
Growth promotion of Prunus rootstocks by root treatment with specific bacterial strains
15
AUTHOR’S PROOFS PDF OUTPUT Plant and Soil 00: 1–15, 2003. © 2003 Kluwer Academic Publishers. Printed in the Netherlands. 1 Growth promotion of Prunus rootstocks by root treatment with specific bacterial strains A. Bonaterra 1 , L. Ruz 1 , E. Badosa 1 , J. Pinochet 2 & E. Montesinos 1,3 1 Institute of Food and Agricultural Technology and CeRTA-CIDSAV, University of Girona, Av. Llu ´ is Santal´ o, 17071 Girona, Spain. 2 Agromillora Catalana, S. A., 08739 Subirats (Barcelona), Spain. 3 Corresponding author ∗ Received 26 July 2002; accepted in revised form 27 March 2003 Key words: dose–response effect, Prunus nursery, Pantoea agglomerans, Pseudomonas fluorescens, in vitro antagonism, secondary metabolites Abstract A collection of bacterial strains obtained from a wide-range origin was screened for ability to promote growth in two types of Prunus rootstocks in a commercial nursery. Only few strains promoted growth significantly and consistently, and a strong specificity for the rootstock cultivar was observed. Irrigation of plants with Pseudomonas fluorescens EPS282 and Pantoea agglomerans EPS427 significantly increased plant height and root weight of the plum Marianna 2624 and the peach–almond hybrid GF-677, respectively. Plant height showed a higher rate of growth in early stages of development (2.6–3.5 times the non-treated controls), but the effect decreased with plant age. However, in aged plants growth promotion was more significant on root weight (1.9 times the non- treated controls) than on plant height. The efficacy of growth promotion and the persistence of strains in the root environment were dependent on the bacterial inoculum concentration applied. Increases in root development were maximum at inoculum concentrations of up to 8 log 10 CFU ml −1 (ca 10 log 10 CFU L −1 of potting mix). Population levels at the optimum inoculum concentration were around 7 log 10 CFU g f.w. −1 root material at early stages of development and decreased to 4 log 10 CFU g f.w. −1 after several months of development. The best plant growth- promoting strains were very diverse in secondary metabolite production and antagonistic ability against several plant pathogens. Introduction Plant growth-promoting rhizobacteria (PGPR) have been used in greenhouse and in the field to improve crop production or as plant enhancers to increase stress tolerance and disease resistance. Pseudomonas, Bacil- lus, Azospirillum, Azotobacter, Enterobacter, Klebsi- ella and Serratia are the most frequent non-symbiotic genera including strains with plant growth promotion activity (Kloepper, 1992). PGPR have been studied in herbaceous plants such as potato, bean, soybean, tomato, cucumber and radish, (Anderson and Guerra, 1985; Brito Alvarez et al. 1995; Burr et al., 1978; Dashti et al., 1997; Leeman et al., 1995; Raupach and Kloepper, 1998). However, few reports include woody ∗ FAX NO: +34-9-7241-8399. Email: [email protected] plants like apple (Caesar and Burr, 1987), citrus (Gardner et al., 1984), and alder (Gutiérrez-Mañero et al., 1996). Plant growth promotion by PGPR may be due to the production and release of plant growth regulator compounds such as indole-3-acetic acid, gibberellins and cytokinins (Barazani and Friedman, 1999; Benizri et al., 2001). In some strains the effect was attributed to the elicitation of root metabolic activities mediated by bacterial surface components, acceleration of ni- trogen fixation activity or phosphate-solubilization in the root environment (Boruah et al., 2002; Dashti et al., 1998). The beneficial effect may be produced by protecting plants against soil-borne pathogens or de- leterious rhizobacteria. Bacteria selected initially for growth promotion in the absence of pathogens may exhibit biological control activity when challenged 10735.tex; 2/05/2003; 12:57; p.1 YB: ICPC: DISK: CP: PIPS NO.: 5126969 (plsokap:bio2fam) v.1.1
Transcript of Growth promotion of Prunus rootstocks by root treatment with specific bacterial strains

AU
TH
OR
’S P
RO
OFS
PDF OUTPUT
Plant and Soil 00: 1–15, 2003.© 2003 Kluwer Academic Publishers. Printed in the Netherlands.
1
Growth promotion of Prunus rootstocks by root treatment with specificbacterial strains
A. Bonaterra1, L. Ruz1, E. Badosa1, J. Pinochet2 & E. Montesinos1,3
1Institute of Food and Agricultural Technology and CeRTA-CIDSAV, University of Girona, Av. Lluis Santalo, 17071Girona, Spain. 2Agromillora Catalana, S. A., 08739 Subirats (Barcelona), Spain. 3Corresponding author∗
Received 26 July 2002; accepted in revised form 27 March 2003
Key words: dose–response effect, Prunus nursery, Pantoea agglomerans, Pseudomonas fluorescens, in vitroantagonism, secondary metabolites
Abstract
A collection of bacterial strains obtained from a wide-range origin was screened for ability to promote growthin two types of Prunus rootstocks in a commercial nursery. Only few strains promoted growth significantly andconsistently, and a strong specificity for the rootstock cultivar was observed. Irrigation of plants with Pseudomonasfluorescens EPS282 and Pantoea agglomerans EPS427 significantly increased plant height and root weight of theplum Marianna 2624 and the peach–almond hybrid GF-677, respectively. Plant height showed a higher rate ofgrowth in early stages of development (2.6–3.5 times the non-treated controls), but the effect decreased withplant age. However, in aged plants growth promotion was more significant on root weight (1.9 times the non-treated controls) than on plant height. The efficacy of growth promotion and the persistence of strains in the rootenvironment were dependent on the bacterial inoculum concentration applied. Increases in root development weremaximum at inoculum concentrations of up to 8 log10 CFU ml−1 (ca 10 log10 CFU L−1 of potting mix). Populationlevels at the optimum inoculum concentration were around 7 log10 CFU g f.w.−1 root material at early stages ofdevelopment and decreased to 4 log10 CFU g f.w.−1 after several months of development. The best plant growth-promoting strains were very diverse in secondary metabolite production and antagonistic ability against severalplant pathogens.
Introduction
Plant growth-promoting rhizobacteria (PGPR) havebeen used in greenhouse and in the field to improvecrop production or as plant enhancers to increase stresstolerance and disease resistance. Pseudomonas, Bacil-lus, Azospirillum, Azotobacter, Enterobacter, Klebsi-ella and Serratia are the most frequent non-symbioticgenera including strains with plant growth promotionactivity (Kloepper, 1992). PGPR have been studiedin herbaceous plants such as potato, bean, soybean,tomato, cucumber and radish, (Anderson and Guerra,1985; Brito Alvarez et al. 1995; Burr et al., 1978;Dashti et al., 1997; Leeman et al., 1995; Raupach andKloepper, 1998). However, few reports include woody
∗ FAX NO: +34-9-7241-8399. Email: [email protected]
plants like apple (Caesar and Burr, 1987), citrus(Gardner et al., 1984), and alder (Gutiérrez-Mañeroet al., 1996).
Plant growth promotion by PGPR may be due tothe production and release of plant growth regulatorcompounds such as indole-3-acetic acid, gibberellinsand cytokinins (Barazani and Friedman, 1999; Benizriet al., 2001). In some strains the effect was attributedto the elicitation of root metabolic activities mediatedby bacterial surface components, acceleration of ni-trogen fixation activity or phosphate-solubilization inthe root environment (Boruah et al., 2002; Dashti etal., 1998). The beneficial effect may be produced byprotecting plants against soil-borne pathogens or de-leterious rhizobacteria. Bacteria selected initially forgrowth promotion in the absence of pathogens mayexhibit biological control activity when challenged
10735.tex; 2/05/2003; 12:57; p.1
YB: ICPC: DISK: CP: PIPS NO.: 5126969 (plsokap:bio2fam) v.1.1

PDF OUTPUT
2
with the pathogen, and the effect of plant growth en-hancement may be due to the control of deleteriousmicroorganisms or nontarget pathogens (Kloepper,1992).
Protection against pathogens may be indirectlyachieved by competition for nutrients and iron or moredirectly due to the synthesis of antibiotic compoundsor HCN (Handelsman and Staab, 1996; Weller et al.,2002). Several reports showed also beneficial effectsdue to the elicitation of systemic resistance in plantsto pathogens (Liu et al., 1995; Maurhofer et al., 1994).
Nursery rootstock production is actually expand-ing, especially for Prunus species which can bemassively produced by in vitro multiplication tech-niques. In this process, plantlets obtained by micro-propagation are first transplanted and acclimatized tochanging environmental conditions, then developedto a suitable size, and finally commercialized dir-ectly or grafted to the appropriate tree species orcultivars. During these stages plant decay or slowgrowth due to insufficient root development may havenegative effects on rootstock production. These prob-lems are usually managed by fertilization, by the useof treatments with plant hormones to increase root de-velopment or with fungicides applications for controlof soil-borne fungal pathogens. However, stimula-tion of root development and pathogen control can beachieved by means of less aggressive methods such astreatment with growth promoting bacteria. The use ofPGPR in plant nurseries has the advantage of acceler-ating the production process by minimizing the timerequired for the lignification of plantlets with the pur-pose of obtaining hardened plants which is essentialfor their future development after transplant into thefield (Caesar and Burr, 1987). Also, rootstock plantletswith well-developed root systems have higher survivalrates at transplant as well as a better establishmentin replant situations (Utkhede and Smith, 1992). Thelatter is considered critical in many fruit tree grow-ing areas in Mediterranean environments. However,there are no reports on the application of plant growthpromoting bacteria in Prunus rootstock production.
