
Nickel distribution in the hyperaccumulator Alyssum serpyllifolium Desf. spp. from the Iberian...
12
This article was downloaded by:[de la Fuente, V.] [de la Fuente, V.] On: 12 June 2007 Access Details: [subscription number 779380235] Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Plant Biosystems - An International Journal Dealing with all Aspects of Plant Biology Official Journal of the Societa Botanica Italiana Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t713737104 Nickel distribution in the hyperaccumulator Alyssum serpyllifolium Desf. spp. from the Iberian Peninsula To cite this Article: de la Fuente, V., Rodríguez, N., Díez-Garretas, B., Rufo, L., Asensi, A. and Amils, R. , 'Nickel distribution in the hyperaccumulator Alyssum serpyllifolium Desf. spp. from the Iberian Peninsula', Plant Biosystems - An International Journal Dealing with all Aspects of Plant Biology, 141:2, 170 - 180 To link to this article: DOI: 10.1080/11263500701401422 URL: http://dx.doi.org/10.1080/11263500701401422 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article maybe used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material. © Taylor and Francis 2007
Transcript of Nickel distribution in the hyperaccumulator Alyssum serpyllifolium Desf. spp. from the Iberian...

This article was downloaded by:[de la Fuente, V.][de la Fuente, V.]
On: 12 June 2007Access Details: [subscription number 779380235]Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Plant Biosystems - An InternationalJournal Dealing with all Aspects ofPlant BiologyOfficial Journal of the Societa Botanica ItalianaPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713737104
Nickel distribution in the hyperaccumulator Alyssumserpyllifolium Desf. spp. from the Iberian Peninsula
To cite this Article: de la Fuente, V., Rodríguez, N., Díez-Garretas, B., Rufo, L.,Asensi, A. and Amils, R. , 'Nickel distribution in the hyperaccumulator Alyssumserpyllifolium Desf. spp. from the Iberian Peninsula', Plant Biosystems - AnInternational Journal Dealing with all Aspects of Plant Biology, 141:2, 170 - 180To link to this article: DOI: 10.1080/11263500701401422
URL: http://dx.doi.org/10.1080/11263500701401422
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article maybe used for research, teaching and private study purposes. Any substantial or systematic reproduction,re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expresslyforbidden.
The publisher does not give any warranty express or implied or make any representation that the contents will becomplete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should beindependently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings,demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with orarising out of the use of this material.
© Taylor and Francis 2007

Dow
nloa
ded
By:
[de
la F
uent
e, V
.] A
t: 12
:17
12 J
une
2007
Nickel distribution in the hyperaccumulator Alyssum serpyllifolium
Desf. spp. from the Iberian Peninsula
V. DE LA FUENTE1, N. RODRIGUEZ2, B. DIEZ-GARRETAS3, L. RUFO1, A. ASENSI3 &
R. AMILS2,4
1Departamento de Biologıa, Universidad Autonoma, Madrid, Spain, 2Centro de Astrobiologıa (INTA-CSIC), Torrejon de
Ardoz, Madrid, Spain, 3Departmento de Biologıa Vegetal, Universidad de Malaga, Malaga, Spain, and 4Centro de Biologıa
Molecular (UAM-CSIC), Universidad Autonoma de Madrid, Madrid, Spain
AbstractThe relative concentration and distribution of nickel (Ni) in vegetative tissues (leaves, stems and trichomes) andreproductive organs (seeds) was studied using energy-dispersive X-ray microanalysis (EDXS) and scanning electronmicroscopy (SEM) in two previously studied Ni-hyperaccumulator subspecies of Alyssum serpyllifolium Desf. growingnaturally in ultramafic soils of the Iberian Peninsula: A. serpyllifolium ssp. lusitanicum Dudley & P. Silva and A. serpyllifoliumssp. malacitanum Rivas Goday ex G. Lopez. Both taxa showed that Ni accumulates preferentially in the leaves, exhibiting ahomogeneous distribution on both epidermis surfaces. The highest Ni concentrations were found inside the epidermal cellsand at the base of trichome stalks. Ni accumulation in seeds was lower than in the vegetative organs. The location of Ni inthese plants allows us to postulate that its accumulation is a protection mechanism against external stress.
Key words: Alyssum serpyllifolium, EDXS, elemental mapping, hyperaccumulation, Iberian Peninsula, nickel
Introduction
The genus Alyssum L. of the Brassicaceae has a large
number of Ni hyperaccumulator taxa. Jaffre et al.
(1976) coined the term ‘hyperaccumulator’ for such
plants, and Brooks with his collaborators extensively
surveyed metal hyperaccumulating plants, especially
for Ni (Brooks et al., 1977). Ni hyperaccumulator
plants represent more than three-quarters of all metal
hyperaccumulators. Forty-eight species have been
reported in ultramafic soils from southwest Europe
to Asia Minor (Brooks, 1998). Ultramafic soils
contain relatively high amounts of Ni that are
potentially toxic to plants, yet they support a highly
specialized flora rich in endemics and apparently
adapted to this condition (Brooks, 1998). Some
species growing in these soils show high concentra-
tions of Ni in the plant tissues, often exceeding those
in the soils (Brooks, 1998).
Exhaustive analytical studies of the flora of many
parts of the world where ultramafic rocks are
significant (Cuba, Caribbean countries, NW
USA, Newfoundland, Brazil, New Caledonia,
Mediterranean Europe, Turkey, central and eastern
Africa, islands of Indonesia and the Philippines, etc.)
have led to the specific identification of more than
300 Ni hyperaccumulating species to date (Baker
et al., 2000; Reeves & Baker, 2000). These have been
reported and discussed elsewhere (Reeves, 1992,
2003; Medina et al., 1994; Reeves et al., 1996, 1999;
Brooks, 1998). However, the mechanism of Ni
hyperaccumulation in plants is still not well under-
stood, in spite of recent advances in the field. As has
been shown for other elements, Ni and the means of
its hyperaccumulation have been described for
epidermal cells in several species, mainly belonging
to the Brassicaceae (Streptanthus Nutt., Alyssum L.
and Thlaspi L. sp. pl.) (Brooks et al., 1981a,b; Reeves
et al., 1981; Reeves, 1992; Heath et al., 1997;
Kramer et al., 1997; Boyd & Moar, 1999; Psaras
et al., 2000; Davis & Boyd, 2000; Kupper et al.,
2001; Asensi et al., 2004). Recent works describing
Ni localization in Alyssum (A. murale Waldst. & Kit.,
A. corsicum Duby, A. pterocarpum Dudley) evidenced
Correspondence: Vicenta de la Fuente, Departamento de Biologıa, Facultad de Ciencias, Universidad Autonoma de Madrid, Cantoblanco, E-28049 Madrid,
Spain. Tel.: þ34 91 4978100. Fax: þ34 91 4978300. E-mail: [email protected]
Plant Biosystems, Vol. 141, No. 2, July 2007, pp. 170 – 180
ISSN 1126-3504 print/ISSN 1724-5575 online ª 2007 Societa Botanica Italiana
DOI: 10.1080/11263500701401422

Dow
nloa
ded
By:
[de
la F
uent
e, V
.] A
t: 12
:17
12 J
une
2007
the existence of high amounts of Ni in epidermal
tissues and emphasized the presence of this element,
together with Mn, in leaf trichomes (Broadhurst
et al., 2004a,b; Mcnear et al., 2005). The study of Ni
metal localization in vegetative and reproductive
organs of other hyperaccumulator plants such as
Berkheya coddii Roessler (Asteraceae) (Robinson
et al., 2003), Senecio coronatus (Thunb.) Harv.
