Cellules souches, hormones sexuelles et régénération thymique
Upload
khangminh22Category
view
4download
0
Citation: Traversari, S.; Cacini, S.;
Nesi, B. Seaweed Extracts as
Substitutes of Synthetic Hormones
for Rooting Promotion in Rose
Cuttings. Horticulturae 2022, 8, 561.
https://doi.org/10.3390/
horticulturae8070561
Academic Editors: Jean Carlos
Cardoso, Wagner Vendrame and
Chao Ma
Received: 9 May 2022
Accepted: 17 June 2022
Published: 21 June 2022
Publisher’s Note: MDPI stays neutral
with regard to jurisdictional claims in
published maps and institutional affil-
iations.
Copyright: © 2022 by the authors.
Licensee MDPI, Basel, Switzerland.
This article is an open access article
distributed under the terms and
conditions of the Creative Commons
Attribution (CC BY) license (https://
creativecommons.org/licenses/by/
4.0/).
horticulturae
Article
Seaweed Extracts as Substitutes of Synthetic Hormones forRooting Promotion in Rose CuttingsSilvia Traversari 1,2 , Sonia Cacini 2,* and Beatrice Nesi 2
1 Research Institute on Terrestrial Ecosystems (IRET), National Research Council (CNR), Via Moruzzi 1,56124 Pisa, Italy; [email protected]
2 CREA Research Centre for Vegetable and Ornamental Crops, Council for Agricultural Research andEconomics, Via dei Fiori 8, 51012 Pescia, Italy; [email protected]
* Correspondence: [email protected]
Abstract: In the horticultural sector, the achievement of an efficient and eco-friendly sustainableproduction of plants is nowadays challenging. Indeed, in plant vegetative propagation of woodyornamentals, the substitution of chemical products used to promote rooting of cuttings with naturalextracts would be a desirable goal. Thus, the aim of this work was to test the replacement of syntheticphytoregulators, such as auxins and brassinosteroids, with biostimulants, such as seaweed extracts,for the rooting promotion of rose cuttings. The rooting rate and biometric parameters of controlcuttings treated with distilled water were compared with those of cuttings treated with synthetichormones, i.e., auxins or 22(S),23(S)-homobrassinolide, or two commercial products based on lowtemperature seaweed extracts, i.e., Kelpak® and Phylgreen. Two scented hybrid tea rose cultivarswere used to assess possible genotype-dependent effects, i.e., ‘Michelangelo®’ and ‘Cosmos®’. Auxinsconfirmed their role in root growth enhancement in ornamental plant cuttings. Like these phytoreg-ulators, Kelpak® improved the survival rate and root biometric parameters of both rose cuttings,highlighting its suitability for the replacement of synthetic products used for rooting promotion inrose propagation. Brassinosteroids showed a species-dependent effect, increasing the root biomassin ‘Cosmos®’ while it resulted as distilled water in ‘Michelangelo®’. Phylgreen did not improve therooting of both rose cuttings, highlighting the necessity of evaluating the applicability and methodol-ogy for this product before its use. In conclusions, our results highlighted the possibility to replacechemical products in rose cutting production.
Keywords: auxins; homobrassinolide; Kelpak®; ornamental plant propagation; Phylgreen
1. Introduction
A key challenge in the production of ornamental plants is the combination of sus-tainable practices and cultivation and propagation efficiency [1]. Plant propagation in thefloricultural sector relies on different techniques, among which the use of stem cuttings isone of the most used for woody plant species, such as roses, also for its timesaving andcost-efficiency [2,3]. This method allows the maintenance of desirable characteristics insuperior rose cultivars, particularly considering their high degree of heterozygosity andpolyploidy, but can be affected by the deep genotypic differences in rooting ability withinthis genus [4]. Indeed, in stem cutting propagation, adventitious root formation is a keystep to be successful and it is promoted by producers in nurseries with the applicationof phytoregulators [5,6]. The most widely spread commercial products consist of aux-ins such as indole-3-butyric acid (IBA) and 1-naphthaleneacetic acid (NAA) [7]. Otherphytoregulators have been shown to have a role in rooting promotion [8], but they arepoorly investigated for commercial applications. Among them, brassinosteroids (BRs) areplant-specific steroid hormones that have a role in both growth promotion, including rootformation, and abiotic stress tolerance [9]. Indeed, BRs increased the degree of rooting and
Horticulturae 2022, 8, 561. https://doi.org/10.3390/horticulturae8070561 https://www.mdpi.com/journal/horticulturae

Horticulturae 2022, 8, 561 2 of 9
root length as well as the total soluble sugars in stem cuttings of two cultivars of barberryornamental plants [10]. However, the small amount and fast metabolization of BRs in planttissues make the production costs of natural compounds too high, and therefore structuraland functional analogues are commonly used, such as the 24-epibrassinolide [11].