The aim of the present work was to find plantgrowth-promoting bacteria suitable for use in commer-cial Prunus rootstock production. This investigationpresents the results of a screening of putative growthpromoting strains of Pseudomonas fluorescens andPantoea agglomerans, and the selection and charac-terization of the best strains. Data on dose–responserelationships and colonization and survival in the rootsystem are provided.
Materials and methods
Bacterial strains
Putative growth promoting bacterial strains were isol-ated from 206 samples taken from aerial parts androots of several plants, including 36 species (sevenfamilies) and 55 locations in the North East of Spainduring the spring and autumn of 1993 and 1994(Montesinos et al., 1996). Plant material includedleaves, flowers, buds and roots. Roots were washedgently with sterile distilled water to remove soil debris.Buds and flowers were cut into small pieces beforeextracting epiphytic bacteria. Epiphytic bacteria wereextracted from plant material by stirring it at 200rpm for 30 min in buffered peptone water (0.1%Bactopeptone in 30 mM potassium phosphate buffer).The extract was serially diluted in potassium phos-phate buffer (70 mM, pH = 7.0) and a 10-µl drop ofeach dilution was spotted onto the surface of King’s Bagar (KB) to detect fluorescent Pseudomonas (Kinget al. 1954) or Miller-Schroth agar plates amendedwith mannitol (MS) to detect Erwinia species (syn.Pantoea) (Miller and Schroth, 1972). Plates wereincubated for 48 h at 25 ◦C. P. fluorescens was prelim-inarily identified in KB agar as fluorescent coloniesunder ultraviolet light (366 nm) and positive for theoxidase test. E. herbicola (syn. P. agglomerans) waspreliminarily identified as typical red–orange colon-ies with a darker central area in MS medium. Morecomplete species identification was performed accord-ing to the Bergey’s Manual of Systematic Bacteri-ology and as reported by several authors (Dye, 1969;Ewing and Fife, 1972; Gavini et al., 1989; Palleroni,1984) using the following additional tests: Gram stain,motility, oxidative/fermentative metabolism, argininedihydrolase, nitrate reduction, levan production, in-dol, gelatin hydrolysis, methyl-red, Voges-Proskauer,tobacco hypersensitivity and ice-nucleation activity.Additional identification of the strains was performedusing the analytical profile index (API) system (API,Biomerieux, France). A total of 186 strains of Pantoeaagglomerans and 252 strains of Pseudomonas fluores-cens, were chosen from the collection. Additionally,18 Pseudomonas species, most pertaining to P. fluor-escens, from culture collections and biocontrol agentsof different origin were used (Table 1).
Spontaneous resistant mutants to 100 µg ml−1 ofrifampicin were used in the colonization and survivalexperiments. The strains used were EPS282, EPS435,EPS512, EPS547, EPS353, EPS427 and EPS453.
10735.tex; 2/05/2003; 12:57; p.2

PDF OUTPUT
3
Table 1. Pseudomonas fluorescens strains provided byother researchers or from culture collections used inthe study
Strain Origin Source1
ATCC13525 Prefilter tanks ATCC
ATCC17513 – ATCC
ATCC17518 Soil ATCC
ATCC17559 – ATCC
ATCC17816 Dahlia ATCC
BL915 Cotton root S. Hill
CHAO Tobacco root G. Defago
JBR1-70 Wheat root J. Raaijmakers
JMP12-84 Wheat root J. Raaijmakers
Ps15 Sugarcane root M. N. Nielsen
Ps31 Sugarcane root M. N. Nielsen
2-79 Wheat root L. Thomashow
Q2−87 Wheat root L. Thomashow
Q4−87 Wheat root L. Thomashow
Qc69-80 Wheat root L. Thomashow
SBW25 Wheat root L. Thomashow
M480-R Sugar beet root M. J. Bailey
WB52 Tomato root W. Fakhouri
1Steve Hill, Novartis Crop Protection, Inc. ResearchTriangle Park, NC, USA; Genevieve Defago, Insti-tute of Plant Sciences/Phytopathology, Swiss Fed-eral Institute of Technology, Zurich, Switzerland; JosJ. Raaijmakers, Laboratory for Phytopathology, Wa-geningen Agricultural University, The Netherlands;Mette Neiendam Nielsen, Department of Ecology andMolecular Biology. Section of Genetics and Micro-biology, Royal Veterinary and Agricultural Univer-sity, Denmark; Linda Thomashow, US Deparmentof Agricultural Research Service, Root Disease andBiological control Research Unit, Washington StateUniversity, USA; Mark J. Bailey, Molecular Micro-bial Ecology, Institute of Virology and EnvironmentalMicrobiology, Oxford, England; Walid Fakhouri, Uni-versität Hohenheim, Institut für Phytomedizin, Stut-gart, Germany.
These mutants were checked for maintenance of themain phenotypic and genotypic characteristics com-pared to the wild type parental strains.
For long-term preservation, bacteria were storedin 20% glycerol at −80 ◦C. For inoculations, strainswere cultivated in LB liquid medium at 25 ◦C for 24h (Maniatis et al., 1982). The cells were pelleted bycentrifugation at 10 000 × g for 15 min, suspendedin sterile phosphate buffer (70 mM potassium phos-phate buffer at pH = 7), and the concentration adjustedto 109 CFU ml−1 using a viable versus absorbanceat 600 nm curve specific for P. agglomerans or P.fluorescens strains. Appropriate dilutions from the ori-
ginal concentration were performed depending on theexperiment.
Plant material
Two types of Prunus rootstocks, the plum Marianna2624 (Prunus cerasifera × P. munsoniana) and the al-mond × peach hybrid GF-677 (P. dulcis × P. persica)were used in the experiments. The plants were pro-duced by in vitro micropropagation in a commercialnursery (Agromillora Catalana, Barcelona, Spain).Most experiments were performed in the nursery facil-ity under greenhouse conditions using standard com-mercial practices. The general process consisted ofin vitro micropropagation and rooting, transplant tosmall minipot trays for acclimatization, followed byseveral additional transplants to pots of increasingsize, and finally grafted with a commercial cultivar formarketing.
Plant growth promotion assays
The screening procedure consisted of two stages. Inthe first stage all the strains were tested in a short-termduration experiment with fast developing and highlyimmature plants. In the second stage strains selectedin the first stage were tested in a longer 3–5 monthsperiod.
(i) First stage selectionA total of 438 strains from our collection and 18strains of P. fluorescens reported as biocontrol agentswere processed first. The strains were tested forgrowth promotion activity in the two Prunus root-stocks, Marianna 2624 and GF-677. In vitro plantswere highly homogeneous and were used after the ac-climatization process (6–8 cm height). Each plantletwas contained in a 60-ml pot. The experimental designconsisted of six replicates per treatment composed ofone plant per replicate. Pots corresponding to the sametreatment were arranged together in trays. Each traycontained a set of nine treatments. Each plant wastreated on 2 February 1999 by irrigation with 10 mlof each bacterial suspension at 108 CFU ml−1 (1010
CFU l−1 of potting mix). The substrate consisted ofBVV potting mix (Prodeasa, Barcelona, Spain) andPindstrup peat-moss substrate (Mosebrug A/S, Den-mark). Fertilizer (200 ppm of 20-10-29 N-P-K) wasapplied every week. A sterile phosphate buffer solu-tion without bacteria was used for non-treated controlplants. This amount of water was enough to wet the
10735.tex; 2/05/2003; 12:57; p.3

PDF OUTPUT
4
potting mix without significant losses by leakage fromthe pots. Plant height was assessed on 5 March 1999after growth promotion approximately 1 month of thetreatment with the putative growth promotion bacteria.
(ii) Second stage selectionFollowing the first stage selection, specificity of root-stock type in many of the most efficient strains wasobserved. Therefore, different strains were tested oneach rootstock.
In the first trial, strains EPS231, EPS282, EPS394,EPS435, EPS547, EPS599 and EPS607 were inocu-lated on Marianna 2624 and strains EPS353, EPS427,EPS453, EPS475 and EPS512 were inoculated on GF-677. The first treatment was performed on 9 February2000 as described for the first stage selection. Plantswere transplanted in 26 March 2000 to larger 300-ml pots, and treated with 50 ml of a 108 CFU ml−1
bacterial suspension per plant (1.7 × 1010 CFU l−1 ofpotting mix). After suitable growth, the plants werefinally transplanted in 30 April 2000 to 40 × 60 ×20-cm plastic boxes containing 25 l of potting mix.Each box contained six plants and was treated with600 ml of a 4.0 × 108 CFU ml−1 of the correspond-ing bacterial suspension (aprox. 1.0 × 1010 CFU l−1
of potting mix). The experimental design consisted ofthree replications (boxes) of six plants per replicationfor each treatment (bacterial strain). Plant height wasmeasured at 3 March, 30 April, 14 June and 7 July.At the end of the assay on 23 July 2000, plant height,fresh weight of aerial part, and root fresh weight weredetermined.
