
Tracing the late Holocene evolution of the NW Iberian upwelling system
21
Tracing the late Holocene evolution of the NW Iberian upwelling system Virgínia Martins a, ⁎ , Jean-Marie Jouanneau b , Olivier Weber b , Fernando Rocha a a Industrial Minerals and Clays Centre (FCT), University of Aveiro, Portugal b Department of Geology and Oceanography, Univ. of Bordeaux I/CNRS, France Received 26 May 2005; received in revised form 21 December 2005; accepted 23 December 2005 Abstract This work analyses data from the OMEX core KSGX 40 (164-cm long) collected in the Galicia Mud Deposit, from the NW Iberian outer continental shelf, off the Ria de Vigo (North Spain). Sediment grain-size and benthic foraminifera distribution patterns indicate a number of substantial changes in sedimentation and also food/oxygen availability to the benthic ecosystem during the last ∼4.8 ka cal BP. Cluster analysis (Q-mode and R-mode) based on the most abundant taxa was used to recognize similarities within the data set. Three main clusters were established by Q-mode cluster analysis. The species distribution pattern of two of these three clusters is clearly related with changes in texture of the sediments. Textural data and benthic foraminifera proxies suggest that between ∼4.8 and 2.2 ka cal BP there was a period of higher hydrodynamism, with more energetic oceanic currents associated with shelf mixed waters. The period since ∼2.2 ka cal BP until present was characterized by generally lower bottom energetic conditions and by a supply of finer sediment, which was richer in organic matter leading to more eutrophic conditions in the benthic environment, mainly between ∼2.2 and 1.2 ka cal BP and since ∼0.5 ka cal BP until the present. Eutrophication was probably determined by strong water column stratified conditions and by coastal upwelling-dominated depositional intervals and/or by higher lateral flux of organic carbon. Since the upwelling increasing periods are in general due the intensification of northerly winds, both most productive upwelling-dominated periods may be attributed to climatic/oceanographic induced changes during the Sub-Atlantic climate. Small sea levels oscillations also should contribute to these results. Eutrophication since the end of Little Ice Age may be related to human influence in coastal areas. © 2006 Published by Elsevier B.V. Keywords: Holocene; NW Iberian upwelling system; stratification; productivity benthic foraminifera proxies 1. Introduction 1.1. Benthic foraminifera as a proxy of productivity Foraminifera are abundant and ubiquitous inhabitants of the marine benthos. The variability of the organic carbon flux to the sea floor has an important environmental influence on the benthic foraminifera (e.g., Altenbach and Sarnthein, 1989; Loubere and Fariduddin, 1999), because surface-derived organic matter is the primary energy source for most of the benthic foraminifera (Wollenburg and Mackensen, 1998). Some benthic foraminifera species react rapidly to the input of organic matter from primary production, exhibiting an increase in population size after Marine Micropaleontology 59 (2006) 35 – 55 www.elsevier.com/locate/marmicro ⁎ Corresponding author. E-mail address: [email protected] (V. Martins). 0377-8398/$ - see front matter © 2006 Published by Elsevier B.V. doi:10.1016/j.marmicro.2005.12.002
Transcript of Tracing the late Holocene evolution of the NW Iberian upwelling system

y 59 (2006) 35–55www.elsevier.com/locate/marmicro
Marine Micropaleontolog
Tracing the late Holocene evolution of the NW Iberianupwelling system
Virgínia Martins a,⁎, Jean-Marie Jouanneau b, Olivier Weber b, Fernando Rocha a
a Industrial Minerals and Clays Centre (FCT), University of Aveiro, Portugalb Department of Geology and Oceanography, Univ. of Bordeaux I/CNRS, France
Received 26 May 2005; received in revised form 21 December 2005; accepted 23 December 2005
Abstract
This work analyses data from the OMEX core KSGX 40 (164-cm long) collected in the Galicia Mud Deposit, from the NWIberian outer continental shelf, off the Ria de Vigo (North Spain). Sediment grain-size and benthic foraminifera distribution patternsindicate a number of substantial changes in sedimentation and also food/oxygen availability to the benthic ecosystem during thelast ∼4.8 ka cal BP. Cluster analysis (Q-mode and R-mode) based on the most abundant taxa was used to recognize similaritieswithin the data set. Three main clusters were established by Q-mode cluster analysis. The species distribution pattern of two ofthese three clusters is clearly related with changes in texture of the sediments. Textural data and benthic foraminifera proxiessuggest that between ∼4.8 and 2.2 ka cal BP there was a period of higher hydrodynamism, with more energetic oceanic currentsassociated with shelf mixed waters. The period since ∼2.2 ka cal BP until present was characterized by generally lower bottomenergetic conditions and by a supply of finer sediment, which was richer in organic matter leading to more eutrophic conditions inthe benthic environment, mainly between ∼2.2 and 1.2 ka cal BP and since ∼0.5 ka cal BP until the present. Eutrophication wasprobably determined by strong water column stratified conditions and by coastal upwelling-dominated depositional intervals and/orby higher lateral flux of organic carbon. Since the upwelling increasing periods are in general due the intensification of northerlywinds, both most productive upwelling-dominated periods may be attributed to climatic/oceanographic induced changes during theSub-Atlantic climate. Small sea levels oscillations also should contribute to these results. Eutrophication since the end of Little IceAge may be related to human influence in coastal areas.© 2006 Published by Elsevier B.V.
Keywords: Holocene; NW Iberian upwelling system; stratification; productivity benthic foraminifera proxies
1. Introduction
1.1. Benthic foraminifera as a proxy of productivity
Foraminifera are abundant and ubiquitous inhabitantsof the marine benthos. The variability of the organic
⁎ Corresponding author.E-mail address: [email protected] (V. Martins).
0377-8398/$ - see front matter © 2006 Published by Elsevier B.V.doi:10.1016/j.marmicro.2005.12.002
carbon flux to the sea floor has an importantenvironmental influence on the benthic foraminifera(e.g., Altenbach and Sarnthein, 1989; Loubere andFariduddin, 1999), because surface-derived organicmatter is the primary energy source for most of thebenthic foraminifera (Wollenburg and Mackensen,1998). Some benthic foraminifera species react rapidlyto the input of organic matter from primary production,exhibiting an increase in population size after

36 V. Martins et al. / Marine Micropaleontology 59 (2006) 35–55
phytodetritus deposition (Murray, 1991). The flux oforganic matter to the sea floor is the main parameter thatcontrols bathymetrically the density and the composi-tion of benthic foraminiferal faunas (Fontanier et al.,2002). The organic flux is also the main parametercontrolling the foraminiferal distribution in sediments(microhabitat) since the supply of metabolisable organicmatter controls the oxygen consumption in the sedimentand the localization of the successive redox fronts(Jorissen et al., 1998; Fontanier et al., 2002). Severalbenthic foraminifera are known to inhabit oxygen-depleted environments (e.g., Jannink et al., 1998;Fontanier et al., 2002). Anoxic conditions do not havea direct lethal effect for many species (e.g., Jannink etal., 1998; Fontanier et al., 2002). The flux of organicmatter to the sea floor is controlled mainly by surface-water productivity, lateral supply of Corg and hydrody-namic conditions. These variables show considerablegeographic variation and reflect global climate process-es, i.e., ocean–atmosphere control of primary produc-tion (Loubere, 1996).
1.2. Regional oceanography
The NW Iberian coast is the northern limit of theEastern North Atlantic upwelling system which isdominated by the Portugal Current System (Ambarand Fiúza, 1994). During summer, the Portugal CoastalCurrent (PCC) is 30–40 km wide, 50–100 m deep andflows southward in the vicinity of the shelf break, beingdriven by upwelling-favourable northerly winds. PCCtransports recently upwelled, cold and nutrient-richEastern North Atlantic Central Water (Fiúza et al., 1982;Fiúza, 1984; Ríos et al., 1992; Pérez et al., 1993). Underupwelling conditions there are also equatorward flowson the shelf surface circulation, originated by the pre-dominance of northerly winds (e.g., Frouin et al., 1990;Huthnance, 1995). On the contrary, a poleward flowingslope undercurrent centred at depths of 150–300 m isprimarily forced by large-scale geopotential gradientsand topography (e.g., Frouin et al., 1990; Huthnance,1995).
The strength and position of the atmospheric pressurecells that rule the North Atlantic climatology, the AzoresHigh and the Iceland Low pressure centres show sea-sonal variations, forcing the along–shore winds respon-sible for the upwelling processes in the Western IberianMargin (Wooster et al., 1976; Blanton et al., 1984;Fiúza, 1984). The predominance of northeasterly windsfrom March/April to September/October is the maincause of upwelling, although the coastline orientationand the properties of subsurface waters also influence
the upwelling phenomena (Wooster et al., 1976; Blantonet al., 1984; Fiúza, 1984). During winter, the strongwinds from S–SW lead to the establishment of a down-welling regime over the shelf, characterized by per-sistent poleward flow (Vitorino et al., 2002).
One of the most characteristic features of the NWIberian upwelling system is a recurrent topography-intensified upwelling centre off Cape Finisterre (43°3′N,93°W; Fig. 1) (e.g., Haynes et al., 1993; Castro et al.,2000) and the formation of upwelling filaments at thebeginning of the upwelling season (Haynes et al., 1993).The upwelling filaments reach their maximum offshoreextent in late September, but have usually disappearedby late October (Álvarez-Salgado et al., 2001a; Hayneset al., 1993). During the upwelling period, primaryproduction is enhanced in shelf areas (Tenore et al.,1995) and inside the Rias Baixas (Hanson et al., 1986;Tilstone et al., 1999) and dissolved organic carbon ac-cumulates in surface layers (Álvarez-Salgado et al.,2001b). According to Abrantes and Moita (1999),during the upwelling regime phytoplankton biomass atthe surface is ten times higher than during the non-upwelling period. Diatoms and coccolithophorid abun-dance distribution patterns in the sediments preservemuch of their original biological spatial variability andreflect the original proportions of production generatedby the occurrence of upwelling, independently of lithol-ogy and/or the relative importance of several geologicalprocesses acting on the West Iberian Shelf.
Under upwelling conditions, the existence of welldeveloped upwelling filaments, such as the oneregularly observed south of Cape Finisterre (Haynes etal., 1993), can promote horizontal transport of bothparticulate and dissolved matter produced during up-welling events from the shelf area into oceanic waters(Álvarez-Salgado et al., 2001a; Smyth et al., 2001).Although during this season sedimentary transfers bythe Galicia shelf-waters are predominantly southwards(Jouanneau et al., 2002). In this region the organiccarbon flux depends not only on oceanic productivitybut also on detritus outwelling from the Rias Baixas,which are recognized to be high productive systems(López-Jamar et al., 1992). The organic carbon flux alsoprobably depends on significant amounts of refractoryterrestrial organic carbon released by coastal erosion andtransported into the studied area. Thus sediments aresupplied by several sources of organic matter to theGalician continental margin.
The aims of this study are to search for changes in theorganic carbon flux to the Galicia Mud Deposit and inferabout some conditioning factors acting on the outershelf off the Ria de Vigo, during the last ∼4.8 ka cal BP.