(Asteraceae) (Przybylowicz et al., 1995), Senecio
anomalochrous Hilliard (Mesjasz-Przybylowicz et al.,
2001), Stackhousia tryonii Bailey (Stackhousiaceae)
(Bhatia et al., 2003), and Sebertia acuminata Pierre ex
Baill. (Sapotaceae) (Perrier et al., 2004), has recently
begun.
There are two well-known Ni hyperaccumulators
from the Brassicaceae that grow naturally on ultra-
mafic soils of the Iberian Peninsula: Alyssum serpylli-
folium ssp. lusitanicum Dudley & P. Silva and
A. serpyllifolium ssp. malacitanum Rivas Goday ex G.
Lopez (Menezes de Sequeira, 1969; Brooks &
Radford, 1978; Brooks et al., 1981a, Asensi et al.,
2004). A. serpyllifolium ssp. malacitanum is a facul-
tative serpentine taxon related to secondary scrub
communities growing on soils of dolomite and
peridotite ultramafic rocks of the Betic Province
(Rivas-Martınez et al., 1991). This taxon is repre-
sentative of the ultramafic chamaephyte vegetation of
southern Spain (Malaga) that belongs to the endemic
alliance Staehelino-Ulicion baetici. It is located in the
western Betic Cordillera of Malaga (Sierra Bermeja,
Palmitera, Alpujata and Carratraca), one of the
largest areas (more than 300 km2) of ultramafic rocks
in the Iberian Peninsula (Asensi et al., 2004). It
constitutes an extensively serpentinized area with
abundant iron oxides produced by hydrothermal
activity (Brooks et al., 1981a). Total Ni concentra-
tions of between 1,532 and 4,254 mg kg71 have been
obtained for soils from these areas (Rufo et al., 2005).
The second subspecies, A. serpyllifolium ssp. lusitani-
cum, belongs to the dwarf scrub serpentine vegetation
growing on the highly Mg-rich ultramafic outcrops
(80 km2) in north-east Portugal (Braganca – Vinhais,
Macedo de Cavaleiros – Mogadoauro) (Asensi et al.,
2004; Sequeira & Pinto da Silva, 1992). Total soil Ni
concentrations between 851 and 2962 mg kg71 have
been reported in these areas (Peterson et al., 2003).
The bioclimatic characterization of both areas corre-
sponds to the Mediterranean macrobioclimate, with a
marked summer drought (Rivas-Martınez et al.,
2002), an extensive dry – subhumid thermomediter-
ranean to dry mesomediterranean level in southern
Spain, and a subhumid supramediterranean to dry
mesomediterranean level in Portugal.
Previous studies tested the Ni tolerance of both
subspecies (Brooks et al., 1981a) in comparison to
the non-accumulator A. serpyllifolium Desf. ssp.
serpyllifolium. They also investigated the chemical
form of Ni in the leaves, establishing a relation
between Ni and malic and malonic acids (Brooks
et al., 1981b), in an attempt to explain the
physiological function of Ni in these taxa. The Ni-
hyperaccumulating capacity of these taxa has been
recently analysed and compared by Asensi et al.,
(2004), who established that the A. serpyllifolium ssp.
malacitanum population of Sierra Bermeja reaches
higher Ni accumulation levels (1,880 – 2,810 mg Ni
kg71 DW) than the Portuguese A. serpyllifolium ssp.
lusitanicum population (1,990 mg Ni kg71 DW).
These Ni concentrations contrast with the 2 mg
kg71 DW Ni found in the non-accumulator
A. serpyllifolium ssp. serpyllifolium from the dolomitic
area of Sierra de Cazulas (Malaga, Spain).
Having established the total concentrations of Ni
in the Iberian subspecies of A. serpyllifolium, the aim
of this work was to determine the comparative
distribution of this element in different plant tissues
using energy-dispersive X-ray microanalysis coupled
to SEM.
Materials and methods
Plant samples
All plant samples are from the natural environment
of ultramafic areas from Sierra Bermeja (Malaga,
Spain) and Braganca (Portugal). Branches, leaves
and fruits from three specimens of A. serpyllifolium
ssp. malacitanum Rivas Goday ex G. Lopez and three
of A. serpyllifolium ssp. lusitanicum Dudley & P. Silva
from Braganca were collected directly in the field,
from a shrub community growing in rocky areas.
Herbarium voucher specimens are preserved in the
MAF and MGC Herbarium (Faculty of Pharmacy,
Universidad Complutense in Madrid, Spain) and in
personal collections. The material from A. serpyllifo-
lium ssp. serpyllifolium was obtained from specimens
preserved in the MAF Herbarium (MAF 82506)
collected in the dolomitic area of Sierra de Cazulas.
Sample preparation for SEM
Micromorphological analysis of leaves, stems and
seeds using SEM – EDXS were performed on the
Iberian subspecies of A. serpyllifolium. There are
epidermal elements in these plants, such as tri-
chomes, which massively cover leaves, cuticles, fruits
and to a lesser degree the stems, and for this reason
we treated trichomes as an independent unit.
It has been suggested that cryogenic SEM-EDXS
techniques (Kupper et al., 2001; Broadhurst et al.,
2004a) coupled with the use of frozen bulk samples
may be ideal to study Ni accumulation since there is
minimal processing of the samples and structural
features can be easily identified. However, according
Nickel distribution in A. serpyllifolium 171

Dow
nloa
ded
By:
[de
la F
uent
e, V
.] A
t: 12
:17
12 J
une
2007
to Psaras et al. (2000) no significant changes in Ni
distribution can be observed when dry instead of
fresh and/or frozen material is used if care is taken to
avoid sample embedding. We chose the Psaras et al.
(2000) method because it allowed us to use material
preserved in herbarium collections.
To obtain micro-morphological SEM images of
leaf and stem cross sections, samples were treated in
order to enhance the visualization of the different
plant tissues. Dry leaf and stem samples were fixed
in situ with formyl acetic alcohol (FAA). After
washing with a 0.1 M phosphate buffer (pH 7.4)
they were dehydrated through a graded ethanol
series. Then, they were cut with a sharp blade and
mounted onto stubs.