In the second trial, a selection of the best strainsaccording to the results obtained in the first trial wasused. Strains EPS282, EPS435, EPS512 and EPS547were inoculated on Marianna 2624. Strains EPS353,EPS427 and EPS453 were inoculated on GF-677.The plants were obtained at approximately 15–20 cmheight (1 month after the acclimatization stage), andwere initially transplanted to 300-ml pots on 21 Feb-ruary 2001. The first treatment was performed aftertransplant by irrigation with 40 ml of the correspond-ing bacterial suspension of 1.0 × 108 CFU ml−1
(1.3 × 1010 CFU l−1 of potting mix). The treatmentwas continued on 8 March, 22 March, 5 April, 19April and 3 May, making a total of five treatments.Treatments were arranged in a completely randomizedblock design with three replicates consisting of sixplants per replicate. Aerial plant height was assessedmonthly upon starting the experiment and aerial plantweight and root fresh weight was assessed at the end
of the assay, 140 days after the beginning of the firstinoculation.
Dose–response assay
Marianna 2624 plantlets aged 1 month were trans-planted to 300-ml pots (one plant per pot) and treatedby irrigation with 50 ml of strain EPS282 at the corres-ponding bacterial concentrations. The first treatmentwas performed on 8 March 2001 and was repeated ap-proximately each 2 weeks until 10 May 2001 includ-ing a total of five treatments. The range of inoculumconcentrations studied was 1 × 104, 1 × 105, 1 × 106,1 × 107, 1 × 108 and 1 × 109 CFU ml−1. The ex-perimental design consisted of three replicates of fiveplants per replicate for each dose. Aerial plant heightwas assessed on 30 March, 5 May and 23 May 2001.Aerial plant weight and root fresh and dry weight weredetermined on 23 May 2001, approximately 3 monthsafter the first treatment.
Assessment of population levels of inoculatedbacterial strains on treated plants
The population levels of the inoculated bacteria con-sisting of spontaneous resistant mutants to rifampicinon treated plants were assessed in the second trial ofthe second stage selection and in the dose–response as-say. In the second trial the strains used were EPS282,EPS435, EPS512 and EPS547 on Marianna 2624 andEPS353, EPS427 and EPS453 on GF-677. In thedose–response assay the strain EPS282 was used totreat plants of Marianna 2624. Population levels of thestrains were assessed at the moment of the first treat-ment (21 February in the second trial and 8 March inthe dose–response assay) and at the end of the assay(5 May in the second trial and 23 May in the dose–response assay). The experimental design consistedof three replicates of one plant per replicate for eachstrain or dose and date assessed.
Samples of plant roots were gently rinsed withsterile distilled water to remove adhering soil particles.Then, roots were extracted in a stomacher for 1 minin buffered peptone water (0.1% Bacto peptone in 30mM potassium phosphate buffer 70 mM, pH 7.0), andsuspensions were 10-fold serially diluted in water. Ap-propriate dilutions were plated onto LB agar platesamended with 100 µg of rifampicin per ml, and incub-ated for 24 h at 22 ◦C. Colonies were counted and datawere transformed to log10 CFU g−1 of fresh weight ofroot.
10735.tex; 2/05/2003; 12:57; p.4

PDF OUTPUT
5
Strain characterization
Some of the best plant growth promoting strains P.fluorescens EPS282, EPS326 and EPS599, P. ag-glomerans EPS427, EPS512, and EPS622, and thebiocontrol agents CHAO, JBR1-70, Ps15 and WB25,were characterized for the in vitro antagonism abilityand the production of several metabolites.
(i) In vitro antagonism assayScreening for in vitro antagonism against the patho-gens Erwinia amylovora, Pseudomonas syringae,Stemphylium vesicarium and Penicillium expansumwas performed using dual cultures with agar overlaysof different culture media. The bacteria and fungalindicator strains were used because they represent im-portant bacteria and fungi which are commonly usedin our laboratory. In vitro antagonism against P. syr-ingae and E. amylovora was determined in glucoseasparagine (GA)(Wodzinski and Paulin, 1994), King’sB and LB agar. GA and KB agar amended with iron(50 µM FeCl3) were also used to prevent siderophoreproduction to detect additional antagonistic phenom-ena. Screening for in vitro antagonism against thepathogens S. vesicarium and P. expansum was per-formed in GA, LB, potato-dextrose agar (PDA) andGA amended with iron. Overlays were prepared bymixing 5 ml of melted 0.7% agar with 0.1 ml of abacterial suspension adjusted to 108 CFU ml−1 ora conidial suspension adjusted to 104 conidia ml−1.Bacterial colonies of the putative antagonists weretransferred to the surface of the overlay agar plateswith sterile toothpicks. The agar plates were incubatedat 25 ◦C and inhibition zones were assessed after 24,48 and 72 h. All the isolates that produced inhibitionzones in at least one medium were considered as posit-ive for a given indicator microorganism in the in vitroantagonism test.
(ii) Metabolite productionHydrogen cyanide production was assessed by thepicrate method (Sneath, 1966). Tubes containing LBagar and a filter paper strip impregnated with 0.5%picric acid and 2% NaCO3 introduced into the test tubescrew cup, were inoculated with the bacteria. Upon in-cubation at 22 ◦C for 3 days production of cyanide wasdetected as a change of the filter paper color from yel-low to orange–brown. Indoleacetic acid (IAA) and re-lated compounds were identified upon colony growthin LB agar amended with 5 mM 1-tryptophan andoverlaid with an 82-mm diameter nitrocellulose mem-
brane disk (Brito Alvarez et al., 1995). Agar plateswere inoculated with bacterial cultures and incubatedat 28 ◦C for 3 days. Then, the membranes were re-moved and placed overlaid on a Whatman filter paperimpregnated with the Salkowski reagent. Strains pro-ducing IAA or analogs substances were identified by apink to red color in the filter surface. Siderophore pro-duction was determined using chrome azurol S (CAS)medium (Schwyn and Neilands, 1987). Chrome azurolS agar plates were inoculated with the strains andincubated at 28 ◦C for 5 days. Strains exhibiting anorange halo were positive for the siderophore pro-duction. Chitinase production was identified in BBLmedium amended with 0.15% colloidal chitin (Fränd-berg and Shnürer, 1998). Chitinolytic bacteria wereidentified as colonies surrounded by clearing zonesafter 5 days of incubation at 30 ◦C. The productionof phenazine-1-carboxylic acid (PCA) and phloroglu-cinol (Phl) was assessed only in P. fluorescens strainsusing a modified high-performance liquid chromato-graphy (HPLC) technique (Alemany, 2001; Keel etal., 1992). Liquid cultures of the strains were pre-pared in LB broth amended with 1% glucose and wereincubated during 48 h at 25 ◦C. A 1-ml aliquot ofthe cultures was centrifuged at 5000 × g during 20min, the supernatant was treated with 50 mg NaCl, 50ml HCl 2 M and 500 µl of ethyl acetate, and cent-rifuged at 10 000 × g during 1 min. An aliquot of400 µl of the supernatant was reduced to dryness un-der vacuum and the dry residue dissolved in 800 µlof acetonitrile–aqueous trichloroacetic acid (40:60) atpH 2.8 for further analysis by HPLC. Samples of 20µl were injected in a Waters HPLC (model 610, Wa-ters, Mildford, Madison, WI, USA) equipped with ananalytical column (5 µm 15 × 0.46 cm) packed withTracer Hypersil ODS attached to a precolumn and op-erating at a flow of 0.8 ml min−1. Measurements wereperformed with a photodiode array detector (model990, Waters). Under these conditions Phl and PCAwere detected respectively at 9 and 7.6 min retentiontime.
Data analysis
To test the significance of the effect of treatments, aone-way analysis of variance was performed. Meanswere separated using the Waller–Duncan test at P ≤0.05. The analysis was performed with the GLMprocedure of the PC-SAS (SAS Institute, Cary, NC).
10735.tex; 2/05/2003; 12:57; p.5

PDF OUTPUT
6
Results
Screening of plant growth promotion activity
The screening on young plants of Prunus rootstockswas very effective in detecting growth promotionstrains. In Marianna 2624, 28% of the strains showeddeleterious effects and inhibited growth, 18% strainswere neutral, and the remaining 54% exhibited growthstimulation in some degree. In GF-677, 22% were de-leterious, 29% were neutral, and 49% showed growthstimulation. These proportions do not differ signi-ficantly between Pseudomonas fluorescens and Pan-toea agglomerans species. However, the intensity ofgrowth promotion was higher in Marianna 2624 thanin GF-677. The more efficient strains caused increasesof 2-fold in plant height and accounted for 15% (69strains) in Marianna 2624, and only 1% (4 strains) inGF-677. From the more efficient strains, 22 strains ofP. agglomerans and 47 of P. fluorescens were active inMarianna 2624, whereas only three P. agglomeransand one P. fluorescens strains were active in GF-677. Among the culture collection strains only four(CHAO, JBR1-70, WB52 and Ps15) caused a signific-ant increase in plant size of 2.1–2.3 times in the plumrootstock, whereas no effect was observed in GF-677(Table 2).