Fig. 1. Satellite Sea-surface temperature image of CCMS-PML Remote Sensing Group of 28/7/1995, with a vertical temperature scale.
37V. Martins et al. / Marine Micropaleontology 59 (2006) 35–55
This takes into view the fact that the impact of upwellingon Quaternary sediments of the Galician Rias andcontinental shelf was studied only superficially (Reyand Diaz del Rio, 1987; López-Jamar et al., 1992; Pregoet al., 1995; Prego and Bao, 1997).
2. Materials and methods
2.1. Sampling
The investigation was carried out on l-cm slices ofcore KSGX 40, 164-cm long, collected during theOMEX (Ocean Margin Exchange Project) oceanograph-ic cruise NO CÔTES DE LA MANCHE — MissionGAMINEX (8/07/1998–19/07/1998), at latitude 42°14′98″N, longitude 09°01′01″Wand sea depth 115 m. Thecore KSGX 40 is a record of the evolution of the GaliciaMud Deposit, located (Fig. 2) on the NW Iberian outercontinental shelf, off the Ria de Vigo (Northwest ofSpain).
2.2. Radiocarbon dating
Three radiocarbon dating by accelerator mass spec-trometry (AMS) ofmixed foraminifera shells (10 to 20mg)
collected in the sediment size fraction N125 μm from39–40, 69–70 and 134–135 cm were carried out by“Beta Analytic Inc.”, Miami, Florida, USA. Data wereupdated to calendar years BP using the CALIB 4.3program of Stuiver et al. (1998). The final resultscorrespond to calibrated ages (calendar years) using 2 σintervals with a standard marine reservoir correction ofapproximately 400 years to the Iberian Margin (Soares,1989; http://radiocarbon.pa.qub.ac.uk/marine/, marinereservoir correction database).
2.3. Grain size
Grain size analysis was carried out on bulk sedi-ment by Laser Diffraction in Particle Size Analysis(Mastersizer S instrument, Malvern Instruments), whichprovided the grain size distribution in the size range0.05 to 878 μm. Grain size analyses were determined inevery 1-cm sediment slices.
2.4. Benthic foraminifera
Foraminifera analyses were performed on 133samples representing 1-cm sediment slices and werecarried out at each centimetre or at intervals of 1 cm. The

Fig. 2. Location of core KSGX 40 in the Galicia Mud Deposit (adapted from Dias et al., 2002).
38 V. Martins et al. / Marine Micropaleontology 59 (2006) 35–55
samples were washed carefully over a set of sieves of 63and 1000 μm. The 1000-μm sieve was used to removelarge fragments of organic material. The N63-μmfractions were dried in an oven at 40 °C, weighed andused for foraminiferal analysis. A minimum number of300 benthic foraminifera have been picked, identified,and counted in each sample. For the determination offoraminifera we followed the taxonomy of Loeblich andTappan (1988) and Ellis and Messina Catalogue (1940–1988) and as well as references cited by Martins andGomes (2004). Very broken specimens whose identifi-cation was not possible were not considered in theestablishment of foraminifera assemblages. Foraminif-eral abundance (number per gram of bulk sediment) wascalculated counting shells from a known weighed sedi-ment split.
The Shannon–Wiener diversity was calculated toidentify changes in the structure of benthic forami-nifera assemblages: H=−∑ pi · ln pi, where pi is theproportion of each species (Shannon and Weaver,1999).
2.5. Statistic treatment of data
The percent value of the most abundant speciesformed the numerical basis for the cluster analysis. Toestablish foraminifera assemblages and sections withsimilar faunal composition, the R-mode (species vs.species) and Q-mode (sample vs. sample) cluster anal-ysis using Pearson correlation and the Ward's distancemethod (Everitt, 1993) were applied to 133 samples and23 of the most abundant species. Both multivariate
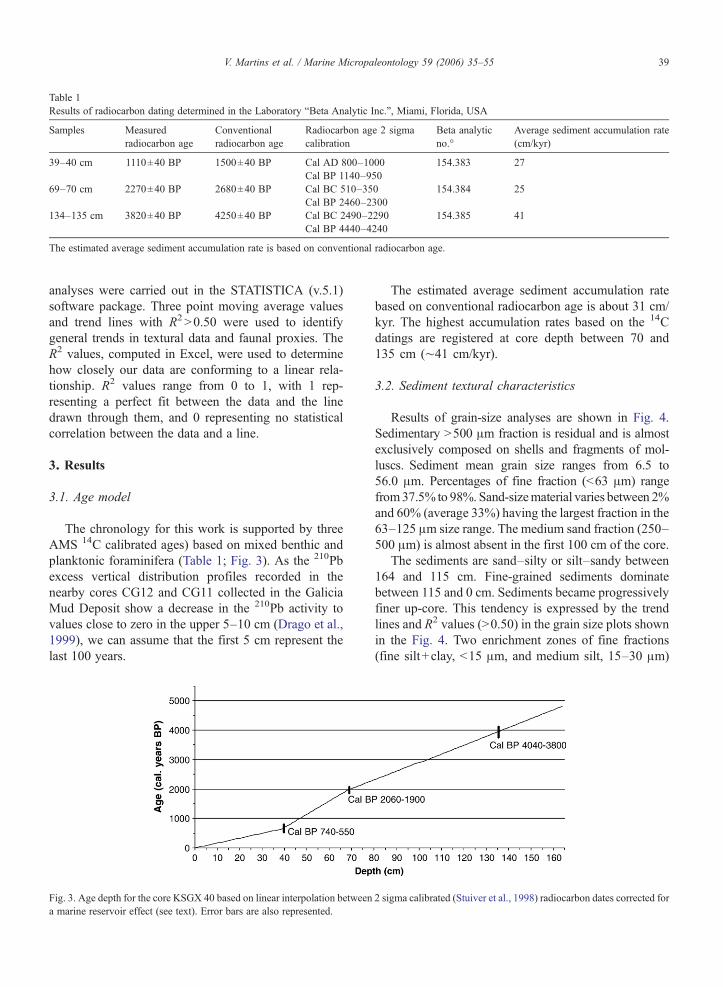
Table 1Results of radiocarbon dating determined in the Laboratory “Beta Analytic Inc.”, Miami, Florida, USA
Samples Measuredradiocarbon age
Conventionalradiocarbon age
Radiocarbon age 2 sigmacalibration
Beta analyticno.°
Average sediment accumulation rate(cm/kyr)
39–40 cm 1110±40 BP 1500±40 BP Cal AD 800–1000 154.383 27Cal BP 1140–950
69–70 cm 2270±40 BP 2680±40 BP Cal BC 510–350 154.384 25Cal BP 2460–2300
134–135 cm 3820±40 BP 4250±40 BP Cal BC 2490–2290 154.385 41Cal BP 4440–4240
The estimated average sediment accumulation rate is based on conventional radiocarbon age.
39V. Martins et al. / Marine Micropaleontology 59 (2006) 35–55
analyses were carried out in the STATISTICA (v.5.1)software package. Three point moving average valuesand trend lines with R2N0.50 were used to identifygeneral trends in textural data and faunal proxies. TheR2 values, computed in Excel, were used to determinehow closely our data are conforming to a linear rela-tionship. R2 values range from 0 to 1, with 1 rep-resenting a perfect fit between the data and the linedrawn through them, and 0 representing no statisticalcorrelation between the data and a line.
3. Results
3.1. Age model
The chronology for this work is supported by threeAMS 14C calibrated ages) based on mixed benthic andplanktonic foraminifera (Table 1; Fig. 3). As the 210Pbexcess vertical distribution profiles recorded in thenearby cores CG12 and CG11 collected in the GaliciaMud Deposit show a decrease in the 210Pb activity tovalues close to zero in the upper 5–10 cm (Drago et al.,1999), we can assume that the first 5 cm represent thelast 100 years.
Fig. 3. Age depth for the core KSGX 40 based on linear interpolation betweena marine reservoir effect (see text). Error bars are also represented.
The estimated average sediment accumulation ratebased on conventional radiocarbon age is about 31 cm/kyr. The highest accumulation rates based on the 14Cdatings are registered at core depth between 70 and135 cm (∼41 cm/kyr).
3.2. Sediment textural characteristics
Results of grain-size analyses are shown in Fig. 4.Sedimentary N500 μm fraction is residual and is almostexclusively composed on shells and fragments of mol-luscs. Sediment mean grain size ranges from 6.5 to56.0 μm. Percentages of fine fraction (b63 μm) rangefrom37.5% to 98%. Sand-sizematerial varies between 2%and 60% (average 33%) having the largest fraction in the63–125 μm size range. The medium sand fraction (250–500 μm) is almost absent in the first 100 cm of the core.
The sediments are sand–silty or silt–sandy between164 and 115 cm. Fine-grained sediments dominatebetween 115 and 0 cm. Sediments became progressivelyfiner up-core. This tendency is expressed by the trendlines and R2 values (N0.50) in the grain size plots shownin the Fig. 4. Two enrichment zones of fine fractions(fine silt+clay, b15 μm, and medium silt, 15–30 μm)
2 sigma calibrated (Stuiver et al., 1998) radiocarbon dates corrected for

Fig. 4. Logs showing textural data for core KSGX 40. Smoothed lines (solid black curves) between data (marks), the mean of the values (grey hatchedline) and some trend lines (solid inclined grey line) are also represented.
Fig. 5. In general the lower section of the core is characterized by higher number of benthic foraminifera (per gram of dry bulk sediment) and values ofthe Shannon–Wiener Index (H). Otherwise the total percentage of infaunal species and intermediate to deep infaunal species is lower in this section.The refereed data have a general inverse pattern in the upper section of the core.
40 V. Martins et al. / Marine Micropaleontology 59 (2006) 35–55

41V. Martins et al. / Marine Micropaleontology 59 (2006) 35–55
can be recognized between 80–50 and 30–0 cm. Thesesections represent two muddy periods, in which fine siltand clay fraction (b15 μm), medium silt fraction (15–30 μm), i.e., the finer fractions of the sediment com-position have a significant decrease while medium sandis almost absent.
3.3. Benthic foraminifera data — vertical profile andR-mode cluster analysis
The absolute abundance of benthic foraminifera(number of specimens per gram of bulk sediment)varied between 160 and 9600. Foraminiferal abundancedecrease progressively upward as sediment mean grainsize decreases (Figs. 4, 5). Highest abundances arefound between 164 and 95 cm in sandier facies. A totalof 354 taxa were recognized. Calcareous hyalineforaminifera species are much more diverse and
Fig. 6. Dendrogram resulting from a cluster analysis based on the relative abufor agglomeration the Ward's method.
abundant (96% to 100%) than porcelaneous (0% to3.2%), and agglutinated (0% to 1.5%) subgroups.
Bolivina/Brizalina spp. (23–67%), Cassidulina/Globocassidulina spp. (3–30%), Bulimina spp. (3–16%), Cibicides spp. (0.3–19%), Fursenkoina/Stain-forthia spp. (0–13%), and Nonionella spp. (0–6%) arethe most abundant taxa. Bolivina/Brizalina spp. (in-cluding mainly Brizalina spathulata, Bolivina ordi-naria, Bolivina pseudoplicata, Brizalina pacifica,Bolivina seminuda and Bolivina difformis) abundancesare higher in finer sediments. This distribution pattern isdetermined mainly by B. ordinaria and B. spathulata,the most abundant species. Cibicides spp. (includingmainly Cibicides ungerianus) are represented more incoarser sediments decreasing their percentage up-core.The relative abundance of Bulimina spp. (includingmainly Bulimina elongata/gibba and Bulimina margin-ata), Cassidulina/Globocassidulina spp. (including
ndance of the 23 most frequent taxa, using the Pearson correlation and

