
In vitro salinity tolerance of two pistachio rootstocks: Pistacia vera L. and P. atlantica Desf
11
Environmental and Experimental Botany 69 (2010) 302–312 Contents lists available at ScienceDirect Environmental and Experimental Botany journal homepage: www.elsevier.com/locate/envexpbot In vitro salinity tolerance of two pistachio rootstocks: Pistacia vera L. and P. atlantica Desf Azza Chelli-Chaabouni a,∗ , Afef Ben Mosbah b , Mohamed Maalej b , Kamel Gargouri c , Radhia Gargouri-Bouzid d , Noureddine Drira b a Research Unity, Genetic Resources and Breeding of Olive tree, Pistachio, and Almond, Olive Tree Institute, PB 1087, 3018 Sfax, Tunisia b Faculty of Sciences of Sfax, Road of Soukra km 4, PB 802, 3038 Sfax, Tunisia c Research Laboratory, Improving of Olive tree and Arboriculture Productivity, Olive Tree Institute, PB 1087, 3018 Sfax, Tunisia d Engineers National School of Sfax, Road of Soukra km 4, PB W, 3038 Sfax, Tunisia article info Article history: Received 10 November 2009 Received in revised form 29 April 2010 Accepted 12 May 2010 Keywords: Pistacia rootstock Salt tolerance Growth Mineral uptake Proline Soluble sugars abstract Seedlings of Pistacia vera L. and Pistacia atlantica Desf. were cultured on hormone-free DKW medium sup- plemented with NaCl. The plants were subjected to low NaCl concentrations ranging from 0 to 80 mM for 45 days or to high salt concentrations (0, 131, and 158.5 mM for P. vera and 0, 131, and 240 mM for P. atlantica) for 25 days. Toxicity symptoms were recorded for seedlings exposed to low NaCl treatments. Plant growth, survival rates, mineral content, as well as proline and soluble sugar contents were deter- mined and evaluated at the end of the culture period. The results indicated that low NaCl treatments yielded no instances of plant death in both species. At high salt conditions, however, significant mortality rates were noted for both species, being 22.86% at 240 mM NaCl for P. atlantica and 25.8% at 158.5 mM NaCl for P. vera. With regards to salinity effects, levels of 60 and 80 mM NaCl induced significant decreases of stem elongation and leaf number in the P. vera species. Salinities between 40 and 80 mM NaCl, how- ever, induced a decrease in the root number of both species. The fresh weights of P. vera and P. atlantica also decreased significantly after 45 days of culture at NaCl concentrations between 40 and 80 mM and after 25 days of culture at 158.5 and 240 mM NaCl, respectively. The sodium and chloride uptake in plant organs seemed to be controlled more efficiently in P. atlantica than in P. vera. In both species, the K + content was noted to undergo a significant decrease when salinity increased. While the K + /Na + ratio was maintained above 2 at low NaCl treatments, it was sharply decreased at high NaCl conditions, suggesting a failure of K–Na selectivity mechanism. The Ca 2+ /Na + ratio decreased significantly at 60 and 80 mM NaCl in P. vera and at 60 mM NaCl for P. atlantica. In both Pistacia species, high NaCl treatments (131–240 mM NaCl) induced a significant increase in proline content. © 2010 Elsevier B.V. All rights reserved. 1. Introduction Salinity is one of the major abiotic stresses that affect plant pro- duction and growth in many arid and semi-arid areas throughout the world (Gebauer et al., 2004). In Tunisia, saline areas cover about three-quarters of the total land surface, with an annual rainfall of less than 400 mm (Anonymous, 1998). Inappropriate crop produc- tion and irrigation systems that employ poor quality water further aggravate the situation through the generation of additional sec- ondary salinisation processes (Gebauer et al., 2004; Türkan and Demiral, 2009). Soil salinity is often reported to have adverse effects on crop growth and productivity particularly through the alteration of the ∗ Corresponding author. Tel.: +216 74 241 240; fax: +216 74 241 033. E-mail address: a [email protected] (A. Chelli-Chaabouni). metabolic processes of plants by inducing changes in the ion tox- icity and osmotic pressure of the tissues. The responses to these alterations are often accompanied by a variety of symptoms, such as the reduction of leaf area, abscission of leaves, increase of leaf thickness and succulence, necrosis of shoot and root, and reduc- tion of internode lengths (Gucci and Tattini, 1997; Kozlowski, 1997; Parida and Das, 2005). The onset of the damage depends on the environmental conditions (e.g. salt composition and concentration, duration of the stress, temperature, and soil quality) as well as on the characteristics of the species (e.g. woody or herbaceous species, age, stage of development, and genotype). Glycophyte species develop different adaptive mechanisms either to exclude salt from their cells or to tolerate it within the cells (Kozlowski, 1997; Parida and Das, 2005). The uptake of large amounts of salt by the plant leads to the increase of the osmotic pressure in the cytosol. Cell homeostasis is maintained by an osmotic adjustment mechanism. It consists of the sequestration 0098-8472/$ – see front matter © 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.envexpbot.2010.05.010
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of In vitro salinity tolerance of two pistachio rootstocks: Pistacia vera L. and P. atlantica Desf

IP
AKa
b
c
d
a
ARRA
KPSGMPS
1
dttltaoD
g
0d
Environmental and Experimental Botany 69 (2010) 302–312
Contents lists available at ScienceDirect
Environmental and Experimental Botany
journa l homepage: www.e lsev ier .com/ locate /envexpbot
n vitro salinity tolerance of two pistachio rootstocks:istacia vera L. and P. atlantica Desf
zza Chelli-Chaabounia,∗, Afef Ben Mosbahb, Mohamed Maalejb,amel Gargouri c, Radhia Gargouri-Bouzidd, Noureddine Drirab
Research Unity, Genetic Resources and Breeding of Olive tree, Pistachio, and Almond, Olive Tree Institute, PB 1087, 3018 Sfax, TunisiaFaculty of Sciences of Sfax, Road of Soukra km 4, PB 802, 3038 Sfax, TunisiaResearch Laboratory, Improving of Olive tree and Arboriculture Productivity, Olive Tree Institute, PB 1087, 3018 Sfax, TunisiaEngineers National School of Sfax, Road of Soukra km 4, PB W, 3038 Sfax, Tunisia
r t i c l e i n f o
rticle history:eceived 10 November 2009eceived in revised form 29 April 2010ccepted 12 May 2010
eywords:istacia rootstockalt tolerancerowthineral uptake
rolineoluble sugars
a b s t r a c t
Seedlings of Pistacia vera L. and Pistacia atlantica Desf. were cultured on hormone-free DKW medium sup-plemented with NaCl. The plants were subjected to low NaCl concentrations ranging from 0 to 80 mMfor 45 days or to high salt concentrations (0, 131, and 158.5 mM for P. vera and 0, 131, and 240 mM for P.atlantica) for 25 days. Toxicity symptoms were recorded for seedlings exposed to low NaCl treatments.Plant growth, survival rates, mineral content, as well as proline and soluble sugar contents were deter-mined and evaluated at the end of the culture period. The results indicated that low NaCl treatmentsyielded no instances of plant death in both species. At high salt conditions, however, significant mortalityrates were noted for both species, being 22.86% at 240 mM NaCl for P. atlantica and 25.8% at 158.5 mMNaCl for P. vera. With regards to salinity effects, levels of 60 and 80 mM NaCl induced significant decreasesof stem elongation and leaf number in the P. vera species. Salinities between 40 and 80 mM NaCl, how-ever, induced a decrease in the root number of both species. The fresh weights of P. vera and P. atlanticaalso decreased significantly after 45 days of culture at NaCl concentrations between 40 and 80 mM and
after 25 days of culture at 158.5 and 240 mM NaCl, respectively. The sodium and chloride uptake in plantorgans seemed to be controlled more efficiently in P. atlantica than in P. vera. In both species, the K+content was noted to undergo a significant decrease when salinity increased. While the K+/Na+ ratio wasmaintained above 2 at low NaCl treatments, it was sharply decreased at high NaCl conditions, suggestinga failure of K–Na selectivity mechanism. The Ca2+/Na+ ratio decreased significantly at 60 and 80 mM NaClin P. vera and at 60 mM NaCl for P. atlantica. In both Pistacia species, high NaCl treatments (131–240 mM
t inc
NaCl) induced a significan. Introduction
Salinity is one of the major abiotic stresses that affect plant pro-uction and growth in many arid and semi-arid areas throughouthe world (Gebauer et al., 2004). In Tunisia, saline areas cover abouthree-quarters of the total land surface, with an annual rainfall ofess than 400 mm (Anonymous, 1998). Inappropriate crop produc-ion and irrigation systems that employ poor quality water furtherggravate the situation through the generation of additional sec-
ndary salinisation processes (Gebauer et al., 2004; Türkan andemiral, 2009).Soil salinity is often reported to have adverse effects on croprowth and productivity particularly through the alteration of the
∗ Corresponding author. Tel.: +216 74 241 240; fax: +216 74 241 033.E-mail address: a [email protected] (A. Chelli-Chaabouni).
098-8472/$ – see front matter © 2010 Elsevier B.V. All rights reserved.oi:10.1016/j.envexpbot.2010.05.010
rease in proline content.© 2010 Elsevier B.V. All rights reserved.
metabolic processes of plants by inducing changes in the ion tox-icity and osmotic pressure of the tissues. The responses to thesealterations are often accompanied by a variety of symptoms, suchas the reduction of leaf area, abscission of leaves, increase of leafthickness and succulence, necrosis of shoot and root, and reduc-tion of internode lengths (Gucci and Tattini, 1997; Kozlowski, 1997;Parida and Das, 2005). The onset of the damage depends on theenvironmental conditions (e.g. salt composition and concentration,duration of the stress, temperature, and soil quality) as well as onthe characteristics of the species (e.g. woody or herbaceous species,age, stage of development, and genotype).
Glycophyte species develop different adaptive mechanisms
either to exclude salt from their cells or to tolerate it within thecells (Kozlowski, 1997; Parida and Das, 2005). The uptake of largeamounts of salt by the plant leads to the increase of the osmoticpressure in the cytosol. Cell homeostasis is maintained by anosmotic adjustment mechanism. It consists of the sequestration
l and
ooii
iuicAsSntaa(fAMi(cotina
icoetttar
ap(oaelec2ltNlsltwbTta
ppls
A. Chelli-Chaabouni et al. / Environmenta
f large quantities of salt ions in the vacuole and/or synthesis ofrganic osmoprotectants (Munns, 2002). These are named compat-ble solutes for they can accumulate to high concentrations withoutnterfering with the metabolism of the plant.
The uptake of large amounts of salt by the plant also leads toonic imbalance. In fact, the disturbance of potassium (K+) nutritionnder salt conditions by potassium–sodium (K+–Na+) interaction
s a common feature in plants and is often associated with K+ defi-iency (Cramer et al., 1987; Slama, 2004; Parida and Das, 2005).
selective uptake and transport of K+ into the shoots is neces-ary for salt tolerance in many higher plants (Cramer et al., 1987).ince a high K+/Na+ ratio in the cytosol is essential to maintainormal cellular functions (Chinnusamy et al., 2005), plant saltolerance strongly depends on the K+ nutrition status (Maathuisnd Amtmann, 1999). The photosynthetic activity and nitrogenssimilation under salt conditions were related to the K+/Na+ ratioTabatabaei, 2006). A high K+/Na+ ratio in the leaves is, there-ore, often considered as a salt-tolerance marker (Maathuis andmtmann, 1999; Chartzoulakis et al., 2002; Dasgan et al., 2002;eena et al., 2003). The calcium (Ca2+) content, on the other hand,
ncreases (Ramoliya et al., 2004; De Pascale et al., 2007) or decreasesGratten and Grieve, 1999; Unno et al., 2002) under salt stressonditions depending on the specific physiology of the plant, thergans, the nature of the stress, as well as its duration. The rolehat Ca2+ plays in the alleviation of salt toxicity is reported tonfluence K+/Na+ selectivity through controlling the Na+ influx viaon-selective ion channels (Cramer et al., 1987; Rengel, 1992; Guccind Tattini, 1997; Melgar et al., 2006; Sotiropoulos, 2007).