The effect of P. agglomerans or P. fluorescensstrains from our collection was dependent on the root-stock type. According to the analysis of variance, theinteraction between strain and rootstock was signific-ant (P < 0.001). Among the P. agglomerans strains,several were effective on both rootstocks cultivars(EPS512, EPS427, EPS560 and EPS475), some wereeffective only in Marianna 2624 (EPS622, EPS394,EPS607, EPS595, EPS435 and EPS547), and otherswere only effective in GF-677 (EPS453, EPS495 andEPS454) (Table 3). The most effective strains in Mari-anna 2624 were EPS622, EPS512 and EPS427 andproduced plant height increases of 3.2–3.5-fold in re-lation to the non-treated control, and in GF-677 wereEPS454 and EPS427 and produced increases of 1.8-fold. Among the P. fluorescens strains several wereeffective on both rootstock types (EPS326, EPS374,EPS353, EPS225 and EPS289), some were effectiveonly in Marianna 2624 (EPS599, EPS231, EPS588,EPS282 and EPS212), and others were only effectivein GF-677 (EPS383 and EPS286) (Table 4). The threemost effective strains in Marianna 2624 were EPS599,EPS326 and EPS231 which caused plant height in-creases of 2.6–2.9-fold versus the non-treated control,
Table 2. Effect of strains of Pseudomonasfluorescens from culture collections or fromother sources on growth promotion of plumMarianna 2624 during the first stage selectionexperiment
Strain Plant height (cm)1
CHAO 10.4 a
JBR1-70 9.7 ab
WB52 9.7 ab
Ps15 9.6 ab
JMP12-84 9.3 abc
M480-R 8.7 abcd
Ps31 8.2 abcd
Q2−87 7.5 abcd
SBW25 6.5 abcde
BL915 6.3 abcde
Q4−87 6.2 abcde
Qc69-80 5.7 abcde
ATCC17816 5.0 bcde
ATCC17559 4.7 bcde
Non-treated control 4.5 cde
2-79 4.1 de
ATCC17513 4.1 de
ATCC17518 3.8 de
ATCC13325 2.3 e
1Values are the mean of six plantlets. Meansfollowed by the same letter are not signi-ficantly different according to the Waller–Duncan test (P = 0.05). The rootstock plant-lets were treated by a single irrigation with10 ml of a bacterial suspension of 108 CFUml−1. The height of the plants was assessed 1month after the treatment.
and in GF-677 were EPS353, EPS326 and EPS383with increases of 1.8-fold versus the non-treated con-trol.
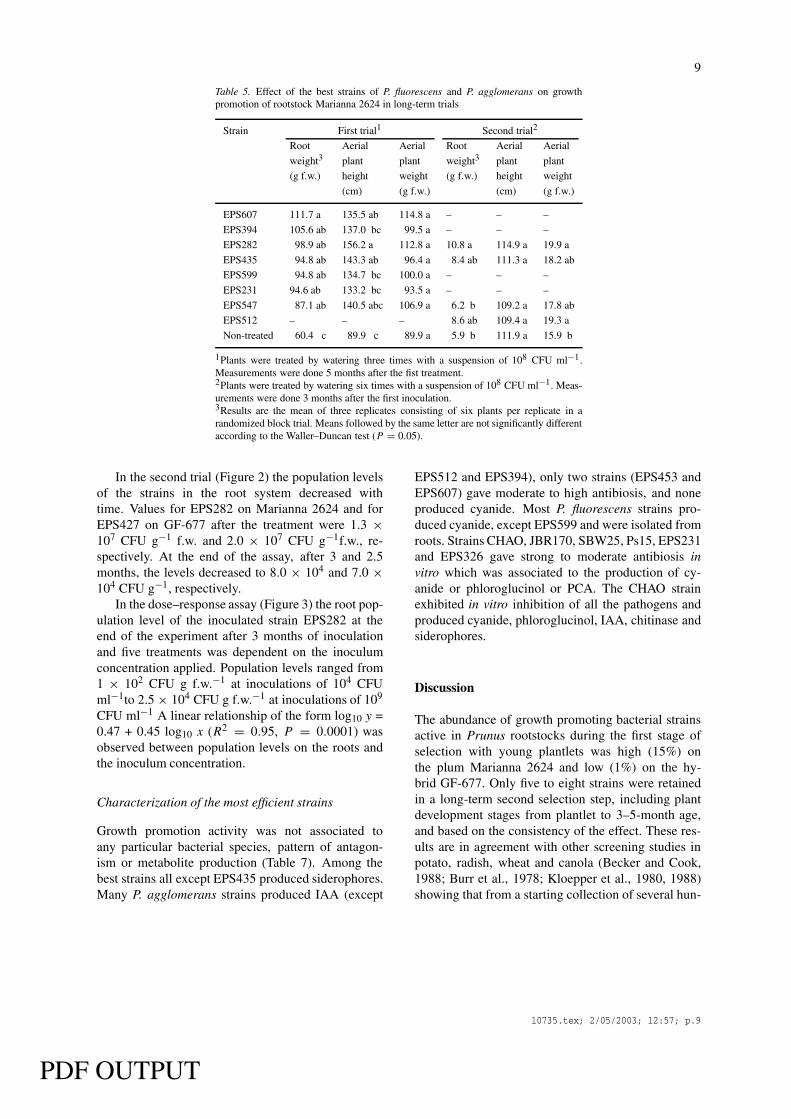
In the second stage selection experiment severalstrains retained from the first screening experimentwere studied. In Marianna 2624 rootstock, the effecton the root weight was highly consistent (Table 5).In the first trial, strains EPS607, EPS394, EPS282,EPS435, EPS599, EPS231 and EPS547 produced sig-nificant increases of root fresh weight (1.9-fold overthe non-treated control) at 5 months after the firstinoculation. However, in the second trial, amongthe strains tested (EPS282, EPS435, EPS547 andEPS512), only strain EPS282 produced significantroot weight promotion (1.8-fold) after 3 months ofplant development. The lower levels of root weightgrowth observed in the second trial were due to theshorter duration of the experiment in the later case.
10735.tex; 2/05/2003; 12:57; p.6
PDF OUTPUT
7
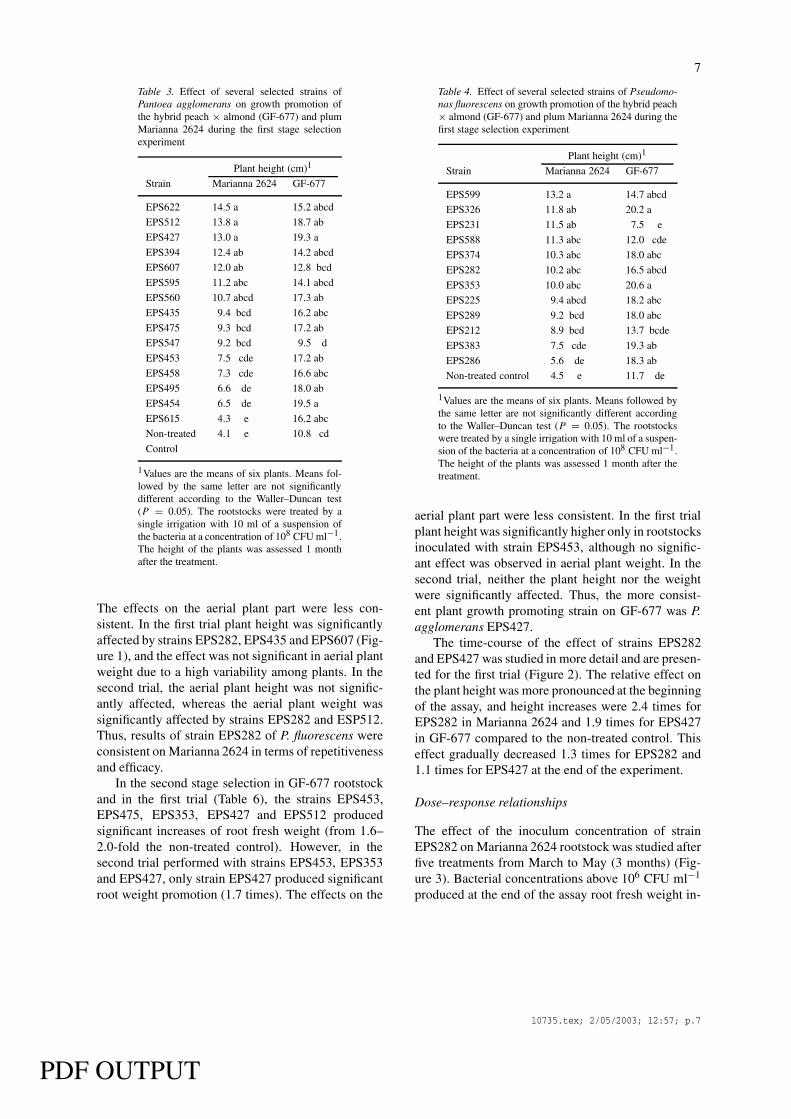
Table 3. Effect of several selected strains ofPantoea agglomerans on growth promotion ofthe hybrid peach × almond (GF-677) and plumMarianna 2624 during the first stage selectionexperiment
Plant height (cm)1
Strain Marianna 2624 GF-677
EPS622 14.5 a 15.2 abcd
EPS512 13.8 a 18.7 ab
EPS427 13.0 a 19.3 a
EPS394 12.4 ab 14.2 abcd
EPS607 12.0 ab 12.8 bcd
EPS595 11.2 abc 14.1 abcd
EPS560 10.7 abcd 17.3 ab
EPS435 9.4 bcd 16.2 abc
EPS475 9.3 bcd 17.2 ab
EPS547 9.2 bcd 9.5 d
EPS453 7.5 cde 17.2 ab
EPS458 7.3 cde 16.6 abc
EPS495 6.6 de 18.0 ab
EPS454 6.5 de 19.5 a
EPS615 4.3 e 16.2 abc
Non-treated 4.1 e 10.8 cd
Control
1Values are the means of six plants. Means fol-lowed by the same letter are not significantlydifferent according to the Waller–Duncan test(P = 0.05). The rootstocks were treated by asingle irrigation with 10 ml of a suspension ofthe bacteria at a concentration of 108 CFU ml−1.The height of the plants was assessed 1 monthafter the treatment.
