
Progress in quantitative analysis of plant hormones - Springer ...
Upload
khangminh22Category
view
1download
0
Two Hormones for One Receptor: Evolution,Biochemistry, Actions, and Pathophysiologyof LH and hCG
Livio Casarini,1,2 Daniele Santi,1,3 Giulia Brigante,1,3 and Manuela Simoni1,2,3
1Department of Biomedical, Metabolic, and Neural Sciences, University of Modena and Reggio Emilia,Modena 41121, Italy; 2Center for Genomic Research, University of Modena and Reggio Emilia, Modena 41121,Italy; and 3Unit of Endocrinology, Department of Medical Specialties, Azienda Ospedaliero-Universitaria,Modena 41125, Italy
ABSTRACT LH and chorionic gonadotropin (CG) are glycoproteins fundamental to sexual development and reproduction. Because they act
on the same receptor (LHCGR), the general consensus has been that LH and human CG (hCG) are equivalent. However, separate evolution
of LHb and hCGb subunits occurred in primates, resulting in two molecules sharing ~85% identity and regulating different physiological
events. Pituitary, pulsatile LH production results in an ~90-minute half-life molecule targeting the gonads to regulate gametogenesis and
androgen synthesis. Trophoblast hCG, the “pregnancy hormone,” exists in several isoforms and glycosylation variants with long half-lives
(hours) and angiogenic potential and acts on luteinized ovarian cells as progestational. The different molecular features of LH and hCG lead
to hormone-specific LHCGR binding and intracellular signaling cascades. In ovarian cells, LH action is preferentially exerted through kinases,
phosphorylated extracellular-regulated kinase 1/2 (pERK1/2) and phosphorylated AKT (also known as protein kinase B), resulting in
irreplaceable proliferative/antiapoptotic signals and partial agonism on progesterone production in vitro. In contrast, hCG displays notable
cAMP/protein kinase A (PKA)-mediated steroidogenic and proapoptotic potential, which is masked by estrogen action in vivo. In vitro data
have been confirmed by a large data set from assisted reproduction, because the steroidogenic potential of hCG positively affects the
number of retrieved oocytes, and LH affects the pregnancy rate (per oocyte number). Leydig cell in vitro exposure to hCG results in
qualitatively similar cAMP/PKA and pERK1/2 activation compared with LH and testosterone. The supposed equivalence of LH and hCG has
been disproved by such data, highlighting their sex-specific functions and thus deeming it an oversight caused by incomplete understanding
of clinical data. (Endocrine Reviews 39: 549 – 592, 2018)
L H and the primate-specific chorionic gonado-tropin (CG) are glycoproteins fundamental for
sexual development and reproduction. Both hormoneshave long been considered equivalent, because theybind the same receptor, the LH chorionic gonado-tropin receptor (LHCGR) (), which is mainlyexpressed in the gonads, and similarly activate theclassical cAMP/protein kinase A (PKA) steroidogenicpathway. In clinical practice, human CG (hCG) is thehormone of choice when LH activity is needed (e.g., inthe treatment of hypogonadotropic hypogonadism(HH) or in assisted reproduction technologies (ARTs),because it is easily purified in high concentration fromurine of pregnant women. Historically, human LH ofpituitary origin was difficult to obtain and lacked fullbiological activity because it embeds a proteolytic siteleading to internally cleaved hormones and thusdisplays one half of the activity of the intact molecule
(, ). Human recombinant LH only recently becameavailable for clinical use in ARTs. However, the ex-perience is insufficient to draw conclusions aboutdifferent, specific indications for LH and hCG inclinical practice. Thus, the idea that human LH andhCG might be used indifferently remains dogmatic.However, because evolution led to the appearance ofseveral copies of the CGB gene in primates, theequivalence between the two gonadotropins with “LHactivity,” interacting with the single receptor, requires acritical reassessment. Physiological considerationssuggest that LH is unique and fundamental for ga-metogenesis regulation. In contrast, the evolutionaryonset of CG genes might be linked to the requirementfor different signals specifically supporting pregnancyin primates. Modern technologies and recent in vitrodata have allowed for the evaluation of multiple sig-naling pathways, activated by a number of LHCGR
ISSN Print: 0163-769X
ISSN Online: 1945-7189
Printed: in USA
Copyright © 2018
Endocrine Society
Received: 23 February 2018
Accepted: 8 June 2018
First Published Online:
13 June 2018
doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv 549
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

interactors (, ). A recent report has suggested that acomplete picture of LH and hCG action might bemissing when exclusively investigating the classicalcAMP/PKA steroidogenic pathway (). In vivo and invitro studies using rodents or rodent-derived cells haveprovided informative results in understanding thephysiology of gonadotropins (, ). However, becauserodents do not have CGB genes, they do not produceCG, and the murine LH receptor is not identical to thehuman receptor (% of the identity between Rattusnorvegicus Lhr and LHCGR). Some recent evidencehas suggested that the human LHCGR possesses aspecific region capable of distinguishing between LHand hCG (). These studies raise the question ofwhether hCG and LH actually are equivalent and fully
interchangeable in humans. In addition, because ro-dents are still being used to calibrate the gonadotropinpreparations used in medical practice () and thephysiology of LH action was mainly obtained frommurine models, another question is whether theprimate-specific nature of the dual ligand systemprovided by LH and CG has been completely char-acterized. Some clinical () and in vitro (, )comparisons of commercial LH and hCG preparationshave been performed in the past years, revealing theexistence of several peculiarities but leaving manyunanswered questions.
We have reviewed the current evidence availableon the similar and different functions of LH andhCG.
Two Ligands for One Receptor: Why?
Ligands and receptors evolved along with their mo-lecular targets, resulting in exclusive, hormone-specificregulation of pathways and physiological functions(). In primates, the existence of LH and hCG asligands for the same receptor suggests that a separationof hormone-specific roles occurred, resulting in dif-ferent, hormone-specific physiological functions.These distinct roles might exist to fulfill differentrequirements for the regulation of fetal develop-ment (in females) and gametogenesis (in bothsexes). These reproductive functions are accompaniedby the prevalent—or even exclusive—time- and sex-dependent presence of only one of the two hormones,pointing toward specific physiological targets rea-sonably regulated by different endocrine signals ().LH- and hCG-specific actions exerted at the molecularlevel are underpinned by different molecular struc-tures associated with each hormone (), which arisefrom a different set of related genes (), along withsource cell-dependent posttranslational modifications(). Although previously considered “equivalent,”both hormones divergently evolved in primates and
are characterized by differences at the genetic, mo-lecular, and physiological level.
Although the specificity of LH and hCG signals hasnot been completely elucidated, it is supported by thedifferent nature of their target cells. In women of fertileage, LH exerts its best-known functions in the ovary,where it mediates proliferative signals in the granulosacells coexpressing the FSH receptor (FSHR) andLHCGR and stimulates androgen synthesis, mainlyandrostenedione, in theca cells exclusively expressingLHCGR. Moreover, LH induces luteinization ofgranulosa cells, progesterone synthesis, and corpusluteum maintenance during the luteal phase of themenstrual cycle. In males, Leydig cells in the testis aretargeted by LH, which induces testosterone pro-duction as the major synthesized androgen. LH istherefore essential for reproduction in both sexes. Incontrast, hCG physiological action is only exerted infemales as a massive progesterone stimulator in thecorpus luteum and mediating placental growth duringpregnancy; it is not produced in males. These con-siderations suggest a sex-specific role for each hor-mone in humans and other primates. Such roles arequite different (gametogenesis vs pregnancy) and most
ESSENTIAL POINTS
· In the past decade, the two hormones LH and human chorionic gonadotropin (hCG) were considered equivalent becausethey bind the same receptor, clearly activating the classically known cAMP/PKA steroidogenic pathway
· Clinical evidence of small or undetectable different outcomes between LH and hCG usage has underlined this concept
· Recent in vitro studies have demonstrated that intracellular signaling, downstream events, and cell fate are specificallymediated by LH and hCG
· LH preferentially activates ERK1/2- and AKT-dependent proliferative signals, and hCG is mainly progestational,supporting the physiological roles of the two hormones
· In the past 20 years, studies comparing the use of commercial LH and hCG preparations in reproductive medicine haveprovided clinical evidence of the differences observed in vitro, confirming the in vitro results
· These data indicate that LH and hCG have unreplaceable roles, overthrowing the previous concept that they areequivalent and revisiting the basis on which clinicians decide on the application of these hormones
550 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

likely not fully interchangeable. Clinical experience hasshown that steroidogenesis and gametogenesis can besupported by hCG administration in both sexes.However, is human (primate) pregnancy sustainableby LH in the absence of hCG? In assisted re-production, hCG is administered to provide LH-likeactivity based on the androgen-stimulating potential ofthe molecule. However, is the action of both moleculeson gamete maturation identical? Compelling resultswere provided by a comparison between LH and hCGadministered to mouse oocytes cultures at the ger-minal vesicle stage in vitro. A greater maturation rateafter hCG treatment was revealed, and LH positivelyaffected early embryonic development (). Thesedata, albeit obtained using mouse tissues, suggest thatdifferent signals are mediated by each ligand, resultingin different effects on the physiological target. This isbut one example of a number of recent in vitro andclinical studies providing a novel, unexpected view ofLH- and hCG-specific roles. In the next paragraphs, wedefine such different roles starting from evolutionaryconsiderations.
Phylogenies and Evolution of LH and hCG
The molecular structures of gonadotropins and theirreceptors are, overall, conserved during evolution andshare similarities with several other ligands and re-ceptors across the phylogenic tree. This hints at acommon ancestral origin of these molecules and thepromiscuity of molecular mechanisms involved inendocrine regulation. Gonadotropins are glycoproteinhormones belonging to the superfamily of cystine knotgrowth factors. Members of the cystine knot growthfactor group share an arrangement of six disulfide-linked cysteine residues that achieve a structurallyrelated “knot” conformation, despite a relatively lowsequence homology (). Glycoprotein hormonespossess a common a and a specific b subunit, as-sembled to form a noncovalently linked heterodimer() acting on specific leucine-rich repeats (LRRs) andrhodopsin-like G protein–coupled receptors (GPCRs).It has been estimated that the evolutionary origin ofglycoprotein hormones and their receptors occurred atthe origin of the metazoans. Hence, they organize theregulation of a wide range of endocrine systems andreproductive and metabolic functions that differenti-ated during evolution (). Such evolutionary issueswere inferred by evaluating the genetic structure ofgonadotropins and their receptor-encoding genes. Anumber of glycoprotein hormones (i.e., gonadotro-pins) and receptor variants developed as an endocrineadaptation to specific environmental conditions andthe physiological changes required to improve re-productive success (fitness) tested by natural selection(). For example, the fertile window length, meno-pausal age, and high pregnancy success might be the
result of species-specific, or even individual-specific,optimizations of reproduction implemented to bestbalance fitness and selective pressure. In primates, theappearance of choriogonadotropic hormones acting asadditional ligands for one receptor suggests that dif-ferent levels of regulation are required to managegametogenesis and pregnancy.
Glycoprotein hormones across the phylogenic treeGlycoprotein hormones and their receptors revealedcommon structural folds, suggesting a commonevolutionary origin of ligand–receptor pairs, likelyresulting in similar and even promiscuous binding andsignaling mechanisms. Binding specificity would bedriven by the protein sequence spanning between theth and th cysteine residues of the hormone,consisting of similar sequences and falling within theLRR domains of their receptors (). In insects, theonly molecule related to vertebrate glycoproteinhormones is the bursicon hormone, an approximately-kDa protein displaying a cystine knot hetero-dimeric structure and proapoptotic activity at theepidermal level (). The analysis of the bursiconhormone-encoding DNA sequence revealed highidentity with glycoprotein hormone-like peptidesfound in silkworms, sea urchins, jellyfish, and corals,suggesting their evolutionary proximity (). Similarhomologies were found by comparing the DNA se-quences encoding the hormone receptors, demon-strating that the ligand–receptor system underwentcoevolution. Coexpression of the LHCGR gene hasbeen coupled to the canonical cAMP pathways intransgenic Drosophila melanogaster, similar to that inhumans, further supporting conservation of the in-tracellular machinery necessary for GPCR signaling ininvertebrates (). An evolutionary step toward ver-tebrates might be provided by a glycoprotein hormoneof adenohypophyseal origin found in hagfish. Thishormone consists of bound a and b subunits and thepresence of their mRNAs matches the developmentalstages of the gonad (), likely providing the earliestpituitary–gonadal system in vertebrates. In basalvertebrates, such as the hagfish, unique glycoproteinhormones regulate all the cognate gonadotropinfunctions found in mammals by acting on distinctreceptors (fshr and lhr) mediating gametogenesis ().This ancestral form of glycoprotein hormone, ho-mologs of which are conserved in vertebrates andsome invertebrates, is known as thyrostimulin andis capable of increasing intracellular cAMP in cellsystems expressing lamprey glycoprotein hor-mone receptor (). The evolutionary history ofmore physiologically specialized gonadotropinsstarted ~ million years ago, when the ancestralgene encoding the thyrostimulin b subunit repeat-edly duplicated, resulting in an LHb-encodinggene from which the TSHb- and FSHb-subunit–encoding genes subsequently originated (). Further
551doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

molecular specializations occurred in primates andequids, in which CGb and CGb-like LHb mole-cules, respectively, developed as gonadotropins ofpregnancy.
Coevolution of glycoprotein ligands and their re-ceptors testifies to the specificity of the glycoprotein-mediated signals demonstrated in cartilaginous fishes,providing an ancient representation of the currentdiversity of reptilian, avian, and mammalian endocrinesystems (). However, endocrine signal promiscuity isretained in some organisms such as zebrafish, wherefsh binds both fshr and lhr, and only lh is specific for itsown receptor (). As a consequence of this in-complete functional differentiation, the endocrinecontrol of spermatogenesis in zebrafish relies on fshrexpression in both Leydig and Sertoli cells (). Fishgonadotropin receptors might be activated by mam-mal cognate ligands and vice versa, suggesting a limitednumber of specific binding residues were conservedduring evolution, even in the case of hCG bindingto fish lhr (). The sharing of interspecific bind-ing capability between gonadotropins and theirreceptors is maintained across fish and mammalsas a legacy of common ancestral evolution. Movingfrom invertebrates to primates, however, the in-creasing complexity and specificity of the glyco-protein hormones and receptors is the requisite ofthe refined function of the pituitary–gonadal axis.The result is increasing endocrine specializationwith species evolution.
LHB gene duplication: appearanceof choriogonadotropinLHb and CGb subunits are encoded by a cluster oftandem genes, located in humans on chromosomeq., which embeds a total of eight genes andpseudogenes (). There are six transcriptionally activeCGB genes () and related promoters (). The LHB/CGB gene cluster reaches the highest complexity inHomo sapiens, and other primates feature a simplerorganization characterized by fewer CGB genes. Be-cause most of the % difference between humans andchimpanzees is due to genomic insertions and de-letions (), CGB genes putatively evolved by repeatedduplications of an ancestral LHB gene common to allspecies of the primate lineage to meet specific phys-iological requirements (, ). High crossover activityputatively occurred in the human LHB/CGB clusterand resulted in sequence inversions generating pal-indromic genes (). One of the most accreditedtheories explains the rise of CGB genes and the in-creasing glycosylation rate of CGb molecules asevolutionary adaptations to the elevated energy de-mands for fetal development. The resulting highlyspecialized regulation of angiogenetic signals andmyometrial invasion is necessary to support hemo-chorial placentation in humans and higher primates(). Thus, six CGB genes are found in humans, four to
five in great apes, and one to three among Macaca,Callicebus, and Aotus (). The highly conserved CGBand CGB gene sequences in humans and great apessuggest they could have relevance for implantation andplacental development in higher primates (). OtherCGB genes (i.e., CGB, CGB, CGB, and CGB) arerecognized as a source of hCG products during humanpregnancy (). Accordingly, simpler primates (e.g.,strepsirrhine) and other mammals have only one LHBgene and feature epitheliochorial placentation char-acterized by the presence of both the uterine epi-thelium and the maternal vascular endotheliumduring pregnancy. The theory explaining the rise ofCGB genes has recently been extended (). Sex-specific functions (i.e., placentation, folliculogenesis,and ovulation in females and spermatogenesis inmales) would be driven by the expression of sex-specific genes belonging to the same genomic clus-ter. In this context, the LHB gene is relegated tomaintenance of physiological functions common toboth males and females, because they could requiresimilar regulation at the intracellular level, and CGBgenes permitted independent evolution of signalsspecifically required for placentation in females, as anexample of sexual dimorphism. This strategy mighthave arisen to solve an intralocus sexual conflict, whichoccurs when traits encoded by the same locus lead toconflicting fitness outcomes between the two sexes(). However, the evolution of placentation in pri-mates might be linked to the gene encoding theglycoprotein hormone a subunit (common glyco-protein a, CGA) (). This gene has two splice vari-ants, one of which is only found in anthropoidprimates with the exonization of an Alu sequence. Theadditional encoding sequence results in an N-terminalextension improving protein stability. Similarly, CGBgenes are characterized by an additional DNA se-quence compared with LHB. Although a certain gradeof similarity is shared among genes encoding glyco-protein hormone b subunits, CGBs display a peculiarextension of ~ nucleotides compatible with amino acids at the C-terminal region (). ThisC-terminal extension (CTP) could have originatedfrom the loss of the stop codon, occurring togetherwith the duplication of an ancestral LHB gene,resulting in inclusion of the 9-untranslated regionwithin the protein coding sequence, which is absent inall known mammalian LHB genes, except for equids(Table ). The human CTP domain contains fourpotential O-linked glycosylation sites, is enriched inserine, threonine, and proline residues (), and can betargeted by specific antibodies owing to the immu-nological potential of the glycosylated sites ().
Classical views of LH and hCG physiologyAlthough the diversification of endocrine axes andthe appearance of CG molecules suggest that differentLH receptor-dependent intracellular functions are
552 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

required to regulate gametogenesis and pregnancy,experimental and clinical observations in women andfemale primates failed to clearly distinguish the actionsof these molecules during gametogenesis () andluteal regression (). Thus, it has been widely ac-cepted that these two molecules are equivalent.However, the evaluation of gonadotropin-specificfunctions in human physiology points to distinct,cell-specific roles for each molecule. In ovariangranulosa cells, proliferative signals directly deliveredby LH are not necessarily exerted through the syn-thesis of androgens, which instead are the mainproduct of LH action in theca cells. In contrast, hCGnaturally only replaces LH in the luteal phase andbeyond, during pregnancy, when steroid production ismainly limited to progesterone.
In humans, gametogenesis progresses owing to anorchestrated regulation by FSH, LH, growth factors,and steroid hormones. In women, the ovarian follicleis the functional unit deputed to oocyte maturationand growth, characterized by a dynamic structure ofsomatic cells surrounding the gamete (). Folli-culogenesis starts ~ weeks after conception, whenthe follicular population comprises four to sevenmillion oocytes at the resting primordial stage. Pools ofthese follicles progressively mature by undergoingmorphological and molecular changes and passingthrough the primary and secondary gonadotropin-independent stages. Subsequently, from pubertal on-set, monthly recruitment is guided by the expression ofFSHR and LHCGR at relatively low, but progressivelyincreasing, levels (), conferring follicular sensitivityto FSH. This is the proliferative signal for granulosacells, which start to replicate and synthesize steroidhormones, inducing antrum formation and support-ing oocyte maturation. At the early follicular phase,
relatively low LH levels are produced, and it has beencommonly accepted that these are enough to mediateandrostenedione synthesis by LHCGR-expressingtheca cells (). In contrast, relatively high levels ofFSHR are exposed at the surface of granulosa cells,which also express low levels of LHCGR (),suggesting a physiological role for the putative in-teraction reported to occur between these two re-ceptors () in modulating gonadotropin signals ().If the coexistence of FSHR and LHCGR has biologicalimportance, it is exquisitely granulosa cell-specific,because FSHR is absent in theca cells. Most impor-tantly, these data suggest that LH- and FSH-dependentsynergetic action in granulosa cells, not provided byeach gonadotropin per se, is required to properly guidefollicular growth. At this stage, the best-known role oftheca cells is still to supply androstenedione, as thesubstrate for estrogen production in granulosa cells,supporting follicular growth and oocyte maturation/metabolism. As pituitary FSH release and follicularFSHR expression decline, LH levels increase, alongwith progression toward the large antral stage, whenfollicular growth is gonadotropin and estrogen de-pendent. However, both FSH and LH production stillcoexist at this stage, with both proliferative and ap-optotic signals occurring in the dominant and atreticfollicles, respectively. In the late follicular phase,LHCGRs fully replace FSHRs in the granulosa cells ofthe dominant follicle to induce both ovulation andthe changes in metabolic state necessary for luteini-zation (i.e., massive, exclusively LH-driven proges-terone production). Therefore, although bothFSHR- and LHCGR-mediated life and death signalsare simultaneously present in granulosa cells, onlyLH-dependent signals of increasing potency are de-livered to theca cells throughout the entire follicular
Table 1. Evolutionary and Genetic Differences Between LH and hCG
Endpoint LH hCG Ref.
Presence In all vertebrates In primates (CGs) (39)
Evolutionary convergent molecules Callithrix jacchus pituitary CG eCG (39, 73)
Genes in nonprimates, n 1 0 (39)
Genes in prosimian, n 1 0 (39)
Genes in Macaca, Callicebus,and Aotus, n
1 1-3 (39)
Genes in Callithrix jacchus, n 0 (one pseudogene) 1 (36, 79)
Genes in great apes, n 1 4-5 (39)
Genes in Homo sapiens, n s 1 6 (36, 39)
Pseudogenes in Homo sapiens, n 0 2 (32)
Highest human gene sequenceidentity (92%)
LHB (PMID NM_000894.2) CGB (GenBank BC041054.1) NA
hCG is referred as the “classical”choriogonadotropin, exceptwhere indicated.Abbreviations: eCG, equinechorionic gonadotropin; NA,not applicable/not available;Ref., reference.
553doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

phase up to ovulation. Thereafter, the second half ofthe menstrual cycle depends on progestational signalstransitorily supported by LH in the corpus luteum.Only in the case of pregnancy is corpus luteumfunction maintained by trophoblast hCG, because theLH levels decrease as a consequence of the negativefeedback exerted at the hypothalamic–pituitary levelby high progesterone concentrations. Therefore,physiologically, the steroidogenic LH activity in theovary is not naturally replaced by hCG before thisstage. The progression of pregnancy features steadilyincreasing progesterone levels, which, during the first to weeks of gestation, result from the hCG action onthe corpus luteum (). It is well known that hCGstimulates the maternal androgen production requiredfor fetal development and pregnancy progression ().Moreover, it seems that both placental and adrenalandrostenedione and testosterone play a role in cer-vical and myometrium remodeling and parturition(), and excessively high androgen levels mightcompromise maternal health (). Apart from thesefunctions, little is known about the direct action ofhCG on androgen synthesis, although the role of thegonadotropin has been classically associated withprogesterone synthesis during pregnancy.
hCG is secreted in high amounts, especially in thefirst trimester of pregnancy, and acts as an essentialand potent steroidogenic factor. Other functions havebeen, however, hypothesized for hCG, including im-munosuppressive and angiogenic functions, especiallyduring the early weeks of pregnancy (), and thecapability of enhancing steroid-mediated signals byactivating cAMP- and extracellular-regulated kinase/ (ERK/)–mediated production of the proges-terone receptor in endometrial cells (). The ex-posure to maternal hCG is crucial for fetal sex steroidproduction and activation of the hypothalamic-pituitary-gonadal (HPG) axis, which affects fertilitypotential in adulthood (). After birth, maternalestrogens decline in the newborn, leading to theincrease of FSH and LH, characterizing the neonatalperiod termed minipuberty. This results in a surge ofpituitary gonadotropins of a magnitude only com-parable to the levels obtained much later duringpuberty ().
FSH and LH signaling are also fundamental formale gametogenesis, because these hormones act onSertoli and Leydig cells, respectively, providing me-chanical and endocrine support for sperm production.The hormonal control of spermatogenesis and, es-pecially, its dependence on FSH or LH, are extremelyspecies-specific among mammals (), although tes-tosterone is, in general, an essential requirement forthe progression of gamete maturation. Testoster-one is produced by Leydig cells on LH stimulationand sustains Sertoli cell function and spermatogene-sis progression, albeit being partially converted to-b-estradiol by the aromatase enzyme, promoting
antiapoptotic signals, likely together with gonado-tropins (, ). Given the steroidogenic role of Leydigcells, which express LHCGR, but not FSHR, it isunderstandable that hCG found clinical utility inreplacing LH functions. Because the use of LH fortreatment of male HH is still limited, even in the era ofrecombinant gonadotropins, no substantial data areavailable to differentiate its action from that of hCG inmales.
Studies that focused on the metabolic fate of go-nadotropins indicated that only ~% of hCG is ex-creted in the urine, and the retained hormone isresorbed and degraded mainly in the kidney and, to alesser extent, the liver and ovary (). In the kidney,these molecules are metabolized to b-core fragmentsdeprived of galactose, sialic acid, and CTP fragment,suggesting that these modifications are required forurinary excretion. Sugar moieties play a key role inestablishing the circulatory half-life of LH and hCG.The routes and rates of LH/CG distribution andelimination were compared in rats and piglets, re-vealing that a high quantity of radiolabeled porcine LHis accumulated in the kidneys within minutes ofinjection, the equine chorionic gonadotropin (eCG)plasma concentration is % after hour, and thehormone is not accumulated in any organ (). Takingtogether, these data indicate that LH is eliminatedfrom serum by renal trapping, resulting in rapid re-moval compared with CG. Other, minor routes forgonadotropin elimination can be found in the liver,through binding of sulfated oligosaccharides to aspecific receptor (SGGnM) expressed in the Kupffercells (). Gonadotropins are subsequently processedin the kidney to be excreted with urine as residual,highly similar LHb and hCGb core structures,identical to the original pituitary and trophoblast coremolecules (). This is due to the relatively highstability of the gonadotropin structure, which is nearlyidentical among LH and CGmolecules of humans andother primates (), suggesting that urinary excretionof highly similar gonadotropin core metabolites mightbe evolutionarily conserved.
In summary, LH and hCG are involved in theregulation of multiple physiological functions, buttheir specificity is underrated due to their actionthrough the same receptor and clinical experiencederived from the use of readily available hCG only inthe treatment of HH and in controlled ovarianstimulation (COS) for ART. In the latter case, ga-metogenesis could be clinically supported by admin-istration of exogenous FSH and hCG, thereby at leastpartially replacing LH action. However, clinical datahave not provided representative models for un-derstanding LH- and hCG-specific functions in vivo todate. For instance, stimulation of multiple oocyteproduction in the clinical setting of COS is farfrom replicating the natural oocyte selection, be-cause it results in multifollicular development in a
554 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

mono-ovulatory species. This effect is due to thepharmacological gonadotropin dosages, which do notnecessarily elicit physiological patterns of estrogenproduction and oocyte selection.
Evolutionary convergence: trophoblast LH andpituitary CGsThe endocrine adaptation to pregnancy results indifferent, interesting evolutionary strategies exhibitedacross the phylogenic tree of mammals. The analysis ofthe lhb genomic locus of several species suggests thatthe CTP fragment might be produced by a number oforganisms by frameshift of the gene transcription ().Data suggest that the concept relating the CTPfragment with placentation of primates could be ex-tended to all mammals () that share the potential toproduce glycoproteins bearing the CTP peptide, whichlikely possesses the key characteristics of hCG. A proofof concept might be provided by the bovine, whichproduces a pregnancy LHb variant featuring aCTP fragment. This molecule is produced by de-cryption of the 9 region of the lhb gene, resulting ina glycoprotein hormone that is, however, poorlyO-glycosylated and displays a shorter half-life com-pared with hCG, not supporting the evolution of CGmolecules in bovines ().
A placental gonadotropin was described in equidslong ago (, ). This hormone is known as eCG,suggesting a similarity to the CG molecules of pri-mates. However, both b subunits of equid pituitaryLH (eLH) and trophoblast eCG are products of thesame lhb gene. They differ in their source of pro-duction and N-linked glycosylation, which is greaterfor eCG than for eLH, with O-linked glycosylationcrucial for both hormones to maintain binding activity(). Moreover, both eCG and eLH have demon-strated binding capability for fshr (), with resultseven greater for eLH than for eCG (), suggesting arole in mediating FSH-like signals. The eCG capabilityof promiscuous fshr activation was described in mostmammals (, ), although it was negligible in horses(). These gonadotropins consist of glycosylationvariants, providing a case for evolutionary convergencebetween equids and primates, with different strategiesadopted to support the same physiological process.
Interesting data were provided by studies of pri-mates, which seem to be an evolutionary counterpartto bovines and equids when considering LH and CG.In , LH bioactivity in the New World marmosetmonkey Callithrix jacchus was demonstrated to beproduced by a pituitary monkey CG sharing ~%identity with hCG (). Monkey CG displays multipleactivities regulating development, gametogenesis, andpregnancy, likely owing to a glycosylation patternsimilar to human LH and a CTP structure similar tohCG. In contrast, the marmoset monkey LHCGRlacks the amino acid sequence encoded by exon of the gene, which corresponds to an extracellular
portion of the receptor (). Although the molecularstructures of gonadotropin and their receptors havebeen detailed in the next section (“Different Sources,Molecular Structures, and Biochemical Properties”),the presence of a CG molecule and the lack of an exon–encoded sequence in C. jacchus LHCGR (alsoknown as LHCGR type II) in the entire New Worldmonkey lineage provide interesting information aboutLH/CG receptor functioning, which reaches itsmaximum complexity in primates (). A previousstudy suggested that the CTP fragment is essential toinduce LHCGR type II activation (). One studydescribed an interesting case of an -year-old malepatient with Leydig cell hypoplasia characterized bythe absence of exon –encoded portion of theLHCGR, who was unresponsive to endogenous LH(). Thus, this patient had relatively high serum LHand very low testosterone levels, delayed pubertaldevelopment, and small testes, indicating a deficit inthe LH signal. However, testosterone biosynthesis andspermatogenesis were recovered with hCG treatment.hCG also induced cAMP production during a func-tional analysis in vitro assessed in exon –deficientLHCGR-transfected cells, which LH failed to stimulatedespite binding to the receptor (). This clinical caseconfirmed the importance of the amino acid regionencoded by the exon of LHCGR to discriminatebetween the two natural ligands, supporting theconcept of coevolution of the ligand–receptor struc-ture as a strategy to regulate gametogenesis and pla-centation in primates (Fig. ).
Although it might be intuitive to find thestructure–function relationship of placental gonado-tropins, the role of hCG and hCGb molecules foundin the human pituitary () is still unknown. Theywere detected in the serum of both men () andwomen () and would be released in a pulsatilefashion. It was postulated that pituitary hCG mole-cules might play a role in the regulation of themenstrual cycle () and ovarian pathogenesis ();however, further evidence is needed to support thisissue.
Different Sources, Molecular Structures, andBiochemical Properties
The genetic differences of LH and hCG re-flect gonadotropin-specific molecular structures,posttranslational modifications, and biochemicalproperties at least partially established in the secretorypathway involving the endoplasmic reticulum andGolgi apparatus of the source cell (). The secretion ofLH by gonadotrope cells of the anterior pituitary iscontrolled by the GnRH (), a peptide produced bythe hypothalamus, under kisspeptin regulation (),in a pulsatile fashion and released into the portalbloodstream. GnRH binds its seven-transmembrane
“A variety of LHb moleculescan be produced in humans.”
555doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

receptor expressed in gonadotrope cells, mainly activat-ing phospholipase C (PLC)-, ERK/-, b-catenin–,calmodulin-, and PKA-dependent signaling (, ).The preferential LH or FSH synthesis depends on thefrequency of GnRH pulses. Low frequencies are linkedto FSH production, and higher frequencies are syn-chronized with waves of LH synthesis (). In women,the preovulatory stage of the menstrual cycle is
characterized by a GnRH surge corresponding to theLH increase, inducing ovulation. Given the depen-dence of gonadotropin production on kisspeptin andGnRH, a new model of ovarian physiology wasproposed, in which follicle maturation and selection,ovulation, and luteal phase occur under the strictcontrol of the neuroendocrine system (). The re-lationship between gonadal and pituitary functions
Figure 1. Genetic, structural, and functional relationships between mammal LHb an CGbmolecules. Mice have a unique Lh moleculeencoded by a single Lhb gene. Mouse Lhb acts through its receptor, which displays the exon 10–encoded portion at the hinge region,and regulates both gametogenesis and pregnancy. A CTP fragment appears as a product deriving from the equid Lhb gene, which is thesource of a placental choriogonadotropin. Although pituitary eLH regulates gametogenesis, eCG is specific for pregnancy in equids.However, they act on a common receptor with the exon 10–encoded sequence. The NewWorld monkey Callithrix jacchus has a peculiarLH/CG system. Both gamete maturation and pregnancy are supported by a unique CGB gene encoding transcripts of both pituitary andplacental origin and displaying the CTP fragment. Physiological functions are ensured by the LH receptor type II, lacking the exon10–encoded portion, which binds the CGbmolecule in replacement of the missing LH. In humans, LH and hCG isoforms are encoded byspecific genes belonging to a genetic cluster. However, pituitary LH lacking a CTP and placental hCG possessing a CTP appear as majorproducts, specifically regulating gametogenesis and pregnancy, respectively. Both human LH and hCG act through the same receptordisplaying the amino acid sequence encoded by exon 10, which is fundamental for activating hormone-specific intracellular signaling.Deletion of LHCGR exon 10–encoded region leads to a truncated receptor capable of binding both LH and hCG but resulting inimpaired LH signaling and male infertility. Production of CGa, encoded by CGA genes, is required for proper dimer formation with b
subunits, although CGas were not included in the image.
556 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