The following tissues were used for the semi-
quantitative SEM-EDX analysis: (i) from leaves,
upper and lower epidermal cell walls and cell
content, mesophyll, and vascular bundles; (ii) from
stems, epidermis, cortex, central cylinder and pith;
and (iii) from seeds, endosperm and testa. Tri-
chomes, which massively cover leaves, fruits and to a
lesser degree stems, were treated as an independent
unit. Sample preparation was done following the
methodology described by Psaras et al., (2000). For
each type of tissue four to five samples were analysed
from each species. Cross-sections of leaf and stem
were cut with a sharp blade and mounted flat on the
surfaces of conductive graphite stubs and sputters,
and then gold-coated in a BIO-RAD SC 502
apparatus for electrical conductivity in order to
prevent charging under the electron beam. Samples
were examined with a Hitachi S-3000N (Japan)
SEM using an acceleration voltage of 20 kV and a
working distance for analysis of 15 mm. During
analysis the sample stage was at room temperature.
The qualitative element composition of samples was
determined by energy dispersive X-Ray microanaly-
sis using an INCAx-sight with a Si-Li Detector
(Oxford, UK), with a detection limit of 10% of the
main element. This instrument is able to detect the
lighter elements, C, O and N, and the quantitative
numerical data of the obtained spectra are referenced
as default to the highest peak obtained in each
spectrum, which in our case generally corresponded
to C. Mapping was displayed using the Microanalysis
Suite, INCA Suite version 3.04 (Oxford, UK)
software. The grid of points for microanalysis was
set at ultra-fine resolution.
Data analysis
Quantitative numerical data were extracted directly
from spectra in % weight. Data analysis was carried
out using Statistica release 6.0 (Statsoft Inc., Tulsa,
USA). Means, standard deviations and standard
errors were calculated. Data were log-transformed
after being tested for normality with the Shapiro Wilk
test (p4 0.05) and for homogeneity of variance with
the Levene test (p4 0.05). In order to test possible
differences between adaxial and abaxial leaf epider-
mis and cuticle metal concentrations we used the
Student’s t-test. Measures from different tissues were
compared by one-way analysis of variance. Least
significant differences (LSD) between means were
calculated only if an F – test was significant at the
0.05 level of probability.
Results
Leaf
The leaf components analysed by SEM – EDXS
were: epidermal cell walls, the interior of epidermal
cells, mesophyll tissue, and vascular bundles (Figure
1). Micro-morphological SEM analyses showed the
presence of trichomes on both surfaces in the two
subspecies analysed. Transverse leaf sections showed
a dominance of palisade parenchyma on both leaf
sides (Figure 1). Stomata were only present on leaf
abaxial surfaces.
No significant differences in Ni, Ca and Mg
concentrations between adaxial and abaxial epider-
mal cell walls and epidermal cells were detected; thus
the two surfaces were treated as a whole.
There were significant differences between Ni
concentration of leaf tissues in the two subspecies
(Table I). The highest relative concentration of Ni
was found inside epidermal cells with 5.75+ 0.97%
(mean+SE) for A. serpyllifolium ssp. malacitanum,
and 4.94+ 0.34% for A. serpyllifolium ssp. lusitani-
cum. Epidermal cell walls, mesophyll tissue and
vascular bundles had significantly lower Ni concen-
trations than the inside of epidermal cells (Table I).
Values obtained for these tissues oscillated around
0.92+ 0.12%, but A. serpyllifolium ssp. malacitanum
they were below the level of instrument detection with
the following Ni distribution pattern: inside epider-
mal cell4 epidermal cell wall – mesophyll (Figure
2A). In the case of A. serpyllifolium ssp. lusitanicum Ni
concentration in epidermal cell walls, and mesophyll
and vascular bundle tissues varied between 0.25+0.13 and 2.14+ 0.37%. Thus, its Ni distribution
pattern was: inside epidermal cell4 epidermal cell
wall4mesophyll – vascular bundle (Figure 2A).
Ca showed a homogeneous distribution in leaf
tissues of both taxa (Figure 2B). As shown in Table I,
only slight differences were found only for the Ca
contained inside epidermal cells of A. serpyllifolium
ssp. lusitanicum, which is higher than the Ca content
in the rest of the leaf tissues of this taxon. Con-
centrations of this element acquired values between
0.33+ 0.06 and 1.12+ 0.31% for A. serpyllifolium
ssp. malacitanum, and 0.18+ 0.05 – 0.65+ 0.08%
172 V. de la Fuente et al.

Dow
nloa
ded
By:
[de
la F
uent
e, V
.] A
t: 12
:17
12 J
une
2007
for A. serpyllifolium ssp. lusitanicum. Often Ca was
present as cystoliths and crystals inside vascular
bundle cells.
Mg concentrations were significantly different in
different leaf tissues of both taxa (Table I), with a
higher concentration inside epidermal cells (Figure
2C). Mg content of leaf tissues of A. serpyllifolium ssp.
lusitanicum were more homogeneous (0.35+ 0.09 –
0.69+ 0.06%) than those of A. serpyllifolium ssp.
malacitanum (0.22+ 0.04 – 1.53+ 0.22%).
Elemental mapping of abaxial leaf surfaces was
carried out to complete the leaf study. Figure 3
shows the distribution of Ni, Ca and Mg in the
different cells of the leaf surface. These results clearly
show that the presence of Ni is homogeneous at the
cellular level, while Mg and Ca are almost absent.
Stem
The epidermis, cortex, central cylinder and pith were
the stem tissues analysed by SEM – EDXS (Figure 4).
Both taxa showed a similar Ni distribution pattern
between the different stem tissues (Figure 2D). The
highest Ni concentrations were found in epidermis
and cortex tissues (0.50+ 0.11 – 0.80+ 0.23%),
with no significant differences between their means
(Table II). Central cylinder and pith Ni concentra-
tions are clearly lower than those of epidermis and
cortex, and ranged from 0.09+ 0.02% to below
detection limits.
Ca distribution in stem tissues of both taxa was
similar (Figure 2E). As shown in Table II, the
highest concentrations of this element were found in
epidermis and cortex tissues (0.83+ 0.18 –
1.69+ 1.01%), while the central cylinder and pith
had lower quantities of Ca (0.10+ 0.03 –
0.60+ 0.18%). Ca crystals were common in pith,
cortex and epidermis of both Alyssum subspecies.
Mg concentrations were significantly different
in different stem tissues of both taxa (Table II). In
both cases, a higher Mg concentration occurred
in epidermis and cortex tissues (Figure 2F). Mg
values ranged from 0.22+ 0.07 – 0.65+ 0.27% for
A. serpyllifolium ssp. malacitanum, and 0.02+ 0.01 –
0.66+ 0.15% for A. serpyllifolium ssp. lusitanicum.