In this context, the roots are often reported to play a key rolen the salt tolerance of plants for they represent the first organs toontrol the uptake and translocation of nutrients and salts through-ut the plant. Despite the direct exposure of these organs to salinenvironment, their growth is less vulnerable to salt than that ofhe shoots (Munns, 2002). Moreover, the accumulation of Na+ inhe roots is an adaptive response used by several woody specieso avoid its toxicity in the shoots (Walker et al., 1987; Picchioni etl., 1990; Gucci and Tattini, 1997). Accordingly, the control of theoot-to-shoot transport of salt can serve as a criterion for tolerance.
In many plants, compatible osmoprotectant metabolites, suchs proline, Glycine-betaine, and soluble sugars, are produced torotect the cells against the damaging effects from salt stressKozlowski, 1997; Sairam and Tyagi, 2004). Salt stressed plantsften accumulate proline in the leaves of perennial (Harinasut etl., 2000; Meena et al., 2003) or non-perennial species (Hernandezt al., 2000; Ben Khaled et al., 2003). Instances of unchanged pro-ine content were, nevertheless, reported in the literature (Tattinit al., 1993). In many species, high accumulation of proline is asso-iated with tolerance to stress (Hare et al., 1998; Sairam and Tyagi,004; Hokmabadi et al., 2005). Accordingly, the ability to accumu-
ate proline has often been proposed as a valuable criterion forhe selection of salt tolerant genotypes (Zid and Grignon, 1991).evertheless, there is uncertainty with regards to the role of pro-
ine in plant salt tolerance as well as its specificity towards salttress (Munns, 2002). In fact, negative connections between pro-ine accumulation and salt tolerance were previously reported inhe literature (Ashraf, 1989; Mademba-SY et al., 2003). Similarly,hile several studies reported the induction of an enhanced solu-
le sugar content by salt stress in many plant species (Gucci andattini, 1997), other works described the instances where levels ofhese carbohydrates were either constant or reduced (Rejskovà etl., 2007).
Considering the damaging effects of salinity on crop growth androductivity, the identification, valorisation, and conservation oflant genetic resources has received considerable attention in the
ast few decades. Of particular interest to the aims of the presenttudy, the application of genetic resources for the breeding of salt
Experimental Botany 69 (2010) 302–312 303
tolerant genotypes is considered as a valuable tool to developsustainable agriculture in dry lands. So far, reports on species prop-agated by grafting often describe salinity tolerance as being relatedto the rootstock (Okubo et al., 2000; Rivera et al., 2003; Huanget al., 2010). In this respect, the selection of tolerant genotypesis needed to enhance productivity. Of special interest, among thecommonly known salt sensitive fruit trees, pistachio (Pistacia veraL.) is considered as relatively tolerant. In fact, there are still varyingreports about the relative degree towards which Pistacia species areconsidered salt tolerant. The reports currently available in the lit-erature presents different levels of salinity tolerance ranging frommoderately sensitive (Sepaskhah and Maftoun, 1988; Picchioni etal., 1990; FAO, 2002) to highly tolerant (Behboudian et al., 1986;Walker et al., 1987). This variation is, in fact, explicable for it reflectsa number of inter and intra specific variations as well as to the effectof different environmental conditions such as the quality of the soiland the temperature. Moreover, and despite their capacity to copewith induced salt stress, few reports attribute the yield reduction ofthese species to increasing salinity (Sepaskhah et al., 1985; Arzaniand Hokmabadi, 2004; Hokmabadi et al., 2005).
Pistachio nut trees are propagated by grafting because therooting of cuttings is very low. In Tunisia, pistachio varieties aretypically grafted on P. vera rootstocks whose good compatibilitywith major cultivars is curtailed by their sensitivity to nematodes.Although the wild species Pistacia atlantica is tolerant to salinity,cold, and nematodes (Ferguson et al., 2005), it has been largely dis-regarded by propagators in last decades in favour of P. vera. This ismainly because of the availability and relatively higher germina-tion rates of seeds from the P. vera species. This low diversificationof rootstock threatens the growth and conservation of pistachiospecies in Tunisia, particularly in terms of environmental stressadaptation. The development of irrigated grounds, which has oftenbeen proposed for the enhancement of productivity, unfortunatelyprovides a double-edged solution that adds to the salinisation ofsoils and the development of pathogens.
In view of the serious concerns described above, special atten-tion should be given to the development of highly performingpistachio rootstocks through the early selection of salt tolerantgenotypes. In this context, a thorough understanding of the mech-anisms adopted by a particular species to avoid toxic salt effectsbecomes a precondition for the attainment of principled andappropriate selection approaches (Ashraf, 2004). Since the fieldevaluation of salt effects is highly correlated with environmentalconditions, in vitro screening techniques would allow for a bettercontrol of culture conditions (Vijayan et al., 2003). So far, the invitro response of many woody plants to induced salinity has beenassessed using whole plants (Bracci et al., 2006) and isolated organs,such as microshoots (Watanabe et al., 2000; Mills et al., 2001; Shibliand Al-Juboory, 2002; Vijayan et al., 2003; Rejskovà et al., 2007;Roussos et al., 2007) and callus (Shibli and Al-Juboory, 2002), as wellas cell suspension cultures (Shibli and Al-Juboory, 2002; Ferreiraand Lima-Costa, 2006). The investigation of the effects of salt onisolated tissues, organs, cells, or calluses may not, however, reflectexactly what may happen in the whole plant (Zid and Grignon,1991; Hernandez et al., 2000). In fact, studies involving the wholeplant can provide information that cannot be obtained otherwise.This includes data on organ interactions, ion transport, antago-nisms, and metabolic activities involved in responses to salt stress.The regeneration via the in vitro organogenesis of selected lines, onthe other hand, can increase the risk of somaclonal variation.
Despite the expressed concerns over the negative impacts of
salinity on plant culture, only few reports have so far investigatedthe salt tolerance of Pistacia species and, to the authors’ knowledge,none of these studies was performed under in vitro conditions.Understanding the mechanisms involved in the response to salin-ity would be highly useful for the rapid and early screening of salt
3 l and E
tsav
srs
2
2
wt(eTfmac
satdfDldwt5cwiTdwo
2
alto
2
glFao
2
ln
04 A. Chelli-Chaabouni et al. / Environmenta
olerant genotypes. The early stage of development usually repre-ents the most salt sensitive phase in woody plants (Shannon etl., 1994). For this reason, young seedlings represent proper andaluable material for the detection of early salt response.
In view of the limited knowledge currently available on thealt tolerance of Pistacia species, the present work investigates andeports on the effects of low and high salt concentrations on theeedlings of two Pistacia species cultured in vitro.
. Materials and methods
.1. Materials and culture conditions
Mature seeds of open-pollinated P. vera and P. atlantica speciesere sampled. P. vera seeds were collected from the experimen-
al station of the Olive Tree Institute, situated in the North of SfaxTunisian South-East area) while the seeds of P. atlantica were gath-red from wild trees in Meknassy (Tunisian Centre-West area).he seedlings obtained from in vitro germination were selectedor their homogeneity and cultivated for one month on a DKW
edium (Driver and Kuniyuki, 1984) supplemented with 7 g l−1
gar and 30 g l−1 sucrose. The pH was adjusted to 5.7 before auto-laving.
Since sodium chloride (NaCl) represents the major source ofalt in irrigation water and soil solutions (Slama, 2004; Türkannd Demiral, 2009), it was used as the source of salt throughouthe experimental assays of this study. Several treatments, withifferent NaCl concentrations and culture durations, were per-ormed on one-month-old seedlings from both species cultured onKW media supplemented with NaCl. The treatments involving
ow NaCl concentrations (0, 20, 40, 60, and 80 mM) were con-ucted for 45 days while those involving high NaCl concentrationsere performed for 25 days. In the latter case, the NaCl concen-
ration was applied at 0 mM, ND50 (NaCl concentration inducing0% of total foliage necrosis of seedlings), and LD50 (NaCl con-entration leading to 50% plant death). The ND50 and LD50 valuesere determined for each species after increasingly administer-
ng concentrations of NaCl (by steps of 20 mM) to the seedlings.he number of seedlings reaching total foliar necrosis and planteath was recorded throughout the culture period. All culturesere incubated at a temperature of 24 ± 2 ◦C and a photoperiod
f 16 h provided by daylight fluorescent lamps.
.2. Survival rates
The rates of the surviving plants involved in all experimentalssays were calculated at the end of the culture period using the fol-owing equation: Number of the survived plants × 100/number ofotal plants. A plant was considered dead when it showed necrosisf all leaves, stems, and roots.
.3. Growth parameters
In the case of low NaCl concentration treatments, the stem elon-ation as well as the number and fresh weights of newly developedeaves, shoots, and roots were determined after 45 days of culture.or the treatments using high NaCl concentrations, the shoot (stemsnd leaves) and root fresh weights were determined after 25 daysf culture.
.4. Toxicity symptoms
For sodium chloride treatments from 0 to 80 mM, the number ofeaves showing total necrosis was counted 3 days after the begin-ing of the experiment and every 5 days throughout the culture
xperimental Botany 69 (2010) 302–312
period. The percentage of altered leaves was determined as follows:number of altered leaves × 100/total leaf number.
2.5. Mineral analyses
Mineral analyses were carried out at the end of the culture peri-ods on dry material. Dry ashing of plant material was obtained at400 ◦C after a drying time of at least 72 h at 70 ◦C until weightstabilisation. The mineralization of the samples was obtainedafter a digestion process in 1N nitric acid solution (HNO3
−). Thesodium (Na+) and potassium (K+) contents were measured usingthe flame emission photometry (Jenway PFP7, Bibby Scientific lim-ited, Staffordshire, UK). Chloride (Cl−) content was analysed bytitration with 0.1N silver nitrate (AgNO3) in the presence of potas-sium bichromate according to a modified colorimetric method ofMohr (Mathieu and Pieltain, 2003). Calcium (Ca2+) content wasanalyzed using an atomic absorption spectrophotometer (AAna-lyst 300, PerkinElmer, Inc., Waltham, MA, USA) only for the plantstreated at NaCl concentration ranging from 0 to 80 mM. All thevalues reported represent the means of at least three replications.
2.6. Biochemical analyses
Proline and soluble sugar contents were determined for theplants that were subjected to high NaCl treatments as described byBartes et al. (1973) and Robyt and White (1987), respectively. Pro-line was extracted from 0.2 g fresh leaves ground in liquid nitrogenand mixed with 10 ml of 3% sulphosalicylic acid. After centrifuga-tion for 15 min at 6000 rpm, the supernatant was added to 2 ml of afreshly prepared ninhydrin acid solution in test tubes. The tubeswere incubated in a 100 ◦C water bath for 1 h and then placedin ice to complete the reaction. Two millilitres of toluene wereadded to the extract, vortexed for 15–20 s, and kept at room tem-perature until the separation of the toluene and aqueous phases.The toluene–chromophore phase was carefully sampled and theabsorbance was measured at 520 nm in a spectrophotometer. Theproline concentration (�moles gfw−1) was determined from a stan-dard curve. Soluble sugars were extracted from 0.2 to 0.4 g freshleaves by heating with 5 ml of 80% methanol solution in a waterbath at 70 ◦C for 30 min. One millilitre of the extract obtained wasmixed with 1 ml of 5% phenol solution and 5 ml of sulphuric acid.The mixed solution was kept to cooling and then vortexed. Theabsorbance was measured at 640 nm in a spectrophotometer. Thevalues obtained were reported to a standard curve of glucose todetermine the soluble sugars content (mg glucose g fw−1).
2.7. Statistics
The data were subjected to one-way ANOVA analyses usingSPSS software. Differences were determined by Duncan’s multi-ple range tests at p ≤ 0.05. The comparison of means of growthparameters was realized separately for each species and after eachculture period because of the phenotypic differences of the species.Comparisons in terms of the mineral and biochemical data wereperformed between all treatments and species. Bi-varied corre-lations were determined according to Pearson test at p ≤ 0.01 orp ≤ 0.05.
3. Results
The application of increasing NaCl concentrations to both Pista-cia species allowed the determination of the ND50 and LD50 values.While a similar ND50 value of 131 mM NaCl was exhibited by bothspecies, the LD50 value of P. vera was 158.5 mM and that of P.atlantica was 240 mM.