42 V. Martins et al. / Marine Micropaleontology 59 (2006) 35–55
mainly Cassidulina laevigata/C. carinata, Cassidulinaminuta, Cassidulina crassa and Globocassidulinasubglobosa) decrease in the middle core section. Higher
percentages of Fursenkoina/Stainforthia spp. [mainlyStainforthia fusiformis (b8.2%), Fursenkoina loeblichi(b7%), Stainforthia complanata (b3.6%)] and

43V. Martins et al. / Marine Micropaleontology 59 (2006) 35–55
Nonionella spp. [mainly Nonionella stella (b4.3%)] canbe observed in the middle core section in coincidencewith higher values of Bolivina/Brizalina spp.
Species diversity evaluated by Shannon–WienerIndex (H) markedly declines in the top 75 cm as infaunalspecies, and namely intermediate to deep infaunalspecies, increased accompanying the decrease ofsediment mean grain size. Infaunal species includesAmmonia beccarii, Bigenerina nodosaria, Bolivina/Brizalina spp., Bulimina spp., C. laevigata/C. carinata,Buliminella tenuata, Chilostomella spp., Fursenkoina/Stainforthia spp., Globobulimina spp., Haynesinadepressula, Hoeglundina elegans, Hyalinea balthica,Melonis spp., Nonion spp., Nonionoides pauperatus,Pseudobolivina fusiformis, Pullenia spp., Rectuvigerinaphlegeri, Sagrina subspinescens, Sphaeroidina bul-loides, Uvigerina spp., Valvulineria bradyana (e.g.,Murray, 1991; Corliss, 1991; Fontanier et al., 2002).Intermediate to deep infaunal species is a sub-group ofthe former including A. beccarii, Bulimina exilis (Rath-burn and Corliss, 1994), Chilostomella spp. (Mackensenand Douglas, 1989), Fursenkoina/Stainforthia spp.(Knudsen and Seidenkrantz, 1994), Bulimina aculeata,B. marginata, Globobulimina spp., H. balthica (Corliss,1991; Fontanier et al., 2002).
R-mode cluster analysis differentiates three mainassemblages represented in the dendrogram of the Fig. 6.The depth profile of these taxa is represented in Fig. 7.Most of the species included in cluster 1 show significantchanges in the section 45–0 cm. B. spathulata decreasesin the middle section of the core like C. laevigata/C.carinata. But C. laevigata/C. carinata has a bimodalabundance becoming one of most frequent species in theupper section of the core. B. marginata shows a verygradual upward increase (R2 =0.50). Otherwise, A.beccarii, H. balthica, Nonium fabum, Uvigerina pere-grina, V. bradyana and Cassidulina teretis increase theirfrequency, in general, suddenly and some timescyclically in the section 45–0 cm. The species of cluster2 increased their percentage during one or both muddyperiods. Textural changes from coarser sediments in corebase to finer sediments in middle and top sections wasaccompanied by the decrease of the species of cluster 3.B. elongata/gibba, Cribrononion gerthi and B. pseudo-plicata were included in this cluster since their
Fig. 7. Percentage of benthic foraminifera taxa (%) included in the cluster 1line) are represented in some plots. Bottom water temperatures and oceanic pthis cluster's species. Percentage of benthic foraminifera taxa (%) included in tvalues (grey hatched line) are represented. This cluster includes higher producbenthic foraminifera taxa (%) included in the cluster 3 of the dendrogram of tsome plots. The abundance of these species could be related with a high eneroxygen rich pore-waters in surficial sediments.
percentage decreased significantly in finer sediments,i.e., in sections 80–50 and 30–0 cm.
3.4. Q-mode cluster analysis
Results of Q-mode cluster analysis are represented inthe dendrogram of Fig. 8. Levels belonging to cluster 3are mainly represented between 164 and 77 cm. From 77to 44 cm predominate samples included in cluster 2.1and, between 30 and 2 cm, samples included in cluster2.2. There are two sections with samples belonging toseveral clusters: between 115–77 and 45–30 cm.
4. Discussion
4.1. Analysing benthic foraminifera clusters andassemblages composition (R-mode cluster analysis)
4.1.1. Species of cluster 1Species of cluster 1 can be related to a rich supply of
organic carbon probably resulting from oceanic produc-tivity. B. spathulata, C. laevigata/C. carinata, B.marginata/aculeata,N. fabum,H. balthica,U. peregrinaand V. bradyana are reported to be characteristic of theupwelling system off Portugal (Levy et al., 1995). B.spathulata is also a marker of upwelling off Namibia(Bremer, 1983).B. marginata are known to have positivecorrelation with nutrient contents (Donnici and Barbero,2002) so the high abundance of this species has beenconsidered a good marker of high productivity waters(Phleger and Soutar, 1973; Lutze and Coulbourn, 1984).B. aculeata is also frequent in areas with calmsedimentation and with a high flux of organic matter(Mackensen et al., 1990; Almogi-Labin et al., 2000). U.peregrina is an epifaunal to shallow infaunal species(Casford et al., 2003) that dominate also in eutrophicbiofacies (De Rijk et al., 2000), being commonlyassociated with higher productivity (e.g., Lutze andCoulbourn, 1984; Rathburn and Corliss, 1994; van derZwaan and Jorissen, 1991; Loubere, 1996).
Ammonia beccarii and Nonium fabum, which are un-common in the lower part of the core, display an abruptincrease pointing to a brief grain size coarser episode at45–30 cm. The increase of this species probably indicatesa rainy period around the period 0.9–0.5 ka cal BP, since
of the dendrogram of the Fig. 6. The mean of the values (grey hatchedroductivity could have been important factors ruling the abundance ofhe cluster 2 of the dendrogram of the Fig. 6. The mean of B. ordinaria'stivity and dysoxic and/or tolerant to periodic anoxia taxa. Percentage ofhe Fig. 6. The mean of the values (grey hatched line) are represented ingy environment and are indicative of lower Corg flux to the bottom and

Fig. 8. Dendrogram resulting from a Q-mode cluster analysis based on the relative abundance of the most frequent taxa in 133 variables (levels incentimetres of the core), using Pearson correlation and for agglomeration the Ward's method. Four main clusters were identified. At left, the tablerepresents core sequences included in each cluster/sub-cluster.
44 V. Martins et al. / Marine Micropaleontology 59 (2006) 35–55

45V. Martins et al. / Marine Micropaleontology 59 (2006) 35–55
A. beccarii is a euryhaline species occurring typically inshallow waters (e.g., Bandy, 1953; Cearreta, 1989; Alveand Murray, 1994; Almogi-Labin et al., 1995; Kim andKennett, 1998) andN. fabum is a quite common species inthe inner to middle shelf (Mendes et al., 2004) occurringuntil a depth of 80 m in surficial sediments of W IberianMargin (Levy et al., 1995; as Nonion commune). Thesepeaks in A. beccarii and N. fabum percentage suggestremobilization from shallower waters. A sheltered shelfassociated with downwelling and rainy conditions, mayhave promoted a higher remobilization in the inner andmiddle shelf and the transport of coarser sediments todeeper oceanic regions. On the NW Iberian shelf theseconditions are nowadays typical of winter, under domi-nant downwelling conditions, during which intense bot-tom nepheloid layers (BNL) can be observed due to riverborne particles supply and remobilization of mid-shelfmuddy sediments (Oliveira et al., 2002). These authorsconcluded that downwelling circulation probably inducestransport of shelf particles to deeper waters in the BNL.
This occurrence was coincident with the abrupt increaseof H. balthica that was followed by the increase of U.peregrina, V. bradyana, B. marginata, C. laevigata/ C.carinata and C. teretis.H. balthica indicate the presence ofcolder water masses on the continental shelf (Murray,1971, 1973) and has been found in Quaternary glacialsediments from the Northeastern Atlantic and Mediterra-nean Sea (Ross, 1984; Caralp, 1988). C. teretis is a rarespecies in superficial sediments of the NW Iberian Margin(Martins and Gomes, 2004). However high abundance ofthis species is typical of the modern shelf sediments in theGreenland, Norwegian, North and Barents Seas, with abottom water temperature of about 0° C (Mackensen andHald, 1988). C. teretis is also abundant in glacial UpperPleistocene sediments in Greenland (e.g., Feyling-Hanssenet al., 1983). So, the peaks of abundance ofH. balthica andC. teretis between 34 and 27 cm (around ∼0.5 ka cal BP)suggest the occurrence of a noticeably bottom water coolerevent. Others species like U. peregrina, V. bradyana, B.marginata and C. laevigata/C. carinata are frequentlydominant in colder bathyal waters (Murray, 1991). Thecyclical enhancements of these species in the upper coresection may indicate recurrent bottom water cooler eventswhich could have occurred since ∼0.5 ka cal BP on theIberian Margin.
Bottom water temperatures and oceanic productivitycould have been important factors ruling the abundanceof the cluster 1 group of species.
4.1.2. Species of cluster 2All the species/taxa of cluster 2 have been reported in
areas of high organic matter flux and low oxygen levels. B.
ordinaria, the dominant species of this group, increased itspercentage as far as sediments became finer reachinggreater percentages in the top 80 cm. This species domi-nates in environments with very depressed levels of oxy-gen, namely in methane hydrates environments (Aharon etal., 2001). Higher abundances of Fursenkoina andStainforthia are also indicative of high productivity/lowoxygen conditions (Leutenegger and Hansen, 1979). Infossil records, they can be found in pre- and post-sapropelassemblages and so they are considered to be tolerant tolow oxic and dysoxic conditions (Jorissen, 1999). S.fusiformis and Stainforthia feylingi are frequently groupedunder the name S. fusiformis (orFursenkoina fusiformis) inthe North Sea, where they dominate in dysoxic sedimentsof the Norwegian fjords (Alve, 1994, 1995). They areopportunists typical of organically enriched sublittoralenvironments where they are sometimes extremely abun-dant (Alve, 1994) in infaunal microhabitats (Moodley,1990;Alve andBernhard, 1995). Considerable populationsof both S. fusiformis (Alve, 1994) and N. stella also havebeen recovered from sulphide habitats (Bernhard andBowser, 1999). Nonionella species show a positivecorrelation with nutrient contents (Donnici and Barbero,2002). They are opportunistic, epifaunal species known toinvade formerly anoxic sea bottoms (e.g., Barmawidjaja etal., 1992; Hohenegger et al., 1993). According to Kitazatoet al. (2000), B. pacifica is a phytodetrital species, with alight-green cytoplasm, which may indicate that they feedupon fresh phytodetritus containing chlorophyla. Thisspecies constructs small and fragile tests that may reflectrapid reproduction and growth in the spring phytodetrituslayer. This species also shows some affinity to degradedorganic matter in the sediment (Altenbach et al., 2003). B.tenuata was also classified as opportunist due to its re-production rate and the rapid exploitation of ephemeralresources in S. Pedro Basin, California (Silva et al.,1996). This species inhabits dysoxic marine sedimentsalong the California margin (Quinterno and Gardner,1987; Bernhard, 1986).
So, cluster 2 includes higher productivity anddysoxic and/or tolerant to periodic anoxia taxa. Thisgroup is more represented in both muddy sections (80–50 and 30–0 cm), which may indicate two higherproductivity periods, which could have led to depressedlevels of oxygen, as suggested by geochemical dataanalyzed byMartins et al. (2006). It is known that higheramounts of food to the benthos fauna can lead to thedevelopment of aerobic organisms consuming oxygenthat can reduce the oxygenation in the sediment pore-water and some times in the water–sediment interface.The higher flux of Corg and the deposition of finersediments can lead to very depressed levels of oxygen

