 20 63.2 200
Cd TREATMENT (fj.M~)Fig. 2. Cadmium concentration in (a) roots and (b) shoots of Thlaspi
caerulescens (hyperaccumulator), bladder campion (indicator), andtomato (susceptible species) grown in nutrient solution with Znadded at a 50:1 ratio to Cd across seven Zn/Cd treatments. Barsrepresent standard errors of values averaged across replications.
c 60 000Nl
Oo30000 -
NXN Bladder CampionI I Tomatoxxx T. caerulescens
a .„ fln. Inl
fl
1
1 I _
•
3.16 31.6 100 316 1000 3160 10000
^ 30 000IDl
cnE
•~" 20 000Crsi
i—oo(/) 10000
N\N Bladder Campioni i TomatoKXX T. caerulescens
:b _ ,_a Nnfi In! In
II
X
3.16 31.6 100 316 1000 3160
Zn TREATMENT (/J.M)10000
Fig. 3. Zinc concentration in (a) roots and (b) shoots of Thlaspicaerulescens (hyperaccumulator), bladder campion (indicator), andtomato (susceptible species) grown in nutrient solution with Znadded at a 50:1 ratio to Cd across seven Zn/Cd treatments. Barsrepresent standard errors of values averaged across replications.

BROWN ET AL.: ZINC AND CADMIUM UPTAKE BY A HYPERACCUMULATOR 129
Zinc and Cadmium TranslocatedTotal Zn and Cd translocated to shoot tissue (concen-
tration of metal in dry matter x dry weight) was calcu-lated. Thlaspi caerulescens translocated significantlymore Zn and Cd to shoots than either bladder campionor tomato, with maximum metals translocated at the3160 u.M Zn/63 \iM Cd treatment (P < 0.001) (Fig. 4).Bladder campion translocated more Cd from solution toshoots than tomato, although translocation of Zn wasnot significantly different.
For T. caerulescens, higher tolerance to both metalswas responsible for higher yields in all but the 3.16 \iMZn treatment for tomato and in all treatments >31.6\iM Zn for bladder campion. Despite generally lowerbiomass, hyperaccumulation of Zn at low solution con-centrations made T. caerulescens more efficient thanbladder campion and tomato at translocating Zn to shoottissue. For phytoremediation purposes, T. caerulescensis much more effective than the other plant species usedin this study.
Shoot/Root Partitioning of Zinc and CadmiumThe lower Zn or Cd tolerance of bladder campion
and tomato compared with T. caerulescens suggest thatinterspecies comparisons of internal distribution of Zn
C- 80
ooX
60
40
20
SS3 Bladder Campion1 1 TomatoKW T. caerulescens
3.16 31.6 100 316 1000 3160
Zn TREATMENT (//M)10000
oOI
M-. -J
4.0
3.5
3.0
2.5
2.0
1.5
1.0
0.5
n n
E5S Bladder CampionI I Tomatoxxx T. caerulescens
.
b _ _ B B j
•
i '0.063 0.632 2 6.32 20 63.2
Cd TREATMENT (//M)200
Fig. 4. (a) Zinc and (b) Cd translocated to harvestable biomass byThlaspi caerulescens (hyperaccumulator), bladder campion (indica-tor), and tomato (susceptible species) grown in nutrient solutionwith Zn added at a 50:1 ratio to Cd across seven Zn/Cd treatments.Bars represent standard errors of values averaged across replica-tions.
and Cd may provide an understanding of different internaltolerance mechanisms (Baker and Walker, 1990.) How-ever, lower tolerance reduced the yield of bladder cam-pion and tomato at higher treatment concentrations, mak-ing comparisons difficult across the range of treatmentsused in this study. Shoot/root ratios of metals are <1when nontolerant or "indicator" species are grown oncontaminated soils (Antonovics et al., 1971; Baker andWalker, 1990; Peterson, 1983).