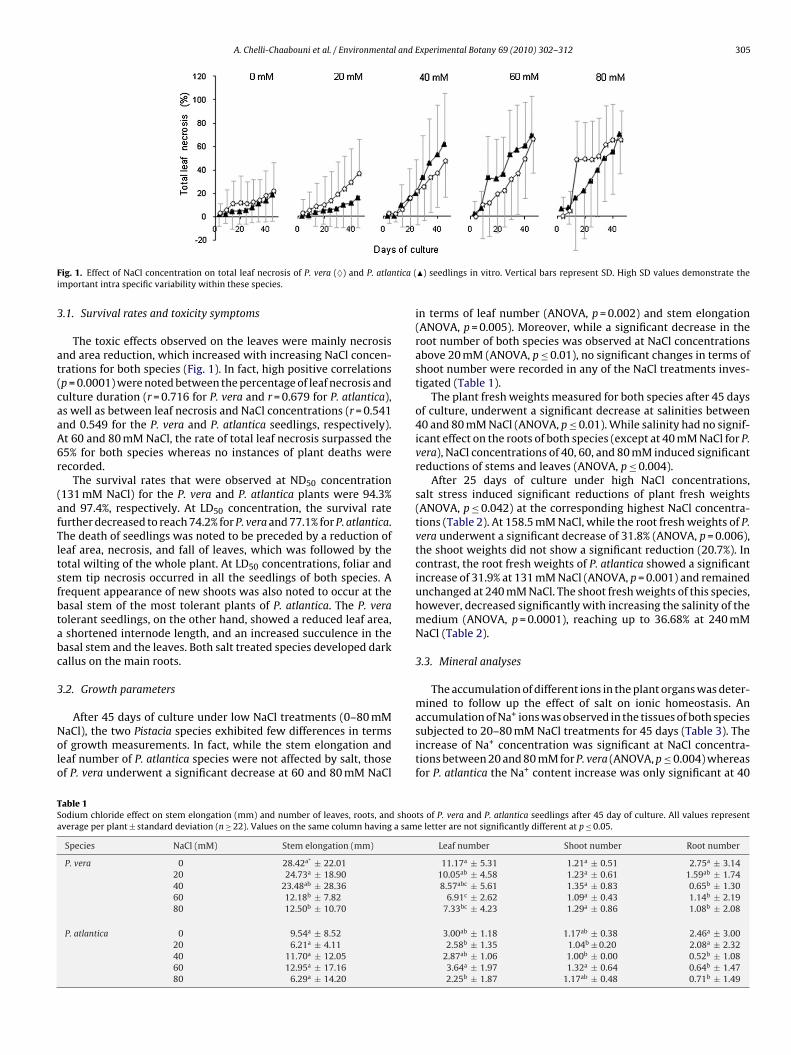
A. Chelli-Chaabouni et al. / Environmental and Experimental Botany 69 (2010) 302–312 305
F ntica (i
3
at(caaA6r
(afTltsfbtabc
3
Nolo
TSa
ig. 1. Effect of NaCl concentration on total leaf necrosis of P. vera (♦) and P. atlamportant intra specific variability within these species.
.1. Survival rates and toxicity symptoms
The toxic effects observed on the leaves were mainly necrosisnd area reduction, which increased with increasing NaCl concen-rations for both species (Fig. 1). In fact, high positive correlationsp = 0.0001) were noted between the percentage of leaf necrosis andulture duration (r = 0.716 for P. vera and r = 0.679 for P. atlantica),s well as between leaf necrosis and NaCl concentrations (r = 0.541nd 0.549 for the P. vera and P. atlantica seedlings, respectively).t 60 and 80 mM NaCl, the rate of total leaf necrosis surpassed the5% for both species whereas no instances of plant deaths wereecorded.
The survival rates that were observed at ND50 concentration131 mM NaCl) for the P. vera and P. atlantica plants were 94.3%nd 97.4%, respectively. At LD50 concentration, the survival rateurther decreased to reach 74.2% for P. vera and 77.1% for P. atlantica.he death of seedlings was noted to be preceded by a reduction ofeaf area, necrosis, and fall of leaves, which was followed by theotal wilting of the whole plant. At LD50 concentrations, foliar andtem tip necrosis occurred in all the seedlings of both species. Arequent appearance of new shoots was also noted to occur at theasal stem of the most tolerant plants of P. atlantica. The P. veraolerant seedlings, on the other hand, showed a reduced leaf area,shortened internode length, and an increased succulence in theasal stem and the leaves. Both salt treated species developed darkallus on the main roots.
.2. Growth parameters
After 45 days of culture under low NaCl treatments (0–80 mMaCl), the two Pistacia species exhibited few differences in termsf growth measurements. In fact, while the stem elongation andeaf number of P. atlantica species were not affected by salt, thosef P. vera underwent a significant decrease at 60 and 80 mM NaCl
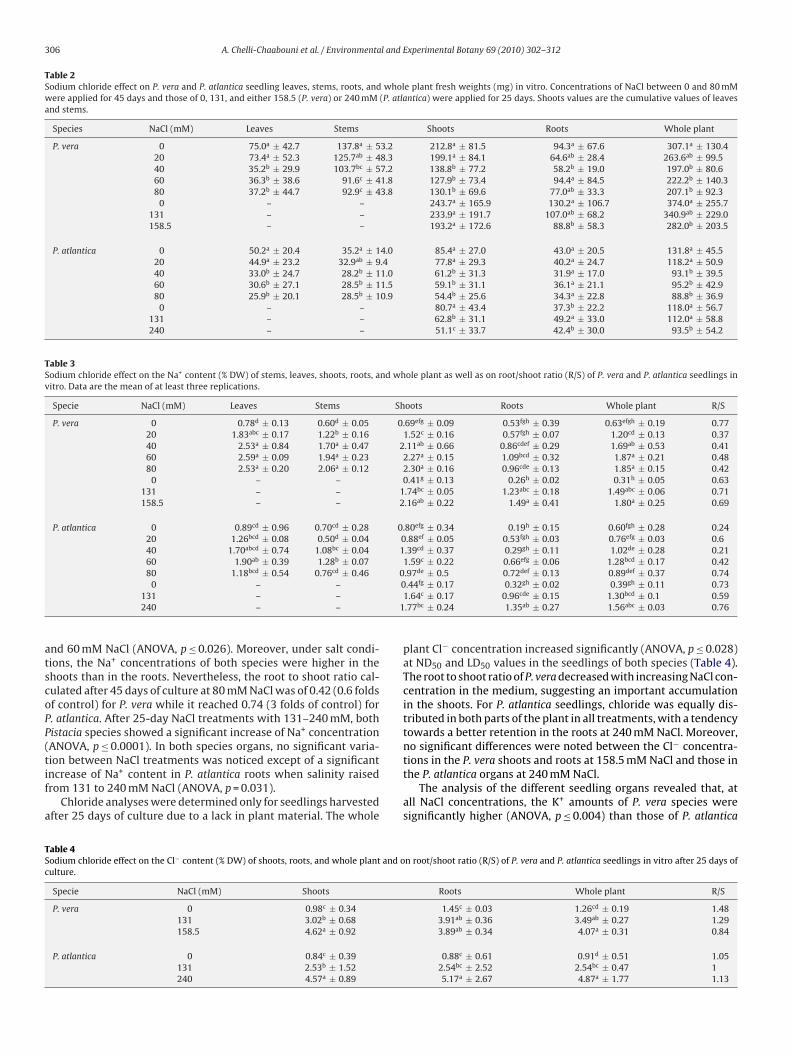
able 1odium chloride effect on stem elongation (mm) and number of leaves, roots, and shooverage per plant ± standard deviation (n ≥ 22). Values on the same column having a sam
Species NaCl (mM) Stem elongation (mm)
P. vera 0 28.42a* ± 22.0120 24.73a ± 18.9040 23.48ab ± 28.3660 12.18b ± 7.8280 12.50b ± 10.70
P. atlantica 0 9.54a ± 8.5220 6.21a ± 4.1140 11.70a ± 12.0560 12.95a ± 17.1680 6.29a ± 14.20
�) seedlings in vitro. Vertical bars represent SD. High SD values demonstrate the
in terms of leaf number (ANOVA, p = 0.002) and stem elongation(ANOVA, p = 0.005). Moreover, while a significant decrease in theroot number of both species was observed at NaCl concentrationsabove 20 mM (ANOVA, p ≤ 0.01), no significant changes in terms ofshoot number were recorded in any of the NaCl treatments inves-tigated (Table 1).
The plant fresh weights measured for both species after 45 daysof culture, underwent a significant decrease at salinities between40 and 80 mM NaCl (ANOVA, p ≤ 0.01). While salinity had no signif-icant effect on the roots of both species (except at 40 mM NaCl for P.vera), NaCl concentrations of 40, 60, and 80 mM induced significantreductions of stems and leaves (ANOVA, p ≤ 0.004).
After 25 days of culture under high NaCl concentrations,salt stress induced significant reductions of plant fresh weights(ANOVA, p ≤ 0.042) at the corresponding highest NaCl concentra-tions (Table 2). At 158.5 mM NaCl, while the root fresh weights of P.vera underwent a significant decrease of 31.8% (ANOVA, p = 0.006),the shoot weights did not show a significant reduction (20.7%). Incontrast, the root fresh weights of P. atlantica showed a significantincrease of 31.9% at 131 mM NaCl (ANOVA, p = 0.001) and remainedunchanged at 240 mM NaCl. The shoot fresh weights of this species,however, decreased significantly with increasing the salinity of themedium (ANOVA, p = 0.0001), reaching up to 36.68% at 240 mMNaCl (Table 2).
3.3. Mineral analyses
The accumulation of different ions in the plant organs was deter-mined to follow up the effect of salt on ionic homeostasis. An
accumulation of Na+ ions was observed in the tissues of both speciessubjected to 20–80 mM NaCl treatments for 45 days (Table 3). Theincrease of Na+ concentration was significant at NaCl concentra-tions between 20 and 80 mM for P. vera (ANOVA, p ≤ 0.004) whereasfor P. atlantica the Na+ content increase was only significant at 40ts of P. vera and P. atlantica seedlings after 45 day of culture. All values represente letter are not significantly different at p ≤ 0.05.
Leaf number Shoot number Root number
11.17a ± 5.31 1.21a ± 0.51 2.75a ± 3.1410.05ab ± 4.58 1.23a ± 0.61 1.59ab ± 1.748.57abc ± 5.61 1.35a ± 0.83 0.65b ± 1.30
6.91c ± 2.62 1.09a ± 0.43 1.14b ± 2.197.33bc ± 4.23 1.29a ± 0.86 1.08b ± 2.08
3.00ab ± 1.18 1.17ab ± 0.38 2.46a ± 3.002.58b ± 1.35 1.04b ± 0.20 2.08a ± 2.32
2.87ab ± 1.06 1.00b ± 0.00 0.52b ± 1.083.64a ± 1.97 1.32a ± 0.64 0.64b ± 1.472.25b ± 1.87 1.17ab ± 0.48 0.71b ± 1.49
306 A. Chelli-Chaabouni et al. / Environmental and Experimental Botany 69 (2010) 302–312
Table 2Sodium chloride effect on P. vera and P. atlantica seedling leaves, stems, roots, and whole plant fresh weights (mg) in vitro. Concentrations of NaCl between 0 and 80 mMwere applied for 45 days and those of 0, 131, and either 158.5 (P. vera) or 240 mM (P. atlantica) were applied for 25 days. Shoots values are the cumulative values of leavesand stems.