Trichomes
Both Alyssum subspecies have many trichomes on leaf
and stem surfaces, which give a tomentose-whitish
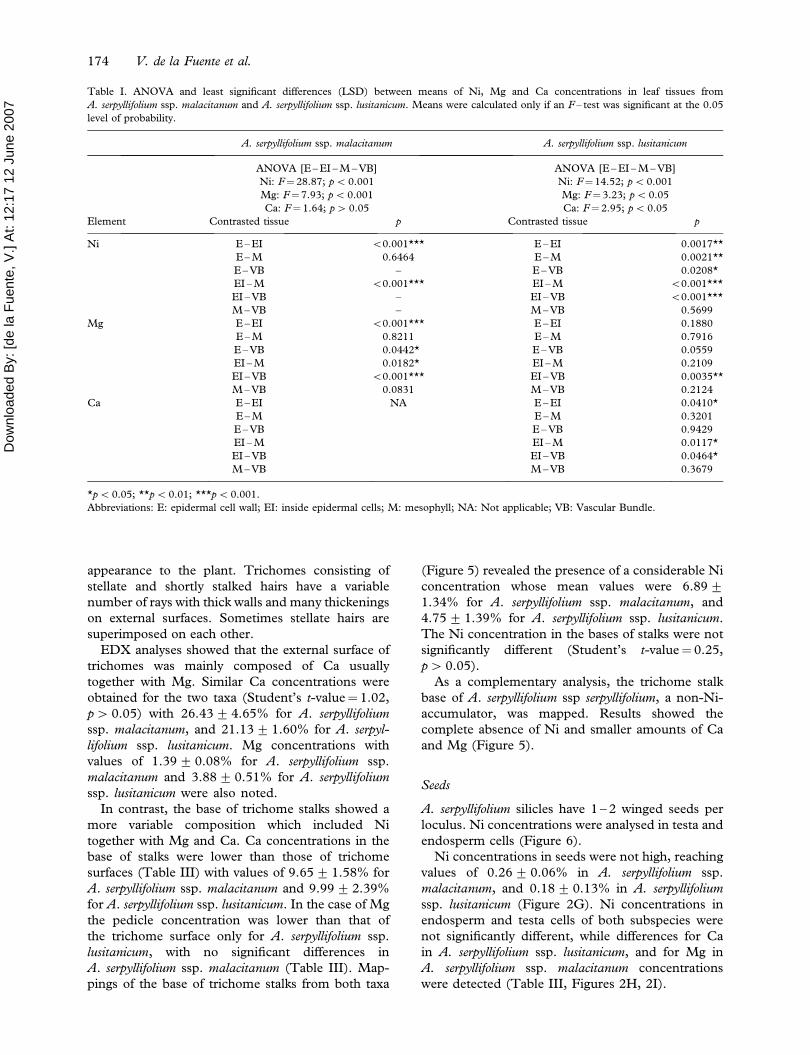
Figure 1. SEM micrographs of a cross section of an Alyssum serpyllifolium ssp. lusitanicum leaf (A, B). Energy-dispersive X-ray microanalysis
of IE (C) and VB (D) of Alyssum serpyllifolium ssp. lusitanicum. Abbreviations: E: epidermal cell walls; IE: inside epidermal cells; M: inside
mesophyll cells; VB: vascular bundle cells; T: trichome.
Nickel distribution in A. serpyllifolium 173
Dow
nloa
ded
By:
[de
la F
uent
e, V
.] A
t: 12
:17
12 J
une
2007
appearance to the plant. Trichomes consisting of
stellate and shortly stalked hairs have a variable
number of rays with thick walls and many thickenings
on external surfaces. Sometimes stellate hairs are
superimposed on each other.
EDX analyses showed that the external surface of
trichomes was mainly composed of Ca usually
together with Mg. Similar Ca concentrations were
obtained for the two taxa (Student’s t-value¼ 1.02,
p4 0.05) with 26.43+ 4.65% for A. serpyllifolium
ssp. malacitanum, and 21.13+ 1.60% for A. serpyl-
lifolium ssp. lusitanicum. Mg concentrations with
values of 1.39+ 0.08% for A. serpyllifolium ssp.
malacitanum and 3.88+ 0.51% for A. serpyllifolium
ssp. lusitanicum were also noted.
In contrast, the base of trichome stalks showed a
more variable composition which included Ni
together with Mg and Ca. Ca concentrations in the
base of stalks were lower than those of trichome
surfaces (Table III) with values of 9.65+ 1.58% for
A. serpyllifolium ssp. malacitanum and 9.99+ 2.39%
for A. serpyllifolium ssp. lusitanicum. In the case of Mg
the pedicle concentration was lower than that of
the trichome surface only for A. serpyllifolium ssp.
lusitanicum, with no significant differences in
A. serpyllifolium ssp. malacitanum (Table III). Map-
pings of the base of trichome stalks from both taxa
(Figure 5) revealed the presence of a considerable Ni
concentration whose mean values were 6.89+1.34% for A. serpyllifolium ssp. malacitanum, and
4.75+ 1.39% for A. serpyllifolium ssp. lusitanicum.
The Ni concentration in the bases of stalks were not
significantly different (Student’s t-value¼ 0.25,
p4 0.05).
As a complementary analysis, the trichome stalk
base of A. serpyllifolium ssp serpyllifolium, a non-Ni-
accumulator, was mapped. Results showed the
complete absence of Ni and smaller amounts of Ca
and Mg (Figure 5).
Seeds
A. serpyllifolium silicles have 1 – 2 winged seeds per
loculus. Ni concentrations were analysed in testa and
endosperm cells (Figure 6).
Ni concentrations in seeds were not high, reaching
values of 0.26+ 0.06% in A. serpyllifolium ssp.
malacitanum, and 0.18+ 0.13% in A. serpyllifolium
ssp. lusitanicum (Figure 2G). Ni concentrations in
endosperm and testa cells of both subspecies were
not significantly different, while differences for Ca
in A. serpyllifolium ssp. lusitanicum, and for Mg in
A. serpyllifolium ssp. malacitanum concentrations
were detected (Table III, Figures 2H, 2I).
Table I. ANOVA and least significant differences (LSD) between means of Ni, Mg and Ca concentrations in leaf tissues from
A. serpyllifolium ssp. malacitanum and A. serpyllifolium ssp. lusitanicum. Means were calculated only if an F – test was significant at the 0.05
level of probability.