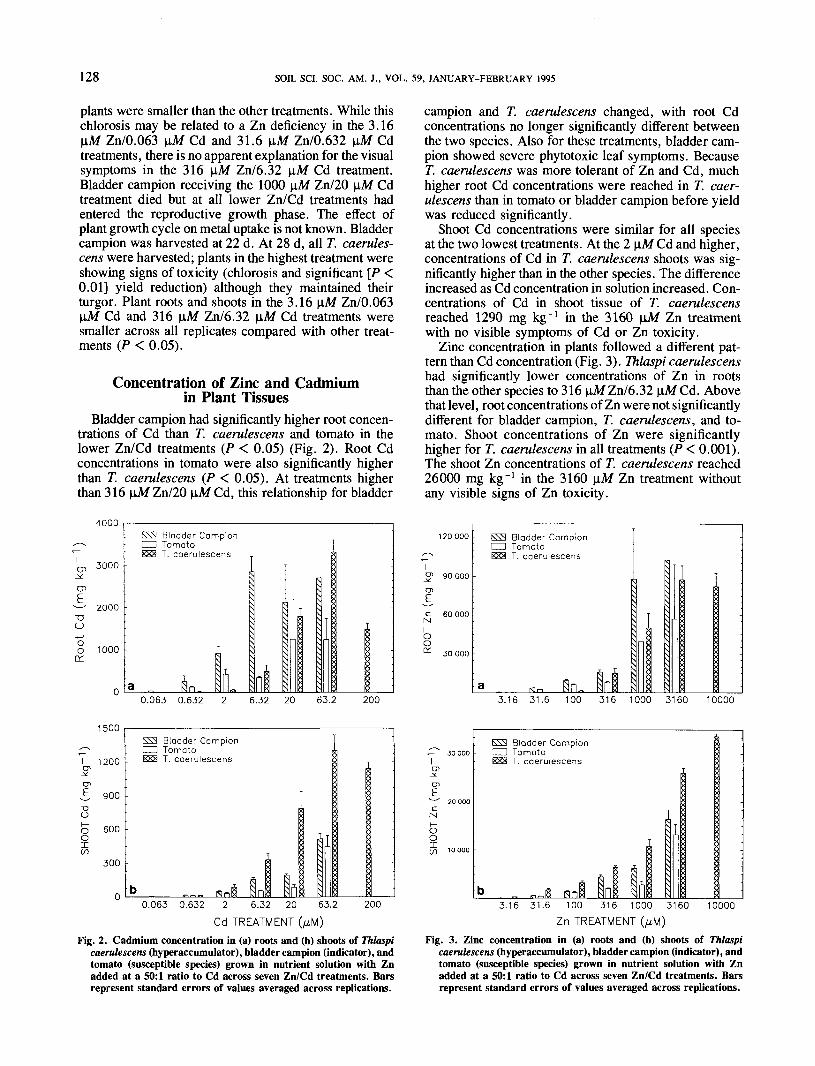
With regard to shoot/root ratios of Zn and Cd, bladdercampion behaved like tomato, with a more severe restric-tion of Zn and Cd translocation to shoots (Fig. 5).Tolerance thresholds (as defined by shoot/root parti-tioning) for both plants were reached at the 316 \iM Zntreatment. Thlaspi caerulescens showed ratio values >1for shoot/root Zn partitioning up to the 316 \iM Zn/6.32\aM Cd treatment. Above 316 \iM Zn, the shoot/rootratio decreased to 0.22 at 1000 uM Zn and graduallyincreased in the higher treatments.
Shoot/root distribution of Cd and Zn followed similarpatterns in T. caerulescens (Fig. 5). Distribution of bothZn and Cd within T. caerulescens was different thandistribution of these metals in bladder campion and to-mato. Thlaspi caerulescens translocated a greater per-centage of Zn and Cd to shoot tissue. A different parti-tioning pattern for both Zn and Cd in T. caerulescenscompared with bladder campion and tomato may suggestthat the internal mechanisms that control the translocation
oorr
oo
0.1
O —— O T. caerulescens• - - • Bladder CampionA- -A Tomato
10 100 1000 10000
1.0
T3oo
Ioo0.1
O —— O T. caerulescens• - -• Bladder CampionA "A Tomato
O
0.1 1.0 10 100
b. CADMIUM TREATMENT (/J.M)1000
Fig. 5. Shoot/root ratios of (a) Zn and (b) Cd concentrations in Thlaspicaerulescens (hyperaccumulator), bladder campion (indicator), andtomato (susceptible species) grown in nutrient solution with Znadded at a 50:1 ratio to Cd across seven Zn/Cd treatments.