was shown by experiments in Lhr knockout andovariectomized mice (). In both of these models,high expression of gonadotropin subunit genes oc-curred, reflecting morphological changes in gonado-trope cells, which display secretory granules larger thanthose in wild-type mice. In contrast, in GnRH-deficient mice expressing low gonadotropin mRNAand protein levels, gonadotrope cells were smaller andfeatured fewer secretory granules. A variety of LHbmolecules can be produced in humans, although thesehave not yet been completely functionally character-ized. They differ in glycosylation patterns, resulting inspecific molecular weights ().
The secretion of hCG by trophoblast cells occursin a nonpulsatile, increasing manner, reaching thepeak around the first trimester of pregnancy, and is notcoupled to GnRH production. A wide variety of hCGisoforms and glycosylation variants are producedduring this period, promoting trophoblast invasion ofmaternal decidua. Most interestingly, and in contrastto LH, hCG production is not subjected to immediatedownregulation by steroid hormones. It could bespeculated that steroidogenic hCG-mediated signalsare constantly delivered during pregnancy as a re-quirement to maintain proper progesterone pro-duction, although the different nature of LH is aimedat more transitory events, such as luteinization, whichneed not be prolonged. A suggestive explanation of themolecular mechanism underlying constant hCG ac-tivity could be provided by the discovery of an~-kDa truncated form of LHCGR, specificallyexpressed by placenta and choriocarcinoma cells, thepresence of which is concomitant with the absence ofhCG downregulation (). The presence of thistruncated receptor would be opposed to the appraisalof full-length (~-kDa) LHCGR at the term placenta,which suggests the existence of a feedback mechanismregulating hCG action via receptor downregulation.Although these findings should be independentlyconfirmed, it is known that hCG production is con-stant but undergoes qualitative dynamic changesthroughout pregnancy. It has been suggested that thepattern and abundance of hCG molecules is in-dividually specific; thus, choriogonadotropin acts as adynamic, autocrine factor that changes qualitativelythroughout the first trimester of pregnancy (). hCGisoforms consist of different polypeptide productsselectively transcribed by CGB genes (), and theoligosaccharide structures differentially linked to thehCG backbone determine the glycosylation variantsand depend on the enzymatic milieu of trophoblastcells, which differs from that of pituitary gonadotropecells ().
The effects of oligosaccharide structures, especiallyO-linked, are apparent in the serum half-life differ-ences between the two hormones, which is minutesfor LH and hours for hCG (). In particular, O-linked glycosylation is abundant in the CTP fragment
of hCG and could be important in determininghormone-specific physiological roles, providing astrategy for developing chimeric gonadotropins withslow metabolic clearance (). In gonadotrope cells,gonadotropin glycosylation is modulated by a numberof hypothalamic and gonadal endocrine factors, suchas estrogens and androgens (). These steroids couldalso affect sialylation and sulfation of the oligosac-charide, further modulating the half-life and bio-potency, thus extending the qualitative range of signalsdelivered to the gonads. In contrast, sulfation of oli-gosaccharides is critical for hormone half-life andbioactivity. This posttranslational modification isevolutionarily preserved among glycoprotein a sub-units, from teleost fishes to mammals (). Finally, itis reasonable, albeit speculative, that the pulsatile re-lease of a short half-lived LH would provide a fine-tuned stimulus optimized to regulate life signalsmediating follicle growth and that hCG moleculeswould trigger the relatively potent stimulus for sus-tained and prolonged progesterone synthesis andangiogenesis. In summary, two different gonadotro-pins, LH and hCG, should be recognized at the re-ceptor level to reach a finer level of regulation, notallowed by a unique ligand, mediating specific in-tracellular signals required to optimize reproductionand development.
Production of a and b subunitsThe production of glycoprotein a subunits is a rate-limiting step for gonadotropin heterodimer formation.The a subunit, encoded by the .-kb CGA gene (),transcripts of which are found in pituitary and pla-centa, allows for glycoprotein hormone heterodimerassembly occurring in the endoplasmic reticulum ofsource cell (). Both hCGa and b subunits arecharacterized by three loops defined by a cysteine knot,and experimental evidence has proved the importanceof the second loop of the a subunit in dimer for-mation. These results were obtained using chimericmolecules, in which the second loops of the hCGa andb subunits were swapped, resulting in b-b homo-dimers capable of receptor binding and activation ofsignal transduction, albeit at lower levels than in wild-type hCG, and inactive a-a homodimers (). Theknowledge of how the formation of glycoproteinhormone heterodimers occurs was improved by sev-eral attempts to develop the crystal structure of hCGduring a -year period from to (–).These models revealed interesting features of themolecule, mostly the seat belt-like structure composedof a segment of the b subunit wrapping around the asubunit loop aL linked by a disulfide bridge betweentwo cysteine residues at positions and of thepolypeptide chain. However, a complete view of thetridimensional dimer structure was strongly hamperedby the oligosaccharide structures bound to hCG.Again, experiments using mutant a subunits lacking
557doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

the disulfide bonds between cysteine residues at po-sitions - and - provided a further step inunderstanding the procedure of gonadotropin heter-odimer formation (). Disruption of these structuresdid not substantially affect hCG or FSH heterodimerformation, although it negatively affected LH heter-odimerization in transfected Chinese hamster ovarian(CHO) cells and in the rat pituitary tumor cell line,GH. In contrast, N-linked glycans at positions and are required for efficient hCGb folding and for-mation of disulfide bonds between the residues atpositions -, -, and - (). These datasuggest that regions recognized by the a subunit forassembly are different in LHb and hCGb.
New insights on the formation of LH andhCG dimers were provided by recent advancementsallowing for the evaluation of molecular interactionsusing a bioinformatic approach. This in silico analysisrevealed that dimer assembly might follow differentb-subunit–specific modes, depending on the presenceor absence of the hydrophobic tail of LHb and hCGb,respectively (). The LHb subunit is not completelyfolded when docking with the a subunit occurs. In suchcases, the a subunit acts as a scaffold, using the cystineknot to enhance b-subunit cystine knot formation. Theheterodimer is then stabilized by the interaction betweenloop of the a subunit and the b-subunit hydrophobictail, which form the seat belt-like structure. This LH-specific mode of dimer formation was named “wrap-around” and differs from the assembly of the hCGa-bdimer, which was named “threading,” to explain themechanism by which the a subunit passes between a bsubunit folded before the docking. An alternativemechanism, similar to the “wraparound”model, was alsoproposed for hCGa-b dimer formation (, ).However, these data suggest that a different heterodimerassembly might have arisen as a strategy to control theproduction of the two hormones. Although hCG as-sembly is efficiently performed, in vitro experiments usingchimeric gonadotropins revealed that LH assembly andsecretion are less vigorous (). The faster rate of hCGsecretion compared with LH secretion presumably re-flects their massively constitutive and GnRH-regulatedsecretory pathways, respectively. The CTP fragment ofhCG plays a role in determining heterodimer formation,because CTP-truncated forms of the chorionic hormoneresulted in a % decreased efficiency of dimerization.Removal of the LHb C-terminal octapeptide increasedthe rate of hormone secretion by transfected CHO cells,confirming the relevance of the C-terminal region of LHand hCG for their physiology.
Previous attempts to produce recombinant LHband hCGb in transfected GH cells in vitro revealedthe role of the N-terminal region in the disulfide-linked aggregation of LHb subunits. In contrast,hCGb was exclusively secreted as a monomericmolecule. Further investigation using mutant go-nadotropins demonstrated that N-glycosylation on
asparagine at position , which LHb lacks, preventshCGb aggregation in culture medium (). Thesedata indicate that glycosylation in the N-terminalregion of the two gonadotropins plays a role in themaintenance of the correct folding and secretion in theabsence of the a subunit (Fig. ). This finding mightexplain the measurable presence of free hCGb sub-units during pregnancy, when high activity of CGBgenes leads to massive protein production and freeLHb subunits are relatively rare ().
LHb subunitLHB gene transcription is stimulated on GnRHbinding to its receptor, activating Gq/ proteins andstimulating phospholipase C to mediate inositol,,-triphosphate and diacylglycerol pathways. Theseintracellular messengers lead to protein kinase C ac-tivation, intracellular calcium increase, and b-cateninsignaling regulating the expression of the LHB gene.The latter is under the control of GATA and chickenovalbumin upstream promoter-transcription factors(, ), and the secretion of LH is regulated by theincrease in intracellular calcium (–). The LHbmonomer was described in the s (). It is an~-kDa glycoprotein of amino acids subjected toposttranslational modifications, which provide .variants of the hormone, with molecular weightsspanning from to and featuring specific bio-activity (). The core fragment is structurally close tothat of hCGb, except for the absence of the CTPfragment. The nomenclature of the amino acid po-sition within LHb and hCGb subunits originatedfrom these first sequence determinations (–),which excluded the -amino acid–long signal pep-tide. A new nomenclature that included the length ofthe signal peptide was proposed later () and wasused to indicate the amino acid positions in thepresent report.
Glycosylation, sulfonation, and sialylation are themain LH modifications, which occur at different ratesduring the ovarian cycle () and with age (). Inparticular, the LHb subunit classically displays one N-glycosylation site at position of the polypeptidechain, and two additional oligosaccharide structuresare linked to the a subunit (). LH heterogeneityconsists of a prevalent glycoform carrying three oli-gosaccharide structures, except during midcycle whendiglycosylated LH is the main LH variant produced.Moreover, the acidic forms of LH molecules increasewith age and are mostly present in elderly women.Because the removal of these oligosaccharide struc-tures by endoglycosidase treatment did not sub-stantially change the LH steroidogenic activity, theirfunctional importance might be related to other as-pects of physiology (). Oligosaccharides can besulfonated or sialylated, resulting in different half-livesof the glycoprotein. Sulfated oligosaccharides consistof branches terminating in SO-GalNAc b,, and
558 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

sialylated oligosaccharides consist of a number of dif-ferent structures featuring two or three branches andone to three sialic acid moieties (). LH mole-cules with two or three sulfonated N-acetylgalactos-amine [SO()-GalNAc] residues have shorter half-livesthan less-sulfonated LH, suggesting their rapid removalby hepatic Kupffer cells. In contrast, higher sialylatedgonadotropin isoforms have extended half-lives, likelyowing to the masking of the sulfonated oligosaccharidesto SGGnM binding and sequestration (, ).However, rather than SO(), sialic acid might be linkedto LH GalNAc residues, likely resulting in an extendedhalf-life ().
hCGb isoforms and glycosylation variantsIn humans, a number of CGb isoforms and variantsare provided by transcription of CGB genes and dif-ferent patterns of glycosylation, respectively. Thesetypes of hCG are known as “classical” hCG, hyper- andhypoglycosylated hCG, nicked isoforms and hCGlacking the CTP fragment, core fragments, and free bsubunits. These molecules represent a palette ofmultiple hCGbs differentially detectable as urinaryproducts by specific immunoassays (, ).
Although in-depth functional characterization ofCGb isoforms in vitro is missing, the expressionpattern of CGB genes might be a determinant of thestatus of pregnancy or miscarriage. Although theregulation of CGB gene transcription is unclear, it islikely under the control of growth factors, cytokines,ligands of the nuclear peroxisome proliferator-activated receptor-g, and steroid hormones, actingthrough activation of cAMP-mediated signals (). Innormal pregnancies, all CGB transcripts were found,especially CGB, CGB, CGB, and CGB, whichachieved - to ,-fold greater expression levelsthan CGB and CGB genes (). An ectopic preg-nancy is characterized by perturbation of CGB ex-pression patterns, featuring a relatively high amount ofCGB and CGB transcripts. In contrast, globallyreduced CGB expression has been associated withmiscarriage. Relatively high amounts of CGB andCGB gene transcripts were found in the testes ofhealthy males, suggesting that they might play a stillunknown role in male reproduction ().
Glycosylation is the posttranslational modificationproviding . hCGb variants, depending on thecombination of glycans specifically elaborated by thesource cell and attaching two potential N- and fourpotential O-glycosylation sites in hCGb (). hCGbisoforms possessing only one N-glycan are known andonly three of four O-glycosylation sites are typicallydecorated with carbohydrate (, ). Moreover,# glycoforms of the a subunit were found, differingby sialylation, oxidation, and N-terminal truncation(). Because hCG and LH are administered forinfertility treatment as a mixture of different glyco-forms resulting from the manufacturing processes and
potentially featuring specific biochemical propertiesand bioactivities, the production of homogeneoushormone samples by chemical synthesis should be ofgreat interest for clinical applications (). Duringpregnancy of aneuploid fetuses, the profile of hCGglycoforms is different from that detectable in preg-nancy of normal karyotyped fetuses (). Althoughfully informative characterization of these hCG gly-coforms in vitro is lacking, it was suggested that theyare related to specific biological activities and func-tions, mainly angiogenic (), in vivo (). Theseconclusions were driven by experiments evaluating cellgrowth and migration in vitro and mediated byhyperglycosylated hCG molecules (hCG-H) (),predominantly produced during the early stages ofpregnancy by extravillous cytotrophoblasts or cho-riocarcinoma, and consistent with the positive regu-lation of early trophoblast invasion (). hCG-Hwould have lower steroidogenic and greater pro-liferative potential than the “classical” form of hCG
Figure 2. Differential free LHb and hCGb subunit release bysomatotrope GH3 cells. Both hormones have an asparagine (N)residue at position 13 of the amino acid chain (N13), which isglycosylated in the hCG molecule. N13 glycosylation preventsthe disulfide-linked aggregation of b subunits and is involved inmaintenance of the correct folding in the absence of the a
subunit. This mechanism might underlie the secretion of freehCGb subunits occurring in pregnancy, when these peptides areproduced in greater amounts than the a subunits, with freeLHb aggregates remaining in the cytoplasm. Vertical lineindicates conserved amino acid residues; two vertical dots,different amino acid residues sharing similar biochemicalproperties; and one dot, biochemically different amino acidresidues.
559doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

produced later (). It should, therefore, be secretedduring the very early days of pregnancy as an essentialmediator of cell proliferation and maternal tissueinvasion by fetal cells. Thus, insufficient hCG-Hduring the first days of pregnancy might be pre-dictive of miscarriage (), highlighting the funda-mental role of this molecule for placentation andembryo development. Similar conclusions were drawnby analyzing the sera of women experiencing recurrentmiscarriages, in which antitrophoblast antibodiesinhibited the hCG-H release by the JEG- cell line invitro (). Alternative, “hyperbranched” glycoformslinked to both a and b subunits are predomi-nantly produced during aberrant pregnancy andby choriocarcinoma rather than in normal preg-nancy, suggesting that the activity of Golgi-processingenzymes occurs differently in malignancy (). Hyper-branching might reflect the presence of free a and bsubunits, by which dimer formation is impaired bylargely tri- and tetra-antennary glycans, in contrast tothose biantennary glycans associated with classicalhCG (). The angiogenetic potential of hyper-glycosylated hCGs is a crucial issue for pregnancysuccess, because proper blood flow is required to avoidembryo hypoxia. This role might be exerted via ac-tivation of cAMP-responsive elements located in thepromoter region of vascular endothelial growthfactor–encoding genes (). It has been proposed thatangiogenic functions and trophoblast invasion ofhCG-H could rely on growth factor-like activitiesputatively mediated through interaction between thehormone and TGFb receptor II, independently ofclassical hCG signaling () (Table ). A similar
mechanism of action was suggested to explain thetumorigenicity of free hCGb subunits in BRCA gene-defective breast cancer (). This is a suggestivehypothesis requiring further confirmation before be-ing accepted, because an independent study revealedthe opposite findings and suggested that experimentalbiases, such as TGFb contamination in the hCGpreparations, might have affected the results ().Although the molecular mechanism underlying theproliferative potential of hCG-H is still unclear, it hasbeen confirmed that these molecules are secreted byseveral tumor cells, even in the male (), likely as aglycosylated product occurring after metabolicreprogramming of tumor cells (). These datasuggest that hCG-H might be a tumor marker andpromoter ().
LH and hCG binding to the human LHCGR:binding differences compared withnonhuman receptorsHuman gonadotropins display binding capability tononhuman receptors. Although indicative of the overallstructure preservation of these ligands and receptorsduring evolution, this is also relevant for clinical treat-ment of human infertility. LH and hCG dosages forclinical purpose are established by evaluating their bi-ological activity against a standard using animal modelsnot expressing the human receptor (), suggestingsome limits to their translation to humans and usage inclinical practice. The first evidence of LH and hCG re-ceptor binding was provided several decades earlier, by invitro studies using rat Leydig cells. These experimentsfound similar sets of binding sites and binding capacity
Table 2. Molecular Differences Between LH and hCG
Endpoint LH hCG Ref.
b amino acid chain length 121 Amino acids 145 Amino acids (98, 101)
Molecular weight ~26-32 KDa ~37 kDa (98, 101)
Highest human protein sequenceidentity (85%)
LHb (PMID NM_000894.2) CGb5 (GenBank: AAI06724.1) NA
LH/hCG-specific amino acidsequences
Absence of CTP 28 amino acid CTP extension (71)
Glycosylations, n 3 (2 N-linked in the a subunit; 1N-linked in the b subunit)
8 (2 N-linked in the a subunit; 2 N- and1 O-linked in the b subunit)
(132)
Total isoforms and glycosylationvariants, n
1 main molecule source of ~39isoforms chromatographicallyseparated
$9 Main molecules (“classical” hCG,hCG-H, nicked hCG, nicked hCGmissing CTP, nicked hCG-H, asialohCG, free hCGb, nicked hCGb,b-core molecule) source of anunknown number of variants
(98, 101)
Molecular targets LHCGR LHCGR; Tb-RII (criticized) (1, 160, 162)
Half-life 25 min 15–462 h (101)
hCG is referred to as the“classical” CG, except asindicated.Abbreviations: NA, notapplicable/not available; Ref.,reference; Tb-RII, TGFbreceptor II.
560 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

for both hormones to the rat Lhr, evaluated by equi-librium association constants (Ka; hCGKa = .3 2
M; LH Ka = . 3 2 M) and association rateconstants (K; hCG K = .3 M/min; LH K .3 M/min) (). However, greater binding affinity wassuggested for hCG compared with LH, owing to itslonger persistence at the receptor level and greater half-time of the bound hormone (hCG, . hours; LH,. hours), resulting in a reduced dissociation rate forhCG. These results suggest that the two hormones in-teract differently with the receptor, as a consequence ofdistinct amino acid sequences. It was observed that themaximal hCG-mediated cAMP increase occurred atrelatively low (,%) Lhr occupancy in rat Leydig cells,adumbrating the existence of “spare” Lhr (). Thisconcept arose by evaluating the discrepancy between hCGbinding and dose–response curves calculated for cAMP,which appeared to be left-shifted. It has been recently usedto explain the putative, steroidogenic activity exerted byLH during the antral phase of folliculogenesis (, ).The putative existence of “spare” LHCGRs in the humanovary can be explained by the requirement for sustainedandrogen synthesis by theca cells, avoiding receptordownregulation by LH. If so, hCG must have a differentaction at the receptor level during pregnancy, when thehormone is massively produced and constantly stimu-lates progesterone synthesis, somehow avoiding LHCGRdownregulation. Although suggestive, these are largelyconjectures, because the existence of “spare” LHCGRs wasonly suggested in vitro. Because hCG is not the naturalligand of rat Lhr, the parallelism between in vitro findingsusing rat Leydig cells and the physiology of the humanovary should be interpreted cautiously (). In con-trast, LH-like signals might be driven by FSH throughFSHR–LHCGR heterodimerization in granulosa cells(), which should be favored by the ~: ratio be-tween the LHCGR and FSHR amounts at the early antralstage (). Moreover, ligand affinity to rat and humanreceptors might not be similar, as suggested by hCG andbovine LH, which both exhibit similar affinities for the ratLhr but different binding affinities for LHCGR (- to,-fold better for hCG than for bovine LH) ().These differences are due to an isoleucine residue fallingwithin the C-terminal end of LRR of LHCGR, com-parable to a serine residue in LRR of the rat Lhr, whichwould determine the LH-specific binding affinity (). Invitro comparisons of human LH, hCG, and some of theirnonhuman, rat, equine, bovine, ovine, and porcinecounterparts in themouse tumor LeydigMLTC cell line,revealed molecule-specific control of adenylate cyclaseactivity, raising the question of whether hCG could be areliable reference ligand in nonhuman LH receptor-expressing systems (). These data suggest that Lhrand LHCGR might not be comparable and might me-diate receptor-specific responses. Although the assump-tion of “spare” ovarian LHCGRs should at least beconfirmed by binding experiments using radiolabeled LHand cells expressing the human LHCGR, it is clear that
most of our knowledge of LH and hCG binding has beenprovided by experiments using cell models expressingnonhuman receptors, relying on the intra- and in-terspecies promiscuity between gonadotropins and theirreceptors but ignoring their differences. Conformationalchanges of LH and hCG occurring on receptor bindingwere first demonstrated using rat gonadal cells (). Theimportance of specific contact sites, such as the inter-cysteine loop sequence of both LHb and hCGb, was alsodemonstrated, revealing that the amino acid region -of these subunits is exposed on the surface of themoleculeand participates in rat Lhr binding (), along with lysineresidues at position of hCGb and position of bothgonadotropins (). However, the hCGa subunit di-rectly interacts with the LHCGR extracellular domain(), participating in hormone receptor binding, and itcannot be excluded that a similar interaction occurs withLH binding.
Informative results regarding LH and hCG bindingaffinity for LHCGR were provided by experimentsevaluating the displacement of radiolabeled hCG byincreasing the molar concentrations of the hormonesincubated together with membrane lysates of LHCGR-transfected COS- cells (). LH displayed an ap-proximately eightfold greater, albeit not statisticallydifferent significantly, half-maximal inhibitory con-centration compared with hCG (IC; hCG IC = .pM; LH IC = . pM), demonstrating a quitesimilar binding affinity for LHCGR between the twogonadotropins. In any case, these experiments shouldbe repeated, evaluating the displacement of radiola-beled LH to draw definitive and clear-cut conclusionsabout the binding features of both molecules. Incontrast, most of the current knowledge depicting theinteraction between LHCGR and its ligands wasprovided by classical experiments using mutatedhuman receptors or chimeric hCG. It is known thathormone b subunits contact specific amino acidresidues of LHCGR LRR b strands, especially and (), which play a role in the formation of a “sled-like”tridimensional structure typical of the gonadotropin-receptor extracellular domain and in hormone bindingand activity (). Analysis of the chimeric hCG b-bdimer confirmed the need for this subunit for receptorbinding. However, this dimer bound two receptormolecules with threefold lower affinity than classicalhCG and failed to elicit any cAMP response ().These results have demonstrated that the a subunit isinvolved in LHCGR binding and activity. Experimentsusing an hCG analog, obtained by fusing the C-ter-minus of the a subunit and the N-terminus of the bsubunit through a CTP fragment, confirmed that boththe N- and the C-terminal portions are involved inreceptor binding and activation (), and the seatbelt-like structure of hCG is only minimally involved inLHCGR binding (). Finally, both the a and bsubunits of hCG possess the first three b-hairpinloops, structurally similar to those of the TNF and the
561doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

nerve growth factor, not involved in receptor binding(). It is remarkable that the corresponding struc-tures of TNF and nerve growth factor were insteadcrucial for interacting with their respective receptors.
Interesting results were provided by mutagenesis ofLHb and hCGb, revealing hormone-specific bio-chemical features embedded in the polypeptidestructure of the two molecules. The glutamine residueat position of the b subunit plays a key role in dimerformation. Substitution of this glutamine with a basicamino acid residue, arginine or lysine, resulted insubunit association decreasing to ,% comparedwith the wild-type and formation of inactive, mutatedLH and hCG dimers, which failed to induce pro-gesterone synthesis in the MA- cell line. Neutral(alanine) or acidic (glutamic acid) residue substitutionat position resulted in mildly (% to %) de-creased subunit association and a lack of mutant LHbinding, although mutant hCG retained full activity(). Although the amino acid residue at position is crucial for both LH and hCG heterodimer forma-tion, structural characteristics intrinsic to the proteinchains result in functional differentiation between LHand hCG despite the similar sequences. Most im-portantly, these data suggest that the two gonado-tropins might bind the receptor differently; however,further mutagenesis experiments are needed to fullyclarify this issue. Overall, they seem to interact simi-larly with the LRR domain (), and a second, less-known, binding site falling within position -and belonging to the hinge region of LHCGRmight beinvolved in hormone-specific contacts. Advancementswere provided using a bioinformatic approach eval-uating the LH- and hCG-specific interaction with thehinge region. In this case, an extroflexion consistingof a sulfated tyrosine located at position (sTyr)would play a key role in discriminating between thetwo hormones, relying on a specific spatial confor-mation of the receptor hinge region (Fig. ). Theprotein segment carrying the sTyr features a “U-shaped” structure displaying proximity between theamino acid sequence encoded by the LHCGR exon and an adjacent helix. Both LHb and hCGb first bindthe LRR domain of the receptor. However, althoughthe spatial occupancy provided by hCG binding toLRRs contributes to the LHCGR conformationalchange by contacting the whole “U-shaped” structure,the smaller size LH needs to interact with the sTyrextroflexion to induce proper conformational as-sembly of the receptor (). The deletion of the aminoacid sequence encoded by exon would result in amodification of the “U-shaped” structure, consisting ofthe shift of the adjacent helix, which spatially replacesthe exon –encoded sequence, shifting Tyr to aspecial position not permissive for LH accommoda-tion. Thus, LH signaling would be impaired () andhCG would retain its functional properties owing topreservation of the “U-shaped” structure binding
and conformational change of LHCGR. These datawere supported by experiments altering the exon–encoded sequence by introducing a double prolinemutation at positions and . In this case, dis-ruption of the “U-shaped” structure with preservationof the proper spatial location of the sTyr residuewould negatively affect hCG but not LH signaling ().
Because LH and hCG display a specific interactionwith LHCGR, different conformational changes of thereceptor can occur, depending on the hormone. In theHEK cell line, coexpression of signaling-deficientLHCGR and binding-deficient LHCGR () allowedfor evaluation of the ligand-induced intermolecularcooperation (). Although treatment by hCG waslinked to full cAMP activation, LH failed to induce anintracellular increase in second messenger (). AnhCG-induced cAMP increase was the result ofbinding-deficient receptor activation by signaling-deficient LHCGR capable of hormone binding,which relies on receptor dimer formation and theligand–receptor complex undergoing specific con-formational changes. This is the so-called LHCGRtransactivation, and it was suggested to occur duringhCG treatment, with LH able to induce mainly self cis-activation of receptors capable of hormone binding(). These results and, in general, receptor cis- andtransactivation were independently supported (,, ), even if the opposite results were alsoreported (), questioning the concept of ligand-specific functional rescue between LHCGR mole-cules ().
In summary, as a result of common evolution,cross-interaction between ligands (LH and hCG vsmurine lh) and receptors (human LHCGR, murinelhr) from different species can be demonstrated in vivoand in vitro, together with species-specific patterns ofligands for one receptor as adaptations to maintaineffective biological responses. However, human LHand hCG display their own specific molecular in-teractions with human and nonhuman receptors,resulting in hormone-dependent modulation of down-stream intracellular signaling and physiology, which isconsidered in detail in the next paragraphs.
LH- and hCG-Specific Intracellular Events
LHCGR binding to its ligands triggers a number ofsubsequent events mediating the activation of multiplesignal transduction pathways (). These events startafter hormone interaction with its high-affinitybinding site located in the extracellular domain ofthe receptor, which, however, is not capable of gen-erating intracellular signaling per se. The bound re-ceptor undergoes a conformational change affectingthe hinge region and, subsequently, the trans-membrane domain. However, contacts between theextracellular domain and loops (), especially the
562 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

second and third extracellular loops, are necessary forproper signaling activation. These secondary, low-affinity contacts established by the hormone com-pelled to interact with extracellular loops and the hingeregion play a key role in signal generation (). Thespatial conformation of the activated LHCGR is linkedto different, independently activated, signaling cas-cades, depending on various cell-specific intracellularinteractors of the receptor, which mainly consist of Gproteins () and b-arrestins (). Although thepresence of LHCGR at the cell surface is linked toweak basal signals existing as an equilibrium betweenstimulatory and inhibitory signals, maximal pro-duction of high-affinity signal occurs on hormonebinding.
Although the intracellular events described areoverall common to all glycoprotein hormone re-ceptors, the existence of ligand–LHCGR-specific in-teractions suggests that qualitatively and quantitativelydifferent patterns of intracellular signaling cascadesmight be differentially activated by LH and hCG.These features could rely on peculiar LHCGR con-formational changes induced by ligands and thepresence of other interacting glycoprotein hormonereceptors and the intracellular enzymatic milieu oftarget cells. All these factors likely contribute jointly todifferentiating LH and hCG physiology.
Classical views and insights onLHCGR-mediated signalingKnowledge of LHCGR-mediated intracellular eventshas progressively increased during the past few de-cades, revealing a complex picture of gonadotropinfunctions not explainable exclusively by the oldconcept of steroidogenesis as the main endpoint ofboth LH and hCG function, exerted via cAMP/PKAactivation and intracellular calcium ion (Ca+) in-crease. This classical assumption presumably origi-nated when cAMP and steroid hormones were themain—or even only—molecules analyzable using thefirst assays available. Modern experimental techniqueshave revealed the existence of several intracellularLHCGR interactions and multiple signaling cascades,requiring a re-evaluation of LH- and hCG-mediatedsignals.
It is common knowledge that LH and hCG inducesimultaneous increases of the second messengercAMP and Ca+ through the LHCGR (). These twoevents occur relatively early after receptor activation,within , minute (, ), and belong to twoseparate, G-protein–dependent signaling pathways(). Spatial conformation of the activated receptorleads to G-protein stimulatory Gas subunit dissoci-ation from the bg dimer, thus activating the adenylylcyclase membrane enzyme, which, in turn, catalyzesthe conversion of ATP into cAMP. This secondmessenger, before its metabolization to AMP byphosphodiesterase enzymes, induces PKA activation
and transcription factor cAMP response element–bindingprotein (CREB) phosphorylation (). However,relatively high intracellular cAMP concentrations werelinked to proapoptotic effects in granulosa cells, alongwith progesterone synthesis and androgen conver-sion to estrogens (, ). In the theca cell, phos-phorylated CREB (pCREB) binds CRE DNA targetsequences, modulating the transcription of steroido-genic enzyme-encoding genes, such as STARDand CYPA, and synthesis of androstenedione.Phosphorylation of the extracellular-regulated kinase/ (ERK/; pERK/) occurs as a downstreamevent to PKA activation in theca cells concomitantwith CREB phosphorylation, inhibition of proges-terone synthesis, and stimulation of androgens syn-thesis by differently modulating the transcription ofgenes encoding steroidogenic enzymes (). Most
© 2018 Endocrine Reviews ENDOCRINE SOCIETY
Intracellular
LHCGR
Extracellulardomain
Serpentinedomain
Exon 10-encoded sequence
LHsTyr331 residue
Intracellular
hCG LH
hCG
Hingeregion
Figure 3. Discrimination of LH- and hCG-mediated signaling stratified by the LHCGR hinge region.Both hormones bind the extracellular domain of the receptor but interact differently with the “U-shaped” portion of the hinge region. (Upper) Although hCG contacts the exon 10–encodedportion, LH spatial conformation leads to the interaction of the hormone with the sTyr331 residue.(Lower) These ligand–receptor interactions result in LHCGR conformational changes associatedwith hormone-specific intracellular signaling. Exon 10 deletion results in the shift of the sTyrresidue impairing the interaction with LH, with a contact point of the “U-shaped” structure of thehinge region with hCG maintained. Thus, exon 10 deletion results in a truncated LHCGR unable tomediate proper LH signaling, albeit retaining both LH and hCG binding capability.
563doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

importantly, ERK signaling is linked to proliferationand viability in all gonadal steroidogenic cells (,) and antiapoptotic processes (), revealing thecentral role of the molecule in regulating GPCR sig-nals, including LH and hCG function, and re-production. However, the activation of pERK/ islinked to several other intracellular processes aftergonadotropin stimulation. It is required for the ste-roidogenic response to LH in certain cell types, such asLeydig cells (), for receptor mRNA downregulation(), and to modulate activation of GPCR kinasesinvolved in receptor phosphorylation and subsequentinternalization by b-arrestins (). The recruitmentof b-arrestins, by itself, is responsible for a secondpERK/ activation () as a likely opposing effectto cAMP proapoptotic events mediated by GPCRs().
Gonadotropin-induced mobilization of intracel-lular Ca+ was first investigated in hCG-treated,transfected cells expressing the murine Lhr and wasassociated with PLC activation (). The signalingcascade is triggered by the Gaq protein activating PLC,with subsequent cleavage of phosphatidylinositol,-bisphosphate to diacyl glycerol and inositol tris-phosphate (IP). IP binds calcium channels located inthe endoplasmic reticulum, resulting in Ca+ releasein a hormone concentration-dependent manner. Al-though both cAMP/PKA and PLC/Ca+ pathways areactivated simultaneously, the hCG half-maximal ef-fective dose (EC) is -fold higher for Ca+ mobi-lization than that needed for cAMP recruitment and isindependent of receptor density, demonstrating thatLHCGR carries a dual signaling potential (). Ca+
binds the calcium-modulated protein calmodulinresulting in downstream activation of calcium-modulatedprotein calmodulin kinases, which control choles-terol transport into mitochondria and steroidogenesis(, ). Moreover, Ca+ signaling was associatedwith proliferative effects in vitro (). Simultaneouswith these events, the bg dimer of G protein mightlead to phosphatidylinositol ,-bisphosphate phos-phorylation to phosphatidylinositol ,-trisphosphateby phosphatidylinositol-,-bisphosphate -kinase,thus activating AKT (also known as protein kinase B).AKT might also be activated through the epidermalgrowth factor (EGF) receptor, exerting antiapoptoticroles and inhibition of CYPA expression (, )and, thus, negatively modulating steroidogenesis, atleast partially. In contrast, activation of AKT andERK/ signaling are necessary for STARD expres-sion, and it is a pathway preserved across mammaliangonads and adrenal glands of mammals to mediategonadotropin- and ACTH-induced steroidogenesis,respectively (). Taken together, gonadotropin sig-naling might stem from a balance between opposingsteroidogenic and proapoptotic vs proliferative andantiapoptotic intracellular events providing the en-docrine regulation of reproduction.
An interesting and relatively recent development isthe dependence of the gonadotropin-mediated sig-naling on receptor concentrations, owing to prefer-ential coupling to b-arrestin/ERK/ and AKTpathways at a relatively low receptor density as analternative to the canonical cAMP/PKA pathway(, , ). These data provide a compelling,albeit speculative, regulatory mechanism that mightcontribute to differentiating gonadotropin signals,depending on the physiological requirements. LHCGRexpression is a dynamic event during the menstrualcycle () and might be associated with different LH-dependent roles. During the follicular and lutealphases of the menstrual cycle, the equilibrium betweenLH-driven proliferative and steroidogenic signalsmight be differentially regulated through LHCGRcoupling, modulating granulosa cell proliferation,luteinization, and androgen or progesterone synthesis.At the early antral stages, LHCGR-mediated signalsmainly consist of the regulation of proliferative signalsdelivered to granulosa cells, and these can occur as aresult of unique patterns of intracellular signalingcascades, plausibly activated by FSHR–LHCGR het-erodimers (, , ). The LH-dependent andro-genic potential of ovarian follicles progressivelyincreases, together with LHCGR expression levels, intheca cells during the late antral stage, suggesting thatreceptor density is linked to specific signaling patterns,relying on variable LHCGR concentration–dependentGas protein coupling accompanied by increasing PLCactivation () and cellular metabolic changes. Thesedata highlight the specificity of LH as an irreplaceableligand of LHCGR during physiological follicularphases.
Agonist-induced desensitization is a well-knownfeature of LHCGR and is characterized by the orga-nization of large receptor aggregates at the cellmembrane () preceding their internalization,which, in turn, is the main determinant of down-regulation () regulated by GPCR kinases, b-arrestins,and other modulators (). Experimental evidencesupports the formation of ligand-dependent LHCGRaggregate structures, suggesting formation of largeLHCGR complexes induced by hCG rather than LHbinding (). LHCGR aggregation might be a de-terminant for localization of a number of receptorswithin endosomes to determine trafficking and recy-cling of these molecules. Persistent cAMP signalingcan be induced by the internalized receptor formingcomplexes with both G proteins and b-arrestins ()and suggests it is likely modulated via the adaptorprotein interacting with pleckstrin homology domainand leucine zipper (). The molecular mechanismunderlying persistent cAMP signaling is of greatphysiological relevance, because it could play a crucialrole in sustaining LH functioning at midcycle in thefemale (), thus bypassing the potential arrest ofsteroidogenic signals due to LHCGR downregulation.
564 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