Species NaCl (mM) Leaves Stems Shoots Roots Whole plant
P. vera 0 75.0a ± 42.7 137.8a ± 53.2 212.8a ± 81.5 94.3a ± 67.6 307.1a ± 130.420 73.4a ± 52.3 125.7ab ± 48.3 199.1a ± 84.1 64.6ab ± 28.4 263.6ab ± 99.540 35.2b ± 29.9 103.7bc ± 57.2 138.8b ± 77.2 58.2b ± 19.0 197.0b ± 80.660 36.3b ± 38.6 91.6c ± 41.8 127.9b ± 73.4 94.4a ± 84.5 222.2b ± 140.380 37.2b ± 44.7 92.9c ± 43.8 130.1b ± 69.6 77.0ab ± 33.3 207.1b ± 92.3
0 – – 243.7a ± 165.9 130.2a ± 106.7 374.0a ± 255.7131 – – 233.9a ± 191.7 107.0ab ± 68.2 340.9ab ± 229.0158.5 – – 193.2a ± 172.6 88.8b ± 58.3 282.0b ± 203.5
P. atlantica 0 50.2a ± 20.4 35.2a ± 14.0 85.4a ± 27.0 43.0a ± 20.5 131.8a ± 45.520 44.9a ± 23.2 32.9ab ± 9.4 77.8a ± 29.3 40.2a ± 24.7 118.2a ± 50.940 33.0b ± 24.7 28.2b ± 11.0 61.2b ± 31.3 31.9a ± 17.0 93.1b ± 39.560 30.6b ± 27.1 28.5b ± 11.5 59.1b ± 31.1 36.1a ± 21.1 95.2b ± 42.980 25.9b ± 20.1 28.5b ± 10.9 54.4b ± 25.6 34.3a ± 22.8 88.8b ± 36.9
0 – – 80.7a ± 43.4 37.3b ± 22.2 118.0a ± 56.7131 – – 62.8b ± 31.1 49.2a ± 33.0 112.0a ± 58.8240 – – 51.1c ± 33.7 42.4b ± 30.0 93.5b ± 54.2
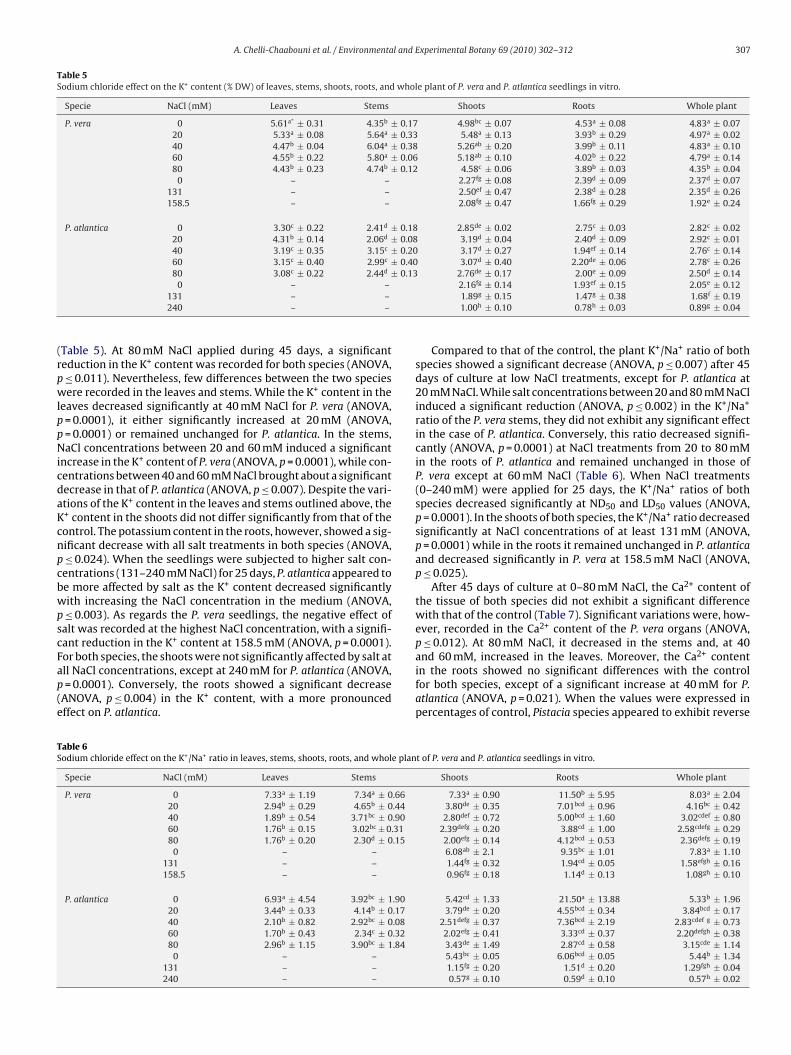
Table 3Sodium chloride effect on the Na+ content (% DW) of stems, leaves, shoots, roots, and whole plant as well as on root/shoot ratio (R/S) of P. vera and P. atlantica seedlings invitro. Data are the mean of at least three replications.
Specie NaCl (mM) Leaves Stems Shoots Roots Whole plant R/S
P. vera 0 0.78d ± 0.13 0.60d ± 0.05 0.69efg ± 0.09 0.53fgh ± 0.39 0.63efgh ± 0.19 0.7720 1.83abc ± 0.17 1.22b ± 0.16 1.52c ± 0.16 0.57fgh ± 0.07 1.20cd ± 0.13 0.3740 2.53a ± 0.84 1.70a ± 0.47 2.11ab ± 0.66 0.86cdef ± 0.29 1.69ab ± 0.53 0.4160 2.59a ± 0.09 1.94a ± 0.23 2.27a ± 0.15 1.09bcd ± 0.32 1.87a ± 0.21 0.4880 2.53a ± 0.20 2.06a ± 0.12 2.30a ± 0.16 0.96cde ± 0.13 1.85a ± 0.15 0.42
0 – – 0.41g ± 0.13 0.26h ± 0.02 0.31h ± 0.05 0.63131 – – 1.74bc ± 0.05 1.23abc ± 0.18 1.49abc ± 0.06 0.71158.5 – – 2.16ab ± 0.22 1.49a ± 0.41 1.80a ± 0.25 0.69
P. atlantica 0 0.89cd ± 0.96 0.70cd ± 0.28 0.80efg ± 0.34 0.19h ± 0.15 0.60fgh ± 0.28 0.2420 1.26bcd ± 0.08 0.50d ± 0.04 0.88ef ± 0.05 0.53fgh ± 0.03 0.76efg ± 0.03 0.640 1.70abcd ± 0.74 1.08bc ± 0.04 1.39cd ± 0.37 0.29gh ± 0.11 1.02de ± 0.28 0.2160 1.90ab ± 0.39 1.28b ± 0.07 1.59c ± 0.22 0.66efg ± 0.06 1.28bcd ± 0.17 0.4280 1.18bcd ± 0.54 0.76cd ± 0.46 0.97de ± 0.5 0.72def ± 0.13 0.89def ± 0.37 0.74
0
1
atscoPP(tif
a
TSc
0 – –131 – –240 – –
nd 60 mM NaCl (ANOVA, p ≤ 0.026). Moreover, under salt condi-ions, the Na+ concentrations of both species were higher in thehoots than in the roots. Nevertheless, the root to shoot ratio cal-ulated after 45 days of culture at 80 mM NaCl was of 0.42 (0.6 foldsf control) for P. vera while it reached 0.74 (3 folds of control) for. atlantica. After 25-day NaCl treatments with 131–240 mM, bothistacia species showed a significant increase of Na+ concentrationANOVA, p ≤ 0.0001). In both species organs, no significant varia-
ion between NaCl treatments was noticed except of a significantncrease of Na+ content in P. atlantica roots when salinity raisedrom 131 to 240 mM NaCl (ANOVA, p = 0.031).Chloride analyses were determined only for seedlings harvestedfter 25 days of culture due to a lack in plant material. The whole
able 4odium chloride effect on the Cl− content (% DW) of shoots, roots, and whole plant and oulture.
Specie NaCl (mM) Shoots
P. vera 0 0.98c ± 0.34131 3.02b ± 0.68158.5 4.62a ± 0.92
P. atlantica 0 0.84c ± 0.39131 2.53b ± 1.52240 4.57a ± 0.89
.44fg ± 0.17 0.32gh ± 0.02 0.39gh ± 0.11 0.731.64c ± 0.17 0.96cde ± 0.15 1.30bcd ± 0.1 0.59.77bc ± 0.24 1.35ab ± 0.27 1.56abc ± 0.03 0.76
plant Cl− concentration increased significantly (ANOVA, p ≤ 0.028)at ND50 and LD50 values in the seedlings of both species (Table 4).The root to shoot ratio of P. vera decreased with increasing NaCl con-centration in the medium, suggesting an important accumulationin the shoots. For P. atlantica seedlings, chloride was equally dis-tributed in both parts of the plant in all treatments, with a tendencytowards a better retention in the roots at 240 mM NaCl. Moreover,no significant differences were noted between the Cl− concentra-
tions in the P. vera shoots and roots at 158.5 mM NaCl and those inthe P. atlantica organs at 240 mM NaCl.The analysis of the different seedling organs revealed that, atall NaCl concentrations, the K+ amounts of P. vera species weresignificantly higher (ANOVA, p ≤ 0.004) than those of P. atlantica
n root/shoot ratio (R/S) of P. vera and P. atlantica seedlings in vitro after 25 days of
Roots Whole plant R/S
1.45c ± 0.03 1.26cd ± 0.19 1.483.91ab ± 0.36 3.49ab ± 0.27 1.293.89ab ± 0.34 4.07a ± 0.31 0.84
0.88c ± 0.61 0.91d ± 0.51 1.052.54bc ± 2.52 2.54bc ± 0.47 15.17a ± 2.67 4.87a ± 1.77 1.13
A. Chelli-Chaabouni et al. / Environmental and Experimental Botany 69 (2010) 302–312 307
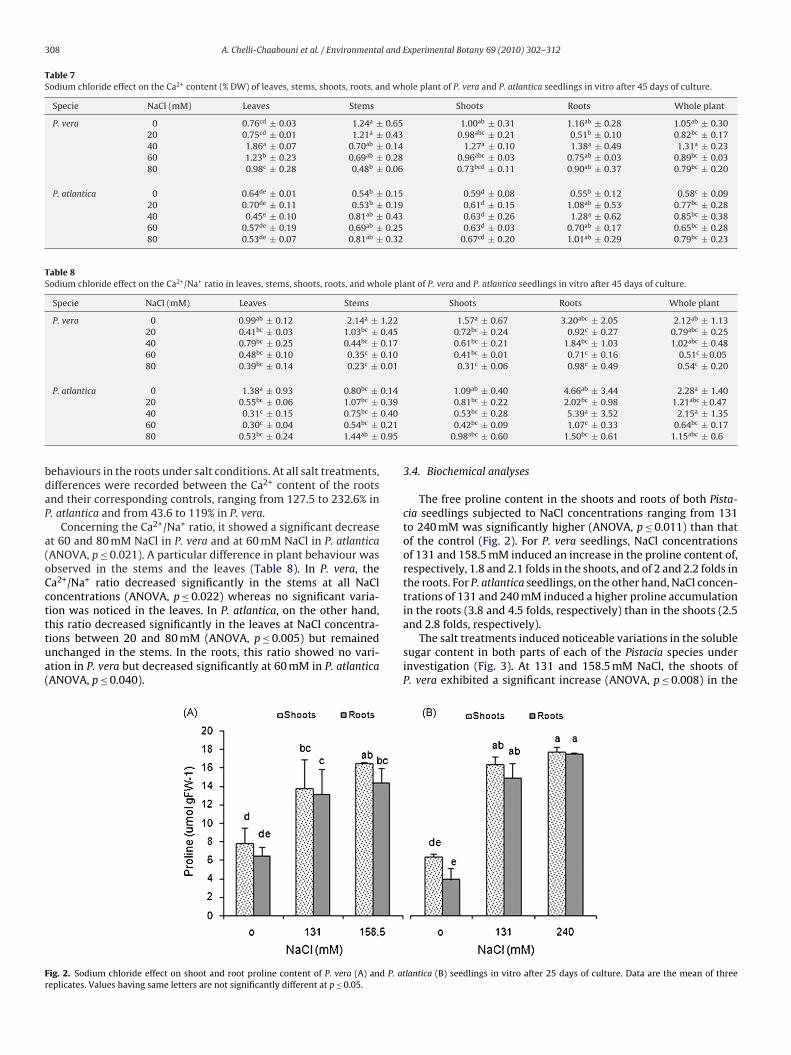
Table 5Sodium chloride effect on the K+ content (% DW) of leaves, stems, shoots, roots, and whole plant of P. vera and P. atlantica seedlings in vitro.