130 SOIL SCI. SOC. AM. J., VOL. 59, JANUARY-FEBRUARY 1995
of Cd and Zn in T. caerulescens may be similar (Bakerand Walker, 1990). Hajar (1987) also found shoot/rootratios >1 for both elements through a range of metaladditions in a pot study. Although his highest Zn ratewas greater than the highest Zn treatment used in thisstudy, the available fraction of total metal added wasnot reported.
Average Zn concentration in plant material (sum ofroot and shoot) was higher for T. caerulescens than theother plants in the 3.16 and 31.6 \iM Zn treatments (datanot shown). This implies that, even at lower solutionconcentrations of Zn, Zn uptake by T. caerulescensfollows a different pattern than bladder campion andtomato. This different uptake pattern was not followedby T. caerulescens for Cd uptake. For Cd, average totalconcentration in plants (sum of root and shoot) washigher for bladder campion in the same treatments. Theseresults may suggest that the specialized mechanism forZn uptake in T. caerulescens does not control Cd uptake.Although this study did not examine uptake of Zn orCd by T. caerulescens when supplied singly, differentaccumulation patterns for Zn and Cd from solution toroot cell plasma may suggest that different mechanismsare involved for each metal. A field study (Brown etal., 1994) showed T. caerulescens shoots with >1000mg Zn kg"1 when grown on uncontaminated soils. Zincconcentrations in T. caerulescens were significantlydifferent (P = 0.01) from other plants used in the study.In the same study, Cd uptake by T. caerulescens wasnot significantly different from bladder campion or lettuce(Latuca saliva L.). These results may indicate that com-petition for uptake between Zn and Cd may not be afactor.
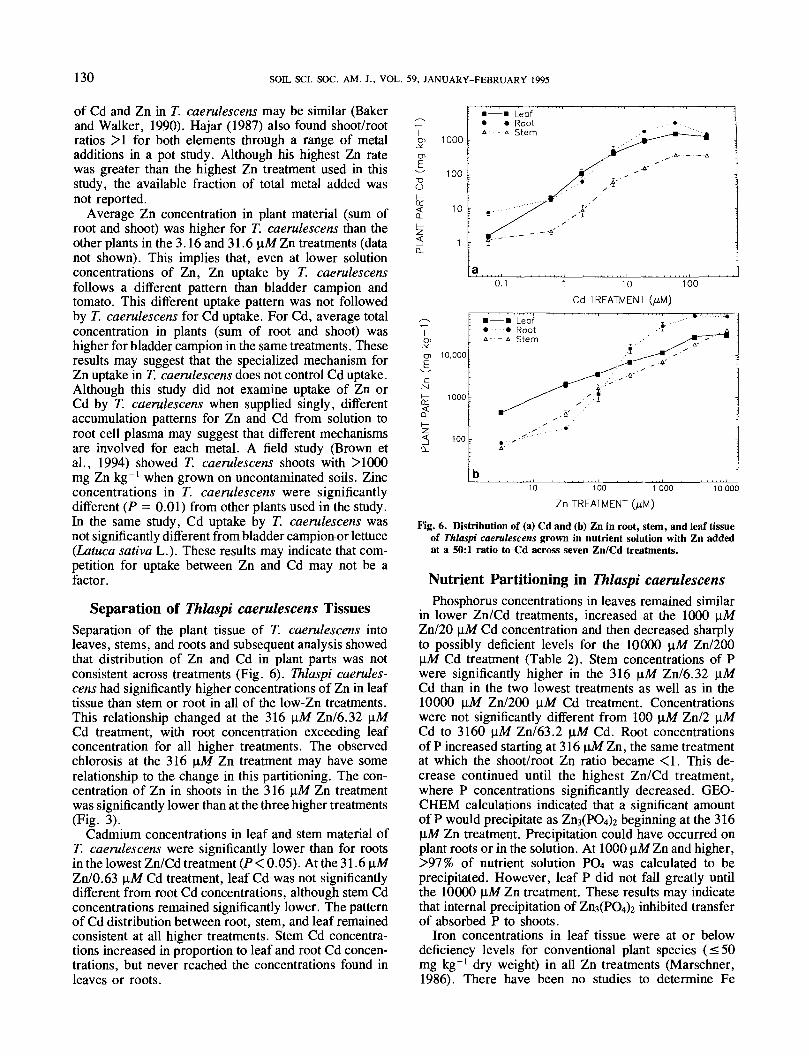
Separation of Thlaspi caerulescens TissuesSeparation of the plant tissue of T. caerulescens intoleaves, stems, and roots and subsequent analysis showedthat distribution of Zn and Cd in plant parts was notconsistent across treatments (Fig. 6). Thlaspi caerules-cens had significantly higher concentrations of Zn in leaftissue than stem or root in all of the low-Zn treatments.This relationship changed at the 316 nM Zn/6.32 \iMCd treatment, with root concentration exceeding leafconcentration for all higher treatments. The observedchlorosis at the 316 uAf Zn treatment may have somerelationship to the change in this partitioning. The con-centration of Zn in shoots in the 316 \iM Zn treatmentwas significantly lower than at the three higher treatments(Fig. 3).