Whether hCG action is mediated through similarreceptor features during pregnancy is unknown. Astudy evaluating whether the LHCGR kinetics of in-ternalization is linked to LH- and hCG-specifictreatment using fluorescent microscopy failed tofind any differences in human primary granulosalutein cells (). However, the existence of hormone-specific LHCGR trafficking cannot be excluded, be-cause hCG displays greater potency in b-arrestin recruitment compared with LH, at least in thetransfected HEK cell line ().
The analysis of LHCGR-mediated signaling cas-cades provides a complex picture of intracellularevents occurring with LH and hCG binding to thisreceptor, suggesting that hormone-specific signalsmight occur at different levels and result in a refined,cell-specific modulation of the biological effect.
The steroidogenic pathwayModulation of the steroidogenic response is of crucialrelevance for the preservation of different physiolog-ical functions, such as gametogenesis and pregnancy,when estrogen, androgen, or progesterone variabilityoccur as major stage- and sex-specific products.Therefore, LH and hCG might be linked to differentcontrols of the steroidogenic pathway, regulatingdifferent and specific functions. Differences in in-tracellular signaling might be induced by LH or hCGbinding to LHCGR, likely depending on hormone-specific conformational changes at the receptor level(). Because these spatial changes affect the in-tracellular portions of LHCGR, resulting in G-proteinactivation, it is plausible that LH and hCG treatment islinked to activation of hormone-specific patterns ofsignaling pathways. Mechanistic in vitro experiments,performed using bioluminescence resonance energytransfer technology and transfected cell models,revealed lower LH- than hCG-dependent levels ofGaq protein activation and intracellular Ca+ increase.In contrast, no differences in Gas protein subunitrecruitment and formation of dimeric or oligomericcomplexes were observed (). These data indicatethat the hormone-induced conformation of the re-ceptor affects activation of LHCGR intracellularinteractors and downstream signaling pathways.
Results indicative of different steroidogenic po-tentials related to LH and hCG were provided by theevaluation of cAMP production in human primarygranulosa luteal cells (), naturally expressing LHCGR.Dose–response experiments showed that hCG is ap-proximately five times more potent than LH in in-ducing cAMP production, a result exacerbated by thedifferent EC values of the two hormones (~ pMfor hCG and pM for LH). This result was laterconfirmed in transfected COS- and HEK cells (,) in the mouse tumor Leydig MLTC cell line ()and in goat granulosa cells () and mouse Leydigprimary cells (), in which hCG exhibited greater
potency than LH, despite different EC values fromthose observed in human primary granulosa cells,likely indicative of cell-specific LHCGR expression andcoupling to intracellular interactors. Equipotent,nonsaturating concentrations (EC) of both gonad-otropins induced similar plateau cAMP levels, reachedin ~ hour but, following, however, different kinetics(). In particular, the LH-induced cAMP response israpid and reaches a plateau after minutes. Incontrast, hCG treatment requires more time, sug-gesting the existence of different regulatory mecha-nisms underlying the steroidogenesis mediated byLHCGR bound to each ligand. However, the trans-lation of in vitro data into physiology is not immediateowing to the several perturbing factors present in vivo,which can lead to biased evaluations and affect datainterpretation. For instance, the addition of FSH to LHand hCG in vitro resulted in a fivefold increase inpotency of hCG in inducing cAMP production but didnot have any effect on the LH-specific response inhuman granulosa cells (). This finding highlightsthe relevance of the coexistence of LH and FSH duringthe follicular phase of the menstrual cycle, whichshould not alter the FSH-mediated steroidogenicsignal. However, the addition of hCG to FSH ad-ministered during COS cycles might not lead to similareffects at the molecular level.
cAMP recruitment reflects downstream pCREBactivation and STARD gene expression, which ismore sustained with hCG treatment, in human pri-mary granulosa cells () and potentiated by FSHcotreatment (). The STARD gene encodes thesteroidogenic acute regulatory protein enzyme mod-ulating cholesterol transport into mitochondria, a rate-limiting step for steroid synthesis. Because these resultswere corroborated by the greater activity of the CRE-reporter gene after hCG, but not LH, stimulation, inboth MLTC and transfected HEK cell lines (),the higher levels of cAMP/PKA pathway activationobtained with hCG vs LH treatment in different cellmodels were likely driven by intrinsic characteristicsof the ligand–receptor complexes, rather than cell-specific intracellular enzymatic environments. Thesedata, therefore, strengthen the hypothesis that hCGhas greater steroidogenic potential than LH, reflectingthe role exerted during pregnancy by CGs in sup-porting massive progesterone production. In contrast,the dual—proliferative and androgen-stimulating—rolesexerted by LH in granulosa and theca cells, respectively,might not require induction of the steroidogenicpathway at the levels necessary in pregnancy. It isreasonable that androstenedione synthesis by theca cellsis mainly supportive for conversion to estradiol, and itslevels are similar for the whole antral phase ().Therefore, high androgenic potential should notbe required to support follicle growth. Long-term( to hours) treatment of granulosa cells withLH resulted in greater CYPA gene expression
“LH and hCG retain differentsteroidogenic potentialsmodulated in a cell-specificmanner.”
565doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

compared with hCG treatment (), although nodifferences were found in the short-term ( to hours)(). The positive action of LH on CYPA geneexpression, which encodes the aromatase enzyme, isconsistent with the requirement for suitable estrogenproduction to support ovarian follicle growth. Incontrast, LH steroidogenic activity should be focusedon oocyte growth by converting progesterone to an-drogens and eventually to estrogens and on transientmaintenance of the corpus luteum, actions requiringonly limited progesterone production as a basal sub-strate for androgen synthesis. In contrast, the role ofhCG in sustaining pregnancy is exerted throughmassive progesterone production, which could beexacerbated in granulosa cells by high levels of cAMP/PKA pathway activation, at least in vitro ().
Because the collection of sufficient primary thecacells and the development of stable theca cell lines() suitable for in vitro experiments are challenging,in vitro evaluation of androgen synthesis is oftenperformed using adrenal or Leydig cell lines. In themouse Leydig tumor-derived MLTC cell line, pro-gesterone dose–response curves produced by LH andhCG treatment resulted in lower EC and higherplateau level achieved with hCG compared with LHtreatment. However, the testosterone dose–responsecurves are similar and differ only from the LH andhCG EC values, reflecting the hormone potencyinferred by cAMP data (). In this cell model,progesterone is a precursor of testosterone, and itssynthesis is strictly connected to steroidogenic acuteregulatory protein activity, the gene expression ofwhich is activated more by hCG than by LH, therebyexplaining the greater hCG-induced plateau level ofprogesterone. Although these data were obtained inLeydig cells, they corroborated hCG function inpregnancy, suggesting that the steroidogenic role ofhCG is focused on the control of efficient, massiveprogesterone production, which could be useless, oreven counterproductive, during the follicular phase.Instead, in the Leydig MLTC cell line, LH- and hCG-mediated testosterone production was similar andreflected the cAMP data, suggesting that synthesis ofthe final steroid (testosterone) is regulated by quan-titative stimuli and substrate availability, with no needfor qualitative control of earlier events.
Scientific studies have provided several datacomparing the effects of pituitary LHs and chorionicgonadotropins of humans and other mammals (,), based on the promiscuity of glycoprotein hor-mones and their receptor systems across the phyla.The differences in cAMP production induced byhuman LH and hCG and mediated by rodent Lhr (,) match those found in human granulosa cells (,). However, downstream LH- and hCG-inducedpCREB, Stard gene expression, and testosteronesynthesis are all highly similar in mouse primaryLeydig cells in vitro (). This effect might be the result
of a balance between stimulatory and inhibitory sig-nals obtained by treatment with human gonadotropinsthrough the cAMP/PKA and ERK/ pathways. Mostimportantly, these data indicate that only a quanti-tatively, but not qualitatively, different intracellularresponse occurs in rodent primary Leydig cells. Thislikely results from their enzymatic equipment, capableof triggering testosterone synthesis such as an “on/offswitch,” more than in discriminating between ste-roidogenic and proliferative signals, such as occurs ingranulosa cells. This concept might find support inobservations recently described in a case report. In ahypogonadal, hypophysectomized man sequentiallytreated by LH and hCG (), both gonadotropinsshowed similar efficacy in inducing testosteroneproduction.
The inconsistency between human granulosa androdent Leydig cells might have resulted from differ-ences in amino acid sequences between LHCGR andmouse Lhr. They share only % identity at the hingeregion (), which is responsible for discriminatingbetween LH- and hCG-mediated signaling (), sug-gesting that the evolutionary divergence between thetwo species impedes the qualitative discrimination ofintracellular signals mediated by human hormonesdespite effective receptor binding. These data providerelevant insights for the so-called Van Hell in vivobioassay (), currently used for inferring gonado-tropin dosages for clinical purpose in humans by onlyevaluating testosterone-dependent endpoints (,). In vivo bioassays aim to assess the biologicalactivity, measured in terms of organ weight gain, ofpharmacological preparations injected into living ro-dents. However, these assays might not be appropriatefor detecting the full spectrum of gonadotropic bio-activity mediated by LHCGR in human cells.
Taken together, LH and hCG retain differentsteroidogenic potentials modulated in a cell-specificmanner. Although a qualitative discriminatory capa-bility between LH- and hCG-mediated signals isdisplayed by granulosa cells, steroidogenesis, mainlyandrogen production, in Leydig and theca cells isregulated by quantitative signals. These data highlightthe specific roles for each gonadotropin in their targetcells, providing experimental evidence that LH andhCG are not equivalent.
Proliferative and proapoptotic signalsProliferative signals control antral follicle growth inthe presence of both FSH and LH (), and hCGphysiologically acts on LHCGR-expressing cells ded-icated to progesterone synthesis. Pregnancy is alsocharacterized by angiogenic events, consistent with cellproliferation, and possibly mediated by hCG-H,growth factors, and steroids. It is, however, plausiblethat LH and “classical” hCG differentially affect cellproliferation, given the specific molecular propertiesrequired to optimize follicle growth and massive
566 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

progesterone production. Discrimination of qualita-tively different LH- and hCG-mediated signaling byLHCGR was demonstrated in human primary gran-ulosa lutein cells by assessing activation of phospho-proteins. Both gonadotropins showed maximumactivation of ERK/ and AKT pathways at a con-centration of pM, resulting, however, in ahormone-specific phosphorylation pattern of thesekinases (). LH treatment resulted in greater and moresustained pERK/ and pAKT activation comparedwith hCG (, ), reflecting the proliferative andantiapoptotic roles (, ) exerted by LH during theantral phase of follicular development. Moreover, therange of effective LH concentrations is widened bythe presence of FSH combined with LH, but not withhCG, confirming that gonadotropin-specific activitiesare potentiated by FSH cotreatment (). LH andhCG displayed different ratios between the cAMPEC and the maximally activating concentrationsrequired for pERK/ and pAKT activation (~: forLH and ~: for hCG) in granulosa cells. These datasuggest that LH has greater proliferative and anti-apoptotic potential than hCG in granulosa cells invitro. This is an example of biased signaling, consistingof activation of different intracellular endpointsevoked by varying concentrations and ligands.
The expression of proliferative and antiapoptoticgenes in granulosa cells reflects hormone-specificsignal transduction. The expression of the growth-promoting AREG gene (), which encodes EGF-similar amphiregulin, and of the CCND gene(), encoding cell cycle regulator, G/S-specificcyclin-D, is upregulated more by LH than by hCG. Incontrast, the high steroidogenic potential of hCG,which relies on efficient cAMP production, is linkedto expression of proapoptotic genes in primarygranulosa cells in vitro, especially in the presence ofFSH. Although both LH and hCG elicited increasedTP proapoptotic gene expression, the procaspase-encoding gene CASP was positively modulated byhCG treatment, although LH treatment failed toobtain similar results, likely due to upregulation of theantiapoptotic X-linked inhibitor of apoptosis protein-encoding gene XIAP (). This finding should beconfirmed in other cell types. However, gene ex-pression data supported a dose-dependent cell viabilitydecrease in human granulosa cells maintained for to hours under hCG stimulation in vitro. However,LH resulted in the opposite effect by counteracting celldeath (, ). Similar conclusions were drawnfrom a study comparing the long-term effects of LHand hCG in goat granulosa cells in vitro. AlthoughhCG induced relatively high cAMP intracellular levelsand reduced cell viability, LH treatment resulted inmarked phosphorylation of ERK/ and an increasedrate of cell proliferation ().
Whether hCG is capable of mediating proapoptoticeffects in vivo, at least during the follicular phase of the
menstrual cycle, should be investigated in-depth, be-cause the molecule was associated with both pro-liferation and inhibition of cancer cell growth (,) and given the proliferative role exerted duringplacentation. It is plausible that hCG-dependent lifesignals are cell specific and sensitive to a particularhormone isoform or glycosylation variant ().However, the proliferative and antiapoptotic signalsshould be transmitted through the ERK/ and AKTpathways (, ), which are preferentially activatedby LH, rather than hCG, at least in human primarygranulosa cells in vitro. hCG displayed greater effi-ciency in recruiting b-arrestin compared with LH,which has a -fold greater EC (hCG EC ~ nM,LH EC nM) and even acts as a partial agonist,not reaching hCG-dependent plateau levels ().Although these data were obtained using the MLTCcell line, they are likely to also be valid in humangranulosa cells, because the recruitment of b-arrestinsis dependent on GPCR activation by agonists ().Most importantly, these data reflect the lower efficacyand efficiency of LH compared with hCG in inducingprogesterone production, indicating that b-arrestinsare involved in steroidogenesis, as demonstrated bythe decreased progesterone levels on depletion ofb-arrestins by small interfering RNA probes ().b-Arrestins are involved in ERK/ phosphorylationand GPCR internalization (), proliferative signalsin granulosa cells (). It is conceivable that relativelyhigh amounts of LH would be required to down-regulate the LHCGR-mediated steroidogenic andproapoptotic signaling (), owing to the less efficientcAMP production induced by LH compared withhCG. These data support the idea that LH has greaterproliferative potential than hCG, at least in granulosacells, reflecting its physiological function as a folliclegrowth regulator (Table ).
Cross-talk between gonadotropin- and steroidhormone-mediated signalingThe effect of LH and hCG on signal transductionmight be precisely revealed by in vitro experiments, inwhich the perturbations and interactions betweensignaling pathways activated by different hormonesand paracrine factors are missing. Proapoptotic effectslinked to the addition of hCG were poorly reproducedin ART cycles in vivo and showed weak effects or evenantithetical results (, ). This concept is also validfor cancer cell growth (, ), which can be bothpositively or negatively susceptible to the presence ofthe hormone, depending on the cell type. Therefore,intracellular effects clearly dissected in vitro could bemasked in vivo.
In the gonads, LH and hCG induce steroid hor-mone production, activating the same pattern of in-tracellular signaling cascades. Steroid compounds,such as glucocorticoids, have been associated with theproliferation of granulosa cells and protection against
“Intracellular effects clearlydissected in vitro could bemasked in vivo.”
567doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

cAMP- and p-induced apoptosis (). Thesemolecules act through activation of pAKT andpERK/ counteracting intracellular death signals.During the follicular phase of the menstrual cycle,gonadotropin functions converge in the production ofprogesterone, which is further converted to androgenin theca cells. Androgen is transferred to granulosacells, where it is transformed to estrogen, leading topotent proliferative effects and inducing follicle growthin vivo. Estradiol activates the AKT pathway in vitro(), and estrogen receptor-b knockout mice arecharacterized by impaired follicular development(). It is reasonable that estradiol could counteractthe proapoptotic signals mediated by high cAMPintracellular levels, obtained during treatment by FSHand hCG administration in COS. This concept wasconfirmed in vitro by adding estradiol to hCG or LHtreatment of human granulosa cell cultures (). Theeffect of the addition of steroid predominates com-pared with that of either LH or hCG alone, triggeringhigh levels of pAKT activation and preventing hCG-dependent procaspase cleavage and decreased cellviability. This molecular mechanism could explainwhy the addition of hCG to FSH in COS for ART hasbeen associated with multiple follicular development,reasonably sustained by high estrogen levels andavoiding natural follicular atresia.
Progesterone is the main steroid produced duringthe luteal phase and by human primary granulosalutein cells in vitro, and it is responsible for theprotection from apoptosis linked to hCG treatment(). In contrast, it is well known that both LH andhCG, as well as progesterone, promote ERK/ andAKT phosphorylation. They also induce expression ofAREG and the epiregulin-encoding gene EREG, twoEGF-like ligands with positive effects on granulosalutein cell viability in vivo and preservation of thecorpus luteum in primates () and rodents (). Thesame intracellular pathways lead to the growth ofprogesterone receptor-expressing cancer cells (),strengthening the evidence for proliferative andantiapoptotic effect mediation by steroid hormonesthrough activation of signaling cascades common togonadotropins. These data support the tumorigenicpotential described in hCGb-overexpressing transgenic
mice, in which human gonadotropin and highlevels of progesterone can assist in the developmentof multiple cancers (, ). However, becauseextragonadal tumors were totally abolished byovariectomy in these mice, it is plausible that thetumorigenic potential of hCG is exerted throughaberrant ovarian functions, rather than by a directgonadotropin effect (). The apoptotic effect seemsto be directly dampened by hCG treatment in Leydigcells (), in which the signals induced by both LHand hCG converge on a similar balance betweencAMP production and ERK/ phosphorylationand downstream steroidogenesis (), suggesting thedifferent nature of LHCGR-mediated signalingwithin the testis and ovary (Fig. ).
Polymorphisms and Mutations
Several single nucleotide polymorphisms (SNPs) andmutations occurring within gonadotropins and theirreceptor genes have been described (, ). They canmodulate or impair the hormonal response or receptorfunction, affecting reproductive function or leading todisease. Generally, spontaneous mutations occur asrandom events and are silent, not resulting in aminoacid changes at the protein level. Some, however, causechanges in the amino acid sequence and are linked toclinical phenotypes. In contrast, SNPs result in mildphenotypes without strong repercussions on the re-productive success of individuals. They have a fre-quency of $% in a given population and contributeto the endocrine ethnic background. Most SNPs andmutations affecting LH and hCG signaling are carriedby LHCGR ().
Among the several SNPs and mutations occurringwithin the LHB/CGB gene cluster, only a few havebeen associated with specific phenotypes. A betterunderstanding of the consequences of LHb and CGbmutations was provided by the study of transgenicmice models. The LHB gene knockout male micepresent with hypogonadism, with low testosteronelevels and hypoplastic Leydig cells, but have normalserum FSH levels; the female mice are infertile andfeature anovulation and degenerated antral follicles
Table 3. Effective Concentration for cAMP, pERK1/2, and pAKT Activation in Primary Mouse Leydig and HumanGranulosa Cells
Primary Cell Type MoleculecAMPEC50, pM
pERK1/2ECmax, pM
pAKTECmax, pM
cAMP/pERK1/2Ratio Ref.
Human granulosa LH 530 6 51 100 100 5:3 (6)
hCG 107 6 14 100 NA 1:1 (6)
Mouse Leydig LH 192 6 54 100 NE 1:9 (8)
hCG 18 6 10 10 NE 1:8 (8)
Abbreviations: ECmax, minimalconcentration maximallyactivating; NA, not activating;NE, not evaluated/assessed; Ref.,reference.
568 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

(). The mouse phenotype partially matches theclinical observations in humans, in whom LHB genemutations lead to a severe, eunuchoid phenotype inmales (). Therefore, mouse models might notprovide a full comprehension of human LH physi-ology. Administration of exogenous FSH is enough tosustain follicular development and ovulation in hy-pophysectomized female mice, and this hormone failsto do the same in Lhr knockout mice (). Thesefindings support the concept that the presence ofgonadotropin receptors, rather than LH, is required tosustain gametogenesis in mice. Transgenic miceoverexpressing both the CGB and the CGA subunitswere expected to replicate the human phenotypelinked to activating mutations of LHCGR (i.e.,asymptomatic women and men developing precociouspuberty and testicular tumors). However, these micefeatured the opposite phenotype, with a normalphenotype in males, and precocious puberty, obesity,and luteinized ovaries with luteomas and hemorrhagiccysts present in females (). However, althoughtransgenic mice might not be fully representative ofhuman physiology, their study has provided new in-sights to comprehend LH/hCG- and LHCGR-specificfunctions emerging from clinical data involving SNPsand mutations (described in the next sections: “LHband hCGb polymorphisms”; “LHb and hCGb mu-tations”; and “Receptor mutations and polymorphicvariants”).
LHb and hCGb polymorphismsFew LHB and CGB gene polymorphisms were studiedin conjunction with their clinical phenotype. Overall,they do not severely affect fertility, and most lackmolecular characterization in vitro, with no clear in-sights provided of their suggested effects in vivo. Oneexception is that provided by the most common LHBgene variant (V-LH), which was discovered in theFinnish population and displays a double polypeptidechange (i.e., tryptophan to arginine at position andisoleucine to threonine at position of the aminoacid chain) (, ) and introduces a glycosylationsite (), hiding the molecule from a specific anti-LHantibody (). V-LH exhibited a reduced serum half-life and bioactivity in vivo compared with wild-typeLH and decreased receptor binding activity and po-tency for progesterone production in vitro () andinduced preferential IP-related signaling, rather thancAMP/PKA (). Perturbation of the signaling cas-cade by V-LH might affect granulosa cell survival andfollicle development. In cumulus cells of heterozygouswomen undergoing ART, high levels of apoptoticmarkers were found, such as the DNA fragmentationindex and cleaved caspase-, and they negativelyinfluenced the success rate of intracytoplasmic sperminjection procedures (). However, increased ex-pression of this variant, identified by SNPs within thepromoter region in linkage disequilibrium, resulting in
© 2018 Endocrine Reviews ENDOCRINE SOCIETY
cAMP/PKA pERK1/2
Cholesterol
Dehydroepiandrosterone
Androstenedione
Testosterone
Theca cell
Leydig cell
Granulosa cell
Steroidogenic pathway cAMP/PKA-pathway
Cholesterol
Dehydroepiandrosterone
pERK1/2, pAKT
Progesterone Apoptotic genes
Cholesterol
Apoptosis
Androstenedione Estrone
Testosterone Estradiol
EGF-like andanti-apoptotic
genes
Cell proliferation
LHCGR-LHCGR homodimer
FSHR-LHCGR heterodimer
FSHR-FSHR homodimer
LH-dependent signal
FSH-dependent signal
hCG-dependent signal
Figure 4. Comparison of LH- and hCG-mediated signaling in the ovary and testis. At the midantralfollicular phase, granulosa cells express both FSHR and LHCGR capable of forming homodimers/heterodimers, with theca cells expressing only LHCGR. In granulosa cells, hCG displays greatersteroidogenic potential than LH, exerted via relatively high levels of cAMP/PKA pathway activation,and this feature is potentiated in the presence of FSH. hCG action is exacerbated by massiveproduction of progesterone, which is converted to androstenedione in the theca cell, andtestosterone is a minor product in the gonads of the female. Androgens serve as a substrate for thearomatase enzyme, which converts them to estrogens with high proliferative and antiapoptoticpotential. Intracellular cAMP increase is linked to proapoptotic stimuli, exacerbated in vitro by theabsence of theca cell-derived substrate for estrogen synthesis. LH displays lower potency than hCGin terms of cAMP/PKA pathway activation, resulting in relatively low steroidogenic andproapoptotic potential. LH signals are preferentially exerted via phosphorylation of ERK1/2 andAKT after the recruitment of G protein and b-arrestins and resulting in proliferative/antiapoptoticevents. LH-specific signals are potentiated in the presence of FSH, which reasonably provides themain steroidogenic stimulus in granulosa cells. Theca cell androgenic potential increases, togetherwith the progression of antral follicle growth and the amount of LHCGR expression, providingsufficient androstenedione to be converted to estradiol. To date, no in vitro studies have comparedthe action of LH and hCG in theca cells. Because Leydig cells are androgenic and exhibit LHCGRexpression, they can be considered the male counterpart to theca cells. hCG is more potent than LHin inducing both cAMP/PKA and pERK1/2 pathway activation but results in a qualitatively similarbalance of stimulatory and inhibitory steroidogenic signals and downstream testosterone synthesis.
569doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

~% greater activity compared with that of the“normal” LHB promoter, compensates for the weakerhormone bioactivity (). The frequency of V-LHwas lower in obese women affected by polycystic ovarysyndrome (PCOS) than in healthy women and non-obese women with PCOS, suggesting that V-LHmightprovide protection from developing symptomaticPCOS in obese women (). Nonetheless, this LHvariant was associated with infertility in homozygousJapanese women (), although Baltic V-LH carriermen affected by idiopathic infertility had higher serumLH levels than healthy men (). V-LH was foundin a -year-old man affected by the fertile eunuchsyndrome (). This patient displayed hypogonadismand normal responses to treatment with exogenousGnRH and hCG. Overall, however, the ethnicity-related clinical features displayed by V-LH and themild phenotypes suggest that it simply represents anexample of phenotypic variations due to geneticpolymorphisms as the basis of human diversity.
An SNP (rs) in exon of the LHB gene wasfound more frequently in South Indian women withPCOS (). This SNP is characterized by the syn-onymous amino acid change “T” to “C” at position of the gene sequence. Its contribution to PCOSpathogenesis is unexplained. It is conceivable that thepolymorphism might affect the expression or func-tions of other molecules, because it is within thepalindromic RUVBL gene sequence, encoding for aprotein interacting with the activating transcriptionfactor (). Several other SNPs within or close to theLHB gene sequence were associated with centralprecocious puberty () or infertility () in women.However, their contribution in defining the endocrinephenotype should be evaluated with other polymor-phisms related to hormones and their receptors.Moreover, these results should be independentlyconfirmed in other populations and supported byfunctional in vitro data, which are still lacking.
Among the CGB gene SNPs and mutations pre-viously described in association with miscarriage ()and discussed in the section “hCGb isoforms andglycosylation variants,” a polymorphism within theCGB gene displayed inefficient assembly whencotransfected with the CGA gene in the CHO cell line.This naturally occurring variant is associated withinfertility in women and is characterized by a valine tomethionine exchange at codon (). Seventy-onehCGb variants deriving from CGB and CGB genesmight be predictive of recurrent miscarriage in Eu-ropean populations (), suggesting the relevance ofthese transcripts in sustaining pregnancy. Moreover,because the heterozygous haplotypes calculated forthese SNPs are relatively frequent, they might besubjected to balancing selection in Europeans. Overall,the associations between CGB gene SNPs and mis-carriage are relatively rare, and the molecular mech-anisms supporting the clinical data are unclarified.
In general, such as in the case of SNPs andcommon variants in other genes, SNPs in LHB andCGB genes are unlikely to be major determinants ofdisease. Rather, they contribute to human phenotypicvariation and, together with other SNPs, might berelevant in discrete genomic clusters associated withparticular reproductive problems, such as PCOS ().
LHb and hCGb mutationsPossibly because of the physiological role played ina specific time-window and in one sex only, inacti-vating hCG mutations are rare and, presumably, in-compatible with successful pregnancy. One of thefirst mutations affecting hCG function was somaticand was found in the a subunit secreted by un-differentiated carcinoma cells of the femoral region(). This mutation consists of a substitution ofglutamic acid by an alanine at position , whichchanges the hydrophobic profile of the molecule,resulting in misfolding and impairment of di-merization with the b subunit. Similar effects at themolecular level have been associated with a genomicCGB gene mutation, leading to a valine to leucineswap at position of the b subunit (). Themutated hCGb is only capable of % dimeric as-sembly yet shows increased potency in inducing cAMPproduction. Nevertheless, enhanced steroidogenicsignaling was not sufficient to replace the loss offunction due to impaired dimerization, because thismutation was found in a Northern European pa-tient affected by recurrent miscarriages. CGB genemutations have also been described and found inindividuals of the same population (), albeitwithout a link to pathogenic phenotypes. A proline toarginine change in position resulted in a twofoldreduction in b-subunit secretion without affecting itsbiological activity, and no effects were associated withan arginine to tryptophan substitution at position .Altogether, these data indicate that genomic mutationswithin the genes encoding for the two major hCGtranscripts result in mild consequences that can betolerated.
Although mutations in the CGB genes pre-dominantly result in miscarriage, mainly due tomisfolding of the b subunit and impaired dimerformation (), LHB gene mutations result in phe-notypes featuring hypogonadism, decreased or im-paired spermatogenesis, delayed puberty, and lowtestosterone levels in males and amenorrhea in fe-males. Such symptoms might be associated with in-fertility in both sexes and might be due to disruption ofLHB splice sites () or in the signal peptides ().Mutations disrupting the cystine knot motif, such as aglycine to aspartic acid mutation at position mightresult in defective heterodimer formation, undetect-able serum LH levels, and hypogonadism (). Asimilar phenotype was described in a patient carrying alysine deletion at position of LHb, which impaired
570 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

release of hormone dimers (). Because mutationswithin the LHB gene do not impair receptor function,long-term administration of exogenous hCG to thesepatients induces virilization, testicular growth, tes-tosterone synthesis, and spermatogenesis, in con-junction with fertility (). These data strengthen thehypothesis that LH and hCG could be equivalent ininducing proper Leydig cell function and fertility,which mainly relies on testosterone synthesis in malesand might be switched on by an on/off molecularmechanism, rather than qualitatively different hor-monal signals (). However, it cannot be excluded thatsignaling cascades are maximally activated by lowamounts of occupied receptors, confirming previousfindings in rodent Leydig cells in vitro (). Certainmutants of LHbmight display residual in vitro activity,sustaining low testosterone synthesis but enough tosupport normal spermatogenesis and fertility ().Studies of siblings carrying the same LHB mutationsthat totally disrupt LH signaling revealed different, sex-specific clinical effects. Although normal pubertalmaturation might be conserved in women, this is notthe case in men (), suggesting that, in women, theregulation of estrogen production needs to be sus-tained by FSH acting on granulosa cells, supported bybasal androgen production originating from thecacells. Moreover, in the ovary, LH-mediated signalsmight plausibly be driven by LHCGR–FSHR heter-odimers activated by FSH at the onset of puberty andduring early antral follicular stages (), althoughLeydig cells are the only LH target and source of theproper testosterone levels required to support malesecondary sex characteristics. Nevertheless, inactivatingmutations of LHb subunit have been associated withovulatory disorders (), suggesting the fundamentalrelevance of proper LH signaling for the ovarian cycle.
Receptor mutations and polymorphic variantsAlthough several SNPs were found within the LHCGRgene, their contribution in determining clinical phe-notypes is mostly weak. An exon SNP, p.NS(rs), was found in association with sper-matogenic damage and is highly prevalent amonginfertile male patients (). However, although thenumber of LHCGR marker SNPs affecting male re-productive function is very low, it is remarkable thatthe very gene is a genetic hot spot for PCOS in the HanChinese population ().
A common polymorphism of LHCGR consists ofthe addition of two amino acid residues, a leucineand a glutamine, at codons and within exon ,originating from a CTCCAG insertion at positions- of the gene (). This receptor variant, knownas “insLQ,” is common among white populationsbut absent in other populations, such as the Japanese.Although the double amino acid residue insertiondoes not severely affect LH- and hCG-mediated cAMPproduction, the polymorphism has been linked to a
more active signal peptide and to adverse outcomes inpatients with breast cancer ().
The LHCGR gene mRNA variants comprise highlyexpressed, primate-specific transcripts, including acryptic exon located between the sixth and seventhexon with unknown function (). This exon isnamed “A” and is responsible for three differentmRNA receptor variants differing in the length basedon the location of stop codons. The resulting mRNAvariants, in addition to the “classical” LHCGR, includetwo truncated forms consisting of exons to and oneA of different length and a full-length transcript withexon A between exons and . Mutations within thisregion are causative of aberrant gene transcription,leading to Leydig cell hypoplasia type II (). Inaddition, SNPs in exon A were associated with tes-tosterone levels in male infertile patients (). Inparticular, the levels of this hormone were more el-evated in G homozygous men than in those carryingthe T allele of the exon A SNP rs. Mutationsof this cryptic exon were discovered for first in ,XYpatients affected by sex development disorders andcharacterized by a female phenotype, a blind-endingvagina, and primary amenorrhea (). Based oncurrent knowledge of human testicular development,the presence of testicular structures and a phenotypelargely resembling testicular feminization (which isdue to the lack of androgen action), suggest that thepatient was not at all responsive to endogenous LHand maternal hCG. Therefore, it is plausible thatisoforms deriving from exon A could be linked to thediscrimination between hCG- and LH-mediated sig-nals. Because our knowledge of the control of humantesticular development by fetal pituitary LH or ma-ternal hCG is limited and provided by individualsaffected by genomic mutations and data from non-human mammals, the putative role of exon A indiscriminating LH and hCG remains unclear.
Interesting data on LH- and hCG-specific functionswere provided by the deletion of the LHCGR exon–encoded sequence, described in the section “Evolu-tionary convergence: trophoblast LH and pituitary CGs.”As discussed, the mutated receptor is capable of trans-mitting hCG, but not appropriate LH, signals, despitebinding both hormones (), resulting in male hypo-gonadism with a normal male phenotype (). Theclinical evidence derived from naturally occurring mu-tations or deletions of LHCGR exons A and hasdemonstrated that these sequences of the receptor areessential for LH and hCG action and could be in-strumental in discriminating between the two hormones,although the mechanism remains largely unknown.
Pathophysiology of LH and hCG
Specific profiles and levels of LH and hCG could beassociated with pathological conditions, such as
“Extragonadal action of LHand hCG might produceclinical effects.”
571doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