In modern agriculture, the substitution of chemical inputs, such as phytoregulators,with alternative natural eco-friendly products presents a key challenge [12] also to meet thetransition towards agroecological production systems and organic farming principles [13].In this context, seaweed extracts constitute a promising alternative solution to the useof synthetic products in the promotion of plant yield and health [14]. Different seaweedspecies have shown their activity as biostimulants, probably related to the presence ofbioactive ingredients such as phytohormones as well as carbohydrates, proteins, and min-eral elements [15,16]. Moreover, seaweed extracts have been shown to be not only naturallyenriched in phytohormones but also able to promote the endogenous biosynthesis of aux-ins, cytokinin, and gibberellins [17]. Beyond their effect as growth promoters, seaweedextracts have also disease suppressing effects [18], potentially reducing the application ofphytochemicals as well. However, the molecular mechanisms behind their function arestill little known. Moreover, new sustainable extraction procedures have been developedto avoid the use of chemicals and preserve the seaweed’s original quality, increasing theirenvironmentally friendly aspects [18]. Despite these promising features, to our knowl-edge, only a few seaweed extracts have been tested for rooting promotion in rose cuttingpropagation, prevalently containing the Ascophyllum nodosum extract [6].
Thus, the aim of this work was to test the replacement of synthetic phytoregulatorswith two seaweed extracts, i.e., Kelpak® and Phylgreen, for promoting the rooting of rosecuttings. The two seaweed products were evaluated in comparison with the absence oftreatment (only distilled water), or two different synthetic phytoregulators, i.e., auxins orBRs. The extracts were tested on two rose cultivars to assess possible differences related togenotype-dependent responses.
2. Materials and Methods2.1. Plant Material and Treatments
The newly produced young herbaceous stems were sampled at the end of March 2021,at the beginning of the growing season, from one-year old plants of two Rosa spp. cultivars,‘Michelangelo®’ and ‘Cosmos®’, both scented hybrid tea Meilland (Meilland International,Le Luc-en-Provence, France), maintained within a greenhouse and pruned on 11 February2021. Rose plants were fertigated with a nutrient solution containing 1.5 g L−1 of Peters®
Excel CalMag Finisher (13-5-20 + 7CaO + MgO + TE, ICL Specialty Fertilizers, Tel Aviv,Israel), maintaining the pH at 5.5–6.0. Single node stem cuttings (3.5–4.5 cm) two weeksold were collected, leaving a half leaf portion to assure transpiration on 29 March. Fivedifferent treatments were applied to both cultivar cuttings (90 cuttings for each treatmentand each cultivar for a total of 900 cuttings): (1) distilled water (H2O); (2) 4000 ppm ofindole-3-butyric acid and 1-naphthaleneacetic acid (Merck KGaA, Darmstadt, German) indistilled water (AUX); (3) 5 ppm of 22(S),23(S)-homobrassinolide (Merck KGaA, Darmstadt,German) in distilled water (BRA); (4) 10% Kelpak® (Agricola Internazionale S.R.L., Pisa,Italy) in distilled water (KEL); (5) 10% Phylgreen (Tradecorp Italia & South East Europe,Bologna, Italy) in distilled water (PHY). Kelpak® contains an Ecklonia maxima extractobtained through a low temperature mechanical extraction avoiding the use of chemicalsand without altering the original product. It has auxins and cytokinins in an optimalbalance for rooting promotion. Phylgreen is made from A. nodosum and, as the Kelpak®,it is also obtained through a low temperature mechanical extraction without chemicals,preserving the product features.
Treatments were applied through the immersion of basal cutting portions (1 cm) for5 s (H2O, AUX, BRA) or 1 min (KEL and PHY) within each specific solution. After thetreatments, cuttings were immediately placed in holes filled with the substrate (1:1 v/vof peat and perlite) and irrigated. Cuttings were sprayed with water 5 times per day for

Horticulturae 2022, 8, 561 3 of 9
3 min for 3 weeks, while after this period they were sprayed 3 times per day for 3 min.Average temperature, humidity, and lighting during the experimental period were 21 ◦C,70%, and 322 Watt m−2. Data were acquired by an EnviroMonitor weather station (DavisInstruments, Hayward, CA, USA).
2.2. Biometric Measures
After 14 weeks from the transplant (30 June), the number of cuttings was evaluated toestablish the rooting rate percentage for each treatment. Nine cuttings were sampled andused as biological replicates for the biometric measures. In particular, the shoot number,shoot length, and root fresh weight (FW) were evaluated. Root images were acquired, androot length and area were measured through Fiji software. Roots were then dried at 75 ◦Cfor 2 days and the dry weight was measured (DW).
2.3. Carbohydrates Analysis
Since a higher soluble sugar concentration has been reported to match with betterrooting performance in rose cuttings [19], carbohydrates were also quantified in stemsduring the rooting period.
For each treatment, 12 cuttings of each cultivar were sampled at the beginning ofthe trial and 4 and 16 days after the treatments. The bark with the emerging callus wascollected, frozen in liquid nitrogen, and stored at −80 ◦C for carbohydrate analysis. Thebark was reduced in powder with liquid nitrogen and used for the determination of solublesugars through the Anthrone colorimetric method (Merck KGaA, Darmstadt, Germany),reading the absorbance at 620 nm against a standard curve of glucose.
2.4. Statistics
Data were tested for normal distribution using the Shapiro–Wilk normality test andthen analyzed by a two-way ANOVA (treatment and cultivar as variables) to assess differ-ences in treatment between the cultivars and then with a one-way ANOVA, followed by aTukey’s post-hoc test, to highlight significant differences between treatments (p ≤ 0.05, 0.01,and 0.001). The statistical analyses and graphs were performed with Prism 9 (GraphPadSoftware, Inc., La Jolla, CA, USA).