Cadmium concentrations in leaf and stem material ofT. caerulescens were significantly lower than for rootsin the lowest Zn/Cd treatment (P < 0.05). At the 31.6 |o,MZn/0.63 ]iM Cd treatment, leaf Cd was not significantlydifferent from root Cd concentrations, although stem Cdconcentrations remained significantly lower. The patternof Cd distribution between root, stem, and leaf remainedconsistent at all higher treatments. Stem Cd concentra-tions increased in proportion to leaf and root Cd concen-trations, but never reached the concentrations found inleaves or roots.
en 1000
T>O
100
10
LeafRootStem
.A- - - -A
0.1 1 10
Cd TREATMENT100
CT 10,000E
QX
Q_I—2<; 100
10 100
Zn TREATMENT1 000 10000
Fig. 6. Distribution of (a) Cd and (b) Zn in root, stem, and leaf tissueof Thlaspi caerulescens grown in nutrient solution with Zn addedat a 50:1 ratio to Cd across seven Zn/Cd treatments.
Nutrient Partitioning in Thlaspi caerulescensPhosphorus concentrations in leaves remained similar
in lower Zn/Cd treatments, increased at the 1000 nMZn/20 \iM Cd concentration and then decreased sharplyto possibly deficient levels for the 10000 \iM Zn/200M.M Cd treatment (Table 2). Stem concentrations of Pwere significantly higher in the 316 \iM Zn/6.32 \iMCd than in the two lowest treatments as well as in the10000 \iM Zn/200 \iM Cd treatment. Concentrationswere not significantly different from 100 |iM Zn/2 \iMCd to 3160 v.M Zn/63.2 u.M Cd. Root concentrationsof P increased starting at 316 \iM Zn, the same treatmentat which the shoot/root Zn ratio became <1. This de-crease continued until the highest Zn/Cd treatment,where P concentrations significantly decreased. GEO-CHEM calculations indicated that a significant amountof P would precipitate as Zn3(PC>4)2 beginning at the 316|0,M Zn treatment. Precipitation could have occurred onplant roots or in the solution. At 1000 uAf Zn and higher,>97% of nutrient solution PO4 was calculated to beprecipitated. However, leaf P did not fall greatly untilthe 10000 \iM Zn treatment. These results may indicatethat internal precipitation of Zn3(PO4)2 inhibited transferof absorbed P to shoots.