The effects on the aerial plant part were less con-sistent. In the first trial plant height was significantlyaffected by strains EPS282, EPS435 and EPS607 (Fig-ure 1), and the effect was not significant in aerial plantweight due to a high variability among plants. In thesecond trial, the aerial plant height was not signific-antly affected, whereas the aerial plant weight wassignificantly affected by strains EPS282 and ESP512.Thus, results of strain EPS282 of P. fluorescens wereconsistent on Marianna 2624 in terms of repetitivenessand efficacy.
In the second stage selection in GF-677 rootstockand in the first trial (Table 6), the strains EPS453,EPS475, EPS353, EPS427 and EPS512 producedsignificant increases of root fresh weight (from 1.6–2.0-fold the non-treated control). However, in thesecond trial performed with strains EPS453, EPS353and EPS427, only strain EPS427 produced significantroot weight promotion (1.7 times). The effects on the
Table 4. Effect of several selected strains of Pseudomo-nas fluorescens on growth promotion of the hybrid peach× almond (GF-677) and plum Marianna 2624 during thefirst stage selection experiment
Plant height (cm)1
Strain Marianna 2624 GF-677
EPS599 13.2 a 14.7 abcd
EPS326 11.8 ab 20.2 a
EPS231 11.5 ab 7.5 e
EPS588 11.3 abc 12.0 cde
EPS374 10.3 abc 18.0 abc
EPS282 10.2 abc 16.5 abcd
EPS353 10.0 abc 20.6 a
EPS225 9.4 abcd 18.2 abc
EPS289 9.2 bcd 18.0 abc
EPS212 8.9 bcd 13.7 bcde
EPS383 7.5 cde 19.3 ab
EPS286 5.6 de 18.3 ab
Non-treated control 4.5 e 11.7 de
1Values are the means of six plants. Means followed bythe same letter are not significantly different accordingto the Waller–Duncan test (P = 0.05). The rootstockswere treated by a single irrigation with 10 ml of a suspen-sion of the bacteria at a concentration of 108 CFU ml−1.The height of the plants was assessed 1 month after thetreatment.
aerial plant part were less consistent. In the first trialplant height was significantly higher only in rootstocksinoculated with strain EPS453, although no signific-ant effect was observed in aerial plant weight. In thesecond trial, neither the plant height nor the weightwere significantly affected. Thus, the more consist-ent plant growth promoting strain on GF-677 was P.agglomerans EPS427.
The time-course of the effect of strains EPS282and EPS427 was studied in more detail and are presen-ted for the first trial (Figure 2). The relative effect onthe plant height was more pronounced at the beginningof the assay, and height increases were 2.4 times forEPS282 in Marianna 2624 and 1.9 times for EPS427in GF-677 compared to the non-treated control. Thiseffect gradually decreased 1.3 times for EPS282 and1.1 times for EPS427 at the end of the experiment.
Dose–response relationships
The effect of the inoculum concentration of strainEPS282 on Marianna 2624 rootstock was studied afterfive treatments from March to May (3 months) (Fig-ure 3). Bacterial concentrations above 106 CFU ml−1
produced at the end of the assay root fresh weight in-
10735.tex; 2/05/2003; 12:57; p.7

PDF OUTPUT
8
Figure 1. Growth stimulation of the plum Marianna 2624 after three treatments by irrigation with a suspension of Pseudomonas fluorescensEPS282 (right) in comparison to the non-treated control (left). The picture corresponds to the first trial of the second selection stage, and toplants aged 3 months.
Figure 2. Time course of plant height in Marianna 2624 root-stock (A) treated with Pseudomonas fluorescens EPS282 (�) andin GF-677 (B) treated with Pantoea agglomerans EPS427 (�) com-pared to non-treated controls (�). Data correspond to the first trialof the second stage selection. The bacterial strains were applied bywatering the plants with a suspension of 108 CFU ml−1 in phos-phate buffer. The treatments were done three times at 20, 60, and120 days from the start of the experiment. Error bars represent theconfidence interval of the mean.
creases of up to 1.7 times compared to the non-treatedcontrol. However, the effect was significant in all para-meters (plant height, root fresh and dry weight) onlyat concentrations of 109 CFU ml−1 and in the twofirst dates assessed (30 March and 5 May). Also, dryfresh weight was less affected by the inoculum con-centration than fresh weight. This may indicate someincrease in root water contents due to the treatmentwith the growth promoting strain.
A direct relationship between root weight and in-oculum concentration was observed. A linear rela-tionship of the form log10 y = 3.4 + 1.18 log10 x(R2 = 0.89, P = 0.0015) was observed between rootfresh weight and inoculum concentration. A relation-ship of the form y = 1.54 + 0.049 log10 x (R2 = 0.87,P = 0.0021) was observed between root dry weightand inoculum concentration.
Colonization and survival of inoculated strains
Low population levels of Pseudomonas resistant torifampicin were detected in the non-treated controlplants as a background level ranging from 101 to 102
CFU g−1 f.w. The origin of these bacteria was un-known. As was reported by other authors (Landa etal., 2002) it may be due to cross contamination duringthe manipulation, watering or even sample processingin the laboratory. Therefore, this background levelwas subtracted from the population levels observedin treated plants. The mean value of the populationlevels assessed in the non-treated controls expressedas log10 CFU g−1 was subtracted from the value ofeach replicate corresponding to each treatment.
10735.tex; 2/05/2003; 12:57; p.8
PDF OUTPUT
9
Table 5. Effect of the best strains of P. fluorescens and P. agglomerans on growthpromotion of rootstock Marianna 2624 in long-term trials
Strain First trial1 Second trial2
Root Aerial Aerial Root Aerial Aerial
weight3 plant plant weight3 plant plant
(g f.w.) height weight (g f.w.) height weight
(cm) (g f.w.) (cm) (g f.w.)
EPS607 111.7 a 135.5 ab 114.8 a – – –
EPS394 105.6 ab 137.0 bc 99.5 a – – –
EPS282 98.9 ab 156.2 a 112.8 a 10.8 a 114.9 a 19.9 a
EPS435 94.8 ab 143.3 ab 96.4 a 8.4 ab 111.3 a 18.2 ab
EPS599 94.8 ab 134.7 bc 100.0 a – – –
EPS231 94.6 ab 133.2 bc 93.5 a – – –
EPS547 87.1 ab 140.5 abc 106.9 a 6.2 b 109.2 a 17.8 ab
EPS512 – – – 8.6 ab 109.4 a 19.3 a
Non-treated 60.4 c 89.9 c 89.9 a 5.9 b 111.9 a 15.9 b
1Plants were treated by watering three times with a suspension of 108 CFU ml−1.Measurements were done 5 months after the fist treatment.2Plants were treated by watering six times with a suspension of 108 CFU ml−1. Meas-urements were done 3 months after the first inoculation.3Results are the mean of three replicates consisting of six plants per replicate in arandomized block trial. Means followed by the same letter are not significantly differentaccording to the Waller–Duncan test (P = 0.05).
In the second trial (Figure 2) the population levelsof the strains in the root system decreased withtime. Values for EPS282 on Marianna 2624 and forEPS427 on GF-677 after the treatment were 1.3 ×107 CFU g−1 f.w. and 2.0 × 107 CFU g−1f.w., re-spectively. At the end of the assay, after 3 and 2.5months, the levels decreased to 8.0 × 104 and 7.0 ×104 CFU g−1, respectively.
In the dose–response assay (Figure 3) the root pop-ulation level of the inoculated strain EPS282 at theend of the experiment after 3 months of inoculationand five treatments was dependent on the inoculumconcentration applied. Population levels ranged from1 × 102 CFU g f.w.−1 at inoculations of 104 CFUml−1to 2.5 × 104 CFU g f.w.−1 at inoculations of 109
CFU ml−1 A linear relationship of the form log10 y =0.47 + 0.45 log10 x (R2 = 0.95, P = 0.0001) wasobserved between population levels on the roots andthe inoculum concentration.
