
Analytical methods for tracing plant hormones
20
REVIEW Analytical methods for tracing plant hormones Fuyou Du & Guihua Ruan & Huwei Liu Received: 9 October 2011 / Revised: 27 November 2011 / Accepted: 28 November 2011 / Published online: 4 January 2012 # Springer-Verlag 2011 Abstract Plant hormones play important roles in regulating numerous aspects of plant growth, development, and response to stress. In the past decade, more analytical methods for the accurate identification and quantitative determination of trace plant hormones have been developed to better our understand- ing of the molecular mechanisms of plant hormones. As sample preparation is often the bottleneck in analysis of plant hormones in biological samples, this review firstly discusses sample preparation techniques after a brief introduction to the classes, roles, and methods used in the analysis of plant hormones. The analytical methods, especially chromatograph- ic techniques and immuno-based methods, are reviewed in detail, and their corresponding advantages, limitations, appli- cations, and prospects are also discussed. This review mainly covers reports published from 2000 to the present on methods for the analysis of plant hormones. Keywords Plant hormones . Sample preparation . Chromatographic techniques . Mass spectrometry . Immuno-based analysis . Trace analysis Introduction Plant hormones are a structurally unrelated collection of small molecules that play a crucial role in regulating numerous aspects of plant growth, development, and response to a wide range of biotic and abiotic stresses [1–8]. The main molecular mechanisms involved in seed germination, cell proliferation, cell differentiation, organ formation, stem elongation, senes- cence, light responsiveness, and responses to undernourish- ment, drought, wounding, or insect and pathogen attack, and so on are all regulated by plant hormones. Thus plant hor- mones are capable of controlling crop plant size and architec- ture [9, 10], increasing crop grain productivity [11–13], and are expected to renovate crop breeding, improve agricultural pro- duction, and resolve global food shortages in the future. Owing to the potential applications in agriculture, medicine, and other fields, many biological scientists have therefore devoted them- selves to the research of plant hormones in hormone biosyn- thesis, metabolism, transport, perception, signaling and response, and tried to elucidate their molecular mechanisms [2–4, 14–17] and to expand their applications [18–23]. On the basis of diverse structures and physiological func- tions, plant hormones are generally grouped into several major classes including auxins, abscisic acid (ABA), cytokinins (CTKs), gibberellins (GAs), ethylene, salicylic acid (SA), jasm- onates (JAs), brassinosteroids (BRs), nitric oxide, strigolac- tones (SLs), and polypeptide hormones, and it is likely that additional plant hormones are yet to be discovered [2, 4, 17]. Chemically, most natural CTKs are adenine derivatives and can be classified by the configuration of their N 6 -side chain as either isoprenoid or aromatic; GAs are a family of tetracyclic diterpe- noid acids, and BRs are a group of naturally occurring poly- hydroxysteroids. Chemical structures of representative plant hormones and amino acid sequences of representative polypep- tide hormones are shown in Fig. 1 and Table 1, respectively. Each class of plant hormones has characteristic biological effects at trace concentrations, and most plant hormones are involved in many different processes throughout plant growth and development; however, they do not act in isolation but are interrelated by synergistic or antagonistic crosstalk so that they modulate each other’ s biosynthesis or responses [5, 8, F. Du (*) : G. Ruan College of Chemistry and Bioengineering, Guilin University of Technology, Guilin 541004, China e-mail: [email protected] H. Liu Institute of Analytical Chemistry, College of Chemistry and Molecular Engineering, Peking University, Beijing 100871, China Anal Bioanal Chem (2012) 403:55–74 DOI 10.1007/s00216-011-5623-x
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Analytical methods for tracing plant hormones

REVIEW
Analytical methods for tracing plant hormones
Fuyou Du & Guihua Ruan & Huwei Liu
Received: 9 October 2011 /Revised: 27 November 2011 /Accepted: 28 November 2011 /Published online: 4 January 2012# Springer-Verlag 2011
Abstract Plant hormones play important roles in regulatingnumerous aspects of plant growth, development, and responseto stress. In the past decade, more analytical methods for theaccurate identification and quantitative determination of traceplant hormones have been developed to better our understand-ing of the molecular mechanisms of plant hormones. Assample preparation is often the bottleneck in analysis of planthormones in biological samples, this review firstly discussessample preparation techniques after a brief introduction to theclasses, roles, and methods used in the analysis of planthormones. The analytical methods, especially chromatograph-ic techniques and immuno-based methods, are reviewed indetail, and their corresponding advantages, limitations, appli-cations, and prospects are also discussed. This review mainlycovers reports published from 2000 to the present on methodsfor the analysis of plant hormones.
Keywords Plant hormones . Sample preparation .
Chromatographic techniques .Mass spectrometry .
Immuno-based analysis . Trace analysis
Introduction
Plant hormones are a structurally unrelated collection of smallmolecules that play a crucial role in regulating numerousaspects of plant growth, development, and response to a wide
range of biotic and abiotic stresses [1–8]. The main molecularmechanisms involved in seed germination, cell proliferation,cell differentiation, organ formation, stem elongation, senes-cence, light responsiveness, and responses to undernourish-ment, drought, wounding, or insect and pathogen attack, andso on are all regulated by plant hormones. Thus plant hor-mones are capable of controlling crop plant size and architec-ture [9, 10], increasing crop grain productivity [11–13], and areexpected to renovate crop breeding, improve agricultural pro-duction, and resolve global food shortages in the future. Owingto the potential applications in agriculture, medicine, and otherfields, many biological scientists have therefore devoted them-selves to the research of plant hormones in hormone biosyn-thesis, metabolism, transport, perception, signaling andresponse, and tried to elucidate their molecular mechanisms[2–4, 14–17] and to expand their applications [18–23].
On the basis of diverse structures and physiological func-tions, plant hormones are generally grouped into several majorclasses including auxins, abscisic acid (ABA), cytokinins(CTKs), gibberellins (GAs), ethylene, salicylic acid (SA), jasm-onates (JAs), brassinosteroids (BRs), nitric oxide, strigolac-tones (SLs), and polypeptide hormones, and it is likely thatadditional plant hormones are yet to be discovered [2, 4, 17].Chemically, most natural CTKs are adenine derivatives and canbe classified by the configuration of theirN6-side chain as eitherisoprenoid or aromatic; GAs are a family of tetracyclic diterpe-noid acids, and BRs are a group of naturally occurring poly-hydroxysteroids. Chemical structures of representative planthormones and amino acid sequences of representative polypep-tide hormones are shown in Fig. 1 and Table 1, respectively.
Each class of plant hormones has characteristic biologicaleffects at trace concentrations, and most plant hormones areinvolved in many different processes throughout plant growthand development; however, they do not act in isolation but areinterrelated by synergistic or antagonistic crosstalk so thatthey modulate each other’s biosynthesis or responses [5, 8,
F. Du (*) :G. RuanCollege of Chemistry and Bioengineering,Guilin University of Technology,Guilin 541004, Chinae-mail: [email protected]
H. LiuInstitute of Analytical Chemistry, College of Chemistryand Molecular Engineering, Peking University,Beijing 100871, China
Anal Bioanal Chem (2012) 403:55–74DOI 10.1007/s00216-011-5623-x
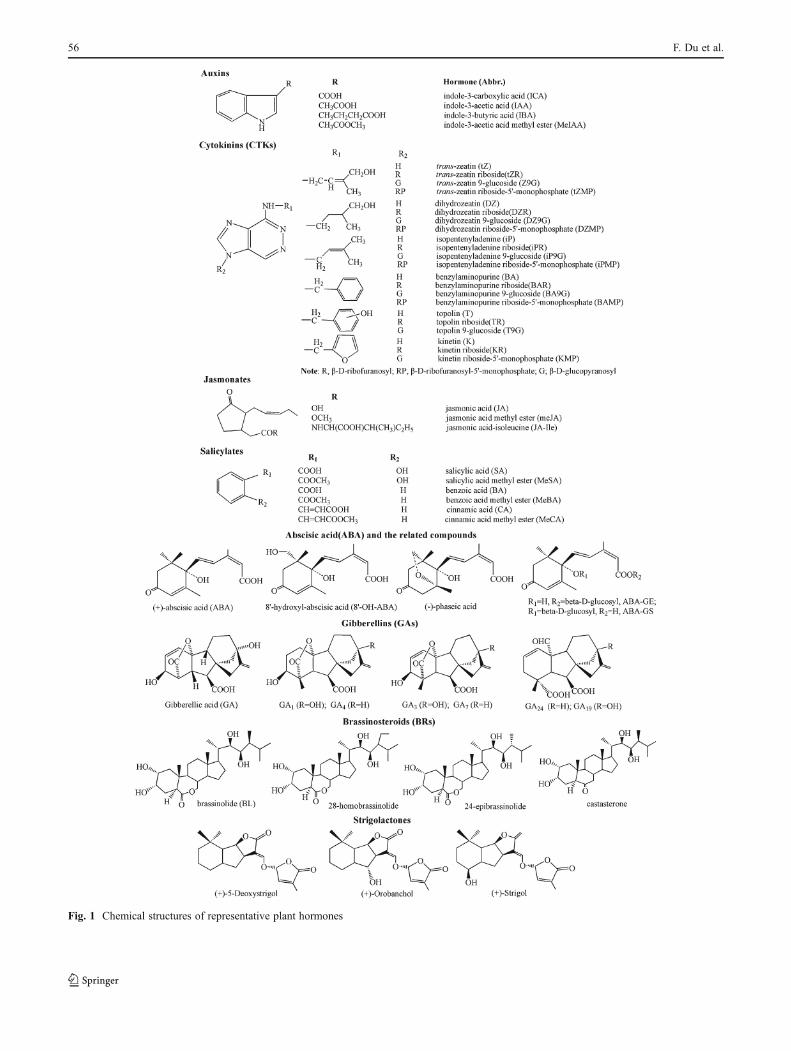
Fig. 1 Chemical structures of representative plant hormones
56 F. Du et al.

26–29]. Therefore, simultaneous quantitative profiling ofmul-tiple classes of plant hormones [30], which provides a basisfor defining additive, synergistic, or antagonistic hormoneactivities and identifying hormone networks regulating plantimmunity, is desired to investigate the concentration, diversi-fication, spacial and temporal distribution of hormones andthe related metabolites in plants, to discovery new hormones,and thus to better understand molecular mechanisms andinteractions of plant hormones.
Plant hormones are typically present in trace amounts inplants, and this makes qualitative and quantitative analysisdifficult. With the development of analytical techniques, moreaccurate and sensitive determination methods for plant hor-mones and related metabolites become available, and severalreview articles have been reported recently [31–35]. The deter-mination of trace plant hormones in complex biological sam-ples often requires extensive sample preparation techniquesprior to instrumental analysis, whether quantitative or qualita-tive. Sample preparation has become a bottleneck in the rapidseparation and ultrasensitive analysis of trace plant hormones,and more sample preparation techniques, including sampling,extraction, purification, and concentration methods, have beendeveloped for analyzing plant hormones [34, 35]. Generally,detection techniques include bioassays, immunoassays, elec-troanalysis, and especially chromatographic methods, such asgas chromatography (GC), capillary electrophoresis (CE), andhigh-performance liquid chromatography (HPLC) coupled
with different detectors. Bioassay is a classical method forhormone analysis [36] and it was first used in the determinationof auxins in 1928. The method is commonly directed to detectone compound, often being too specific to allow effectiveidentification of multiple analytes. Additionally, it is time con-suming, has poor repeatability, and is not very accurate, andtherefore has not attracted much attention in recent years.Immunoassays, including radioimmunoassays, enzyme-linkedimmunosorbent assays (ELISAs), avidin–biotin amplifiedELISA, scintillation proximity assays (SPAs), and immunocy-tochemical techniques, can be used as sensitive, viable alterna-tive assays and have been employed for plant hormonedeterminations [37–39]. A limited number of immunoassaykits are available commercially. However, the specific recog-nition involved in immunoassays has meant that their use in thesimultaneous determination of two or more classes of planthormones in a given sample is challenging. Chromatographicmethods, especially when coupled with mass spectrometry(MS), have shown good separation and qualitative abilitiesand are powerful tools for the detection of hormone contentsin complex plant samples. These studies started from the anal-ysis of one compound or one specified class [31, 32, 40], andgradually extended to two or more classes of plant hormonesand related compounds [30, 41–44]. These high-throughputand comprehensive approaches are vital for functionalgenomics research and for understanding the role ofhormone-induced signaling networks [45–47], and there is an
Table 1 Amino acid sequencesof representative polypeptidehormones [17, 24, 25]
Polypeptide hormone Amino acid sequence
Tomato systemin (TomSys) AVQSKPPSKRDPPKMQTD
Potato systemin I (PotSys I) AVHSTPPSKRDPPKMQTD
Potato systemin II (PotSys I) AAHSTPPSKRDPPKMQTD
Nightshade systemin AVRSTPPPKRDPPKMQTD
Pepper systemin (PepSys) AVHSTPPSKRPPPKMQTD
Tobacco systemin I RGANLPOOSOASSOOSKE
Tobacco systemin II NRKPLSOOSOKPADGQRP
Tobacco systemin III GRHDSVLPOOSOKTD
LeHypSys I RTOYKTOOOOTSSSOTHQ
LeHypSys II GRHDYVASOOOOKPQDEQRQ
LeHypSys III GRHDSVLPOOSOKTD
LeHypSys III GRHDSVLPOOSOKTD
PhHypSys I RSLHKSOOOTOKPSDEQGQ
PhHypSys II RHDYHLSOOOAOKPADHTGQ
PhHypSys III RGKRLPOOAOEYDPOYHQ
Alfalfa RALF ATTKYISYGALQRNTVPXSRRGASYYN
Asparagus PSK-α Y(SO3H)IY(SO3H)TQ
Asparagus PSK-β Y(SO3H)IY(SO3H)T
CLAVATA3 RTVP(OH)SGP(OH)DPLHH
POLARIS MKPRLCFNFRRRSISPCYISISYLLVAKLFKLFKIH
SCR (SP11) NLMKRCTRGFRKLGKCTTLEEEKCKTLYPRGQCTCSDSKMNTHSCDCKSC
Analytical methods for tracing plant hormones 57