hypogonadism, cancer, and endocrine disorders. Theseeffects could result from excessive or low hormoneactivity, altering normal physiology and leading to awide range of clinical phenotypes. Because gonado-tropins modulate cell growth, it is reasonable to as-sume that their action is linked to the proliferation ofcancer cells and tumorigenesis. Moreover, extra-gonadal action of LH and hCG might produce clinicaleffects. For instance, the pregnancies of fetuses affectedby trisomy are typically characterized by high levelsof hCG in the maternal serum, despite the low hCGsynthesis capability by the placenta (). This ap-parent paradox might be caused by high activity ofsialyltransferase- and fucosyltransferase- enzymes intrisomy trophoblast cells, resulting in highly gly-cosylated and acidic hCG molecules, displaying re-duced activity in vitro. hCG synthesis peaks at~ weeks in trisomy pregnancies and declines tolower levels than in chromosomally normal preg-nancies. Regardless, because no association betweensialylated hCG isoforms and trisomy pregnancy wasfound (, ), although fucosylation was not in-vestigated, any conclusions should be interpretedcarefully. However, highly glycosylated molecules aremore persistent owing to their increased half-life ().Moreover, high hCG levels during pregnancy couldinduce aberrant expression of LHCGR in the adrenalglands of the mother, resulting in the increased risk ofadrenal hyperplasia and transient Cushing syndromedue to cortisol release in response to the hCG-inducedsignals (). Several other clinical effects related toaberrant LH and hCG signaling (i.e., hypogonadism,precocious puberty, PCOS, miscarriage, and cancer,are described in the following subsections).
Hypogonadism and precocious pubertyHypogonadism is a pathology characterized by de-creased gonadal activity and hormone productionand can be caused by impairment of LH-mediatedsignaling (). In males, the disease can arise fromthe testis (primary) or from dysfunction of thehypothalamic–pituitary unit (secondary or centralhypogonadism) (), although the cause for severalpatients with hypogonadism will remain idiopathic,suggesting a possible polygenic nature (). Thephenotype associated with an impaired HPG axismight vary depending on the severity of the disease.Most severe cases can be linked to rare autosomalrecessive conditions, such as inactivating LHCGRmutations, typically interfering with the developmentof male external genitalia and testicular descent andresulting in phenotypically female, ,XY patients withLeydig cell hypoplasia type . The disease of thesepatients will be unresponsive to both endogenousLH and exogenous hCG administration and is char-acterized by primary hypogonadism and sexualdifferentiation disorder, featuring the absence ofLeydig cells, the lack of masculinization and pubertal
maturation with a female-like phenotype and externalgenitalia. Milder phenotypes could be linked to LHBmutations affecting hormone function, resulting ininfertile individuals with male external genitalia andmaldescended testes, a micropenis, and/or hypospa-dias. Given the presence of a functional LHCGR, hCGtherapy showed efficacy in inducing testosteroneproduction and might restore fertility. The clinicalpicture is strictly connected to testosterone levels,insufficient to support male sexual development. Thetotal blockade of Leydig cell function is largely at-tributed to mutations impairing receptor transportto the cell membrane () and activation of theG-protein–dependent signaling cascades (). How-ever, mutations affecting LH binding have also beenreported (). An inactivating homozygous mutationwas found in a ,XY hypogonadic patient withdelayed puberty, a normal prepubertal male pheno-type, and descended testes, displaying a glutamineto arginine substitution at codon of LHb ().The resulting b subunit was capable of forming aheterodimer with the a subunit but failed to bind thereceptor in vitro. This patient was treated with long-term hCG administration, resulting in testicular en-largement, virilization, and spermatogenesis. Similarphenotypes and histories were found in consanguin-eous, hypogonadic patients bearing the deletion oflysine at position of the LHb, linked to intracellularretention of the hormone (). These data allow forthe comparison of the effects of mutations impairingLHCGR or LHb functions in the male. Although thefirst are linked to severe, female-like phenotypes, LHbmutations are compatible with the male phenotypeand exogenous hCG-induced spermatogenesis ().Taken together, these findings suggest that maternalhCG, together with the presence of intact LHCGR,can, at least in part, compensate for the absence of fetalpituitary LH and support the development of a malephenotype, although the presence of maldescendedtestes and a micropenis suggest that proper productionof functional LH molecules is required to fully supportsecondary sex characteristics.
Inactivating mutations within the LHB gene havebeen described in infertile females, in whom thephenotype displayed normal external genitalia andspontaneous breast and pubic hair development atpuberty. Menarche was delayed or even normal;however, these patients were oligo-amenorrheic andcharacterized by failure in achieving ovulation andnormal LH, estradiol, and progesterone levels in theovulatory or luteal phase. Because women with LHBmutations have functional LHCGRs, they can suc-cessfully be treated with LH or hCG (), in contrastto hypogonadic women with LHCGR-inactivatingmutations, which will be unresponsive to gonado-tropin treatment.
Precocious puberty is defined as the onset of pu-berty at a relatively young age and can result from
572 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

several factors, such as hypothalamic or pituitarydysfunction, McCune-Albright syndrome, or sexhormone–secreting tumors. LH signaling is one of thefactors regulating the onset of puberty. However, al-though LHCGR-activating mutations resulting insustained tonic cAMP production due to an aber-rant ratio between the Gas (stimulating) and Gai(inhibitory) protein activation (), were linked tofamilial, male-limited, precocious puberty, no phe-notype has been observed in females. No LHb mu-tations are known to cause excessive LH activity.
Polycystic ovary syndromePCOS is a common endocrine disorder affecting % to% of women worldwide and defined as the co-existence of at least two of three typical features:polycystic ovaries, ovulatory dysfunction, and hyper-androgenism (). Other symptoms can occur inassociation with the disease, such as type diabetes,metabolic syndrome, adrenal dysfunction, obesity,and/or insulin resistance (). Because PCOS haslikely maintained an overall constant prevalence overcenturies, albeit linked to anovulatory subfertility, ithas been addressed as an evolutionary paradox (,). Intralocus sexual conflict was proposed to ex-plain the persistence of genetic loci linked to increasedreproductive success in males in conjunction with arisk of developing the disease in females (, ).PCOS has a polygenic nature, and genome-wide as-sociation studies found both FSHR and LHCGR genesto be two of several hot spots for the disease (, ),suggesting the relevance of gonadotropin signalingin its pathogenesis. However, endocrine-disruptingchemicals modulating sex hormone–dependent sig-nals might be associated with the disease ().Hyperandrogenism is a major determinant of thedisease and is indicative of excessive androgens pro-duced by theca cells exposed to relatively high LHlevels. Moreover, the androgen and estradiol responseto FSH stimulation is greater in those with PCOS thanin healthy women, suggesting that molecular mech-anisms regulating paracrine signals between granulosaand theca cells are amplified in individuals carrying thedisease (). As a consequence of the high estrogenlevels, the feedback mechanism regulating pulsatilegonadotropin production is altered in women withPCOS, resulting in low FSH levels, a high LH/FSHratio, and impaired follicle selection and ovulation(). In humans, cyclic gonadotropin productionrecovers after treatment with the estrogen receptorantagonist clomiphene citrate, which temporarily re-stores proper feedback mechanisms at the pituitarylevel (). Polycystic ovaries and adrenal disturbancesare recapitulated in the phenotype of transgenic, LH-overexpressing female mice, supporting the role playedby this hormone in PCOS pathogenesis (). hCG-overexpressing female mice displayed a slightly dif-ferent ovarian phenotype, characterized by multiple
corpora lutea and enhanced estradiol, progesterone,and testosterone levels, in addition to prolactin-linkedadenomas (). Different phenotypes of female miceoverexpressing LH and hCG strongly support the viewof a different in vivo action of these two gonadotropinsin rodents.
Appearance of the polycystic ovary is one of thecharacteristics that can be used for the diagnosis ofPCOS and consists of the recruitment of several fol-licles reaching the antral stages without completingmaturation. Provided that LHCGR is of central im-portance for the understanding of the disease, thecacell androgenic function and granulosa cell pro-liferation are modulated by different genes (), suchas DENNDA, INSR, and RABB, whose functions areinvolved in modulating proliferative signals throughthe activation of AKT and ERK/ pathways. Theactivity of these kinases is increased in the ovary ofwomen with PCOS (), compatible with exaggeratedLH- and estrogen-dependent stimuli. SNPs within theLHB gene might also be linked to increased PCOSrisk. This is the case for the two SNPs inducing atryptophan-to-arginine change at position and anisoleucine-to-threonine change at position of LHb(), which are in linkage disequilibrium and mightcontribute to elevated testosterone levels in Brazilianwomen with PCOS. Moreover, it was suggested thatthe V-LH variant might have been a contributingfactor in the development of the disease in a Japanesewoman (), although another study failed to find asimilar association in Turkish women (). This issueshould be further investigated and independentlyconfirmed in other populations. Nonetheless, it issuggestive of altered functioning of proliferative andandrogenic LH activity. Most importantly, the gly-cosylation profile of LH molecules might changedepending on the steroid hormone levels, influencinggonadotropin activity (). Although the molecularmechanism underlying the control of biologicallyactive LH isoform production by the pituitary isunknown, age-specific profiles of LH glycosylationwere reported among women with PCOS. Mainlyalkaline LH species were found in adult patients withPCOS (), although basic isoforms featuring high invitro activity were predominant among adolescent girlsaffected by the disease and positively correlated withandrogen levels (). It is plausible that highly bio-active LH isoforms will result in elevated androgenlevels, and this hypothesis is consistent with the in-hibition of steroid synthesis by GnRH antagonists inwomen with PCOS ().
Given their chronic overexposure to “LH activity,”women with PCOS will require particular attentionwhen ovarian stimulation by exogenous gonadotro-pins is required in the framework of ART. Concurrentwith the presence of high estradiol levels and thedevelopment of excess antral follicles, which can leadto an excessive response to gonadotropin stimulation
“Women with PCOS willrequire particular attentionwhen ovarian stimulation byexogenous gonadotropin isrequired in the framework ofART.”
573doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

(), women with PCOS might have an increased riskof developing ovarian hyperstimulation syndrome(OHSS) and multiple pregnancies with ART ().Thus, ovulation induction with a GnRH agonist hasbeen proposed as a better choice than hCG (, ),consistent with the high steroidogenic potential of thechoriogonadotropin, which could be linked to OHSS.However, the matter is still under debate, and theclinical symptom spectrum related to PCOS, togetherwith ovarian morphology, might be a determinant forthe response to hCG exposure (), even in preg-nancy, when the levels of endogenous-free hCGbmolecules differ between healthy women and thosewith PCOS (). A clinical study comparing ARTbetween white and Yoruba women from WesternNigeria, triggered by hCG, revealed higher estradiollevels and the prevalence of symptoms typically relatedto PCOS among African women, who have a greaterrisk of OHSS and twin pregnancies. This findingsuggests that ethnicity could be a determinant forPCOS. The anovulatory condition linked to the diseasemight lead to maintenance of a follicular reserve for alonger time compared with healthy women, and thisconcept has been corroborated by the convergingeffects of genes regulating the age of menopause andLH levels (). PCOS could therefore be viewed as anevolutionary strategy to prolong the fertile window atthe cost of fewer ovulatory cycles (). Althoughsuggestive, this hypothesis should be examined fur-ther, because no evidence of evolutionary advantageswas experimentally demonstrated and subfertilitylinked to PCOS might result in substantial effects inmodern times. Today, especially in Western societies,fertility and pregnancy are sought at a relatively latereproductive female age (), and ovarian stimulationmight be performed in the framework of ART usingcommercial hormone preparations, a setting quitedifferent from a natural cycle in a nonmedicalizedsociety.
MiscarriageMost miscarriages occur within the first trimester ofpregnancy and are mainly due to placental or fetalabnormalities. Embryo aneuploidies are typical causesof miscarriage () and other clinical conditions, suchas diabetes and obesity, could be risk factors for poorpregnancy outcomes (). These data point to theimportance of proper metabolic and endocrinefunction in regulating embryo implantation and de-velopment. The action of LH is central in folliclegrowth and uterine preparation, and hCG is requiredto support pregnancy, with both these aspects affectingpregnancy success. A dual function could also beassumed for LH action. It is linked to estrogen-mediated proliferative events in the uterine epithe-lial cells during the follicular phase, resulting inincreased endometrial thickness, and it becomesmainly steroidogenic during the luteal phase, when the
increase in progesterone levels is associated with thesecretory action in the uterus. These changes in ste-roidogenic potential are accompanied by the - to-fold increase in LHCGR expression, from the earlyantral stage through to ovulation (), and are veryintense in the corpus luteum (). Antral folliclegrowth requires proliferative signals preferentially anddirectly exerted by LH, as demonstrated in vitro, andestradiol, which increases via the cAMP/PKA pathway,together with the receptor number. Thus, estradiol canbe used as a marker for successful pregnancy outcomes(). The maintenance of large LHCGR numbers inthe corpus luteum reasonably predisposes later, massiveprogesterone synthesis induced by LH and, in the caseof pregnancy, hCG. Thus, LH and hCG should ensureproper progesterone levels, which, in turn, will mediatethe events preparatory to, and supportive of, preg-nancy. Women experiencing recurrent miscarriagesmight achieve better pregnancy outcomes throughprogesterone supplementation (). These data il-lustrate the role of both LH and hCG in endometrialpreparation and maintenance of the corpus luteumthrough adequate steroid production. It is reasonablethat endometrial preparation by the addition of LH inART cycles might provide adequate stimuli leading tolower pregnancy loss (), although hCG could notprovide similar activity and leads to different effects inpregnancy outcomes (). As the hormone of preg-nancy, hCG exerts its specific functions during the firsttrimester, resulting in angiogenetic effects and in-creased progesterone synthesis. These roles are irre-placeable, as demonstrated by reports describing theassociation between recurrent miscarriage and mu-tations of the hCGb-encoding genes, which impairsignal transduction and alter the biochemical prop-erties of the molecule (). In women with a suc-cessful pregnancy, the serum and urinary levels of thehormone will be different than those measured inwomen with a miscarriage (). Thus, specific hCGfunctions and support of embryo development alsodepends on hormone levels.
CancerBecause LH and hCG might activate intracellularsignaling cascades regulating cell proliferation andantiapoptotic events, it was suspected that they mightretain tumorigenic potential and be involved in cancerformation via LHCGR-induced signals (). Thestudy of mouse models revealed that tumorigenesis atgonadal and extragonadal sites is related to excessivegonadotropin levels (). Similar conclusions havebeen suggested by the study of CGB gene mutations,which lead to aberrant hCG production and havebeen linked to gestational trophoblastic neoplasia(). An interesting case report provided evidence ofhCG-dependent tumorigenesis. A woman undergoingchemotherapy for metastatic renal cell carcinomabecame pregnant during the treatment break period
574 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

and experienced dramatic growth of the tumor ().This event was consistent with hCG production andLHCGR expression in cancer cells. In addition, clinicalabortion coincided with rapid tumor regression,suggesting that the angiogenic potential of high hCGwas proportional to the hormonal levels. A relation-ship between cancer cell growth and the production ofhCGb and hCG-H molecules could be suspected(), because they are the major isoforms producedduring the first weeks of pregnancy. Their tumorigenicpotential might rely on the ERK/ and AKT path-ways, which is surprising because these pathways arepreferentially activated by LH rather than hCG.Overall, hyperglycosylated hCG molecules were sus-pected to induce proliferative signals required to en-hance maternal immune cell modulation, embryoimplantation, and trophoblast invasion at the earlypregnancy stages (). In vitro findings confirmed theproliferative potential of hCGb and hCG-H ().These data are consistent with the hypothesis thathCGb and hCG-H exert intracellular signaling dif-ferent from “classical,” dimeric hCG () and thattheir action is focused on activation of cAMP/PKA-mediated steroidogenic signals (, ), fundamentalfor massive progesterone production during preg-nancy. This molecular mechanism could be the basisof a protective effect against breast cancer (), be-cause relatively high intracellular cAMP levels couldtrigger the activation of apoptotic processes in certaincell types (, ), including breast cancer cells ()and testicular germ cell tumors (). Moreover, highLHCGR expression levels might result in increasedsensitivity to hCG-induced proapoptotic signals,suggesting that receptor levels are a prognostic value,at least for some ovarian cancers (). In contrast,hCG steroidogenic potential might be transposed invivo by increased production of estrogen metabolitesand growth factors displaying proangiogenic andproliferative activity () and, thus, indirectly in-ducing variable effects, depending on the enzymaticmilieu of the target cells.
“Overload” of steroidogenic, LHCGR-inducedsignals could also be provided by excessive LH ac-tivity. The data from mouse models overexpressinghuman LH has proved that gonadotropin tumorigenicpotential is a result of hyperstimulated ovaries, whichare induced to produce high estrogen levels ().Similar results were observed in hCG-overexpressingmice (), demonstrating that, in rodents, high levelsof both human gonadotropins produce comparablesteroid-dependent effects in vivo, as suggested usingmouse Leydig cells in vitro (). Gonadectomy ofcertain mouse strains has been linked to Lhr- andestrogen-independent adrenal tumors, which arephenotypically different from those developed in miceoverexpressing LH, hCG, or Fshr (, ), sug-gesting that the adrenal gland might be a target ofgonadotropin action (). These data are suggestive
of molecular mechanisms putatively linking adrenaltumors and the high LH levels encountered duringhuman menopause. A gonadotropin-responsive ad-renocortical adenoma was reported in a menopausalwoman, with relatively high testosterone levels inresponse to stimulation of cancer cells by endogenousLH (). Similar effects might arise in LHCGR-overexpressing adrenal tumors, which were found insome cases involving menopausal and pregnantwomen (). Large amounts of receptors wouldtrigger aberrant activation of the Wnt pathway bybinding endogenous gonadotropins, stimulating dif-ferentiation of adrenocortical cells. In addition, similartumorigenic effects might be due to activating LHCGRmutations in males, providing high basal cAMP andinositol phosphate levels (). In such cases, bothexcessive testosterone synthesis and proliferative sig-nals might be linked to Leydig cell tumors and toprostate cancer, and downregulation of the HPG axisby GnRH antagonists might be applied as clinicaltreatment to counteract aberrant cell growth ().
In conclusion, causality between gonadotropinsand cancer is not sufficiently supported by the sci-entific data. Tumorigenesis might instead be linked tothe action of steroids, which, in certain cases, aresynthesized by tumor cells and induce proliferativeevents. In contrast, cancer cells can display aberranttranscription of gonadotropins and their receptorgenes, as well as other hormones and receptors (),such as steroids, GnRH, growth factors, and others. Inparticular, the presence of a specific receptor expres-sion per se might not be indicative of cell function,because the opposite effects could depend on quan-titative expression of GPCRs (). Given the tumor-specific nature of the expression of these factors, theresulting effects are unpredictable. The complexity oftumor cell metabolism provides a unique picture thatshould be evaluated on a case-by-case basis, with thespecific treatment choice entrusted to clinician decision.
Clinical Applications
Gonadotropins are clinically used in both sexes whenendogenous production is impaired, such as in the caseof HPG disruption. In hypogonadotropic (e.g., sec-ondary or central) hypogonadism, gonadotropinadministration represents the most physiologicaltherapeutic approach. The administration of steroids(i.e., estradiol and/or progesterone in women or tes-tosterone in men) is preferred in selected cases.However, when physiological HPG activity is to berestored and direct stimulation of the gonads isnecessary, gonadotropin administration becomesmandatory. This therapy consists of daily or weeklysubcutaneous or intramuscular injections of biologicalcompounds, which are more expensive than steroidsand cumbersome for the patient. Despite these
“The complexity of tumor cellmetabolism provides a uniquepicture that should beevaluated on a case-by-casebasis.”
575doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

challenges, this is the only therapeutic approach thatcan mimic the complex balance of gonadotropinstimulation of the gonad.
Although the most frequent gonadotropin appli-cation is in COS for ART, this approach remainslargely empirical and not sustained by strong scientificevidence. Moreover, the business related to thistreatment makes the current pharmacological schemesfor COS driven widely by industry, rather than byscience. No scientific interest has been evident inestablishing the best gonadotropin combination inclinical practice during the past few decades. However,HH, characterized by the lack of endogenous go-nadotropin secretion, represents the best in vivomodelto compare the different actions of gonadotropins toevaluate their kinetics and efficacy. In this section, wehave assessed the current LH/hCG biological com-pounds and their clinical applications.
Urinary and recombinant preparationsLH and hCG are both available as registered drugs(Table ). Their biological activity (i.e., calibration) isevaluated by in vivo bioassays, assessing the gonado-tropin effects in living rats or mice (). The Van Hellbioassay is the standard method in pharmacopeia toassess gonadotropin bioactivity (). This method,developed in the s, is based on the daily sub-cutaneous injection for days of a fixed gonadotropindose in - to -day-old immature male rats until thefinal measurement of seminal vesicle weight gain(). Both LH and hCG are calibrated using the VanHell method, comparing their bioactivity to an in-ternational standard (). This method has two mainlimitations. First, the animal model used is unable todiscriminate between LH and hCG. Second, thisbioassay evaluates gonadotropin steroidogenic activityand not the full spectrum of molecular actions ().Mouse Leydig cells were recently used in vitro to detect
biological differences between LH and hCG. In ad-dition to the qualitative differences previously dem-onstrated in human granulosa cells (, , ), LHand hCG resulted in quantitatively different earlyintracellular actions of the cAMP/PKA pathways andsteroidogenesis (). These results suggest that the VanHell method might not evaluate gonadotropin bio-activity correctly, and certainly not fully, because itonly evaluates testosterone-dependent endpoints (,). Thus, calibration of LH and hCG compoundsusing the Van Hell bioassay does not consider thedifferences in molecular action described.
Historically, the first gonadotropin isolationdates back to the third decade of the th century,when two compounds with FSH and LH activitieswere extracted from the urine of pregnant andpostmenopausal women and named prolan A and B,respectively (). Only in the s were the earlygonadotropin compounds produced, first from humanpituitaries and then from urine when the humanmenopausal gonadotropin (hMG) was purified frompostmenopausal women (). These first compoundsshowed a FSH/LH activity ratio of :, ensured by bothLH molecules and supplementation of hCG derivedfrom the urine of pregnant women. However, a largeamount of impurities was detected, consisting of LHsubunits, growth factors, glycoproteins, binding pro-teins, and immunoglobulins (). All these residuescaused high batch-to-batch variability and could in-fluence hMG biological action, reducing the efficacyand exposing patients to possible adverse events ().The urinary purification process was subsequentlyimproved, eliminating all residual extraneous activitiesbut leading to progressive LH loss. To maintain theFSH/LH ratio :, LH activity was replaced by urinaryhCG (). Thus, in highly purified hMG compounds,the impurity percentage decreased to %. hCGmolecules represent % of the remaining protein
Table 4. Available Drugs With LH and/or hCG Activity According to Current Regulatory Agencies
Preparation Provider Molecule Source Immunoreactivity
Gonasi HP® IBSA CGa Urinary hCG
Humegon® Organon Inc. hMG Urinary FSH + LH + hCG
Luveris® Merck KGaA r-LH Recombinant LH
Menogon® Ferring Pharmaceuticals hMG Urinary FSH + LH + hCG
Menopur® Ferring Pharmaceuticals hMG Urinary FSH + LH + hCG
Ovitrelle® Merck KGaA CGa Recombinant hCG
Pergogreen® Serono hMG Urinary FSH + LH + hCG
Pergonal® Serono hMG Urinary FSH + LH + hCG
Pregnyl® MSD-Organon CGa Urinary hCG
576 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

content (). Hence, hMG compounds with thegreatest purity were defined as those containing thelowest amount of LH and the greatest hCG concen-tration (). However, hCGmolecules added to hMGare calibrated in vivo in rodents against an LH stan-dard. Thus, the amount of hCG in the final compoundis the number of molecules capable of producing abiological effect equivalent to that of the LH standard.This process depends largely on the half-life and doesnot consider the molecular differences between LHand hCG. Therefore, the number of hCG molecules inthe final preparation could be significantly lower thanthe number of LH molecules needed to obtain thesame biological action, with a disequilibrium, in molarterms, between receptor and ligand. This concept mustnot be overlooked, because the different number ofLH/hCG molecules competing with the same numberof receptors could be relevant in terms of LHCGRoccupancy and activation ().
Overall, although FSH is easily obtained by urinepurification, LH is lost during the chromatographysteps, reducing the efficiency of obtaining urine-derived LH preparations. In contrast, urine hCG (u-hCG) preparations were readily developed because ofthe abundance of hCG in the urine of pregnantwomen (). Thus, urine-derived biological com-pounds containing LH alone were not available untilthe advent of DNA technologies. Only urine hCG(u-hCG) compounds have been used to obtain LHactivity in clinical practice. Recombinant DNA tech-nologies led to the production of gonadotropins inCHO cell lines, with high rates of safety and consis-tency. Currently, six recombinant gonadotropins areavailable: follitropin a, follitropin b, follitropin g,lutropin a (r-LH), chorionic gonadotropin a (r-hCG),and corifollitropin a (). r-LH and r-hCG possessgreater purity than the urine-derived counterpartsand consist of a mixture of isoforms exhibiting ahigh degree of glycosylation heterogeneity, structur-ally and biologically comparable to that of endoge-nous gonadotropins, with slight differences due toposttranslational modifications ().
In addition to classical gonadotropins, recombi-nant DNA technology has allowed for the creation ofnew chimeric compounds, such as long-acting FSH(corifollitropin a) (). In this case, the FSH bsubunit was coupled to the CTP of the hCG b subunit(, ). Corifollitropin shows in vitro and in vivopharmacological activity comparable to that of folli-tropin g but with a longer half-life (~ hours) ().This in an interesting example of how DNA tech-nologies could mix several gonadotropin features inthe same compound.
Different clinical effects: ovarian stimulation,luteal support, and other aspectsThe gonadotropin administration in COS during ARTis the current in vivo model in which the different
clinical effects of biological compounds containing LHand hCG can be evaluated. In this setting, although atruly standardized approach does not exist, FSH ad-ministration is always provided to obtain multi-follicular growth (). In addition to this, LH or hCGadministration can be added to support FSH actions,mimicking the physiological concerted action of LHand FSH. LH physiologically regulates folliculargrowth, stimulating theca cell production of andro-gens, which, in turn, serve as a substrate for estradiolproduction in granulosa cells (). In the late fol-licular phase, the estradiol increase acts through apositive feedback mechanism causing the LH surgeneeded for ovulation (). Finally, LH stimulatesprogesterone production from the corpus luteum,which is then maintained by hCG if pregnancy occurs.To mimic this complex gonadotropin-mediated pro-cess during COS, regulatory agencies have allowed forthe addition of LH, hCG, or hMG to FSH, becausethey are still considered equivalent in clinical terms.No specific guidelines are available to select whichpatient might benefit from, and should be treatedwith, a specific combination. The selection remainslargely empirical, if not arbitrary.
In contrast, hCG is generally used at the end of theCOS phase to trigger final oocyte maturation. Phys-iologically, however, it is LH that is responsible forfinal follicular maturation and follicle rupture withoocyte expulsion from the follicle (). Despite thisdifference between COS protocols and physiology,mainly due to the historical availability of hCG but notof LH, the final ART outcome did not change whenu-hCG or r-hCG was compared with LH (). Thus,no specific evidence supports the use of LH instead ofhCG for triggering ovulation. In , the use of aGnRH agonist to induce an endogenous LH surgesufficient to trigger ovulation was proposed (). Thisalternative to hCG was not exploited until the use ofGnRH antagonists for pituitary downregulation inCOS, with a shorter and reversible action comparedwith the agonists (, ). Thus, with GnRH an-tagonists, the pituitary gland remains responsive and asingle GnRH agonist bolus is sufficient to displace theantagonist, activate the receptor, and induce endog-enous LH release, mimicking the physiological mid-cycle gonadotropin surge (). This surge consistsof a short ascending limb of ~ hours and a longdescending limb of ~ hours (). This pattern isslightly different from the physiological midcyclesurge, which lasts hours and shows three consec-utive phases (). Although GnRH agonist triggeringcould be more physiological and could reduce theadverse event risk (, ), neither large retro-spective studies () nor randomized clinical trials(RCTs) (, ) found differences between GnRHagonists and hCG for triggering ovulation. Thus,specific evidence favoring one or another ovulationtrigger scheme is not available for the final ART
577doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

outcome. In particular, specific hCG actions are ob-tained with this treatment, such as sustained pro-gesterone production, angiogenesis promotion, bloodflow and nutrition to the fetus (), umbilical corddevelopment, and uterine growth synchronization(). Thus, hCG substitution by LH or a GnRHagonist for triggering results in the loss of all theseessential hCG activities after fertilization. Thus, when aGnRH agonist trigger is used, appropriate lutealsupport with progesterone and estradiol should beconsidered (). Although this issue has not beenfully clarified and needs further investigation, it hasbeen established that luteal support with progesteroneis used in all cycles. Some investigators proposedmodified luteal support when a GnRH agonist triggeris chosen, using hCG either in one -IU singlebolus () or in repeated - to -IU boluses(), or with the addition of LH (). However, arecent meta-analysis showed that GnRH agonisttrigger use, together with luteal support by LH activity,leads to similar ART outcomes compared with clinicalprotocols using the hCG trigger (). However, theanalysis showed notably high heterogeneity in theresults, limiting its clinical significance.
COS represents the best in vivo example in whichgonadotropin combinations are used in pharmaco-logical doses, although the various protocols have notbeen based on physiology or sound scientific evidence.LH and hCG have been used interchangeably in this“unphysiological” context, and it is surprising how wellART has worked despite the sometimes disparate and“creative” stimulation protocols. It is difficult to drawphysiological conclusions about LH and hCG fromthe supraphysiological setting of COS. HH shouldrepresent a better model to understand the differenteffects in vivo of LH and hCG, if they exist.
Clinical experience with LH and hCG: the presentand the futureLH activity is needed in clinical practice in themanagement of both female and male hypogonadismwhen estrogen and androgen production should bestimulated through the direct action on theca andLeydig cells, respectively. Although both LH and hCGare currently commercially available, historically, onlyhCG has been used, because it was the only readilyavailable LHCGR ligand. In theory, both LH and hCGcould be used now despite the complete lack of sci-entific evidence in favor of one vs the other. This is truein many countries for female hypogonadism, but notfor male hypogonadism, because only hCG admin-istration is permitted by regulatory agencies for thelatter. It has been widely demonstrated that hCG isefficient at restoring eugonadism in HH men. In thecase of male hypogonadal hypogonadism, the standardtherapeutic approach has been based on hCG ad-ministration since the s (). In this setting,convincing evidence about the efficacy of hCG
administration is available, with different dosages andschemes, ranging from IU every other day ()to $ IU two times weekly (). However, thereason hCG administration has been preferred to LHin clinical practice resides in the historic availability ofcommercial preparations, rather than systematic,evidence-based demonstrations. No clinical trials haveyet compared LH to hCG in male HH. Only a singlecase has been reported, comparing a daily low hCGdose ( IU) to r-LH ( IU daily) in a man with HHafter surgery for a pituitary adenoma (). In thatpatient, either hCG or r-LH restored eugonadismwithout exhibiting a substantial difference. A carefulcomparison of the in vivo action of LH and hCG inHH men is needed. In healthy men with pituitarysuppression from a GnRH antagonist, it was dem-onstrated that a daily LH dose of . IU restoredtestosterone levels to the normal range (). That trialfound no differences between bolus and pulsatileadministration, suggesting that the physiologicalpulsatile LH secretion pattern is not strictly needed toobtain Leydig cell stimulation (). These two ex-amples indicate that low doses of LH could be suf-ficient to stimulate testicular androgen production toan extent similar to that obtained by apparently muchhigher doses of hCG ( IU weekly for hCG vs .IU weekly for LH), revealing that the dosage of LHnecessary to stimulate physiological testosteroneproduction might be much lower than expected. If so,this would clearly indicate a differential action of LHand hCG in vivo in human males. Clinical studies withLH in HH men are urgently needed to explore thisfinding.
In women, the main LH clinical application re-mains COS during ART. Several trials have evaluateddifferent ART outcomes using LH instead of hCG tosupport follicular growth (Fig. ) (). Despite theirlarge numbers, these studies showed high heteroge-neity and low quality, hindering the development ofstandard protocols based on scientific evidence. Thus,the gonadotropin stimulation during COS, claimed tobe “personalized” by ART physicians, remains a pe-culiar example of personalized medicine, in thatpersonalization is not based on the selection of theappropriate stimulus according to objective criteriabut, rather, on the “personal” beliefs of the prescriber.This challenge is further complicated by the increasingnumber of young infertile couples seeking ART andthe corresponding cost of gonadotropins and pro-cedures, which leaves the pharmacological approachwidely driven by the industry.
In this confusing setting, several clinicians haveproposed the addition of LH activity to FSH duringCOS. This relatively new combination has been widelydebated in the reported studies, in particular regardingthose with a poor response and women of advancedage (–). Available evidence does not clearlysupport the hypothesis of increased pregnancy rates
578 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

using LH combined with FSH in unselected women(), and the number of reported meta-analyses al-most outweigh the number of RCTs available on thistopic, suggesting the difficulty in designing an ob-jective meta-analysis to identify the best COS ap-proach, distinguishing the actual benefit of onetreatment compared with another (). Apart fromthe paucity of adequately powered studies and the ex-tremely high heterogeneity of the treatments used, thistopic has been weakened by the heterogeneity of theendpoints evaluated in RCTs (). The vast majority ofRCTs and meta-analyses have focused on the ARTclinical outcome. However, when the effectiveness of theaddition of LH and/or hCG is the topic of investigation,the first endpoint to be evaluated must be the ovarianresponse (i.e., the first measurable parameter of go-nadotropin action). Pregnancy and live birth rates, incontrast, are the final results of ART and influenced by anincreasing number of factors (e.g., sperm contribution,endometrial receptivity and implantation, placenta). Arecent meta-analysis combined the reported results,considering all gonadotropin combinations and all ARToutcomes, demonstrating that FSH alone resulted ingreater oocyte numbers compared with hMG or FSHplus LH (). The addition of LH activity is useful forreducing the amount of FSH needed and to improveoocyte quality but only when LH is used rather thanhCG. This could be a further suggestion that by reducingthe FSH doses used, the FSH activity in granulosa cells isshifted from the proapoptotic to the proliferativepathways, such as those activated by LH (, , ).Only five studies have directly compared LH to hCG;however, none clearly reported the hCG dosage com-bined with FSH, preventing any estimation of the realexposure of the ovary to LH/hCG in terms of the molarrelationship between LH or hCG and the LHCGR.
With the limitation of the pharmacological, ratherthan the physiological, setting, these results suggestthat it is difficult to establish the correct amount of LHto be used in clinical practice, which might be differentdepending on the clinical setting (HH or COS) andthe patient’s sex. In addition, considering the diffi-culty of measuring in vivo LH bioactivity using ap-proved bioassays, the debate remains fully open.The correct appraisal of the clinical application ofLH should probably combine in vitro and in vivodemonstrations.
Complications of LH and hCG therapyIn the evaluation of possible clinical applications of LHand hCG, adverse events must also be considered. Themost relevant iatrogenic COS adverse event remainsOHSS, which is the final result of an exaggerated ovarianresponse (). It rarely occurs when ART is applied tounselected women (incidence of % to %); however, itsincidence increases when high-risk women are treat-ed (). This risk is associated with a low body massindex, young age, high estradiol serum levels, high IL
concentrations, high vascular EGF, and renin–angiotensinsystem activation, although specific predictive pa-rameters are not yet available (). Moreover, eitherthe elevated estradiol serum levels obtained at the dayof ovulation or the large number of follicles developedduring COS could predispose to the development ofOHSS (). These OHSS-predisposing conditionscould be related to gonadotropin action and should becarefully evaluated to predict the occurrence of adverseART events.
hCG, rather than LH, is generally used to triggerovulation. In addition to differences at the molecularlevel, LH and hCG have different circulating half-lives(). The sustained luteotropic activity induced by hCGcould lead to side effects through the release of vasoactivesubstances (e.g., vascular EGF) and prostaglandins actingdirectly on the ovarian follicles. These direct and indirecteffects could result in OHSS (). Triggering ovulationwith a GnRH agonist instead of a classical hCG bolusseemed to reduce the OHSS risk (, ). A recentmeta-analysis reported a reduced OHSS rate with aGnRH agonist compared with hCG-triggered cycles,although no statistical significance was reached ().These findings have confirmed the lack of clearknowledge of OHSS pathophysiology. To enhance theknowledge of underlying pathophysiological mecha-nisms and to reduce the incidence of this adverse event,the two different clinical OHSS presentations should beconsidered separately (). First, early OHSS occurswithin days after hCG administration as a final,prolonged hCG effect on already stimulated ovaries. ThisOHSS clinical picture could probably benefit from re-placement of the hCG trigger by a GnRH agonist. LateOHSS occurs. days after triggering, representing theovarian response to the endogenous hCG increase afterfertilization (). This late complication does not benefitfrom hCG substitution, and further research is needed tobetter understand this condition and its prevention.
0.6
0.0
-0.6
Favo
rs F
SH
alon
eFa
vors
gon
adot
ropi
nco
mbi
natio
ns
Oocytes
MIIoocytes
Implantationrate
Embryos Pregnancy rate
© 2018 Endocrine Reviews ENDOCRINE SOCIETY
FSH versus FSH+LH
FSH versus FSH+hCG
Figure 5. Overall modelillustrating effects of LH vshCG supplementation toFSH on ART outcomes.Scatter plots were obtainedby meta-analysis (11) andindicate mean differences foreach outcome evaluated.Data were interpolated usingpolynomial function; 95%confidence intervals notshown. Adapted, underCreative Commons CC BYlicense, from Santi et al.(11).
579doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