Iron concentrations in leaf tissue were at or belowdeficiency levels for conventional plant species (<50mg kg"1 dry weight) in all Zn treatments (Marschner,1986). There have been no studies to determine Fe
BROWN ET AL.: ZINC AND CADMIUM UPTAKE BY A HYPERACCUMULATOR 131
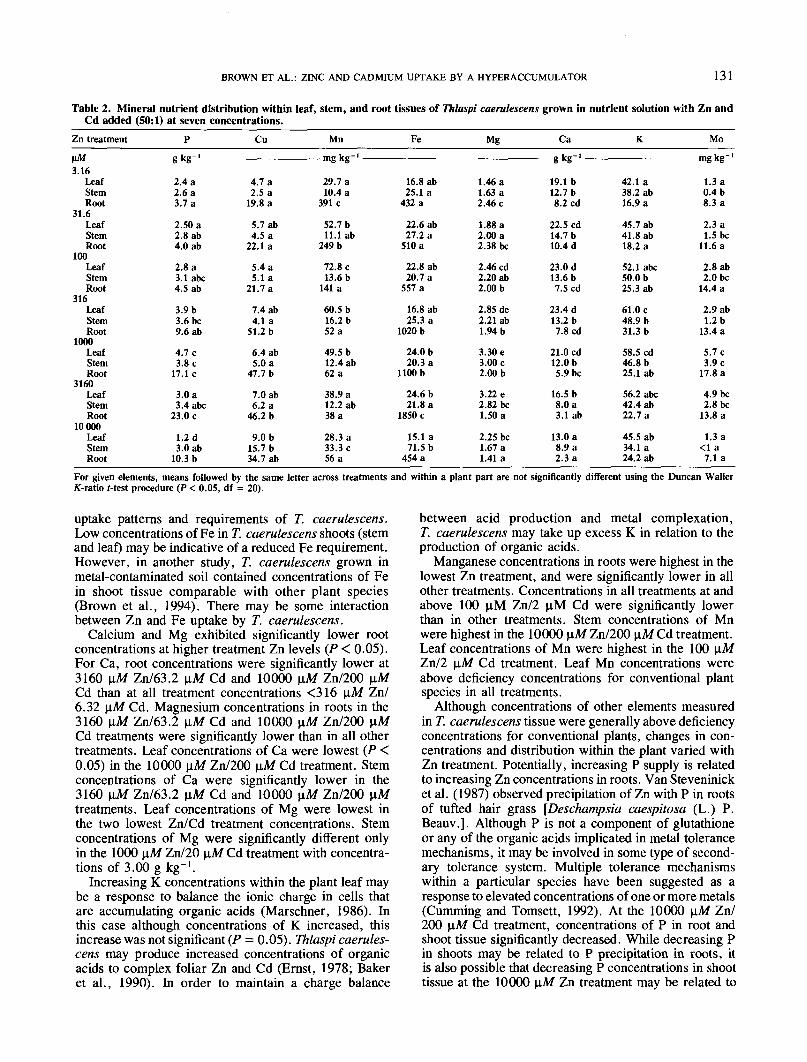
Table 2. Mineral nutrient distribution within leaf, stem, and root tissues of Thlaspi caerulescens grown in nutrient solution with Zn andCd added (50:1) at seven concentrations.