A. serpyllifolium ssp. malacitanum A. serpyllifolium ssp. lusitanicum
ANOVA [E – EI – M – VB] ANOVA [E – EI – M – VB]
Ni: F¼28.87; p5 0.001 Ni: F¼14.52; p50.001
Mg: F¼7.93; p5 0.001 Mg: F¼3.23; p50.05
Ca: F¼ 1.64; p4 0.05 Ca: F¼2.95; p5 0.05
Element Contrasted tissue p Contrasted tissue p
Ni E – EI 50.001*** E – EI 0.0017**
E – M 0.6464 E – M 0.0021**
E – VB – E – VB 0.0208*
EI – M 50.001*** EI – M 50.001***
EI – VB – EI – VB 50.001***
M – VB – M – VB 0.5699
Mg E – EI 50.001*** E – EI 0.1880
E – M 0.8211 E – M 0.7916
E – VB 0.0442* E – VB 0.0559
EI – M 0.0182* EI – M 0.2109
EI – VB 50.001*** EI – VB 0.0035**
M – VB 0.0831 M – VB 0.2124
Ca E – EI NA E – EI 0.0410*
E – M E – M 0.3201
E – VB E – VB 0.9429
EI – M EI – M 0.0117*
EI – VB EI – VB 0.0464*
M – VB M – VB 0.3679
*p50.05; **p50.01; ***p5 0.001.
Abbreviations: E: epidermal cell wall; EI: inside epidermal cells; M: mesophyll; NA: Not applicable; VB: Vascular Bundle.
174 V. de la Fuente et al.

Dow
nloa
ded
By:
[de
la F
uent
e, V
.] A
t: 12
:17
12 J
une
2007
Discussion
This is the first report concerning Ni localization in
populations of A. serpyllifolium ssp. lusitanicum and
A. serpyllifolium ssp. malacitanum growing naturally
on ultramafic areas of the Iberian Peninsula.
Present results show that structural features of
the different analysed tissues are preserved, and
that the distribution of Ni in these structures has a
pattern similar to that reported by Kupper et al.
(2001), Broadhurst et al. (2004a) and Mcnear
et al. (2005) in other Alyssum Ni-hyperaccumulator
species.
Both analysed taxa showed a similar Ni distribu-
tion pattern among tissues of the different vegetative
and reproductive organs selected for the study. In
both Alyssum species, the Ni abundance in leaves is
in the following order: inside epidermal cells4 epi-
dermal cell walls4mesophyll and vascular bundle
cells. The large amount of Ni found in epidermal
cells is probably located inside vacuoles, as reported
by Brooks et al., (1981b) in these two Alyssum
Iberian subspecies, and recently confirmed in other
Ni-hyperaccumulating Mediterranean Brassicaceae
(Kupper et al., 2001; Psaras et al., 2000; Broadhurst
et al., 2004a; Mcnear et al., 2005) and species from
other families (Kramer at al. 1997; Bidwell et al.,
2004). Ni was found to be homogeneously distrib-
uted on both epidermal surfaces. If was also
preferentially located in epidermis and cortex stem
tissues although in lower concentrations than in
leaves. The preferential location in external tissues
Figure 2. (A) Ni concentrations in leaf tissues of A. serpyllifolium ssp. malacitanum and A. serpyllifolium ssp. lusitanicum. (B) Ca concentrations
in leaf tissues of A. serpyllifolium ssp. malacitanum and A. serpyllifolium ssp. lusitanicum. (C) Mg concentrations of leaf tissues A. serpyllifolium
ssp. malacitanum and A. serpyllifolium ssp. lusitanicum. (D) Ni concentrations in stem tissues of A. serpyllifolium ssp. malacitanum and
A. serpyllifolium ssp. lusitanicum. (E) Ca concentrations in stem tissues of A. serpyllifolium ssp. malacitanum and A. serpyllifolium ssp.
lusitanicum. (F) Mg concentrations of stem tissues of A. serpyllifolium ssp. malacitanum and A. serpyllifolium ssp. lusitanicum. (G) Ni
concentrations in seeds tissues of A. serpyllifolium ssp. malacitanum and A. serpyllifolium ssp. lusitanicum. (H) Ca concentrations in seeds
tissues of A. serpyllifolium ssp. malacitanum and A. serpyllifolium ssp. lusitanicum. (I) Mg concentrations in seeds tissues of A. serpyllifolium ssp.
malacitanum and A. serpyllifolium ssp. lusitanicum. Means and standard errors. All concentrations are expressed in %. Leaf: n¼ 53 – 6; Stem:
n¼34 – 11; Seeds: n¼ 12 – 3; Trichomes: n¼ 17 – 3. Abbreviations: C: cortex; CC: central cylinder; E: epidermal cell wall; EI: inside
epidermal cells; En: endosperm; M: mesophyll; P: pith; T: testa; VB: vascular bundle.
Nickel distribution in A. serpyllifolium 175

Dow
nloa
ded
By:
[de
la F
uent
e, V
.] A
t: 12
:17
12 J
une
2007
may indicate the existence of efficient selective Ni
translocation mechanisms.
Previous studies have shown discrepancies con-
cerning the localization of Ni in the trichomes of
Alyssum species. Kramer et al. (1997) suggested that
Ni was preferentially accumulated in the epidermal
trichomes. Psaras et al. (2000) reported that Ni was
excluded from trichomes of various Alyssum species
from Greece. The latter observation was supported by
Kupper et al. (2001) in two Alyssum species, although
these authors pointed to the presence of Ni in the basal
part of trichomes after staining them with dimethyl-
glyoxime. Recent reports on Ni location in Alyssum
showed the existence of large amounts of Ni in
epidermal tissues, thereby emphasizing the presence
of this element, together with Mn in leaf trichomes
(Broadhurst et al., 2004a,b; Mcnear et al., 2005).
In the present study, EDX scans clearly revealed
the presence of Ni in the base of trichomes stalks. Ni
concentrates on the basal surface and in the interior
of the pedicle of trichomes. This distribution is not
easy to observe because it requires that the trichomes
be kept in an inverted position, and not in their
normal position (Psaras et al., 2000), due to the high
Ca content of the cell walls that impedes electron
beam penetration. Ca is the most abundant element
in trichomes, and Ni is distributed under the Ca,
which occupies a more external position. Ni was not
detected in the external thickening of trichome cell
walls, which have a higher Ca concentration when
compared with other morphological structures of the
plant. The existence of calcareous deposits, calcified
cells walls and trichomes are restricted to a compara-
tively small number of families, Brassicaceae
amongst them (Metcalfe, 1983). Accumulation of
other metals (Zn and Cd) in the base of trichomes
has been observed for another Brassicaceae, Arabi-
dopsis halleri L., by Kupper et al. (2000) and Zhao
et al. (2000).