3. Results3.1. Survival Percentage and Biometric Measures
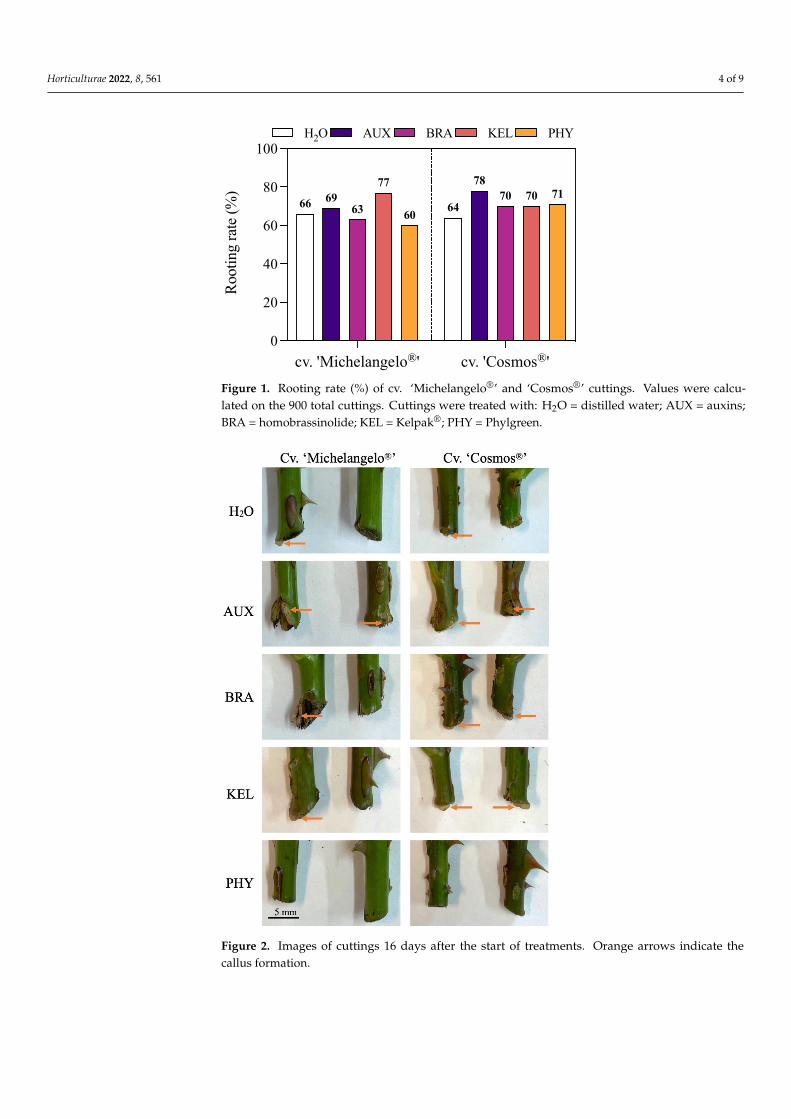
Control cuttings of both cultivars treated with distilled H2O showed a rooting percent-age on average of 65% (Figure 1). Cuttings treated with BRA and PHY had a slightly lowerrooting rate than control cuttings in cv. ‘Michelangelo®’. The highest rooting percentagein this cultivar was measured under KEL treatment (77%, + 19% in comparison to controlcuttings). On the contrary, all treatments increased the rooting percentage in cv. ‘Cosmos®’,particularly the cuttings treated with AUX, which had a survival rate 22% higher than thecontrol ones.
After 16 days from the treatments, callus formation was mostly present in the lowestpart of cuttings treated with AUX, BRA, and KEL (Figure 2). Callus was absent fromcuttings treated with PHY, while it was a small portion in those treated with H2O.
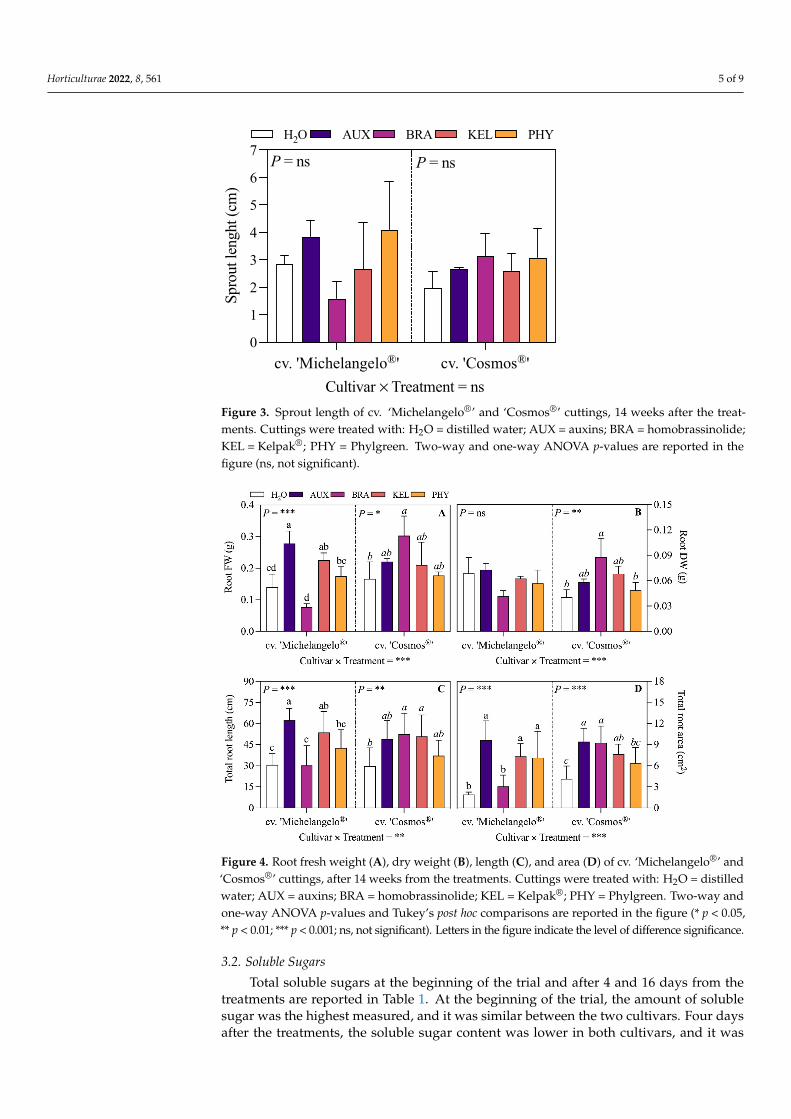
Every cutting produced one single sprout and sprout length was not significantlydifferent between treatments in both cultivars (Figure 3).