46 V. Martins et al. / Marine Micropaleontology 59 (2006) 35–55
and, perhaps, to anoxia in the sediments, due thedifficulty of oxygen renewal in pore water.
4.1.3. Species of cluster 3Cluster 3 includes species that prefer elevated epiben-
thic habitats under the impact of near-bottom currents,C.ungerianus (like others Cibicides species), Gavelinopsispraegeri, C. gerthi (as Elphidium) (Murray, 1991;Barmawidjaja et al., 1995). Cibicides species live inelevated habitats attached to firm substrates and areindicative of stronger bottom currents (Rasmussen et al.,2002). All these species have a significant positive cor-relation with sandier facies. C. ungerianus and G. prae-geri are known to be suspension feeders (Murray, 1991)and are favoured by coarse-grained sediments with lowconcentrations of organic carbon due the presence ofenergetic bottom currents (Barmawidjaja et al., 1995).
Another group of benthic foraminifera included incluster 3 which has been correlated with coarser sedi-ments and/or lower organic carbon flux is C. minuta, G.subglobosa and C. crassa. C. minuta is a dominantspecies on the middle to lower shelf (82–451 m waterdepth) of the Gulf of Guinea, in coarse-grained andorganic rich sediments (sed. org. C 0.1–3.5 wt.%) (Al-tenbach et al., 2003). G. subglobosa is also an op-portunist, suspended feeder species (Linke and Lutze,1993; Altenbach, 1992). C. crassa is an epifaunal orshallow infaunal species (Dulk et al., 2000) and is as-sociated with lower productivity (Fariduddin and Lou-bere, 1997). The standing stock of this species is reducedin areas with higher Corg content (Altenbach et al., 2003).
This cluster also includes B. pseudoplicata (b20%), adominant species with C. ungerianus, as well as B.elongata/gibba (b12%) and B. difformis (b6%). Boli-vinids and buliminids occur in a range of oceanic set-tings but when they occur in high abundance they mayindicate high nutrient levels and/or dysoxia (Mackensenet al., 1990; Murray, 1991). However in this core thespecies of both these groups have a distribution patterndifferentiated. The relative abundance, for instance, ofB. spathulata, B. pseudoplicata, B. difformis and B.elongata/gibba slightly decrease in the muddy sections(80–50 cm, and/or 30–0 cm) where others bolivinidsand buliminids, like B. ordinaria, B. pacifica and B.marginata, increase. This suggests a differentiated be-haviour of the species of both these groups to theenvironmental stress. According to Donnici and Barbero(2002), B. elongata for instance shows a positive cor-relation with nutrient contents but a negative one withsubstrate grain size.
Others epifaunal species with a planoconvex mor-phology, typical for the epiphytic microhabitat, which
have been excluded from cluster analysis due to theirlow abundance, are also present mainly in coarser sedi-ments: Hanzawaia nitidula, Lobatula lobatula, Lamar-ckina haliotidea, Lepidodeuterammina ochracea,Paumotua terebra, Planorbulina mediterranensis andRemaneica helgolandica. Others species with the samedistribution pattern can also be added to this group, likeTrifarina angulosa that prefer elevated epibenthic habi-tats under the impact of near-bottom currents (Jorissen,1987; Altenbach, 1992; Linke and Lutze, 1993; Stigteret al., 1998), Textularia spp., keeled Elphidium spp. andQuinqueloculina spp. (Altenbach et al., 2003; Murray,1991) which prefer coarser sediments and high speed ofbottom currents. In the Elphidium group, species such asE. macellum var. aculeatum, E. crispum, E. jenseni (asE. fichtellianum) have also an epiphytic style of life(Jorissen, 1987) so they are favoured by active bottomcurrents and limited by the lower oxygen content (Lan-ger et al., 1989; Saffert and Thomas, 1998). Globocas-sidulina rossensis (as C. obtusa) also has a similarprofile along the core to these species. It has beenrecorded together with L. lobatula (as Cibicideslobatulus) and T. angulosa in the warm Atlantic surfacewater along the shelf and upper slope of Norway and incoarse sediments due to winnowing by bottom currents(Sejrup et al., 1981).
The higher abundance of all these taxa in the section164–70 cm, in which sediments are coarser, could berelated with a high energy environment and may indi-cate the presence of mixed waters on the shelf and areindicative of lower Corg flux to the bottom and oxygenrich pore-waters in surficial sediments.
4.2. Analysing Q-mode cluster analysis
Q-mode cluster analysis differentiates four mainclusters. In the cluster 2 can be considered two otherssub-clusters (2.1 and 2.2). Whereas the species of the R-mode cluster 3 are related to the Q-mode cluster 3, mainlyrepresented between 164 and 77 cm, the R-mode cluster 2are mainly related to the Q-mode cluster 2.1corresponding to the section 77–44 cm. Otherwise theR-mode cluster 2 are mainly related to the Q-mode cluster2.2 including samples between 30 and 2 cm. So weinterpret the section between 164 and 77 cm, asrepresenting a period in which a sheltered shelf withmixedwaters prevailed.Although between 115 and 77 cmshort periods of prevalence of mixed waters and low fluxof Corg to the bottom seems to be intercalated with shortperiods of low hydrodynamic bottom waters conditionsand high input of Corg to the bottom. This period can beinterpreted as a transition to a longer period characterized

47V. Martins et al. / Marine Micropaleontology 59 (2006) 35–55
by high oceanic productivity but low hydrodynamic bot-tom waters over the core site registered in the section 77–44 cm, which evolved to another with similar character-istics (signed by textural and faunal data) between 30 and2 cm.
4.3. Looking for long-term variability in organic matterflux to the bottom
The previous findings that suggested the use of‘phytodetritus species’ as proxies for pulsed inputs oforganic matter to the sea floor (e.g., Smart et al., 1994;Thomas et al., 1995; Thomas and Gooday, 1996) encou-raged us to look for evidence of long-term variability ofprimary production in the NW Iberian Margin during thelast 4.8 ka cal BP, based on benthic foraminifera re-cognized to be good markers of Corg flux and currentvelocity. The percentage of these groups and some texturaldata were plotted against calibrated age BP (Fig. 9).
The group of high velocity currents and low flux ofCorg marker includes the total percentage of epifaunalspecies some of them known to be suspension feeders,like Cibicides spp., G. praegeri, C. gerthi, C. minuta, C.crassa, G. subglobosa, H. nitidula, G. rossensis, L.lobatula, L. haliotidea, L. ochracea, P. terebra, P.
Fig. 9. Sedimentological data and benthic foraminifera proxies of low andoccurred two most important periods of deposition of finer and richer in organ(marks), the mean of the values (grey hatched line) and trend lines (solid inc
mediterranensis, R. helgolandica, T. angulosa, Elphi-dium spp., Textularia spp. and Quinqueloculina spp., aswas discussed in 4.1.3. The percentage of this group ismarked by a decreasing trend line (with R2 =0.75) fromthe base, where they achieve the maximum value of45%, until the core top, where a minimum value of 2%is registered, following the pattern of sediments finefractions content.
The group of high flux of Corg marker includes thetotal percentage of the taxa integrated in R-mode cluster2, i.e., B. ordinaria, B. pacifica, B. tenuata, Nonionellaspp., and Fursenkoina/Stainforthia spp., as well as somespecies of R-mode cluster 1, like B. marginata/aculeata,B. spathulata, U. peregrina and V. bradyana as wasanalyzed in 4.1.2 and 4.1.1. This group has a generalinverse trend profile of the one of lower productivityproxy, and has a negative correlation with the sedimentmean grain size. Greater values of high productivityproxy are achieved after ∼2.2 ka cal BP.
The decline up core of lower productivity epifaunalspecies, as well as sedimentary sand fractions content,was followed by the marked increase of high productivity(and hypoxia-tolerant species) and sedimentary finefractions content. Benthic foraminifera proxies supportthe fact that the higher flux in organic carbon was
high productivity. These data suggest that in the last ∼4.8 ka cal BPic matter sediments. Smoothed lines (solid black curves) between datalined grey line) are also represented.

48 V. Martins et al. / Marine Micropaleontology 59 (2006) 35–55
coincident with the deposition of finer sediments. Thehigher deposition of Corg occurred during the both muddyevents, i.e., between 2.2 and 1.2 ka cal BP and after 0.5 kacal BP. This feature agrees with Jouanneau et al. (2002)observations in several cores sampled in the NW IberianShelf (Galicia region), in which maximum concentrationsof Corg are found in the finest facies.
4.4. Consequences of high deposition of Corg in benthicforaminifera assemblages
During both the muddy events the higher deposition ofCorg may have led to depressed levels of oxygen beneaththe interface water–sediments and to early diageneticprocesses of several redox-sensitive elements, like Mn,Fe, Cu, Ni, Cr, Co, Zn, Ni and Pb as observed by Martinset al. (2006). These conditions (in the upper core section)were marked by the decrease in density and diversity andby the increase in dominance (a small number of speciesaccount for much of the assemblage) in benthicforaminifera assemblages and by the clear increase ininfaunal species, namely intermediate to deep infaunalspecies. According to Fontanier et al. (2003) the deepmicrohabitat of several infaunal taxa may be caused bytheir low competitiveness in the more attractive sedimentsurface niches, which are much richer in easily metabo-lisable organic matter. According to these authors, theoxygenation of the pore waters does not have a directcontrol on the vertical distribution of most foraminiferaltaxa. Most species are able to adapt to severe dysoxia oreven anoxia, since sufficient high quality food particlesare available. However, the continuation of anoxia forweeks or months, in combination with H2S, prohibits thesustenance of large foraminiferal populations (Bernhardand SenGupta, 1999), since it is possible that foraminiferabecame dormant during exposure to adverse conditionssuch as oxygen depletion (Bernhard and Alve, 1996)although some species are able to survive in microxia andanoxia environments for considerable periods of time(Bernhard, 1993). So the upward decline in faunal densityand changing in population structure may be due toenvironmental stress settled on by longer periods ofanoxic conditions, in combination with H2S, in pore wa-ters near the sediments surface. These conditions seem tobe more intense during the deposition of both muddydeposits (Martins et al., 2005, 2006).
4.5. Sources of Corg to the Galicia Mud Deposit
The main patterns of phytoplankton biomass in up-welling systems are related to water-column stratifica-tion, nutrient availability, and the intensity and
persistence of upwelling conditions. The introductionof nutrients from the nearby continental areas throughriver runoff is also a process that can increase oceanicproductivity.
At present the upwelling season occurs from March–April to September–October (Wooster et al., 1976;Fiúza et al., 1982; Fiúza, 1983), when the weakening ofthe Greenland Low Pressure Centre and strengtheningand northward displacement of the Azores HighPressure Centre promotes northerly winds. During therest of the year the winds are predominantly from thesouth and south–west due to the intensification of theGreenland Low Pressure Centre, thus favouring down-welling. The upwelling enhances the primary produc-tion in Galician shelf areas (Tenore et al., 1995) andinside the Rias Baixas (Hanson et al., 1986; Tilstone etal., 1999) as well as the accumulation of organic carbonin surface sedimentary layers (Álvarez-Salgado et al.,2001b). The Rias Baixas and continental shelf off Ga-licia are high productivity regions (Blanton et al., 1984;López-Jamar et al., 1992), mainly due to upwelling(Prego, 1993). Bode et al. (1996) estimated the totalannual primary production in the Galicia area in 220 Cm−2 yr−1. Off the Rias Baixas (3400 km2), the resultantexport per unit of shelf area is roughly 100 g C m−2 yr−1
(Álvarez-Salgado et al., 2001a) which represents asmuch as 45% of the total annual primary production inthe study area (Bode et al., 1996). So the outwelling oforganic detritus from the Rias Baixas, which is typicaland very intense during the upwelling season, bring intothe continental shelf large quantities of nutrients(Álvarez-Salgado et al., 2001b). The higher flux oforganic matter in this zone can result, not only fromlocal oceanic biological production, but also fromrelatively fresh organic carbon exported from the RiasBaixas biological productivity (Estrada, 1984; Bode etal., 1994; Álvarez-Salgado et al., 1997). An importantupwelling centre located at Cape Finisterre (North ofthis region), where marine organic matter production ishigher (e.g., Haynes et al., 1993; Castro et al., 2000),may account with another supplementary source oforganic matter to the southern areas, during the periodsof intensified upwelling. Since, NWwind intensificationforces equatorward displacement of the surface wind-mixed layer with a consequent redistribution of organicmatter with north sources. The upwelling centre, offCape Finisterre (NW Iberian Upwelling System) is amajor route for the export of primary production fromshelf waters in the Galicia region (McClain et al., 1986).
The increase of infaunal species, namely intermedi-ate to deeper infauna, in the upper core section, after∼2.2 ka cal BP, and particularly in both muddy events,