increased demand for a highly sensitive, high-throughput, andcomprehensive analytical approach to elucidate the hormonalsignaling networks, functions, and dynamics.
This review present summarizes the current analyticalmethods for plant hormone analyses, including sample prep-aration, chromatographic analysis, immuno-based analysis,and others. This review also discusses the advantages, limi-tations, and applications of the developed approaches, and thefuture challenges involved in the detection of plant hormones.
Sample preparation
Sample preparation impacts nearly all the subsequent assaysteps and is hence critical for the unequivocal identification,confirmation, and quantification of analytes, especially thosepresent at trace or ultra-trace levels in complex matrices [48,49]. In addition, as analytical chemistry grows, sample prepa-ration gradually becomes a major part of the analysis, capableof taking up to 80% of the total time of a complete separation-based analytical process [50]. Therefore, sample preparation isa crucial process for trace plant hormone analysis.
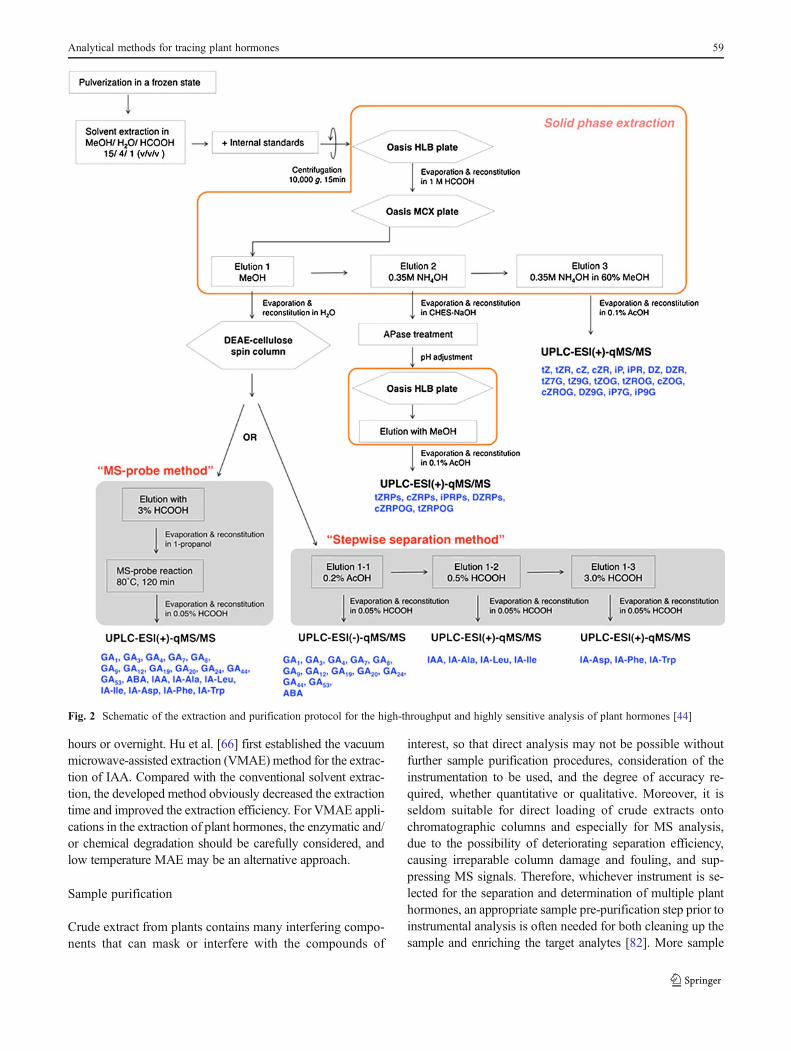
Individual steps of the procedures used in sample prepa-ration are important for the determination of analytes. Forthe analysis of plant hormones, sample preparation general-ly comprises several procedures, including sampling, freeze-drying, comminution, homogenization, exhaustive extrac-tion from the matrix, and subsequent removal of co-extracted interferences by several cleanup steps prior toinstrumental analysis. Figure 2 shows a typical comprehen-sive procedure used for the extraction, purification, andenrichment of multiple classes of plant hormones [44].
Sample extraction
The preparation and collection of plant samples for the analysisof trace analytes is a key step before sample extraction [51, 52].The concentrations and distribution of plant hormones in planttissues are affected by temperature, watering regime, air humid-ity, light intensity, and potassium chlorate concentration [8,53–55]. Therefore, the preparation conditions of plant samplesfor the analysis of trace plant hormones are rigidly controlled. Inaddition, wounding can induce changes in plant hormone levels;hence, it is important to freeze the collected fresh tissues in liquidnitrogen immediately after detaching the tissues from plants.And plant tissues need to be accurately weighed as dried powderbefore they are defrosted [30]. During the experimental process-es, enzymatic and/or chemical degradation and metabolism ofplant hormones must be effectively restrained to avoid thevariable concentrations, and thus appropriate extraction solvent,effective extraction technique, and optimal experimental condi-tions should be selected in the determination of plant hormones.
An ideal extraction solvent would provide high extrac-tion efficiency for all plant hormones in a given sample in asingle experiment. The modified Bieleski’s solvent (methanol/formic acid/water 15:1:4) is a good choice for the simultaneousextraction of multiple classes of plant hormones, includingauxins, CTKs, ABA, JAs, and a number of related metabolites[44, 56, 57], because it allows the enzymatic degradation ofplant hormones to be blocked without extracting large quanti-ties of lipids. Attention should be paid to the solvent, whichmay still release free plant hormones from their correspondingnucleosides, because this affects the accuracy and reliability ofthe determination of endogenous plant hormones. 1-Propanol/H2O/concentrated HCl [30, 42, 58], methanol/water [59], andacetonitrile [60] were also used as solvents in the simultaneousextraction of auxins, CTKs, ABA, GAs, JAs, salicylates, andtheir related metabolites in plant tissues.
For the extraction of individual plant hormones, diversesolvents such as methanol, methanol/water, acetone, acetone/water, propanol, propanol/water, neutral or acid buffer, and themodified Bieleski’s solvent are broadly used. Hydrolysis andother chemical degradation during the extraction process mustalso be considered. For example, indole-3-acetic acid (IAA)and IAA conjugates are the main natural auxins commonlyoccurring in all vascular and lower plants [15, 61], free auxinswere commonly extracted with 1-propanol/H2O/concentratedHCl [30, 42], modified Bieleski’s solvent [44], acetone/water[62, 63], methanol/water [64–67], isopropanol/acetic acid [68],isopropanol/imidazole buffer (pH 7) [40, 69, 70]), and evenionic liquids [71]. For the determination of total auxins, thehydrolysis of auxin conjugates in the crude extract was essen-tial, and an equal volume of 14 M NaOH was generally addedto the crude extract and then incubated at 100 °C for 3 h [69].Moreover, to prevent auxins from oxidation and degradation,addition of antioxidants during the extraction process is neces-sary [65–67]. For CTKs extraction, the modified Bieleski’ssolvent was superior to Bieleski’s solvent (methanol/chloroform/water/formic acid 12:5:2:1, v/v/v/v), methanol,ethanol, perchloric acid, and their mixtures, and gave thehighest responses for deuterated CTKs (used as test com-pounds) in plant extracts and sufficiently suppressed the de-phosphorylation of CTK mononucleotides [72]. Methanol[73], methanol/water [74, 75], and methanol/chloroform [76]were available for BRs extraction. For the extraction of plantpolypeptide hormones, cold water has been applied to extractsystemins [77–79], and 1% trifluoroacetic acid aqueous solu-tion was used to extract proline-rich systemins andhydroxyproline-rich glycopeptides [80, 81]. In fact, the selec-tion of the extraction solvent depends not only on the physico-chemical properties of the target plant hormones but also on thetypes of plant materials. The optimal solvent should be selectedby performing certain experiments.
Generally, plant hormones were extracted from solid plantsamples by maceration at 4 °C, even at −20 °C for several
58 F. Du et al.
hours or overnight. Hu et al. [66] first established the vacuummicrowave-assisted extraction (VMAE) method for the extrac-tion of IAA. Compared with the conventional solvent extrac-tion, the developed method obviously decreased the extractiontime and improved the extraction efficiency. For VMAE appli-cations in the extraction of plant hormones, the enzymatic and/or chemical degradation should be carefully considered, andlow temperature MAE may be an alternative approach.
Sample purification
Crude extract from plants contains many interfering compo-nents that can mask or interfere with the compounds of
interest, so that direct analysis may not be possible withoutfurther sample purification procedures, consideration of theinstrumentation to be used, and the degree of accuracy re-quired, whether quantitative or qualitative. Moreover, it isseldom suitable for direct loading of crude extracts ontochromatographic columns and especially for MS analysis,due to the possibility of deteriorating separation efficiency,causing irreparable column damage and fouling, and sup-pressing MS signals. Therefore, whichever instrument is se-lected for the separation and determination of multiple planthormones, an appropriate sample pre-purification step prior toinstrumental analysis is often needed for both cleaning up thesample and enriching the target analytes [82]. More sample
Fig. 2 Schematic of the extraction and purification protocol for the high-throughput and highly sensitive analysis of plant hormones [44]
Analytical methods for tracing plant hormones 59

purification techniques, including liquid–liquid extraction,solid-phase extraction, molecularly imprinted extraction, andimmunoaffinity purification have been developed to furtherpurify trace plant hormones from crude extracts.
Liquid–liquid extraction
Liquid–liquid extraction (LLE) is based on the solubilitydifference of target analytes between two liquid phases. Themost classical extraction is performed in separating funnels toextract analytes from an aqueous sample solution into a non-polar or less polar organic solvent. Many organic solvents,such as hexane, diethyl ether, butanol, ethyl acetate, dichloro-methane/isopropanol, and methylene chloride/1-propanol,have been used to purify plant hormones on the basis of thephysicochemical properties of the target hormones [42,83–88]. However, traditional LLE involves many extractionsteps and thus is time- and solvent-consuming work. It alsoreduces the extraction efficiency of water-soluble plant hor-mones because of serious emulsification. To resolve the issue,Absalan et al. [71] developed a novel LLE method by usingimidazolium-based ionic liquids as the extractant. The methodwas less time consuming, more efficient, and cheaper for thepurification of IBAwith [C4mim][PF6], and the IBA recoverywas higher than 96%.
Recently, liquid–liquid microextraction (LLME) in twoapplication modes (hollow fiber–LLME [89] and dispersiveliquid–liquid microextraction (DLLME) [90, 91]) has beenapplied for the further purification of plant hormones. Wuand Hu [89] established a simple, selective, sensitive, andinexpensive method of hollow fiber-based liquid–liquid–liquid microextraction (HF–LLLME) for the simultaneousextraction and concentration of four acidic plant hormones(IAA, ABA, SA, and JA) from natural coconut juice sam-ples. The HF–LLLME method provided good enrichmentfactors (48- to 243-fold) and high recoveries (88.3–119.1%)for the target plant hormones, along with excellent samplecleanup capability. DLLME has been applied for the simul-taneous extraction and concentration of major classes ofplant growth regulators (GA, ABA, SA, IAA, IBA, indole-3-propionic acid (IPA), 1-naphthylacetic acid (NAA), andkinetin riboside (KR)) from crude extract, and exhibited sev-eral merits including high enrichment efficiency and reducedsample preparation and analysis time [89, 90]. However, bothCHCl3 (extraction solvent) and acetone (disperser solvent) aretoxic, volatile, and flammable, so safe and environmentallybenign solvents are desired in the LLME purification of planthormones.
Solid-phase extraction
Compared with LLE, solid-phase extraction (SPE) benefitsfrom low intrinsic costs, shorter processing times, low
solvent consumption, and simpler processing procedures,and it thus is widely used to extract and enrich analytes onthe basis of diverse interactions, including adsorption, hy-drogen bonding, polar and nonpolar interactions, cation,anion exchange or size exclusion [92]. As for SPE, theselection of an appropriate SPE sorbent is a key to success,and it depends on understanding the mechanism of interac-tion between the sorbent and the analyte of interest. Thatunderstanding in turn depends on knowledge about theproperties of both sorbents and analytes, including theirhydrophobic, polar, acidic/basic, and ionogenic properties.SPE with various kinds of sorbents is a common procedurefor the further purification of plant hormones from crudeplant extracts by using different separation modes, includingion exchange [93, 94], reversed phase (C18 [56, 59, 72,95–98]; Oasis HLB [44, 56, 99]), and mixed mode (OasisMCX [44, 56, 72, 97, 100] and Oasis MAX [99, 101]).Generally, more than one kind of sorbent is used to effec-tively extract and enrich trace plant hormones from crudeextract, which might comprise complex interference com-pounds, and C18-based sorbents are the most widely used forthe pre-purifications.
SPE with Oasis MCX sorbents was selected to separateCTKs from auxin and ABA, and further separate CTK ribo-tides from CTK bases, ribosides, and glucosides, whichdepended on the mixed reversed-phase (RP) and cation-exchange interactions between plant hormones and the OasisMCX sorbents [100]. SPE with Oasis MAX sorbent, which isanion-exchange, water-wettable, mixed-mode SPE sorbent,was selected to separate and purify acidic plant hormones withanion-exchange groups such as auxin and ABA from crudeextracts [101]. In addition, SPE with a C18 Sep-Pak cartridgewas successfully applied to pre-concentrate multiple classes ofplant hormones and their metabolites from crude extracts [63,65], an approach which is mainly based on the hydrophobicinteractions between C18 Sep-Pak sorbents and plant hor-mones. Ma et al. [96] developed a pre-concentration methodby using a single C18 SPE cartridge to enrich six classes ofputative endogenous plant hormones from coconut water withreasonably high recoveries of 67–95%. In fact, sometimesonly a single C18 SPE cartridge is not enough to purifymultiple classes of plant hormones before LC–MS analysis.Izumi et al. [56] demonstrated that sample purification of fourclasses of plant hormones and their derivatives using OasisHLB and Oasis MCX cartridges significantly decreased theion-suppressing effects of biological matrix as compared withthe purification using only Oasis HLB cartridge; this indicatedthat the mixed-mode cartridges with both RP and anion-exchange characteristics are able to significantly enhance thesensitivity of MS detection and they can retain different clas-ses of plant hormones effectively. For the simultaneous ex-traction and concentration of 43 molecular species of CTKs,auxins, ABA, and GAs from a crude extract, the crude extract
60 F. Du et al.