Treatments had a different effect on the root biometric parameters of the two rosecultivars (Figure 4). The root FW (Figure 4A) was higher under AUX and KEL treatments incv. ‘Michelangelo®’ (+99 and 62%, respectively) and under BRA treatment in cv. ‘Cosmos®’(+82%). The same trend was shown in root DW (Figure 4B) by cv. ‘Cosmos®’. Root length(Figure 4C) was higher under AUX and KEL treatments in cv. ‘Michelangelo®’ (+103 and75%, respectively) and under BRA and PHY treatments in cv. ‘Cosmos®’ (+76 and 71%,respectively). Root area (Figure 4D) was higher under AUX, KEL, and PHY treatments incv. ‘Michelangelo®’ and under AUX, BRA, and PHY treatments in cv. ‘Cosmos®’.
Horticulturae 2022, 8, 561 4 of 9
Horticulturae 2022, 8, x FOR PEER REVIEW 4 of 9
‘Cosmos®’ (+ 82%). The same trend was shown in root DW (Figure 4B) by cv. ‘Cosmos®’. Root length (Figure 4C) was higher under AUX and KEL treatments in cv. ‘Michelangelo®’ (+ 103 and 75%, respectively) and under BRA and PHY treatments in cv. ‘Cosmos®’ (+ 76 and 71%, respectively). Root area (Figure 4D) was higher under AUX, KEL, and PHY treatments in cv. ‘Michelangelo®’ and under AUX, BRA, and PHY treatments in cv. ‘Cos-mos®’.
Figure 1. Rooting rate (%) of cv. ‘Michelangelo®’ and ‘Cosmos®’ cuttings. Values were calculated on the 900 total cuttings. Cuttings were treated with: H2O = distilled water; AUX = auxins; BRA = ho-mobrassinolide; KEL = Kelpak®; PHY = Phylgreen.
cv. 'Michelangelo®' cv. 'Cosmos®'0
20
40
60
80
100
66 6469
78
6370
7770
60
71
Root
ing
rate
(%)
H2O AUX BRA KEL PHY
Figure 1. Rooting rate (%) of cv. ‘Michelangelo®’ and ‘Cosmos®’ cuttings. Values were calcu-lated on the 900 total cuttings. Cuttings were treated with: H2O = distilled water; AUX = auxins;BRA = homobrassinolide; KEL = Kelpak®; PHY = Phylgreen.
Horticulturae 2022, 8, x FOR PEER REVIEW 4 of 9
‘Cosmos®’ (+ 82%). The same trend was shown in root DW (Figure 4B) by cv. ‘Cosmos®’. Root length (Figure 4C) was higher under AUX and KEL treatments in cv. ‘Michelangelo®’ (+ 103 and 75%, respectively) and under BRA and PHY treatments in cv. ‘Cosmos®’ (+ 76 and 71%, respectively). Root area (Figure 4D) was higher under AUX, KEL, and PHY treatments in cv. ‘Michelangelo®’ and under AUX, BRA, and PHY treatments in cv. ‘Cos-mos®’.
Figure 1. Rooting rate (%) of cv. ‘Michelangelo®’ and ‘Cosmos®’ cuttings. Values were calculated on the 900 total cuttings. Cuttings were treated with: H2O = distilled water; AUX = auxins; BRA = ho-mobrassinolide; KEL = Kelpak®; PHY = Phylgreen.
cv. 'Michelangelo®' cv. 'Cosmos®'0
20
40
60
80
100
66 6469
78
6370
7770
60
71Ro
otin
g ra
te (%
)
H2O AUX BRA KEL PHY
Figure 2. Images of cuttings 16 days after the start of treatments. Orange arrows indicate thecallus formation.
Horticulturae 2022, 8, 561 5 of 9
Horticulturae 2022, 8, x FOR PEER REVIEW 5 of 9
Figure 2. Images of cuttings 16 days after the start of treatments. Orange arrows indicate the callus formation.