49V. Martins et al. / Marine Micropaleontology 59 (2006) 35–55
should be caused not only by high quality food but alsoby more degraded organic matter. It is known thatforaminiferal faunas respond to rapid changes in theorganic flux or bottom water oxygenation by a shift oftheir microhabitat structure (Barmawidjaja et al., 1992;Ohga and Kitazato, 1997). While some species ofbenthic foraminifera can feed on a wide range of foodparticles (Loubere and Fariduddin, 1999) and cantolerate lower food quality, feeding on material withlow nutritional value, others have preference to highquality food, labile and easily metabolized organicmatter (produced in the oceanic water column), at thesurface sediment (Caralp, 1984). Labile organic matteris concentrated at the sediment surface and will becomeincreasingly refractory in deeper sediment layers. Forthis reason, interspecies differences in vertical distri-bution may also reflect differences in nutritive be-haviour (Jorissen et al., 1995). Jorissen et al. (1998) andFontanier et al. (2003) suggested that deep infaunalforaminifera can feed on the decay products of lowquality sedimentary organic matter, made available bybacterial anaerobic degradation or on the bacterialstocks themselves around important redox boundaries.
So the benthic foraminifera proxies allow us tosuppose that the organic carbon flux to the Galicia MudDeposit was provided not only by the oceanicproductivity (fresh organic matter) but also from otherssources, like by detritus outwelling (relatively freshorganic matter) from the Lower Rias recognized to behigh productivity systems or from other shelf sources.Terrestrial refractory, organic carbon released by coastalerosion and transported to the studied area probably isnot used so much by bacterial and foraminiferal faunas.So we can suppose that in the upper core section, after∼2.2 ka cal BP, and particularly in both muddy events,in which the intermediate to deep infauna percentageincreased, the contribution in Corg provided from di-versified sources should be higher. However all thesemain sources of Corg used as food by benthic fauna maybe directly or indirectly related to the increasing in theupwelling activity.
Nevertheless mainly in the last century (probablyregistered in the upper 5/10 cm) eutrophication may bedue also to anthropogenic activity. The human density incoastal areas increased in the last century, leading to anincrease of Corg in coastal systems. So whereas the ben-thic foraminifera high productivity proxies may signal anincrease in upwelling intensity during the Little Ice Age,in the last century, the signal of high oceanic productivityshould be masked by eueutrophication induced byhuman activities in coastal areas of this region. So wecan suppose that the upwelling activity may be higher in
the last 2.2 ka cal BP, but mainly between 2.2 and 1–2 kacal BP and 0.5–0.1 ka cal BP.
4.6. Evidence of water column stratification in the last4.8 ka cal BP
Scott et al. (2003), Scourse et al. (2002) and Evans etal. (2002) recently published studies of Holocene sea-sonal stratification: the firsts two in the Celtic Sea andthe last in North Sea, northeast England (NW EuropeanContinental Shelf). In these studies similar benthic fo-raminiferal assemblages variations were described. Mo-dern oceanographic conditions in these both regionsresult in the development of seasonally stratified watersduring the annual heating cycle. This process occurs inthe central to northern North Sea region resulting inlarge areas of stratified water. In this region frontalsystems development determine a high surface produc-tivity during the transition between stratified and mixedwaters.
According to these authors in the Celtic and NorthSeas, the mixed water assemblage is characterised byC. lobatulus, Textularia bocki, Ammonia batavus (A.beccarii, in this work) and Quinqueloculina seminulum.In the studied core the most important species related tomixed water conditions are C. ungerianus and G.praegeri, which are mainly represented on the corebase decreasing upward through section 164–77 cm andbeing almost absent between 77 and 0 cm (Fig. 7). Thesespecies, like others belonging to Cibicides, are known tobe related to conditions of full oxygenation and rela-tively high velocity flows. They are commonly associa-ted with sands or muddy sands in areas of high currentactivity which are, by definition, mobile (Scourse et al.,2002). G. praegeri is another species actually confinedto mixed waters. The distribution pattern of C.ungerianus and G. praegeri among others species (in-cluded in the cluster 3) also suggests a decreasing inmixed water conditions at the core site since ∼4.8 ka calBP. These higher hydrodynamical conditions, favour-able to a coarser detrital sedimentation, arrested thesettling of organic matter of the overlying productivityand/or of lateral transport to the sea floor, which alsoseems to have been lower. As a result sediments pore-waters remained more oxic (Martins et al., 2005, 2006).
These authors also identified several species whichcan be used as key indicators of stratified conditionswhich include Bolivina group, B. marginata, Nonionellaturgida and S. fusiformis, among others species (Fig. 7).In the core KSGX 40 the percentage of these taxa ishigher in the top 77 cm, i.e., after ∼2.2 ka cal BP. TheBolivina/Brizalina group is predominant in this section.

50 V. Martins et al. / Marine Micropaleontology 59 (2006) 35–55
B. marginata exhibits a progressive increase from thebase to this core top indicating also the evolution to amore seasonally stratified water column in the lateperiod, after ∼0.5 ka cal BP, represented in the top 30cm. S. fusiformis is a species whose abundance is ingeneral very low at mixed water sites (Moodley, 1990).This species has been also identified as a frontalindicator in the Celtic Sea (Scourse et al., 2002). Thepresence of this species in the upper core section canalso suggest that oceanic fronts started to develop at, orclose to, the core site since that time. Nowadays,convergent fronts located over 100 m isobath candevelop during the transition between the upwelling anddownwelling regime, off the W coast of Galicia, due theinteraction between the slope poleward flow and thesurface seaward flow from the Rias Baixas (Álvarez-Salgado et al., 2001a, 2003; Huthnance et al., 2002;Nogueira et al., 2003). This current confines the waterfrom the Rias Baixas on the continental shelf, whoseoutwelling is typical and very intense during theupwelling season (Teixeira et al., 2003) and candiminish the strength of shelf–ocean exchanges pro-cesses (Alvarez-Salgado et al., 2003; Nogueira et al.,2003). This situation should favour the deposition offiner sediments richer in organic matter on the outershelf.
Faunal and sedimentological evolution suggest theoccurrence of a period characterized by an oceanogra-phic regime favourable to the establishment of mixedwaters conditions between ∼4.8 and 2.2 ka cal BP andmore stratified shelf waters conditions since ∼2.2 ka calBP. Although, between∼3 and 2.2 ka cal BP, the coarsersedimentary fractions decline up-core compensated bythe progressive increase in finer fractions (b30 μm).This sedimentological change coincided with cyclicalreductions in Cibicides abundance and by others lowproductivity taxa, but was compensated by symmetricalincreases in high productivity fauna. This period isinterpreted as a transition to the muddy event charac-terized by the abrupt increase in fine fractions (b30 μm)that took place between ∼2.2 and ∼1.2 ka cal BP.Faunal composition in this muddy event is characterizedby the significant decrease of Cibicides and Gavelinop-sis and by the increase of B. ordinaria, Fursenkoina/Stainforthia, B. pacifica, Nonionella spp. and B.tenuata. During this period there may have been pre-vailed strong stratified water column conditions, sincesome of these taxa are indicators of thermal stratificationof water column (according to Scourse et al., 2002;Evans et al., 2002). This period may be contemporane-ous with a higher Corg flux to the marine benthic envi-ronment, probably in the sequence of upwelling
intensification, as was discussed. This period evolvedto another, between ∼1.2 and 0.5 ka cal BP, character-ized by the slight increase of sediment mean grain size,mainly due to the increase of coarse silt. This texturalchange was accompanied by a slight increase in benthicforaminifera abundance and diversity, and by a smalldecrease in infaunal percentage and in values of highproductivity proxy. Sedimentological and faunal char-acteristics suggest that this period should correspond toa less well-stratified shelf sea. The enhanced speed ofbottom currents should result in a slight decrease inorganic matter flux to the sea floor, which may result ina decrease in high productivity fauna. This faunal de-crease may also testify a less productive period probablycorrelated with a small decline in upwelling intensityand frequency.
The sediment fine fraction increased again in the top30 cm (since 500 yr cal BP). This change is characterizedby low benthic foraminifera abundance and diversity andhigh infaunal percentages, namely in intermediate todeep infaunal species. In this period the peaks ofFursenkoina/Stainforthia, composed mainly on S. fusi-formis, a species indicative of stratified water column(according to Scourse et al., 2002; Evans et al., 2002), arerecurrent. Although B. marginata, another species indi-cative of fully stratified water (according to the sameauthors), has augmented their relative abundance, as wellas B. ordinaria. So, sedimentological and faunal charac-teristics suggest the occurrence of another period ofmajor stratified conditions also associated with a highinput in Corg to the bottom.
4.7. Factors determining such oceanographic conditions
Nowadays the water column on the NW IberianMargin is generally mixed in winter and stratified insummer, with phytoplankton blooms occurring in thetransition between mixing and stratification periods(Varela et al., 2003). Stratification in the water columnis mainly controlled by solar heating in summer but canalso occur in winter (the season in which rain is moreintense) due to river discharge (Kudela et al., 2005).Duringwinter conditions the shelf is nearly homogeneousexcept for surface stratification due to riverine outflow(Huthnance et al., 2002). Stratification starts intermittent-ly from late March until early June, becomes shallowerthrough June, and deepens by September (Huthnance etal., 2001). The degree of thermal stratification evolvesfrom the weakest stratification in spring to the strongest inlate summer–early autumn (Kudela et al., 2005).Although on the NW Iberian Margin intermittent up-welling events from spring to autumn, especially