was pre-purified by using the SPE with Oasis HLB plate.Mixed mode SPE with Oasis MCX plate was then used tofurther purify the obtained sample, resulting in simultaneousconcentration and separation of 43 hormones [44]. Similarly,SPE with Sep-Pak plus C18 and Oasis MCX cartridges wereused in turn to extract and concentrate auxins, CTKs, andABA from the crude extracts of two sets of Nicotiana glaucaand N. langsdorffii samples [57]. In addition, SPE with poly-dimethylsiloxane film was successfully used for the extractionof MeJA in leaf tissues [102].
In order to purify CTK ribotides, bases, ribosides, andglucosides from crude extracts, the mixed mode Oasis MCXSPE followed by C18 SPE was used and exhibited highextraction and concentration efficiency [97, 100]. However,for the relatively highly polar CTK nucleotides, the anion-exchange SPE column was effective in separating CTKnucleotides from CTK bases and sugar conjugates [94]. Adual-step SPE method has therefore been developed for pre-concentration and purification of CTK nucleotides usingOasis HLB and MAX cartridges with high recovery andless interfering substances in the final extracts [99].
For effective pre-purification of plant polypeptide hor-mones, a diethylaminoethyl (DEAE) cellulose column, RPC18 flash chromatography, Sephadex G25 gel filtration, CMSephadex chromatography, RP C18 column, strong cation-exchange HPLC, and RP C18 HPLC were applied once ortwice [80, 81]. Du et al. [79] selected semipreparative C18
liquid chromatography to pre-purify systemins from crudeextracts, which could protect the immunoaffinity column(IAC) from blocking by interferences and increased the enrich-ment efficiency of IAC.
Solid-phase microextraction (SPME) is a simple, time-efficient, and solvent-free sample preparation technique. Itintegrates sampling, extraction, concentration, and sampleintroduction into a single step and facilitates coupling betweenSPME and GC, GC–MS, LC, LC–MS, and other instruments[103], and has been successfully applied to the purification ofplant hormones [104–107]. For headspace (HS)-SPME, thevolatility of analyte was important in the extraction of planthormones [104, 105]. To improve SPME extraction efficien-cies, fiber type, pH value, salinity, extraction time, and tem-perature were optimized in the simultaneous extraction ofIAA, ABA, indole-3-butyric acid (IBA), and NAA from crudesamples, and the four plant hormones were selectivelyextracted with high recoveries by using Carbowax-coatedfibers under the optimal conditions [106]. Liu et al. [107]developed a polymer monolith-based SPME method to purifyCTKs from crude extracts of Oryza sativa, Arabidopsisthaliana, and oil seed rape tissues. The obtained resultsshowed that the proposed SPME method was suitable for thesimultaneous, selective purification of five CTKs. Although anumber of SPME fibers are now commercially available, theselective, high-throughput SPME fibers still are promising in
the simultaneous extraction of plant hormones from planttissues.
Molecularly imprinted extraction
Molecularly imprinted polymers (MIPs) are synthetic materialswith artificially generated recognition sites able to specificallybind a target molecule in preference to other closely relatedcompounds, and therefore MIPs have attracted extensive at-tention and been widely applied in many fields, such as SPE,chemical sensors, and artificial antibodies owing to their de-sired selectivity, physical robustness, thermal stability, as wellas low cost and easy preparation [108–110]. However, thereare few reports about the application of MIPs to the purifica-tion of plant hormones. Kugimiya and Takeuchi [111, 112]prepared an IAA-imprinted polymer as an SPEmaterial, whichexhibited a significant selectivity for IAA. The affinity of MIPwas obviously influenced by the functional monomer as aresult of electrostatic and hydrogen-bonding interactions withthe template molecule, and the MIP prepared with N,N-dime-thylaminoethyl methacrylate showed higher affinity and selec-tivity for IAA than the polymer prepared with methacrylicacid. Further investigations have indicated that the MIP mem-brane prepared with 9-vinyladenine had higher permselectivityfor IAA in comparison with the imprinted membrane madewith methacrylic acid, and exhibited higher transport selectiv-ity for the template molecule IAA than IBA or kinetin [113]. Liet al. [66, 67] developed an enhanced IAA imprinting methodbased on binary functional monomers, 4-vinylpyridine (4-VP),and β-cyclodextrin (β-CD). The magnetic molecularlyimprinted polymer (mag-MIP) beads, which were preparedby microwave heating initiated suspension polymerization,exhibited improved recognition ability toward IAA, as com-pared with the mag-MIP beads prepared with the individualmonomer separately. Themag-MiP beads selectively extractedand concentrated IAA and IBA from the crude extracts of planttissues, and were applied to investigate the developmentalprofiles of IAA concentration in pea seeds and rice seedsduring seed germination.
Recently, monolithic molecularly imprinted solid-phase ex-traction (MI-SPE) was first applied to extract and enrich CTKsfrom the crude extract [114]. The prepared MIP monolithexhibited specific recognition for kinetin (K), kinetin glucoside(KR), trans-zeatin (tZ), and meta-topolin (mT) in comparisonwith the nonimprinted polymer (NIP) monolith. Under theoptimized conditions, the mean recoveries of K, KR, tZ, andmT were 85.9%, 79.3%, 73.5%, and 70.1% with relativestandard deviations (RSDs) less than 15%, respectively, whichsuggested that MI-SPE is a potential technique for the purifi-cation of CTKs. Owing to the high selectivity of MI-SPE,however, the MI-SPE approach is unsuitable for the simulta-neous extraction of multiple classes of plant hormones.
Analytical methods for tracing plant hormones 61

Immunoaffinity extraction
Immunoaffinity extraction (IAE) is a high-affinity and highlyselective sample preparation technique based upon antigen–antibody interactions (molecular recognition), which enablesselective extraction and the concentration of individual com-pounds or classes of compounds from liquid matrices in onestep or the sample purification of extracts from solid matrices,and increases the sensitivity of the final instrumental determi-nations [115]. IAE provides unique and powerful techniquesfor trace analysis and has been successfully applied for thefurther sample purification of plant hormones [93, 116–121].However, obtaining selective antibodies for low molecularmass plant hormones is more difficult; therefore, before immu-nization plant hormones have to be modified by bonding to acarrier macromolecule, usually a protein such as bovine serumalbumin (BSA), to form antigens. On the other hand, theselection of the hapten is still an important variable when tryingto obtain antibodies with the required specificity, especiallywhen antibodies are targeted to recognizing a whole class ofstructurally related compounds. For obtaining antibodies withhigh specificity for aromatic CTKs, ortho-topolin riboside [93]or a synthetic benzylaminopurine [6-(2-hydroxy-3-methoxy-benzylamino)purine riboside (2OH3MeOBAPR)] [116] wasselected as the hapten and BSAwas used as a carrier protein.The obtained monoclonal antibodies against 2OH3MeOBAPRshowed very weak cross-reactivity (<0.1%) with all naturalisoprenoid CTKs, which allowed the specific depuration andefficient concentration of aromatic CTKs. Novák et al. [119]developed a simple, high-throughput batch IAE micropurifica-tion procedure for extracting a wide range of naturally occur-ring CTKs (bases, ribosides, O- and N-glucosides, andnucleotides) from plant tissues in solutions. The immunopur-ification procedure presented here provides possibilities toautomate CTK analysis because the whole extraction and pu-rification procedure can be performed in an Eppendorf tube.Liang et al. [121] developed a novel class-specific immunoaf-finity column (IAC) for the natural isoprenoid CTKs by usingtrans-zeatin riboside as the hapten to generate a completeantigen. The prepared IAC based on the polyclonal antibodiesagainst trans-zeatin riboside exhibited high specificity andcapacity for isoprenoid CTKs and achieved simultaneous ex-traction and enrichment of eight endogenous isoprenoid CTKsfrom A. thaliana leaves.
Recently, an IAC based on polyclonal antibodies againstsystemin was prepared and exhibited high specificity, highbinding capacity, and good reusability for systemins, andwas successfully applied to extract and enrich polypeptidehormones from Solanaceous plants [79]. However, the pu-rification efficiency of the IAC would decrease after beingused six times for the purification of systemins from realsamples, and this lack of durability suggested that IAC wasunsuitable for high-throughput analysis. To improve the
durability and reusability of IAC, a pre-cleanup process withC18-based SPE [117–120], mixed-mode SPE [121], or semi-preparative HPLC [79] was necessary in the purification ofplant hormones from real plant tissues. Nevertheless, IAChas great potential for the purification of plant hormonesbecause of its high selectivity and high sensitivity.
Other sample preparation methods
Besides the above sample preparation techniques, vapor-phaseextraction (VPE) is a promising approach to purify planthormones and improve GC–MS analysis, and a detailed pro-cedure of the VPE technique has been described [42, 122,123]. The method is regarded as one of the most robustmethods for plant hormone profiling by GC–MS, because itis a simple, sensitive, reliable, and highly efficient method,which based on a one-step extraction, phase partitioning,methylation with HCl/methanol, and collection of methylatedand, thus, volatilized compounds on Super Q filters, therebyomitting further purification steps. The avoidance of multiplecleaning steps by trapping volatile compounds from crudemethylated extracts reduces the loss of compounds as indicatedby the high recovery rate for JA and SA.
Chromatographic analysis
After purification and enrichment, the simultaneous, accuratedetermination of individual or multiple classes of plant hor-mones in the samples is still difficult due to their complicatedmatrix, but the high separation efficiency of chromatographyand its ability to be combined with different detectors makes itan ideal technique for separating plant hormones from thematrix and to provide improved quantitative results [35]. Mod-ern analysis of plant hormones relies heavily on chromato-graphic techniques, including GC, CE, and especially liquidchromatography (LC) in combination with different detectors.
LC and LC–MS analysis
LC has emerged as a powerful separation tool with highresolution, and has been widely used for the analysis of traceplant hormones since the early 1970s. LC coupled with vari-ous types of detectors, including UV–vis detectors, fluores-cence detection, and especially MS, is the most commonlyapplied method for hormone determination, and some excel-lent reviews on this topic have reported in special journalissues [31–35].
LC–UV was developed for the simultaneous determinationof various classes of plant hormones, including IAA, IBA,ABA, GA, SA, JA, and CTKs in natural plant samples [89,96]. However, the method was not suitable for the determina-tion of some plant hormones (e.g., JAs) which are not very
62 F. Du et al.

sensitive in UV detection. An alternative means to the tradi-tional UV method is fluorescence detection (FLD) [90,124–126]. JAs were derivatized using 9-anthryldiazomethaneand thenwere determined by LC–FLD; the agreement betweenthe LC–tandem mass spectrometry (MS/MS) and LC–FLDresults confirmed the specificity and accuracy of the proposedLC–FLDmethod [124]. For the simultaneous determination ofendogenous plant hormones, including IBA, NAA, and 2,4-dichlorophenoxyacetic acid (2,4-DA), 6-oxy(acetylpiperazine)fluorescein (APF) was used as a new fluorescent labelingreagent and the proposed LC–FLD method was applied tothe determination of the exogenous plant hormones in thecrude extracts of vegetable samples without extra purificationand enrichment with high recoveries [126]. To increase theselectivity and reliability, Dobrev et al. [100] developed a two-dimensional liquid chromatography (2D-LC) method in “heartcutting” mode, which efficiently removed the contaminantsco-eluting with IAA and ABA. The quantification by 2D-HPLC with online UV (ABA) and FLD (IAA) detection isstatistically identical (with 95% confidence) to those measuredby GC–MS.
Generally, LC–MS may be combined with single-quadrupole or triple-quadrupole instruments (QqQ), quad-rupole linear ion trap (QqLIT), time-of-flight (ToF) spec-trometers, or hybrid quadrupole ToF instruments. LC–MS ismost widely employed for the simultaneous qualitative andquantitative profiling of multiple classes of plant hormonesbecause of the high sensitivity and selectivity of MS [33].The capacity to simultaneously qualify and quantify multi-ple classes of plant hormones will facilitate the study ofhormone function and crosstalk.
A sensitive method for the quantitative analysis of allnatural isoprenoid CTKs in plant material by electrospraysingle-quadrupole mass spectrometry has been developedand a baseline chromatographic separation of 20 nonderiv-atized naturally occurring CTKs has been obtained [93].Compound identification and quantification largely dependupon separation and the uniqueness of the ions selected formonitoring. Using a single quadrupole instrument, it may benecessary to confirm LC–MS selectivity by employing othersupplementary detection methods such as UV, fluorescence,or immunoactivity assay. Compared with the full scan MSmode, MS/MS with selected reaction monitoring (SRM),single ion recording (SIR), and especially multiple reactionmonitoring (MRM) mode provide better selectivity, specific-ity, and sensitivity for the simultaneous analysis and quantifi-cation of individual or multiple classes of plant hormones[33].
Liquid chromatography–electrospray ionization–tandemmass spectrometry (LC–ESI–MS/MS) has emerged as aneffective method for plant hormone analysis. This approachhas been applied for the simultaneous determination ofindividual or multiple classes of plant hormones and their
metabolites [30, 43, 56, 58, 63, 85, 88, 95, 96, 127–132].For example, Chiwocha et al. [63] developed a highlyselective and sensitive LC–ESI–MS/MS method for thesimultaneous analysis and quantification of plant hormonesand metabolites from four of the nine major classes (auxins,CTKs, ABA, and GAs) using deuterium-labeled analogs asinternal standards in a single run. The proposed method wasspecific and sensitive because all components are partiallypurified using LC, followed by ESI–MS/MS detection inMRM mode, and it was successfully applied to the analysisof plant hormones and hormonal metabolites associatedwith seed dormancy and germination in lettuce. Pan et al.[30, 58] described a rapid and sensitive LC–ESI–MS/MSmethod for the simultaneous quantification of seven majorclasses of plant hormones including auxins, CTKs, ABA,GAs, JAs, SAs, and relevant methyl esters in crude plantextracts without purification or derivatization. After doubleextraction, the sample was subjected to analysis with LC–MS/MS without further derivatization, which realized thedifferentiation between hormone esters and their acid formfrom complicated biological matrices.
In order to rapidly and effectively separate multiple classesof plant hormones and improve the MS sensitivity, micro-column liquid chromatography (mLC) [95], capillary liquidchromatography (capillary-LC), nanoflow liquid chromatog-raphy (nano-LC) [56], and especially ultra-performance liquidchromatography (UPLC) [44, 119] are receiving much atten-tion owing to their excellent performance in terms of highchromatographic resolution, high peak capacity, high sensi-tivity, and rapid speed of analysis. Izumi et al. [56] developeda highly sensitive and comprehensive analytical approach forthe simultaneous determination of four classes of plant hor-mones and their derivatives, including auxins, CTKs, ABA,and GAs, based on nanoflow liquid chromatography–electro-spray ionization–ion trap mass spectrometry (nano-LC–ESI–IT-MS/MS) under MRM mode detection. The optimizednano-LC–ESI–IT-MS/MS method showed ca. 5- to 10-foldgreater sensitivity than capillary-LC–ESI–IT-MS/MS, and thedetection limits (S/N 3) of several plant hormones were in thesub-femtomole range. Kojima et al. [44] established a highlysensitive and high-throughput method for the simultaneousanalysis of 43 molecular species of CTKs, auxins, ABA, andGAs (Fig. 2). This method consists of an automatic liquidhandling system for SPE and UPLC coupled with a tandemquadrupole mass spectrometer (qMS/MS) equipped with anelectrospray interface (UPLC–ESI–qMS/MS). In order to im-prove the detection limit of negatively charged compounds,such as GAs, Kojima et al. chemically derivatized fractionscontaining auxins, ABA, and GAs with bromocholine, whichhas a quaternary ammonium functional group. The developedmethod only needed 1 to 100 mg fresh weight of plant tissuesto determine plant hormone profiles and could analyze 180plant samples simultaneously.
Analytical methods for tracing plant hormones 63