Zn treatment
\iM3.16
LeafStemRoot
31.6LeafStemRoot
100LeafStemRoot
316LeafStemRoot
1000LeafStemRoot
3160LeafStemRoot
10000LeafStemRoot
Pe kff- '5 Ke
2.4 a2.6 a3.7 a
2.50 a2.8 ab4.0 ab
2.8 a3.1 abc4.5 ab
3.9 b3.6 be9.6 ab
4.7 c3.8 c
17.1 c
3.0 a3.4 abc
23.0 c
1.2 d3.0 ab
10.3 b
Cu
4.7 a2.5 a
19.8 a
5.7 ab4.5 a
22.1 a
5.4 a5.1 a
21.7 a
7.4 ab4.1 a
51.2 b
6.4 ab5.0 a
47.7 b
7.0 ab6.2 a
46.2 b
9.0 b15.7 b34.7 ab
Mn
—— mg kg"' ——
29.7 a10.4 a
391 c
52.7 b11.1 ab
249 b
72.8 c13.6 b
141 a
60.5 b16.2 b52 a
49.5 b12.4 ab62 a
38.9 a12.2 ab38 a
28.3 a33.3 c56 a
Fe
16.8 ab25.1 a
432 a
22.6 ab27.2 a
510 a
22.8 ab20.7 a
557 a
16.8 ab25.3 a
1020 b
24.0 b20.3 a
1100 b
24.6 b21.8 a
1850 c
15.1 a71.5 b
454 a
Mg
1.46 a1.63 a2.46 c
1.88 a2.00 a2.38 be
2.46 ed2.20 ab2.00 b
2.85 de2.21 ab1.94 b
3.30 e3.00 c2.00 b
3.22 e2.82 be1.50 a
2.25 be1.67 a1.41 a
Cagkg-'
19.1 b12.7 b8.2 ed
22.5 ed14.7 b10.4 d
23.0 d13.6 b7.5 ed
23.4 d13.2 b7.8 ed
21.0 ed12.0 b5.9 be
16.5 b8.0 a3.1 ab
13.0 a8.9 a2.3 a
K
42.1 a38.2 ab16.9 a
45.7 ab41.8 ab18.2 a
52.1 abc50.0 b25.3 ab
61.0 c48.9 b31.3 b
58.5 ed46.8 b25.1 ab
56.2 abc42.4 ab22.7 a
45.5 ab34.1 a24.2 ab
Mo
mgkg-'
1.3 a0.4 b8.3 a
2.3 a1.5 be
11.6 a
2.8 ab2.0 be
14.4 a
2.9 ab1.2 b
13.4 a
5.7 c3.9 c
17.8 a
4.9 be2.8 be
13.8 a
1.3 a<la7.1 a
For given elements, means followed by the same letter across treatments and within a plant part are not significantly different using the Duncan WallerK-ratio (-test procedure (P < 0.05, df = 20).
uptake patterns and requirements of T. caerulescens.Low concentrations of Fe in T. caerulescens shoots (stemand leaf) may be indicative of a reduced Fe requirement.However, in another study, T. caerulescens grown inmetal-contaminated soil contained concentrations of Fein shoot tissue comparable with other plant species(Brown et al., 1994). There may be some interactionbetween Zn and Fe uptake by T. caerulescens.
Calcium and Mg exhibited significantly lower rootconcentrations at higher treatment Zn levels (P < 0.05).For Ca, root concentrations were significantly lower at3160 \aM Zn/63.2 \iM Cd and 10000 \iM Zn/200 \iMCd than at all treatment concentrations <316 \iM Zn/6.32 \\,M Cd. Magnesium concentrations in roots in the3160 \iM Zn/63.2 \iM Cd and 10000 \nM Zn/200 \iMCd treatments were significantly lower than in all othertreatments. Leaf concentrations of Ca were lowest (P <0.05) in the 10000 \iM Zn/200 \iM Cd treatment. Stemconcentrations of Ca were significantly lower in the3160 nM Zn/63.2 \iM Cd and 10000 u.M Zn/200 U.Mtreatments. Leaf concentrations of Mg were lowest inthe two lowest Zn/Cd treatment concentrations. Stemconcentrations of Mg were significantly different onlyin the 1000 \iM Zn/20 \iM Cd treatment with concentra-tions of 3.00 g kg-'.
Increasing K concentrations within the plant leaf maybe a response to balance the ionic charge in cells thatare accumulating organic acids (Marschner, 1986). Inthis case although concentrations of K increased, thisincrease was not significant (P = 0.05). Thlaspi caerules-cens may produce increased concentrations of organicacids to complex foliar Zn and Cd (Ernst, 1978; Bakeret al., 1990). In order to maintain a charge balance
between acid production and metal complexation,T. caerulescens may take up excess K in relation to theproduction of organic acids.
Manganese concentrations in roots were highest in thelowest Zn treatment, and were significantly lower in allother treatments. Concentrations in all treatments at andabove 100 uM Zn/2 nM Cd were significantly lowerthan in other treatments. Stem concentrations of Mnwere highest in the 10000 \iM Zn/200 \iM Cd treatment.Leaf concentrations of Mn were highest in the 100 \i,MZn/2 |iM Cd treatment. Leaf Mn concentrations wereabove deficiency concentrations for conventional plantspecies in all treatments.