51V. Martins et al. / Marine Micropaleontology 59 (2006) 35–55
important during summer, interrupt the stratification orraise the thermocline, injecting nutrients and allowingsummer phytoplankton blooms (Varela et al., 2003 andreferences herein). Maximum values of chlorophyllusually occur in spring and summer for the Iberian sys-tems, although high chlorophyll values may also be re-corded at other times of year (Kudela et al., 2005).
Textural data agree with a prevalence of a spring–summer regime on the NW Iberian Margin, during theperiod registered in the upper section of the core butmainly during both the muddy events. On the other handthe prevalence of a winter regime associated with heavierrains and to downwelling conditions under SW windsmay benefit the transport and deposition of coarsersediments in deeper areas of the shelf. For the depositionof these events a lower velocity of bottomoceanic currentsis necessary which could be driven by the prevalence of aspring–summer regime and/or by sea-level rise influenc-ing the development convergent fronts located near theactual 100 m isobath. The development of these frontsmay also have eased the deposition of finer sedimentsricher in Corg during these events. Evans et al. (2002) alsoobserved that tidally stratified water is largely controlledby water depth, which would have been forced by post-glacial sea-level rise. After 4000–5000 yr BP the rates ofsea level rise were strongly attenuated (Bao et al., 1999),but our data also suggest that the influence of small sealevel oscillations during the both high productivity events,may be influenced the deposition of suchmuddy intervals.
These two main intervals (∼2.2–1.2 and∼0.5–0.1 kacal BP) occurred during two recent colder periods of theSub-Atlantic chronozone. The first stratification periodtook place after the transition between the Sub-Borealand Sub-Atlantic climates, probably during the DarkAges. The second one prevailed during the Little IceAge. According to Bond et al. (1992), there are a goodcorrelation between colder periods and stronger north-erly winds in the Nordic Seas. Otherwise, the maindriving force in upwelling systems is wind stress (Pitcheret al., 1998). In the NW Iberian Margin the upwellingintensity is related to the strengthening of northerlywinds. The last higher productivity period was precededaround 0.5 ka cal BP by abrupt peaks of benthic fora-minifera species related to colder bottom waters. Thistransitional interval, between Medieval Warm Periodand Little Ice Age, marked the evolution to cyclicalbottom waters cooling and progressively less oxygenat-ed sediments.
Data analysed in this work allowed the identificationin the NW Iberian Margin of two main periods in whichprevailed a stratified water column interpolated withlonger events of intensified upwelling favouring the
deposition of finer sediments richer in organic matter, inthe last 4.8 ka cal BP. This model is very similar to thatpresented by Evans et al. (2002) for the shelf sea of-fshore northeast England. This corroborates the relationof our results with the climatic influence. Insolation,wind stress and oceanic conditions may have a deter-minant influence in the analysed results.
5. Conclusions
The distribution pattern of textural and faunal datapresented in this work points to a general relationshipbetween fine-grained sediments and high nutrient contentduring the last∼4.8 ka cal BP. The sea-floor environmentsignificantly altered over this period due to differentoceanographic conditions probably under the influence ofclimatic oscillations and small sea level oscillations,which determined changes in the water column stratifi-cation and in the upwelling pattern, as well as in currentvelocities and in sediment supply to the Galicia MudDeposit. The benthic regime on the outer shelf off the Riade Vigo changed from a high hydrodynamic and moremesotrophic environment at 4.8 ka cal BP to a calmer andmore eutrophic environment after ∼2.2 ka cal BP. Twoperiods of more stratified conditions also could havetaken place: the first between ∼2.2 and 1.2 ka cal BP andthe second since ∼0.5 ka cal BP coinciding with twomuddy events. During the both muddy periods, the higherflux in organic matter to the bottom could be coupled withtwo long-term water column stratification and upwellingdominated periods. Whereas the water column stratifica-tion is mainly controlled by solar heating, in summer theupwelling increase is caused by the intensification ofnortherly winds, which in general are strengthened duringthe colder climate events. Both periods characterized byhigher flux in organic matter to the bottom may beattributed to climatic/oceanographic induced changesduring the Sub-Atlantic climate, namely throughout theDark Ages and the Little Ice Age, during which shouldhave occurred some contrasting climatic conditions. Asconvergent fronts located near the actual 100 m isobathcan also promote the deposition of finer sediments richerin organic matter, it can be supposed that small sea levelsoscillations should have influenced the position of theseoceanic fronts and the water column stratification in thestudied site. Eutrophication since the end of Little IceAge might be also influenced by human influence.
Acknowledgements
Wewould like to thank the editor AndreasMackensenand the reviewers Joachim Schönfeld and particularly

52 V. Martins et al. / Marine Micropaleontology 59 (2006) 35–55
John Murray for providing constructive and veryimportant suggestions to the discussion of data analysedin this manuscript and for helping with language issues.We are grateful to José Sebastián Carrión, of theDepartment of Plant Biology, Faculty of Biology,University of Murcia, Spain for their valuable commentsto the manuscript. The authors also wish to express theirthanks to all people involved in the Mission GAMINEX(8/07/1998–19/07/1998) of the oceanographic cruiseNO CÔTES DE LAMANCHE – in which was collectedthe OMEX (Ocean Margin Exchange Project) coreKSGX 40.
References
Abrantes, F., Moita, M.T., 1999. Water column and recent sedi-ment data on diatoms and coccolithophorids, off Portugal, con-firm sediment record of upwelling events. Oceanol. Acta 22,319–336.
Aharon, P., Hackworth, M., Platon, E., Wheeler, C., Sen Gupta, B.,2001. Isotope records of recent benthic foraminifera from hydrate-bearing sediments: methane-hydrate dissociation effects. GSAAnnual Meeting, Paper n.°, pp. 66–70.
Almogi-Labin, A., Siman-Tov, R., Rosenfeld, A., Debard, E., 1995.Occurrence and distribution of the foraminifer Ammonia beccariitepida (Cushman) in water bodies, recent and quaternary, of theDead Sea Rift, Israel. Mar. Micropaleontol. 6, 153–159.
Almogi-Labin, A., Schmiedl, G., Hemleben, C., Siman-Tov, R., Segl,M., Meischner, D., 2000. The influence of the NE winter monsoonon productivity changes in the Gulf of Aden, NW Arabian Sea,during the last 530 ka as recorded by foraminifera. Mar.Micropaleontol. 40, 295–319.
Altenbach, A.V., 1992. Short term processes and patterns in theforaminiferal response to organic flux rates. Mar. Micropaleontol.19, 119–129.
Altenbach, A., Sarnthein, M., 1989. Productivity record in benthicforaminifera. In: Berger, W.H., Smetacek, V.S., Wefer, G. (Eds.),Productivity of the Ocean: Present and Past. Wiley, pp. 255–269.
Altenbach, A.V., Lutze, G.F., Schiebel, R., Schönfeld, J., 2003. Impactof interrelated and interdependent ecological controls on benthicforaminifera: an example from the Gulf of Guinea. Palaeogeogr.Palaeoclimatol. Palaeoecol. 197, 213–238.
Álvarez-Salgado, X.A., Castro, C.G., Pérez, F.F., Fraga, F., 1997.Nutrient mineralization patterns in shelf waters of the WesternIberian upwelling. Cont. Shelf Res. 17, 1247–1270.
Álvarez-Salgado, X.A., Doval, M.D., Borges, A.V., Joint, I.,Frankignoulle, M., Woodward, E.M.S., Figueiras, F.G., 2001a.Off-shelf fluxes of labile materials by an upwelling filament in theNW Iberian Upwelling System. Prog. Oceanogr. 51, 321–337.
Álvarez-Salgado, X.A., Gago, X.A.J., Míguez, B.M., Pérez, F.F.,2001b. Net ecosystem production of dissolved organic carbon in acoastal upwelling system: the Ría de Vigo, Iberian Margin of theNorth Atlantic. Limnol. Oceanogr. 46, 135–147.
Alvarez-Salgado, X.A., Figueiras, F.G., Pérez, F.F., Groom, S.,Nogueira, E., Borges, A.V., Chou, L., Castro, C.G., Moncoiffé, G.,Ríos, A.F., Miller, A.E.J., Frankignoulle, M., Savidge, G., Wollast,R., 2003. The Portugal coastal counter current off NW Spain: newinsights on its biogeochemical variability. Prog. Oceanogr. 56,281–321.
Alve, E., 1994. Opportunistic features of the foraminifer Stainforthiafusiformis Williamson: evidence from Frierfjord, Norway. J.Micropalaeontol. 13, 24.
Alve, E., 1995. Benthic foraminiferal distribution and recolonizationof formerly anoxic environments in Drammensfjord, southernNorway. Mar. Micropaleontol. 25, 169–186.
Alve, E., Bernhard, J.M., 1995. Vertical migratory response of benthicforaminifera to controlled oxygen concentrations in an experi-mental mesocosm. Mar. Ecol., Progr. Ser. 116, 137–151.
Alve, E., Murray, J.W., 1994. Ecology and taphonomy of benthicforaminifera in a temperate mesotidal inlet. J. Foraminiferal Res.24 (1), 18–27.
Ambar, I., Fiúza, A.F.G., 1994. Some features of the Portugal currentsystem: a poleward slope undercurrent, an upwelling related summersouthward flow and an autumn–winter poleward coastal surfacecurrent. In: Katsaros, K.B., Fiúza, A.F.G., Ambar, I. (Eds.),Proceedings of the Second International Conference on Air–SeaInteraction and on Meteorology and Oceanography of the CoastalZone.AmericanMeteorological Society, Boston, USA, pp. 286–287.
Bandy, O.L., 1953. Ecology and paleoecology of some Californiaforaminifera. Part I. The frequency distribution of recentforaminifera off California. J. Paleontol. 27 (2), 161–182.
Bao, R., Freitas, M.C., Andrade, C., 1999. Separating eustatic fromlocal effects: a late-Holocene record of coastal change in AlbufeiraLagoon, Portugal. Holocene 9, 341–352.
Barmawidjaja, D., Jorissen, F.J., Puskaric, S., van der Zwaan, G.J.,1992. Microhabitat selection by benthic foraminifera in thenorthern Adriatic Sea. J. Foraminiferal Res. 22, 297–317.
Barmawidjaja, D.M., van der Zwaan, G.J., Jorissen, F.J., Puskaric, S.,1995. 150 years of eutrophication in the northern Adriatic Sea:evidence from a benthic foraminiferal record. Mar. Geol. 122,367–384.
Bernhard, J.M., 1986. Characteristic assemblages and morphologies ofbenthic foraminifera from anoxic, organic-rich deposits: Jurassicthrough Holocene. J. Foraminiferal Res. 16 (3), 207–215.
Bernhard, J.M., 1993. Experimental and field evidence of Antarcticforaminiferal tolerance to anoxia and hydrogen sulfide. Mar.Micropaleontol. 20, 203–213.
Bernhard, J.M., Alve, E., 1996. Survival, ATP pool, andultrastructural characterization of foraminifera from Drammen-sfjord (Norway): response to anoxia. Mar. Micropaleontol. 28,5–17.
Bernhard, J.M., Bowser, S.S., 1999. Benthic foraminifera of dysoxicsediments: chloroplast sequestration and functional morphology.Earth-Sci. Rev. 46, 149–165.
Bernhard, J.M., Sen Gupta, B.K., 1999. Foraminifera of oxygen-depleted environments. In: Sen Gupta, B.K. (Ed.), ModernForaminifera. Kluwer Academic Publishers, pp. 201–216.
Blanton, J.O., Atkinson, L.P., Fernande, F., Castillejo, F., Lavin, A.,1984. Coastal upwelling off the Rias Bajas. Galicia, northwestSpain. I. Hydrographic studies. Rapp. P.-V. Reun. Cons. Int.Explor. Mer (CIEM) 183, 79–90.
Bode, A., Casas, B., Varela, M., 1994. Size-fractioned primaryproductivity in the Galician shelf (NW Spain): netplankton versusnanoplakton dominance. Sci. Mar. 58, 131–141.
Bode, A., Casas, B., Fernández, E., Marañon, E., Serret, P., Varela, M.,1996. Phytoplankton biomass and production in shelf waters offNW Spain: spatial and seasonal variability in relation to upwelling.Hydrobiologia 341, 225–234.
Bond, G., Heinrich, H., Broecker, W., Labeyrie, L., Mcmanus, J.,Andrews, J., Houn, S., Jantschick, R., Clasen, S., Simet, C.,Tedesco, K., Klas, M., Bonani, G., Ivy, S., 1992. Evidence for