In addition to OHSS, other clinical COS compli-cations should be considered. Among these, the mostfrequent adverse effect is cycle cancellation, whichoccurs in .% to .% of women of advanced age(). This event leads to important socioeconomicconsequences owing to the need to repeat COS,further exposing women to the risk of OHSS and anincreasing economic burden. The main reason forcycle cancellation remains an inadequate response togonadotropin stimulation. This probably results fromboth the lack of standardization in COS protocols andthe high heterogeneity of the clinical responses. Theonly chance to reduce the cycle cancellation rate is todevelop truly, evidence-based personalization of COSschemes, which will require rigorous clinical studies.
In HH men, the only adverse event that must beconsidered during hCG/LH treatment is excessive serumtestosterone levels, with possible negative effects on redblood cells, liver, and prostate gland. However, this effectis the result of androgen action on sensitive tissues ratherthan a direct consequence of hCG administration. It willbe interesting to explore whether HH treatment by LHrather than hCG will reduce the rate of adverse events.Similar conclusions can be applied to the pharmaco-logical treatment of cryptorchidism. Cryptorchidism isidentified by the absence of one or both undescendedtestes to the scrotum and is linked to germ cell loss,infertility, and increased testicular cancer risk (). Thedescent of the testes can be induced by hCG admin-istration or surgery. Treatment of cryptorchidism withhCG was associated with germ cell apoptosis and im-paired reproductive function in the adult (). Thiseffect is consistent with the cAMP/PKA-dependentproapoptotic potential of hCG and might plausibly bedampened by administering LH instead of CG. How-ever, the use of LH as a treatment of cryptorchidism hasnever been reported.
Beyond Reproduction: Effects of hCG onThyroid and Adrenal Glands
Thyroid hormones and TSH are involved in metabolismand development regulation, and increasing evidence hashighlighted a role for these molecules even in re-production (), revealing cross-talk between the go-nadal and thyroid axes in vertebrates (). Highstructural and biochemical similarities between gonad-otropins and TSH (), and their receptors, underliesuch interactions. The binding affinity of LHCGR andTSH receptor (TSHR) for their respective ligands relieson amino acidic residue at positions - of theextracellular domains. These were considered to lead tochimeric receptors for hormone cross-interaction studiesin the past (). hCG–TSHR chimeras revealed -foldcAMP increases in transfected HEK cells, and theeffect was mimicked by high concentrations (. 3
to ng/mL) () of hCG acting on wild-type TSHR in
vitro, demonstrating cross-activation of the canonicalcAMP/PKA pathway () and iodide uptake and tri-iodothyronine secretion in cultured human thyrocytes(). An in vitro comparison of LH and hCG using aCHO cell line permanently expressing TSHR cDNA(CHO-JP cells) revealed that LH is more potent thanhCG in inducing an intracellular cAMP increase, and aparallel line analysis demonstrated the overall equiva-lency of .mMhCG equal to .mMLH equal to .32 mM TSH (). A mutant hCG lacking the CTPfragment displayed similar potency to LH, suggestingthat the C-terminal peptide might act as a protectivefactor to prevent hyperthyroidism due to hCG cross-reactivity during pregnancy, when circulating concen-trations of this hormone are extremely high. However,both LH and hCG are able to displace TSH bound to itsreceptor, thereby revealing an unexpected, ~-foldgreater, thyrotropic activity in LH compared with hCG,equivalent to . and .mIU of TSH activity per . IUof LH and hCG, respectively (). In CHO-JP cells,enzymatic digestion of hCG resulted in deglycosylatedand/or desialylated molecules with greater potency inactivating cAMP compared with classical hCG (),suggestive of plausible evolutionary conservation of theTSH dependence of thyroid function during the earlyweeks of pregnancy. Similar results were obtained withhCG preparations from patients with trophoblasticdisease. These studies support the concept of thyrotropicaction of gonadotropins, especially hCG, which reachesrelatively high serum levels during pregnancy or inpatients affected by hCG-secreting tumors. It is wellknown that the increase in hCG levels during the th toth weeks of pregnancy matches the decrease in TSHlevels and sustains continuous thyroid hormone pro-duction (). This likely depicts the dual function ofhCG, consisting of negative feedback exerted throughthyroid hormones at the pituitary and supporting thy-roid function. These effects are emphasized during twinpregnancies, which features prolonged, higher hCGlevels compared with single pregnancies and more fre-quently increased free T and suppressed TSH levels(). In contrast, insufficient thyroid stimulation couldresult in low thyroid hormone levels and could result inspontaneous abortion (). Taken together, the meta-bolic regulation by thyroid hormones might be, in part,dependent on hCG during pregnancy and might beessential to support fetal development.
Because TSHR and thyroid hormone receptors areexpressed in endometrial cells, their ligands might beinvolved in the physiological regulation of this tissue(), although clear-cut evidence remains to befound. Given the relatively high potency of LH ininducing TSHR-mediated cAMP production in vitro(, ), a role for gonadotropins exerted at the levelof the female reproductive system through the TSHRcannot be excluded. LH and TSH release by the pi-tuitary gland seems to be synchronized, at least duringthe menstrual cycle, and is largely measurable as a
580 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

serum concentration peak of both hormones ().Also, it is known that hypothyroidism before pubertyleads to delayed sexual maturity, implying thyroidhormone action on gonadal function (). However,the promiscuity of endocrine signals was demon-strated even in simple vertebrates, such as fish (),suggesting that it is a conserved feature regulatingcertain physiological functions. It is not surprising thathypo- and hyperthyroidism are associated withmenstrual disturbances, anovulation, and subfertilityin females (). The cross-talk between thyroid andgonadal functions is far from being thoroughly elu-cidated. The relatively recent discovery of thyro-stimulin (), a gonadotropin-like molecule acting onthe TSHR and considered the most ancestral glyco-protein hormone, has provided information onovarian functioning. The production of thyrostimulinby oocytes has been described, suggesting that thismolecule, rather than TSH, might act as a paracrinefactor in the ovary by binding TSHR expressed ingranulosa cells (), where it would induce pro-liferative signals, simultaneously activating cAMP/PKA, ERK/, and AKT pathways (). Althoughsuggestive, these findings should be confirmed byclinical evidence, which is required to understand thewhole picture of the thyroid–gonadal interaction.
Interactions between LH/hCG and the adrenalgland were described in healthy women during themenopausal transition, as an effect linked to increasingserum LH concentrations and, during the first tri-mester of pregnancy, to hCG production. The adrenalgland intriguingly shares a common developmentalorigin with gonadal cells (). It is characterized bydifferent types of cells secreting mainly glucocorti-coids, as well as androgens and a small amount ofestrogens. During the menopausal transition, the lackof progesterone-negative feedback to the pituitary,occurring with the arrest of follicular maturation,results in increasing levels of LH molecules act-ing on LHCGR expressed in adrenal cortical cells(). The adrenal gland response consists of thesynthesis of pregnenolone metabolites obtained viathe D-steroidogenic pathway (), mainly dehy-droepiandrosterone and androstenediol (), as well ascortisol (). Most importantly, chronically elevated LHlevels are suspected to be linked to an increased risk ofthe development of adrenal tumors (), althoughfurther studies are required to elucidate the effect of LHon adrenal function. Long-term elevated LH levels in-duce Lhr expression in mouse adrenal glands, a first steprequired for initiating cortical tumor development (),and it is possible that such an effect also occurs inhumans (). hCG action in the adrenal gland wasdiscussed in the section “Pathophysiology of LH andhCG.” In certain cases, increased hCG has been sus-pected to be linked to adrenal LHCGR expressionduring pregnancy, resulting in high cortisol levelsand transitory Cushing syndrome (). However,
hCG-stimulated synthesis of adrenal steroids is marginalin healthy women () and transient Cushing syn-drome is a relatively rare event.
Conclusions
The evolution of different glycoprotein hormones andtheir receptors accompanied the separation of distinctendocrine axes to optimize endocrine and metabolicfunctions, which developed independently in verte-brates. Interspecies ligand–hormone binding is pos-sible, depending on the phylogenetic distance of thespecies and suggesting evolutionary conservation ofkey amino acid residues fundamental for the preser-vation of biochemical properties of the proteins. In-traspecies interactions between endocrine axes are alsopossible, owing to the common developmental originof the hormone target cells or as a method of opti-mizing certain metabolic functions. However, thecomplexity of the physiological mechanisms mediat-ing placentation of certain mammals, such as equidsand primates, required a more profound regulatorylevel, making the role of gonadotropins of pituitaryand chorionic origin distinguishable. It is plausible thatthis complexity achieved its maximal expression inhumans, with the largest number of genes encodingfor LHb/CGb molecules likely evolving from a singleancestral gene. Although these hormones might existin more than one isoform and glycosylation variant,each displaying specific biochemical properties, theirphysiological roles might be separated according tospecific functions required for gamete maturation andpregnancy support (Table ).
LH production is maintained in both males andfemales during the fertile age, when their functiondepends on the specific enzymatic milieu of targetcells. In women, the known androgenic role of LHis fulfilled by an overall flattened production ofandrostenedione by theca cells, as the major steroidserving as a substrate to be converted to the pro-liferative factor, estradiol, by granulosa cells under FSHstimulation. However, both progestational and pro-liferative LH-dependent effects have been shown to berelevant, specifically in granulosa cells, by in vitroexperiments, revealing previously unknown regulatorymolecular mechanisms, suggesting that the afore-mentioned metabolic functions are fundamental forproper oocyte maturation. These roles are exertedthrough a fine-tuned modulation of the balance be-tween the steroidogenic cAMP/PKA pathway on theone hand and proliferative/antiapoptotic signals me-diated through ERK/ and AKT pathway activationon the other, thus exhibiting the capability of biasedsignaling mediated under synergistic modulationexerted by FSH. Hence, female folliculogenesis isassisted by two LH and one FSH target cells andcharacterized by both LHCGR and FSHR expression
“Clinical comparisons of LHand hCG in those with HH areneeded to clarify manyoutstanding questions.”
581doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022
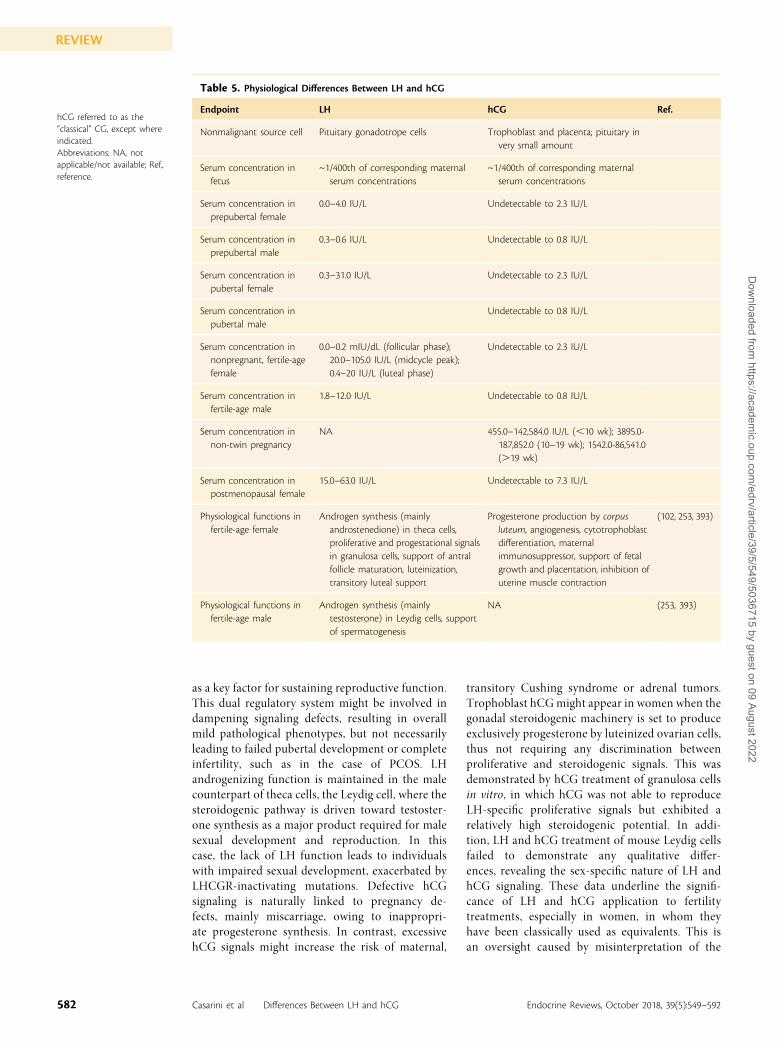
as a key factor for sustaining reproductive function.This dual regulatory system might be involved indampening signaling defects, resulting in overallmild pathological phenotypes, but not necessarilyleading to failed pubertal development or completeinfertility, such as in the case of PCOS. LHandrogenizing function is maintained in the malecounterpart of theca cells, the Leydig cell, where thesteroidogenic pathway is driven toward testoster-one synthesis as a major product required for malesexual development and reproduction. In thiscase, the lack of LH function leads to individualswith impaired sexual development, exacerbated byLHCGR-inactivating mutations. Defective hCGsignaling is naturally linked to pregnancy de-fects, mainly miscarriage, owing to inappropri-ate progesterone synthesis. In contrast, excessivehCG signals might increase the risk of maternal,
transitory Cushing syndrome or adrenal tumors.Trophoblast hCG might appear in women when thegonadal steroidogenic machinery is set to produceexclusively progesterone by luteinized ovarian cells,thus not requiring any discrimination betweenproliferative and steroidogenic signals. This wasdemonstrated by hCG treatment of granulosa cellsin vitro, in which hCG was not able to reproduceLH-specific proliferative signals but exhibited arelatively high steroidogenic potential. In addi-tion, LH and hCG treatment of mouse Leydig cellsfailed to demonstrate any qualitative differ-ences, revealing the sex-specific nature of LH andhCG signaling. These data underline the signifi-cance of LH and hCG application to fertilitytreatments, especially in women, in whom theyhave been classically used as equivalents. This isan oversight caused by misinterpretation of the
Table 5. Physiological Differences Between LH and hCG
Endpoint LH hCG Ref.
Nonmalignant source cell Pituitary gonadotrope cells Trophoblast and placenta; pituitary invery small amount
Serum concentration infetus
~1/400th of corresponding maternalserum concentrations
~1/400th of corresponding maternalserum concentrations
Serum concentration inprepubertal female
0.0–4.0 IU/L Undetectable to 2.3 IU/L
Serum concentration inprepubertal male
0.3–0.6 IU/L Undetectable to 0.8 IU/L
Serum concentration inpubertal female
0.3–31.0 IU/L Undetectable to 2.3 IU/L
Serum concentration inpubertal male
Undetectable to 0.8 IU/L
Serum concentration innonpregnant, fertile-agefemale
0.0–0.2 mIU/dL (follicular phase);20.0–105.0 IU/L (midcycle peak);0.4–20 IU/L (luteal phase)
Undetectable to 2.3 IU/L
Serum concentration infertile-age male
1.8–12.0 IU/L Undetectable to 0.8 IU/L
Serum concentration innon-twin pregnancy
NA 455.0–142,584.0 IU/L (,10 wk); 3895.0-187,852.0 (10–19 wk); 1542.0-86,541.0(.19 wk)
Serum concentration inpostmenopausal female
15.0–63.0 IU/L Undetectable to 7.3 IU/L
Physiological functions infertile-age female
Androgen synthesis (mainlyandrostenedione) in theca cells,proliferative and progestational signalsin granulosa cells, support of antralfollicle maturation, luteinization,transitory luteal support
Progesterone production by corpusluteum, angiogenesis, cytotrophoblastdifferentiation, maternalimmunosuppressor, support of fetalgrowth and placentation, inhibition ofuterine muscle contraction
(102, 253, 393)
Physiological functions infertile-age male
Androgen synthesis (mainlytestosterone) in Leydig cells, supportof spermatogenesis
NA (253, 393)
hCG referred to as the“classical” CG, except whereindicated.Abbreviations: NA, notapplicable/not available; Ref.,reference.
582 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

clinical data provided by studies performed in theART context, in which the LH- and hCG-specificeffects are masked by those that occur after themassive estrogen production induced to achieve
multifollicular development, which does not nor-mally occur in humans. Clinical comparisons of LHand hCG in those with HH is needed to clarifymany outstanding questions.
References1. Ascoli M, Fanelli F, Segaloff DL. The lutropin/
choriogonadotropin receptor, a 2002 perspective.Endocr Rev. 2002;23(2):141–174.
2. Stockell Hartree A, Shownkeen RC. Studies of hu-man pituitary LH containing internally cleaved betasubunit. J Mol Endocrinol. 1991;6(1):101–109.
3. Ward DN, Glenn SD, Nahm HS, Wen T. Charac-terization of cleavage products in selected humanlutropin preparations: a protease-sensitive site inhuman lutropin beta subunit. Int J Pept Protein Res.1986;27(1):70–78.
4. Lohse MJ, Benovic JL, Codina J, Caron MG, LefkowitzRJ. beta-Arrestin: a protein that regulates beta-adrenergic receptor function. Science. 1990;248(4962):1547–1550.
5. Galet C, Ascoli M. Arrestin-3 is essential for theactivation of Fyn by the luteinizing hormone re-ceptor (LHR) in MA-10 cells. Cell Signal. 2008;20(10):1822–1829.
6. Casarini L, Lispi M, Longobardi S, Milosa F, La MarcaA, Tagliasacchi D, Pignatti E, Simoni M. LH and hCGaction on the same receptor results in quantita-tively and qualitatively different intracellular sig-nalling. PLoS One. 2012;7(10):e46682.
7. Rulli SB, Kuorelahti A, Karaer O, Pelliniemi LJ,Poutanen M, Huhtaniemi I. Reproductive distur-bances, pituitary lactotrope adenomas, and mam-mary gland tumors in transgenic female miceproducing high levels of human chorionic gonad-otropin. Endocrinology. 2002;143(10):4084–4095.
8. Riccetti L, De Pascali F, Gilioli L, Potı F, Giva LB,Marino M, Tagliavini S, Trenti T, Fanelli F, MezzulloM, Pagotto U, Simoni M, Casarini L. Human LH andhCG stimulate differently the early signallingpathways but result in equal testosterone synthesisin mouse Leydig cells in vitro. Reprod Biol Endocrinol.2017;15(1):2.
9. Grzesik P, Kreuchwig A, Rutz C, Furkert J, Wiesner B,Schuelein R, Kleinau G, Gromoll J, Krause G. Dif-ferences in signal activation by LH and hCG aremediated by the LH/CG receptor’s extracellularhinge region. Front Endocrinol (Lausanne). 2015;6:140.
10. Robyn C, Diczfalusy E. Bioassay of antigonadotrophicsera. 2. Assay of the human chorionic gonadotro-phin (HCG) and luteinising hormone (LH) neutral-ising potencies. Acta Endocrinol (Copenh). 1968;59(2):261–276.
11. Santi D, Casarini L, Alviggi C, Simoni M. Efficacy offollicle-stimulating hormone (FSH) alone, FSH +luteinizing hormone, human menopausal gonado-tropin or FSH + human chorionic gonadotropin onassisted reproductive technology outcomes in the“personalized” medicine era: a meta-analysis. FrontEndocrinol (Lausanne). 2017;8:114.
12. Huhtaniemi IT, Catt KJ. Differential binding affinitiesof rat testis luteinizing hormone (LH) receptors forhuman chorionic gonadotropin, human LH, andovine LH. Endocrinology. 1981;108(5):1931–1938.
13. Schoneberg T, Hofreiter M, Schulz A, Rompler H.Learning from the past: evolution of GPCR func-tions. Trends Pharmacol Sci. 2007;28(3):117–121.
14. Policastro PF, Daniels-McQueen S, Carle G, Boime I.A map of the hCG beta-LH beta gene cluster. J BiolChem. 1986;261(13):5907–5916.
15. Cole LA. hCG and hyperglycosylated hCG in theestablishment and evolution of hemochorial pla-centation. J Reprod Immunol. 2009;82(2):112–118.
16. Dinopoulou V, Drakakis P, Kefala S, Kiapekou E,Bletsa R, Anagnostou E, Kallianidis K, Loutradis D.Effect of recombinant-LH and hCG in the absenceof FSH on in vitro maturation (IVM) fertilization andearly embryonic development of mouse germinalvesicle (GV)-stage oocytes. Reprod Biol. 2016;16(2):138–146.
17. Sun PD, Davies DR. The cystine-knot growth-factorsuperfamily. Annu Rev Biophys Biomol Struct. 1995;24(1):269–291.
18. Bousfield GR, Dias JA. Synthesis and secretion ofgonadotropins including structure-function corre-lates. Rev Endocr Metab Disord. 2011;12(4):289–302.
19. Roch GJ, Sherwood NM. Glycoprotein hormonesand their receptors emerged at the origin ofmetazoans. Genome Biol Evol. 2014;6(6):1466–1479.
20. Casarini L, Santi D, Marino M. Impact of genepolymorphisms of gonadotropins and their re-ceptors on human reproductive success. Re-production. 2015;150(6):R175–R184.
21. Moyle WR, Campbell RK, Myers RV, Bernard MP,Han Y, Wang X. Co-evolution of ligand-receptorpairs. Nature. 1994;368(6468):251–255.
22. Dewey EM, McNabb SL, Ewer J, Kuo GR, TakanishiCL, Truman JW, Honegger H-W. Identificationof the gene encoding bursicon, an insect neuro-peptide responsible for cuticle sclerotization andwing spreading. Curr Biol. 2004;14(13):1208–1213.
23. Graves J, Markman S, Alegranti Y, Gechtler J,Johnson RI, Cagan R, Ben-Menahem D. The LH/CGreceptor activates canonical signaling pathwaywhen expressed in Drosophila. Mol Cell Endocrinol.2015;413:145–156.
24. Uchida K, Moriyama S, Chiba H, Shimotani T,Honda K, Miki M, Takahashi A, Sower SA, Nozaki M.Evolutionary origin of a functional gonadotropin inthe pituitary of the most primitive vertebrate,hagfish. Proc Natl Acad Sci USA. 2010;107(36):15832–15837.
25. Huhtaniemi I. A short evolutionary history of FSH-stimulated spermatogenesis. Hormones (Athens).2015;14(4):468–478.
26. Sower SA, Decatur WA, Hausken KN, Marquis TJ,Barton SL, Gargan J, Freamat M, Wilmot M,Hollander L, Hall JA, Nozaki M, Shpilman M, Levavi-Sivan B. Emergence of an ancestral glycoproteinhormone in the pituitary of the sea lamprey, a basalvertebrate. Endocrinology. 2015;156(8):3026–3037.
27. Li MD, Ford JJ. A comprehensive evolutionaryanalysis based on nucleotide and amino acid se-quences of the alpha- and beta-subunits of glyco-protein hormone gene family. J Endocrinol. 1998;156(3):529–542.
28. Buechi HB, Bridgham JT. Evolution of specificity incartilaginous fish glycoprotein hormones and re-ceptors. Gen Comp Endocrinol. 2017;246:309–320.
29. Bogerd J, Granneman JCM, Schulz RW, Vischer HF.Fish FSH receptors bind LH: how to make thehuman FSH receptor to be more fishy? Gen CompEndocrinol. 2005;142(1-2):34–43.
30. Garcıa-Lopez A, de Jonge H, Nobrega RH, de WaalPP, van Dijk W, Hemrika W, Taranger GL, Bogerd J,Schulz RW. Studies in zebrafish reveal unusualcellular expression patterns of gonadotropin re-ceptor messenger ribonucleic acids in the testis andunexpected functional differentiation of the go-nadotropins. Endocrinology. 2010;151(5):2349–2360.
31. Aizen J, Kowalsman N, Kobayashi M, Hollander L,Sohn YC, Yoshizaki G, Niv MY, Levavi-Sivan B. Ex-perimental and computational study of inter- andintra- species specificity of gonadotropins for vari-ous gonadotropin receptors. Mol Cell Endocrinol.2012;364(1-2):89–100.
32. Julier C, Weil D, Couillin P, Cote JC, Nguyen VC,Foubert C, Boue A, Thirion JP, Kaplan JC, Junien C.The beta chorionic gonadotropin-beta luteinizinggene cluster maps to human chromosome 19. HumGenet. 1984;67(2):174–177.
33. Bo M, Boime I. Identification of the transcriptionallyactive genes of the chorionic gonadotropin betagene cluster in vivo. J Biol Chem. 1992;267(5):3179–3184.
34. Hollenberg AN, Pestell RG, Albanese C, Boers ME,Jameson JL. Multiple promoter elements in thehuman chorionic gonadotropin beta subunit genesdistinguish their expression from the luteinizinghormone beta gene. Mol Cell Endocrinol. 1994;106(1-2):111–119.
35. Britten RJ. Divergence between samples of chim-panzee and human DNA sequences is 5%, countingindels. Proc Natl Acad Sci USA. 2002;99(21):13633–13635.
36. Nagirnaja L, Rull K, Uuskula L, Hallast P, GrigorovaM,Laan M. Genomics and genetics of gonadotropinbeta-subunit genes: Unique FSHB and duplicatedLHB/CGB loci. Mol Cell Endocrinol. 2010;329(1-2):4–16.
37. Hallast P, Saarela J, Palotie A, Laan M. High di-vergence in primate-specific duplicated regions:human and chimpanzee chorionic gonadotropinbeta genes. BMC Evol Biol. 2008;8(1):195.
38. Hallast P, Nagirnaja L, Margus T, Laan M. Segmentalduplications and gene conversion: human lutei-nizing hormone/chorionic gonadotropin beta genecluster. Genome Res. 2005;15(11):1535–1546.
39. Maston GA, Ruvolo M. Chorionic gonadotropinhas a recent origin within primates and an evolu-tionary history of selection.Mol Biol Evol. 2002;19(3):320–335.
40. Hallast P, Rull K, Laan M. The evolution and ge-nomic landscape of CGB1 and CGB2 genes.Mol CellEndocrinol. 2007;260-262:2–11.
41. Rull K, Laan M. Expression of b-subunit of HCGgenes during normal and failed pregnancy. HumReprod. 2005;20(12):3360–3368.
42. Pennell TM, Morrow EH. Two sexes, one genome:the evolutionary dynamics of intralocus sexualconflict. Ecol Evol. 2013;3(6):1819–1834.
583doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