Time-of-flight mass spectrometry (ToF-MS) in combina-tion with LC is more often used in high-resolution mode(typical mass error <2 mDa), which provides better discrim-ination of background and more accurate identification ofunknown peaks in a sample even if analytical standards arenot available. Therefore, a liquid chromatography–quadru-pole time-of-flight tandem mass spectrometry (LC–QToF-MS/MS) method was proposed for the determination oftrace plant hormones [55, 79, 133–136]. Du et al. [79]developed a superior method for the accurate identificationand quantitative determination of systemins in Solanaceousplants with IAC purification and enrichment followed byLC online coupled to electrospray quadrupole time-of-flightmass spectrometry (LC–ESI–QToF-MS) without addition ofinternal standards, which suggested that ESI–QToF-MS/MSwas suitable for wide-range polypeptide hormone profilingbecause of its higher mass accuracy and better resolution.Under the optimized conditions, the developed methodwas successfully applied to the determination of TomSysin tomato leaves and PotSys I and PotSys II in potatoleaves.
Moreover, it is important to determinate different chiralplant hormones to understand the molecular mechanisms ofstereoisomers in plant growth, development, and response tostress [137–139]. For example, small changes in the struc-ture of JA or other JAs will result in hormone inactivation oractivity through epimerization, which implies that the spe-cific bioactivities of JA and other JAs are dependent onchemical form [138, 139]. The reported chiral HPLC meth-od could only distinguish (−)-JA ((−)-(3R,7R)-JA) and (+)-JA ((+)-(3S,7S)-JA) but was powerless for all four stereo-isomers, i.e., (−)-JA, (+)-JA, (−)-epi-JA ((−)-(3S,7R)-JA),and (+)-epi-JA ((+)-(3R,7S)-JA) [140]. Blanch et al. [141]developed a method based on the use of permethyl-β-CDHPLC for the enantioselective resolution of the four stereo-isomers of methyl jasmonate with no need for the priorformation of the diastereoisomers. Recently, Han et al.[136] established a novel chiral liquid chromatography–quadrupole time-of-flight mass spectrometry (cLC–QToF-MS) method for the simultaneous discrimination and directanalysis of all four JA stereoisomers without the need forderivatization. Four JA stereoisomers were distinguishedfrom each other and satisfactory results were obtained in termsof sensitivity, linearity, and repeatability. For enantiseparationand determination of chiral volatile plant hormones, an onlineRPLC–GC via through-oven transfer adsorption−desorption(TOTAD) (RPLC–TOTAD-GC) method was proposed andwas suitable for the determination of the levels and enantio-meric composition of chiral plant hormones in plant samples[142, 143].
Generally, the effect of the matrix on the determination oftrace plant hormones was obvious [144], and thus internalstandards were necessary for accurate evaluation of method
recovery and ensuring more reliable results in target quantifi-cation. Isotope-labeled compounds (e.g., deuterium-labeledstandards) are appropriate to be internal standards, as theyhave almost the same physical and chemical properties astheir corresponding analytes, which provides correction foranalytes loss during sample preparation and also compensatesthe suppression of ion yield caused by co-eluting componentsin the sample matrix [33, 145]. However, sometimes properinternal standards are unavailable for the determination ofplant hormones, and the synthesis of internal standards bringsextra workload for the whole analysis.
GC and GC–MS analysis
GC separation depends on the hormone’s different phys-iochemical properties, such as boiling points and molec-ular polarity, and different physiochemical interactionswith the GC column [33]. GC coupled to UV detection,fluorescence detection, or MS have been proposed anddemonstrated as powerful tools for profiling plant hor-mones and related metabolites, and GC–MS is the mostfrequently used tool owing to its high sensitivity andselectivity [31–35, 42, 146].
Prior to GC–MS analysis, appropriate derivatization (eithermanual or robotic) is an important process for plant hormoneanalysis as it not only enhances volatility but also sometimesimproves the stability of many compounds as well as facili-tates GC separation and MS ionization [33]. The widely usedreagents for the derivatization of plant hormones includeethereal diazomethane [41], trimethylsilyldiazomethane [42,123], diazomethane [63], pentafluorobenzyl bromide [83],HCl/methanol [122], N-methyl-N-(tert-butyldimethylsilyl)tri-fluoroacetamide (MTBSTFA) [147], N,O-bis(trimethylsilyl)trifluoroacetamide (BSTFA)/trimethylchlorosilane (TMCS)[148]. Compared with trifluoroacetylation, pentafluorobenzy-lation, methylation, and trimethylsilylation, the MTBSTFAderivatization protocol was the superior in the comprehensivemulti-targeted quantification of plant hormones in terms of theprospective sensitivity of MS, higher selectivity of detection,the purity of the reagent, and the stability of derivatives;however, the MTBSTFA method has low sensitivity forGAs and monosaccharide conjugates [147]. In contrast, ethe-real diazomethane was suitable for the derivatization of GAs[84]; therefore, diverse derivatization reagents were used forthe simultaneous determination and identification of multipleplant hormones [84, 123].
In addition, the choice of ionization method is also veryimportant in the GC–MS analysis of hormone compounds.Chemical ionization (CI) and electron ionization (EI) are twowidely used ionization methods for the GC–MS analysis ofplant hormones. Compared with EI, CI uses lower energy,affords less fragment ions, and usually yields an easily iden-tifiable pseudomolecular ion in the mass spectra. EI yields
64 F. Du et al.

more fragment ions and therefore a molecular fingerprint withstructural information. Schmelz et al. [42] present a facile,rapid, and inclusive analytical method that quantifies multipleplant hormones, phytotoxin coronatine (COR), and volatileorganic compounds (VOCs) simultaneously in small amountsof plant tissue by using GC–MS in the CImode. The proposedmethod was applied to study the changes in an array ofphysiological and ecological signals that accumulate in planttissues during abiotic and biotic stress, which illustrated thepotential and simplicity of this approach in quantifying sig-naling crosstalk interactions that occur at the level of synthesisand accumulation. The same GC–MS method was used forthe direct quantitative analysis of plant hormones and theirmethyl esters in plant extracts, and it is advantageous foranalyzing methyl ester and acid forms by a single preparationon a single chromatogram after silylation [148]. GC–MS in EImode was developed to simultaneously determine the concen-trations of IAA, ABA, JA, and dihydrojasmonic acid (DHJA)in plant tissues [65]. To better understand both auxin signalingand metabolic homeostasis, Barkawi et al. [146] recentlyreported a protocol for the precise, high-throughput determi-nation of free IAA in plant tissue by isotope dilution analysisusing GC–MS in EI mode. According to the protocol, theamount of tissue required is relatively small (25 mg of freshweight) and one can process more than 500 samples per weekusing an automated system. Compared with the HPLC–MS/MS method for the simultaneous determination of multipleclasses of plant hormones, GC–MS was disadvantageous interms of the determination coefficients of calibration curves,reproducible recoveries, limits of detection, sample prepara-tion, and sample throughput [57].
GC–MS/MS has been applied for the determination ofplant hormones [41, 42, 123]. Müller et al. [41] developed asensitive and accurate multiplex GC–MS/MS technique toquantify IAA, ABA, JA, 12-oxophytodienoic acid (OPDA),and SA in plant tissues and whole-plant organ-distributionmaps of multiple hormones with considerably higher spatialresolution and comparatively high temporal resolution for A.thaliana were generated. Acquisition of appropriate internalstandards and choice of derivatization agents are the primarylimiting factors in the expansion of GC and GC–MSmethods.
CE and CE–MS analysis
CE is a simple and effective alternative technology for thesample cleanup and separation of complex biological andchemical mixtures. CE with UV [87, 97, 149–153], laser-induced fluorescence detection (LIF) [60, 153–156], electro-chemiluminescent (ECL) [153, 157], and especially MSdetection [59, 99, 153, 158–162] provide a powerful toolfor the highly sensitive profiling of plant hormones and has
been applied for the simultaneous determination of multipleplant hormones and related metabolites.
Different CE modes, including micellar electrokinetic capil-lary chromatography (MECC), capillary-zone electrophoresis(CZE), and capillary electrochromatography (CEC), were usedfor the separation of plant hormones, and MEKC has betterseparation selectivity than CZE, because surfactant is added tobuffer solution to form micelles. However, the MEKC–UVmethod is unable to directly detect low amounts of plant hor-mones in plant tissues as a result of the low sensitivity of UVdetection. Therefore, MECC combined with some online con-centration techniques such as large volume sample stacking wasdeveloped to analyze multiple plant hormones including GA,ABA, IAA, NNA, 2,4-DA, kinetin-6-furfurylaminopurine, andN-benzyladenine in tobacco flowers [87] and CTKs in coconutwater [97, 163]. Compared with the conventional injectionmode, online concentration techniques achieved higher enrich-ment factors for these hormones and improved the sensitivity ofCE–UV [87, 149].
To enhance the selectivity and sensitivity of the CE meth-od, CE–LIF has been developed for the simultaneous deter-mination of individual or multiple classes of plant hormones.Prior to CE–LIF analysis, generally, plant hormones werederivatized with a fluorescent labeling reagent such as 6-oxy-(acetypiperazine) fluorescein [60], 8-aminopyrene-1,3,6-trisulfonate [154], and 5-bromomethylfluorescein [155]. Re-cently, Bai et al. [156] developed a novel CE–LIF methodbased on the in-capillary noncovalent labeling of quantumdots (QDs) for the determination of plant polypeptide hor-mones without expensive instrumentation, complicated pre-capillary derivatization, or large consumption of samples andreagents. The QDs-labeled TomSys was completely separatedfrom free QDs, which ensured the reliable quantitative mea-surement of TomSys. Moreover, this in-capillary labelingapproach was also compatible with large volume injection ofTomSys, resulting in the sample stacking and online concen-tration of the analytes.
Compared with CE–UV and CE–LIT methods, CE–MSexhibits higher selectivity and sensitivity in the analysis ofplant hormones, and it thus another technique which iscomplementary to the LC–MS method. However, the directcoupling of MEKC to MS is limited because of nonvolatilemicelles, which are detrimental to the MS signal. To resolvethis issue, a partial filling (PF) technique was developed forpreventing the nonvolatile micelles from entering the MSion source. A PF-MEKC directly coupled to MS (PF-MEKC–MS) method was proposed and has been utilizedto simultaneously separate and determine 13 structurallysimilar CTKs in banana pulp samples [160] and eight GAsin coconut water [161]. Recently, a sensitive PF-MEKC–QToF-MS method was developed for the simultaneousenantioseparation of all four JA stereoisomers, and discrim-ination of them in racemic JA without the need for a single
Analytical methods for tracing plant hormones 65

pure isomer [162]. To separate JA stereoisomers, the suc-cessive coating technique was applied by modifying thecapillary with multiple ionic polymer layers of polybrene–dextran sulfate–polybrene. The proposed method was thensuccessfully applied for the analysis of the naturally occur-ring JA stereoisomers in wounded tobacco leaves, and en-dogenous (−)-JA and (+)-epi-JA at different concentrationswere found.
CEC, which combines the efficiency of CE and the selec-tivity of LC, has also been developed to simultaneously analyzemultiple plant hormones by using an amino-coated capillary[59], monolithic silica-ODS capillary [149], Hypersil C18
packed capillary [150], or cationic polymer-coated capillary[159] to reverse electroosmotic flow. The pressurized capillaryelectrochromatography (pCEC) with UV method has beenproposed and successfully applied to the simultaneous analysisof multiple endogenetic and ectogenic plant hormones includ-ing auxin, ABA, GA, and CTKs [149, 150]. A simple, rapid,and sensitive CEC–MSmethod was proposed and validated forthe separation and simultaneous quantification of acidic planthormones [59, 159]. To improve the detection sensitivity ofacidic plant hormones, 3-bromoactonyltrimethylammoniumbromide (BTA) was used as a new mass probe in the CEC–ToF-MS method, and satisfactory identification and quantifica-tion of plant hormones were obtained [59]. In addition, a CZE–MS method was developed to analyze CTK nucleotides incoconut water under a volatile buffer system [99, 158]. Com-paredwithHPLC,CZE ismore suited for the direct separation ofcharged CTK nucleotides without any derivatization step [99].
Immuno-based methods
Techniques based on the detection of antibody–antigen (Ab–Ag) interaction, such as radioimmunoassay (RIA), ELISA,and SPA, have been widely used to develop plant hormonedetection methods, although the methods sometimes sufferfrom problems (e.g., complex sample preparation, cross-reactivity, imperfect validation, and variable results for plantsample analyses), and several review articles were publishedrecently [31, 32, 34].
ELISAs are available for the detection of major planthormones, including CTKs [37, 164–166], JAs [164, 167],ABA [164, 168–170], GAs [164, 171, 172], IAA [164, 173],24-epicastasterone, and related brassinolide analogs [74, 174],and some commercial ELISA kits are available. Comparedwith conventional indirect competitive ELISA (icELISA)methods, a simplified icELISA method, in which the analyte,antibody, and enzyme-labeled second antibody are added inone step, was established for the analysis of zeatin ribosideand two other small molecules, and offered several advantagesincluding being simpler, less time consuming, and more sen-sitive [165]. However, the results of Deng et al. [167] indicat-ed that direct competitive ELISA (dcELISA) was more
sensitive than either the conventional or simplified icELISAin the analysis of jasmonates in plants. Generally, the ELISAapproach is less sensitive than HPLC–MS methods for theanalyses of plant hormones from native plant tissues [74]. Inaddition, Lee et al. [172] developed a new detection methodfor bioactive gibberellin A4 (GA4) in living cells based onantibody VH/VL (heavy and light chains, respectively) inter-action, which make it feasible to realize the in situ and in vivodetection of plant hormones in living organisms. QD–anti-body (or aptamer) conjugates, which combine the specificityof antibodies or aptamers with the unique photophysical prop-erties and in vivo bioimaging capabilities of QDs [175, 176],have potential applications in the ultrasensitive, in situ, andreal-time detection of plant hormones in living organisms.
Besides the techniques mentioned above, biosensors arereceiving much attention owing to their unique propertiesand potential application in plant hormone determinations[177]. Immunosensors are biosensors that use antibodies orantigens as the specific sensing element and provideconcentration-dependent signals, and have been adaptedfor the detection of plant hormones. Electrochemical bio-sensors combine the sensitivity of electroanalytical methodswith the inherent bioselectivity of the biological component(e.g., antibody or aptamer), and are commonly used immu-nosensors. These immunosensors presented good specificityand high sensitivity and have been successfully applied tothe analysis of IAA [178, 179], ABA [180–182], CTKs [39,183, 184], and GA [185]. In addition, immobilization pro-tocols, biosensor regeneration, signal amplification, equip-ment miniaturization, and antibody (or aptamer) propertiesare considered in the development and applications of theseimmunosensors. For example, to develop an amperometricimmunosensor for ABA, an approximate 10-nm-thick goldlayer was first sputtered uniformly onto the electrode sur-face, and then gold nanoparticles were grown directly on thegold layer for antibody adsorption by immersing the elec-trode into H2AuCl4 solution [181], or an anti-ABA antibodywas adsorbed directly on a porous nanogold film [180].
Although immuno-based methods are excellent for thedetection of plant hormones because of their high selectivityand sensitivity, the specificity of antibodies restricts theapplications of the method in the simultaneous detection ofmultiple classes plant hormones. On the other hand, becausethe immuno-based methods are less dependent on largeequipment and are more amenable to miniaturization, theyare more portable and suitable for in situ screening of targethormones in plant tissues [34].
Other analytical methods
Electrochemical techniques are suitable for the analysis ofplant hormones because of their simplicity, convenience, and
66 F. Du et al.