53V. Martins et al. / Marine Micropaleontology 59 (2006) 35–55
massive discharges of icebergs into the North Atlantic oceanduring the last glacial period. Nature 360, 245–249.
Bremer, J.M., 1983. Biogenic sediments on the southwest African(Namibian) continental margin. In: Thiede, J., Suess, E. (Eds.),Coastal Upwelling, Its Sediment Record, Part B. NATOConferences Series, vol. 4 (10B). Plenum Press, New York,pp. 423–430.
Caralp, M.H., 1984. Impact de la matière organique dans des zones deforte productivité sur certains foraminifères benthiques. Oceanol.Acta 7 (4), 509–515.
Caralp, M.H., 1988. Late Glacial to recent deep-sea benthicforaminifera from the northeastern Atlantic (Cadiz Gulf) andwestern Mediterranean (Alboran Sea): paleooceanographic results.Mar. Micropaleontol. 13, 265–289.
Casford, J.S.L., Rohling, E.J., Abu-Zied, R.H., Fontanier, C., Jorissen,F.J., Leng, M.J., Schmiedl, G., Thomson, J., 2003. A dynamicconcept for eastern Mediterranean circulation and oxygenationduring sapropel formation. Palaeogeogr. Palaeoclimatol. Palaeoe-col. 190, 103–119.
Castro, C.G., Pérez, F.F., Álvarez-Salgado, X.A., Fraga, F., 2000.Coupling between the thermohaline, chemical and biological fieldsduring two contrasting upwelling events off the NW IberianPeninsula. Cont. Shelf Res. 20, 189–210.
Cearreta, A., 1989. Foraminiferal assemblages in the Ria of SanVicente de la Barquera (Cantabria, Spain). Rev. Esp. Micro-paleontol. 12 (1), 67–80.
Corliss, B.H., 1991. Morphology and microhabitat preferences ofbenthic foraminifera from the northwest Atlantic Ocean. Mar.Micropaleontol. 17, 195–236.
De Rijk, S., Jorissen, F.J., Rohling, E.J., Troelstra, S.R., 2000. Organicflux control on bathymetric zonation of Mediterranean benthicforaminifera. Mar. Micropaleontol. 40, 151–166.
Dias, J.M.A., Gonzalez, R., Garcia, C., Diaz-del-Rio, V., 2002.Sediment distribution patterns on the Galicia-Minho continentalshelf. Prog. Oceanog. 52 (2–4), 215–231.
Donnici, S., Barbero, R.S., 2002. The benthic foraminiferal commu-nities of the northern Adriatic continental shelf. Mar. Micro-paleontol. 44, 93–123.
Drago, T., Araújo, F., Valério, P., Weber, O., Jouanneau, J.M., 1999.Geomorphological control of fine sedimentation on the northernPortuguese shelf. Bol. Inst. Esp. Oceanogr. 15 (1–4), 111–122.
Dulk, M., Reichart, G.J., van Heyst, S., Zachariasse, W.J., van derZwaan, G.J., 2000. Benthic foraminifera as proxies of organicmatter flux and bottom water oxygenation? A case history from thenorthern Arabian Sea. Palaeogeogr. Palaeoclimatol. Palaeoecol.161, 337–359.
Ellis, B.F., Messina, A.R., 1940–1988. Catalogue of index smallerforaminifera. The American Museum of Natural History, vol. 4.New York.
Estrada, M., 1984. Phytoplankton distribution and composition of thecoast of Galicia (northwest of Spain). J. Plankton Res. 6,417–434.
Evans, J.R., Austin, W.E.N., Brew, D.S., Wilkinson, I.P., Kennedy,H.A., 2002. Holocene shelf sea evolution offshore northeastEngland. Mar. Geol. 191 (3–4), 147–164.
Everitt, B.S., 1993. Cluster Analysis. Edward Arnold, London.Fariduddin, M., Loubere, P., 1997. The surface ocean productivity
response of deeper water benthic foraminifera in the AtlanticOcean. Mar. Micropaleontol. 32, 289–310.
Feyling-Hanssen, R.W., Funder, K.S., Peterson, K.S., 1983. The LodinElv formation: a Plio-Pleistocene occurrence in Greenland. Geol.Soc. Den. Bull. 31, 81–106.
Fiúza, A., 1983. Upwelling patterns off Portugal. In: Suess, E., Thiede,J. (Eds.), Coastal Upwelling — Its Sediment Record, Part A.Plenum Press, pp. 85–98.
Fiúza, A., 1984. Hidrologia e dinâmica das águas costeiras dePortugal. PhD thesis, Lisbon Univ., 294 pp.
Fiúza, A., Macedo, M., Guerreiro, R., 1982. Climatological space andtime variation of the Portuguese coastal upwelling. Oceanol. Acta5 (1), 31–40.
Fontanier, C., Jorissen, F.J., Licari, L., Alexandre, A., Anschutz, P.,Carbonel, P., 2002. Live benthic foraminiferal faunas from the Bayof Biscay: faunal density, composition, and microhabitats. Deep-Sea Res. Part I 49 (4), 751–785.
Fontanier, C., Jorissen, F.J., Chaillou, G., David, C., Anschutz, P.,Lafon, V., 2003. Seasonal and interannual variability of benthicforaminiferal faunas at 550 m depth in the Bay of Biscay. Deep-SeaRes. I 50, 457–494.
Frouin, R., Fiuza, A.F.G., Ambar, I., Boyd, T.J., 1990. Observations ofa poleward surface current off the coast of Portugal and Spainduring winter. J. Geophys. Res. 95, 679–691.
Hanson, R.B., Alvarez-Ossorio, M.T., Cal, R., Campos, M.J., Roman,M., Santiago, G., Varela, M., Yoder, J.A., 1986. Plankton responsefollowing a spring upwelling event in the Ria de Arosa, Spain.Mar. Ecol., Prog. Ser. 32, 101–113.
Haynes, R., Barton, E.D., Pilling, I., 1993. Development, persistenceand variability of upwelling filaments off the Atlantic coast of theIberian Peninsula. J. Geophys. Res. 98, 22681–22692.
Hohenegger, J., Piller, W.E., Baal, C., 1993. Horizontal and verticalspatial microdistribution of foraminifers in the shallow subtidal Gulfof Trieste, northern Adriatic Sea. J. Foraminiferal Res. 23, 79–101.
Huthnance, JM., 1995. Circulation, exchange and water masses at theocean margin: the role of physical processes at the shelf edge. Prog.Oceanogr. 35, 353–431.
Huthnance, J.M., Coelho, H., Griths, C.R., Knight, P.J., Rees, A.P.,Sinha, B., Vangriesheim, A., White, M., Chatwin, P.G., 2001.Physical structures, advection and mixing in the region of Gobanspur. Deep-Sea Res. II 48, 2979–3021.
Huthnance, J.M., Van Aken, H.M., White, M., Barton, E.D., Le Cann,B., Ferreira Coelho, E., Alvarez Fanjul, E., Miller, P., Vitorino, J.,2002. Ocean margin exchange–water flux estimates. J. Mar. Syst.32, 107–137.
Jannink, N.T., Zachariasse, W.J., van der Zwaan, G.J., 1998. Living(Rose Bengal stained) benthic foraminifera from the Pakistancontinental margin (northern Arabian Sea). Deep-Sea Res. I 45,1483–1513.
Jorissen, F.J., 1987. The distribution of benthic foraminifera in theAdriatic Sea. Mar. Micropaleontol. 12, 21–48.
Jorissen, F.J., 1999. Benthic foraminiferal successions late QuaternaryMediterranean sapropels. Mar. Geol. 153, 91–101.
Jorissen, F.J., Stigter, H.C., Widmark, J.G.V., 1995. A conceptualmodel explaining benthic foraminiferal microhabitats. Mar.Micropaleontol. 26, 3–15.
Jorissen, F.J., Wittling, I., Peypouquet, J.P., Rabouille, C., Relexans,J.C., 1998. Live benthic foraminiferal faunas off Cape Blanc,NW-Africa: community structure and microhabitats. Deep-SeaRes. I 45, 2157–2188.
Jouanneau, J.M., Weber, O., Drago, T., Rodrigues, A., Oliveira, A.,Dias, J.M.A., Garcia, C., Schmidt, S., Reyss, J.L., 2002. Recentsedimentation and sedimentary budgets on the western Iberianshelf. Prog. Oceanogr. 52 (2–4), 261–275.
Kim, J.-M., Kennett, J.P., 1998. Paleoenvironmental changes associ-ated with the Holocene marine transgression, Yellow Sea(Hwanghae). Mar. Micropaleontol. 34, 71–89.