Characterization of the most efficient strains
Growth promotion activity was not associated toany particular bacterial species, pattern of antagon-ism or metabolite production (Table 7). Among thebest strains all except EPS435 produced siderophores.Many P. agglomerans strains produced IAA (except
EPS512 and EPS394), only two strains (EPS453 andEPS607) gave moderate to high antibiosis, and noneproduced cyanide. Most P. fluorescens strains pro-duced cyanide, except EPS599 and were isolated fromroots. Strains CHAO, JBR170, SBW25, Ps15, EPS231and EPS326 gave strong to moderate antibiosis invitro which was associated to the production of cy-anide or phloroglucinol or PCA. The CHAO strainexhibited in vitro inhibition of all the pathogens andproduced cyanide, phloroglucinol, IAA, chitinase andsiderophores.
Discussion
The abundance of growth promoting bacterial strainsactive in Prunus rootstocks during the first stage ofselection with young plantlets was high (15%) onthe plum Marianna 2624 and low (1%) on the hy-brid GF-677. Only five to eight strains were retainedin a long-term second selection step, including plantdevelopment stages from plantlet to 3–5-month age,and based on the consistency of the effect. These res-ults are in agreement with other screening studies inpotato, radish, wheat and canola (Becker and Cook,1988; Burr et al., 1978; Kloepper et al., 1980, 1988)showing that from a starting collection of several hun-
10735.tex; 2/05/2003; 12:57; p.9

PDF OUTPUT
10
Table 6. Effect of the best strains of P. fluorescens and P. agglomerans on growthpromotion of rootstock GF-677 in long term trials
Strain First trial1 Second trial2
Root Aerial Aerial Root Aerial Aerial
weight3 plant plant weight3 plant plant
(g f.w.) height weight (g f.w.) height weight
(cm) (g f.w.) (cm) (g f.w.)
EPS453 83.5 a 173.6 ab 160.6 ab 3.8 ab 79.8 ab 8.3 ab
EPS475 77.4 a 160.9 abc 170.3 ab – – –
EPS353 76.2 a 155.5 bc 143.1 ab 4.1 ab 68.1 b 7.1 b
EPS427 72.6 ab 156.0 bc 132.2 b 4.5 a 72.1 ab 7.6 ab
EPS512 67.2 ab 160.0 bc 143.4 ab – – –
Non-treated 42.3 c 148.0 c 129.8 b 2.6 b 74.5 ab 8.3 ab
1Plants were treated by watering three times with a suspension of 108 CFU ml−1.Measurements were done 5 months after the fist treatment.2Plants were treated by watering six times with the corresponding bacterial suspensionat 108 CFU ml−1. Measurements were done 3 months after the first inoculation.3Results are the mean of three replicates consisting of six plants per replicate.Means followed by the same letter are not significantly different according to theWaller–Duncan test (P = 0.05).
Table 7. Antagonism and secondary metabolite production of the best growth promoting strains in Prunus rootstocks
Species Strain Origin1 In vitro antagonism2 Secondary metabolites3
Plant Organ Pe Ea Ps Sv HCN IAA Sid Chit Phlo Pca
P. fluorescens CHAO Tobacco Root + + + + + + + + + −JBR1-70 Wheat Root + + + + + − + − + −SBW25 Tomato Root + + − + + − + − − −Ps15 Sugarcane Root − + + + + − + − + −EPS231 Pear Root + + + − + − + − − −EPS326 Lactuca Root + + + − + − + − − −EPS599 Crataegus Flower + + − − − − + − − −EPS282 Legume Root − + − − + − + − − −EPS353 Pear Bud − + − − + − + − − −
P. agglomerans EPS453 Peach Flower + + + + − + + − nd nd
EPS607 Pear Leaf − + + + − + + − nd nd
EPS512 Apple Bud − + − − − − + − nd nd
EPS547 Plum Flower − + − − − + + − nd nd
EPS394 Cherry Flower − − − − − − + − nd nd
EPS427 Apricot Flower − − − − − + + − nd nd
EPS435 Quince Bud − − − − − + − − nd nd
EPS475 Peach Bud − − − − − + + − nd nd
EPS622 Apple Leaf − − − − − + + − nd nd
1Plant and organ of origin.2In vitro antagonism against P. expansum (Pe), E. amylovora (Ea), P. syringae (Ps) and S. vesicarium (Sv).3Secondary metabolite production: hidrogen cyanide (HCN), indoleacetic acid (IAA), siderophores (Sid), chitinases (Chit),phloroglucinol (Phlo) and phenazine (PCA). nd, not determined.
10735.tex; 2/05/2003; 12:57; p.10

PDF OUTPUT
11
Figure 3. Effect of the inoculum concentration of Pseudomonasfluorescens EPS282 on EPS282 root population levels (A), and onplant height (B) and root weight (C) of plum Marianna 2624 . Thetreatments were done by watering five times the plants with 50 mlof the bacterial suspension in phosphate buffer. Plant height wasassessed three times at March 30 (�), May 5 (�) and May 23 (�),2001. Root fresh (�) and dry (�) weight and bacterial populationlevels were assessed at the end of the assay. Error bars represent theconfidence interval of the mean.
dreds of strains, generally a 20–25% exhibited certaindegree of growth promotion activity, but in the finalsteps of selection, based on efficacy and consistencyonly 1–3% were suitable for further use.
In the present study, more strains of P. fluorescensthan of P. agglomerans were effective as rootstockgrowth promoters when applied by irrigation. Thismay be due to a better adaptation of P. fluorescensto the soil root environment than P. agglomerans ac-cording to several reports which have described that P.fluorescens is commonly isolated from the root system(O’Sullivan and O’Gara, 1992), whereas P. agglomer-
ans is more frequent in the aerial plant part (Lindow etal., 1983).
One of the factors influencing the growth pro-motion effect on Prunus rootstocks is the inoculumconcentration of the bacterial strain used for irriga-tion. A dose–response relationship was found betweenthe intensity of root growth promotion and the in-oculum concentration. This dose–response depend-ence observed was in agreement with the dose–response relationships observed between biologicalcontrol levels and dose of biocontrol agent (Johnson,1994; Montesinos and Bonaterra, 1996) or diseaselevels and pathogen dose (Vanderplank, 1975). Thebest concentration of strain EPS282 in Marianna 2624was 109 CFU ml−1 because it produced increases inboth the aerial plant height and root weight. How-ever, a concentration of 108 CFU ml−1 seems moresuitable from the practical point of view. This dosecorrespond to 1010 CFU l−1 of potting mix and waswithin the range of dose reported generally as suitablein many studies (Bahme et al., 1988; Utkhede andSmith, 1992).
Increases of plant height after treatment withgrowth promotion strains were intense during the firstmonth of plant development, but the effect decreasedwith plant age. The strains Pseudomonas fluorescensEPS282 and Pantoea agglomerans EPS427 producedheight increases of up to 3.5 times in young plantletsand of root increases of about 1.9 times. Growth stim-ulation effects reported in the literature by the use ofPGPR strains are similar to those reported by us onPrunus rootstocks. However most reports deal withherbaceous plants, including chick-pea, soybean, sug-arcane, pea, and canola (Dileep Kumar and Dube,1992; Dileep Kumar et al., 2001; Kloepper et al.,1988; Mirza et al., 2001). Information on growth stim-ulation in woody plants is less abundant and limited toa few species. Weight increases on M7 and M26 applerootstocks of 1.5 to 2.0 times were observed aftertreatment with fluorescent Pseudomonas and someEnterobacteria in greenhouse and field trials (Caesarand Burr, 1987). Shoot growth increases of 1.2 to2.0 times upon treatment with Bacillus subtilis strainEBW-4 were reported in apple trees in a 4-year study(Utkhede and Smith, 1992). Also, plant weight in-creases of 1.8 times were reported on Casuarina aftertreatment with Azospirillum (Rodriguez-Barrueco etal., 1991).
The observation that the effect of growth stimula-tion on Prunus rootstocks decreased with plant age hasbeen previously described in other studies with differ-
10735.tex; 2/05/2003; 12:57; p.11

PDF OUTPUT
12
ent plant species. This may be attributed to the fact thatthe conditions of rootstock rhizosphere may changeduring the growing period, and thus, the efficacy ofa strain can be reduced or can give some inconsistencyin its performance (Gagné et al., 1993; Germida andWalley, 1996; Kloepper et al., 1988).
The growth-promoting strains persisted reasonablywell in the Prunus root environment, and its popula-tion level was affected by the inoculum concentrationapplied and by the stage of plant development. Dur-ing the long-term experiment consisting of repeatedinoculation at 108 CFU ml−1, strains EPS282 andEPS427 had a population level of 107 CFU g f.w.−1 atthe young plantlet stage but decreased to 105 CFU gf.w.−1, when plants mature at the end of the assay.The decrease in population levels is related to the de-crease of the plant promotion effect observed at theend of the experiments. This reduction of the bac-terial population seems to be common to other PGPRstudies and may indicate that at the end of the as-say the rhizosphere conditions were not optimal forthe PGPR development or may be due to a displace-ment of the PGPR strains by other autochthonousbacteria. The population levels reported by us on rootsof treated Prunus rootstocks are comparable to the4.8–5.3 log10CFU g f.w.−1 found on bean (Andersonand Guerra, 1985), 3.1–4.1 log10 CFU g f.w.−1 ob-served on potato (Bahme et al., 1988) or 4.1 log10CFU g f.w.−1 reported on maize after inoculation withPGPR bacteria (Chabot et al., 1996).