low cost. Several electrochemical techniques have been devel-oped for the quantitative determination of IAA in plant tissuesby using electrodes modified with nano-Au/multi-walled car-bon nanotubes (MWNTs)/chitosan [186], CeCl3/dihexadecylhydrogen phosphate [187], or reduced graphene oxide/poly(safranine T) [188]. Gan et al. [189, 190] designed aMeJA electrochemical sensor based on phosphotungsticacid/graphene oxide (PTA/GO) nanohybrid or nano-montmorillonite. Compared with common gold electrodes,the electrodesmodifiedwith copper nanoparticles could detectSA at a higher sensitivity [191]. Recently, Kowalska et al.[192] designed an electrochemical biosensor for the detectionof the plant hormone CTK based on the interactions betweenCTK dehydrogenase (CKX) and the N6-side chain of CTKs.Prussian Blue (PrB) was electrodeposited on Pt microelectr-odes prior to deposition of the purified CKX enzyme entrap-ment in a sol–gel matrix. The prepared biosensor wassensitive to micromolar concentrations of several CTKs.
With the advancement of electrochemical techniques, elec-trochemical biosensors are increasingly being investigated toperform in situ, real-time measurements of plant hormones.McLamore et al. [193] developed a novel self-referencingelectrochemical microsensor to noninvasively monitor in vivoIAAmovement near the surface of Zea mays roots without theaddition of exogenous IAA. Enhanced sensor surface modifi-cation, decoupling of acquired signals, and integrated fluxanalyses were combined to provide direct, noninvasive, real-time quantification of endogenous IAA movement in B73maize inbred and brachytic2 (br2) auxin transport mutantroots, which suggested that the proposed technique is usefulfor in situ, real-time, in vivo monitoring of plant hormones insurface tissues and for better understanding the biochemicalmechanisms of plant hormones involved in plant growth anddevelopment. In addition, Napier’s research group [194] hasalso developed surface plasmon resonance (SPR) as a genericimmunosensor platform for real-time plant hormone sensing,testing it for both ABA and auxin.
In addition, some spectral methods have been employedfor the determination of plant hormones in plant tissues. Anexcitation–emission matrix fluorescence coupled to asecond-order calibration method was proposed to simulta-neously detect ABA and GA in ginkgo leaves and buds[195], and to directly determine IAA in soil [196]. Visible/shortwave near-infrared (Vis/SW NIR) spectroscopy com-bined with some chemometrics methods including partialleast-square regression (PLSR) and stepwise multiple linearregression (SMLR) was developed as a nondestructive andrapid tool for the determination of ethylene content in to-matoes [197]. Guo et al. [198] described a colorimetricmethod for the selective detection and differentiation ofIAA and IBA. From the color changes of IAA and IBAafter Ehrlich reaction, the identification of auxin at themicromolar level can be achieved even with the naked eye.
Moreover, in situ imaging techniques are desired for thedetection of plant hormones. The distribution of auxin in theroot cap was visualized by in situ imaging with fluorescencesignal from green fluorescent protein (GFP) [199]. Becausethe growth-related pH dynamics were dependent on auxin,and specifically on auxin transport mediated by the auxininflux carrier AUX1 in cells of the lateral root cap and rootepidermis [200], the proposed approach that integrated con-focal microscopy and automated computer vision-based im-age analysis could be used to detect auxin in plant tissues.
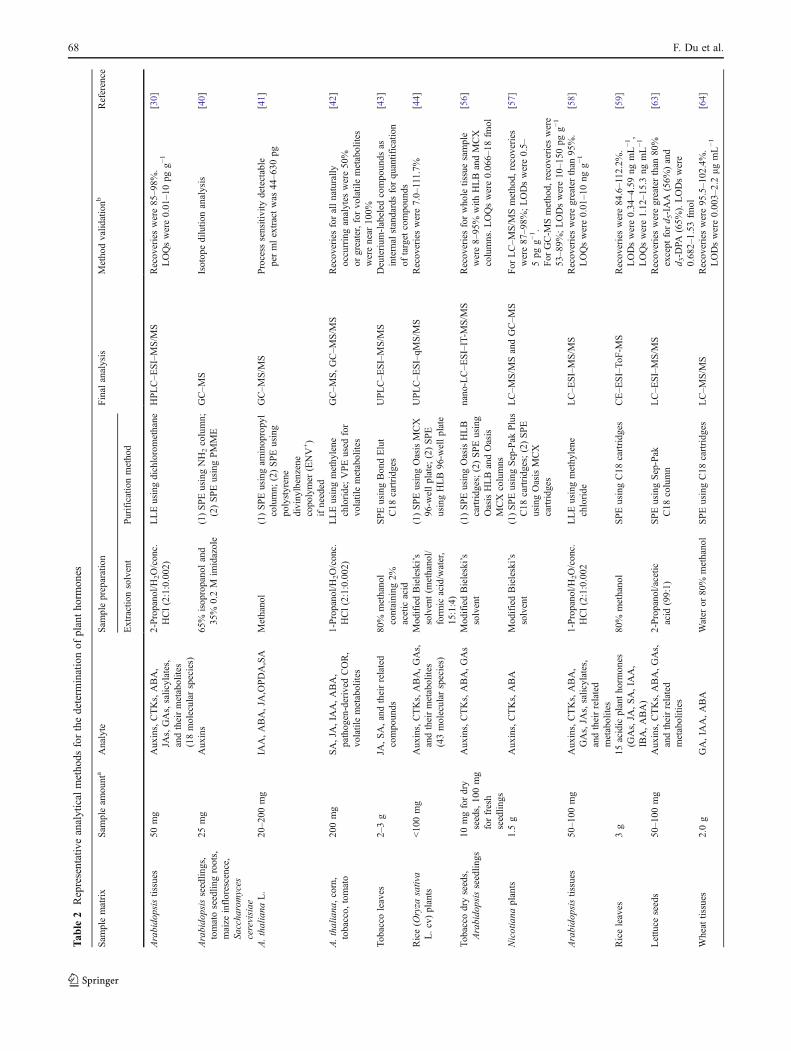
Table 2 presents a summary of representative analyticalmethods for the determination of plant hormones.
Applications
During the past 10 years, our knowledge of the molecularmechanisms of plant hormones, including hormone biosyn-thesis, metabolism, transport, perception, signaling, and re-sponse, has grown exponentially [2–8, 102, 201], which isinvaluable for modern agriculture [11–13]. Modern analyticalmethods provide a new detection platform for the study ofplant hormones.
To elucidate the possible biosynthetic pathways of isopren-oid CTKs in plants, Liang et al. [121] developed protocols forthe extraction, purification, and detection of isoprenoid CTKsin plant tissues based on mixed-mode SPE pre-cleanup, IAE,and LC–QToF-MS detection, which allowed simultaneousquantification of eight endogenous isoprenoid CTKs in A.thaliana leaves. The LC–QToF-MS method was also appliedto detect IAA–myo-inositol in roots of Arabidopsis, and theresults showed that four IAA–myo-inositol isomers existedboth in Arabidopsis roots and siliques, whereas two isomersexisted in maize kernels [202]. The analysis of data fromIAA–myo-inositol and free IAA provided evidence for thestudy of the essential role of free myo-inositol in cotyledondevelopment. LC–QToF-MS has also been applied to studythe methylation of GAs in plants [203].
In recent years, LC–ESI–MS/MS has been widely appliedin the study of molecular mechanisms of plant hormones [33,46, 47, 204–206]. To investigate the role of plant autophagyand SA signaling under nutrient-rich conditions, the methodbased on the combination of SPE with LC–ESI–MS/MS(SPE–LC–ESI–MS/MS) was adopted to measure the levelsof endogenous levels of SA, IAA, ABA, JA, JA-Ile, GA1, andGA4 in wild type and autophagy defective mutants and then tostudy the related roles of SA signaling [204]. The obtainedresults implied that plant autophagy operates a novel negativefeedback loop modulating SA signaling to negatively regulatesenescence and immunity-related programmed cell death. Thedeveloped SPE–LC–ESI–MS/MS method [205] was used toanalyze GAs, ABA, IAA, iP, tZ, DHZ, JA, JA-Ile, and SA,and the UPLC–QToF-MS method [206] was selected to
Analytical methods for tracing plant hormones 67
Tab
le2
Representativeanalytical
metho
dsforthedeterm
inationof
plantho
rmon
es
Sam
plematrix
Sam
pleam
ount
aAnalyte
Sam
plepreparation
Final
analysis
Methodvalid
ationb
Reference
Extractionsolvent
Purificationmethod
Arabidopsistissues
50mg
Auxins,CTKs,ABA,
JAs,GAs,salicylates,
andtheirmetabolites
(18molecular
species)
2-Propanol/H
2O/conc.
HCl(2:1:0.002)
LLEusingdichloromethane
HPLC–ESI–MS/M
SRecoverieswere85
–98%.
LOQswere0.01
–10
pgg−
1[30]
Arabidopsisseedlin
gs,
tomatoseedlin
groots,
maize
inflorescence,
Saccharomyces
cerevisiae
25mg
Auxins
65%
isopropanoland
35%
0.2M
imidazole
(1)SPEusingNH2column;
(2)SPEusingPMME
GC–MS
Isotopedilutio
nanalysis
[40]
A.thaliana
L.
20–200mg
IAA,ABA,JA
,OPDA,SA
Methanol
(1)SPEusingam
inopropyl
column;
(2)SPEusing
polystyrene
divinylbenzene
copolymer
(ENV+)
ifneeded
GC–MS/M
SProcess
sensitivity
detectable
permlextractwas
44–630pg
[41]
A.thaliana
,corn,
tobacco,
tomato
200mg
SA,JA
,IA
A,ABA,
pathogen-derived
COR,
volatilemetabolites
1-Propanol/H
2O/conc.
HCl(2:1:0.002)
LLEusingmethylene
chloride;VPEused
for
volatilemetabolites
GC–MS,GC–MS/M
SRecoveriesforallnaturally
occurringanalytes
were50%
orgreater,forvolatilemetabolites
werenear
100%
[42]
Tobaccoleaves
2–3g
JA,SA,andtheirrelated
compounds
80%
methanol
containing
2%acetic
acid
SPEusingBondElut
C18
cartridges
UPLC–ESI–MS/M
SDeuterium
-labeled
compounds
asinternal
standardsforquantification
oftarget
compounds
[43]
Rice(O
ryza
sativa
L.cv)plants
<100mg
Auxins,CTKs,ABA,GAs,
andtheirmetabolites
(43molecular
species)
ModifiedBieleski’s
solvent(m
ethanol/
form
icacid/water,
15:1:4)
(1)SPEusingOasisMCX
96-w
ellplate;
(2)SPE
usingHLB96-w
ellplate
UPLC–ESI–qM
S/M
SRecoverieswere7.0–
111.7%
[44]
Tobaccodryseeds,
Arabidopsisseedlin
gs10
mgfordry
seeds,100mg
forfresh
seedlin
gs
Auxins,CTKs,ABA,GAs
ModifiedBieleski’s
solvent
(1)SPEusingOasisHLB
cartridges;(2)SPEusing
OasisHLBandOasis
MCX
columns
nano-LC–ESI–IT-M
S/M
SRecoveriesforwhole
tissuesample
were8–
95%
with
HLBandMCX
columns.LOQswere0.066–
18fm
ol
[56]
Nicotiana
plants
1.5g
Auxins,CTKs,ABA
ModifiedBieleski’s
solvent
(1)SPEusingSep-Pak
Plus
C18
cartridges;(2)SPE
usingOasisMCX
cartridges
LC–MS/M
SandGC–MS
For
LC–MS/M
Smethod,
recoveries
were87
–98%;LODswere0.5–
5pg
g−1.
For
GC-M
Smethod,
recoveries
were
53–89%;LODswere10–150pg
g−1
[57]
Arabidopsistissues
50–100mg
Auxins,CTKs,ABA,
GAs,JA
s,salicylates,
andtheirrelated
metabolites
1-Propanol/H
2O/conc.
HCl(2:1:0.002
LLEusingmethylene
chloride
LC–ESI–MS/M
SRecoveriesweregreaterthan
95%.
LOQswere0.01
–10
ngg−
1[58]
Riceleaves
3g
15acidic
planthorm
ones
(GAs,JA
,SA,IA
A,
IBA,ABA)
80%
methanol
SPEusingC18
cartridges
CE–ESI–To
F-M
SRecoverieswere84.6–112.2%
.LODswere0.34
–4.59
ngmL−1,
LOQswere1.12
–15.3
ngmL−1
[59]
Lettuce
seeds
50–100mg
Auxins,CTKs,ABA,GAs,
andtheirrelated
metabolities
2-Propanol/acetic
acid
(99:1)
SPEusingSep-Pak
C18
column
LC–ESI–MS/M
SRecoveriesweregreaterthan
80%
except
ford 5-IAA
(56%
)and
d 3-D
PA(65%
).LODswere
0.682–
1.53
fmol
[63]
Wheat
tissues
2.0g
GA,IA
A,ABA
Water
or80%
methanol
SPEusingC18
cartridges
LC–MS/M
SRecoverieswere95.5–102.4%
.LODswere0.003–
2.2μg
mL−1
[64]
68 F. Du et al.
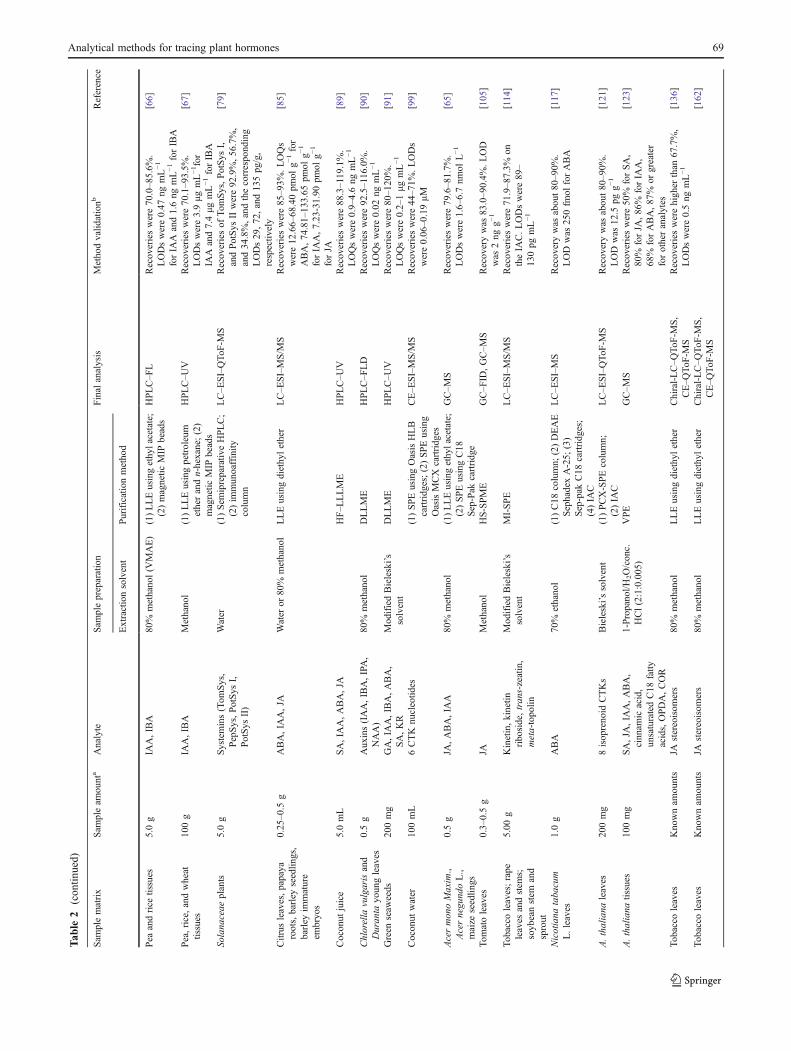
Tab
le2
(con
tinued)
Sam
plematrix
Sam
pleam
ount
aAnalyte
Sam
plepreparation
Final
analysis
Methodvalid
ationb
Reference
Extractionsolvent
Purificationmethod
Pea
andrice
tissues
5.0g
IAA,IBA
80%
methanol(V
MAE)
(1)LLEusingethylacetate;
(2)magnetic
MIP
beads
HPLC–FL
Recoverieswere70.0–85.6%.
LODswere0.47
ngmL−1
forIA
Aand1.6ng
mL−1forIBA
[66]
Pea,rice,andwheat
tissues
100g
IAA,IBA
Methanol
(1)LLEusingpetroleum
etherandn-hexane;(2)
magnetic
MIP
beads
HPLC–UV
Recoverieswere70.1–93.5%.
LODswere3.9μg
mL−1for
IAA
and7.4μg
mL−1forIBA
[67]
Solanaceae
plants
5.0g
Systemins(Tom
Sys,
PepSys,PotSys
I,PotSys
II)
Water
(1)Sem
ipreparativ
eHPLC;
(2)im
munoaffinity
column
LC–ESI–QToF
-MS
Recoveriesof
Tom
Sys,PotSys
I,andPotSys
IIwere92.9%,56.7%,
and34.8%,andthecorresponding
LODs29,72,and135pg/g,
respectiv
ely
[79]
Citrus
leaves,papaya
roots,barley
seedlin
gs,
barley
immature
embryos
0.25
–0.5g
ABA,IA
A,JA
Water
or80%
methanol
LLEusingdiethylether
LC–ESI–MS/M
SRecoverieswere85
–93%.LOQs
were12.66–
68.40pm
olg−
1for
ABA,74.81–
133.65
pmol
g−1
forIA
A,7.23-31.90
pmol
g−1
forJA
[85]
Coconut
juice
5.0mL
SA,IA
A,ABA,JA
HF–LLLME
HPLC–UV
Recoverieswere88.3–119.1%
.LOQswere0.9–
4.6ng
mL−1
[89]
Chlorella
vulgarisand
Duranta
youngleaves
0.5g
Auxins(IAA,IBA,IPA,
NAA)
80%
methanol
DLLME
HPLC–FLD
Recoverieswere92.5–116.0%
.LOQswere0.02
ngmL−1
[90]
Green
seaw
eeds
200mg
GA,IA
A,IBA,ABA,
SA,KR
ModifiedBieleski’s
solvent
DLLME
HPLC–UV
Recoverieswere80
–120%
.LOQswere0.2–
1μg
mL−1
[91]
Coconut
water
100mL
6CTK
nucleotid
es(1)SPEusingOasisHLB
cartridges;(2)SPEusing
OasisMCX
cartridges
CE–ESI–MS/M
SRecoverieswere44
–71%.LODs
were0.06
–0.19
μM[99]
AcermonoMaxim.,
AcernegundoL.,
maize
seedlin
gs
0.5g
JA,ABA,IA
A80%
methanol
(1)LLEusingethylacetate;
(2)SPEusingC18
Sep-Pak
cartridge
GC–MS
Recoverieswere79.6–81.7%.
LODswere1.6–
6.7nm
olL−1
[65]
Tomatoleaves
0.3–
0.5g
JAMethanol
HS-SPME
GC–FID
,GC–MS
Recoverywas
83.0–90.4%.LOD
was
2ng
g−1
[105
]
Tobaccoleaves;rape
leaves
andstem
s;soybeanstem
and
sprout
5.00
gKinetin,kinetin
riboside,trans-zeatin,
meta-topolin
ModifiedBieleski’s
solvent
MI-SPE
LC–ESI–MS/M
SRecoverieswere71.9–87.3%
ontheIA
C.LODswere89
–
130pg
mL−1
[114
]
Nicotiana
tabacum
L.leaves
1.0g
ABA
70%
ethanol
(1)C18
column;
(2)DEAE
SephadexA-25;
(3)
Sep-pak
C18
cartridges;
(4)IA
C
LC–ESI–MS
Recoverywas
about80
–90%.
LOD
was
250fm
olforABA
[117
]
A.thaliana
leaves
200mg
8isoprenoid
CTKs
Bieleski’ssolvent
(1)PCX-SPEcolumn;
(2)IA
CLC–ESI–QToF
-MS
Recoverywas
about80
–90%.
LOD
was
12.5
pgg−
1[121
]
A.thaliana
tissues
100mg
SA,JA
,IA
A,ABA,
cinnam
icacid,
unsaturatedC18
fatty
acids,OPDA,COR
1-Propanol/H
2O/conc.
HCl(2:1:0.005)
VPE
GC–MS
Recoverieswere50%
forSA,
80%
forJA
,86%
forIA
A,
68%
forABA,87%
orgreater
forotheranalytes
[123
]
Tobaccoleaves
Knownam
ounts
JAstereoisom
ers
80%
methanol
LLEusingdiethylether
Chiral-LC–QToF
-MS,
CE–QToF
-MS
Recoverieswerehigher
than
67.7%,
LODswere0.5ng
mL−1
[136
]
Tobaccoleaves
Knownam
ounts
JAstereoisom
ers
80%
methanol
LLEusingdiethylether
Chiral-LC–QToF
-MS,
CE–QToF
-MS
[162
]
Analytical methods for tracing plant hormones 69
analyze ABA–GE in different tissues over the course of seeddevelopment. The obtained data suggested that ABA metab-olism depends on developmental stages and tissues, and thatABA interacts with other hormones to regulate seed develop-mental processes [205]. In addition, Koo et al. [207] investi-gated the levels JA-Ile and 12OH-JA-Ile inArabidopsis leavesby LC–MS/MS before and after wounding, which was helpfulto elucidate the mediation roles of cytochrome P450CYP94B3 on catabolism and inactivation of JA-Ile.
To investigate the spatial and temporal changes of planthormones’ response to biotic and abiotic stresses, the meth-od based on SPE pre-cleanup, IAC purification, and LC–MSanalysis (SPE–IAC–LC–MS) was applied [208–210]. Stirket al. [209] investigated the changes in CTKs and auxinconcentrations in seaweed concentrates when stored at anelevated temperature by using SPE–IAC–LC–MS. Theobtained results showed that the CTKs were found to in-crease with storage, whereas the auxins were less stable,decreasing with storage. The same method was also used toidentify and quantify endogenous CTKs in intact pea seed-ling roots and then study the spatial and temporal changes ofCTKs in developing pea roots, which may provide insightsinto the role of the various CTKs during germination andseedling establishment [210]. Moreover, an SPE–UPLC–ESI–qMS/MS method was used to study the organ distribu-tions of hormone species in rice and to identify interactionsamong the four major hormones in the rice GA signalingmutants, gid1-3, gid2-1, and slr1 [44]. The hormone profil-ing data and transcriptome data in the GA signaling mutantsobtained by the SPE–UPLC–ESI–qMS/MS method allowedresearchers to analyze relationships between changes in geneexpression and hormone metabolism, and to discover novelgenes involved in regulating plant hormone action and me-tabolism. In fact, other developed analytical methods (e.g.,GC–MS) have been applied in the study of plant hormones[211–213].
Conclusion and trends
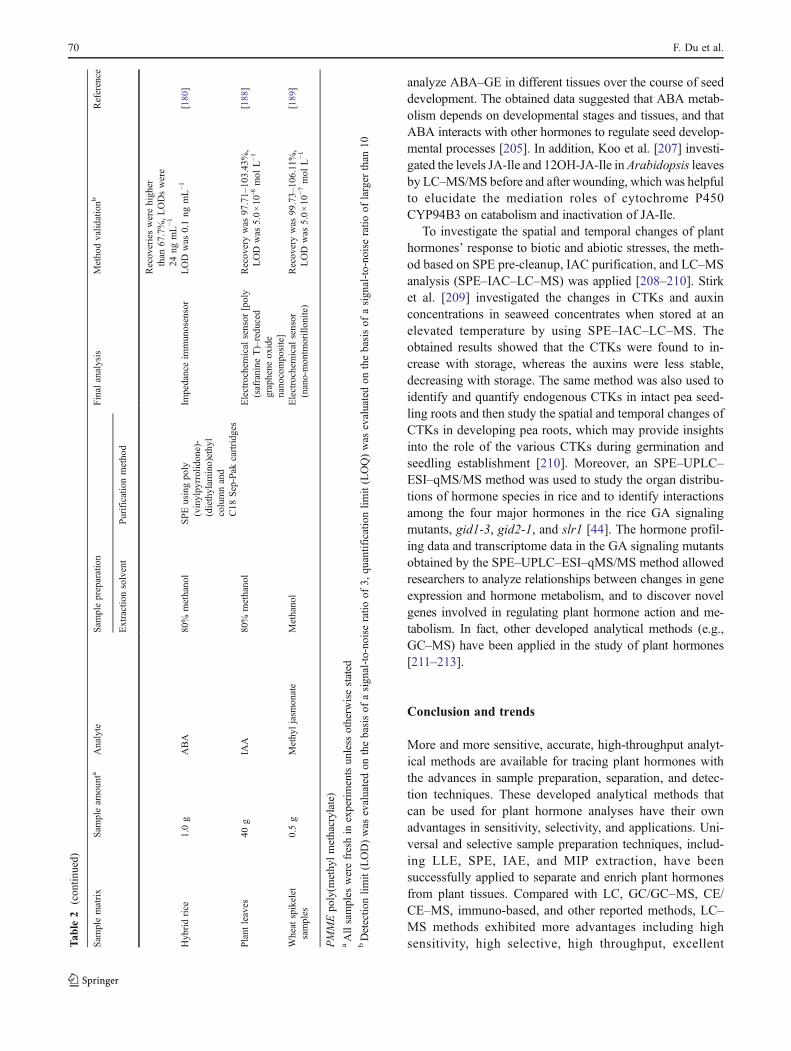
More and more sensitive, accurate, high-throughput analyt-ical methods are available for tracing plant hormones withthe advances in sample preparation, separation, and detec-tion techniques. These developed analytical methods thatcan be used for plant hormone analyses have their ownadvantages in sensitivity, selectivity, and applications. Uni-versal and selective sample preparation techniques, includ-ing LLE, SPE, IAE, and MIP extraction, have beensuccessfully applied to separate and enrich plant hormonesfrom plant tissues. Compared with LC, GC/GC–MS, CE/CE–MS, immuno-based, and other reported methods, LC–MS methods exhibited more advantages including highsensitivity, high selective, high throughput, excellentT
able
2(con
tinued)
Sam
plematrix
Sam
pleam
ount
aAnalyte
Sam
plepreparation
Final
analysis
Methodvalid
ationb
Reference
Extractionsolvent
Purificationmethod
Recoverieswerehigher
than
67.7%,LODswere
24ng
mL−1
Hybridrice
1.0g
ABA
80%
methanol
SPEusingpoly
(vinylpyrrolidone)-
(diethylam
ino)ethyl
columnand
C18
Sep-Pak
cartridges
Impedanceim
munosensor
LOD
was
0.1ng
mL−1
[180
]
Plant
leaves
40g
IAA
80%
methanol
Electrochem
ical
sensor
[poly
(safranine
T)–reduced
graphene
oxide
nanocomposite]
Recoverywas
97.71–
103.43%,
LOD
was
5.0×10
-8mol
L−1
[188
]
Wheat
spikelet
samples
0.5g
Methyljasm
onate
Methanol
Electrochem
ical
sensor
(nano-montm
orillonite)
Recoverywas
99.73–
106.11%,
LOD
was
5.0×10
−7mol
L−1
[189
]
PMMEpo
ly(m
ethy
lmethacrylate)
aAllsamples
werefreshin
experimentsun
less
otherw
isestated
bDetectio
nlim
it(LOD)was
evaluatedon
thebasisof
asign
al-to-no
iseratio
of3,
quantificationlim
it(LOQ)was
evaluatedon
thebasisof
asign
al-to-noiseratio
oflarger
than
10
70 F. Du et al.