Information regarding the presence of Ni in seeds
is not frequently found (Przybylowicz et al., 1995;
Psaras & Manetas, 2001; Bhatia et al., 2003). We
report here the localization of Ni in the testa and
endosperm of the Iberian Alyssum taxa studied. Ni
concentration in seeds was lower than in vegetative
organs, as previously described (Brooks, 1998;
Psaras & Manetas, 2001).
Different interpretations have been proposed for
metal hyperaccumulation in plants. Several authors
interpreted Ni accumulation as a defense mechanism
against herbivore and pathogen attacks (Boyd &
Martens, 1998). Robinson et al. (2003), among
others, disagree with this interpretation because of
Figure 4. SEM micrograph of a cross section of Alyssum
serpyllifolium ssp. lusitanicum stem. Abbreviations: C: cortex; CC:
central cylinder; E: epidermis; P: pith.
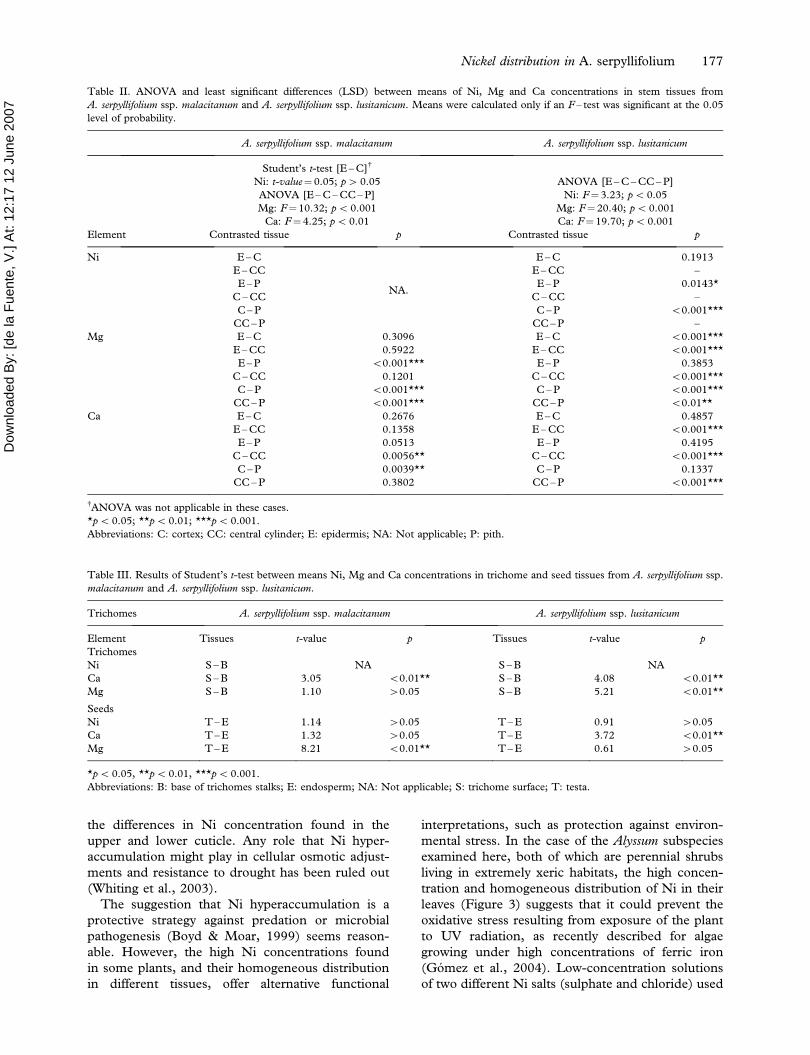
Figure 3. SEM micrograph of an abaxial surface of leaf epidermis of Alyssum serpyllifolium ssp. lusitanicum. The dot maps show the
distribution of Ni (blue), Mg (orange) and Ca (green) by EDXS analyses. For colour figures, please visit http://www.informaworld.com/
TPLB.
176 V. de la Fuente et al.
Dow
nloa
ded
By:
[de
la F
uent
e, V
.] A
t: 12
:17
12 J
une
2007
the differences in Ni concentration found in the
upper and lower cuticle. Any role that Ni hyper-
accumulation might play in cellular osmotic adjust-
ments and resistance to drought has been ruled out
(Whiting et al., 2003).
The suggestion that Ni hyperaccumulation is a
protective strategy against predation or microbial
pathogenesis (Boyd & Moar, 1999) seems reason-
able. However, the high Ni concentrations found
in some plants, and their homogeneous distribution
in different tissues, offer alternative functional
interpretations, such as protection against environ-
mental stress. In the case of the Alyssum subspecies
examined here, both of which are perennial shrubs
living in extremely xeric habitats, the high concen-
tration and homogeneous distribution of Ni in their
leaves (Figure 3) suggests that it could prevent the
oxidative stress resulting from exposure of the plant
to UV radiation, as recently described for algae
growing under high concentrations of ferric iron
(Gomez et al., 2004). Low-concentration solutions
of two different Ni salts (sulphate and chloride) used
Table II. ANOVA and least significant differences (LSD) between means of Ni, Mg and Ca concentrations in stem tissues from
A. serpyllifolium ssp. malacitanum and A. serpyllifolium ssp. lusitanicum. Means were calculated only if an F – test was significant at the 0.05
level of probability.
A. serpyllifolium ssp. malacitanum A. serpyllifolium ssp. lusitanicum
Student’s t-test [E – C]{
Ni: t-value¼ 0.05; p4 0.05 ANOVA [E – C – CC – P]
ANOVA [E – C – CC – P] Ni: F¼3.23; p50.05
Mg: F¼10.32; p5 0.001 Mg: F¼20.40; p50.001
Ca: F¼ 4.25; p5 0.01 Ca: F¼19.70; p5 0.001
Element Contrasted tissue p Contrasted tissue p
Ni E – C
NA.
E – C 0.1913
E – CC E – CC –
E – P E – P 0.0143*
C – CC C – CC –
C – P C – P 50.001***
CC – P CC – P –
Mg E – C 0.3096 E – C 50.001***
E – CC 0.5922 E – CC 50.001***
E – P 50.001*** E – P 0.3853
C – CC 0.1201 C – CC 50.001***
C – P 50.001*** C – P 50.001***
CC – P 50.001*** CC – P 50.01**
Ca E – C 0.2676 E – C 0.4857
E – CC 0.1358 E – CC 50.001***
E – P 0.0513 E – P 0.4195
C – CC 0.0056** C – CC 50.001***
C – P 0.0039** C – P 0.1337
CC – P 0.3802 CC – P 50.001***
{ANOVA was not applicable in these cases.
*p50.05; **p50.01; ***p5 0.001.
Abbreviations: C: cortex; CC: central cylinder; E: epidermis; NA: Not applicable; P: pith.
Table III. Results of Student’s t-test between means Ni, Mg and Ca concentrations in trichome and seed tissues from A. serpyllifolium ssp.
malacitanum and A. serpyllifolium ssp. lusitanicum.