Figure 3. Sprout length of cv. ‘Michelangelo®’ and ‘Cosmos®’ cuttings, 14 weeks after the treatments. Cuttings were treated with: H2O = distilled water; AUX = auxins; BRA = homobrassinolide; KEL = Kelpak®; PHY = Phylgreen. Two-way and one-way ANOVA p-values are reported in the figure (ns, not significant).
Figure 4. Root fresh weight (A), dry weight (B), length (C), and area (D) of cv. ‘Michelangelo®’ and ‘Cosmos®’ cuttings, after 14 weeks from the treatments. Cuttings were treated with: H2O = distilled water; AUX = auxins; BRA = homobrassinolide; KEL = Kelpak®; PHY = Phylgreen. Two-way and one-way ANOVA p-values and Tukey’s post hoc comparisons are reported in the figure (* p < 0.05, ** p < 0.01; *** p < 0.001; ns, not significant). Letters in the figure indicate the level of difference significance.
cv. 'Michelangelo®' cv. 'Cosmos®'0
1
2
3
4
5
6
7
Spro
ut le
nght
(cm
)
H2O AUX BRA KEL PHY
Cultivar × Treatment = ns
P = ns P = ns
Figure 3. Sprout length of cv. ‘Michelangelo®’ and ‘Cosmos®’ cuttings, 14 weeks after the treat-ments. Cuttings were treated with: H2O = distilled water; AUX = auxins; BRA = homobrassinolide;KEL = Kelpak®; PHY = Phylgreen. Two-way and one-way ANOVA p-values are reported in thefigure (ns, not significant).
Horticulturae 2022, 8, x FOR PEER REVIEW 5 of 9
Figure 2. Images of cuttings 16 days after the start of treatments. Orange arrows indicate the callus formation.
Figure 3. Sprout length of cv. ‘Michelangelo®’ and ‘Cosmos®’ cuttings, 14 weeks after the treatments. Cuttings were treated with: H2O = distilled water; AUX = auxins; BRA = homobrassinolide; KEL = Kelpak®; PHY = Phylgreen. Two-way and one-way ANOVA p-values are reported in the figure (ns, not significant).
Figure 4. Root fresh weight (A), dry weight (B), length (C), and area (D) of cv. ‘Michelangelo®’ and ‘Cosmos®’ cuttings, after 14 weeks from the treatments. Cuttings were treated with: H2O = distilled water; AUX = auxins; BRA = homobrassinolide; KEL = Kelpak®; PHY = Phylgreen. Two-way and one-way ANOVA p-values and Tukey’s post hoc comparisons are reported in the figure (* p < 0.05, ** p < 0.01; *** p < 0.001; ns, not significant). Letters in the figure indicate the level of difference significance.
cv. 'Michelangelo®' cv. 'Cosmos®'0
1
2
3
4
5
6
7
Spro
ut le
nght
(cm
)
H2O AUX BRA KEL PHY
Cultivar × Treatment = ns
P = ns P = ns
Figure 4. Root fresh weight (A), dry weight (B), length (C), and area (D) of cv. ‘Michelangelo®’ and‘Cosmos®’ cuttings, after 14 weeks from the treatments. Cuttings were treated with: H2O = distilledwater; AUX = auxins; BRA = homobrassinolide; KEL = Kelpak®; PHY = Phylgreen. Two-way andone-way ANOVA p-values and Tukey’s post hoc comparisons are reported in the figure (* p < 0.05,** p < 0.01; *** p < 0.001; ns, not significant). Letters in the figure indicate the level of difference significance.
3.2. Soluble Sugars
Total soluble sugars at the beginning of the trial and after 4 and 16 days from thetreatments are reported in Table 1. At the beginning of the trial, the amount of solublesugar was the highest measured, and it was similar between the two cultivars. Four daysafter the treatments, the soluble sugar content was lower in both cultivars, and it was

Horticulturae 2022, 8, 561 6 of 9
not significantly different between the treatments. After 16 days from the treatments, thesoluble sugar content was lower than the previous days in all treatments and, moreover, itwas lower under PHY treatment than in control cuttings in cv. ‘Michelangelo®’.
Table 1. Soluble sugars within the bark and the emerging callus of cv. ‘Michelangelo®’ (M)and ‘Cosmos®’ (C) cuttings, 0, 4, and 16 days after the treatments. Cuttings were treated with:H2O = distilled water; AUX = auxins; BRA = homobrassinolide; KEL = Kelpak®; PHY = Phylgreen.Two-way and one-way ANOVA p-values and Tukey’s post hoc comparisons are reported in the table(* p < 0.05; ** p < 0.01; *** p < 0.001; ns, not significant).