43. Chen H, Chen L, Wu Y, Shen H, Yang G, Deng C.The exonization and functionalization of an Alu-Jelement in the protein coding region of glyco-protein hormone alpha gene represent a novelmechanism to the evolution of hemochorial pla-centation in primates. Mol Biol Evol. 2017;34(12):3216–3231.
44. Fiddes JC, Goodman HM. The cDNA for the beta-subunit of human chorionic gonadotropin sug-gests evolution of a gene by readthrough into the39-untranslated region. Nature. 1980;286(5774):684–687.
45. Bousfield GR, Butnev VY, Butnev VY. Identificationof twelve O-glycosylation sites in equine chorionicgonadotropin beta and equine luteinizing hormoness by solid-phase Edman degradation. Biol Reprod.2001;64(1):136–147.
46. Bidart JM, Bellet DH, Alberici GF, Van Besien F,Bohuon C. The immune response to a syntheticpeptide analogous to the 109-145 beta hCG carboxyl-terminus is directed against two major and two minorregions. Mol Immunol. 1987;24(4):339–345.
47. Mak SMJ, Wong WY, Chung HS, Chung PW, KongGWS, Li TC, Cheung LP. Effect of mid-follicularphase recombinant LH versus urinary HCG supple-mentation in poor ovarian responders undergoingIVF—a prospective double-blinded randomizedstudy. Reprod Biomed Online. 2017;34(3):258–266.
48. Zeleznik AJ. In vivo responses of the primate corpusluteum to luteinizing hormone and chorionic go-nadotropin. Proc Natl Acad Sci USA. 1998;95(18):11002–11007.
49. Wallace WHB, Kelsey TW. Human ovarian reservefrom conception to the menopause. PLoS One.2010;5(1):e8772.
50. Yung Y, Aviel-Ronen S, Maman E, Rubinstein N,Avivi C, Orvieto R, Hourvitz A. Localization ofluteinizing hormone receptor protein in the humanovary. Mol Hum Reprod. 2014;20(9):844–849.
51. Chappel SC, Howles C. Reevaluation of the roles ofluteinizing hormone and follicle-stimulating hor-mone in the ovulatory process. Hum Reprod. 1991;6(9):1206–1212.
52. Jeppesen JV, Kristensen SG, Nielsen ME, HumaidanP, Dal Canto M, Fadini R, Schmidt KT, Ernst E, YdingAndersen C. LH-receptor gene expression in humangranulosa and cumulus cells from antral and pre-ovulatory follicles. J Clin Endocrinol Metab. 2012;97(8):E1524–E1531.
53. Ji I, Lee C, Song Y, Conn PM, Ji TH. Cis- and trans-activation of hormone receptors: the LH receptor.Mol Endocrinol. 2002;16(6):1299–1308.
54. Jonas KC, Chen S, Virta M, Mora J, Franks S,Huhtaniemi I, Hanyaloglu AC. Temporal reprog-ramming of calcium signalling via crosstalk of go-nadotrophin receptors that associate as functionallyasymmetric heteromers. Sci Rep. 2018;8(1):2239.
55. Tal R, Taylor HS, Burney RO, Mooney SB, GiudiceLC. Endocrinology of pregnancy. In: De Groot LJ,Chrousos G, Dungan K, Feingold KR, Grossman A,Hershman JM, Koch C, Korbonits M, McLachlan R,New M, Purnell J, Rebar R, Singer F, Vinik A, eds.Endotext. South Dartmouth, MA: MDText.com, Inc.;2000.
56. Makieva S, Saunders PTK, Norman JE. Androgens inpregnancy: roles in parturition. Hum Reprod Update.2014;20(4):542–559.
57. Chen T, Surcel H-M, Lundin E, Kaasila M, Lakso H-A,Schock H, Kaaks R, Koskela P, Grankvist K, HallmansG, Pukkala E, Zeleniuch-Jacquotte A, Toniolo P,Lehtinen M, Lukanova A. Circulating sex steroidsduring pregnancy and maternal risk of non-epithelial ovarian cancer. Cancer Epidemiol Bio-markers Prev. 2011;20(2):324–336.
58. Cole LA. hCG physiology. Placenta. 2013;34(12):1257.
59. Tapia-Pizarro A, Archiles S, Argandoña F, Valencia C,Zavaleta K, Cecilia Johnson M, Gonzalez-Ramos R,Devoto L. hCG activates Epac-Erk1/2 signalingregulating progesterone receptor expression andfunction in human endometrial stromal cells. MolHum Reprod. 2017;23(6):393–405.
60. Pitteloud N, Dwyer A. Hormonal control of sper-matogenesis in men: therapeutic aspects inhypogonadotropic hypogonadism. Ann Endocrinol(Paris). 2014;75(2):98–100.
61. Rey RA. Mini-puberty and true puberty: differencesin testicular function. Ann Endocrinol (Paris). 2014;75(2):58–63.
62. Pentikainen V, Erkkila K, Suomalainen L, ParvinenM, Dunkel L. Estradiol acts as a germ cell survivalfactor in the human testis in vitro. J Clin EndocrinolMetab. 2000;85(5):2057–2067.
63. Pareek TK, Joshi AR, Sanyal A, Dighe RR. Insightsinto male germ cell apoptosis due to depletion ofgonadotropins caused by GnRH antagonists. Ap-optosis. 2007;12(6):1085–1100.
64. Nisula BC, Blithe DL, Akar A, Lefort G, Wehmann RE.Metabolic fate of human choriogonadotropin.J Steroid Biochem. 1989;33(4B)733–737.
65. Klett D, Bernard S, Lecompte F, Leroux H, MagallonT, Locatelli A, Lepape A, Combarnous Y. Fast renaltrapping of porcine luteinizing hormone (pLH)shown by 123I-scintigraphic imaging in rats explainsits short circulatory half-life. Reprod Biol Endocrinol.2003;1(1):64.
66. Fiete D, Srivastava V, Hindsgaul O, Baenziger JU. Ahepatic reticuloendothelial cell receptor specific forSO4-4GalNAc beta 1,4GlcNAc beta 1,2Man alphathat mediates rapid clearance of lutropin. Cell. 1991;67(6):1103–1110.
67. Birken S, Chen Y, Gawinowicz MA, Agosto GM,Canfield RE, Hartree AS. Structure and significanceof human luteinizing hormone-beta core fragmentpurified from human pituitary extracts. Endocri-nology. 1993;133(3):985–989.
68. Birken S, Gawinowicz MA, Maydelman Y, MilgromY. Metabolism of gonadotropins: comparisons ofthe primary structures of the human pituitary andurinary LH beta cores and the chimpanzee CG betacore demonstrate universality of core production.J Endocrinol. 2001;171(1):131–141.
69. Ben-Menahem D, Grotjan HE. Strategies for con-struction of luteinizing hormone beta subunitanalogs with carboxyl terminal extensions in non-primate, non-equid mammalian species. Mol CellEndocrinol. 2007;260-262:205–211.
70. Bousfield GR, Liu WK, Sugino H, Ward DN.Structural studies on equine glycoprotein hor-mones: amino acid sequence of equine lutropinbeta-subunit. J Biol Chem. 1987;262(18):8610–8620.
71. Gabay R, Rozen S, Samokovlisky A, Amor Y,Rosenfeld R, Kohen F, Amsterdam A, Berger P, Ben-Menahem D. The role of the 39 region of mam-malian gonadotropin b subunit gene in theluteinizing hormone to chorionic gonadotropinevolution.Mol Cell Endocrinol. 2014;382(2):781–790.
72. Allen WR, Moor RM. The origin of the equineendometrial cups. I. Production of PMSG by fetaltrophoblast cells. J Reprod Fertil. 1972;29(2):313–316.
73. Sherman GB, Wolfe MW, Farmerie TA, Clay CM,Threadgill DS, Sharp DC, Nilson JH. A single geneencodes the beta-subunits of equine luteinizinghormone and chorionic gonadotropin. Mol Endo-crinol. 1992;6(6):951–959.
74. Butnev VY, Gotschall RR, Baker VL, Moore WT,Bousfield GR. Negative influence of O-linked
oligosaccharides of high molecular weight equinechorionic gonadotropin on its luteinizing hormoneand follicle-stimulating hormone receptor-bindingactivities. Endocrinology. 1996;137(6):2530–2542.
75. Butney VY, Gotschall RR, Butnev VY, Baker VL, MooreWT, Bousfield GR. Hormone-specific inhibitory in-fluence of alpha-subunit Asn56 oligosaccharide onin vitro subunit association and follicle-stimulatinghormone receptor binding of equine gonadotropins.Biol Reprod. 1998;58(2):458–469.
76. Guillou F, Combarnous Y. Purification of equinegonadotropins and comparative study of their acid-dissociation and receptor-binding specificity. Bio-chim Biophys Acta. 1983;755(2):229–236.
77. Stewart F, Allen WR. The binding of FSH, LH andPMSG to equine gonadal tissues. J Reprod FertilSuppl. 1979;(27):431–440.
78. Stewart F, Allen WR. Biological functions and re-ceptor binding activities of equine chorionic go-nadotrophins. J Reprod Fertil. 1981;62(2):527–536.
79. Simula AP, Amato F, Faast R, Lopata A, Berka J,Norman RJ. Luteinizing hormone/chorionic go-nadotropin bioactivity in the common marmoset(Callithrix jacchus) is due to a chorionic gonado-tropin molecule with a structure intermediatebetween human chorionic gonadotropin and hu-man luteinizing hormone. Biol Reprod. 1995;53(2):380–389.
80. Gromoll J, Wistuba J, Terwort N, Godmann M,Muller T, Simoni M. A new subclass of the lutei-nizing hormone/chorionic gonadotropin receptorlacking exon 10 messenger RNA in the New Worldmonkey (Platyrrhini) lineage. Biol Reprod. 2003;69(1):75–80.
81. Troppmann B, Kleinau G, Krause G, Gromoll J.Structural and functional plasticity of the lutei-nizing hormone/choriogonadotrophin receptor.Hum Reprod Update. 2013;19(5):583–602.
82. Muller T, Gromoll J, Simula AP, Norman R, Sandhowe-Klaverkamp R, Simoni M. The carboxyterminal pep-tide of chorionic gonadotropin facilitates activation ofthe marmoset LH receptor. Exp Clin Endocrinol Di-abetes. 2004;112(10):574–579.
83. Gromoll J, Eiholzer U, Nieschlag E, Simoni M. Malehypogonadism caused by homozygous deletion ofexon 10 of the luteinizing hormone (LH) receptor:differential action of human chorionic gonado-tropin and LH. J Clin Endocrinol Metab. 2000;85(6):2281–2286.
84. Muller T, Gromoll J, Simoni M. Absence of exon 10of the human luteinizing hormone (LH) receptorimpairs LH, but not human chorionic gonadotropinaction. J Clin Endocrinol Metab. 2003;88(5):2242–2249.
85. Hoermann R, Spoettl G,Moncayo R,Mann K. Evidencefor the presence of human chorionic gonadotropin(hCG) and free beta-subunit of hCG in the humanpituitary. J Clin Endocrinol Metab. 1990;71(1):179–186.
86. Odell WD, Griffin J. Pulsatile secretion of humanchorionic gonadotropin in normal adults. N Engl JMed. 1987;317(27):1688–1691.
87. Odell WD, Griffin J. Pulsatile secretion of chorionicgonadotropin during the normal menstrual cycle.J Clin Endocrinol Metab. 1989;69(3):528–532.
88. Cole LA, Gutierrez JM. Production of humanchorionic gonadotropin during the normal men-strual cycle. J Reprod Med. 2009;54(4):245–250.
89. Merhi Z, Pollack SE. Pituitary origin of persistentlyelevated human chorionic gonadotropin in a pa-tient with gonadal failure. Fertil Steril. 2013;99(1):293–296.
90. Bielinska M, Rzymkiewicz D, Boime I. Humanluteinizing hormone and chorionic gonadotropinare targeted to a regulated secretory pathway inGH3 cells. Mol Endocrinol. 1994;8(7):919–928.
584 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

91. Kochman K, Kerdelhue B, Zor U, Jutisz M. Studies ofenzymatic degradation of luteinizing hormone-releasing hormone by different tissues. FEBS Lett.1975;50(2):190–194.
92. Skorupskaite K, George JT, Anderson RA. Thekisspeptin-GnRH pathway in human reproductivehealth and disease. Hum Reprod Update. 2014;20(4):485–500.
93. Gardner S, Maudsley S, Millar RP, Pawson AJ. Nu-clear stabilization of b-catenin and inactivation ofglycogen synthase kinase-3b by gonadotropin-releasing hormone: targeting Wnt signaling in thepituitary gonadotrope. Mol Endocrinol. 2007;21(12):3028–3038.
94. Thompson IR, Kaiser UB. GnRH pulse frequency-dependent differential regulation of LH and FSHgene expression. Mol Cell Endocrinol. 2014;385(1-2):28–35.
95. Jayes FC, Britt JH, Esbenshade KL. Role ofgonadotropin-releasing hormone pulse frequencyin differential regulation of gonadotropins in thegilt. Biol Reprod. 1997;56(4):1012–1019.
96. Lunenfeld B, Buhler K. The neuro control of theovarian cycle—a hypothesis. Gynecol Endocrinol.2018;34(4):278–282.
97. Abel MH, Widen A, Wang X, Huhtaniemi I,Pakarinen P, Kumar TR, Christian HC. Pituitarygonadotrophic hormone synthesis, secretion, sub-unit gene expression and cell structure in normaland follicle-stimulating hormone b knockout,follicle-stimulating hormone receptor knockout,luteinising hormone receptor knockout, hypogonadaland ovariectomised female mice. J Neuroendocrinol.2014;26(11):785–795.
98. Stanton PG, Pozvek G, Burgon PG, Robertson DM,Hearn MT. Isolation and characterization of humanLH isoforms. J Endocrinol. 1993;138(3):529–543.
99. Merz WE. The primate placenta and humanchorionic gonadotropin. Exp Clin Endocrinol. 1994;102(3):222–234.
100. Evans J, Salamonsen LA, Menkhorst E, Dimitriadis E.Dynamic changes in hyperglycosylated humanchorionic gonadotrophin throughout the first tri-mester of pregnancy and its role in early placen-tation. Hum Reprod. 2015;30(5):1029–1038.
101. Cole LA. hCG, five independent molecules. ClinChim Acta. 2012;413(1-2):48–65.
102. Cole LA. hCG, the wonder of today’s science. ReprodBiol Endocrinol. 2012;10(1):24.
103. Mann K, Lamerz R, Hellmann T, Kumper HJ, StaehlerG, Karl HJ. Use of human chorionic gonadotropinand alpha-fetoprotein radioimmunoassays: speci-ficity and apparent half-life determination afterdelivery and in patients with germ cell tumors.Oncodev Biol Med. 1980;1(4-5):301–312.
104. Furuhashi M, Shikone T, Fares FA, Sugahara T,Hsueh AJ, Boime I. Fusing the carboxy-terminalpeptide of the chorionic gonadotropin (CG)beta-subunit to the common alpha-subunit: re-tention of O-linked glycosylation and enhancedin vivo bioactivity of chimeric human CG. MolEndocrinol. 1995;9(1):54–63.
105. Ulloa-Aguirre A, Maldonado A, Damian-MatsumuraP, Timossi C. Endocrine regulation of gonadotropinglycosylation. Arch Med Res. 2001;32(6):520–532.
106. Manzella SM, Dharmesh SM, Beranek MC, SwansonP, Baenziger JU. Evolutionary conservation of thesulfated oligosaccharides on vertebrate glycopro-tein hormones that control circulatory half-life. J BiolChem. 1995;270(37):21665–21671.
107. Fiddes JC, Goodman HM. Isolation, cloning andsequence analysis of the cDNA for the alpha-subunit of human chorionic gonadotropin. Na-ture. 1979;281(5730):351–356.
108. Xing Y, Myers RV, Cao D, Lin W, Jiang M, BernardMP, Moyle WR. Glycoprotein hormone assembly inthe endoplasmic reticulum: I. The glycosylated endof human alpha-subunit loop 2 is threadedthrough a beta-subunit hole. J Biol Chem. 2004;279(34):35426–35436.
109. Moyle WR, Myers RV, Wang Y, Han Y, Lin W, KelleyGL, Ehrlich PH, Rao SN, Bernard MP. Functionalhomodimeric glycoprotein hormones: implicationsfor hormone action and evolution. Chem Biol. 1998;5(5):241–254.
110. Lustbader JW, Birken S, Pileggi NF, Kolks MA, PollakS, Cuff ME, Yang W, Hendrickson WA, Canfield RE.Crystallization and characterization of humanchorionic gonadotropin in chemically deglycosy-lated and enzymatically desialylated states. Bio-chemistry. 1989;28(24):9239–9243.
111. Lapthorn AJ, Harris DC, Littlejohn A, Lustbader JW,Canfield RE, Machin KJ, Morgan FJ, Isaacs NW.Crystal structure of human chorionic gonadotropin.Nature. 1994;369(6480):455–461.
112. Wu H, Lustbader JW, Liu Y, Canfield RE,Hendrickson WA. Structure of human chorionicgonadotropin at 2.6 A resolution from MADanalysis of the selenomethionyl protein. Structure.1994;2(6):545–558.
113. Furuhashi M, Suzuki S, Suganuma N. Disulfidebonds 7-31 and 59-87 of the alpha-subunit play adifferent role in assembly of human chorionicgonadotropin and lutropin. Endocrinology. 1996;137(10):4196–4200.
114. Feng W, Matzuk MM, Mountjoy K, Bedows E,Ruddon RW, Boime I. The asparagine-linked oli-gosaccharides of the human chorionic gonado-tropin beta subunit facilitate correct disulfide bondpairing. J Biol Chem. 1995;270(20):11851–11859.
115. Bernard MP, Lin W, Kholodovych V, Moyle WR.Human lutropin (hLH) and choriogonadotropin(CG) are assembled by different pathways: a modelof hLH assembly. J Biol Chem. 2014;289(20):14360–14369.
116. Darling RJ, Wilken JA, Miller-Lindholm AK, UrlacherTM, Ruddon RW, Sherman SA, Bedows E. Func-tional contributions of noncysteine residues withinthe cystine knots of human chorionic gonadotropinsubunits. J Biol Chem. 2001;276(14):10692–10699.
117. Ruddon RW, Sherman SA, Bedows E. Protein foldingin the endoplasmic reticulum: lessons from thehuman chorionic gonadotropin b subunit. ProteinSci. 1996;5(8):1443–1452.
118. Muyan M, Furuhashi M, Sugahara T, Boime I. Thecarboxy-terminal region of the beta-subunits ofluteinizing hormone and chorionic gonadotropindifferentially influence secretion and assembly ofthe heterodimers. Mol Endocrinol. 1996;10(12):1678–1687.
119. Suzuki S, Furuhashi M, Suganuma N. AdditionalN-glycosylation at Asn(13) rescues the human LHbeta-subunit from disulfide-linked aggregation. MolCell Endocrinol. 2000;160(1-2):157–163.
120. Corless CL, Matzuk MM, Ramabhadran TV,Krichevsky A, Boime I. Gonadotropin beta subunitsdetermine the rate of assembly and the oligosac-charide processing of hormone dimer in transfectedcells. J Cell Biol. 1987;104(5):1173–1181.
121. Lo A, Zheng W, Gong Y, Crochet JR, Halvorson LM.GATA transcription factors regulate LHb geneexpression. J Mol Endocrinol. 2011;47(1):45–58.
122. Zheng W, Horton CD, Kim J, Halvorson LM. Theorphan nuclear receptors COUP-TFI and COUP-TFII regulate expression of the gonadotropin LHbgene. Mol Cell Endocrinol. 2010;330(1-2):59–71.
123. Millar RP. GnRHs and GnRH receptors. AnimReprod Sci. 2005;88(1-2):5–28.
124. Caunt CJ, Finch AR, Sedgley KR, Oakley L, LuttrellLM, McArdle CA. Arrestin-mediated ERK activation bygonadotropin-releasing hormone receptors: receptor-specific activation mechanisms and compartmentali-zation. J Biol Chem. 2006;281(5):2701–2710.
125. Salisbury TB, Binder AK, Grammer JC, Nilson JH.Maximal activity of the luteinizing hormoneb-subunit gene requires b-catenin. Mol Endocrinol.2007;21(4):963–971.
126. Sairam MR, Li CH. Human pituitary lutropin: iso-lation, properties, and the complete amino acidsequence of the beta-subunit. Biochim Biophys Acta.1975;412(1):70–81.
127. Morgan FJ, Birken S, Canfield RE. The amino acidsequence of human chorionic gonadotropin: thealpha subunit and beta subunit. J Biol Chem. 1975;250(13):5247–5258.
128. Talmadge K, Boorstein WR, Fiddes JC. The humangenome contains seven genes for the beta-subunitof chorionic gonadotropin but only one gene forthe beta-subunit of luteinizing hormone. DNA.1983;2(4):281–289.
129. den Dunnen JT, Antonarakis SE. Mutation no-menclature extensions and suggestions to describecomplex mutations: a discussion. Hum Mutat. 2000;15(1):7–12.
130. Wide L, Eriksson K. Dynamic changes in glycosyl-ation and glycan composition of serum FSH and LHduring natural ovarian stimulation. Ups J Med Sci.2013;118(3):153–164.
131. Wide L. Median charge and charge heterogeneity ofhuman pituitary FSH, LH and TSH. II. Relationship tosex and age. Acta Endocrinol (Copenh). 1985;109(2):190–197.
132. Esteves SC, Alviggi C. The role of LH in controlledovarian stimulation. In: Ghumman S, ed. Principlesand Practice of Controlled Ovarian Stimulation inART. New Delhi: Springer India; 2015:171–196.
133. Cole LA, Metsch LA, Grotjan HE Jr. Significantsteroidogenic activity of luteinizing hormone ismaintained after enzymatic removal of oligosac-charides. Mol Endocrinol. 1987;1(9):621–627.
134. Green ED, Baenziger JU. Asparagine-linked oligo-saccharides on lutropin, follitropin, and thyrotropin.I. Structural elucidation of the sulfated and sialy-lated oligosaccharides on bovine, ovine, and humanpituitary glycoprotein hormones. J Biol Chem. 1988;263(1):25–35.
135. Wide L, Eriksson K, Sluss PM, Hall JE. Serum half-lifeof pituitary gonadotropins is decreased by sulfo-nation and increased by sialylation in women. J ClinEndocrinol Metab. 2009;94(3):958–964.
136. Burgon PG, Stanton PG, Robertson DM. In vivobioactivities and clearance patterns of highly pu-rified human luteinizing hormone isoforms. Endo-crinology. 1996;137(11):4827–4836.
137. Weisshaar G, Hiyama J, Renwick AG, Nimtz M. NMRinvestigations of the N-linked oligosaccharides atindividual glycosylation sites of human lutropin. EurJ Biochem. 1991;195(1):257–268.
138. Cole LA. Immunoassay of human chorionic go-nadotropin, its free subunits, and metabolites. ClinChem. 1997;43(12):2233–2243.
139. Birken S, Kovalevskaya G, O’Connor J. Metabolismof hCG and hLH to multiple urinary forms. Mol CellEndocrinol. 1996;125(1-2):121–131.
140. Handschuh K, Guibourdenche J, Cocquebert M,Tsatsaris V, Vidaud M, Evain-Brion D, Fournier T.Expression and regulation by PPARgamma of hCGalpha- and beta-subunits: comparison betweenvillous and invasive extravillous trophoblastic cells.Placenta. 2009;30(12):1016–1022.
141. Rull K, Hallast P, Uuskula L, Jackson J, Punab M,Salumets A, Campbell RK, Laan M. Fine-scale
585doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

quantification of HCG beta gene transcription inhuman trophoblastic and non-malignant non-trophoblastic tissues. Mol Hum Reprod. 2008;14(1):23–31.
142. Zhu H, Qiu C, Ruth AC, Keire DA, Ye H. A LC-MS all-in-one workflow for site-specific location, identifi-cation and quantification of N-/O-glycosylation inhuman chorionic gonadotropin drug products.AAPS J. 2017;19(3):846–855.
143. Liu C, Bowers LD. Mass spectrometric character-ization of the b-subunit of human chorionic go-nadotropin. J Mass Spectrom. 1997;32(1):33–42.
144. Valmu L, Alfthan H, Hotakainen K, Birken S, StenmanU-H. Site-specific glycan analysis of human chorionicgonadotropin beta-subunit from malignancies andpregnancy by liquid chromatography—electrospraymass spectrometry. Glycobiology. 2006;16(12):1207–1218.
145. Toll H, Berger P, Hofmann A, Hildebrandt A,Oberacher H, Lenhof HP, Huber CG. Glycosyla-tion patterns of human chorionic gonadotropinrevealed by liquid chromatography-mass spec-trometry and bioinformatics. Electrophoresis. 2006;27(13):2734–2746.
146. Fernandez-Tejada A, Vadola PA, Danishefsky SJ.Chemical synthesis of the b-subunit of humanluteinizing (hLH) and chorionic gonadotropin(hCG) glycoprotein hormones. J Am Chem Soc.2014;136(23):8450–8458.
147. Brun S, Leguy MC, Bruneel A, Fournier T, AnselemO, Guibourdenche J. hCG in screening for aneu-ploidy: a possible role for its glycoforms? Placenta.2014;35(6):425–427.
148. Berndt S, Perrier d’Hauterive S, Blacher S, Pequeux C,Lorquet S, Munaut C, Applanat M, Herve MA,Lamande N, Corvol P, van den Brule F, Frankenne F,Poutanen M, Huhtaniemi I, Geenen V, Noel A,Foidart J-M. Angiogenic activity of human chorionicgonadotropin through LH receptor activation onendothelial and epithelial cells of the endometrium.FASEB J. 2006;20(14):2630–2632.
149. Fournier T, Guibourdenche J, Evain-Brion D. Review:hCGs: different sources of production, differentglycoforms and functions. Placenta. 2015;36(Suppl 1):S60–S65.
150. Prast J, Saleh L, Husslein H, Sonderegger S, Helmer H,Knofler M. Human chorionic gonadotropin stim-ulates trophoblast invasion through extracellularlyregulated kinase and AKT signaling. Endocrinology.2008;149(3):979–987.
151. Guibourdenche J, Handschuh K, Tsatsaris V,Gerbaud P, Leguy MC, Muller F, Brion DE, FournierT. Hyperglycosylated hCG is a marker of earlyhuman trophoblast invasion. J Clin EndocrinolMetab. 2010;95(10):E240–E244.
152. Cole LA. Hyperglycosylated hCG. Placenta. 2007;28(10):977–986.
153. Kovalevskaya G, Birken S, Kakuma T, Ozaki N, Sauer M,Lindheim S, Cohen M, Kelly A, Schlatterer J, O’ConnorJF. Differential expression of human chorionic go-nadotropin (hCG) glycosylation isoforms in failing andcontinuing pregnancies: preliminary characterizationof the hyperglycosylated hCG epitope. J Endocrinol.2002;172(3):497–506.
154. von Schonfeldt V, Rogenhofer N, Ruf K, Thaler CJ,Jeschke U. Sera of patients with recurrent mis-carriages containing anti-trophoblast antibodies(ATAB) reduce hCG and progesterone productionin trophoblast cells in vitro. J Reprod Immunol. 2016;117:52–56.
155. Elliott MM, Kardana A, Lustbader JW, Cole LA. Car-bohydrate and peptide structure of the alpha- andbeta-subunits of human chorionic gonadotropin fromnormal and aberrant pregnancy and choriocarcinoma.Endocrine. 1997;7(1):15–32.
156. Blithe DL. N-linked oligosaccharides on free alphainterfere with its ability to combine with humanchorionic gonadotropin-beta subunit. J Biol Chem.1990;265(35):21951–21956.
157. Brouillet S, Hoffmann P, Chauvet S, Salomon A,Chamboredon S, Sergent F, Benharouga M, Feige JJ,Alfaidy N. Revisiting the role of hCG: new regulationof the angiogenic factor EG-VEGF and its receptors.Cell Mol Life Sci. 2012;69(9):1537–1550.
158. Berndt S, Blacher S, Munaut C, Detilleux J, Perrierd’Hauterive S, Huhtaniemi I, Evain-Brion D, Noel A,Fournier T, Foidart J-M. Hyperglycosylated humanchorionic gonadotropin stimulates angiogenesisthrough TGF-b receptor activation. FASEB J. 2013;27(4):1309–1321.
159. Sengodan SK, Nadhan R, Nair RS, Hemalatha SK,Somasundaram V, Sushama RR, Rajan A, Latha NR,Varghese GR, Thankappan RK, Kumar JM, Chil A,Anilkumar TV, Srinivas P. BRCA1 regulation onb-hCG: a mechanism for tumorigenicity in BRCA1defective breast cancer. Oncogenesis. 2017;6(9):e376.
160. Koistinen H, Hautala L, Koli K, Stenman U-H. Ab-sence of TGF-b receptor activation by highly pu-rified hCG preparations. Mol Endocrinol. 2015;29(12):1787–1791.
161. Lempiainen A, Sankila A, Hotakainen K, Haglund C,Blomqvist C, Stenman U-H. Expression of humanchorionic gonadotropin in testicular germ cell tu-mors. Urol Oncol. 2014;32(5):727–734.
162. Varki A, Kannagi R, Toole B, Stanley P. Glycosylationchanges in cancer. In: Varki A, Cummings RD, EskoJD, Stanley P, Hart GW, Aebi M, Darvill AG,Kinoshita T, Packer NH, Prestegard JH, Schnaar RL,Seeberger PH, eds. Essentials of Glycobiology. 3rd ed.Cold Spring Harbor, NY: Cold Spring Harbor Lab-oratory Press 2017.
163. Cole LA, Butler SA. Hyperglycosylated human cho-rionic gonadotropin and human chorionic gonado-tropin free beta-subunit: tumor markers and tumorpromoters. J Reprod Med. 2008;53(7):499–512.
164. Van Hell JG, Matthijsen R, Overbeek GA. Effects ofhuman menopausal gonadotrophin preparations indifferent bioassay methods. Acta Endocrinol(Copenh). 1964;47:409–418.
165. Catt KJ, Dufau ML. Spare gonadotrophin receptorsin rat testis. Nat New Biol. 1973;244(137):219–221.
166. Younis JS. Commentary: efficacy of follicle-stimulating hormone (FSH) alone, FSH + luteiniz-ing hormone, human menopausal gonadotropin orFSH + human chorionic gonadotropin on assistedreproductive technology outcomes in the “per-sonalized” medicine era: a meta-analysis. FrontEndocrinol (Lausanne). 2017;8:264.
167. Casarini L, Santi D, Simoni M, Potı F. “Spare”luteinizing hormone receptors: facts and fiction.Trends Endocrinol Metab. 2018;29(4):208–217.
168. Mazurkiewicz JE, Herrick-Davis K, Barroso M, Ulloa-Aguirre A, Lindau-Shepard B, Thomas RM, Dias JA.Single-molecule analyses of fully functional fluo-rescent protein-tagged follitropin receptor revealhomodimerization and specific heterodimerizationwith lutropin receptor. Biol Reprod. 2015;92(4):100.
169. Cosowsky L, Lin W, Han Y, Bernard MP, CampbellRK, Moyle WR. Influence of subunit interactions onlutropin specificity. Implications for studies of gly-coprotein hormone function. J Biol Chem. 1997;272(6):3309–3314.
170. Galet C, Ascoli M. The differential binding affinitiesof the luteinizing hormone (LH)/choriogonadotropinreceptor for LH and choriogonadotropin are dic-tated by different extracellular domain residues.MolEndocrinol. 2005;19(5):1263–1276.
171. Nguyen TD, Filliatreau L, Klett D, Combarnous Y.Comparative effects of sub-stimulating concentrations
of non-human versus human Luteinizing Hormones(LH) or chorionic gonadotropins (CG) on adenylatecyclase activation by forskolin inMLTC cells.Gen CompEndocrinol. 2018;261:23–30.
172. Moyle WR, Pressey A, Dean-Emig D, Anderson DM,Demeter M, Lustbader J, Ehrlich P. Detection ofconformational changes in human chorionic go-nadotropin upon binding to rat gonadal receptors.J Biol Chem. 1987;262(35):16920–16926.
173. Keutmann HT, Charlesworth MC, Mason KA,Ostrea T, Johnson L, Ryan RJ. A receptor-bindingregion in human choriogonadotropin/lutropin betasubunit. Proc Natl Acad Sci USA. 1987;84(7):2038–2042.
174. Xia H, Huang J, Chen TM, Puett D. Lysine residues 2and 104 of the human chorionic gonadotrophinbeta subunit influence receptor binding. J MolEndocrinol. 1993;10(3):337–343.
175. Hong S, Ji I, Ji TH. The a-subunit of human cho-riogonadotropin interacts with the exodomain ofthe luteinizing hormone/choriogonadotropin re-ceptor. Endocrinology. 1999;140(6):2486–2493.
176. Vischer HF, Granneman JCM, Noordam MJ,Mosselman S, Bogerd J. Ligand selectivity of go-nadotropin receptors: role of the beta-strands ofextracellular leucine-rich repeats 3 and 6 of thehuman luteinizing hormone receptor. J Biol Chem.2003;278(18):15505–15513.
177. Smits G, Campillo M, Govaerts C, Janssens V, RichterC, Vassart G, Pardo L, Costagliola S. Glycoproteinhormone receptors: determinants in leucine-richrepeats responsible for ligand specificity. EMBO J.2003;22(11):2692–2703.
178. Roy S, Setlur S, Gadkari RA, Krishnamurthy HN,Dighe RR. Translational fusion of two beta-subunitsof human chorionic gonadotropin results in pro-duction of a novel antagonist of the hormone.Endocrinology. 2007;148(8):3977–3986.
179. Narayan P, Gray J, Puett D. A biologically activesingle chain human chorionic gonadotropin analogwith altered receptor binding properties. Endocri-nology. 2000;141(1):67–71.
180. Bernard MP, Lin W, Cao D, Myers RV, Xing Y, MoyleWR. Only a portion of the small seatbelt loop inhuman choriogonadotropin appears capable ofcontacting the lutropin receptor. J Biol Chem. 2004;279(43):44438–44441.
181. Shao K, Purohit S, Bahl OP. Effect of modification ofall loop regions in the alpha- and beta-subunits ofhuman choriogonadotropin on its signal trans-duction activity. Mol Cell Endocrinol. 1996;122(2):173–182.
182. Hu SB, Johnson L, Roche PC, Keutmann HT. Afunctional determinant in human luteinizing hor-mone and chorionic gonadotropin: differential ef-fect of mutations about beta-GLN-54. Endocrinology.1997;138(4):1627–1633.
183. Jiang X, Dias JA, He X. Structural biology of glyco-protein hormones and their receptors: insights tosignaling. Mol Cell Endocrinol. 2014;382(1):424–451.
184. Rivero-Muller A, Chou Y-Y, Ji I, Lajic S, HanyalogluAC, Jonas K, Rahman N, Ji TH, Huhtaniemi I. Rescueof defective G protein-coupled receptor functionin vivo by intermolecular cooperation. Proc NatlAcad Sci USA. 2010;107(5):2319–2324.
185. Grzesik P, Teichmann A, Furkert J, Rutz C, Wiesner B,Kleinau G, Schulein R, Gromoll J, Krause G. Differencesbetween lutropin-mediated and choriogonadotropin-mediated receptor activation. FEBS J. 2014;281(5):1479–1492.
186. Lee C, Ji I, Ryu K, Song Y, Conn PM, Ji TH. Twodefective heterozygous luteinizing hormone re-ceptors can rescue hormone action. J Biol Chem.2002;277(18):15795–15800.
586 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