Trichomes A. serpyllifolium ssp. malacitanum A. serpyllifolium ssp. lusitanicum
Element Tissues t-value p Tissues t-value p
Trichomes
Ni S – B NA S – B NA
Ca S – B 3.05 50.01** S – B 4.08 50.01**
Mg S – B 1.10 40.05 S – B 5.21 50.01**
Seeds
Ni T – E 1.14 40.05 T – E 0.91 40.05
Ca T – E 1.32 40.05 T – E 3.72 50.01**
Mg T – E 8.21 50.01** T – E 0.61 40.05
*p50.05, **p50.01, ***p5 0.001.
Abbreviations: B: base of trichomes stalks; E: endosperm; NA: Not applicable; S: trichome surface; T: testa.
Nickel distribution in A. serpyllifolium 177

Dow
nloa
ded
By:
[de
la F
uent
e, V
.] A
t: 12
:17
12 J
une
2007
Figure 5. SEM micrographs of the lower surface of a trichome of A. serpyllifolium ssp. mlacitanum (A), A. serpyllifolium ssp. lusitanicum (B),
A. serpyllifolium ssp. serpyllifolium (C). The dot maps show the distribution of Ni (blue), Mg (orange) and Ca (green) by EDXS analyses. For
colour figures, please visit http://www.informaworld.com/TPLB.
Figure 6. SEM micrographs of: (A) a seed of A. serpyllifolium ssp. lusitanicum. (B) micromorphology of seed epidermal surface of
A. serpyllifolium ssp. malacitanum. (C) cross section of A. serpyllifolium ssp. lusitanicum seed. (D) epidermal wing surface of a seed of
A. serpyllifolium ssp. lusitanicum. Abbreviations: E: endosperm; T: testa.
178 V. de la Fuente et al.

Dow
nloa
ded
By:
[de
la F
uent
e, V
.] A
t: 12
:17
12 J
une
2007
to study transmittance showed a high percentage of
absorbance in specific regions of the UV range,
suggesting that an even distribution of this metal on
the leaves might have a protective effect. Work is in
progress to evaluate this additional function for Ni in
hyperaccumulator Alyssum subspecies grown in xero-
edaphic climatic conditions.
References
Asensi A, Rodrıguez N, Dıez-Garretas B, Amils R, de la Fuente V.
2004. Nickel hyperaccumulation of some subspecies of Alyssum
serpyllifolium (Brassicaceae) from ultramafic soils of Iberian
Peninsula. In: Boyd RS, Baker AJM, Proctor J, editors.
Ultramafic rocks: their soils, vegetation and fauna. Proceedings
of the IV International Conference on Serpentine Ecology, 2003,
April 21 – 26 Havana, Cuba. Science Reviews. pp. 263 – 265.
Baker AJM, McGrath SP, Reeves RD, Smith JAC. 2000. Metal
hyperaccumulator plants: a review of the ecology and
physiology of a biochemical resource for phytoremediation of
metal-polluted soils. In: Terry N, Nanuelos G, editors.
Phytoremediation of contaminated soil and water. Boca Raton,
USA: Lewis Publishers. pp. 85 – 107.
Bhatia NP, Orlic I, Siegele R, Ashwath N, Bajer AJM, Walsh KB.
2003. Elemental mapping using PIXE shows the main pathway
of nickel movement is principally simplistic within the fruit of
the hyperaccumulator Stackhousia tryonii. New Phytol
160:479 – 488.
Bidwell SD, Crawford SA, Woodrow IE, Sommer-Knudsen J,
Marshall AT. 2004. Sub-cellular localization of Ni in the
hyperaccumulator, Hybanthus floribundus (Lindley) F. Muell.
Plant Cell Environ 27:705 – 716.
Boyd RS, Martens SN. 1998. Nickel hyperaccumulated by Thlaspi
montanum var. montanum is acutely toxic to an insect hervibore.
Oikos 70:21 – 25.
Boyd RS, Moar WJ. 1999. The defensive function of Ni in plants:
response of the polyphagos herbivore Spodoptera exigua
(Lepidoptera: Noctuidae) to hyperaccumulator and accumu-
lator species of Streptanthus (Brassicaceae). Oecologia
118:218 – 224.
Broadhurst CL, Chaney RL, Angle JA, Erbe EF, Maugel TK.
2004a. Nickel localization and response to increasing Ni soil
levels in leaves of the Ni hyperaccumulator Alyssum murale.
Plant Soil 265:225 – 242.
Broadhurst CL, Chaney RL, Angle JS, Erbe EF, Murphy CA.
2004b. Simultaneous hyperaccumulation of Nickel, Manga-
nese and Calcium in Alyssum leaf trichomes. Environ Sci
Technol 28:5797 – 5802.
Brooks RR. 1998. Geobotany and hyperaccumulators. In: Brooks
RR, editor. Plants that hyperaccumulate heavy metals. New
York, USA: CAB International. pp. 55 – 94.
Brooks RR, Randford CC. 1978. Nickel accumulation by
European species of the genus Alyssum. Proc Royal Soc Lon
Ser B 200:217 – 224.
Brooks RR, Lee J, Reeves RD, Jaffre T. 1977. Detection of
nickeliferous rocks by analysis of herbarium specimens of
indicator plants. J Geochem Explor 7:49 – 57.
Brooks RR, Shaw S, Asensi A. 1981a. Some observations on the
ecology, metal uptake and nickel tolerance of Alyssum
serpyllifolium subspecies from the Iberian Peninsula. Vegetatio
45:183 – 188.
Brooks RR, Shaw S, Asensi A. 1981b. The chemical form and
physiological function of nickel in some Iberian Alyssum
species. Physiol Plantarum 51:167 – 170.
Davis MA, Boyd RS. 2000. Dynamics of Ni-based defence and
organic defences in the Ni hyperaccumulator, Streptanthus
polygaloides (Brassicaceae). New Phytol 146:211 – 217.
Gomez F, Grau A, Vazquez L, Amils R. 2004. UV radiation effects
over microorganisms and study of protective agents. Proceed-
ings of the III European Workshop on Exo-Astrobiology,
Mars, The Search for Life. Madrid, Spain, 18 – 20 November
2003. pp. 21 – 26.
Heath SM, Southworth D, D’allura JA. 1997. Localization of
nickel in epidermal subsidiary cells of leaves of Thalspi
montanum var. siskiyouense (Brassicaceae) using energy-
dispersive X-Ray microanalysis. Int J Plant Sci 158:184 –
188.
Jaffre T, Brooks RR, Lee J, Reeves RD. 1976. Sebertia acuminata a
hyperaccumulator of nickel from New Caledonia. Science
193:579 – 580.
Kramer U, Grime GW, Smith JAC, Hawes CR, Baker AJM. 1997.