Day Cv. Treatment p
H2O AUX BRA KEL PHY T C × T
0M 18.0 ± 1.27 − − − − − −C 20.8 ± 1.79 − − − − − −
4M 8.0 ± 0.95 7.2 ± 1.79 10.0 ± 5.60 8.8 ± 2.19 7.9 ± 3.77 ns nsC 12.3 ± 2.00 14.3 ± 2.71 10.7 ± 1.23 12.6 ± 1.10 14.8 ± 4.00 ns
16M 2.9 ± 1.16 ab 3.8 ± 1.23 a 0.3 ± 0.16 b 0.8 ± 0.82 ab 3.3 ± 1.37 ab *
***C 9.7 ± 1.84 a 11.0 ± 2.19 a 8.2 ± 2.26 a 9.5 ± 2.34 a 1.6 ± 0.16 b **
4. Discussion
The rose is one of the most important and valuable ornamental shrubs worldwide [20].Its propagation is usually achieved by cuttings with a rooting efficiency varying from 0 to100% [4]. The cvs. ‘Cosmos®’ and ‘Michelangelo®’ showed a rooting percentage of about65%, highlighting a medium rooting ability. The possible failures in root formation areusually overcome by producers with the application of plant growth regulators [3]. Inparticular, the rooting promotion of cuttings using auxins is widely used in commercialplant propagation [7]. Indeed, AUX treatment showed an increase in survival percentage,callus formation, and root biometric parameters in both cultivar cuttings in comparisonwith the control conditions, i.e., only distilled water. Other authors reported an increasefrom 33 to 65% in rooting of R. centifolia medial cuttings in the same substrate using3500 ppm of auxins [21] or from 67 to 75% or 81% in rooting of R. damascena cuttings insand using 200 mg dm−3 IAA or 25 mg dm−3 of NAA, respectively [22]. Thus, the aim ofthis work was to find alternative substances to auxins for the promotion of cutting rootingsuitable for rose propagation.
BRs have already shown their role in rooting promotion of ornamental plant cut-tings [10], but commercial synthetic products are still based principally on auxins and, toour knowledge, they have never been tested on rose cuttings. BRs have shown contrastingeffects on rooting in woody cuttings of different plant genotypes [23,24]. In addition, inour experimental conditions, 22(S),23(S)-homobrassinolide showed opposite effects on thetwo rose cultivars. The cuttings of cv. ‘Michelangelo®’ treated with BRA had a similarbehavior to those treated with only distilled water, while in cv. ‘Cosmos®’ this treatmentshowed the best results in terms of root biometric parameters. Thus, BRs were found to beunsuitable for propagation in operational conditions since testing on every specific rosegenotype is required. The worst performance in rooting under BRA treatment retrievedin cv. ‘Michelangelo®’ matched with a low amount in soluble sugars within the emergingcallus 16 days after the treatment. Since a positive correlation between root number andsoluble sugar concentration was reported in other rose cultivar cuttings [19], this resultmight support a less susceptibility of cv. ‘Michelangelo®’ to rooting under BRA treatment.Indeed, other authors also highlighted that a depletion of carbohydrates reduced callus androot formation in single-node leafy stem cuttings [25]. However, a constitutional decreasein soluble sugars during the first 15 days has already been reported within the bark ofcuttings of Zizyphus jujuba Mill. [26] and could explain the lower level of soluble sugarsfound in all rose cuttings in comparison with the beginning of the trial.

Horticulturae 2022, 8, 561 7 of 9
Biostimulants like seaweed extracts seem to be a promising alternative to syntheticproducts in horticulture [14]. Seaweed extracts have been successfully used for the promo-tion of rooting in several ornamental species. Specifically, Kelpak® has been demonstratedto promote the growth of leafy pelargonium cuttings [27]. In our experimental conditions,this natural product increased the rooting percentage and the root biometric parameters ofboth rose cultivar cuttings. In particular, under this treatment, the cuttings had the highestrooting percentage in cv. ‘Michelangelo®’. Kelpak® has been shown to contain abscisicacid, gibberellins, and BRs in higher concentrations than in kelp E. maxima tissues, andthese hormones can directly contribute to the promotion of plant growth [28]. Indeed,Kelpak® treatment increased the concentration of cytokinin and bioactive compounds inaerial parts of Eucomis autumnalis [29], as well as increased rooting rate of cuttings of Prunusmarianna stockplants [30]. On the contrary, Phylgreen did not significantly improve therooting performance of both cultivar cuttings, while in previous studies, the use of a 40%concentrated A. nodosum extract increased the rooting of Passiflora actinia by about 10% [31]and the rhizogenesis in hypocotyl slice of Prunus domestica under in vitro propagationexperiments by about 30% [32]. The specific application of Phylgreen on Arabidopsis thalianaplants promoted abiotic stress resistance and growth if applied repeatedly throughout thecropping cycle [33], therefore highlighting the need to test the time and method of applica-tion. Indeed, a lower concentration of soluble sugars was measured under this treatmentin cv. ‘Cosmos®’, probably highlighting a low metabolic activity. The limited informationavailable in the literature does not allow further argumentation on this biostimulant sincecontrasting results were observed on rose cuttings with respect to other investigated species.However, the results highlighted the importance of evaluating the type of seaweed to treateach specific ornamental species on the basis of the application method.
Interestingly, root DW was not particularly altered by any treatment, highlightingthat the root biometric variations were principally related to increased water content andthus cell elongation. Indeed, the role of auxins in promoting cell elongation throughturgor-driven processes is already known [34].