Specie NaCl (mM) Leaves Stems Shoots Roots Whole plant
P. vera 0 5.61a* ± 0.31 4.35b ± 0.17 4.98bc ± 0.07 4.53a ± 0.08 4.83a ± 0.0720 5.33a ± 0.08 5.64a ± 0.33 5.48a ± 0.13 3.93b ± 0.29 4.97a ± 0.0240 4.47b ± 0.04 6.04a ± 0.38 5.26ab ± 0.20 3.99b ± 0.11 4.83a ± 0.1060 4.55b ± 0.22 5.80a ± 0.06 5.18ab ± 0.10 4.02b ± 0.22 4.79a ± 0.1480 4.43b ± 0.23 4.74b ± 0.12 4.58c ± 0.06 3.89b ± 0.03 4.35b ± 0.04
0 – – 2.27fg ± 0.08 2.39d ± 0.09 2.37d ± 0.07131 – – 2.50ef ± 0.47 2.38d ± 0.28 2.35d ± 0.26158.5 – – 2.08fg ± 0.47 1.66fg ± 0.29 1.92e ± 0.24
P. atlantica 0 3.30c ± 0.22 2.41d ± 0.18 2.85de ± 0.02 2.75c ± 0.03 2.82c ± 0.0220 4.31b ± 0.14 2.06d ± 0.08 3.19d ± 0.04 2.40d ± 0.09 2.92c ± 0.0140 3.19c ± 0.35 3.15c ± 0.20 3.17d ± 0.27 1.94ef ± 0.14 2.76c ± 0.1460 3.15c ± 0.40 2.99c ± 0.40 3.07d ± 0.40 2.20de ± 0.06 2.78c ± 0.2680 3.08c ± 0.22 2.44d ± 0.13 2.76de ± 0.17 2.00e ± 0.09 2.50d ± 0.14
(rpwlppNicdaKcnpcbwpscFap(e
TS
0 – –131 – –240 – –
Table 5). At 80 mM NaCl applied during 45 days, a significanteduction in the K+ content was recorded for both species (ANOVA,≤ 0.011). Nevertheless, few differences between the two speciesere recorded in the leaves and stems. While the K+ content in the
eaves decreased significantly at 40 mM NaCl for P. vera (ANOVA,= 0.0001), it either significantly increased at 20 mM (ANOVA,= 0.0001) or remained unchanged for P. atlantica. In the stems,aCl concentrations between 20 and 60 mM induced a significant
ncrease in the K+ content of P. vera (ANOVA, p = 0.0001), while con-entrations between 40 and 60 mM NaCl brought about a significantecrease in that of P. atlantica (ANOVA, p ≤ 0.007). Despite the vari-tions of the K+ content in the leaves and stems outlined above, the+ content in the shoots did not differ significantly from that of theontrol. The potassium content in the roots, however, showed a sig-ificant decrease with all salt treatments in both species (ANOVA,≤ 0.024). When the seedlings were subjected to higher salt con-entrations (131–240 mM NaCl) for 25 days, P. atlantica appeared toe more affected by salt as the K+ content decreased significantlyith increasing the NaCl concentration in the medium (ANOVA,≤ 0.003). As regards the P. vera seedlings, the negative effect of
alt was recorded at the highest NaCl concentration, with a signifi-ant reduction in the K+ content at 158.5 mM (ANOVA, p = 0.0001).
or both species, the shoots were not significantly affected by salt atll NaCl concentrations, except at 240 mM for P. atlantica (ANOVA,= 0.0001). Conversely, the roots showed a significant decreaseANOVA, p ≤ 0.004) in the K+ content, with a more pronouncedffect on P. atlantica.
able 6odium chloride effect on the K+/Na+ ratio in leaves, stems, shoots, roots, and whole plan
Specie NaCl (mM) Leaves Stems
P. vera 0 7.33a ± 1.19 7.34a ± 0.6620 2.94b ± 0.29 4.65b ± 0.4440 1.89b ± 0.54 3.71bc ± 0.9060 1.76b ± 0.15 3.02bc ± 0.3180 1.76b ± 0.20 2.30d ± 0.15
0 – –131 – –158.5 – –
P. atlantica 0 6.93a ± 4.54 3.92bc ± 1.9020 3.44b ± 0.33 4.14b ± 0.1740 2.10b ± 0.82 2.92bc ± 0.0860 1.70b ± 0.43 2.34c ± 0.3280 2.96b ± 1.15 3.90bc ± 1.84
0 – –131 – –240 – –
2.16fg ± 0.14 1.93ef ± 0.15 2.05e ± 0.121.89g ± 0.15 1.47g ± 0.38 1.68f ± 0.191.00h ± 0.10 0.78h ± 0.03 0.89g ± 0.04
Compared to that of the control, the plant K+/Na+ ratio of bothspecies showed a significant decrease (ANOVA, p ≤ 0.007) after 45days of culture at low NaCl treatments, except for P. atlantica at20 mM NaCl. While salt concentrations between 20 and 80 mM NaClinduced a significant reduction (ANOVA, p ≤ 0.002) in the K+/Na+
ratio of the P. vera stems, they did not exhibit any significant effectin the case of P. atlantica. Conversely, this ratio decreased signifi-cantly (ANOVA, p = 0.0001) at NaCl treatments from 20 to 80 mMin the roots of P. atlantica and remained unchanged in those ofP. vera except at 60 mM NaCl (Table 6). When NaCl treatments(0–240 mM) were applied for 25 days, the K+/Na+ ratios of bothspecies decreased significantly at ND50 and LD50 values (ANOVA,p = 0.0001). In the shoots of both species, the K+/Na+ ratio decreasedsignificantly at NaCl concentrations of at least 131 mM (ANOVA,p = 0.0001) while in the roots it remained unchanged in P. atlanticaand decreased significantly in P. vera at 158.5 mM NaCl (ANOVA,p ≤ 0.025).
After 45 days of culture at 0–80 mM NaCl, the Ca2+ content ofthe tissue of both species did not exhibit a significant differencewith that of the control (Table 7). Significant variations were, how-ever, recorded in the Ca2+ content of the P. vera organs (ANOVA,p ≤ 0.012). At 80 mM NaCl, it decreased in the stems and, at 40
2+
and 60 mM, increased in the leaves. Moreover, the Ca contentin the roots showed no significant differences with the controlfor both species, except of a significant increase at 40 mM for P.atlantica (ANOVA, p = 0.021). When the values were expressed inpercentages of control, Pistacia species appeared to exhibit reverset of P. vera and P. atlantica seedlings in vitro.
Shoots Roots Whole plant
7.33a ± 0.90 11.50b ± 5.95 8.03a ± 2.043.80de ± 0.35 7.01bcd ± 0.96 4.16bc ± 0.422.80def ± 0.72 5.00bcd ± 1.60 3.02cdef ± 0.80
2.39defg ± 0.20 3.88cd ± 1.00 2.58cdefg ± 0.292.00efg ± 0.14 4.12bcd ± 0.53 2.36defg ± 0.196.08ab ± 2.1 9.35bc ± 1.01 7.83a ± 1.101.44fg ± 0.32 1.94cd ± 0.05 1.58efgh ± 0.160.96fg ± 0.18 1.14d ± 0.13 1.08gh ± 0.10
5.42cd ± 1.33 21.50a ± 13.88 5.33b ± 1.963.79de ± 0.20 4.55bcd ± 0.34 3.84bcd ± 0.17
2.51defg ± 0.37 7.36bcd ± 2.19 2.83cdef g ± 0.732.02efg ± 0.41 3.33cd ± 0.37 2.20defgh ± 0.383.43de ± 1.49 2.87cd ± 0.58 3.15cde ± 1.145.43bc ± 0.05 6.06bcd ± 0.05 5.44b ± 1.341.15fg ± 0.20 1.51d ± 0.20 1.29fgh ± 0.040.57g ± 0.10 0.59d ± 0.10 0.57h ± 0.02
308 A. Chelli-Chaabouni et al. / Environmental and Experimental Botany 69 (2010) 302–312
Table 7Sodium chloride effect on the Ca2+ content (% DW) of leaves, stems, shoots, roots, and whole plant of P. vera and P. atlantica seedlings in vitro after 45 days of culture.
Specie NaCl (mM) Leaves Stems Shoots Roots Whole plant
P. vera 0 0.76cd ± 0.03 1.24a ± 0.65 1.00ab ± 0.31 1.16ab ± 0.28 1.05ab ± 0.3020 0.75cd ± 0.01 1.21a ± 0.43 0.98abc ± 0.21 0.51b ± 0.10 0.82bc ± 0.1740 1.86a ± 0.07 0.70ab ± 0.14 1.27a ± 0.10 1.38a ± 0.49 1.31a ± 0.2360 1.23b ± 0.23 0.69ab ± 0.28 0.96abc ± 0.03 0.75ab ± 0.03 0.89bc ± 0.0380 0.98c ± 0.28 0.48b ± 0.06 0.73bcd ± 0.11 0.90ab ± 0.37 0.79bc ± 0.20
P. atlantica 0 0.64de ± 0.01 0.54b ± 0.15 0.59d ± 0.08 0.55b ± 0.12 0.58c ± 0.0920 0.70de ± 0.11 0.53b ± 0.19 0.61d ± 0.15 1.08ab ± 0.53 0.77bc ± 0.2840 0.45e ± 0.10 0.81ab ± 0.43 0.63d ± 0.26 1.28a ± 0.62 0.85bc ± 0.3860 0.57de ± 0.19 0.69ab ± 0.25 0.63d ± 0.03 0.70ab ± 0.17 0.65bc ± 0.2880 0.53de ± 0.07 0.81ab ± 0.32 0.67cd ± 0.20 1.01ab ± 0.29 0.79bc ± 0.23
Table 8Sodium chloride effect on the Ca2+/Na+ ratio in leaves, stems, shoots, roots, and whole plant of P. vera and P. atlantica seedlings in vitro after 45 days of culture.
Specie NaCl (mM) Leaves Stems Shoots Roots Whole plant
P. vera 0 0.99ab ± 0.12 2.14a ± 1.22 1.57a ± 0.67 3.20abc ± 2.05 2.12ab ± 1.1320 0.41bc ± 0.03 1.03bc ± 0.45 0.72bc ± 0.24 0.92c ± 0.27 0.79abc ± 0.2540 0.79bc ± 0.25 0.44bc ± 0.17 0.61bc ± 0.21 1.84bc ± 1.03 1.02abc ± 0.4860 0.48bc ± 0.10 0.35c ± 0.10 0.41bc ± 0.01 0.71c ± 0.16 0.51c ± 0.0580 0.39bc ± 0.14 0.23c ± 0.01 0.31c ± 0.06 0.98c ± 0.49 0.54c ± 0.20
P. atlantica 0 1.38a ± 0.93 0.80bc ± 0.14 1.09ab ± 0.40 4.66ab ± 3.44 2.28a ± 1.40bc bc 0.39 bc bc abc
0.400.210.95
bdaP
a(oCctttua(
Fr
20 0.55 ± 0.06 1.07 ±40 0.31c ± 0.15 0.75bc ±60 0.30c ± 0.04 0.54bc ±80 0.53bc ± 0.24 1.44ab ±
ehaviours in the roots under salt conditions. At all salt treatments,ifferences were recorded between the Ca2+ content of the rootsnd their corresponding controls, ranging from 127.5 to 232.6% in. atlantica and from 43.6 to 119% in P. vera.
Concerning the Ca2+/Na+ ratio, it showed a significant decreaset 60 and 80 mM NaCl in P. vera and at 60 mM NaCl in P. atlanticaANOVA, p ≤ 0.021). A particular difference in plant behaviour wasbserved in the stems and the leaves (Table 8). In P. vera, thea2+/Na+ ratio decreased significantly in the stems at all NaCloncentrations (ANOVA, p ≤ 0.022) whereas no significant varia-ion was noticed in the leaves. In P. atlantica, on the other hand,
his ratio decreased significantly in the leaves at NaCl concentra-ions between 20 and 80 mM (ANOVA, p ≤ 0.005) but remainednchanged in the stems. In the roots, this ratio showed no vari-tion in P. vera but decreased significantly at 60 mM in P. atlanticaANOVA, p ≤ 0.040).ig. 2. Sodium chloride effect on shoot and root proline content of P. vera (A) and P. ateplicates. Values having same letters are not significantly different at p ≤ 0.05.