Micro-PIXE as a technique for studying nickel localization in
the leaves of the hyperaccumulator plant Alyssum lesbiacum.
Nucl Instrum Meth Phys Res 130:346 – 350.
Kupper H, Lombi E, Zhao FJ and McGrath SP. 2000. Cellular
compartmentation of cadmium and zinc in relation to other
elements in the hyperaccumulator Arabidopsis halleri. Planta
212:75 – 84.
Kupper H, Lombi E, Zhao FJ, Wieshammer G, McGrath SP.
2001. Cellular compartimentation of nickel in the hyperaccu-
mulator Alyssum lesbiacum, Alyssum bertolonii and Thlaspi
goensigense. J Exp Bot 52:2291 – 2300.
Mcnear DH, Peltier JRE, Everhart J, Chaney RL, Sutton S,
Newvile M, et al. 2005. Application of quantitative fluores-
cence and absorption – edge computed microtomography to
image metal compartmentalization in Alyssum murale. Environ
Sci Technol 39:2210 – 2218.
Medina E, Cuevas E, Figueroa J, Lugo AE. 1994. Mineral content
of leaves from trees growing on serpentine soils under
contrasting rainfall regimes in Puerto Rico. Plant Soil
158:13 – 21.
Menezes de Sequeira EM. 1969. Toxicity and movement of heavy
metals in serpentinic soils (north – eastern Portugal). Agrono-
mia Lusit 30:115 – 154.
Mesjasz-Przybylowicz J, Grodzinska K, Przybylowicz WJ, Godzik
B, Szarek-Lukaszewska G. 2001. Nuclear microprobe studies
of elemental distribution in seeds of Biscutella laevigata L. from
zinc wastes in Olkus, Poland. Nucl Instrum Meth Phys Res
181:634 – 639.
Metacalfe CR. 1983. Calcareous deposits, calcified cell walls,
cystoliths, and similar structures. In: Metacalfe CR, Chalk L,
editors. Anatomy of the dicotyledons Vol (II): Wood structure
and conclusion of the general introduction. Oxford, UK:
Clarendon Press. pp. 94 – 108.
Perrier N, Colin F, Jaffre T, Ambrosi JP, Rose J, Bottero JY. 2004.
Nickel speciation in Sebertia acuminata, a plant growing on a
lateritic soil of New Caledonia. CR Geosci 336:567 – 577.
Peterson LR, Trivett V, Baker AJM, Aguiar C, Pollard J. 2003.
Spread of metals through an invertebrate food chain as
influenced by a plant that hyperaccumulates nickel. Chemoe-
cology 13:103 – 108.
Przybylowicz WJ, Pineda CA, Prozesky VM, Mesjasz-Przybylo-
wicz J. 1995. Investigation of Ni hyperaccumulation by true
elemental imaging. Nucl Instrum Meth Phys Res 104:176 –
181.
Psaras GK, Manetas Y. 2001. Nickel localization in seeds of the
metal hyperaccumulator Thlaspi pindicum Hausskn. Ann Bot
88:513 – 516.
Psaras GK, Cosntantinidis TH, Cotsopoulos B, Manetas Y. 2000.
Relative abundance of nickel in the leaf epidermis of eight
hyperaccumulator: evidence that the metal is excluded from
guard cells and trichomes. Ann Bot 86:73 – 78.
Reeves RD. 1992. The hyperaccumulation of nickel by serpentine
plants. In: Baker AJM, Proctor J, Reeves RD, editors. The
vegetation of ultramafic (serpentine) soils. Andover, UK:
Intercept Ltd. pp. 253 – 277.
Nickel distribution in A. serpyllifolium 179

Dow
nloa
ded
By:
[de
la F
uent
e, V
.] A
t: 12
:17
12 J
une
2007
Reeves RD. 2003. Tropical hyperaccumulators of metals and their
potential for phytoextraction. Plant Soil 249:57 – 65.
Reeves RD, Baker AJM. 2000. Metal-accumulating plants. In:
Raskin I, Ensley BD, editors. Phytoremediation of toxic
metals: using plants to clean up the environment. New York,
USA: John Wiley & Sons. pp. 193 – 229.
Reeves RD, Brooks RR, MacFarlane RM. 1981. Nickel uptake by
Californian Streptanthus and Caulanthus with particular refer-
ence to the hyperaccumulator, Streptanthus polygaloides Gray
(Brassicaceae). Am J Bot 68:708 – 712.
Reeves RD, Baker AJM, Borhidi A, Berazaın R. 1996. Nickel-
accumulating plants from the ancient serpentine soils of Cuba.
New Phytol 133:217 – 224.
Reeves RD, Baker AJM, Borhidi A, Berazaın R. 1999. Nickel
hyperaccumulation in the serpentine flora of Cuba. Ann Bot
London 83:29 – 38.
Rivas-Martınez S, Asensi A, Molero J, Valle F. 1991. Endemismos
vasculares de Andalucıa. Rivasgodaya 6:5 – 76.
Rivas-Martınez S, Dıaz ET, Fernandez-Gonzalez F, Izco J, Loidi
J, Lousa M, et al. 2002. Vascular plant communities of Spain
and Portugal. Addenda to the syntaxonomical checklist of
2001. Intinera Geobot 15:5 – 432.
Robinson BH, Lombi E, Zhao FJ, McGrath SP. 2003. Uptake and
distribution of nickel and other metals in the hyperaccumulator
Berkheya coddii. New Phytol 158:279 – 285.
Rufo L, Rodrıguez N, de la Fuente V. 2005. Analisis comparado
de metales en suelos y plantas de la Sierra Bermeja. In:
Jimenez – Ballesta R, Alvarez Gonzalez AM, editors. II
Simposio Nacional de Control de la Degradacion de Suelos:
Comunicaciones. 1st ed. July 2005. Madrid, Spain: Universi-
dad Autonoma de Madrid. pp. 197 – 201.
Sequeira EM, Pinto da Silva AR. 1992. Ecology of serpentinized
areas of north-east Portugal. In: Roberts A, Proctor J, editors.
The ecology of areas with serpentinized rocks. A world view.
The Netherlands: Kluwer Academic Publishers. pp. 169 – 197.
Whiting SN, Neumann PM, Baker AJM. 2003. Nickel and zinc
hyperaccumulation by Alyssum murale and Thlaspi caerulescens
(Brassicaceae) do not enhance survival and whole-plant growth
under drought stress. Plant Cell Env 26:351 – 360.
Zhao FJ, Lombi E, Breedon T, McGrath SP. 2000. Zinc
hyperaccumulation and cellular distribution in Arabidopsis
halleri. Plant Cell Environ 23:507 – 517.
180 V. de la Fuente et al.




