5. Conclusions
In conclusion, Kelpak® was shown to improve both rooting percentage and rootbiometric parameters of both rose cultivar cuttings, highlighting its suitability for thereplacement of synthetic products used for rooting promotion. BRs showed a species-dependent effect, highlighting the necessity of evaluating their applicability before their useas alternative synthetic phytoregulators to auxins. Phylgreen did not improve the rootingperformance of rose cuttings, indicating that this natural product cannot be suitable for rosepropagation using this methodology. Since the use of authorized hormonal agrochemicalsis very limited in some countries and is getting even lower every year, this study offers anew possible product also available for organic systems. Further efforts must be conductedto clarify the biochemical and molecular mechanisms of action of this seaweed extract.
Author Contributions: Conceptualization and investigation, S.T., S.C. and B.N.; trial set up andsampling, S.T., S.C. and B.N.; formal analysis and data curation, S.T.; funding acquisition, S.C. andB.N.; Writing—original draft, S.T.; Writing—review & editing, S.T., S.C. and B.N. All authors haveread and agreed to the published version of the manuscript.
Funding: This research was funded by the Italian Ministry of Agriculture, Food and Forestry Policies,sub-project ‘Tecnologie digitali integrate per il rafforzamento sostenibile di produzioni e trasfor-mazioni agroalimentari (AgroFiliere)’, AgriDigit program (DM 36503.7305.2018 of 20 December 2018).
Institutional Review Board Statement: Not applicable.
Informed Consent Statement: Not applicable.
Data Availability Statement: The data presented in this study are available on request from thecorresponding author.

Horticulturae 2022, 8, 561 8 of 9
Acknowledgments: The authors would like to thank Monica Michelotti and Paolo Bini for theirtechnical support during the trial.
Conflicts of Interest: The authors declare no conflict of interest.
References1. Cardoso, J.C.; Vendrame, W.A. Innovation in propagation and cultivation of ornamental plants. Horticulturae 2022, 8, 229.
[CrossRef]2. Kentelky, E.; Jucan, D.; Cantor, M.; Szekely-Varga, Z. Efficacy of different concentrations of NAA on selected ornamental woody
shrubs cuttings. Horticulturae 2021, 7, 464. [CrossRef]3. Akhtar, M.S.; Khan, M.A.; Riaz, A.; Younis, A. Response of different rose species to different root promoting hormones. Pak. J.
Agric. Sci. 2002, 39, 297–299.4. Nguyen, T.H.N.; Tänzer, S.; Rudeck, J.; Winkelmann, T.; Debener, T. Genetic analysis of adventitious root formation in vivo and
in vitro in a diversity panel of roses. Sci. Hortic. 2020, 266, 109277. [CrossRef]5. Zhang, W.; Fan, J.; Tan, Q.; Zhao, M.; Cao, F. Mechanisms underlying the regulation of root formation in Malus hupehensis stem
cuttings by using exogenous hormones. J. Plant Growth Regul. 2017, 36, 174–185. [CrossRef]6. Paradikovic, N.; Teklic, T.; Zeljkovic, S.; Lisjak, M.; Špoljarevic, M. Biostimulants research in some horticultural plant species—A
review. Food Energy Secur. 2019, 8, e00162. [CrossRef]7. Blythe, E.K.; Sibley, J.L.; Tilt, K.M.; Ruter, J.M. Methods of auxin application in cutting propagation: A review of 70 years of
scientific discovery and commercial practice. J. Environ. Hortic. 2007, 25, 166–185. [CrossRef]8. Betti, C.; Della Rovere, F.; Piacentini, D.; Fattorini, L.; Falasca, G.; Altamura, M.M. Jasmonates, ethylene and brassinosteroids
control adventitious and lateral rooting as stress avoidance responses to heavy metals and metalloids. Biomolecules 2021, 11, 77.[CrossRef]
9. Vardhini, B.V. Modifications of morphological and anatomical characteristics of plants by application of brassinosteroids undervarious abiotic stress conditions—A review. Plant Gene 2017, 11, 70–89. [CrossRef]
10. Pacholczak, A.; Zajaczkowska, M.; Nowakowska, K. The Effect of brassinosteroids on rooting of stem cuttings in two barberry(Berberis thunbergii L.) cultivars. Agronomy 2021, 11, 699. [CrossRef]
11. Moreno-Castillo, E.; Ramírez-Echemendía, D.P.; Hernández-Campoalegre, G.; Mesa-Tejeda, D.; Coll-Manchado, F.; Coll-García, Y.In silico identification of new potentially active brassinosteroid analogues. Steroids 2018, 138, 35–42. [CrossRef] [PubMed]
12. Xu, L.; Geelen, D. Developing biostimulants from agro-food and industrial by-products. Front. Plant Sci. 2018, 9, 1567. [CrossRef][PubMed]
13. Brzozowski, L.; Mazourek, M.A. Sustainable agricultural future relies on the transition to organic agroecological pest management.Sustainability 2018, 10, 2023. [CrossRef]
14. Baltazar, M.; Correia, S.; Guinan, K.J.; Sujeeth, N.; Bragança, R.; Gonçalves, B. Recent advances in the molecular effects ofbiostimulants in plants: An overview. Biomolecules 2021, 11, 1096. [CrossRef]
15. Dmytryk, A.; Chojnacka, K. Algae as fertilizers, biostimulants, and regulators of plant growth. In Algae biomass: Characteristics andApplications; Chojnacka, K., Wieczorek, P.P., Schroeder, G., Michalak, I., Eds.; Springer: Cham, Switzerland, 2018; pp. 115–122.