accuracy, and favorable precision, and can potentially be anextremely useful approach in achieving comprehensivequantitative plant hormone profiles. Electrochemical bio-sensors and in situ imaging techniques have also attractedconsiderable attention in the detection of plant hormones.
To better elucidate and understand molecular mechanismsof plant hormones in plants, an ideal analytical method isdesired to simultaneously investigate the concentrations,diversifications, and spacial and temporal distributions ofmultiple—even all classes of—plant hormones (hormone pro-files, hormone–metabolome or hormonome [44]) and therelated metabolites in given sample in a single experiment,and to discover more new plant hormones. This issue may beaddressed in the future, though more challenges still remain inseveral aspects. The main trends and challenges in the devel-opment of analytical methods for studying plant hormones arelisted below:
& To develop highly sensitive and high-throughput methodsfor monitoring plant hormone profiles. Plant hormoneshave different chemical and structural properties, and theydo not act in isolation but are highly interrelated by syn-ergistic or antagonistic crosstalk so that they modulateeach other’s biosynthesis or signaling. Therefore, it isnecessary to develop a comprehensive analytical methodfor monitoring all classes of plant hormones, along withtheir metabolites. According to the results of Pan et al.[30], Schmelz et al. [42], Kojima et al. [44], and Izumi etal. [56], the greatest challenge for the simultaneous deter-mination of multiple classes of plant hormones is to de-velop simple, rapid, effective, and universal samplepreparation techniques.
& To develop automatic, miniature, and integrated analyt-ical platforms for high-efficiency and high-throughputseparation and identification of multiple classes of planthormones. Most analytical methods at present are notautomatic and integrated methods. The development ofautomated sample preparation techniques [44, 146] andthe improvement of the coupling of sampling and detec-tion techniques are still challenges.
& To develop ultrasensitive, noninvasive, in vivo, in situ, andreal-time detection methods for tracing plant hormones.The concentrations of plant hormones are very low, usuallyat the nano- to femtomolar level. Furthermore, the concen-trations, diversifications, and distributions of plant hor-mones in plant samples are variable in terms of plantgrowth, development, and response to stresses. So, in situsampling techniques should avoid wounding plant samplesas little as possible, and new analytical instruments shouldhave the capability of ultrasensitive, real-time, and in situdetection. In vivo SPME is an alternative technique for thenondestructive and on-site sampling of plant tissues [214,215]. Electrochemicalmicro- (or nano-)probes [177], SPR-
based biosensors [194, 216, 217], in situ MS imagingtechniques [218], and atomic and molecular MS [219]are promising techniques for the in vivo, in situ, and real-time detection of plant hormones.
Acknowledgements This work was supported by the National NaturalScience Foundation of China (Grants 20905005 and 21065003), by theChina Postdoctoral Science Foundation (Grant 201003008), and by theDoctoral Research Foundation of Guilin University of Technology (Grant002401003314).
References
1. Teale WD, Paponov IA, Palme K (2006) Nat Rev Mol Cell Biol7:847–859
2. Santner A, Estelle M (2009) Nature 459:1071–10783. Bari R, Jones JDG (2009) Plant Mol Biol 69:473–4884. Santner A, Calderon-Villalobos LIA, Estelle M (2009) Nat Chem
Biol 5:301–3075. Pieterse CMJ, Leon-Reyes A, der Ent SV, Van Wees SCM (2009)
Nat Chem Biol 5:308–3166. Sánchez-Rodríguez C, Rubio-Somoza I, Sibout R, Persson S
(2010) Trends Plant Sci 15:291–3017. Davies PJ (2010) Plant hormones: biosynthesis, signal transduc-
tion, action, 3rd edn. Springer, Berlin, pp 717–7408. Peleg Z, Blumwald E (2011) Curr Opin Plant Biol 14:290–2959. Peng J, Richards DE, Hartley NM, Murphy GP, Devos KM,
Flintham JE, Beales J, Fish LJ, Worland AJ, Pelica F, SudhakarD, Christou P, Snape JW, Gale MD, Harberd NP (1999) Nature400:256–261
10. Spielmeyer W, Ellis MH, Chandler PM (2002) Proc Nat Acad SciUSA 99:9043–9048
11. Ashikari M, Sakakibara H, Lin SY, Yamamoto T, Takashi T,Nishimura A, Angeles ER, Qian Q, Kitano H, Matsuoka M(2005) Science 309:741–745
12. Wu CY, Trieu A, Radhakrishnan P, Kwok SF, Harris S, Zhang K,Wang JL, Wan JM, Zhai HQ, Takatsuto S, Matsumoto S, FujiokaS, Feldmann KA, Pennell RI (2008) Plant Cell 20:2130–2145
13. Divi UK, Krishna P (2009) New Biotechnol 26:131–13614. McSteen P, Zhao YD (2008) Dev Cell 14:467–47315. Zhao YD (2010) Annu Rev Plant Biol 61:49–6416. Lindsey K, Casson S, Chilley P (2002) Trends Plant Sci 7:78–8317. Bahyrycz A, Konopińska D (2007) J Pept Sci 13:787–79718. Flescher E (2007) Cancer Lett 245:1–1019. Malíková J, Swaczynová J, Kolář Z, Strnad M (2008) Phytochem
69:418–42620. Cohen S, Flescher E (2009) Phytochem 70:1600–160921. Voller J, Zatloukal M, Lenobel R, Doležal K, Béreš T, Kryštof V,
Spíchal L, Niemann P, Džubák P, Hajdúch M, Strnad M (2010)Phytochem 71:1350–1359
22. Bassaganya-Riera J, Skoneczka J, Kingston DGJ, Krishnan A,Misyak SA, Guri AJ, Pereira A, Carter AB, Minorsky P, TumarkinR, Hontecillas R (2010) Curr Med Chem 17:467–478
23. Li HH, Hao RL, Wu SS, Guo PC, Chen CJ, Pan LP, Ni H (2011)Biochem Pharmacol 82:701–712
24. Pearce G, Moura DS, Stratmann J, Ryan CA (2001) Proc NatlAcad Sci USA 98:12843–12847
25. Pearce G, Siems WF, Bhattacharya R, Chen YC, Ryan CA (2007)J Biol Chem 282:17777–17784
26. Robert-Seilaniantz A, Grant M, Jones JDG (2011) Annu RevPhytopathol 49:317–343
Analytical methods for tracing plant hormones 71