187. Jeoung M, Lee C, Ji I, Ji TH. Trans-activation, cis-activation and signal selection of gonadotropin re-ceptors. Mol Cell Endocrinol. 2007;260-262:137–143.
188. Zhang M, Guan R, Segaloff DL. Revisiting andquestioning functional rescue between dimerizedLH receptor mutants. Mol Endocrinol. 2012;26(4):655–668.
189. Alvarez CA, Narayan P, Huang J, Puett D. Charac-terization of a region of the lutropin receptor ex-tracellular domain near transmembrane helix 1 thatis important in ligand-mediated signaling. Endocri-nology. 1999;140(4):1775–1782.
190. Ji TH, Ryu KS, Gilchrist R, Ji I. Interaction, signalgeneration, signal divergence, and signal trans-duction of LH/CG and the receptor. Recent ProgHorm Res. 1997;52:431–453, discussion 454.
191. Herrlich A, Kuhn B, Grosse R, Schmid A, Schultz G,Gudermann T. Involvement of Gs and Gi proteinsin dual coupling of the luteinizing hormone re-ceptor to adenylyl cyclase and phospholipase C.J Biol Chem. 1996;271(28):16764–16772.
192. Nakamura K, Lazari MF, Li S, Korgaonkar C, AscoliM. Role of the rate of internalization of the agonist-receptor complex on the agonist-induced down-regulation of the lutropin/choriogonadotropinreceptor. Mol Endocrinol. 1999;13(8):1295–1304.
193. Gudermann T, Nichols C, Levy FO, Birnbaumer M,Birnbaumer L. Ca2+ mobilization by the LH re-ceptor expressed in Xenopus oocytes independentof 39,59-cyclic adenosine monophosphate forma-tion: evidence for parallel activation of two signalingpathways. Mol Endocrinol. 1992;6(2):272–278.
194. Trehan A, Rotgers E, Coffey ET, Huhtaniemi I,Rivero-Muller A. CANDLES, an assay for monitoringGPCR induced cAMP generation in cell cultures.Cell Commun Signal. 2014;12(1):70.
195. Ayoub MA, Landomiel F, Gallay N, Jegot G, PouponA, Crepieux P, Reiter E. Assessing gonadotropinreceptor function by resonance energy transfer-based assays. Front Endocrinol (Lausanne). 2015;6:130.
196. Montminy MR, Bilezikjian LM. Binding of a nu-clear protein to the cyclic-AMP response elementof the somatostatin gene. Nature. 1987;328(6126):175–178.
197. Casarini L, Reiter E, Simoni M. b-Arrestins regulategonadotropin receptor-mediated cell proliferationand apoptosis by controlling different FSHR orLHCGR intracellular signaling in the hGL5 cell line.Mol Cell Endocrinol. 2016;437:11–21.
198. Zwain IH, Amato P. cAMP-induced apoptosis ingranulosa cells is associated with up-regulation ofP53 and bax and down-regulation of clusterin.Endocr Res. 2001;27(1-2):233–249.
199. Tajima K, Yoshii K, Fukuda S, Orisaka M, MiyamotoK, Amsterdam A, Kotsuji F. Luteinizing hormone-induced extracellular-signal regulated kinase acti-vation differently modulates progesterone andandrostenedione production in bovine theca cells.Endocrinology. 2005;146(7):2903–2910.
200. Menon B, Franzo-Romain M, Damanpour S, MenonKMJ. Luteinizing hormone receptor mRNA down-regulation is mediated through ERK-dependentinduction of RNA binding protein. Mol Endo-crinol. 2011;25(2):282–290.
201. Ren X-R, Reiter E, Ahn S, Kim J, Chen W, LefkowitzRJ. Different G protein-coupled receptor kinasesgovern G protein and beta-arrestin-mediated sig-naling of V2 vasopressin receptor. Proc Natl Acad SciUSA. 2005;102(5):1448–1453.
202. Tai P, Shiraishi K, Ascoli M. Activation of thelutropin/choriogonadotropin receptor inhibits ap-optosis of immature Leydig cells in primary culture.Endocrinology. 2009;150(8):3766–3773.
203. Martinat N, Crepieux P, Reiter E, Guillou F. Extra-cellular signal-regulated kinases (ERK) 1, 2 are re-quired for luteinizing hormone (LH)-inducedsteroidogenesis in primary Leydig cells and controlsteroidogenic acute regulatory (StAR) expression.Reprod Nutr Dev. 2005;45(1):101–108.
204. Tohgo A, Choy EW, Gesty-Palmer D, Pierce KL,Laporte S, Oakley RH, Caron MG, Lefkowitz RJ,Luttrell LM. The stability of the G protein-coupledreceptor-b-arrestin interaction determines themechanism and functional consequence of ERKactivation. J Biol Chem. 2003;278(8):6258–6267.
205. Revankar CM, Vines CM, Cimino DF, Prossnitz ER.Arrestins block G protein-coupled receptor-mediatedapoptosis. J Biol Chem. 2004;279(23):24578–24584.
206. Gudermann T, Birnbaumer M, Birnbaumer L. Evi-dence for dual coupling of the murine luteinizinghormone receptor to adenylyl cyclase and phos-phoinositide breakdown and Ca2+ mobilization:studies with the cloned murine luteinizing hor-mone receptor expressed in L cells. J Biol Chem.1992;267(7):4479–4488.
207. Zhu X, Gilbert S, Birnbaumer M, Birnbaumer L. Dualsignaling potential is common among Gs-coupledreceptors and dependent on receptor density. MolPharmacol. 1994;46(3):460–469.
208. Manna PR, Pakarinen P, El-Hefnawy T, Huhtaniemi IT.Functional assessment of the calcium messengersystem in cultured mouse Leydig tumor cells: regu-lation of human chorionic gonadotropin-inducedexpression of the steroidogenic acute regulatoryprotein. Endocrinology. 1999;140(4):1739–1751.
209. Pezzi V, Clark BJ, Ando S, Stocco DM, Rainey WE.Role of calmodulin-dependent protein kinase II inthe acute stimulation of aldosterone production.J Steroid Biochem Mol Biol. 1996;58(4):417–424.
210. Mertens-Walker I, Bolitho C, Baxter RC, Marsh DJ.Gonadotropin-induced ovarian cancer cell migra-tion and proliferation require extracellular signal-regulated kinase 1/2 activation regulated by calciumand protein kinase Cdelta. Endocr Relat Cancer.2010;17(2):335–349.
211. Andric N, Ascoli M. Mutations of the lutropin/choriogonadotropin receptor that do not activatethe phosphoinositide cascade allow hCG to inducearomatase expression in immature rat granulosacells. Mol Cell Endocrinol. 2008;285(1-2):62–72.
212. Light A, Hammes SR. Membrane receptor cross talkin steroidogenesis: recent insights and clinical im-plications. Steroids. 2013;78(6):633–638.
213. Tranchant T, Durand G, Gauthier C, Crepieux P,Ulloa-Aguirre A, Royere D, Reiter E. Preferentialb-arrestin signalling at low receptor densityrevealed by functional characterization of the hu-man FSH receptor A189 V mutation. Mol CellEndocrinol. 2011;331(1):109–118.
214. Donadeu FX, Ascoli M. The differential effects of thegonadotropin receptors on aromatase expression inprimary cultures of immature rat granulosa cells arehighly dependent on the density of receptorsexpressed and the activation of the inositol phosphatecascade. Endocrinology. 2005;146(9):3907–3916.
215. Lyga S, Volpe S, Werthmann RC, Gotz K, SungkawornT, Lohse MJ, Calebiro D. Persistent cAMP signaling byinternalized LH receptors in ovarian follicles. Endo-crinology. 2016;157(4):1613–1621.
216. Hunzicker-Dunn M, Barisas G, Song J, Roess DA.Membrane organization of luteinizing hormonereceptors differs between actively signaling anddesensitized receptors. J Biol Chem. 2003;278(44):42744–42749.
217. Hirakawa T, Galet C, Kishi M, Ascoli M. GIPC bindsto the human lutropin receptor (hLHR) through anunusual PDZ domain binding motif, and it regulates
the sorting of the internalized human choriogo-nadotropin and the density of cell surface hLHR.J Biol Chem. 2003;278(49):49348–49357.
218. Roess DA, Horvat RD, Munnelly H, Barisas BG.Luteinizing hormone receptors are self-associated inthe plasma membrane. Endocrinology. 2000;141(12):4518–4523.
219. Thomsen ARB, Plouffe B, Cahill TJ III, Shukla AK,Tarrasch JT, Dosey AM, Kahsai AW, Strachan RT,Pani B, Mahoney JP, Huang L, Breton B, HeydenreichFM, Sunahara RK, Skiniotis G, Bouvier M, LefkowitzRJ. GPCR-G protein-b-arrestin super-complex me-diates sustained G protein signaling. Cell. 2016;166(4):907–919.
220. Sposini S, Jean-Alphonse FG, Ayoub MA, Oqua A,West C, Lavery S, Brosens JJ, Reiter E, Hanyaloglu AC.Integration of GPCR signaling and sorting from veryearly endosomes via opposing APPL1 mechanisms.Cell Reports. 2017;21(10):2855–2867.
221. Riccetti L, Yvinec R, Klett D, Gallay N, CombarnousY, Reiter E, Simoni M, Casarini L, AyoubMA. Humanluteinizing hormone and chorionic gonadotropindisplay biased agonism at the LH and LH/CG re-ceptors. Sci Rep. 2017;7(1):940.
222. Jonas KC, Fanelli F, Huhtaniemi IT, Hanyaloglu AC.Single molecule analysis of functionally asymmetricG protein-coupled receptor (GPCR) oligomers re-veals diverse spatial and structural assemblies. J BiolChem. 2015;290(7):3875–3892.
223. Gupta C, Chapekar T, Chhabra Y, Singh P, Sinha S,Luthra K. Differential response to sustained stim-ulation by hCG & LH on goat ovarian granulosacells. Indian J Med Res. 2012;135:331–340.
224. Casarini L, Riccetti L, De Pascali F, Nicoli A, TagliaviniS, Trenti T, La Sala GB, Simoni M. Follicle-stimulatinghormone potentiates the steroidogenic activity ofchorionic gonadotropin and the anti-apoptoticactivity of luteinizing hormone in human granulosa-lutein cells in vitro. Mol Cell Endocrinol. 2016;422:103–114.
225. Casarini L, Riccetti L, De Pascali F, Gilioli L, MarinoM, Vecchi E, Morini D, Nicoli A, La Sala GB, SimoniM. Estrogen modulates specific life and deathsignals induced by LH and hCG in human primarygranulosa cells in vitro. Int J Mol Sci. 2017;18(5):E926.
226. Kristensen SG, Mamsen LS, Jeppesen JV, Bøtkjær JA,Pors SE, Borgbo T, Ernst E, Macklon KT, AndersenCY. Hallmarks of human small antral follicle de-velopment: implications for regulation of ovariansteroidogenesis and selection of the dominantfollicle. Front Endocrinol (Lausanne). 2018;8:376.
227. Wickenheisser JK, Nelson-DeGrave VL, McAllisterJM. Human ovarian theca cells in culture. TrendsEndocrinol Metab. 2006;17(2):65–71.
228. Saint-Dizier M, Foulon-Gauze F, Lecompte F,Combarnous Y, Chopineau M. The additionalN-glycosylation site of the equine LH/CG receptor isnot responsible for the limited cyclic AMP pathwayactivation by equine chorionic gonadotropin rela-tive to luteinizing hormone. Reprod Biol. 2011;11(2):157–164.
229. Klett D, Meslin P, Relav L, Nguyen TMD, Mariot J,Jegot G, Cahoreau C, Combarnous Y. Low re-versibility of intracellular cAMP accumulation inmouse Leydig tumor cells (MLTC-1) stimulated byhuman Luteinizing Hormone (hLH) and ChorionicGonadotropin (hCG).Mol Cell Endocrinol. 2016;434:144–153.
230. Santi D, Spaggiari G, Casarini L, Fanelli F, Mezzullo M,Pagotto U, Granata ARM, Carani C, Simoni M.Central hypogonadism due to a giant, “silent” FSH-secreting, atypical pituitary adenoma: effects ofadenoma dissection and short-term Leydig cellstimulation by luteinizing hormone (LH) and
587doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

human chorionic gonadotropin (hCG). Aging Male.2017;20(2):96–101.
231. Tyrey L. Human chorionic gonadotropin: propertiesand assay methods. Semin Oncol. 1995;22(2):121–129.
232. Christin-Maitre S, Vasseur C, Fauser B, Bouchard P.Bioassays of gonadotropins. Methods. 2000;21(1):51–57.
233. Qiu M, Quan F, Han C, Wu B, Liu J, Yang Z, Su F,Zhang Y. Effects of granulosa cells on steroido-genesis, proliferation and apoptosis of stromal cellsand theca cells derived from the goat ovary. J SteroidBiochem Mol Biol. 2013;138:325–333.
234. Brown C, LaRocca J, Pietruska J, Ota M, Anderson L,Smith SD, Weston P, Rasoulpour T, Hixon ML.Subfertility caused by altered follicular developmentand oocyte growth in female mice lacking PKBalpha/Akt1. Biol Reprod. 2010;82(2):246–256.
235. Rao CV. Protective effects of human chorionicgonadotropin against breast cancer: how can weuse this information to prevent/treat the disease?Reprod Sci. 2017;24(8):1102–1110.
236. Pennanen P, Peltonen S, Kallionpaa RA, Peltonen J.The effect of estradiol, testosterone, and humanchorionic gonadotropin on the proliferation ofSchwann cells with NF1 +/- or NF1 -/- genotypederived from human cutaneous neurofibromas[published online ahead of print November 28,2017]. Mol Cell Biochem.
237. Kayampilly PP, Menon KMJ. AMPK activationby dihydrotestosterone reduces FSH-stimulatedcell proliferation in rat granulosa cells by inhibit-ing ERK signaling pathway. Endocrinology. 2012;153(6):2831–2838.
238. Lohse MJ, Hoffmann C. Arrestin interactions with Gprotein-coupled receptors. Handb Exp Pharmacol.2014;219:15–56.
239. Bradbury FA, Menon KMJ. Evidence that constitu-tively active luteinizing hormone/human chorionicgonadotropin receptors are rapidly internalized.Biochemistry. 1999;38(27):8703–8712.
240. Check JH, Davies E, Brasile D, Choe JK, Amui J. Aprospective comparison of in vitro fertilization (IVF)outcome following controlled ovarian hyperstim-ulation (COH) regimens using follitropin alphaexclusively or with the addition of low dose humanchorionic gonadotropin (hCG) and ganirelix. ClinExp Obstet Gynecol. 2009;36(4):217–218.
241. Berkkanoglu M, Isikoglu M, Aydin D, Ozgur K. Clinicaleffects of ovulation induction with recombinantfollicle-stimulating hormone supplemented withrecombinant luteinizing hormone or low-doserecombinant human chorionic gonadotropin in themidfollicular phase in microdose cycles in poor re-sponders. Fertil Steril. 2007;88(3):665–669.
242. Sasson R, Shinder V, Dantes A, Land A, AmsterdamA. Activation of multiple signal transductionpathways by glucocorticoids: protection of ovarianfollicular cells against apoptosis. Biochem BiophysRes Commun. 2003;311(4):1047–1056.
243. Li Q, He H, Zhang Y-L, Li X-M, Guo X, Huo R, Bi Y, Li J,Fan H-Y, Sha J. Phosphoinositide 3-kinase p110dmediates estrogen- and FSH-stimulated ovarian folliclegrowth. Mol Endocrinol. 2013;27(9):1468–1482.
244. Couse JF, Yates MM, Deroo BJ, Korach KS. Estrogenreceptor-b is critical to granulosa cell differentiationand the ovulatory response to gonadotropins. En-docrinology. 2005;146(8):3247–3262.
245. Hirata R, Hojo T, Sano M, Hayashi N, Okuda K.Potential role of hCG in apoptosis of humanluteinized granulosa cells. J Reprod Dev. 2015;61(1):67–73.
246. Puttabyatappa M, Brogan RS, Vandevoort CA,Chaffin CL. EGF-like ligands mediate progesterone’s
anti-apoptotic action on macaque granulosa cells.Biol Reprod. 2013;88(1):18.
247. Kariagina A, Xie J, Leipprandt JR, Haslam SZ.Amphiregulin mediates estrogen, progesterone, andEGFR signaling in the normal rat mammary glandand in hormone-dependent rat mammary cancers.Horm Cancer. 2010;1(5):229–244.
248. Salazar M, Lerma-Ortiz A, Hooks GM, Ashley AK,Ashley RL. Progestin-mediated activation of MAPKand AKT in nuclear progesterone receptor negativebreast epithelial cells: the role of membrane pro-gesterone receptors. Gene. 2016;591(1):6–13.
249. Faraoni EY, Camilletti MA, Abeledo-Machado A,Ratner LD, De Fino F, Huhtaniemi I, Rulli SB, Dıaz-Torga G. Sex differences in the development ofprolactinoma in mice overexpressing hCGb: role ofTGFb1. J Endocrinol. 2017;232(3):535–546.
250. Huhtaniemi I, Rulli S, Ahtiainen P, Poutanen M.Multiple sites of tumorigenesis in transgenic miceoverproducing hCG. Mol Cell Endocrinol. 2005;234(1-2):117–126.
251. Yamashita S, Tai P, Charron J, Ko C, Ascoli M. TheLeydig cell MEK/ERK pathway is critical formaintaining a functional population of adult Leydigcells and for fertility. Mol Endocrinol. 2011;25(7):1211–1222.
252. Casarini L, Pignatti E, Simoni M. Effects of poly-morphisms in gonadotropin and gonadotropinreceptor genes on reproductive function. RevEndocr Metab Disord. 2011;12(4):303–321.
253. Casarini L, Huhtaniemi IT, Simoni M, Rivero-MullerA. Gonadotropin receptors. In: Simoni M,Huhtaniemi IT, eds. Endocrinology of the Testis andMale Reproduction. Endocrinology. Cham: Springer;2017:123–168.
254. Ma X, Dong Y, Matzuk MM, Kumar TR. Targeteddisruption of luteinizing hormone beta-subunitleads to hypogonadism, defects in gonadal ste-roidogenesis, and infertility. Proc Natl Acad Sci USA.2004;101(49):17294–17299.
255. Lofrano-Porto A, Barra GB, Giacomini LA,Nascimento PP, Latronico AC, Casulari LA, da RochaNeves FA. Luteinizing hormone beta mutation andhypogonadism in men and women. N Engl J Med.2007;357(9):897–904.
256. Pakarainen T, Zhang F-P, Nurmi L, Poutanen M,Huhtaniemi I. Knockout of luteinizing hormonereceptor abolishes the effects of follicle-stimulatinghormone on preovulatory maturation and ovula-tion of mouse graafian follicles.Mol Endocrinol. 2005;19(10):2591–2602.
257. Pettersson K, Ding YQ, Huhtaniemi I. An immu-nologically anomalous luteinizing hormone variantin a healthy woman. J Clin Endocrinol Metab. 1992;74(1):164–171.
258. Nilsson C, Jiang M, Pettersson K, Iitia A, Makela M,Simonsen H, Easteal S, Herrera RJ, Huhtaniemi I. De-termination of a common genetic variant of luteinizinghormone using DNA hybridization and immunoassays.Clin Endocrinol (Oxf). 1998;49(3):369–376.
259. Suganuma N, Furui K, Kikkawa F, Tomoda Y,Furuhashi M. Effects of the mutations (Trp8 –.Arg and Ile15 –. Thr) in human luteinizing hor-mone (LH) beta-subunit on LH bioactivity in vitroand in vivo. Endocrinology. 1996;137(3):831–838.
260. Furui K, SuganumaN, Tsukahara S, Asada Y, Kikkawa F,Tanaka M, Ozawa T, Tomoda Y. Identification of twopoint mutations in the gene coding luteinizing hor-mone (LH) beta-subunit, associated with immuno-logically anomalous LH variants. J Clin EndocrinolMetab. 1994;78(1):107–113.
261. Liao WX, Goh HH, Roy AC. Functional character-ization of a natural variant of luteinizing hormone.Hum Genet. 2002;111(2):219–224.
262. Jiang M, Lamminen T, Pakarinen P, Hellman J,Manna P, Herrera RJ, Huhtaniemi I. A novel Ala(-3)Thr mutation in the signal peptide of humanluteinizing hormone beta-subunit: potentiation ofthe inositol phosphate signalling pathway and at-tenuation of the adenylate cyclase pathway byrecombinant variant hormone. Mol Hum Reprod.2002;8(3):201–212.
263. Bosco L, Ruvolo G, Luparello C, Ferrari S, Valerio D,Santi D, Piomboni P, Sarcina E, Lispi M, RoccheriMC. Gene expression and apoptosis levels in cu-mulus cells of patients with polymorphisms of FSHRand LHB undergoing in vitro fertilization program.Cell Physiol Biochem. 2017;43(6):2391–2404.
264. Jiang M, Pakarinen P, Zhang FP, El-Hefnawy T,Koskimies P, Pettersson K, Huhtaniemi I. A com-mon polymorphic allele of the human luteinizinghormone beta-subunit gene: additional mutationsand differential function of the promoter sequence.Hum Mol Genet. 1999;8(11):2037–2046.
265. Tapanainen JS, Koivunen R, Fauser BC, Taylor AE,Clayton RN, Rajkowa M, White D, Franks S, AnttilaL, Pettersson KS, Huhtaniemi IT. A new contributingfactor to polycystic ovary syndrome: the geneticvariant of luteinizing hormone. J Clin EndocrinolMetab. 1999;84(5):1711–1715.
266. Liu S, Ogata T, Maruyama T, Yoshimura Y, IshizukaB. Association of common LH variant with hy-perfunctional promoter in a Japanese infertilewoman. Endocr J. 2005;52(6):781–784.
267. Punab AM, Grigorova M, Punab M, Adler M, KuuraT, Poolamets O, Vihljajev V, Zilaitiene B, Erenpreiss J,Matulevicius V, Laan M. Carriers of V-LH among1593 Baltic men have significantly higher serum LH.Andrology. 2015;3(3):512–519.
268. Shiraishi K, Naito K. Fertile eunuch syndrome withthe mutations (Trp8Arg and Ile15Thr) in the betasubunit of luteinizing hormone. Endocr J. 2003;50(6):733–737.
269. Dasgupta S, Sirisha PVS, Neelaveni K, Anuradha K,Sudhakar G, Reddy BM. Role of luteinizing hormoneb-subunit gene variants among South Indianwomen with polycystic ovary syndrome. Gene. 2012;494(1):51–56.
270. Cho SG, Bhoumik A, Broday L, Ivanov V, RosensteinB, Ronai Z. TIP49b, a regulator of activating tran-scription factor 2 response to stress and DNAdamage. Mol Cell Biol. 2001;21(24):8398–8413.
271. Zhao Y, Chen T, Zhou Y, Li K, Xiao J. An associationstudy between the genetic polymorphisms withinGnRHI, LHb and FSHb genes and central pre-cocious puberty in Chinese girls. Neurosci Lett. 2010;486(3):188–192.
272. Lamminen T, Jiang M, Manna PR, Pakarinen P,Simonsen H, Herrera RJ, Huhtaniemi I. Functionalstudy of a recombinant form of human LHbeta-subunit variant carrying the Gly(102)Ser mutationfound in Asian populations. Mol Hum Reprod. 2002;8(10):887–892.
273. Miller-Lindholm AK, Bedows E, Bartels CF, Ramey J,Maclin V, Ruddon RW. A naturally occurring ge-netic variant in the human chorionic gonadotropin-beta gene 5 is assembly inefficient. Endocrinology.1999;140(8):3496–3506.
274. Rull K, Nagirnaja L, Ulander V-M, Kelgo P, Margus T,Kaare M, Aittomaki K, Laan M. Chorionic gonad-otropin beta-gene variants are associated with re-current miscarriage in two European populations.J Clin Endocrinol Metab. 2008;93(12):4697–4706.
275. Casarini L, Brigante G. The polycystic ovarysyndrome evolutionary paradox: a genome-wideassociation studies-based, in silico, evolutionaryexplanation. J Clin Endocrinol Metab. 2014;99(11):E2412–E2420.
588 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

276. Nishimura R, Shin J, Ji I, Middaugh CR, Kruggel W,Lewis RV, Ji TH. A single amino acid substitution inan ectopic alpha subunit of a human carcinomachoriogonadotropin. J Biol Chem. 1986;261(23):10475–10477.
277. Nagirnaja L, Venclovas C, Rull K, Jonas KC, PeltoketoH, Christiansen OB, Kairys V, Kivi G, Steffensen R,Huhtaniemi IT, Laan M. Structural and functionalanalysis of rare missense mutations in humanchorionic gonadotrophin b-subunit. Mol HumReprod. 2012;18(8):379–390.
278. Basciani S, Watanabe M, Mariani S, Passeri M,Persichetti A, Fiore D, Scotto d’Abusco A, Caprio M,Lenzi A, Fabbri A, Gnessi L. Hypogonadism in apatient with two novel mutations of the luteinizinghormone b-subunit gene expressed in a compoundheterozygous form. J Clin Endocrinol Metab. 2012;97(9):3031–3038.
279. Valdes-Socin H, Salvi R, Daly AF, Gaillard RC,Quatresooz P, Tebeu P-M, Pralong FP, Beckers A.Hypogonadism in a patient with a mutation in theluteinizing hormone beta-subunit gene. N Engl JMed. 2004;351(25):2619–2625.
280. Potorac I, Rivero-Muller A, Trehan A, Kiełbus M,Jozwiak K, Pralong F, Hafidi A, Thiry A, Menage J-J,Huhtaniemi I, Beckers A, Daly AF. A vital region forhuman glycoprotein hormone trafficking revealedby an LHB mutation. J Endocrinol. 2016;231(3):197–207.
281. Valdes-Socin H, Salvi R, Thiry A, Daly AF, Pralong FP,Gaillard R, Beckers A. Testicular effects of isolatedluteinizing hormone deficiency and reversal bylong-term human chorionic gonadotropin treat-ment. J Clin Endocrinol Metab. 2009;94(1):3–4.
282. Achard C, Courtillot C, Lahuna O, Meduri G, SoufirJ-C, Liere P, Bachelot A, Benyounes H, SchumacherM, Kuttenn F, Touraine P, Misrahi M. Normalspermatogenesis in a man with mutant luteinizinghormone. N Engl J Med. 2009;361(19):1856–1863.
283. Takahashi K, Karino K, Kanasaki H, Kurioka H, OzakiT, Yonehara T, Miyazaki K. Influence of missensemutation and silent mutation of LHbeta-subunitgene in Japanese patients with ovulatory disorders.Eur J Hum Genet. 2003;11(5):402–408.
284. Simoni M, Tuttelmann F, Michel C, Bockenfeld Y,Nieschlag E, Gromoll J. Polymorphisms of theluteinizing hormone/chorionic gonadotropin re-ceptor gene: association with maldescended testesand male infertility. Pharmacogenet Genomics. 2008;18(3):193–200.
285. Chen Z-J, Zhao H, He L, Shi Y, Qin Y, Shi Y, Li Z, YouL, Zhao J, Liu J, Liang X, Zhao X, Zhao J, Sun Y, ZhangB, Jiang H, Zhao D, Bian Y, Gao X, Geng L, Li Y, ZhuD, Sun X, Xu JE, Hao C, Ren CE, Zhang Y, Chen S,Zhang W, Yang A, Yan J, Li Y, Ma J, Zhao Y.Genome-wide association study identifies suscep-tibility loci for polycystic ovary syndrome onchromosome 2p16.3, 2p21 and 9q33.3. Nat Genet.2011;43(1):55–59.
286. Piersma D, Berns EMJJ, Verhoef-Post M, UitterlindenAG, Braakman I, Pols HAP, Themmen APN. Acommon polymorphism renders the luteinizinghormone receptor protein more active by im-proving signal peptide function and predicts ad-verse outcome in breast cancer patients. J ClinEndocrinol Metab. 2006;91(4):1470–1476.
287. Rodien P, Cetani F, Costagliola S, Tonacchera M,Duprez L, Minegishi T, Govaerts C, Vassart G. Evi-dences for an allelic variant of the human LC/CGreceptor rather than a gene duplication: functionalcomparison of wild-type and variant receptors.J Clin Endocrinol Metab. 1998;83(12):4431–4434.
288. Kossack N, Simoni M, Richter-Unruh A, ThemmenAPN, Gromoll J. Mutations in a novel, cryptic exon
of the luteinizing hormone/chorionic gonadotropinreceptor gene cause male pseudohermaphroditism.PLoS Med. 2008;5(4):e88.
289. Kossack N, Troppmann B, Richter-Unruh A, KleinauG, Gromoll J. Aberrant transcription of the LHCGRgene caused by a mutation in exon 6A leads toLeydig cell hypoplasia type II. Mol Cell Endocrinol.2013;366(1):59–67.
290. Liu W, Han B, ZhuW, Cheng T, Fan M, Wu J, Yang Y,Zhu H, Si J, Lyu Q, Chai W, Zhao S, Song H, Kuang Y,Qiao J. Polymorphism in the alternative donor siteof the cryptic exon of LHCGR: functional conse-quences and associations with testosterone level.Sci Rep. 2017;7:45699.
291. Frendo J-L, Guibourdenche J, Pidoux G, Vidaud M,Luton D, Giovangrandi Y, Porquet D, Muller F,Evain-Brion D. Trophoblast production of a weaklybioactive human chorionic gonadotropin in tri-somy 21-affected pregnancy. J Clin EndocrinolMetab. 2004;89(2):727–732.
292. Sutton JM, Cole LA. Sialic acid-deficient invasivetrophoblast antigen (sd-ITA): a new urinary variantfor gestational Down syndrome screening. PrenatDiagn. 2004;24(3):194–197.
293. Talbot JA, Spencer K, Abushoufa RA. Detection ofmaternal serum hCG glycoform variants in thesecond trimester of pregnancies affected by Downsyndrome using a lectin immunoassay. PrenatDiagn. 2003;23(1):1–5.
294. Plockinger U, Chrusciel M, Doroszko M, Saeger W,Blankenstein O, Weizsacker K, Kroiss M, HauptmannK, Radke C, Pollinger A, Tiling N, Steinmuller T,Huhtaniemi I, Quinkler M, Bertherat J, Lacroix A,Rahman N. Functional implications of LH/hCG re-ceptors in pregnancy-induced Cushing syndrome.J Endocr Soc. 2017;1(1):57–71.
295. Latronico AC, Anasti J, Arnhold IJ, Rapaport R,Mendonca BB, Bloise W, Castro M, Tsigos C,Chrousos GP. Brief report: testicular and ovarianresistance to luteinizing hormone caused by inac-tivating mutations of the luteinizing hormone-receptor gene. N Engl J Med. 1996;334(8):507–512.
296. Basaria S. Male hypogonadism. Lancet. 2014;383(9924):1250–1263.
297. Topaloglu AK. Update on the genetics of idiopathichypogonadotropic hypogonadism. J Clin Res PediatrEndocrinol. 2017;9(Suppl 2):113–122.
298. Vezzoli V, Duminuco P, Vottero A, Kleinau G,Schulein R, Minari R, Bassi I, Bernasconi S, Persani L,Bonomi M. A new variant in signal peptide of thehuman luteinizing hormone receptor (LHCGR) af-fects receptor biogenesis causing Leydig cell hy-poplasia. Hum Mol Genet. 2015;24(21):6003–6012.
299. Kremer H, Kraaij R, Toledo SP, Post M, Fridman JB,Hayashida CY, van Reen M, Milgrom E, Ropers HH,Mariman E, Themmen APN, Brunner HG. Malepseudohermaphroditism due to a homozygousmissense mutation of the luteinizing hormonereceptor gene. Nat Genet. 1995;9(2):160–164.
300. Arnhold IJ, Lofrano-Porto A, Latronico AC. Inactivatingmutations of luteinizing hormone b-subunit orluteinizing hormone receptor cause oligo-amenorrheaand infertility in women. Horm Res. 2009;71(2):75–82.
301. Weiss J, Axelrod L, Whitcomb RW, Harris PE,Crowley WF, Jameson JL. Hypogonadism causedby a single amino acid substitution in the betasubunit of luteinizing hormone. N Engl J Med. 1992;326(3):179–183.
302. Yang X, Ochin H, Shu L, Liu J, Shen J, Liu J, Lin C, CuiY. Homozygous nonsense mutation Trp28X in theLHB gene causes male hypogonadism. J AssistReprod Genet. 2018;35(5):913–919.
303. Lee C, Ji I, Ji TH. Distinct mechanisms of cAMPinduction by constitutively activating LH receptor
and wild-type LH receptor activated by hCG. En-docrine. 2004;25(2):111–115.
304. Azziz R, Carmina E, Chen Z, Dunaif A, Laven JSE,Legro RS, Lizneva D, Natterson-Horowtiz B, TeedeHJ, Yildiz BO. Polycystic ovary syndrome. Nat Rev DisPrimers. 2016;2:16057.
305. Rosenfield RL, Ehrmann DA. The pathogenesis ofpolycystic ovary syndrome (PCOS): the hypothesisof PCOS as functional ovarian hyperandrogenismrevisited. Endocr Rev. 2016;37(5):467–520.
306. Unluturk U, Sezgin E, Yildiz BO. Evolutionary de-terminants of polycystic ovary syndrome: part 1.Fertil Steril. 2016;106(1):33–41.
307. Fessler DMT, Natterson-Horowitz B, Azziz R. Evo-lutionary determinants of polycystic ovary syn-drome: part 2. Fertil Steril. 2016;106(1):42–47.
308. Casarini L, Simoni M, Brigante G. Is polycystic ovarysyndrome a sexual conflict? A review. ReprodBiomed Online. 2016;32(4):350–361.
309. Shi Y, Zhao H, Shi Y, Cao Y, Yang D, Li Z, Zhang B,Liang X, Li T, Chen J, Shen J, Zhao J, You L, Gao X,Zhu D, Zhao X, Yan Y, Qin Y, Li W, Yan J, Wang Q,Zhao J, Geng L, Ma J, Zhao Y, He G, Zhang A, Zou S,Yang A, Liu J, Li W, Li B, Wan C, Qin Y, Shi J, Yang J,Jiang H, Xu JE, Qi X, Sun Y, Zhang Y, Hao C, Ju X,Zhao D, Ren CE, Li X, Zhang W, Zhang Y, Zhang J,Wu D, Zhang C, He L, Chen Z-J. Genome-wideassociation study identifies eight new risk loci forpolycystic ovary syndrome. Nat Genet. 2012;44(9):1020–1025.
310. Chen Z-J, Zhao H, He L, Shi Y, Qin Y, Shi Y, Li Z, YouL, Zhao J, Liu J, Liang X, Zhao X, Zhao J, Sun Y, ZhangB, Jiang H, Zhao D, Bian Y, Gao X, Geng L, Li Y, ZhuD, Sun X, Xu JE, Hao C, Ren CE, Zhang Y, Chen S,Zhang W, Yang A, Yan J, Li Y, Ma J, Zhao Y.Genome-wide association study identifies suscep-tibility loci for polycystic ovary syndrome onchromosome 2p16.3, 2p21 and 9q33.3. Nat Genet.2011;43(1):55–59.
311. Palioura E, Diamanti-Kandarakis E. Polycystic ovarysyndrome (PCOS) and endocrine disruptingchemicals (EDCs). Rev Endocr Metab Disord. 2015;16(4):365–371.
312. Wachs DS, Coffler MS, Malcom PJ, Shimasaki S,Chang RJ. Increased androgen response to follicle-stimulating hormone administration in womenwith polycystic ovary syndrome. J Clin EndocrinolMetab. 2008;93(5):1827–1833.
313. Franks S, Adams J, Mason H, Polson D. Ovulatorydisorders in women with polycystic ovary syn-drome. Clin Obstet Gynaecol. 1985;12(3):605–632.
314. Dickey RP, Holtkamp DE. Development, pharma-cology and clinical experience with clomiphenecitrate. Hum Reprod Update. 1996;2(6):483–506.
315. Kero J, Poutanen M, Zhang F-P, Rahman N, McNicolAM, Nilson JH, Keri RA, Huhtaniemi IT. Elevatedluteinizing hormone induces expression of its re-ceptor and promotes steroidogenesis in the adrenalcortex. J Clin Invest. 2000;105(5):633–641.
316. McAllister JM, Legro RS, Modi BP, Strauss JF III.Functional genomics of PCOS: from GWAS tomolecular mechanisms. Trends Endocrinol Metab.2015;26(3):118–124.
317. Nelson-Degrave VL, Wickenheisser JK, Hendricks KL,Asano T, Fujishiro M, Legro RS, Kimball SR, Strauss JFIII, McAllister JM. Alterations in mitogen-activatedprotein kinase kinase and extracellular regulatedkinase signaling in theca cells contribute to ex-cessive androgen production in polycystic ovarysyndrome. Mol Endocrinol. 2005;19(2):379–390.
318. Batista MCP, Duarte E de F, Borba MD dos R, ZinglerE, Mangussi-Gomes J, dos Santos BTA, de MoraesOL, Hayashida SAY, Baracat EC, da Rocha Neves FA,Maciel GA, Bachega TA, Barra GB, Lofrano-Porto A.
589doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