16. Patel, J.S.; Mukherjee, A. Seaweed and associated products: Natural biostimulant for improvement of plant health. In EmergingTrends in Plant Pathology; Singh, K.P., Jahagirdar, S., Sarma, B.K., Eds.; Springer: Singapore, 2021; pp. 317–330.
17. Ali, O.; Ramsubhag, A.; Jayaraman, J. Biostimulant properties of seaweed extracts in plants: Implications towards sustainablecrop production. Plants 2021, 10, 531. [CrossRef]
18. Kisvarga, S.; Farkas, D.; Boronkay, G.; Neményi, A.; Orlóci, L. Effects of biostimulants in horticulture, with emphasis onornamental plant production. Agronomy 2022, 12, 1043. [CrossRef]
19. Otiende, M.A.; Nyabundi, J.O.; Ngamau, K.; Opala, P. Effects of cutting position of rose rootstock cultivars on rooting and itsrelationship with mineral nutrient content and endogenous carbohydrates. Sci. Hortic. 2017, 225, 204–212. [CrossRef]
20. Vazquez-Iglesias, I.; Ochoa-Corona, F.M.; Tang, J.; Robinson, R.; Clover, G.R.; Fox, A.; Boonham, N. Facing Rose rosette virus: Arisk to European rose cultivation. Plant Pathol. 2020, 69, 1603–1617. [CrossRef]
21. Al-Saqri, F.; Alderson, P.G. Effects of IBA, cutting type and rooting media on rooting of Rosa centifolia. J. Hortic. Sci. 1996, 71,729–737. [CrossRef]
22. Pati, P.K.; Prakash, O.; Sharma, M.; Sood, A.; Ahuja, P.S. Growth performance of cuttings raised from in vitro and in vivopropagated stock plants of Rosa damascena Mill. Biol. Plant. 2004, 48, 609–611. [CrossRef]
23. Bannoud, F.; Bellini, C. Adventitious rooting in Populus species: Update and perspectives. Front. Plant Sci. 2021, 12, 918.[CrossRef]
24. Farazi, E.; Afshari, H.; Hokm Abadi, H. Effect of different concentrations of brassinosteroid on physiomorphological characteristicsof five pistachio genotypes (Pistacia vera L). J. Nuts 2015, 6, 143–153.
25. Costa, J.M.; Heuvelink, E.; Van de Pol, P. Propagation by cuttings. In Reference Module in Life Sciences; Roitberg, B.D., Ed.; Elsevier:Amsterdam, The Netherlands, 2017.
26. Shao, F.; Wang, S.; Huang, W.; Liu, Z. Effects of IBA on the rooting of branch cuttings of Chinese jujube (Zizyphus jujuba Mill.) andchanges to nutrients and endogenous hormones. J. For. Res. 2018, 29, 1557–1567. [CrossRef]

Horticulturae 2022, 8, 561 9 of 9
27. Krajnc, A.U.; Ivanus, A.; Kristl, J.; Susek, A. Seaweed extract elicits the metabolic responses in leaves and enhances growth ofPelargonium cuttings. Eur. J. Hortic. Sci. 2012, 77, 170–181.
28. Stirk, W.A.; Tarkowská, D.; Turecová, V.; Strnad, M.; Van Staden, J. Abscisic acid, gibberellins and brassinosteroids in Kelpak®,a commercial seaweed extract made from Ecklonia maxima. J. Appl. Phycol. 2014, 26, 561–567. [CrossRef]
29. Aremu, A.O.; Placková, L.; Gruz, J.; Bíba, O.; Novák, O.; Stirk, W.A.; Doleza, K.; Van Staden, J. Seaweed-derived biostimulant(Kelpak®) influences endogenous cytokinins and bioactive compounds in hydroponically grown Eucomis autumnalis. J. PlantGrowth Regul. 2016, 35, 151–162. [CrossRef]
30. Szabó, V.; Sárvári, A.; Hrotkó, K. Treatment of stockplants with biostimulators and their effects on cutting propagation of Prunusmarianna ‘GF 8-1’. Acta Hortic. 2011, 923, 277–282. [CrossRef]
31. Gomes, E.N.; Vieira, L.M.; Tomasi, J.D.C.; Tomazzoli, M.M.; Grunennvaldt, R.L.; Fagundes, C.D.M.; Machado, R.C.B. Brownseaweed extract enhances rooting and roots growth on Passiflora actinia Hook stem cuttings. Ornam. Hortic. 2018, 24, 269–276.[CrossRef]
32. Faize, M.; Faize, L.; Burgos, L.; Critchley, A.; Albuquerque, N. Application of Ascophyllum nodosum-based soluble extract onmicropropagation and regeneration of Nicotiana benthamiana and Prunus domestica. Plants 2021, 10, 1354. [CrossRef]
33. Fleming, T.R.; Fleming, C.C.; Levy, C.C.; Repiso, C.; Hennequart, F.; Nolasco, J.B.; Liu, F. Biostimulants enhance growth anddrought tolerance in Arabidopsis thaliana and exhibit chemical priming action. Ann. Appl. Biol. 2019, 174, 153–165. [CrossRef]
34. Majda, M.; Robert, S. The role of auxin in cell wall expansion. Int. J. Mol. Sci. 2018, 19, 951. [CrossRef] [PubMed]
Copyright © 2022 FDOKUMEN