A high specificity was observed between severalgrowth-promoting strains and the type of rootstock.Strains of P. fluorescens EPS383 and EPS286 wereonly active in GF-677, whereas strains EPS231 andEPS588 were only active in Marianna 2624. Addi-tionally, differences in efficacy were observed amongstrains being active on both rootstock types, but ingeneral, the efficacy was higher in Marianna than inGF-677. Similar results describing strain–host plantspecificity have been reported in other plant systemssuch as several herbaceous crops (Howie and Echandi,1983; Kloepper, 1996). This specificity appears to berelated to the different composition of the rhizosphere-exudates depending on the plant species which affectthe levels of colonization and subsequently the effic-acy of the PGPR strains (Latour et al., 1996), or tothe specific compounds present in the root-exudatesthat may stimulate in the bacteria the synthesis ofsecondary metabolites implicated in the plant growthpromotion (van Overbeek and van Elsas, 1995).
The best strains which exhibited growth promotionof Prunus rootstock were very diverse in secondarymetabolite production and antagonistic ability againstsome plant pathogens. However, taking into accountall strains with plant growth promotion activity, acharacteristic pattern of antagonism or metabolite pro-duction was not observed. This is in agreement withother studies that have found a wide functional andgenetic diversity between PGPR strains (Ellis et al.,2000; Rainey et al., 1994). However, in some strainscertain phenotypic characteristics could be associatedto rootstock growth promotion. Production of IAA invitro by strain EPS427 and most of the P. agglom-erans strains may be implicated in the promotion ofrootstock growth. Other reports indicate that whilemany plant growth-promoting bacteria can produceat least small amounts of IAA, high IAA producersare often inhibitory to root growth (Barazani andFriedman, 1999). Therefore, the role of IAA is notalways consistent with the plant growth promotionamong bacteria and may depend on the plant speciesbecause plant responses to hormones vary consider-ably (Gutierrez-Mañero et al., 1996). However, morespecific research should be conducted to prove its dir-ect implication. The production of cyanide by strainEPS282 and by most of the P. fluorescens strainsgreatly contrasts with its ability to promote growth.Cyanide production can increase the competitive fit-ness of the PGPR strains and enhance the suppressionof deleterious rhizobacteria (Mazzola et al., 1992) orcan stimulate the exudation of nutrients from plantroots (Åström, 1991). However, cyanide productionhas been reported to cause deleterious effects on plantdevelopment by inhibition of root growth on someplant–PGPR systems (Schippers et al., 1991). Moststrains produced siderophores. Siderophores can se-quester iron in the rhizosphere making it unavailableto other rhizosphere microorganisms, then increasingthe fitness of the PGPR strains (Kloepper et al., 1980).
A very striking example of growth promotionstrain of rootstock plants, is P. fluorescens CHAO,which produced also IAA and siderophores. TheCHAO strain exhibited a wide range spectrum ofantagonism and produce several antimicrobial com-pounds in vitro such as cyanide, phloroglucinol, andchitinase. Other strains like JBR1-70, SBW25, PS15,EPS231, EPS453, EPS607 and EPS326 also werestrongly antagonistic to the bacteria and fungal indic-ator strains used. It seems that the antagonistic activitycan be an interesting trait because many PGPR strainsin which the mode of action has been investigated
10735.tex; 2/05/2003; 12:57; p.12

PDF OUTPUT
13
appear to enhance plant growth promotion indirectlyby reducing populations of deleterious rhizobacteria(Kloepper, 1992). However, some studies pointed thata broad-spectrum of antibiosis has not always correl-ated well with growth promotion activity (Burr et al.,1978).
The screening procedure used in our study wasperformed in a pathogen-free environment in a com-mercial nursery, and no artificial inoculation of rootpathogens was done. Thus, the theoretical benefits ofcertain antagonistic properties or antimetabolite pro-duction in some strains, may not be expressed inthe environmental conditions used. It is expected thatgrowth-promoting strains with high antimicrobial po-tential will provide additional advantages to Prunusrootstock plants in the presence of root pathogens, ordeleterious bacteria. However, further studies are re-quired to show if the suppression of specific pathogensof Prunus rootstocks from the root system increase theeffect of growth promotion. To investigate the involve-ment of bacterial metabolites in rootstock growth-promotion, experiments using reductionist moleculartechniques will be performed.
Finally, the fact that several strains of P. agglomer-ans and P. fluorescens were effective and consistentlyenhanced development of Prunus rootstocks after ir-rigation with relatively diluted bacterial suspensions,opens the possibility of its use in commercial nurs-eries. The effect of these strains on plant rootstockdevelopment is particularly interesting because an op-timal growth during the first year is essential for goodestablishment in the field and also it is an advantage toshorten the time required for plant production. How-ever, additional work has to be done to set-up suitablemethods of mass production and formulation of thebacteria to be used in practice.
Acknowledgements
We are grateful to colleagues and institutions whosekindly provided the reference strains used in thepresent work (Table 1). We also acknowledge J.Pereda, L. Aguiño, O. Montojo, and C. Fernándezfor skillful assistance in laboratory and nursery work.This work was funded in part by project PETRI 95-0266-OP of the Comisión Interministerial de Cienciay Tecnología of Spain.
References
Alemany J 2001 Characterization of metabolites produced byPseudomonas fluorescens effective in the biological control offungal phytopathogens. PhD Thesis. University of Girona.
Anderson A J and Guerra D 1985 Responses of bean to root col-onization with Pseudomonas putida in a hydroponic system.Phytopathology 75, 992–995.
Åström B 1991 Role of bacterial cyanide production in differ-ential reactions of plant cultivars to deleterious rhizospherepseudomonads. Plant Soil 133, 93–100.
Bahme J B, Schroth M N, Van Gundy S D, Weinhold A R andTolentino D M 1988 Effect of inocula delivery systems onrhizobacterial colonization of underground organs of potato.Phytopathology 78, 534–542.
Barazani O and Friedman J 1999 Is IAA the major root growthfactor secreted from plant-growth-mediating bacteria? J. Chem.Ecol. 25, 2397–2406.
Becker J O and Cook R J 1988 Role of siderophores in suppressionof Pythium species and production of increased-growth responseof wheat by fluorescent pseudomonads. Phytopathology 78, 778–782.
Benizri E, Baudoin E and Guckert A 2001 Root colonization byinoculated plant growth-promoting rhizobacteria. Biocontrol Sci.Technol. 11, 557–574.
Boruah H P and Dileep Kumar B S 2002 Biological activity ofsecondary metabolites produced by a strain of Pseudomonasfluorescens. Folia Microbiol. 47, 359–363.
Brito Alvarez M A, Gagné S and Antoun H 1995 Effect of composton rhizosphere microflora of the tomato and on the incidence ofplant growth-promoting rhizobacteria. Appl. Environ. Microbiol.61, 194–199.
Burr T J, Schroth M N and Suslow T V 1978. Increased potatoyields by treatments of seed pieces with specific strains ofPseudomonas fluorescens and P. putida. Phytopathology 68,1377–1383.
Caesar A J and Burr T J 1987 Growth promotion of apple seedlingsand rootstocks by specific strains of bacteria. Phytopathology 77,1583–1588.
Chabot R, Antoun H, Kloepper J W and Beauchamp C J 1996 Rootcolonization of maize and lettuce by bioluminiscent Rhizobiumleguminosarum biovar phaseoli. Appl. Environ. Microbiol. 62,2767–2772.
Dashti N, Zhang F, Hynes R and Smith D L 1997 Application ofplant growth-promoting rhizobacteria to soybean (Glycine max(L.) Merr.) increases protein and dry matter yield under short-season conditions. Plant Soil 188, 33–41.
Dashti N, Zhang F, Hynes R and Smith D L 1998 Plant growth pro-moting rhizobacteria accelerate nodulation and increase nitrogenfixation activity by field grown soybean [Glycine max (L.) Merr.]under short season conditions. Plant Soil 200, 205–213.
Dileep Kumar B S and Dube H C 1992 Seed bacterization with afluorescent Pseudomonas for enhanced plant growth, yield anddisease control. Soil Biol. Biochem. 24, 539–542
Dileep Kumar B S, Berggren I and Mårtenson A M 2001 Potentialfor improving pea production by co-inoculation with fluorescentPseudomonas and Rhizobium. Plant Soil 229, 25–34.
Dye D W 1969 A taxonomic study of the genus Erwinia. III The“herbicola” group. N. Z. J. Sci.12, 833–839.
Ellis R J, Timms-Wilson T M and Bailey M J 2000. Identificationof conserved traits in fluorescent pseudomonads with antifungalactivity. Environ. Microbiol. 2, 274–284.