0.81 ± 0.22 2.02 ± 0.98 1.21 ± 0.470.53bc ± 0.28 5.39a ± 3.52 2.15a ± 1.350.42bc ± 0.09 1.07c ± 0.33 0.64bc ± 0.17
0.98abc ± 0.60 1.50bc ± 0.61 1.15abc ± 0.6
3.4. Biochemical analyses
The free proline content in the shoots and roots of both Pista-cia seedlings subjected to NaCl concentrations ranging from 131to 240 mM was significantly higher (ANOVA, p ≤ 0.011) than thatof the control (Fig. 2). For P. vera seedlings, NaCl concentrationsof 131 and 158.5 mM induced an increase in the proline content of,respectively, 1.8 and 2.1 folds in the shoots, and of 2 and 2.2 folds inthe roots. For P. atlantica seedlings, on the other hand, NaCl concen-trations of 131 and 240 mM induced a higher proline accumulationin the roots (3.8 and 4.5 folds, respectively) than in the shoots (2.5
and 2.8 folds, respectively).The salt treatments induced noticeable variations in the solublesugar content in both parts of each of the Pistacia species underinvestigation (Fig. 3). At 131 and 158.5 mM NaCl, the shoots ofP. vera exhibited a significant increase (ANOVA, p ≤ 0.008) in the
lantica (B) seedlings in vitro after 25 days of culture. Data are the mean of three
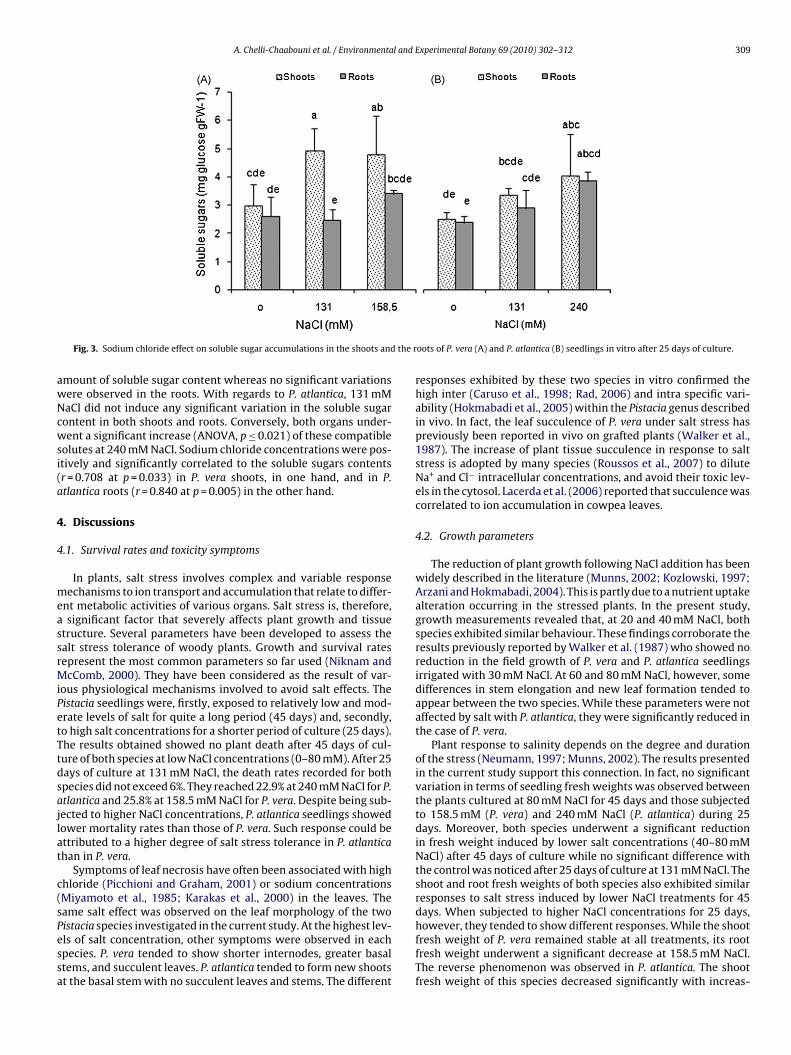
A. Chelli-Chaabouni et al. / Environmental and Experimental Botany 69 (2010) 302–312 309
the r
awNcwsi(a
4
4
meassrMiPetTtdsajlat
c(sPessa
Fig. 3. Sodium chloride effect on soluble sugar accumulations in the shoots and
mount of soluble sugar content whereas no significant variationsere observed in the roots. With regards to P. atlantica, 131 mMaCl did not induce any significant variation in the soluble sugarontent in both shoots and roots. Conversely, both organs under-ent a significant increase (ANOVA, p ≤ 0.021) of these compatible
olutes at 240 mM NaCl. Sodium chloride concentrations were pos-tively and significantly correlated to the soluble sugars contentsr = 0.708 at p = 0.033) in P. vera shoots, in one hand, and in P.tlantica roots (r = 0.840 at p = 0.005) in the other hand.
. Discussions
.1. Survival rates and toxicity symptoms
In plants, salt stress involves complex and variable responseechanisms to ion transport and accumulation that relate to differ-
nt metabolic activities of various organs. Salt stress is, therefore,significant factor that severely affects plant growth and tissue
tructure. Several parameters have been developed to assess thealt stress tolerance of woody plants. Growth and survival ratesepresent the most common parameters so far used (Niknam andcComb, 2000). They have been considered as the result of var-
ous physiological mechanisms involved to avoid salt effects. Theistacia seedlings were, firstly, exposed to relatively low and mod-rate levels of salt for quite a long period (45 days) and, secondly,o high salt concentrations for a shorter period of culture (25 days).he results obtained showed no plant death after 45 days of cul-ure of both species at low NaCl concentrations (0–80 mM). After 25ays of culture at 131 mM NaCl, the death rates recorded for bothpecies did not exceed 6%. They reached 22.9% at 240 mM NaCl for P.tlantica and 25.8% at 158.5 mM NaCl for P. vera. Despite being sub-ected to higher NaCl concentrations, P. atlantica seedlings showedower mortality rates than those of P. vera. Such response could bettributed to a higher degree of salt stress tolerance in P. atlanticahan in P. vera.
Symptoms of leaf necrosis have often been associated with highhloride (Picchioni and Graham, 2001) or sodium concentrationsMiyamoto et al., 1985; Karakas et al., 2000) in the leaves. Theame salt effect was observed on the leaf morphology of the two
istacia species investigated in the current study. At the highest lev-ls of salt concentration, other symptoms were observed in eachpecies. P. vera tended to show shorter internodes, greater basaltems, and succulent leaves. P. atlantica tended to form new shootst the basal stem with no succulent leaves and stems. The differentoots of P. vera (A) and P. atlantica (B) seedlings in vitro after 25 days of culture.
responses exhibited by these two species in vitro confirmed thehigh inter (Caruso et al., 1998; Rad, 2006) and intra specific vari-ability (Hokmabadi et al., 2005) within the Pistacia genus describedin vivo. In fact, the leaf succulence of P. vera under salt stress haspreviously been reported in vivo on grafted plants (Walker et al.,1987). The increase of plant tissue succulence in response to saltstress is adopted by many species (Roussos et al., 2007) to diluteNa+ and Cl− intracellular concentrations, and avoid their toxic lev-els in the cytosol. Lacerda et al. (2006) reported that succulence wascorrelated to ion accumulation in cowpea leaves.
4.2. Growth parameters
The reduction of plant growth following NaCl addition has beenwidely described in the literature (Munns, 2002; Kozlowski, 1997;Arzani and Hokmabadi, 2004). This is partly due to a nutrient uptakealteration occurring in the stressed plants. In the present study,growth measurements revealed that, at 20 and 40 mM NaCl, bothspecies exhibited similar behaviour. These findings corroborate theresults previously reported by Walker et al. (1987) who showed noreduction in the field growth of P. vera and P. atlantica seedlingsirrigated with 30 mM NaCl. At 60 and 80 mM NaCl, however, somedifferences in stem elongation and new leaf formation tended toappear between the two species. While these parameters were notaffected by salt with P. atlantica, they were significantly reduced inthe case of P. vera.
Plant response to salinity depends on the degree and durationof the stress (Neumann, 1997; Munns, 2002). The results presentedin the current study support this connection. In fact, no significantvariation in terms of seedling fresh weights was observed betweenthe plants cultured at 80 mM NaCl for 45 days and those subjectedto 158.5 mM (P. vera) and 240 mM NaCl (P. atlantica) during 25days. Moreover, both species underwent a significant reductionin fresh weight induced by lower salt concentrations (40–80 mMNaCl) after 45 days of culture while no significant difference withthe control was noticed after 25 days of culture at 131 mM NaCl. Theshoot and root fresh weights of both species also exhibited similarresponses to salt stress induced by lower NaCl treatments for 45days. When subjected to higher NaCl concentrations for 25 days,
however, they tended to show different responses. While the shootfresh weight of P. vera remained stable at all treatments, its rootfresh weight underwent a significant decrease at 158.5 mM NaCl.The reverse phenomenon was observed in P. atlantica. The shootfresh weight of this species decreased significantly with increas-
3 l and E
is2hhahtiaaipfwos
4
idc1NtoiMrlwttctweaitra
bNdsatfaoawc(
ttpma1c
10 A. Chelli-Chaabouni et al. / Environmenta
ng NaCl in the medium while its root fresh weight underwent aignificant increase at 131 mM NaCl and showed no variation at40 mM. At 131 mM NaCl, the root to shoot ratio of P. atlantica wasigher by up to 1.7 folds than that of P. vera. These findings suggest aigher water uptake rate in P. vera NaCl treated shoots. This uptakellowed the ion dilution to avoid the toxic level in the cytosol. Thisypothesis, also suggested by Behboudian et al. (1986), is consis-ent with the morphological symptoms of hypehydricity observedn the P. vera basal shoots. Furthermore, the significant increasend absence of variation in the root fresh weights of P. atlanticat 131 and 240 mM NaCl, respectively, strongly suggest that saltnduced the development of woody roots in P. atlantica, which wasreviously reported by Walker et al. (1987). Similarly, while theresh weights of tolerant clones of in vitro cultured jojoba shootsere previously reported to increase or remain unaffected, those
f sensitive clones were noted to decrease when cultured on mediaupplemented with different chloride salts (Mills et al., 2001).
.3. Mineral analyses
Under salt conditions, the Na+ content of both Pistacia speciesn the shoots was higher than in the roots. These findings are inisagreement with results previously reported on older seedlingsultivated in the field (Walker et al., 1987; Picchioni and Miyamoto,990) and nursery conditions (Chelli Chaabouni et al., 2008) wherea+ concentration was described to be higher in the roots than in
he shoots. The transport and accumulation of Na+ in the leavesften characterize tolerant includer species that compartmental-ze toxic ions in the vacuoles (Zid and Grignon, 1991; Niknam and
cComb, 2000; Ashraf, 2004). The values recorded for root to shootatios reflected the includer trait of both species as they accumu-ated high amounts of Na+ ions in the leaves. Under salt conditions,
hile P. atlantica seemed to retain more Na+ in the roots than inhe shoots, P. vera allowed their transport in the shoots. The reten-ion of Na+ in the roots of P. atlantica helped to avoid reaching highoncentrations in the leaves that may affect plant metabolism. Inhis respect, Tabatabaei (2006) previously reported similar resultsith olive tree cultivars subjected to high salt stress where salt tol-
rance in olive trees was hypothesized to be associated with thebility to reduce the uptake and/or transport of saline ions. Sim-larly, other previous reports suggest that salt tolerance is likelyo be attributed to the control of salt translocation to the leavesather than to the absorption of salt (Chartzoulakis, 2005; Ghars etl., 2008).
Furthermore, the findings from chloride analyses showed thatoth Pistacia seedlings did not exhibit significant variations at allaCl concentrations. The root to shoot ratio was also noted toecrease with increasing salinity in P. vera but to remain relativelytable for P. atlantica. These results may be attributed to a greaterbility displayed by P. atlantica to control chloride translocationo the shoots using a more effective salt tolerance mechanism. Inact, older P. atlantica seedlings that were subjected to 150 mM Cl−
nd 90 mM Na+ were previously reported to retain higher amountsf Na+ and Cl− in feeder roots (Walker et al., 1987). Similarly, P.tlantica and the hybrid PGII (P. atlantica × P. integerrima) irrigatedith a saline solution were described to exhibit higher chloride
oncentrations in the roots than that retained by P. terebinthusPicchioni and Miyamoto, 1990).