Although concentrations of other elements measuredin T. caerulescens tissue were generally above deficiencyconcentrations for conventional plants, changes in con-centrations and distribution within the plant varied withZn treatment. Potentially, increasing P supply is relatedto increasing Zn concentrations in roots. Van Steveninicket al. (1987) observed precipitation of Zn with P in rootsof tufted hair grass [Deschampsia caespitosa (L.) P.Beauv.]. Although P is not a component of glutathioneor any of the organic acids implicated in metal tolerancemechanisms, it may be involved in some type of second-ary tolerance system. Multiple tolerance mechanismswithin a particular species have been suggested as aresponse to elevated concentrations of one or more metals(Cumming and Tomsett, 1992). At the 10000 \iM Zn/200 iiM Cd treatment, concentrations of P in root andshoot tissue significantly decreased. While decreasing Pin shoots may be related to P precipitation in roots, itis also possible that decreasing P concentrations in shoottissue at the 10000 u.Af Zn treatment may be related to

132 SOIL SCI. SOC. AM. J., VOL. 59, JANUARY-FEBRUARY 1995
decreasing concentrations of Mg, Mo, and Ca and theobserved chlorosis and growth suppression. These find-ings indicate that T. caerulescens's tolerance limits wereexceeded in our study. Possibly, solution concentrationsof Zn were high enough in this treatment to cause someprecipitation of Zn3(PO4)2, and limit plant availabilityof P. The exact cause of toxicity is not known nor couldwe clearly define the elemental interactions involvedsince the precipitation of P could have occurred eitherwithin the symplasm or within the root apoplasm.
CONCLUSIONSThlaspi caerulescens shows much greater accumu-
lation of, and tolerance to, both Zn and Cd than theother plant species studied. Toxicity was evident forT. caerulescens receiving the 10000 ]iM Zn/200 uJtf Cdtreatment, with shoot Zn and Cd concentrations of 32500and 1270 mg kg"1, respectively. In this study, the 50:1Zn/Cd ratio was representative of the ratio of theseelements found on sites contaminated by mining andsmelting activities. Further studies with different Zn/Cdratios are necessary to describe potential uptake patternsof T. caerulescens grown on sites contaminated by otheractivities.
Thlaspi caerulescens was much more efficient thanbladder campion or tomato at translocating both elementsfrom solution to shoots. Except for low biomass, themetal-accumulating properties of T. caerulescens arethose necessary for plants that could be used for phyto-remediation of metal-contaminated soils. Concentrationsof Zn in harvestable shoots are high enough that planttissue could be treated as a low-grade ore (Chaney,1983). Zinc and Cd could be recycled by smelting driedT. caerulescens shoots. Low yield and slow growth rateare the two major factors that limit the potential forthe phytoremediation of Zn/Cd-contaminated soils bysuccessive croppings of T. caerulescens. In addition,T. caerulescens is a low-growing plant, which makesany mechanical harvesting problematic. Based on themore extensive literature on Ni hyperaccumulators (>200species are known), there is no inherent requirement forslow growth in metal hyperaccumulator species (Bakerand Brooks, 1989). Our results suggest T. caerulescensplants or genes from this species could be used to developphytoremediation technology for Zn- and Cd-contam-inated soils. A higher yielding crop with similar Zn- andCd-accumulating properties could be used to remediateZn/Cd-contaminated soils in situ within a shorter timeframe.
ACKNOWLEDGMENTSWe gratefully acknowledge the assistance of C.E. Green
and M.L. Leech in collecting and increasing the bladder cam-pion seed from Palmerton, of C.E. Green for her advice onnutrient solution methodology, and of Ed Henke.

SOLTANPOUR ET AL.: DRIS AND NUTRIENT SUFFICIENCY RANGE FOR CORN 133
Copyright © 2022 FDOKUMEN
