54 V. Martins et al. / Marine Micropaleontology 59 (2006) 35–55
Kitazato, H., Shirayama, Y., Nakatsuka, T., Fujiwara, S., Shimanaga,M., Kato, Y., Okada, Y., Kanda, J., Yamaoka, A., Masuzawa, T.,Suzuki, K., 2000. Seasonal phytodetritus deposition and responsesof bathyal benthic foraminiferal populations in Sagami Bay, Japan:preliminary results from “Project Sagami 1996–1999”. Mar.Micropaleontol. 40, 135–149.
Knudsen, K.L., Seidenkrantz, M.-S., 1994. Stainforthia feylingi: newspecies from Arctic to subarctic environments, previouslyrecorded as Fursenkoina schreiberziana (Cszjek). ContributionsCushman Foundation Foraminiferal Res., Special Pub., vol. 32,pp. 5–13.
Kudela, R., Pitcher, G., Probyn, T., Figueiras, F., Moita, T., Trainer, V.,2005. Coastal in harmful algal blooms upwelling systems.Oceanography 18 (2), 185–197.
Langer, M.R., Hottinger, L., Huber, B., 1989. Functional morphologyin low-diverse benthic foraminiferal assemblages from tidal flats ofthe North Sea. Senkenb. Marit. 20, 81–99.
Leutenegger, S., Hansen, H.J., 1979. Ultrastructural and radio-tracer studies of pore-function in foraminifera. Mar. Biol. 54,11–16.
Levy, A., Mathieu, R., Poignant, A., Rosset-Moulinier, M., Ubaldo,M.L., Lebreiro, S., 1995. Foraminiferes Actuels de la MargeContinentale Portuguaise-inventaire et distribution. Mem. Inst.Geol. Min., Lisbon 32 (116 pp.).
Linke, P., Lutze, G.F., 1993. Microhabitat preferences of benthicforaminifera — a static concept or a dynamic adaptation tooptimize food acquisition? Mar. Micropaleontol. 20, 215–234.
Loeblich, A.R., Tappan, H., 1988. Foraminiferal Genera and TheirClassification. Van Nostrand Reinhold, New York. 970 pp.
López-Jamar, E., Cal, R.M., Gonzalez, G., Hanson, R.B., Rey, J.,Santiago, G., Tenore, K.R., 1992. Upwelling and outwellingeffects on the benthic regime of the continental shelf off Galicia,NW Spain. J. Mar. Res. 50, 465–488.
Loubere, P., 1996. Surface ocean productivity and bottom wateroxygen signals in deep-water benthic foraminiferal assemblages.Mar. Micropaleontol. 28, 247–261.
Loubere, P., Fariduddin, M., 1999. Benthic foraminifera and the fluxof organic carbon to the seabed. In: Sen Gupta, B.K. (Ed.),Modern Foraminifera. Kluwer Academic Publishers, Dordrecht,pp. 181–199.
Lutze, G.F., Coulbourn, W.T., 1984. Recent benthic foraminifera fromthe continental margin off Northwest Africa: community structureand distribution. Mar. Micropaleontol. 8, 361–401.
Mackensen, A., Douglas, R.G., 1989. Down-core distribution of liveand dead deep-water benthic foraminifera in box cores from theWeddell Sea and the California continental margin. Deep-Sea Res.36 (6), 879–900.
Mackensen, A., Hald, M., 1988. Cassidulina teretis Tappan andCassidulina laevigata D'Orbigny: their modern and lateQuaternary distribution in Northern Seas. J. Foraminiferal Res.8 (1), 16–24.
Mackensen, A., Grobe, H., Kuhn, G., Fütterer, D.K., 1990. Benthicforaminiferal assemblages from the eastern Weddel Sea between68° and 73°S: distribution, ecology and fossilization potential.Mar. Micropaleontol. 16, 241–283.
Martins, V., Gomes, V., 2004. In: Gomes, C. (Ed.), Foraminíferosda Margem Continental NW Ibérica: Sistemática, Ecologia.377 pp.
Martins, V., Jouanneau, J., Weber, J., Patinha, C., Silva, E.F., Terroso,D., Dias, J.A., Rocha, F., 2005. Holocene record of productivity inthe NW Iberian continental shelf. Cuad. Lab. Xeol. Laxe 30,99–124.
Martins, V., Patinha, C., Ferreira da Silva, E., Jouanneau, J.-M.,Weber, O., Rocha, F., 2006. Holocene record of productivity inthe NW Iberian continental shelf. J. Geochem. Explor. 88 (1–3),408–411.
McClain, C.R., Chao, S., Atkinson, L.P., Blanton, J.O., Castillejo, F.,1986. Wind driven upwelling in the vicinity of Cape Finisterre,Spain. J. Geophys. Res. 91, 8470–8486.
Mendes, I., Gonzalez, R., Lobo, F., Dias, J.M.A., Martins, V., 2004.Factors influencing the distribution of recent benthic foraminiferaon the Guadiana shelf (southwestern Iberia). Mar. Micropaleontol.51, 171–192.
Moodley, L., 1990. Southern North Sea seafloor and subsurface dis-tribution of living benthic foraminifera. Neth. J. Sea Res. 27,57–71.
Murray, J.W., 1971. An Atlas of British Recent Foraminiferids.Heinemann Educ. Books, London. 244 pp.
Murray, J.W., 1973. Distribution and Ecology of Living BenthicForaminiferids. Heinemann Educational Books, London. 274 pp.
Murray, J.W., 1991. Ecology and palaeoecology of benthic foraminif-era. Longman Scientific and Technical, London. 397 pp.
Nogueira, E., González-Nuevo, G., Morán, X.A.G., Varela, M., Bode,A., 2003. Hydrological structures along the N and NW IberianShelf during the winter–spring transition. 4rd Symposium on theIberian Atlantic Continental Margin, Vigo, Spain. Thalassas 19b,65–66.
Ohga, T., Kitazato, H., 1997. Seasonal changes in bathyal foraminif-eral populations in response to the flux of organic matter (SagamiBay, Japan). Terra Nova 9, 28–31.
Oliveira, A., Rocha, F., Rodrigues, A., Jouanneau, J., Dias,A.,Weber,O.,Gomes, C., 2002. Clayminerals from the sedimentary cover from thenorthwest Iberian shelf. Prog. Oceanogr. 52 (2–4), 233–247.
Pérez, F.F., Mouriño, C., Fraga, F., Ríos, A.F., 1993. Displacement ofwater masses and remineralization rates off the Iberian Peninsulaby nutrient anomalies. J. Mar. Res. 51, 869–892.
Phleger, F.B., Soutar, A., 1973. Production of benthic foraminifera inthree east Pacific oxygen minima. Micropalaeontology 19 (1),110–115.
Pitcher, G.C., Boyd, A.J., Horstman, D.A., Mitchell-Innes, B.A., 1998.Subsurface dino-flagellate populations, frontal blooms and theformation of red tide in the southern Benguela upwelling system.Mar. Ecol. Prog. Ser. 172, 243–264.
Prego, R., 1993. General aspects of carbon biogeochemistry in the Ria deVigo, northwestern Spain. Geochim. Cosmochim. Acta 57,2,041–2,052.
Prego, R., Bao, R., 1997. Upwelling influence on the Galician coast:silicate in shelf water and underlying surface sediments. Cont.Shelf Res. 17 (3), 307–318.
Prego, R., Bao, R., Howland, R., 1995. The biogeochemical cycling ofdissolved silicate in a Galician ria. Aphelia 42, 301–318.
Quinterno, P.J., Gardner, J.V., 1987. Benthic foraminifers on thecontinental shelf and upper slope, Russian river area, northernCalifornia. J. Foraminiferal Res. 17 (2), 132–152.
Rasmussen, T.L., Thomsen, E., Troelstra, S.R., 2002. Millennial-scaleglacial variability versus Holocene stability: changes in plankticand benthic foraminifera faunas and ocean circulation in the NorthAtlantic during the last 60000 years. Mar. Micropaleontol. 47,143–176.
Rathburn, A.E., Corliss, B.H., 1994. The ecology of living (stained)deep-sea benthic foraminifera from the Sulu Sea. Paleoceanogra-phy 9 (1), 87–150.
Rey, J., Diaz del Rio, V., 1987. Structure of recent sedimentary units inthe Galician continental shelf. Cuad. Lab. Xeol. Laxe 12, 35–45.

55V. Martins et al. / Marine Micropaleontology 59 (2006) 35–55
Ríos, A.F., Pérez, F.F., Fraga, F., 1992. Water masses in the upper andmiddle North Atlantic Ocean east of the Azores. Deep-Sea Res. 39,645–658.
Ross, C.R., 1984. Hyalinea balthica and its late Quaternary paleo-climatic implications: strait of Sicily. J. Foraminiferal Res. 14 (2),134–139.
Saffert, H., Thomas, E., 1998. Living foraminifera and totalpopulations in salt marsh peat cores: Kelsey March (Clinton, CT)and the Great Marshes (Barnstable, MA). Mar. Micropaleontol. 33,175–202.
Scott, G.A., Scourse, J.D., Austin, W.E.N., 2003. The distribution ofbenthic foraminifera in the Celtic Sea: the significance of seasonalstratification. J. Foraminiferal Res. 33, 32–61.
Scourse, J.D., Austin, W.E.N., Long, B.T., Assinder, D.J., Huws, D.,2002. Holocene evolution of seasonal stratification in the CelticSea: refined age model, mixing depths and foraminiferalstratigraphy. Mar. Geol. 191, 119–145.
Sejrup, H. -P., Fleran, T., Hald, M., Beck, L., Hagen, J., Miljeteig, I.,Morvik, I., Norvik, O., 1981. Benthonic foraminifera in surfacesamples from the Norwegian continental margin between 62° Nand 65° N. J. Foraminiferal Res. 11 (4), 277–295.
Shannon, C., Weaver, W., 1999. The Mathematical Theory ofCommunication, 5.a Ed. University of Illinois Press. 144 pp.
Silva, K.A., Corliss, B.H., Rathburn, A.E., Thunell, R.C., 1996.Seasonality of living benthic foraminifera from the San PedroBasin, California Borderland. J. Foraminiferal Res. 26, 71–93.
Smart, C.M., King, S., Gooday, A.J., Murray, J.W., Thomas, E., 1994.A benthic foraminiferal proxy for pulsed organic matter palaeo-fluxes. Mar. Micropaleontol. 23, 89–99.
Smyth, T.J., Miller, P.I., Groom, S.B., Lavender, S.J., 2001. Remotesensing of sea surface temperature and chlorophyll duringLagrangian experiments at the Iberian margin. Prog. Oceanogr.51 (2–4), 269–281.
Soares, A.M., 1989. Efeito de Reservatório Oceânico Nas ÁguasCosteiras de Portugal Continental. ICEN-LNETI, Departamentode Química. 135 pp.
Stigter, H.C., Jorissen, E.J., van der Zwaan, G.J., 1998. Bathymetricdistribution and microhabitat partionating of live (Rose BengalStained) benthic foraminifera along a shelf to bathial transect in thesouthern Adriatic Sea. J. Foraminiferal Res. 28 (1), 40–65.
Stuiver, M., Reimer, P.J., Bard, E., Beck, J.W., Burr, G.S., Hughen,K.A., Kromer, B., McCormac, F.G., van der Plich, J., Spurk, M.,1998. INTCAL98 radiocarbon age calibration, 24000–0 cal. BP.Radiocarbon 40, 1041–1083.
Teixeira, I., Figueiras, F.G., Castro, C.G., 2003. Hydrography,microplankton composition and primary production associatedwith the transition period between upwelling and downwellingseasons in Galicia waters (NW Spain). 4rd Symposium on theIberian Atlantic Continental Margin, Vigo, Spain. Thalassas 19a,201.
Tenore, K.R., Alonso-Noval, M., Alvarez-Ossorio, M., Atkinson, L.P.,Cabanas, J.M., Cal, R.M., Campos, H.J., Castillejo, F., Chesney,E.J., Gonzalez, N., Hanson, R.B., Mcclain, C.R., Miranda, A.,Roman, M.R., Sánchez, J., Santiago, G., Valdés, L., Varela, M.,Yoder, J., 1995. Fisheries and oceanography off Galicia, NWSpain: mesoscale spatial and temporal changes in physical pro-cesses and resultant patterns of biological productivity. J. Geophys.Res. 100 (C6), 10943–10966.
Thomas, E., Gooday, A.J., 1996. Cenozoic deep-sea benthicforaminifera: tracers for changes in oceanic productivity. Geology24, 355–358.
Thomas, E., Booth, L., Maslin, M., Shackleton, N.J., 1995.Northeastern Atlantic benthic foraminifera during the last 45,000years: changes in productivity seen from the bottom up.Paleoceanography 10, 545–562.
Tilstone, G.H., Figueiras, F.G., Fermín, E.G., Arbones, B., 1999.Significance of nanophytoplankton photosynthesis and primaryproduction in a coastal upwelling system (Ria de Vigo, NW Spain).Mar. Ecol. Prog. Ser. 183, 13–27.
van der Zwaan, G.J., Jorissen, F.J., 1991. Biofacial patterns in river-induced shelf anoxia. In: Tyson, R.V., Pearson, T.H. (Eds.),Modern and Ancient Continental Shelf Anoxia. Geol. Soc. LondonSpec. Publ., vol. 58, pp. 65–82.
Varela, M., Bode, A., Lorenzo, J., 2003. Differences in speciescomposition between spring and summer phytoplankton bloomsinduced by upwelling in the northwest Iberian shelf. 4rdSymposium on the Iberian Atlantic Continental Margin, Vigo,Spain. Thalassas 19a, 202.
Vitorino, J., Oliveira, A., Jouanneau, J.M., Drago, T., 2002. Winterdynamics on the northern Portuguese shelf. Part 1: physicalprocesses. Prog. Oceanogr. 52 (2–4), 129–153.
Wollenburg, J.E., Mackensen, A., 1998. Living benthic foraminifersfrom the central Arctic Ocean: faunal composition, standing stockand diversity. Mar. Micropaleontol. 34, 153–185.
Wooster, W.S., Bakun, A., Mclain, D.R., 1976. The seasonalupwelling cycle along the eastern boundary of the North Atlantic.J. Mar. Res. 34 (2), 131–141.




