Potassium ion is the major cation within plants which coun-erbalances the negative charge of anions. The K+ ion stabiliseshe pH, osmotic potential, and turgor pressure within cells. It also
lays a crucial role in the activation of the enzymes involved in theetabolism and synthesis of proteins and carbohydrates, as wells in the regulation of stomata movement and tropism (George,993). Moreover, K+ ions contribute to the osmotic adjustment ofells in salt stressed plants. In this respect, the control treatments
xperimental Botany 69 (2010) 302–312
presented in the current study revealed that P. vera seedlings main-tained higher K+ and Ca2+ concentrations than those of P. atlantica.This latter species appeared to display a lower nutrient uptake thanP. vera, which may explain the faster growth rate exhibited by P.vera at the very early stage of development. After 45 days of cultureat 0–60 mM NaCl, the K+ content of both Pistacia seedlings remainedsimilar to that of the control. A significant decrease was observedat 80 mM. This reduction in ion uptake, however, did not exceed9.9% in P. vera and 11.3% in P. atlantica, indicating that potassiumnutrition was not greatly affected by NaCl treatments. These find-ings corroborate previous reports that classified Pistacia species assalt tolerant when compared to other fruit trees (Behboudian et al.,1986; Picchioni et al., 1990; Tattini et al., 2006, 2008). This reductionwas also observed to be more pronounced in the leaves (21%) thanin the roots (14%) of P. vera. The reverse was, however, observedwith P. atlantica (7% in leaves and 27.3% in roots). When high salttreatments were administered for 25 days, the differences in theK+ uptake of these species became more pronounced. A higher K+
content reduction was noticed in P. atlantica than in P. vera organsparticularly at 240 mM NaCl where the reduction exceeded 53%.The Na–K antagonistic effect in the roots might have contributedto the decrease in the K+ uptake and its translocation into theplant.
At the whole plant level, the K+/Na+ ratio was maintained above2 in all the treatments applied to both species for 45 days. Seeingthat this ability to maintain high K+/Na+ ratio in the leaves was pre-viously observed in salt tolerant species (Maathuis and Amtmann,1999; Chartzoulakis et al., 2002; Dasgan et al., 2002; Meena et al.,2003; Türkan and Demiral, 2009), the Pistacia species tested can beconsidered as salt tolerant under the in vitro conditions presentedin the current work. This ratio, however, felt down after 25 daysof high salt concentrations (DL50), reaching a minimal value of 1(Maathuis and Amtmann, 1999) in P. vera or even lower values inP. atlantica. As suggested by Greenway and Munns (1980), thesevalues of K+/Na+ ratio correspond to the limit of optimal proteinsynthesis efficiency. The decrease described above can, therefore,be attributed to a failure in the K–Na selectivity mechanism causedby the accumulation of Na+. Accordingly, at NaCl concentrationsabove 131 mM, the in vitro tested Pistacia species seem to havereached the threshold of salt tolerance.
Calcium is known to play a key role in maintaining the integrityof the plasma membrane in the root cells, thus limiting the toxiceffect of Na+ (Rengel, 1992; Gucci and Tattini, 1997). It acts as asecondary messenger in the regulation of signal transduction path-ways for the response to abiotic stress and in the promotion ofK+/Na+ selectivity (Rengel, 1992; Maathuis and Amtmann, 1999).At the end of the culture period, and at NaCl concentrations from0 to 80 mM, the Ca2+ content remained unchanged in both Pistaciaspecies, except for a significant decrease at 80 mM NaCl in P. vera.Despite their success in maintaining constant levels of tissue Ca2+,both species differed in the partitioning of this ion in their differ-ent organs. At all salt treatments, the Ca2+ concentration of P. verawas maintained constant or significantly higher than that of thecontrol in the leaves. It also stabilized or decreased significantly inthe stems and the roots. These results are in agreement with thosepreviously reported by Hokmabadi et al. (2005). The increase ofCa2+ in the leaves was also previously described for other species(Ramoliya et al., 2004; De Pascale et al., 2007). The increased levelof calcium in the leaves of P. vera may be explained by the translo-cation of Ca2+ from the roots to the leaves so as to alleviate the salttoxicity in this organ (Cramer et al., 1987). The important decrease
of Ca2+ in the shoots, on the other hand, was reported as a salt sen-sitivity trait in Phalaris arundinacea and Cucumis sativus (Unno etal., 2002). Under NaCl treatments, the Ca2+ content of P. atlanticawas not significantly different from that of the control, except fora significant increase in the roots at 40 mM NaCl.
l and
0d6ci
4
twbiTWt(aewa
di(tiw2atootetst(toi(eidacc
5
isriaomhcat
A. Chelli-Chaabouni et al. / Environmenta
The data obtained after the subjection of the seedlings to–80 mM NaCl for 45 days showed that the Ca2+/Na+ ratioecreased significantly at 60 and 80 mM NaCl in P. vera and at0 mM NaCl in P. atlantica. In fact, high Ca2+/Na+ ratios have beenonsidered as characteristic traits when screening for salt tolerancen plants (Dasgan et al., 2002; Yildiz et al., 2008).
.4. Biochemical analyses
Two compatible solutes, proline and soluble sugars, known forheir osmoprotectant activity in plants subjected to salt stress,ere investigated. After 25 days of culture, salinity levels ranging
etween 0 and 240 mM NaCl were noted to induce a significantncrease in the free proline content of both Pistacia species in vitro.heses results are in agreement with those previously reported byalker et al. (1988) for P. vera cv. Kerman in glasshouse condi-
ions. Similar results were also obtained under in vitro conditionsHarinasut et al., 2000). Besides, Hokmabadi et al. (2005) observedhigh accumulation of leaf proline in the most tolerant P. vera vari-ty. Similarly, in vitro high accumulation of proline under salinityas previously reported for the salt tolerant Citrus variety (Ferreira
nd Lima-Costa, 2006).The increase or decrease of soluble sugar content have also been
escribed as responses to salinity in many plant species eithern vivo (Meena et al., 2003; Yang and Guizhu, 2007) or in vitroWatanabe et al., 2000; Rochdi et al., 2003; Rejskovà et al., 2007). Inhe present study, the total soluble carbohydrate content of P. verancreased significantly in the shoots at 131 and 158.5 mM NaCl,
hile this content increase was noticed at NaCl concentrations of40 mM in P. atlantica shoots and roots. A positive correlation waslso recorded between the soluble sugar content and NaCl concen-ration in the roots for P. atlantica and in the shoots for P. vera. Thesergans showed higher accumulations of Na+ and greater reductionsf K+ ions, which provide further support to the hypothesis thathese compatible solutes play a major role in osmoregulation (Haret al., 1998; Parida and Das, 2005). It is worth noting, however,hat the variation of total carbohydrate content alone may not beufficient to evaluate the effect of salt stress on carbohydrate sta-us. In fact, the endogenous variation of particular carbohydratese.g. mannitol, sucrose, and glucose) may provide further informa-ion about the specific carbohydrates involved in the salt tolerancef these species and could lead to the determination of specificndicators of salt tolerance. The results described by Rejskovà et al.2007) support this position for not only did they illustrate differ-nt behaviours in the total carbohydrate contents of olive shootsn vitro under salt and low temperature stress, but also portrayedifferent variations of specific carbohydrates. Tattini et al. (2002)lso reported on a negative correlation between mannitol and Na+
oncentrations in Phyllirea latifolia subjected to different salt con-entrations.
. Conclusion
The findings presented in the current paper demonstrate that,n terms of the parameters being investigated and in compari-on to previous reports available in the literature, the in vitroesponses of P. vera and P. atlantica to NaCl treatment reflected sim-lar behaviours to those achieved under in vivo conditions. Theylso indicate that some reactions are activated from early devel-pmental stages and provide further information on the possible
echanisms employed by these species to avoid salt toxicity. Theigher salt tolerance of P. atlantica observed in vitro seems to beorrelated with a higher survival rate, a lower growth reduction,nd a lesser reduction in terms of the K+/Na+ and Ca2+/Na+ selec-ivity ratios in growing tissues. The variation between the different
Experimental Botany 69 (2010) 302–312 311
organs analysed in terms of growth rates and ion accumulationsconfirmed that the consideration of the whole plant is a prerequi-site for the investigation of the salt tolerance of these species. It alsocorroborates the view that the roots and vascular tissues play a keyrole in the adaptive response to salt stress. Finally, the great intra-specific variability observed particularly in the growth parametersis useful in the early screening for salt tolerance.
Acknowledgments
The authors wish to thank Dr. Ahmed Rebai, from the SfaxBiotechnology Center for his constructive review of the statisti-cal analyses, Dr. Hatem Ben Taher from the Olive Tree Institute forhis critical review and criticism of the current paper, Nabil Soua,Noura Hadrich, and Samira Kallel-Dhouib for their valuable techni-cal assistance, and Mr. Anouar Smaoui from the English Departmentat the Sfax Faculty of Science for accepting to proofread and polishthe language of the current manuscript.
References
Anonymous, 1998. National report. The state of the environment. Republic ofTunisia. Ministry for the environment and land use planning, 44 pp.
Arzani, K., Hokmabadi, H., 2004. Effects of boron and sodium chloride concentrationon growth and performance of some young pistachio rootstock seedlings. ActaHort. 658, 407–412.
Ashraf, M., 1989. The effect of NaCl on water relations, chlorophyll, and protein andproline contents of two cultivars of blackgram (Vigna mungo L.). Plant Soil 119,205–210.
Ashraf, M., 2004. Some important physiological criteria for salt tolerance in plants.Flora 199, 361–376. http://www.elsevier.de/flora.
Bartes, L.S., Waldren, R.P., Teare, I.D., 1973. Rapid determination of free prolinewater-stress studies. Plant Soil 39, 205–207.
Behboudian, M.H., Walker, R.R., Törökfalvy, E., 1986. Effects of water stress andsalinity on photosynthesis of pistachio. Sci. Hort. 29, 251–261.
Ben Khaled, L., Gomez, A.M., Honrubia, M., Oihabi, A., 2003. Effet du stress salinen milieu hydroponique sur le trèfle inoculé par le Rhizobium. Agronomie 23,553–560.
Bracci, T., Minnocci, A., Sebastiani, L., 2006. Screening of salt tolerant Olea europaeaL. seedlings by in vitro technology. In: Ismail-Alaoui, M., Roussos, S., Perraud-Gaine, I. (Eds.), Biotechnology and Quality of Olive Tree Products around theMediterranean Basin, Actes, pp. 147–154.
Caruso, T., Iannini, C., Monastra, F., Zakynthinos, G., Rouskas, D., Barone, E., Marra,F.P., Sottile, F., Battle, I., Vargas, F., Romero, M., Padulosi, S., Greco, C.I., Sabina,M.R., Martelli, G., Ak, B.E., Laghezali, M., 1998. Genetic and phenotypic diversityin Pistachio (P. vera L.) germplasm collected in Mediterranean Countries. ActaHort. 470, 168–178.
Chartzoulakis, K.S., 2005. Salinity and olive: growth, salt tolerance, photosynthesis,and yield. Agric. Water Manage. 78, 108–121.
Chartzoulakis, K., Loupassaki, M., Bertaki, M., Androulakis, I., 2002. Effects of NaClsalinity on growth, ion content and CO2 assimilation rate of six olive cultivars.Sci. Hort. 96, 235–247.
Chelli Chaabouni, A., Hammami, M., Gouia, M., Gargouri, K., Gargouri, R., Drira, N.,2008. Effect of salt stress on Pistacia atlantica seedlings at nursery conditions.XIV GREMPA Meeting, Athens, Abstract, p. 103.
Chinnusamy, V., Jagendorf, A., Zhu, J.K., 2005. Understanding and improving salttolerance in plants. Crop Sci. 45, 437–448.
Cramer, G.R., Lynch, J., Laüchli, A., Epstein, E., 1987. Influx of Na+, K+, and Ca2+
into roots of salt-stressed cotton seedlings: effects of supplemental Ca2+. PlantPhysiol. 83, 510–516.
Dasgan, H.Y., Aktas, H., Abak, K., Cakmak, I., 2002. Determination of screening tech-niques to salinity tolerance in tomatoes and investigation of genotype responses.Plant Sci. 163, 695–703.
De Pascale, S., Martino, A., Raimondi, G., Maggio, A., 2007. Comparative analysisof water and salt stress-induced modifications of quality parameters in cherrytomatoes. J. Hort. Sci. Biotech. 82 (2), 283–289.
Driver, J.A., Kuniyuki, A.H., 1984. In vitro propagation of paradox walnut rootstock.Hort. Sci. 19, 507–509.
FAO, 2002. Agricultural drainage water management in arid and semi-arid areas.http://www.fao.org/documents/show cdr.asp?url file=/DOCREP/005/Y4263E/y4263e0e.htm.
Ferguson, L., Sanden, B., Grattan, S., Epstein, L., Klueger, B., 2005. The orchard. Pis-tachio rootstocks. In: Ferguson, L., Beede, R. H., Freeman, M.W., Haviland, D.R.,
Holtz, B.A., Kallsen, C.E., Coviello, J. (Eds.), Pistachio Production Manual, 4th Edi-tion, Fruit and Nut Research and Information Center, University of California,USA, pp. 67–73.Ferreira, A.L., Lima-Costa, M.E., 2006. Metabolic responses to salt stress in cell sus-pension cultures of sensitive and resistant Citrus. J. Hort. Sci. Biotech. 81 (6),983–988.