27. Argueso CT, Raines T, Kieber JJ (2010) Curr Opin Plant Biol13:533–539
28. Ross JJ, Weston DE, Davidson SE, Reid JB (2011) Physiol Plant141:299–309
29. Simon S, Petrášek J (2011) Plant Sci 180:454–46030. Pan XQ, Welti R, Wang XM (2010) Nat Protoc 5:986–99231. Tarkowski P, Ge LY, Yong JWH, Tan SN (2009) TrAC Trends
Anal Chem 28:323–33532. Liang Y, Zhao MP, Liu HW (2009) Chinese J Anal Chem
37:1232–123933. Pan XQ, Wang XM (2009) J Chromatogr B 877:2806–281334. Bai Y, Du FY, Bai Y, Liu HW (2010) Anal Methods 2:1867–187335. Fu JH, Sun XH, Wang JD, Chu JF, Yan CY (2011) Chinese Sci
Bull 56:355–36636. Volksch B, Bublitz F, FritscheW (1989) J Basic Microb 29:463–46837. Weiler EW (1984) Ann Rev Plant Physiol 35:85–9538. Hedden P (1993) Ann Rev Plant Physiol Plant Mol Biol 44:107–
12939. Székács A, Hegedűs G, Tóbiás I, Pogány M, Barna B (2000)
Anal Chim Acta 421:135–14640. Barkawi LS, Tam YY, Tillman JA, Normanly J, Cohen JD (2010)
Nat Protoc 5:1109–111841. Müller A, Düchting P, Weiler EW (2002) Planta 216:44–5642. Schmelz EA, Engelberth J, Alborn HT, O’Donnell P, Sammons
M, Toshima H, Tumlinson JH III (2003) Proc Natl Acad Sci USA100:10552–10557
43. Matsuura H, Aoi A, Satou C, Nakaya M, Masuta C, Nabeta K(2009) Plant Growth Regul 57:293–301
44. Kojima M, Kamada-Nobusada T, Komatsu H, Takei K, Kuroha T,Mizutani M, Ashikari M, Ueguchi-Tanaka M, Matsuoka M,Suzuki K, Sakakibara H (2009) Plant Cell Physiol 50:1201–1214
45. Garnica M, Houdusse F, Zamarreño AM, Garcia-Mina JM (2010)J Plant Physiol 167:1264–1272
46. Nishiyama R, Watanabe Y, Fujita Y, Le DT, Kojima M, Werner T,Vankova R, Yamaguchi-Shinozaki K, Shinozaki K, Kakimoto T,Sakakibara H, Schmülling T, Tran LSP (2011) Plant Cell23:2169–2183
47. Lackman P, González-GuzmánM, Tilleman S, Carqueijeiro I, PérezAC, Moses T, Seo M, Kanno Y, Häkkinen ST, Van Montagu MCE,Thevelein JM, Maaheimo H, Oksman-Caldentey KM, RodriguezPL, Rischer H, Goossens A (2011) Proc Natl Acad Sci USA108:5891–5896
48. Ridgway K, Lalljie SPD, Smith RM (2007) J Chromatogr A1153:36–53
49. Tobiszewski M, Mechlińska A, Zygmunt B, Namieśnik J (2009)TrAC Trends Anal Chem 28:943–951
50. Chen Y, Guo ZP, Wang XY, Qiu CG (2008) J Chromatogr A1184:191–219
51. Zygmunt B, Namiesnik J (2003) J Chromatogr Sci 41:109–11652. Romanik G, Gilgenast E, Przyjazny A, Kamiński M (2007) J
Biochem Biophys Methods 70:253–26153. Alcázar R, Parker JE (2011) Trends Plant Sci. doi:10.1016/j.
tplants.2011.09.00154. Qaderi MM, Kurepin LV, Reid DM (2012) Environ Exp Bot
75:107–11355. Susawaengsup C, Rayanakorn M, Wongpornchai S, Wangkarn S
(2011) Talanta 85:897–90556. Izumi Y, Okazawa A, Bamba T, Kobayashi A, Fukusaki E (2009)
Anal Chim Acta 648:215–22557. Giannarelli S, Muscatello B, Bogani P, Spiriti MM, Buiatti M,
Fuoco R (2010) Anal Biochem 398:60–6858. Pan XQ, Welti R, Wang XM (2008) Phytochem 69:1773–178159. Chen ML, Huang YQ, Liu JQ, Yuan BF, Feng YQ (2011) J
Chromatogr B 879:938–94460. Chen H, Guo XF, Zhang HS, Wang H (2011) J Chromatogr B
879:1802–1808
61. Bajguz A, Piotrowska A (2009) Phytochem 70:957–96962. Vine JH, Noiton D, Plummer JA, Baleriola-Lucas C, Mullins MG
(1987) Plant Physiol 85:419–42263. Chiwocha SDS, Abrams SR, Ambrose SJ, Cutler AJ, Loewen M,
Ross ARS, Kermode AR (2003) Plant J 35:405–41764. Hou SJ, Zhu J, Ding MY, Lv GH (2008) Talanta 76:798–80265. Zhang FJ, Jin YJ, Xu XY, Lu RC, Chen HJ (2008) Phytochem
Anal 19:560–56766. Hu YL, Li YW, Zhang Y, Li GK, Chen YQ (2011) Anal Bioanal
Chem 399:3367–337467. Zhang Y, Li YW, Hu YL, Li GK, Chen YQ (2010) J Chromatogr
A 1217:7337–734468. Sorce C, Picciarelli P, Calistri G, Lercari B, Ceccarelli N (2008) J
Plant Physiol 165:482–48969. Ribnicky DM, Cohen JD, Hu WS, Cooke TJ (2002) Planta
214:505–50970. Barkawi LS, Tam YY, Tillman JA, Pederson B, Calio J, Al-Amier
H, Emerick M, Normanly J, Cohen JD (2008) Anal Biochem372:177–188
71. Absalan G, Akhond M, Sheikhian L (2008) Talanta 77:407–41172. Hoyerová K, Gaudinová A, Malbeck J, Dobrev PI, Kocábek T,
Blanka Š, Trávníčková A, Kamínek M (2006) Phytochem67:1151–1159
73. Gupta D, Bhardwaj R, Nagar PK, Kaur S (2004) Plant GrowthRegul 43:97–100
74. Swaczynová J, Novák O, Hauserová E, Fuksová K, Šíša M,Kohout L, Strnad M (2007) J Plant Growth Regul 26:1–14
75. Sondhi N, Bhardwaj R, Kaur S, Kumar N, Singh B (2008) PlantGrowth Regul 54:217–224
76. Bajguz A (2009) J Plant Physiol 166:1946–194977. Pearce G, Strydom D, Johnson S, Ryan CA (1991) Science
253:895–89778. Constabel CP, Yip L, Ryan CA (1998) Plant Mol Biol 36:55–6279. Du FY, Bai Y, Bai Y, Liu HW (2010) Anal Chem 82:9374–938380. Pearce G, Moura DS, Stratman J, Ryan CA (2001) Nature
411:817–82081. Pearce G, Bhattacharya R, Chen YC, Barona G, Yamaguchi Y,
Ryan CA (2009) Plant Physiol 150:1422–143382. Smith RM (2003) J Chromatogr A 1000:3–2783. Duffield PH, Netting AG (2001) Anal Biochem 289:251–25984. Blake PS, Taylor JM, Finch-Savage WE (2002) Plant Growth
Regul 37:119–12585. Durgbanshi A, Arbona V, Pozo O, Miersch O, Sancho JV, Mez-
Cadenas AG (2005) J Agric Food Chem 53:8437–844286. PacákováV, Štulík K, VlasákováV, BřezinováA (1997) J Chromatogr
A 764:331–33587. Liu BF, Zhong XH, Lu YT (2002) J Chromatogr A 945:257–26588. Xie W, Han C, Zheng ZQ, Chen XM, Qian Y, Ding HY, Shi L, Lv
CH (2011) Food Chem 127:890–89289. Wu YL, Hu B (2009) J Chromatogr A 1216:7657–766390. Lu QM, Chen LH, Lu MH, Chen GN, Zhang L (2010) J Agric
Food Chem 58:2763–277091. Gupta V, Kumar M, Brahmbhatt H, Reddy CRK, Seth A, Jha B
(2011) Plant Physiol Biochem 49:1259–126392. Poole CF (2003) TrAC Trends Anal Chem 22:362–37393. Novák O, Tarkowski P, Tarkowská D, Doležal K, Lenobel R,
Strnad M (2003) Anal Chim Acta 480:207–21894. Takei K, Yamaya T, Sakakibara H (2003) J Plant Res 116:265–26995. van Rhijn JA, Heskamp HH, Davelaar E, Jordi W, Leloux MS,
Brinkman UAT (2001) J Chromatogr A 929:31–4296. Ma Z, Ge LY, Lee ASY, Yong JWH, Tan SN, Ong ES (2008)
Anal Chim Acta 610:274–28197. Ge LY, Yong JWH, Tan SN, Yang XH, Ong ES (2004) J Chromatogr
A 1048:119–12698. Li YH, Wei F, Dong XY, Peng JH, Liu SY, Chen H (2011)
Phytochem Anal 22:442–449
72 F. Du et al.