10735.tex; 2/05/2003; 12:57; p.13

PDF OUTPUT
14
Ewing W H and Fife M A 1972 Enterobacter agglomerans (Beijer-inck) comb. nov. (the herbicola-lathyri bacteria). Int. J. Syst.Bacteriol. 22, 4–11.
Frändberg and Shnürer D 1998 Antifungal activity of chitinolyticbacteria isolated from airtight stored cereal grain. Can. J. Micro-biol. 44, 121–127.
Gagné S, Dehbi L, Le Quéré D, Cayer F, Morin J L, Lemay L,Fournier N 1993 Increase of greenhouse tomato fruit yields byplant growth-promoting rhizobacteria (PGPR) inoculated intothe peat-based growing media. Soil Biol. Biochem. 25, 269–272.
Gardner J M, Chandler J L and Feldman A W 1984 Growthpromotion and inhibition by antibiotic-producing fluorescentpseudomonads on citrus roots. Plant Soil 77, 103–113.
Gavini F, Mergaert J, Beji A, Mielcarek C, Izard D, Kersters K,de Ley J 1989 Transfer of Enterobacter agglomerans (Beijerink1888) Ewing and Fife 1972 to Pantoea gen. nov. as Pantoeaagglomerans comb. nov. and description of Pantoea dispersasp.nov. Int. J. Syst. Bacteriol. 39, 337–345.
Germida J J and Walley F L 1996 Plant growth-promoting rhizobac-teria alter rooting patterns and arbuscular mycorrhizal fungicolonization of field-grown spring wheat. Biol. Fertil. Soils. 23,113–129.
Gutierrez-Mañero F J, Acero N, Lucas J A and Probanza A 1996The influence of native rhizobacteria on European alder (Alnusglutinosa (L.) Gaertn.) growth. Plant Soil 182, 67–74.
Handelsman J and Staab E V 1996 Biocontrol of soilborne plantpathogens. Plant Cell 8, 1855–1869
Howie W J and Echandi E 1983 Rhizobacteria: Influence of cul-tivar and soil type on plant growth and yield of potato. Soil Biol.Biochem. 15, 127–132.
Johnson K B 1994 Dose-response relationships and inundativebiological control. Phytopathology 84, 780–784.
Keel C, Schinder U, Maurhofer M, Voisard M, Laville J, BurgerU, Wirthner P, Haas D and Défago G 1992 Suppresion of rootdiseases by Pseudomonas fluorescens CHAO: importance of thebacterial secondary metabolite 2,4-diacetylphloroglucinol. Mol.Plant-Microbe Interact. 5, 4–13.
King E O, Ward M K and Rainey D E 1954 Two simple media forthe demonstration of the pyocyanin and fluoroscein. J. Lab. Clin.Med. 44, 301–307
Kloepper J W 1992 Plant growth-promoting rhizobacteria as biolo-gical control agents, p. 255–274. In F. Blaine Metting, Jr. (ed.),Soil Microbial Ecology. Marcel Dekker, New York.
Kloepper J W 1996 Host specificity in microbe-microbe interac-tions. Bioscience 46, 406–409.
Kloepper J W, Schroth M N and Miller T D 1980 Effects ofrhizosphere colonization by plant growth-promoting rhizobac-teria on potato plant development and yield. Phytopathology 70,1078–1082.
Kloepper J W, Hume D J, Scher F M, Singleton C, Tipping B,Laliberté M, Frauley K, Kutchaw T, Simonson C, Lifshitz R,Zaleska I and Lee L 1988 Plant growth-promoting rhizobacteriaon canola (rapeseed). Plant Dis. 72, 42–46.
Landa B B, Mavrodi O V, Raaijmakers J M, McSpadden Gardener BB, Thomashow L S and Weller D M 2002 Differential ability ofgenotypes of 2,4-diacetylphloroglucinol-producing Pseudomo-nas fluorescens strains to colonize the roots of pea plants. Appl.Environ. Microbiol. 68, 3226–3237.
Latour X, Corberand T, Laguerre G, Allard F and Lemanceau P1996 The composition of fluorescent pseudomonad populationassociated with roots is influenced by plant and soil type. Appl.Environ. Microbiol. 62, 2449–2456.
Leeman M, van Pelt J A, Hendrickx M J, Scheffer R J, Bakker PA H M and Schippers B 1995 Biocontrol of Fusarium wilt of
radish in commercial greenhouse trials by seed treatment withPseudomonas fluorescens WCS374. Phytopathology 85, 1301–1305.
Lindow S E, Arny D C and Upper C D 1983 Biological control offrost injury: An isolate of Erwinia herbicola antagonistic to icenucleation active bacteria. Phytopathology 73, 1097–1102.
Liu L, Kloepper J W and Tuzun S 1995 Induction of systemicresistance in cucumber against bacterial angular leaf spot byplant-promoting rhizobacteria. Phytopathology 85, 843–847.
Maniatis T, Fritsch E F and Sambrook J 1982 Molecular Cloning: Alaboratory manual. Cold Spring Harbor Laboratory, Cold SpringHarbor, NY.
Maurhofer M, Hase C, Meuwly P, Métraux J-P and Défago G1994 Induction of systemic resistance of tobaccco to tobacconecrosis virus by the root-colonizing Pseudomonas fluorescensstrain CHAO: Influence of the gacA gene and of pyoverdineproduction. Phytopathology 84, 139–146.
Mazzola M, Cook R J, Thomashow L S, Weller D M and PiersonIII L S 1992 Contribution of phenazine antibiotic biosynthesis tothe ecological competence of fluorescents pseudomonads in soilhabitats. Appl. Environ. Microbiol. 58, 2616–2624.
Miller T D and Schroth M N 1972 Monitoring the epiphytic pop-ulation of Erwinia amylovora on pear with selective media.Phytopathology 62, 1175–1182.
Mirza M S, Ahmad W, Latif F, Haurat J, Bally R, Normand P andMalik K A 2001 Isolation, partial characterization, and the effectof plant growth-promoting bacteria (PGPB) on micro-propagatedsugarcane in vitro. Plant Soil 237, 47–54.
Montesinos E and Bonaterra A 1996 Dose-response models in bio-logical control of plant pathogens: An empirical verification.Phytopathology 86, 464–472.
Montesinos E, Bonaterra A, Ophir Y and Beer S V 1996 Antag-onism of selected bacterial strains to Stemphylium vesicariumand biological control of brown spot of pear under controlledenvironment conditions. Phytopathology 86, 856–863.
O’Sullivan D J and O’Gara F 1992 Traits of fluorescent Pseudo-monas spp. involved in suppression of plant root pathogens.Microbiol. Rev. 56, 662–676.
Palleroni N J 1984 Genus I Pseudomonas. In Bergey’s Manual ofDeterminative Bacteriology, 8 ed. Eds. N R Krieg and J G Holtpp. 141–199. Williams & Wilkins Co., Baltimore, MD.
Rainey P B, Bailey M J and Thompson I P 1994 Phenotypic andgenotypic diversity of fluorescent pseudomonads isolated fromfield grown sugarbeet. Microbiology 140, 2315–2331.
Raupach G S and Kloepper J W 1998 Mixtures of plant growthpromoting rhizobacteria enhance biological control of multiplecucumber pathogens. Phytopathology 88, 1158–1164.
Rodríguez-Barrueco C, Cervantes E, Subbarao N S and Rodríguez-Caceres E 1991 Growth promoting effect of Azospirillum brasi-lense on Casuarina cunninghamiana Miq. seedlings. Plant Soil.135, 121–124.
Schippers B A, Bakker W, Bakker P and Van Peer R 1991 Benefi-cial and deleterious effects of HCN-producing pseudomonads onrhizosphere interactions. Plant Soil 129, 75–83.
Schwyn B and Neilands J B 1987 Universal chemical assay for thedetection and the determination of siderophores. Anal. Biochem.160, 47–56.
Sneath P H A 1966. Identification methods applied to Chromobac-terium. pp. 15–20. In Identification Methods for Microbiologist,Part A. Eds B M Gibb and F A Skinner pp. 15–20. AcademicPress, London.
Utkhede R S and Smith E M 1992 Promotion of apple tree growthand fruit production by the EBW-4 strain of Bacillus subtilis inapple replant disease soil. Can. J. Microbiol. 38, 1270–1273.
10735.tex; 2/05/2003; 12:57; p.14

PDF OUTPUT
15
Vanderplank J E 1975 Principles of Plant Infection. Academic Press,New York.
van Overbeek L S and Van Elsas J D 1995 Root exudate-inducedpromoter activity in Pseudomonas fluorescens mutants in thewheat rhizosphere. Appl. Environ. Microbiol. 61, 890–898.
Weller D M, Raaijmakers J M, McSpadden Gardener B B and
Thomashow L S 2002 Microbial populations responsible forspecific soil suppressivesess to plant pathogens. Annu. Rev.Phytopathol. 40, 309–348.
Wodzinski R S and Paulin J P 1994 Frequency and diversity of an-tibiotic production by putative Erwinia herbicola strains. Appl.Bacteriol. 76, 603–607.
10735.tex; 2/05/2003; 12:57; p.15




