3 l and E
G
G
G
G
G
G
H
H
H
H
H
K
K
L
M
M
M
M
M
M
M
M
N
N
O
P
P
P
R
12 A. Chelli-Chaabouni et al. / Environmenta
ebauer, J., El-Siddig, K., Salih, A.A., Ebert, G., 2004. Tamarindus indica L. seedlingsare moderately salt tolerant when exposed to NaCl-induced salinity. Sci. Hort.103, 1–8.
eorge, E.F., 1993. Plant Propagation by Tissue Culture—Part 1: The Technology, 2nded. Exegetics Ltd.
hars, M.A., Parre, E., Debez, A., Bordenave, M., Richard, L., Leport, L., Bouchereau,A., Savouré, A., Abdelly, C., 2008. Comparative salt tolerance analysis betweenArabidopsis thaliana and Thellungiella halophila, with special emphasis onK+/Na+ selectivity and proline accumulation. J. Plant Physiol. 165, 588–599.
ratten, S.R., Grieve, C.M., 1999. Salinity mineral nutrient relation in horticulturalcrops. Sci. Hort. 78, 127–157.
reenway, H., Munns, R., 1980. Mechanisms of salt tolerance in nonhalophytes.Annu. Rev. Plant Physiol. 31, 149–190.
ucci, R., Tattini, M., 1997. Salinity tolerance in olive. In: Janik. J. (Ed.), HorticulturalReviews, John Wiley & sons, Inc., USA, vol. 21, pp. 177–214.
are, P.D., Cress, W.A., Van Staden, J., 1998. Dissecting the roles of osmolyte accu-mulation during stress. Plant Cell Environ. 21, 535–553.
arinasut, P., Srisunak, S., Pitukchaisopol, S., Charoensataporn, R., 2000. Mechanismsof adaptation to increasing salinity of mulberry: proline content and ascorbateperoxydase activity in leaves of multiple shoots. ScienceAsia 26, 207–211.
ernandez, S., Deleu, C., Larher, F., 2000. Accumulation de proline dans les tissuesfoliaires de tomate en réponse à la salinité. C. R. Acad. Sci. Paris, Life Sci. 323,551–557.
okmabadi, H., Arzani, K., Grierson, P.F., 2005. Growth, chemical composition, andcarbon isotope discrimination of pistachio (Pistacia vera L.) rootstock seedlingsin response to salinity. Aust. J. Agric. Res. 56, 135–144.
uang, Y., Bie, Z., He, S., Hua, B., Zhen, A., Liu, Z., 2010. Improving cucumber toleranceto major nutrients induced salinity by grafting onto Cucurbita fucifolia. Environ.Exp. Bot. 69, 32–38.
arakas, B., Bianco, R.L., Rieger, M., 2000. Association of leaf marginal scorch withsodium accumulation in salt-stressed peach. HortScience 35, 83–84.
ozlowski, T.T., 1997. Responses of woody plants to flooding and salinity. Tree Phys-iology Monograph No. 1, Heron Publishing, Victoria, Canada. pp. 1–29.
acerda, C.F., Assis Junior, J.O., Lemos Filho, L.C.A., De Oliveira, T.S., Guimaraes,F.V.A., Gomes-Filho, E., Prisco, J.T., Bezerra, M.A., 2006. Morpho-physiologicalresponses of cowpea leaves to salt stress. Braz. J. Plant Physiol. 18 (4), 455–465(doi:10.1590/S1677-04202006000400003).
aathuis, F.J.M., Amtmann, A., 1999. K+ nutrition and Na+ toxicity: the basis ofcellular K+/Na+ ratios. Ann. Bot. 84, 123–133.
ademba-SY, F., Bouchereau, A., Larher, F.R., 2003. Proline accumulation in culti-vated citrus and its relationship with salt tolerance. J. Hort. Sci. Biotech. 78 (5),617–623.
athieu, C., Pieltain, F., 2003. Analyse chimique des sols. Méthodes choisies.Lavoisier.
eena, S.K., Gupta, N.K., Gupta, S., Khandelwal, S.K., Sastry, E.V.D., 2003. Effect ofsodium chloride on the growth and gaz exchange of young Ziziphus seedlingrootstocks. J. Hort. Sci. Biotech. 78 (4), 454–457.
elgar, J.C., Benlloch, M., Fernandez-Escobar, R., 2006. Calcium increases sodiumexclusion in olive plants. Sci. Hort. 109, 303–305.
ills, D., Zhang, G., Benzioni, A., 2001. Effect of different salts and of ABA ongrowth and mineral uptake in jojoba shoots grown in vitro. J. Plant Physiol.158, 1031–1039.
iyamoto, S., Gobran, G.R., Piela, K., 1985. Salt effects on seedling growth and ionuptake of three pecan rootstock cultivars. Agron. J. 77, 383–388.
unns, R., 2002. Comparative physiology of water and salt stress. Plant Cell Environ.25, 239–250.
eumann, P., 1997. Salinity resistance and plant growth revisited. Plant Cell Environ.20, 1193–1198.
iknam, S.R., McComb, J., 2000. Salt tolerance screening of selected Australianwoody species—a review. Forest Ecol. Manag. 139, 1–19.
kubo, M., Furukawa, Y., Sakuratini, T., 2000. Growth, flowring and leaf proper-ties of pear cultivars grafted on two Asian pear rootstock seedlings under NaClirrigation. Sci. Hort. 85, 91–101.
arida, A.K., Das, A.B., 2005. Salt tolerance and salinity effects on plant: a review.Ecotoxicol. Environ. Safe. 60, 324–349.
icchioni, G.A., Miyamoto, S., Storey, J.B., 1990. Salt effects on growth and ion uptake
of pistachio rootstock seedlings. J. Am. Soc. Hort. Sci. 115, 647–653.icchioni, G.A., Graham, C.J., 2001. Salinity, growth and ion uptake selectivity ofcontainer-growth Crataegus opaca. Sci. Hort. 90 (1), 151–166.
ad, H.M., 2006. Evaluation of salt resistance in two pistachio species with regardto growth, indices of seedlings, seedlings survival and some ecophysiologicalcharacteristics. Acta Hort. 726, 231–239.
xperimental Botany 69 (2010) 302–312
Ramoliya, P.J., Patel, H.M., Pandey, A.N., 2004. Effect of salinization of soil on growthand macro- and micro-nutrient accumulation in seedlings of Salvadora persica(Salvadoraceae). Forest Ecol. Manag. 202, 181–193.
Rejskovà, A., Patkovà, L., Stodulkovà, E., Lipavskà, H., 2007. The effect of abioticstresses on carbohydrate status of olive shoots (Olea europaea L.) under in vitroconditions. J. Plant Physiol. 164, 174–184.
Rengel, Z., 1992. The role of calcium in salt toxicity. Plant Cell Environ. 15, 625–632.Rivera, R.M., Ruiz, J.M., Romero, L., 2003. Role of grafting in horticultural plants under
stress conditions. Food, Agr. Environ. 1, 70–74.Robyt, J., White, B., 1987. Biochemical Techniques: Theory and Practice. Waveland
Press, Prospect Heights, IL (ISBN 0-88133-556-8).Rochdi, A., El Yacoubi, H., Rachidai, A., 2003. Comportement vis-à-vis de la salinité
de cals de porte-greffes d’agrumes Citrus aurantium. Citrange troyer et Poncirustrifoliata: évaluation de critères certifiant la réponse des agrumes au stress salin.Agronomie 23, 643–649.
Roussos, P.A., Gasparatos, D., Tsantili, E., Pontikis, C.A., 2007. Mineral nutrition ofjojoba explants in vitro under sodium chloride salinity. Sci. Hort. 114, 59–66.
Sairam, R.K., Tyagi, A., 2004. Physiology and molecular biology of salinity stresstolerance in plants. Curr. Sci. 86 (3), 407–421.
Sepaskhah, A.R., Maftoun, M., Karimian, N., 1985. Growth and chemical compositionof pistachio as affected by salinity and applied iron. J. Hort. Sci. 60, 115–121.
Sepaskhah, A.R., Maftoun, M., 1988. Relative salt tolerance of pistachio cultivars. J.Hort. Sci. 63, 157–162.
Shannon, M.C., Grieve, C.M., Francois, L.E., 1994. Whole-plant response to salinity.In: Wilkinson, R.E. (Ed.), Plant–Environment Interactions. Marcel Dekker, NewYork, pp. 199–244.
Shibli, R.A., Al-Juboory, K., 2002. Comparative responses of “Nabali” olivemicroshoots, callus and suspension cell cultures to salinity and water deficit.J. Plant Nutr. 25 (1), 61–74.
Slama, F., 2004. La salinité et la production végétale. Centre de Publication Univer-sitaire, p. 163.
Sotiropoulos, T.E., 2007. Effect of NaCl and CaCl2 on growth and contents of minerals,chlorophyll, proline and sugars in apple rootstock M 4 cultured in vitro. Biol.Plantarum 51 (1), 177–180.
Tabatabaei, S.J., 2006. Effects of salinity and N on growth, photosynthesis and Nstatus of olive (Olea europaea L.) trees. Sci. Hort. 108, 432–438.
Tattini, M., Heimler, D., Traversi, M.L., Pieroni, A., 1993. Polyamine analysis in saltstressed plants of olive (Olea europaea L.). J. Hort. Sci. 68, 613–617.
Tattini, M., Montagni, G., Traversi, M.L., 2002. Gas exchange, water relations andosmotic adjustement in Phyllirea latifolia grown at various salinity concentra-tions. Tree Physiol. 22, 403–412.
Tattini, M., Remorini, D., Pinelli, P., Agati, G., Saracini, E., Traversi, M.L., Massai,R., 2006. Morpho-anatomical, physiological and biochemical adjustements inresponse to root zone salinity stress and high solar radiation in two Mediter-ranean evergreen shrubs, Myrtus communis and Pistacia lentiscus. New Phytol.170 (4), 779–794.
Tattini, M., Melgar, J.C., Traversi, M.L., 2008. Responses of Olea europea to high salin-ity: a brief-ecophysiological-review. Adv. Hort. Sci. 22 (3), 159–173.
Türkan, I., Demiral, T., 2009. Recent developments in understanding salinity toler-ance. Environ. Exp. Bot. 67, 2–9.
Unno, H., Maeda, Y., Yamamoto, S., Okamoto, M., Takenaga, H., 2002. Relationshipbetween salt tolerance and Ca2+ retention among plant species. Japan J. Soil Sci.Plant Nutr. 73, 715–718.
Vijayan, K., Chakraborti, S.P., Ghosh, P.D., 2003. In vitro screening of mulberry (Morusspp.) for salinity tolerance. Plant Cell Rep. 22 (5), 350–357.
Walker, R.R., Törökfalvy, E., Behboodian, M.H., 1987. Uptake and distribution of chlo-ride. Sodium and potassium ions and growth of salt-treated pistachio plants.Aust. J. Agric. Res. 38, 383–394.
Walker, R.R., Törökfalvy, E., Behboodian, M.H., 1988. Photosynthetic rates and solutepartitioning in relation to growth of salt-treated pistachio plants (P. vera cv.Kerman). Aust. J. Plant Physiol. 15, 787–798.
Watanabe, S., Kojima, K., Ide, Y., Sasaki, S., 2000. Effects of saline and osmotic stresson proline and sugar accumulation in Populus euphratica in vitro. Plant Cell Tis.Org. Cult. 63 (3), 199–206.
Yang, L., Guizhu, C., 2007. Physiological adaptability of three mangrove species tosalt stress. Acta Ecol. Sin. 27 (6), 2208–2214.
Yildiz, M., Uzal, Ö., Yilmaz, H., 2008. Consequences of NaCl Salinity on growth
and ion accumulation in selected strawberry cultivars. Eur. J. Hort. Sci. 73,69–72.Zid, E., Grignon, C., 1991. Les tests de sélection précoce pour la résistance des plantesaux stress. Cas des stress salin et hydrique. In: AUPELF-UREF (Ed.), L’améliorationdes plantes pour l’adaptation aux milieux arides. John Libbey Eurotext, Paris, pp.91–108.