Trp28Arg/Ile35Thr LHB gene variants are associatedwith elevated testosterone levels in women withpolycystic ovary syndrome. Gene. 2014;550(1):68–73.
319. Kurioka H, Takahashi K, Irikoma M, Okada M, OzakiT, Ueda T, Miyazaki K. Diagnostic difficulty inpolycystic ovary syndrome due to an LH-beta-subunit variant. Eur J Endocrinol. 1999;140(3):235–238.
320. Elter K, Erel CT, Cine N, Ozbek U, Hacihanefioglu B,Ertungealp E. Role of the mutations Trp8 =. Argand Ile15 =. Thr of the human luteinizing hor-mone beta-subunit in women with polycystic ovarysyndrome. Fertil Steril. 1999;71(3):425–430.
321. Lobo RA, Shoupe D, Chang SP, Campeau J. Thecontrol of bioactive luteinizing hormone secretionin women with polycystic ovary syndrome. Am JObstet Gynecol. 1984;148(4):423–428.
322. Ding YQ, Huhtaniemi I. Preponderance of basicisoforms of serum luteinizing hormone (LH) is as-sociated with the high bio/immune ratio of LH inhealthy women and in women with polycysticovarian disease. Hum Reprod. 1991;6(3):346–350.
323. Ropelato MG, Garcia-Rudaz MC, Castro-FernandezC, Ulloa-Aguirre A, Escobar ME, Barontini M,Veldhuis JD. A preponderance of basic luteinizinghormone (LH) isoforms accompanies inappropriatehypersecretion of both basal and pulsatile LH inadolescents with polycystic ovarian syndrome. J ClinEndocrinol Metab. 1999;84(12):4629–4636.
324. Hayes FJ, Taylor AE, Martin KA, Hall JE. Use ofa gonadotropin-releasing hormone antagonist asa physiologic probe in polycystic ovary syn-drome: assessment of neuroendocrine and an-drogen dynamics. J Clin Endocrinol Metab. 1998;83(7):2343–2349.
325. Balen AH. Ovulation induction in the managementof anovulatory polycystic ovary syndrome. Mol CellEndocrinol. 2013;373(1-2):77–82.
326. Engmann L, Siano L, Schmidt D, Nulsen J, Maier D,Benadiva C. GnRH agonist to induce oocyte mat-uration during IVF in patients at high risk of OHSS.Reprod Biomed Online. 2006;13(5):639–644.
327. Krishna D, Dhoble S, Praneesh G, Rathore S,Upadhaya A, Rao K. Gonadotropin-releasing hor-mone agonist trigger is a better alternative thanhuman chorionic gonadotropin in PCOS un-dergoing IVF cycles for an OHSS free clinic: a ran-domized control trial. J Hum Reprod Sci. 2016;9(3):164–172.
328. Cela V, Obino MER, Alberga Y, Pinelli S,Sergiampietri C, Casarosa E, Simi G, Papini F, ArtiniPG. Ovarian response to controlled ovarian stim-ulation in women with different polycystic ovarysyndrome phenotypes. Gynecol Endocrinol. 2018;34(6):518–523.
329. Karsli MF, Gultekin IB, Cakmak B, Yeral MI, SeckinKD, Alt Nboga O, Kucukozkan T. Do we needreadjustment of the biochemical parameters in firsttrimester combined aneuploidy screening test inwomen with polycystic ovary syndrome? PrenatDiagn. 2014;34(11):1073–1076.
330. Saxena R, Bjonnes AC, Georgopoulos NA, Koika V,Panidis D, Welt CK. Gene variants associated withage at menopause are also associated with poly-cystic ovary syndrome, gonadotrophins and ovarianvolume. Hum Reprod. 2015;30(7):1697–1703.
331. Corbett S, Morin-Papunen L. The polycystic ovarysyndrome and recent human evolution. Mol CellEndocrinol. 2013;373(1-2):39–50.
332. Kajii T, Ferrier A, Niikawa N, Takahara H, Ohama K,Avirachan S. Anatomic and chromosomal anom-alies in 639 spontaneous abortuses. Hum Genet.1980;55(1):87–98.
333. Abell SK, Nankervis A, Khan KS, Teede HJ. Type 1and type 2 diabetes preconception and in preg-nancy: health impacts, influence of obesity andlifestyle, and principles of management. SeminReprod Med. 2016;34(2):110–120.
334. Saner-Amigh K, Mayhew BA, Mantero F, Schiavi F,White PC, Rao CV, Rainey WE. Elevated expressionof luteinizing hormone receptor in aldosterone-producing adenomas. J Clin Endocrinol Metab.2006;91(3):1136–1142.
335. Melnick AP, Pereira N, Murphy EM, Rosenwaks Z,Spandorfer SD. How low is too low? Cycle day 28estradiol levels and pregnancy outcomes. Fertil Steril.2016;105(4):905–909.e1.
336. Saccone G, Schoen C, Franasiak JM, Scott RT Jr,Berghella V. Supplementation with progestogens inthe first trimester of pregnancy to prevent mis-carriage in women with unexplained recurrentmiscarriage: a systematic review and meta-analysisof randomized, controlled trials. Fertil Steril. 2017;107(2):430–438.e3.
337. Chen C-D, Chiang Y-T, Yang P-K, Chen M-J, ChangC-H, Yang Y-S, Chen S-U. Frequency of low serumLH is associated with increased early pregnancy lossin IVF/ICSI cycles. Reprod Biomed Online. 2016;33(4):449–457.
338. Lohstroh PN, Overstreet JW, Stewart DR, NakajimaST, Cragun JR, Boyers SP, Lasley BL. Secretion andexcretion of human chorionic gonadotropin duringearly pregnancy. Fertil Steril. 2005;83(4):1000–1011.
339. Iles RK, Delves PJ, Butler SA. Does hCG or hCGbplay a role in cancer cell biology?Mol Cell Endocrinol.2010;329(1-2):62–70.
340. Layman LC, Edwards JL, Osborne WE, Peak DB,Gallup DG, Tho SP, Reindollar RH, Roach DJ,McDonough PG, Lanclos KD. Human chorionicgonadotrophin-beta gene sequences in womenwith disorders of HCG production. Mol HumReprod. 1997;3(4):315–320.
341. Mangel L, Bıro K, Battyani I, Gocze P, Tornoczky T,Kalman E. A case study on the potential angiogeniceffect of human chorionic gonadotropin hormonein rapid progression and spontaneous regression ofmetastatic renal cell carcinoma during pregnancyand after surgical abortion. BMC Cancer. 2015;15(1):1013.
342. Głodek A, Jankowska A. CGB activates ERK andAKT kinases in cancer cells via LHCGR-independentmechanism. Tumour Biol. 2014;35(6):5467–5479.
343. Evans J. Hyperglycosylated hCG: a unique humanimplantation and invasion factor. Am J ReprodImmunol. 2016;75(3):333–340.
344. Lee C-L, Chiu PCN, Hautala L, Salo T, Yeung WSB,Stenman U-H, Koistinen H. Human chorionic go-nadotropin and its free b-subunit stimulate tro-phoblast invasion independent of LH/hCGreceptor. Mol Cell Endocrinol. 2013;375(1-2):43–52.
345. Gehring C, Siepmann T, Heidegger H, Jeschke U. Thecontroversial role of human chorionic gonadotro-pin in the development of breast cancer and othertypes of tumors. Breast. 2016;26:135–140.
346. Aharoni D, Dantes A, Oren M, Amsterdam A.cAMP-mediated signals as determinants for apo-ptosis in primary granulosa cells. Exp Cell Res. 1995;218(1):271–282.
347. Amsterdam A, Keren-Tal I, Aharoni D. Cross-talkbetween cAMP and p53-generated signals in in-duction of differentiation and apoptosis in ste-roidogenic granulosa cells. Steroids. 1996;61(4):252–256.
348. Kumar N, Gupta S, Dabral S, Singh S, Sehrawat S.Role of exchange protein directly activated bycAMP (EPAC1) in breast cancer cell migration andapoptosis.Mol Cell Biochem. 2017;430(1-2):115–125.
349. Lenhard M, Tsvilina A, Schumacher L, Kupka M,Ditsch N, Mayr D, Friese K, Jeschke U. Humanchorionic gonadotropin and its relation to grade,stage and patient survival in ovarian cancer. BMCCancer. 2012;12(1):2.
350. Henrıquez S, Kohen P, Xu X, Veenstra TD, Muñoz A,Palomino WA, Strauss JF III, Devoto L. Estrogenmetabolites in human corpus luteum physiology:differential effects on angiogenic activity. Fertil Steril.2016;106(1):230–237.e1.
351. Mohammad HP, Abbud RA, Parlow AF, Lewin JS,Nilson JH. Targeted overexpression of luteinizinghormone causes ovary-dependent functional ade-nomas restricted to cells of the Pit-1 lineage. En-docrinology. 2003;144(10):4626–4636.
352. Ratner LD, Gonzalez B, Ahtiainen P, Di Giorgio NP,Poutanen M, Calandra RS, Huhtaniemi IT, Rulli SB.Short-term pharmacological suppression of thehyperprolactinemia of infertile hCG-overproducingfemale mice persistently restores their fertility. En-docrinology. 2012;153(12):5980–5992.
353. Bernichtein S, Peltoketo H, Huhtaniemi I. Adrenalhyperplasia and tumours in mice in connectionwith aberrant pituitary-gonadal function. Mol CellEndocrinol. 2009;300(1-2):164–168.
354. Doroszko M, Chrusciel M, Belling K, Vuorenoja S,Dalgaard M, Leffers H, Nielsen HB, Huhtaniemi I,Toppari J, Rahman NA. Novel genes involved inpathophysiology of gonadotropin-dependent ad-renal tumors in mice. Mol Cell Endocrinol. 2017;444:9–18.
355. Bernichtein S, Alevizaki M, Huhtaniemi I. Is theadrenal cortex a target for gonadotropins? TrendsEndocrinol Metab. 2008;19(7):231–238.
356. de Lange WE, Pratt JJ, Doorenbos H. A gonado-trophin responsive testosterone producing adre-nocortical adenoma and high gonadotrophin levelsin an elderly woman. Clin Endocrinol (Oxf). 1980;12(1):21–28.
357. Teo AED, Garg S, Shaikh LH, Zhou J, Karet Frankl FE,Gurnell M, Happerfield L, Marker A, Bienz M, AzizanEAB, Brown MJ. Pregnancy, primary aldosteronism,and adrenal CTNNB1mutations. N Engl J Med. 2015;373(15):1429–1436.
358. Liu G, Duranteau L, Carel J-C, Monroe J, Doyle DA,Shenker A. Leydig-cell tumors caused by an acti-vating mutation of the gene encoding the lutei-nizing hormone receptor. N Engl J Med. 1999;341(23):1731–1736.
359. Newton CL, Anderson RC, Millar RP. Therapeuticneuroendocrine agonist and antagonist analogs ofhypothalamic neuropeptides as modulators of thehypothalamic-pituitary-gonadal axis. Endocr Dev.2016;30:106–129.
360. Lacroix A, Bourdeau I, Lampron A, Mazzuco TL,Tremblay J, Hamet P. Aberrant G-protein coupledreceptor expression in relation to adrenocorticaloverfunction. Clin Endocrinol (Oxf). 2010;73(1):1–15.
361. Baer G, Loumaye E. Comparison of recombinanthuman luteinising hormone (r-hLH) and humanmenopausal gonadotropin (hMG) in assisted re-productive technology. Curr Med Res Opin. 2003;19(2):83–88.
362. Riccetti L, Klett D, Ayoub MA, Boulo T, Pignatti E,Tagliavini S, Varani M, Trenti T, Nicoli A,Capodanno F, La Sala GB, Reiter E, Simoni M,Casarini L. Heterogeneous hCG and hMG com-mercial preparations result in different intracellularsignalling but induce a similar long-term pro-gesterone response in vitro. Mol Hum Reprod. 2017;23(10):685–697.
363. Stocco DM, Wang X, Jo Y, Manna PR. Multiplesignaling pathways regulating steroidogenesis andsteroidogenic acute regulatory protein expression:
590 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

more complicated than we thought. Mol Endo-crinol. 2005;19(11):2647–2659.
364. Evans HM, Simpson ME, Austin PR. The recognitionand comparison of prolan and prolan-like sub-stances. J Exp Med. 1933;58(5):561–568.
365. Johnsen SG, Hamburger C. Studies on urinary go-nadotrophins. IV. Qualitative difference betweenthe international reference preparation for humanmenopausal gonadotrophin and other extracts ofurine from postmenopausal women. Acta Endo-crinol (Copenh). 1959;32:497–508.
366. Giudice E, Crisci C, Eshkol A, Papoian R. Compo-sition of commercial gonadotrophin preparationsextracted from human post-menopausal urine:characterization of non-gonadotrophin proteins.Hum Reprod. 1994;9(12):2291–2299.
367. Wolfenson C, Groisman J, Couto AS, Hedenfalk M,Cortvrindt RG, Smitz JE, Jespersen S. Batch-to-batchconsistency of human-derived gonadotrophinpreparations compared with recombinant prepa-rations. Reprod Biomed Online. 2005;10(4):442–454.
368. van de Weijer BHM, Mulders JWM, Bos ES, VerhaertPDEM, van den Hooven HW. Compositional ana-lyses of a human menopausal gonadotrophinpreparation extracted from urine (menotropin).Identification of some of its major impurities.Reprod Biomed Online. 2003;7(5):547–557.
369. Zegers-Hochschild F, Fernandez E, Mackenna A,Fabres C, Altieri E, Lopez T. The empty folliclesyndrome: a pharmaceutical industry syndrome.Hum Reprod. 1995;10(9):2262–2265.
370. Casarini L, Brigante G, Simoni M, Santi D. Clinicalapplications of gonadotropins in the female:assisted reproduction and beyond. Prog Mol BiolTransl Sci. 2016;143:85–119.
371. Perlman S, van den Hazel B, Christiansen J, Gram-Nielsen S, Jeppesen CB, Andersen KV, Halkier T,Okkels S, Schambye HT. Glycosylation of anN-terminal extension prolongs the half-life andincreases the in vivo activity of follicle stimulatinghormone. J Clin Endocrinol Metab. 2003;88(7):3227–3235.
372. Fares FA, Suganuma N, Nishimori K, LaPolt PS,Hsueh AJ, Boime I. Design of a long-acting follitropinagonist by fusing the C-terminal sequence of thechorionic gonadotropin beta subunit to the folli-tropin beta subunit. Proc Natl Acad Sci USA. 1992;89(10):4304–4308.
373. Klein J, Lobel L, Pollak S, Lustbader B, Ogden RT,Sauer MV, Lustbader JW. Development and char-acterization of a long-acting recombinant hFSHagonist. Hum Reprod. 2003;18(1):50–56.
374. Devroey P, Fauser BC, Platteau P, Beckers NG, DhontM, Mannaerts BM. Induction of multiple folliculardevelopment by a single dose of long-actingrecombinant follicle-stimulating hormone (FSH-CTP, corifollitropin alfa) for controlled ovarianstimulation before in vitro fertilization. J ClinEndocrinol Metab. 2004;89(5):2062–2070.
375. Edwards RG, Steptoe PC. Induction of folliculargrowth, ovulation and luteinization in the humanovary. J Reprod Fertil Suppl. 1975;(22):121–163.
376. Cole LA. Biological functions of hCG and hCG-related molecules. Reprod Biol Endocrinol. 2010;8(1):102.
377. Wide L, Bakos O. More basic forms of both humanfollicle-stimulating hormone and luteinizing hor-mone in serum at midcycle compared with thefollicular or luteal phase. J Clin Endocrinol Metab.1993;76(4):885–889.
378. Al-Inany HG, Abou-Setta AM, Aboulghar MA,Mansour RT, Serour GI. HMG versus rFSH forovulation induction in developing countries: a cost-effectiveness analysis based on the results of a
recent meta-analysis. Reprod Biomed Online. 2006;12(2):163–169.
379. Nakano R, Mizuno T, Kotsuji F, Katayama K, WshioM,Tojo S. “Triggering” of ovulation after infusion ofsynthetic luteinizing hormone releasing factor (LRF).Acta Obstet Gynecol Scand. 1973;52(3):269–272.
380. Diedrich K, Diedrich C, Santos E, Zoll C, al-Hasani S,Reissmann T, Krebs D, Klingmuller D. Suppression ofthe endogenous luteinizing hormone surge by thegonadotrophin-releasing hormone antagonistcetrorelix during ovarian stimulation. Hum Reprod.1994;9(5):788–791.
381. Albano C, Smitz J, Camus M, Riethmuller-Winzen H,Van Steirteghem A, Devroey P. Comparison ofdifferent doses of gonadotropin-releasing hormoneantagonist cetrorelix during controlled ovarianhyperstimulation. Fertil Steril. 1997;67(5):917–922.
382. Itskovitz J, Boldes R, Levron J, Erlik Y, Kahana L,Brandes JM. Induction of preovulatory luteinizinghormone surge and prevention of ovarian hyper-stimulation syndrome by gonadotropin-releasinghormone agonist. Fertil Steril. 1991;56(2):213–220.
383. Hoff JD, Quigley ME, Yen SS. Hormonal dynamics atmidcycle: a reevaluation. J Clin Endocrinol Metab.1983;57(4):792–796.
384. Haahr T, Roque M, Esteves SC, Humaidan P. GnRHagonist trigger and LH activity luteal phase supportversus hCG trigger and conventional luteal phasesupport in fresh embryo transfer IVF/ICSI cycles—asystematic PRISMA review and meta-analysis. FrontEndocrinol (Lausanne). 2017;8:116.
385. Castillo JC, Humaidan P, Bernabeu R. Pharmaceu-tical options for triggering of final oocyte matu-ration in ART. Biomed Res Int. 2014;2014:580171.
386. Bodri D, Guillen JJ, Galindo A, Mataro D, Pujol A,Coll O. Triggering with human chorionic gonado-tropin or a gonadotropin-releasing hormone agonistin gonadotropin-releasing hormone antagonist-treated oocyte donor cycles: findings of a largeretrospective cohort study. Fertil Steril. 2009;91(2):365–371.
387. Acevedo B, Gomez-Palomares JL, Ricciarelli E,Hernandez ER. Triggering ovulationwith gonadotropin-releasing hormone agonists does not compromiseembryo implantation rates. Fertil Steril. 2006;86(6):1682–1687.
388. Melo M, Busso CE, Bellver J, Alama P, Garrido N,Meseguer M, Pellicer A, Remohı J. GnRH agonistversus recombinant HCG in an oocyte donationprogramme: a randomized, prospective, controlled,assessor-blind study. Reprod Biomed Online. 2009;19(4):486–492.
389. Benadiva C, Engmann L. Intensive luteal phasesupport after GnRH agonist trigger: it does help.Reprod Biomed Online. 2012;25(3):329–330.
390. Humaidan P, Polyzos NP, Alsbjerg B, Erb K,Mikkelsen AL, Elbaek HO, Papanikolaou EG,Andersen CY. GnRHa trigger and individualizedluteal phase hCG support according to ovarianresponse to stimulation: two prospective ran-domized controlled multi-centre studies in IVFpatients. Hum Reprod. 2013;28(9):2511–2521.
391. Castillo JC, Dolz M, Bienvenido E, Abad L, Casañ EM,Bonilla-Musoles F. Cycles triggered with GnRHagonist: exploring low-dose HCG for luteal support.Reprod Biomed Online. 2010;20(2):175–181.
392. Papanikolaou EG, Verpoest W, Fatemi H, Tarlatzis B,Devroey P, Tournaye H. A novel method of lutealsupplementation with recombinant luteinizinghormone when a gonadotropin-releasing hormoneagonist is used instead of human chorionic go-nadotropin for ovulation triggering: a randomizedprospective proof of concept study. Fertil Steril.2011;95(3):1174–1177.
393. Santi D, Corona G. Primary and secondary hypo-gonadism. In: Simoni M, Huhtaniemi IT, eds. En-docrinology of the Testis and Male Reproduction.Endocrinology. Cham: Springer; 2017:687–747.
394. Barrio R, de Luis D, Alonso M, Lamas A, Moreno JC.Induction of puberty with human chorionic go-nadotropin and follicle-stimulating hormone inadolescent males with hypogonadotropic hypo-gonadism. Fertil Steril. 1999;71(2):244–248.
395. Kobori Y, Suzuki K, Iwahata T, Shin T, Sato R, NishioK, Yagi H, Arai G, Soh S, Okada H. Hormonal therapy(hCG and rhFSH) for infertile men with adult-onsetidiopathic hypogonadotropic hypogonadism. SystBiol Reprod Med. 2015;61(2):110–112.
396. Veldhuis JD, Liu PY, Takahashi PY, Weist SM,Wigham JR. Analysis of the impact of intravenousLH pulses versus continuous LH infusion on tes-tosterone secretion during GnRH-receptor block-ade. Am J Physiol Regul Integr Comp Physiol. 2012;303(10):R994–R1002.
397. Mochtar MH, Van der Veen F, Ziech M, van WelyM, Musters A. Recombinant luteinizing hormone(rLH) for controlled ovarian hyperstimulation inassisted reproductive cycles. Cochrane DatabaseSyst Rev. 2007;(2):CD005070.
398. Mochtar MH, Danhof NA, Ayeleke RO, Van derVeen F, van Wely M. Recombinant luteinizinghormone (rLH) and recombinant follicle stimulat-ing hormone (rFSH) for ovarian stimulation in IVF/ICSI cycles. Cochrane Database Syst Rev. 2017;5:CD005070.
399. Legro RS, Kunselman AR. A good meta-analysis ishard to find. Fertil Steril. 2012;97(5):1048–1049.
400. Konig TE, van der Houwen LEE, Lambalk CB.Recombinant LH supplementation in women of 35years and older undergoing IVF? Fertil Steril. 2012;98(1):e10–e12.
401. Smith V, Osianlis T, Vollenhoven B. Prevention ofovarian hyperstimulation syndrome: a review.Obstet Gynecol Int. 2015;2015:1–10.
402. Nastri CO, Ferriani RA, Rocha IA, Martins WP.Ovarian hyperstimulation syndrome: pathophysi-ology and prevention. J Assist Reprod Genet. 2010;27(2-3):121–128.
403. Youssef MA, Van der Veen F, Al-Inany HG, MochtarMH, Griesinger G, Nagi Mohesen M, AboulfoutouhI, van Wely M. Gonadotropin-releasing hormoneagonist versus HCG for oocyte triggering in antagonist-assisted reproductive technology. Cochrane Data-base Syst Rev. 2014;(10):CD008046.
404. Ozgur K, Humaidan P, Coetzee K. SegmentedART—thenew era in ART? Reprod Biol. 2016;16(2):91–103.
405. La Marca A, Sunkara SK. Individualization of con-trolled ovarian stimulation in IVF using ovarianreserve markers: from theory to practice. HumReprod Update. 2014;20(1):124–140.
406. Wood HM, Elder JS. Cryptorchidism and testicularcancer: separating fact from fiction. J Urol. 2009;181(2):452–461.
407. Dunkel L, Taskinen S, Hovatta O, Tilly JL, WikstromS. Germ cell apoptosis after treatment of cryptor-chidism with human chorionic gonadotropin isassociated with impaired reproductive function inthe adult. J Clin Invest. 1997;100(9):2341–2346.
408. Colicchia M, Campagnolo L, Baldini E, Ulisse S,Valensise H, Moretti C. Molecular basis of thyro-tropin and thyroid hormone action during im-plantation and early development. Hum ReprodUpdate. 2014;20(6):884–904.
409. Duarte-Guterman P, Navarro-Martın L, Trudeau VL.Mechanisms of crosstalk between endocrine sys-tems: regulation of sex steroid hormone synthesisand action by thyroid hormones. Gen CompEndocrinol. 2014;203:69–85.
591doi: 10.1210/er.2018-00065 https://academic.oup.com/edrv
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022

410. Cornell JS, Pierce JG. The subunits of human pi-tuitary thyroid-stimulating hormone: isolation,properties, and composition. J Biol Chem. 1973;248(12):4327–4333.
411. Nagayama Y, Russo D, Chazenbalk GD, WadsworthHL, Rapoport B. Extracellular domain chimeras ofthe TSH and LH/CG receptors reveal the mid-region (amino acids 171-260) to play a vital rolein high affinity TSH binding. Biochem Biophys ResCommun. 1990;173(3):1150–1156.
412. Tomer Y, Huber GK, Davies TF. Human chorionicgonadotropin (hCG) interacts directly withrecombinant human TSH receptors. J Clin Endo-crinol Metab. 1992;74(6):1477–1479.
413. Schubert RL, Narayan P, Puett D. Specificity ofcognate ligand-receptor interactions: fusion pro-teins of human chorionic gonadotropin and theheptahelical receptors for human luteinizing hor-mone, thyroid-stimulating hormone, and follicle-stimulating hormone. Endocrinology. 2003;144(1):129–137.
414. Kraiem Z, Sadeh O, Blithe DL, Nisula BC. Humanchorionic gonadotropin stimulates thyroid hor-mone secretion, iodide uptake, organification, andadenosine 39,59-monophosphate formation incultured human thyrocytes. J Clin Endocrinol Metab.1994;79(2):595–599.
415. Yoshimura M, Hershman JM, Pang XP, Berg L,Pekary AE. Activation of the thyrotropin (TSH)receptor by human chorionic gonadotropin andluteinizing hormone in Chinese hamster ovary cellsexpressing functional human TSH receptors. J ClinEndocrinol Metab. 1993;77(4):1009–1013.
416. Carayon P, Lefort G, Nisula B. Interaction of humanchorionic gonadotropin and human luteinizinghormone with human thyroid membranes. Endo-crinology. 1980;106(6):1907–1916.
417. Yoshimura M, Pekary AE, Pang XP, Berg L, Cole LA,Kardana A, Hershman JM. Effect of peptide nickingin the human chorionic gonadotropin beta-subuniton stimulation of recombinant human thyroid-stimulating hormone receptors. Eur J Endocrinol.1994;130(1):92–96.
418. Harada A, Hershman JM, Reed AW, Braunstein GD,Dignam WJ, Derzko C, Friedman S, Jewelewicz R,Pekary AE. Comparison of thyroid stimulators andthyroid hormone concentrations in the sera ofpregnant women. J Clin Endocrinol Metab. 1979;48(5):793–797.
419. Grun JP, Meuris S, De Nayer P, Glinoer D. The thy-rotrophic role of human chorionic gonadotrophin(hCG) in the early stages of twin (versus single)pregnancies. Clin Endocrinol (Oxf). 1997;46(6):719–725.
420. Ross HA, Exalto N, Kloppenborg PW, Benraad TJ.Thyroid hormone binding in early pregnancy andthe risk of spontaneous abortion. Eur J ObstetGynecol Reprod Biol. 1989;32(2):129–136.
421. Aghajanova L, Stavreus-Evers A, Lindeberg M,Landgren B-M, Sparre LS, Hovatta O. Thyroid-stimulating hormone receptor and thyroid hor-mone receptors are involved in human endometrialphysiology. Fertil Steril. 2011;95(1):230–237, 237.e1–237.e2.
422. Benvenga S, Di Bari F, Granese R, Antonelli A. Serumthyrotropin and phase of the menstrual cycle. FrontEndocrinol (Lausanne). 2017;8:250.
423. Weber G, Vigone MC, Stroppa L, Chiumello G.Thyroid function and puberty. J Pediatr EndocrinolMetab. 2003;16(Suppl 2):253–257.
424. Koutras DA. Disturbances of menstruation inthyroid disease. Ann N Y Acad Sci. 1997;816(1Adolescent Gy):280–284.
425. Nakabayashi K, Matsumi H, Bhalla A, Bae J,Mosselman S, Hsu SY, Hsueh AJW. Thyrostimulin, aheterodimer of two new human glycoprotein hor-mone subunits, activates the thyroid-stimulatinghormone receptor. J Clin Invest. 2002;109(11):1445–1452.
426. Sun S-C, Hsu P-J, Wu F-J, Li S-H, Lu C-H, Luo C-W.Thyrostimulin, but not thyroid-stimulating hor-mone (TSH), acts as a paracrine regulator to acti-vate the TSH receptor in mammalian ovary. J BiolChem. 2010;285(6):3758–3765.
427. Huang W-L, Li Z, Lin T-Y, Wang S-W, Wu F-J, Luo C-W. Thyrostimulin-TSHR signaling promotes theproliferation of NIH:OVCAR-3 ovarian cancer cellsvia trans-regulation of the EGFR pathway. Sci Rep.2016;6(1):27471.
428. Val P, Lefrançois-Martinez A-M, Veyssiere G,Martinez A. SF-1 a key player in the developmentand differentiation of steroidogenic tissues. NuclRecept. 2003;1(1):8.
429. Lasley BL, Crawford SL, McConnell DS. Ovarianadrenal interactions during the menopausal tran-sition. Minerva Ginecol. 2013;65(6):641–651.
430. Miller WL, Auchus RJ. The molecular biology, bio-chemistry, and physiology of human steroidogen-esis and its disorders. Endocr Rev. 2011;32(1):81–151.
431. Rao CV. Human adrenal LH/hCG receptors andwhat they could mean for adrenal physiology andpathology.Mol Cell Endocrinol. 2010;329(1-2):33–36.
432. Vuorenoja S, Rivero-Muller A, Kiiveri S, Bielinska M,Heikinheimo M, Wilson DB, Huhtaniemi IT, RahmanNA. Adrenocortical tumorigenesis, luteinizing hor-mone receptor and transcription factors GATA-4 andGATA-6. Mol Cell Endocrinol. 2007;269(1-2):38–45.
433. Mazzuco TL, Chabre O, Feige J-J, Thomas M. Ab-errant expression of human luteinizing hormonereceptor by adrenocortical cells is sufficient toprovoke both hyperplasia and Cushing’s syndromefeatures. J Clin Endocrinol Metab. 2006;91(1):196–203.
434. Piltonen T, Koivunen R, Morin-Papunen L,Ruokonen A, Huhtaniemi IT, Tapanainen JS.Ovarian and adrenal steroid production: regulatoryrole of LH/HCG. Hum Reprod. 2002;17(3):620–624.
AcknowledgmentsWe thank Eric A. Zizzi for language editing.Financial Support: This work was supported by a grant
to the Department of Biomedical, Metabolic, and NeuralSciences, University of Modena and Reggio Emilia (Italy), fromthe Italian Ministry of University and Research in the De-partments of Excellence Programme.Correspondence and Reprint Requests: Manuela
Simoni, MD, PhD, Unit of Endocrinology, Department ofBiomedical, Metabolic, and Neural Sciences, University ofModena and Reggio Emilia, via P. Giardini 1355, Modena41126, Italy. E-mail: [email protected] Summary: The authors have nothing to
disclose.
AbbreviationsART, assisted reproduction technology; Ca2+, calcium ion;CG, chorionic gonadotropin; CGA, common glycoprotein a;CHO, Chinese hamster ovarian; COS, controlled ovarianstimulation; CREB, cAMP response element–binding protein;CTP, C-terminal extension; eCG, equine chorionic gonado-tropin; EGF, epidermal growth factor; eLH, equid pituitary LH;ERK1/2, extracellular-regulated kinase 1/2; FSHR, FSH re-ceptor; GPCR, G protein–coupled receptor; hCG, humanchorionic gonadotropin; hCG-H, hyperglycosylated humanchorionic gonadotropin molecules; HH, hypogonadotropichypogonadism; hMG, human menopausal gonadotropin;HPG, hypothalamic-pituitary-gonadal; IP3, inositol tris-phosphate; K1, association rate constant; Ka, equilibriumassociation constant; LHCGR, LH chorionic gonadotropinreceptor; LRR, leucine-rich repeat; OHSS, ovarian hyper-stimulation syndrome; pCREB, phosphorylated cAMP re-sponse element–binding protein; PCOS, polycystic ovarysyndrome; pERK1/2, phosphorylated extracellular-regulatedkinase 1/2; PKA, protein kinase A; PLC, phospholipase C; RCT,randomized clinical trial; r-hCG, chorionic gonadotropin a; r-LH, lutropin a; SNP, single nucleotide polymorphism;sTyr331, sulfated tyrosine located at position 331; TSHR, TSHreceptor; u-hCG, urine human chorionic gonadotropin; V-LH,LHB gene variant.
592 Casarini et al Differences Between LH and hCG Endocrine Reviews, October 2018, 39(5):549–592
REVIEWD
ownloaded from
https://academic.oup.com
/edrv/article/39/5/549/5036715 by guest on 09 August 2022
Copyright © 2022 FDOKUMEN




