99. Ge LY, Yong JWH, Tan SN, Yang XH, Ong ES (2006) J ChromatogrA 1133:322–331
100. Dobrev PI, Kamínek M (2002) J Chromatogr A 950:21–29101. Dobrev PI, Havlíček L, Vágner M, Malbeck J, Kamínek M
(2005) J Chromatogr A 1075:159–166102. Wei F, Cheng JH, Liu SY, Huang JY, Wang Z, Dong XY, Li PP,
Kong FP, Wu Y, Li YH, Feng YQ, Chen H (2010) PhytochemAnal 21:290–297
103. Risticevic S, Niri VH, Vuckovic D, Pawliszyn J (2009) AnalBioanal Chem 393:781–795
104. Meyer R, Rautenbach GF, Dubery IA (2003) Phytochem Anal14:155–159
105. Zadra C, Borgogni A, Marucchini C (2006) J Agric Food Chem54:9317–9321
106. Liu HT, Li YF, Luan TG, Lan CY, ShuWS (2007) Chromatographia66:515–520
107. Liu Z, Wei F, Feng YQ (2010) Anal Methods 2:1676–1685108. Turiel E, Martín-Esteban A (2010) Anal Chim Acta 668:87–99109. Beltran A, Borrull F, Cormack PAG, Marcé RM (2010) TrAC
Trends Anal Chem 29:1363–1375110. Chen LX, Xu SF, Li JH (2011) Chem Soc Rev 40:2922–2942111. Kugimiya A, Takeuchi T (1999) Anal Sci 15:29–33112. Kugimiya A, Takeuchi T (1999) Anal Chim Acta 395:251–255113. Chen CB, Chen YJ, Zhou J, Wu CH (2006) Anal Chim Acta
569:58–65114. Du FY, Ruan GH, Liang SH, Xie FW, Liu HW (2011) J
Chromatogr A115. Hennion MC, Pichon V (2003) J Chromatogr A 1000:29–52116. Hauserová E, Swaczynová J, Doležal K, Lenobel R, Popa I,
Hajdúch M, Vydra D, Fuksová K, Strnad M (2005) J ChromatogrA 1100:116–125
117. Hradecká V, Novák O, Havlíček L, Strnad M (2007) J ChromatogrB 847:162–173
118. Pĕnčík A, Rolčík J, Novák O, Magnus V, Barták P, Buchtík R,Salopek-Sondi B, Strnad M (2009) Talanta 80:651–655
119. Novák O, Hauserová E, Amakorová P, Doležal K, Strnad M(2008) Phytochem 69:2214–2224
120. Simerský R, Novák O, Morris DA, Pouzar V, Strnad M (2009) JPlant Growth Regul 28:125–136
121. Liang Y, Zhu XC, Zhao MP, Liu HW (2011) Methods.doi:10.1016/j.ymeth.2011.08.006
122. Engelberth J, Schmelz EA, Alborn HT, Cardoza YJ, Huang J,Tumlinson JH (2003) Anal Biochem 312:242–250
123. Schmelz EA, Engelberth J, Tumlinson JH, Block A, Alborn HT(2004) Plant J 39:790–808
124. Kristl J, Veber M, Krajnicic B, Oresnik K, Slekovec M (2005)Anal Bioanal Chem 325:886–893
125. Rozhon W, Petutschnig E, Wrzaczek M, Jonak C (2005) AnalBioanal Chem 382:1620–1627
126. Chen H, Zhang ZX, Zhang GM, Guo XF, Zhang HS, Wang H(2010) J Agric Food Chem 58:4560–4564
127. Gómez-Cadenas A, Pozo OJ, García-Augustín P, Sancho JV(2002) Phytochem Anal 13:228–234
128. Segarra G, Jauregui O, Casanova E, Trillas I (2006) Phytochem67:395–401
129. Ge LY, Yong JWH, Goh NK, Chia LS, Tan SN, Ong ES (2005) JChromatogr B 829:26–34
130. Chen WQ, Gai Y, Liu SC, Wang RX, Jiang XN (2010) J IntegrPlant Biol 52:925–932
131. Tarkowski P, Václavíková K, Novák O, Pertry I, Hanuš J, WhenhamR, Vereecke D, ŠebelaM, StrnadM (2010) Anal ChimActa 680:86–91
132. Huo FF, Bai Y, Liu HW (2010) Rapid Commun Mass Spectrom24:3325–3334
133. Fletcher AT, Mader JC (2007) J Plant Growth Regul 26:351–361
134. Du FY, Bai Y, Bai Y, Liu HW (2011) Chinese J Anal Chem39:1147–1152
135. Huang YQ, Liu JQ, Gong HY, Yang J, Li YS, Feng YQ (2011)Analyst 136:1515–1522
136. Han YH, Zhou ZG, Wu HL, Nie HG, Lei R, Bai Y, Liu HW(2011) Anal Chem submitted
137. Wasternack C (2007) Ann Bot 100:681–697138. Fonseca S, Chico JM, Solano R (2009) Curr Opin Plant Biol
12:539–547139. Fonseca S, Chini A, Hamberg M, Adie B, Porzel A, Kramell R,
Miersch O, Wasternack C, Solano R (2009) Nat Chem Biol5:344–350
140. Krammell R, Schneider G, Miersch O (1996) Phytochem Anal7:209–212
141. Blanch GP, Flores G, del Mar Caja M, del Castillo MLR (2009) JSep Sci 32:180–184
142. del Mar Caja M, Blanch GP, del Castillo MLR (2008) J AgricFood Chem 56:5475–5479
143. de la Peña Moreno F, Blanch GP, Flores G, del Castillo MLR(2010) J Chromatogr A 1217:1083–1088
144. Fan SF, Wang XP, Li PW, Zhang Q, Zhang W (2011) J Sep Sci34:640–650
145. López-Carbonell M, Gabasa M, Jáuregui O (2009) Plant PhysiolBioch 47:256–261
146. Barkawi LS, Tam YY, Tillman JA, Normanly J, Cohen JD (2010)Nat Protoc 5:1609–1618
147. Birkemeyer C, Kolasa A, Kopka J (2003) J Chromatogr A993:89–102
148. Koo YJ, Yoon E, Song JT, Seo HS, Kim JH, Lee YW, Lee JS,Cheong JJ, Choi YD (2008) J Chromatogr B 863:80–87
149. Wang SJ, Jia L, Xing D, Chen DP, Zhao JS (2008) J Sep Sci31:859–864
150. Lu QM, Zhang L, Chen LH, Lu MH, Tong P, Chen GN (2010) JSep Sci 33:651–657
151. Wang YH, Tan HR, Gao LP, Si XY, Shao L (2008) Acta LaserBiol Sinica 17:840–845
152. Hui LJ, Ma Y (2011) Chromatographia 73:507–515153. Ge LY, Tan S, Yong JWH, Tan SN (2006) Electrophoresis
27:4779–4791154. Liu X,Ma L, Lin YW, LuYT (2003) J Chromatogr A 1021:209–213155. Zhang ZL, Liu X, Li DF, Lu YT (2005) Anal Bioanal Chem
382:1616–1619156. Bai Y, Du FY, Yang YY, Bai Y, Liu HW (2011) J Sep Sci
34:2893–2900157. Yin XB, Guo JM, Wei W (2010) J Chromatogr A 1217:1399–
1406158. Ge L, Yong JWH, Tan SN, Ong ES (2006) Electrophoresis 27
(11):2171–2181159. Ge LY, Peh CYC, Yong JWH, Tan SN, Hua L, Ong ES (2007) J
Chromatogr A 1159:242–249160. Ge L, Tan SN, Yong JWH, Hua L, Ong ES (2008) Electrophoresis
29(10):2024–2032161. Ge LY, Yong JWH, Tan SN, Hua L, Ong ES (2008) Electrophoresis
29:2126–2134162. Han Y, Bai Y, Xiao Y, Du F, Liang Y, Tan Z, Zhao M, Liu H
(2011) Electrophoresis 32:2693–2699163. Ge LY, Yong JWH, Tan SN, Yang XH, Ong ES (2005)
Electrophoresis 26:1768–1777164. Zhang H, Tan GL, Yang LN, Yang JC, Zhang JH, Zhao BH
(2009) Plant Physiol Biochem 47:195–204165. Zhao J, Li G, Yi GX, Wang BM, Deng AX, Nan TG, Li ZH, Li
QX (2006) Anal Chim Acta 571:79–85166. Guthrie JW, Hamula CLA, Zhang HQ, Le XC (2006) Methods
38:324–330167. Deng AX, Tan WM, He SP, Liu W, Nan TG, Li ZH, Wang BM,
Li QX (2008) J Integr Plant Biol 50:1046–1052
Analytical methods for tracing plant hormones 73

168. Nagamune K, Hicks LM, Fux B, Brossier F, Chini EN, Sibley LD(2008) Nature 451:207–210
169. Gubler F, Hughes T, Waterhouse P, Jacobsen J (2008) PlantPhysiol 147:886–896
170. Zhou GH, Wang P, Yuan J, Qiu T, He ZK (2011) Sci China Chem54:1298–1303
171. Suzuki Y, Mizuno T, Urakami E, Yamaguchi I, Asami T (2008)Plant Biotechnol J 6:355–367
172. Lee YG, Asami T, Yamaguchi I, Ueda H, Suzuki Y (2008)Biochem Biophys Res Commun 376:134–138
173. Chen Q, Qi WB, Reiter RJ, Wei W, Wang BM (2009) J PlantPhysiol 166:324–328
174. Khripach VA, Zhabinskii VN, Litvinovskaya RP (2011) Immuno-assays of brassinosteroids. In: Hayat S, Ahmad A (eds) Brassi-nosteroids: a class of plant hormone. Springer, Dordrecht, pp375–392
175. Zrazhevskiy P, Sena M, Gao XH (2010) Chem Soc Rev 39:4326–4354
176. Kuang H, Zhao Y, Ma W, Xu L, Wang L, Xu C (2011) TrACTrends Anal Chem 30:1620–1636
177. Sadanandom A, Napier RM (2010) Curr Opin Plant Biol 13:736–743
178. Li J, Wu ZW, Xiao LT, Zeng GM, Huang GH, Shen GL, Yu RQ(2002) Anal Sci 18:403–407
179. Li J, Xiao LT, Zeng GM, Huang GH, Shen GL, Yu RQ (2003)Anal Chim Acta 494:177–185
180. Li YW, Xia K, Wang RZ, Jiang JH, Xiao LT (2008) Anal BioanalChem 391:2869–2874
181. Wang RZ, Li YW, Li Q, Shen GL, Xiao LT (2009) Anal Lett 42(17):2893–2904
182. Li Q, Wang RZ, Huang ZG, Li HS, Xiao LT (2010) ChineseChem Lett 21:472–475
183. Li J, Xiao LT, Zeng GM, Huang GH, Shen GL, Yu RQ (2003)Anal Biochem 321:89–95
184. Casanova E, Valdés AE, Fernández B, Moysset L, Trillas MI(2004) J Plant Physiol 161:95–104
185. Li J, Xiao LT, Zeng GM, Huang GH, Shen GL, Yu RQ (2005) JAgric Food Chem 53:1348–1353
186. Zhang XY, Liu XM, Liu WL, Yang M, Zhang ZQ (2010) Chem JChinese Univ Chin 31:33–37
187. Yang YJ, Xiong XW, HouKK, Hu SS (2011) Russian J Electrochem47:47–52
188. Gan T, Hu CG, Chen ZL, Hu SH (2011) Talanta 85:310–316189. Gan T, Hu CG, Chen ZL, Hu SH (2010) J Agric Food Chem
58:8942–8947190. Gan T, Hu CG, Chen ZL, Hu SH (2010) Sens Actuators B Chem
151:8–14191. Wang Z, Wei F, Liu SY, Xu Q, Huang JY, Dong XY, Yu JH, Yang
Q, Zhao YD, Chen H (2010) Talanta 80:1277–1281192. Kowalska M, Tian FM, Šmehilová M, Galuszka P, Frébort I,
Napier R, Dale N (2011) Anal Chim Acta 701:218–223193. McLamore ES, Diggs A, Marzal PC, Shi J, Blakeslee JJ, Peer
WA, Murphy AS, Porterfield DM (2010) Plant J 63:1004–1016194. Napier R (2011) Plant hormone sensing, group homepage. http://
www2.warwick.ac.uk/fac/sci/lifesci/research/planthormonesensing.Accessed 8 Oct 2011
195. Li YN, Wu HL, Nie JF, Li SF, Yu YJ, Zhang SR, Yu RQ (2009)Anal Methods 1:115–122
196. Li YN, Wu HL, Zhu SH, Nie JF, Yu YJ, Wang XM, Yu RQ(2009) Anal Sci 25:83–88
197. Xie LJ, Ying YB, Ying TJ (2009) Chemometr Intell Lab 97:141–145
198. Guo JM, Xin YY, Yin XB (2010) J Agric Food Chem 58:6556–6561
199. Blilou I, Xu J, Wildwater M, Willemsen V, Paponov I, Friml J,Heidstra R, Aida M, Palme K, Scheres B (2005) Nature 433:39–44
200. Monshausen GB, Miller ND, Murphy AS, Gilroy S (2011) Plant J65:309–318
201. Depuydt S, Hardtke CS (2011) Curr Biol 21:R365–R373202. Luo Y, Qin GJ, Zhang J, Liang Y, Song YQ, Zhao MP, Tsuge T,
Aoyama T, Liu JJ, Gu HY, Qu LJ (2011) Plant Cell 23:1352–1372
203. Varbanova M, Yamaguchi S, Yang Y, McKelvey K, Hanada A,Borochov R, Yu F, Jikumaru Y, Ross J, Cortes D, Ma CJ, Noel JP,Mander L, Shulaev V, Kamiya Y, Rodermel S, Weiss D, PicherskyE (2007) Plant Cell 19:32–45
204. Yoshimoto K, Jikumaru Y, Kamiya Y, Kusano M, Consonni C,Panstruga R, Ohsumi Y, Shirasu K (2009) Plant Cell 21:2914–2927
205. Kanno Y, Jikumaru Y, Hanada A, Nambara E, Abrams SR,Kamiya Y, Seo M (2010) Plant Cell Physiol 51:1988–2001
206. Saika H, OkamotoM,Miyoshi K, Kushiro T, Shinoda S, JikumaruY,Fujimoto M, Arikawa T, Takahashi H, Ando M, Arimura S, MiyaoA, Hirochika H, Kamiya Y, Tsutsumi N, Nambara E, Nakazono M(2007) Plant Cell Physiol 48:287–298
207. Koo AJK, Cooke TF, Howe GA (2011) Proc Natl Acad Sci U S A108:9298–9303
208. Mader J, Emery RJN, Turnbull CGN (2003) Physiol Plant119:295–308
209. Stirk WA, Arthur GD, Lourens AF, Novák O, Strnad M, vanStaden J (2004) J Appl Phycol 16:31–39
210. Stirk WA, Novák O, Václavíková K, Tarkowski P, Strnad M, vanStaden J (2008) Planta 227:1279–1289
211. Hyun Y, Choi S, Hwang HJ, Yu J, Nam SJ, Ko J, Park JY, SeoYS, Kim EY, Ryu SB, Kim WT, Lee YH, Kang H, Lee I (2008)Dev Cell 14:183–192
212. Urano K, Kurihara Y, Seki M, Shinozaki K (2010) Curr OpinPlant Biol 13:132–138
213. Ortiz-Castro R, Díaz-Pérez C, Martínez-Trujillo M, del Río RE,Campos-García J, López-Bucio J (2011) Proc Natl Acad Sci USA108:7253–7258
214. Ouyang G, Vuckovic D, Pawliszyn J (2011) Chem Rev111:2784–2814
215. Duan CF, Shen Z, Wu DP, Guan YF (2011) TrAC Trends AnalChem 30:1568–1574
216. Hoa XD, Kirk AG, Tabrizian M (2007) Biosens Bioelectron23:151–160
217. Mayer KM, Hafner JH (2011) Chem Rev 111:3828–3857218. Liu J, Wang H, Cooks RG, Ouyang Z (2011) Anal Chem
83:7608–7613219. Careri M, Mangia A (2011) Anal Bioanal Chem 399:2585–2595
74 F. Du et al.




















